一种脑海绵状血管瘤细胞模型的构建方法及其在新药筛选中的应用与流程
本发明属于生物医药技术领域,尤其涉及一种脑海绵状血管瘤细胞模型的构建方法及其在新药筛选中的应用。
背景技术:
脑海绵状血管瘤(cerebral carvernous malformation,CCM)是发生在中枢神经系统的一种血管错构畸形,是先天性脑血管畸形的一种,约占脑血管畸形的10%~20%,人群患病率约0.1%~4%。
CCM由缺乏肌层和弹性纤维的大小不等的海绵状血管窦组成,团状毛细血管扩张,内壁为单层扁平的内皮细胞(endothelial cells,ECs),缺乏周细胞(pericyte,PCs)覆盖,无正常的血管基膜。由于血管内皮屏障发生改变,导致血管通透性增高,红细胞外渗,并继发脑实质炎症反应,从而显著增加中风、癫痫、神经功能缺失的风险。
CCM分为散发性和常染色体显性遗传,遗传性CCM与3个致病基因种系突变相关,包括CCM1(KRIT1,7q11.2-q21)、CCM2(MGC4607、7p15-p13)和CCM3(PDCD10、3q25.2-q27)。90%以上的CCM病人携带这3个致病基因的突变。近十几年来,3个CCM致病基因种系突变的研究报道非常多。由于CCM病灶组织内体细胞基因突变的发现提出了“二次打击”机制(two-hit mechanism)学说,认为其参与了脑海绵状血管瘤的发病机制,导致表达于脑海绵状血管瘤毛细血管腔内皮细胞的致病基因编码蛋白完全失去功能。这种学说最近已经在小鼠动物模型中被证实。出生后特异的失活内皮细胞的Ccm基因,会导致大脑出现CCM样的损伤。虽然CCM发病机制尚不清楚,但分子遗传学研究为我们深入理解CCM的发病机制和临床治疗提供了重要依据。鉴于目前缺乏有效的治疗药物,手术切除是CCM的唯一治疗手段,因此,研究CCM的发病机制,寻找新的有 效药物是当前的研究热点。
药物筛选模型是寻找和发现新药物的重要条件之一。目前药物的筛选方法主要在包括体内和体外筛选方法,体内筛选方法成本高,操作难度大,不利于高通量的药物筛选;体外筛选方法主要是在分子和细胞水平上实现的,具有材料用量少,作用机制明确,易用于大规模筛选等优点,是药物筛选的主要方法。但目前尚无公认的CCM细胞模型,因此,有必要建立一种CCM细胞模型,加速CCM药物开发进程。
技术实现要素:
一方面,本发明的目的在于克服现有技术存在的不足之处而提供了一种脑海绵状血管瘤细胞模型的构建方法。本发明的细胞模型具有血管内皮细胞出芽的增多、扩大的管腔形成和减少的周细胞募集等表型,符合人脑海绵状血管瘤的病理特点,非常适合用于人脑海绵状血管畸形发病机制研究和开发相应的治疗药物,也可用于药物的筛选、药效观察和药效评价。
本发明采用的技术方案为:一种脑海绵状血管瘤细胞模型的构建方法,包括:
将转染了CCM3基因siRNAs的血管内皮细胞接种在单层融合的成纤维细胞之上共培养;或
将转染了CCM3基因siRNAs的血管内皮细胞包被在微载体球珠上,然后将微载体球珠包埋在纤维蛋白胶里,继而在纤维蛋白胶上层铺上成纤维细胞共培养。
作为对上述技术方案的进一步改进,所述血管内皮细胞表达绿色荧光蛋白(GFP)。GFP作为一种标记蛋白,其内源荧光基团在受到紫外光或蓝光激发时可高效发射清晰可见的绿光。所述的GFP还包括在野生型GFP基础上进行改进的蛋白,比如进行基因优化(如去除隐蔽性内含子等);或通过氨基酸替换提高GFP脱辅基蛋白在高温(如37℃)条件下的正确折叠,以及改变GFP光谱特性等。因此,作为本发明的优选方式,所述的GFP是增效的绿色荧光蛋白(EGFP),EGFP为对绿色荧光蛋白进行改进后的蛋白,与GFP具有很高的同源性,在表达后发光的效果更为理想。本领域人员很容易获得GFP或EGFP的序列,例如,可从pEGFP-N1质粒上通过PCR或酶切的方式获得。
作为对上述技术方案的进一步改进,所述共培养的时间为7至14天。
作为对上述技术方案的进一步改进,所述微载体球珠为cytodex球珠。
作为对上述技术方案的进一步改进,所述微载体球珠还同时包埋有正常的周细胞。
作为对上述技术方案的更进一步改进,所述血管内皮细胞与周细胞的包埋数量比例为2:1。全身系统各器官ECs:PCs的比例不一,其中脑和视网膜中ECs:PCs的比例最高,接近1:1至2:1,形成独特的血脑屏障和血视网膜屏障,我们选择ECs:PCs为2:1的比例是因为周细胞比血管内皮细胞长得快,在此比例下能更好地模仿体内脑和视网膜中血管内皮细胞的出芽,而在ECs:PCs较低的比例(如1:1)条件下周细胞限制内皮细胞的出芽。
作为对上述技术方案的进一步改进,所述血管内皮细胞表达绿色荧光蛋白,所述正常周细胞表达红色荧光蛋白(mCherry)。
作为对上述技术方案的进一步改进,所述血管内皮细胞为HBMVECs(人脑微血管内皮细胞)。
作为对上述技术方案的进一步改进,所述CCM3基因siRNA为以下三个siRNA的混合物:
sc-62084A:
正义链序列:GGAUAUAGCUAGUGCAAUAtt,
反义链序列:UAUUGCACUAGCUAUAUCCtt;
sc-62084B:
正义链序列:CAACCGACUAAUUCAUCAAtt,
反义链序列:UUGAUGAAUUAGUCGGUUGtt;
sc-62084C:
正义链序列:GGAUUAUUGCCAUCUUACAtt,
反义链序列:UGUAAGAUGGCAAUAAUCCtt。
另一方面,本发明还提供了根据所述的脑海绵状血管瘤细胞模型的构建方法得到的脑海绵状血管瘤细胞模型。
又一方面,本发明还提供了一种筛选治疗脑海绵状血管瘤的潜在物质的方法,包括:
1)将候选物质与所述脑海绵状血管瘤细胞模型接触;
2)检测所述候选物质对脑海绵状血管瘤细胞模型的影响,若所述候选物质可抑制因CCM3缺失引起的血管内皮细胞出芽的增加和扩大的管腔的形成,则表明所述候选物质是用于治疗脑海绵状血管瘤的潜在物质。
作为对上述技术方案的进一步改进,所述步骤1)包括:在测试组中,将候选物质加入到所述脑海绵状血管瘤细胞模型中;和/或
步骤2)包括:检测测试组的体系中所述候选物质对脑海绵状血管瘤细胞模型的影响,并与对照组比较,其中所述的对照组是不添加所述候选物质的所述脑海绵状血管瘤细胞模型;如果测试组中候选物质抑制因CCM3缺失引起的血管内皮细胞出芽的增加和扩大的管腔的形成在统计学上高于对照组,就表明该候选物是治疗脑海绵状的潜在物质。
作为对上述技术方案的进一步改进,所述候选物质选自siRNA、抗体、化合物。
当所述候选物质为siRNA时,所述步骤1)具体为将候选物质转染至所述脑海绵状血管瘤细胞模型中;当所述候选物质为抗体或化合物时,所述步骤1)具体为将候选物质加入到所述脑海绵状血管瘤细胞模型中。
相对于现有技术,本发明的有益效果为:
因此,本发明脑海绵状血管瘤细胞模型具有血管内皮细胞出芽的增加、扩大的管腔形成和减少的周细胞募集等表型,符合人脑海绵状血管瘤的病理特点,非常适合用于人脑海绵状血管畸形发病机制研究和开发相应的治疗药物,也可 用于药物的筛选、药效观察和药效评价。
本发明设计合理,提供的筛选和评价系统结构完善,具有很高的应用价值,而且设备简单、经济,适合于在早期开展对于治疗心血管疾病药物(siRNA、抗体、化合物)的筛选。
附图说明
图1显示了器官型的血管生成实验结果;表达绿色荧光蛋白EGFP和/或siRNAs转染的HBMVECs被种在单层融合的成纤维细胞之上,在7至14天共培养过程中观察管腔形成过程;VE-cadherin和collagen IV免疫荧光染色显示CCM3缺失的内皮细胞出芽和管腔形成明显增加,如图1A所示;定量分析显示血管分支的数量、平均管腔直径、和成管的面积(collagen IV覆盖的区域)在CCM3敲低组明显增加(如图1A、B所示);因CCM3缺失引起的增加的EC出芽和扩大的管腔形成不受正常IgG处理的影响,但能被同时沉默UNC13B表达或用ANGPT2中和抗体所抑制(如图1A、B所示);其中,图1A中的箭指示出芽分支;图1B中的三幅图分别从左至右依次显示siCtrl、siUNC13B、siCCM3、siCCM3/siUNC13B和siCCM3/αAngpt2等五组的血管分支的数量、平均管腔直径、和成管的面积的相应参数值及定量分析比较。
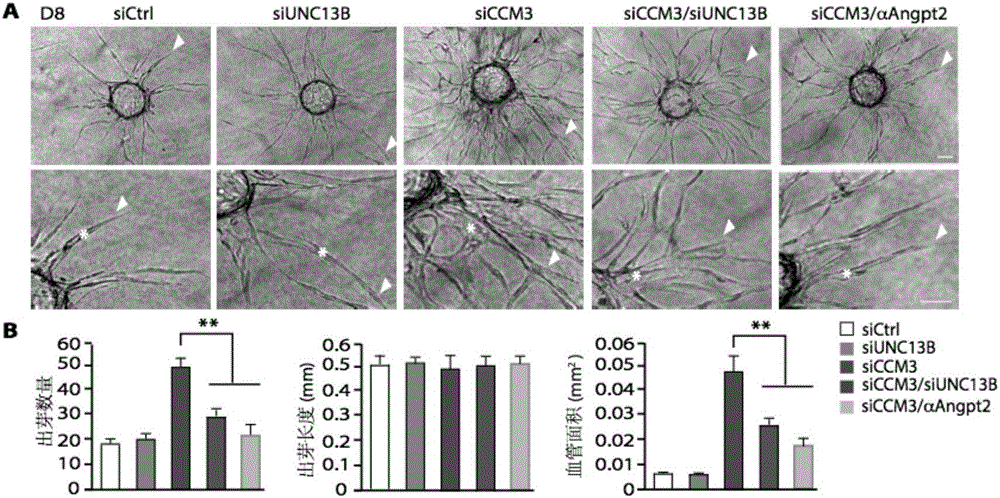
图2显示了3D出芽实验结果;该实验中ECs包被在cytodex球珠上,然后cytodex球珠被埋在fibrin胶里,继而在胶上层铺上成纤维细胞;我们发现,因CCM3敲低引起的增加的EC出芽和扩大的管腔形成能被同时沉默UNC13B表达或用ANGPT2中和抗体所抑制(如图2A、B所示);其中,图2A中的箭头指示出芽,星号指示管腔;图2B中的三幅图分别从左至右依次显示siCtrl、siUNC13B、siCCM3、siCCM3/siUNC13B和siCCM3/αAngpt2等五组的血管出芽数量、出芽长度、和成管面积的相应参数值及定量分析比较。
图3显示了周细胞募集结果;为了同时观察出芽过程中的EC和PC,我们把表达绿色荧光蛋白EGFP的HBMVEC转染不同的siRNAs(包括Ctrl或CCM3或UNC13B)和表达红色荧光蛋白mCherry的正常HBMVPC同时包被在cytodex 球珠上。在出芽过程中,PC被招募到正常的EC上,但CCM3敲低的EC不能招募PC(如图3A所示)。然而,在EC同时沉默UNC13B表达或用ANGPT2中和抗体或两者的结合能恢复正常的PC募集到出芽EC上(如图3A、B所示);其中,图3A中的箭指示PC能被招募到EC上,箭头指示CCM3敲低组的EC不能招募PC;图3B中的三幅图分别从左至右依次显示siCtrl、siUNC13B、siCCM3、siCCM3/siUNC13B、siCCM3/αAngpt2、siCCM3/siUNC13B/αAngpt2等六组的血管出芽数量、和周细胞覆盖率的相应参数值及定量分析比较。
具体实施方式
为更好的说明本发明的目的、技术方案和优点,下面将结合具体实施例对本发明作进一步说明。
Ccm3与CCM3可替换使用;Ang-2、Angpt2、ANGPT2可替换使用;Unc13b、Unc13B、UNC13B可替换使用;αAngpt2即Angpt2中和抗体。
实施例
1、材料与方法
1.1 Organotypic血管生成实验
我们采用经改进的Organotypic血管生成实验(Organotypic血管生成实验可参见:Hetheridge C,Mavria G and Mellor H.Uses of the in vitro endothelial-fibroblast organotypic co-culture assay in angiogenesis research.Biochem Soc Trans.2011;39:1597-600.)。用逆转录病毒(MOI 20)转染HBMVECs使其表达绿色荧光蛋白(EGFP),2天后再转染siRNAs到细胞体内;或直接给未转染EGFP的HBMVECs转染siRNAs,1天后消化HBMVECs,并在预先铺上2×104人成纤维细胞并达到融合的24孔板内每孔接种含或不含EGFP表达的经siRNAs转染的8.5×103的HBMVECs。我们每两天给细胞换EGM2培养液,并在2至14天在荧光显微镜下直接通过绿色荧光蛋白观察小管形成,或通过VE-cadherin和collagen IV免疫荧光染色观察小管形成。
1.2三维出芽实验
用LipofectamineRNAiMAX(Invitrogen)转染siCtrl,siUNC13B,siCCM3或siCCM3与siUNC13B的联合到HBMVECs作用8小时或过夜,接着给HBMVECs换成新鲜的微血管内皮细胞生长液(Microvascular Endothelial Cell Growth Medium-2MV,Lonza)培养。我们参照文献(可参见:Nakatsu MN and Hughes CC.An optimized three-dimensional in vitro model for the analysis of angiogenesis.Methods Enzymol.2008;443:65-82.)进行三维出芽实验。细胞转染24小时后,即被消化并与一定数量的微载体珠(C3275,Sigma)混匀在含有1.5MLEGM2-MV培养液的流式细胞管里,放进培养箱培养,4小时内每隔20分钟轻拍打管壁,以使最终每个微载体珠包被有接近400个细胞。第二天,把这些包被有HBMVECs的微载体珠包埋在2mg/ml的纤维蛋白胶里,继而把20,000/孔的成纤维细胞铺在纤维蛋白胶上层,并给每孔细胞1.0ML含或不含ANGPT2中和抗体(1μg/ml或10μg/ml;美国Genentech公司或广州道瑞医药科技有限公司)的EGM2-MV培养液。每两天给细胞换液并从2至13天通过显微镜观察细胞出芽状况、管腔形成、血管分支形成及融合过程。
1.13 HBMVECs和HBMVPCs共培养系统的三维出芽实验
我们参照文献进行血管内皮细胞和周细胞的相互作用的三维出芽实验(可参见:Waters,J.P.,et al.In vitro self-assembly of human pericyte-supported endothelial microvessels in three-dimensional coculture:a simple model for interrogating endothelial-pericyte interactions.J Vasc Res 50,324-331(2013);Chang,W.G.,Andrejecsk,J.W.,Kluger,M.S.,Saltzman,W.M.&Pober,J.S.Pericytes modulate endothelial sprouting.Cardiovasc Res 100,492-500(2013);Scheppke,L.,et al.Notch promotes vascular maturation by inducing integrin-mediated smooth muscle cell adhesion to the endothelial basement membrane.Blood 119,2149-2158(2012).)。首先用逆转录病毒(MOI 20)转染HBMVECs使其表达绿色荧光蛋白(EGFP),用腺病毒(MOI 20)转染HBMVPCs使其表达mCherry。继而用LipofectamineRNAiMAX(Invitrogen)转染siCtrl,siUNC13B,siCCM3或siCCM3与siUNC13B的联合到HBMVECs。细胞转染24小时后,即被消化并按ECs:PCs(2:1)的比例与一定数量的微载体珠(C3275,Sigma)混匀,培养在含有1.5ML EGM2-MV培养液的流式细胞管里,放进培养箱,4小时内每隔20分钟轻拍打管壁,以使最终每个微载体珠包被有接近400个细胞。我们选择ECs:PCs(2:1) 的比例是因为周细胞比血管内皮细胞长得快,且在ECs:PCs(1:1)的比例条件下周细胞限制内皮细胞的出芽。第二天,把这些包被有HBMVECs的微载体珠包埋在2mg/ml的纤维蛋白胶里,继而把20,000/孔的成纤维细胞铺在纤维蛋白胶上层,并给每孔细胞1.0ML含或不含ANGPT2中和抗体(1μg/ml或10μg/ml;美国Genentech公司或广州道瑞医药科技有限公司)的EGM2-MV培养液。每两天给细胞换液并从2至8天通过荧光显微镜观察内皮细胞出芽状况、血管内皮细胞形成小管的周细胞的覆盖率。
我们所用的siRNA由美国Santa Cruz公司合成,人的UNC13B基因和CCM3基因的siRNA均为三个有效siRNA的混合物。
UNC13B基因的siRNA为以下三个siRNA的混合物:
sc-42022A:
正义链序列:GGAUUGCGCUGAAGACUAUtt
反义链序列:AUAGUCUUCAGCGCAAUCCtt
sc-42022B:
正义链序列:GACAUCAAGUCAAGAGUAAtt
反义链序列:UUACUCUUGACUUGAUGUCtt
sc-42022C:
正义链序列:GGAUCGACCUCUCUACAUAtt
反义链序列:UAUGUAGAGAGGUCGAUCCtt
人的CCM3基因siRNA为以下三个有效siRNA的混合物:
sc-62084A:
正义链序列:GGAUAUAGCUAGUGCAAUAtt,
反义链序列:UAUUGCACUAGCUAUAUCCtt;
sc-62084B:
正义链序列:CAACCGACUAAUUCAUCAAtt,
反义链序列:UUGAUGAAUUAGUCGGUUGtt;
sc-62084C:
正义链序列:GGAUUAUUGCCAUCUUACAtt,
反义链序列:UGUAAGAUGGCAAUAAUCCtt。
2.结论
我们用体外实验EC出芽模型和管腔形成模型分析CCM3对内皮出芽和管腔形成的影响。表达绿色荧光蛋白EGFP和/或siRNAs转染的HBMVECs被种在单层融合的成纤维细胞之上,在7至14天共培养过程中观察管腔形成过程。VE-cadherin和collagen IV免疫荧光染色显示CCM3缺失的内皮细胞出芽和管腔形成明显增加(如图1A所示)。定量分析显示血管分支的数量、平均管腔直径、和成管的面积(collagen IV覆盖的区域)在CCM3敲低组明显增加(如图1A、B所示)。因CCM3缺失引起的增加的EC出芽和扩大的管腔形成不受正常IgG处理的影响,但能被同时沉默UNC13B表达或用ANGPT2中和抗体所抑制(如图1A、B所示)。
为了进一步证实CCM3调控EC出芽和管腔形成,我们用3D出芽实验来验证,该实验中ECs包被在cytodex球珠上,然后cytodex球珠被埋在fibrin胶里,继而在胶上层铺上成纤维细胞。我们发现,因CCM3敲低引起的增加的EC出芽和扩大的管腔形成能被同时沉默UNC13B表达或用ANGPT2中和抗体所抑制(如图2A、B所示)。
3D出芽实验也是被用来研究周细胞或平滑肌细胞募集和血管成熟的一个系统。为了同时观察出芽过程中的EC和PC,我们把表达绿色荧光蛋白EGFP的HBMVEC转染不同的siRNAs(包括Ctrl或CCM3或UNC13B)和表达红色荧光蛋白mCherry的正常HBMVPC以2:1的比例同时包被在cytodex球珠上。在出芽过程中,PC被招募到正常的EC上,但CCM3敲低的EC不能招募PC。然而,在EC同时沉默UNC13B表达或用ANGPT2中和抗体或两者的结合能恢复正常的PC募集到出芽EC上(如图3A、B所示)。我们的结果提示CCM3能通过抑制EC中ANGPT2的胞吐作用,维护正常EC细胞连接、EC管腔形成和PC募集。
3.结论分析
(1)用体外实验EC出芽模型和管腔形成模型分析CCM3对内皮出芽和管腔形成的影响。定量分析显示血管分支的数量、平均管腔直径、和成管的面积(collagen IV覆盖的区域)在CCM3敲低组明显增加。
(2)为了进一步证实CCM3调控EC出芽和管腔形成,我们用3D出芽实 验来验证,我们发现,因CCM3敲低引起EC出芽的增加和扩大的管腔形成。
(3)为了同时观察出芽过程中的EC和PC,证实CCM3调控周细胞募集,我们把表达绿色荧光蛋白EGFP的HBMVEC转染CCM3siRNAs和表达红色荧光蛋白mCherry的正常HBMVPC同时包被在cytodex球珠上。在出芽过程中,PC被招募到正常的EC上,但CCM3敲低的EC不能招募PC。
(4)体外实验EC出芽模型和管腔形成模型中,因CCM3缺失引起的增加的EC出芽和扩大的管腔形成能被同时沉默UNC13B表达所抑制;3D出芽实验中因CCM3敲低引起EC出芽的增加和扩大的管腔形成能被同时沉默UNC13B表达所抑制;周细胞募集实验中沉默UNC13B表达能将PC募集到CCM3敲低的EC上。在CCM疾病中使用UNC13B siRNA维护正常EC出芽、EC管腔形成和PC募集,UNC13B siRNA可以作为CCM候选新药。
(5)体外实验EC出芽模型和管腔形成模型中,因CCM3缺失引起的增加的EC出芽和扩大的管腔形成不受正常IgG处理的影响,但能被ANGPT2中和抗体所抑制;3D出芽实验中因CCM3敲低引起EC出芽的增加和扩大的管腔形成能用ANGPT2中和抗体抑制;周细胞募集实验中用ANGPT2中和抗体能将PC募集到CCM3敲低的EC上。在CCM疾病中使用ANGPT2中和抗体维护正常EC出芽、EC管腔形成和PC募集,ANGPT2中和抗体可以作为CCM候选新药。
以上研究证实在体外细胞模型中血管内皮细胞CCM3基因缺失能导致EC细胞连接的破坏、血管内皮细胞出芽的增加、扩大的管腔形成和减少的周细胞募集,这些EC表型与CCM疾病表型类似,能反应CCM疾病的进程,是筛选CCM治疗新药的良好平台,可以用于包括siRNA、抗体、化合物等在内的各种新药筛选。
最后所应当说明的是,以上实施例仅用以说明本发明的技术方案而非对本发明保护范围的限制,尽管参照较佳实施例对本发明作了详细说明,本领域的普通技术人员应当理解,可以对本发明的技术方案进行修改或者等同替换,而不脱离本发明技术方案的实质和范围。
- 还没有人留言评论。精彩留言会获得点赞!