HUWE1抑制剂BI8622的新应用的制作方法
本发明涉及医药技术领域,具体涉及一种huwe1抑制剂bi8622的新应用。
背景技术:
炎症小体激活在宿主的炎症发生、抵抗病原体入侵,自身免疫性疾病发生中都发挥重要作用。经典的炎症小体包括nlrp3,aim2和nlrc4炎症小体,其激活分别通过识别不同的刺激物,最终都会导致细胞半胱氨酸蛋白酶-1(caspase-1)的激活,从而诱导细胞产生成熟的白介素1β(il-1β),白介素18(il-18),激活的caspase-1可裂解消化道皮肤素d(gsdmd),导致细胞焦亡的发生(inflammasomes:mechanismofaction,roleindisease,andtherapeuticshaitaoguo,justinbcallaway&jennyp-yting;naturemedicinevolume21,pages677–687(2015).)。炎症小体激活异常与多种疾病发生相关,通过靶向炎症小体激活的药物研发越来越多。
huwe1抑制剂bi8622是一类小分子化合物,结构式如下:
bi8622由stefaniepeter等人筛选得到,通过泛素化修饰功能可特异性抑制泛素连接酶huwe1,并发现其对肿瘤发生发展有一定的抑制作用(peters,bultinckj,myantk,jaenickela,walzs,müllerj,etal.tumorcell-specificinhibitionofmycfunctionusingsmallmoleculeinhibitorsofthehuwe1ubiquitinligase.embomolmed.2014;6:1525–1541.),但未见有研究报道huwe1抑制剂bi8622可用于抑制炎症小体激活。
技术实现要素:
本发明所要解决的技术问题是提供一种huwe1抑制剂bi8622在制备抑制炎症小体激活药物中的应用。
本发明的bi8622其化学结构式如下:
本发明中,所述炎症小体包括nlrp3、aim2和nlrc4。
本发明中,所述nlrp3炎症小体由脂多糖和三磷酸腺苷共同激活,所述的aim2炎症小体由弗朗西斯菌或dsdna激活,所述的nlrc4炎症小体由沙门氏菌激活,所述的nlrp3和aim2炎症小体同时由李斯特菌激活。
本发明提供了一种抑制炎症小体激活的药物,所述的抑制炎症小体激活药物包括活性成分bi8622及药物学上可接受的载体。
本发明提供了一种huwe1抑制剂bi8622在制备痛风、ii型糖尿病或系统性红斑狼疮的炎症性疾病药物中的应用。
本发明提供了一种痛风、ii型糖尿病或系统性红斑狼疮的炎症性疾病药物,所述的药物包括活性成分bi8622及药物学上可接受的载体。
本发明的技术方案与现有技术相比,本发明的有益效果为:
本发明将上述bi8622对小鼠原代bmdm细胞、人单核巨噬细胞系thp1细胞和人原代pbmc细胞分别进行了对激活的nlrp3、aim2和nlrc4炎性小体及细胞死亡的影响,结果表明,bi8622无论是在小鼠bmdm细胞,还是人单核巨噬细胞系thp1以及人原代pbmc细胞中,均可以显著抑制炎症小体的激活,并且显著降低细胞死亡事件的发生。
本发明首次证实huwe1抑制剂bi8622的新用途:可用于抑制验证小体的激活,表明bi8622在治疗炎症小体异常激活疾病(如痛风,ii型糖尿病,系统性红斑狼疮等炎症性疾病)中具有潜在药用价值。
附图说明
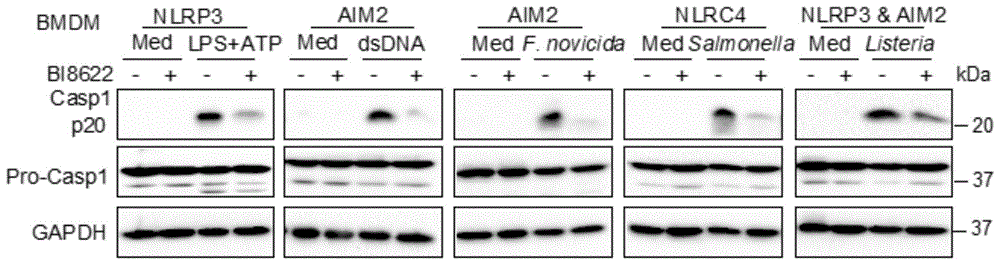
图1为通过bi8622处理组和对照组处理后,小鼠原代bmdm细胞中活化的caspase1p20的表达的对比图;
图2为通过bi8622处理组和对照组处理后,小鼠原代bmdm细胞中il-1β表达水平的对比图;
图3为通过bi8622处理组和对照组处理后,小鼠原代bmdm细胞中tnf表达水平的对比图;
图4为通过bi8622处理组和对照组处理后,小鼠原代bmdm细胞中il-6表达水平的对比图;
图5为通过bi8622处理组和对照组处理后,小鼠原代bmdm细胞死亡比例的对比图;
图6为通过bi8622处理组和对照组处理后,人thp1细胞il-1β表达水平的对比图;
图7为通过bi8622处理组和对照组处理后,人thp1细胞tnf表达水平的对比图;
图8为通过bi8622处理组和对照组处理后,人thp1细胞il-6表达水平的对比图;
图9为通过bi8622处理组和对照组处理后,人thp1细胞死亡比例的对比图;
图10为通过bi8622处理组和对照组处理后,人原代pbmc细胞il-1β表达水平的对比图;
图11为通过bi8622处理组和对照组处理后,人原代pbmc细胞tnf表达水平的对比图;
图12为通过bi8622处理组和对照组处理后,人原代pbmc细胞il-6表达水平的对比图;
图13为通过bi8622处理组和对照组处理后,人原代pbmc细胞死亡比例的对比图。
具体实施方式
本发明的具体实施例中,健康c57/bl小鼠由中国科学院昆明动物研究所提供[实验动物许可证号:syxk(滇)k2013-0013]。
本发明中的huwe1抑制剂bi8622购自medchemexpress公司,货号hy-120929。
本发明中所用的人单核巨噬细胞系thp1来源于中国科学院昆明动物研究所细胞库,也可采用本领域中的其他单核巨噬细胞细胞。
nlrp3炎症小体通常由外源分子如革兰氏阴性菌细胞壁组分、核酸、穿孔素,内源性分子如三磷酸腺苷(atp)、尿酸结晶等所激活(cassels.l.,eisenbarths.c.,iyers.s.,etal.,thenalp3inflammasomeisessentialforthedevelopmentofsilicosis.proc.natl.acad.sci.usa,2008,105(26):9035-9040.kannegantit.-d.,body-malapelm.,amera.,etal.criticalroleforcryopyrin/nalp3inactivationofcaspase-1inresponsetoviralinfectionanddouble-strandedrna.j.biol.chem.,2006,281(48):36560-36568.)。
nlrc4炎症小体由沙门氏菌(salmonella)感染所激活(mariathasans.,newtonk.,monackd.m.,etal.differentialactivationoftheinflammasomebycaspase-1adaptorsascandipaf.nature,2004,430(6996):213-218.)。
aim2炎症小体由弗朗西斯菌及双链dna(dsdna)诱导激活(fernandes-alnemrit.,yuj.-w.,julianac.,etal.,theaim2inflammasomeiscriticalforinnateimmunitytofrancisellatularensis.nat.immunol.2010,11:385-393.bürckstümmert.,baumannc.,blümls.,etal.anorthogonalproteomic-genomicscreenidentifiesaim2asacytoplasmicdnasensorfortheinflammasome.nat.immunol.2009,10:266-272.)。
在本发明中,若无特殊说明,所有的原料组分均为本领域技术人员熟知的市售商品。
下面将结合本发明中的实施例,对本发明中的技术方案进行清楚、完整地描述。显然,所描述的实施例仅仅是本发明一部分实施例,而不是全部的实施例。基于本发明中的实施例,本领域普通技术人员在没有做出创造性劳动前提下所获得的所有其他实施例,都属于本发明保护的范围。
实施例1
bi8622对小鼠原代bmdm细胞中nlrp3,aim2,nlrc4三种激活的炎症小体及细胞死亡的影响试验。
材料与方法:
小鼠原代bmdm细胞的制备:将适龄成年(如4-6周)的c57/bl小鼠安乐死,在无菌条件下分离股骨、胫骨并置于pbs缓冲液中,用手术刀刮去骨表面的肌肉、软骨及骨骺,将骨两端切开露出骨髓腔后,用1ml注射器吸取dmem-f12培养基冲洗骨髓腔,收集细胞,细胞吹打均匀后,计数并调整细胞密度为1-2million/ml,在dmem-f12完全培养基中添加20ng/ml的m-csf生长因子,将细胞接种在细胞培养板中,在5%co2、37℃的co2培养箱内培养,静置培养5天后收取贴壁细胞行相应的实验。
受试化合物bi8622用二甲基亚砜(dmso)配制成浓度为10mm的储存液,处理细胞前用dmem-f12完全培养基稀释成10um的工作浓度,对照组选用0.1%dmso处理,实验组bi8622和对照组处理时间均为3h,随后进行后续炎症小体刺激物刺激与检测实验。
炎症小体刺激物为:1)激活nlrp3炎症小体的脂多糖(lps)+三磷酸腺苷(atp);2)激活aim2炎症小体的弗朗西斯菌(f.novicida)及双链dna(dsdna);3)激活nlrc4炎症小体的沙门氏菌(salmonella);4)既可以激活nlrp3炎症小体又可激活aim2炎症小体的李斯特菌(listeria)。
具体的刺激方式:用终浓度为500ng/ml的lps刺激1h,终浓度5mm的atp刺激15min,从而激活nlrp3炎症小体;用终浓度为1.5ugx-fect转染dsdna刺激1h或用200moi的f.novicida刺激12h,从而激活aim2炎症小体;用终浓度为3moi的salmonella刺激2h,从而激活nlrc4炎症小体;用终浓度为30moi的listeria刺激3h,从而同时激活nlrp3和aim2炎症小体。
同时选用未添加刺激物的培养上清(media)用作0h对照组。
运用westernblot检测培养上清中半胱氨酸蛋白酶-1(caspase-1)激活切割产物p20的表达水平以及细胞裂解液中caspase-1前体的表达水平;同时用酶联免疫法(elisa)检测白介素1β(il-1β)、肿瘤坏死因子(tnf)和白介素6(il-6)的表达水平;通过检测培养上清中乳酸脱氢酶(ldh)的分泌水平进行细胞死亡情况检测(ldh法)。
结果表明,在小鼠原代bmdm细胞中,bi8622处理组细胞与dmso对照组细胞相比,在lps+atp、dsdna、f.novicida、salmonella以及listeria刺激处理后其活化的caspase-1p20的表达均显著下降(图1,其中med组代表未添加刺激物的培养上清对照组,casp1代表caspase-1)。elisa检测结果表明,lps+atp、f.novicida、salmonella以及listeria刺激处理后,bi8622处理组细胞il-1β的分泌明显低于对照组细胞,dsdna处理组细胞中il-1β表达水平较低,两组细胞间没有差异(图2);而两组细胞tnf的表达水平在上述几种处理方式中均没有明显差异(图3);il-6的表达除在f.novicida、salmonella刺激后bi8622处理组比对照组有所下降,其他刺激处理两组细胞均没有明显差异(图4)。ldh检测结果表明,上述几种刺激方式中bi8622处理组细胞细胞死亡均低于dmso对照组细胞(图5)。以上结果表明,bi8622在小鼠bmdm细胞中可显著抑制nlrp3,aim2和nlrc4炎症小体的激活,降低细胞的死亡。
实施例2
bi8622对人thp1细胞中激活的nlrp3,aim2,nlrc4三种炎症小体及细胞死亡的影响试验。
thp1细胞为人单核巨噬细胞,培养条件为1640培养基加10%fbs,具体的培养方法按照常规方法即可。受试化合物bi8622用二甲基亚砜(dmso)配制成浓度为10mm的储存液,处理细胞前用1640完全培养基稀释成10um的工作浓度,对照组选用0.1%dmso处理,实验组bi8622和对照组处理时间均为3h,随后进行后续炎症小体刺激物刺激与检测实验。
炎症小体刺激物为:1)激活nlrp3炎症小体的脂多糖(lps)+三磷酸腺苷(atp);2)激活aim2炎症小体的双链dna(dsdna);3)激活nlrc4炎症小体的沙门氏菌(salmonella)。
具体的刺激方式:用终浓度为500ng/ml的lps刺激1h,终浓度5mm的atp刺激15min,从而激活nlrp3炎症小体;用终浓度为1.5ugx-fect转染dsdna刺激1h,从而激活aim2炎症小体;用终浓度为3moi的salmonella刺激2h,从而激活nlrc4炎症小体。
同时选用未添加刺激物的培养上清(media)用作0h对照组。
运用elisa法来检测il-1β、tnf和il-6的表达,采用ldh法检测细胞死亡情况。
结果表明,采用lps+atp、dsdna、salmonella刺激处理thp1细胞后,bi8622处理组细胞il-1β的分泌明显低于对照组细胞(图6);bi8622处理组细胞tnf及il-6的表达水平同样低于对照组细胞(图7);细胞中tnf的表达水平在上述几种处理方式中均没有明显差异(图8);ldh法检测同样发现bi8622处理组thp1细胞在lps+atp、dsdna、salmonella处理后细胞死亡明显低于对照组细胞(图9)。结果表明,同小鼠bmdm细胞类似,bi8622在人thp1细胞中可显著抑制nlrp3,aim2和nlrc4炎症小体的激活,可明显降低细胞的死亡。
实施例3
bi8622对人原代pbmc细胞中激活的nlrp3,aim2,nlrc4三种炎症小体及细胞死亡的影响试验。
从正常供者体内取出外周血10ml,用密度梯度离心法分离得到外周血单个核细胞(pbmc)[中国科学院昆明动物研究所生命科学伦理委员会审批,编号:smkx-20191201-01]。将分离得到的pbmc细胞在1640培养基+10%fbs中培养,具体的培养方法按照常规方法即可。受试化合物bi8622用二甲基亚砜(dmso)配制成浓度为10mm的储存液,处理细胞前用1640完全培养基稀释成10um的工作浓度,对照组选用0.1%dmso处理,实验组bi8622和对照组处理时间均为3h,随后进行后续炎症小体刺激物刺激与检测实验。选用刺激物及具体刺激方式具体参见实施例2,同时选用未添加刺激物的培养上清(media)用作0h对照组。运用elisa法来检测il1-β、tnf和il-6的表达,采用ldh法检测细胞死亡情况。
结果表明,采用lps+atp、dsdna、salmonella刺激处理pbmc细胞后,bi8622处理组细胞il-1β的分泌明显低于对照组细胞(图10);bi8622处理组细胞tnf的表达在lps+atp和salmonella刺激后低于对照组pbmc细胞,dsdna刺激两组细胞tnf表达无明显差异(图11);bi8622处理组pbmcil-6的表达水平在salmonella刺激后明显低于对照组,而lps+atp和dsdna刺激后无明显差异(图12)。ldh检测同样发现bi8622处理组人原代pbmc在lps+atp、dsdna、salmonella处理后细胞ldh分泌水平明显低于对照组pbmc,提示bi8622处理组人原代pbmc死亡明显低于对照组(图13)。以上结果表明,bi8622在人原代pbmc中可显著抑制nlrp3,aim2和nlrc4炎症小体的激活,可明显降低细胞的死亡。
通过实施例中的结果可知,本发明首次证实huwe1抑制剂bi8622的新用途:bi8622既可以显著抑制炎症小体的激活,又显著降低细胞死亡事件的发生。
以上所述仅是本发明的优选实施方式,应当指出,对于本技术领域的普通技术人员来说,在不脱离本发明原理的前提下,还可以做出若干改进和润饰,这些改进和润饰也应视为本发明的保护范围。
- 还没有人留言评论。精彩留言会获得点赞!