一株干巴菌菌株及其发酵得到的干巴菌菌丝体锌多糖及应用的制作方法
本发明涉及菌丝体多糖
技术领域:
,特别涉及一株干巴菌菌株,还涉及此菌株液体发酵得到的干巴菌菌丝体锌多糖,及干巴菌菌丝体锌多糖的应用。
背景技术:
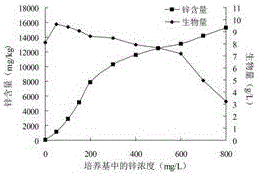
:锌是人体内执行生理功能最多的必须微量元素,广泛参与酶活性调控、物质代谢、基因表达调控、免疫系统调控、激素调节等一系列生物学过程。但是锌元素不能在体内合成和储存,只能通过每天补充获得,而每天在膳食中获得的锌元素往往不能满足人们的生理需要。据报道,世界上50%的人口面临锌摄入量不足的风险。缺锌引起的营养失衡问题是目前世界上面临的主要营养健康问题之一。由于锌元素的重要生理作用,多年来人们一直致力于研发更好的补锌产品,补锌产品不断的更新换代。第一代补锌产品为硫酸锌等无机锌盐。无机锌盐生物利用率低,服用后副作用较多,已经被逐渐淘汰。第二代补锌产品为葡萄糖酸锌等有机酸锌。有机酸锌会产生肠胃刺激,对其他微量元素的吸收容易产生拮抗作用,更不适于糖尿病人补锌。第三代补锌产品为氨基酸螯合锌。氨基酸螯合锌的生产工艺并不完善,存在蛋白原料成本高、蛋白分解要求高、有机溶剂消耗量大和对环境污染严重等问题。目前,生物态有机锌是补锌产品研制与开发的主流。微生物可以将环境中的无机锌吸收转化为微生物细胞内的生物态有机锌,这种生物态有机锌没有任何毒副作用、更加易于吸收、生物利用率高。食用真菌有很强的吸收转化微量元素的特性,以食用真菌为载体进行微量元素的生物富集已经逐渐引起人们的重视。食用真菌富集锌元素可以通过固体栽培获得富锌子实体或液体发酵培养获得富锌菌丝体这两条途径实现。其中,液体发酵培养有明显的优越性。液体发酵不仅具有生产周期短,菌丝体产量高,多糖等活性成分含量高,对锌等金属元素的有效富集率高的优点。而且可以进行大规模生产和连续化生产,没有实现人工栽培的食用真菌品种也可以通过液体发酵技术获得有价值的代谢产物。因此,液体发酵技术有利于食用真菌产业从劳动密集型产业向技术密集型产业转变,并有利于提高食用真菌产品的附加值。近年来的研究表明,食用真菌吸收富集锌元素后会形成锌蛋白和锌多糖等富锌有机物,从而将环境中的无机锌吸收转化形成锌多糖。食用真菌多糖本身具有降血糖、降血脂、抗衰老、抗氧化、提高免疫力、提高记忆力、抗肿瘤、保护肝脏等广泛的生物学功效。而锌多糖是食用真菌将无机锌转化成有机锌的有效形式之一,并且它是一种有机锌-多糖配合物,兼有多糖与有机锌两者的活性,表现出高于单一多糖或单一锌的生物活性。本课题组研究发现干巴菌在液体发酵培养时菌丝体生长旺盛,对锌元素的耐受和吸收能力强。干巴菌(ThelephoraganbajunZang)为革菌属(Thelephora)真菌,异香浓郁,珍贵稀少,仅仅分布在我国滇中800~2300m高海拔地区的弱酸性红壤地面上,与松树有外生菌根关系,目前人工栽培尚未成功。干巴菌不仅口感好而且营养价值极高,可以增强人体的免疫功能,每年7~10月份干巴菌上市時总是供不应求,优质干巴菌市场价格如今已经上涨到了2000元/kg。由于其分布的局限性,国内外对干巴菌的研究极为有限,主要集中在干巴菌的分类、菌种分离、菌种鉴定和生态环境等方面。因此,干巴菌是一种具有极高经济价值及营养价值的亟待研究开发的菌种资源。CN103719833A公开了一种富硒干巴菌粉,以干巴菌菌丝为原料,经过干燥、研磨步骤制备得到。但其说明书中没有记载硒的存在形式,也就无法判断此种形式的硒是否易被人体吸收。目前未见干巴菌液体发酵制备干巴菌菌丝体锌多糖及活性研究的报道。技术实现要素:为了解决以上现有技术中补锌产品效果差、不易吸收、肠胃刺激、生物利用度不高的问题,本申请提供了一株菌丝体锌多糖产量高并且锌多糖中锌元素含量高的干巴菌(ThelephoraganbajunZang)菌株TG-1。本发明还提供了干巴菌菌株TG-1发酵得到的干巴菌菌丝体锌多糖及制备方法。本发明还提供了干巴菌菌丝体锌多糖的应用。本发明是通过以下步骤得到的:一株干巴菌菌株TG-1,其保藏编号为CGMCCNo.12977。干巴菌菌株TG-1发酵得到的干巴菌菌丝体锌多糖,干巴菌菌株TG-1CGMCCNo.12977在添加有锌盐的液体培养基中发酵,菌丝体经过分离,提取干巴菌菌丝体锌多糖。干巴菌菌丝体锌多糖的锌含量为1000~15000mg/kg。所述的干巴菌菌丝体锌多糖,液体培养基中优选锌盐为硫酸锌、醋酸锌、氯化锌或氧化锌。所述的干巴菌菌丝体锌多糖,优选添加有锌盐的液体培养基中含有马铃薯150~250g/L,葡萄糖15~25g/L,硫酸镁0.5~2.5g/L,磷酸二氢钾0.5~2.5g/L,锌浓度50~600mg/L,在15~35℃发酵培养。所述的干巴菌菌丝体锌多糖,菌丝体经过分离,提取干巴菌菌丝体锌多糖的优选操作如下:(1)将菌丝体分离后冻干或烘干,并粉碎得到干巴菌富锌菌丝体粉末;(2)干巴菌富锌菌丝体粉末与去离子水以1﹕10~1﹕30的比例混合,调节pH值为7~9,超声波破碎,超声功率为100~800w,超声时间为5~20min,然后置于40~100℃水浴中,提取1~3h,离心取上清液;(3)离心后的富锌菌丝体残渣按步骤(2)再提取2~4次,合并上清液;(4)采用下述方法中的一种:①将上清液浓缩至原体积的3/4~1/2,上清液中加入1~3倍体积乙醇,静置至沉淀完全析出,3000~15000r/min离心5~15min,弃上清并于40~60℃条件下烘干,获得干巴菌锌多糖。②将上清液浓缩至原体积的3/4~1/2,上清液中加入1~3倍体积乙醇,静置至沉淀完全析出,3000~15000r/min离心5~15min,弃上清并于40~60℃条件下烘干,烘干后再次溶解,离心(3000~15000r/min,5~15min),上清液除蛋白,冻干,获得干巴菌锌多糖。所述的干巴菌菌丝体锌多糖在制备补锌抗衰老保健品中的用途。所述的用途,优选所述的保健品是具有清除自由基、抗氧化作用的保健品。本发明的有益效果:1)在液体发酵时干巴菌菌株TG-1具有很好的锌元素吸收转化能力,锌元素耐受能力(锌耐受浓度可以达到50~600mg/L),以及旺盛的菌丝体生长能力。干巴菌TG-1可以很好的富集锌元素,制备锌多糖,锌多糖中锌含量可以达到1000~15000mg/kg。因此,干巴菌菌株TG-1对干巴菌菌丝体锌多糖的生产以及提高锌多糖中锌的含量具有非常重要的意义;2)利用液体发酵途径,制备干巴菌菌丝体锌多糖,为干巴菌的资源化利用提供了一条有效途径;3)干巴菌菌丝体锌多糖具有显著的还原力和清除超氧阴离子自由基、羟基自由基和DPPH自由基的能力,可以显著提高衰老小鼠体内的抗氧化酶活性,同时降低小鼠体内的脂质过氧化水平。菌种保藏信息保藏时间:2016年9月23日,保藏单位:中国微生物菌种保藏管理委员会普通微生物中心,保藏编号:CGMCCNo.12977,保藏单位地址:北京市朝阳区北辰西路1号院3号中国科学院微生物研究所,邮政编码:100101分类命名:干巴菌Thelephoraganbajun。附图说明图1干巴菌菌株TG-1锌元素的富集能力,图2干巴菌菌株TG-2锌元素的富集能力,图3干巴菌菌株TG-3锌元素的富集能力,图4干巴菌菌株TG-4锌元素的富集能力,图5锌多糖MZPS和多糖MPS的抗氧化能力(A:还原力;B:羟基自由基清除能力),图6锌多糖MZPS和多糖MPS对衰老小鼠SOD水平的影响(NC为空白对照组,MC为衰老模型组,I,II,III为MPS低、中、高剂量组,IV,V,VI为MZPS低、中、高剂量组),图7锌多糖MZPS和多糖MPS对衰老小鼠T-AOC水平的影响(NC为空白对照组,MC为衰老模型组,I,II,III为MPS低、中、高剂量组,IV,V,VI为MZPS低、中、高剂量组),图8锌多糖MZPS和多糖MPS对衰老小鼠LPO含量的影响(NC为空白对照组,MC为衰老模型组,I,II,III为MPS低、中、高剂量组,IV,V,VI为MZPS低、中、高剂量组),图9锌多糖MZPS和多糖MPS对衰老小鼠MDA含量的影响(NC为空白对照组,MC为衰老模型组,I,II,III为MPS低、中、高剂量组,IV,V,VI为MZPS低、中、高剂量组)。具体实施方式下面通过具体的实施例对本发明进行详细说明,但这些例举性实施方式的用途和目的仅用来例举本发明,并非对本发明的实际保护范围构成任何形式的任何限定,更非将本发明的保护范围局限于此。实施例1干巴菌菌株的获得分别于云南省不同地区采集4个新鲜干巴菌子实体,用75%酒精进行表面消毒,再用无菌水冲洗数次后,用灭菌镊子撕取干巴菌内部组织小块接种到含氯霉素的马铃薯葡萄糖(PDA)培养基平板上,移入24℃培养箱中培养,待长出菌丝后,反复纯化,直至获得干巴菌纯培养物。于4个干巴菌子实体中分别筛选获得4株干巴菌菌种,并分别命名为TG-1、TG-2、TG-3、TG-4。实施例2干巴菌菌株TG-1、TG-2、TG-3、TG-4富锌能力分析分别取0.5cm2干巴菌菌种TG-1、TG-2、TG-3、TG-4接入PDA液体培养基中于摇床24℃震荡培养7d,然后分别将其转接入含有不同浓度锌的PDA液体培养基中(锌浓度为0、50、100、150、200、300、400、500、600、700、800mg/L,以ZnSO4·7H2O的形式添加),每个梯度5个重复,500mL三角瓶装液量为250mL,于摇床24℃震荡培养10d。发酵结束后,离心(10000r/min,10min)分离干巴菌菌丝体,并用去离子水洗涤菌丝体3遍。将菌丝体于55℃烘箱中烘干,测定菌丝体生物量及菌丝体中锌元素的含量。图1~4分别为TG-1、TG-2、TG-3、TG-4发酵时培养基中锌浓度与菌丝体生物量及锌含量的关系图。培养基中适当浓度的锌元素促进干巴菌菌丝体的生长,而锌浓度过高时产生抑制作用。干巴菌TG-1菌丝体生长旺盛,锌元素耐受能力强,菌丝体中锌元素含量高,具有良好的锌元素富集能力。干巴菌TG-2与TG-1相比菌丝的生长能力较差,菌丝体生物量较低。干巴菌TG-3与TG-1相比锌元素的耐受能力较差。而干巴菌TG-4菌丝体中锌元素的含量较TG-1低。因此,干巴菌菌株TG-1是富集锌元素的理想菌株,当锌浓度为50mg/L時,干巴菌菌丝体生物量最高,为9.63g/L。当锌浓度小于等于300mg/L時,锌元素起到促进干巴菌菌丝体生长的作用。当锌浓度大于等于400mg/L時,锌元素逐渐产生毒性作用,抑制干巴菌菌丝体的生长。实施例3干巴菌的富锌液体发酵进行发酵罐培养实验。①菌种的活化。分别将干巴菌TG-1、TG-2、TG-3、TG-4接种于活化PDA培养基平板上,24℃培养7d。②种子培养。分别取0.5cm2干巴菌活化菌种接入富锌PDA液体培养基中(马铃薯200g/L,葡萄糖20g/L,硫酸镁1g/L,磷酸二氢钾1.5g/L,锌浓度为150mg/L,以ZnSO4·7H2O的形式添加),500mL三角瓶装液量为300mL,然后于摇床24℃振荡培养10d。③发酵罐培养。装量:7.5L;接种量:10%;发酵温度:24℃;搅拌速度:350r/min;泡沫控制:接种后加两滴消泡剂。③发酵过程中pH值及DO值的测定。利用pH电极及溶氧电极自动在线测定并记录数据。④富锌菌丝体的分离。将干巴菌富锌培养物离心(10000r/min,10min),获得富锌菌丝体,反复离心洗涤菌丝体3遍后,将其置于烘箱中,55℃烘干。在干巴菌的发酵过程中,pH值逐步降低,由发酵初期的4.77逐步降低至3.86。发酵过程中菌体大量繁殖并且胞外多糖大量产生,从30h开始,培养物逐渐变得粘稠,溶氧率降低较快,发酵结束时溶氧率降至20%左右。经计算干巴菌TG-1、TG-2、TG-3、TG-4富锌菌丝体的产量分别为9.25g/L、5.96g/L、6.35g/L和7.08g/L。因此,发酵罐培养时干巴菌TG-1富锌菌丝体的产量最高。实施例4干巴菌菌丝体锌多糖的提取培养基中锌浓度为150mg/L时,分别获得菌株TG-1、TG-2、TG-3、TG-4的菌丝体,将菌丝体分别粉碎,按1:20的固液比加入去离子水,并调溶液pH=8,于超声波细胞破碎仪中(400w,15min)超声破碎,然后置于90℃水浴中2h(每隔15min搅动一次),离心(10000r/min,10min),以上步骤共重复4次,合并上清液并过140目筛,加入2倍体积的乙醇,静置24h后,离心(10000r/min,10min),收集沉淀,于55℃烘箱中烘干,再次溶于适量去离子水中,离心取上清液,加入Sevag试剂,充分振摇15min,除去变性蛋白质层及下层有机相,如此反复去蛋白3次后,冻干并称重。计算干巴菌菌丝体锌多糖的产量及锌多糖的锌含量,见表1。表1干巴菌TG-1、TG-2、TG-3、TG-4的锌多糖产量及其锌含量锌多糖产量(g/L)锌含量(mg/kg)TG-10.734300TG-20.513980TG-30.494040TG-40.583230由表1可见,干巴菌菌株TG-1的锌多糖产量及锌多糖中的锌含量显著高于菌株TG-2、TG-3、TG-4。实施例5还原力及羟基自由基清除能力的测定(1)还原力的测定采用普鲁士兰法进行还原力的测定。将TG-1菌株发酵得到的干巴菌菌丝体多糖(Myceliapolysaccharide,MPS)和干巴菌菌丝体锌多糖(Myceliazincpolysaccharide,MZPS)分别用去离子水进行梯度稀释,配置一系列不同浓度的多糖水溶液。取1mL多糖样品于试管中,加入2.5mL磷酸盐缓冲液(pH6.6,0.2mol/L)和1mL铁氰化钾溶液(10g/L),混匀后置于50℃恒温水浴锅中20min。水浴结束后,再加入2mL三氯乙酸(100g/L)和1.2mL三氯化铁溶液(1g/L)。混匀后测定700nm处吸光度,并用2,6-二叔丁-4-甲基苯酚(2,6-di-tert-butyl-4-methylphenol,BHT)作为阳性对照。(2)羟基自由基清除能力的测定采用Fenton法进行羟基自由基清除能力的测定。配置一系列不同浓度的多糖水溶液。将1mL硫酸亚铁溶液(9mmol/L),1mL水杨酸乙醇溶液(9mmol/L),1mL多糖样品溶液和1mL过氧化氢溶液(8.8mmol/L)于试管中混合均匀,37℃水浴30min。离心(5000r/min,10min)后取上清液于510nm处测定吸光度,并用BHT作为阳性对照。按照以下公式计算羟基自由基清除率(Scavengingrate,SR),SR(%)=(A0–A)/A0×100。其中,A0为空白对照的吸光度,A为多糖样品的吸光度。羟基自由基的清除能力以EC50值表示,EC50值代表羟基自由基清除50%時所需要的样品浓度。由图5(A)可见,随着多糖浓度的增加,还原力逐渐增强。在浓度为5000mg/L时,MPS、MZPS和BHT的还原力分别为1.91,2.08和1.27,三者还原力的强弱顺序为:MZPS>MPS>BHT。因此,MZPS和MPS都具有一定的还原力,且MZPS的还原力高于MPS。由图5(B)可见,MZPS,MPS和BHT的EC50值分别为875.44,1501.32和320.91mg/L。当浓度高于1000mg/L时MZPS的超氧阴离子自由基清除能力超过了BHT,三者清除能力大小为:MZPS>BHT>MPS。当浓度达到4000mg/L时,MPS的超氧阴离子自由基清除能力超过了BHT,三者清除能力大小为:MZPS>MPS>BHT。因此,MZPS和MPS都具有清除超氧阴离子自由基的能力,且MZPS的清除能力高于MPS。实施例6干巴菌锌多糖的体内抗氧化实验(1)动物模型构建与药物干预将小鼠随机分为8组(每组10只),即空白对照组(NC)、衰老模型组(MC)、MPS高、中、低干预组(I,II,III)和MZPS高、中、低干预组(IV,V,VI),适应性饲养一周后按照以下方法开始实验。(NC)空白对照组:灌胃去离子水(10mL/kg),同时腹腔注射生理盐水(5mL/kg)。(MC)衰老模型组:灌胃去离子水(10mL/kg),同时腹腔注射D-半乳糖(150mg/kg/d)。(I,II,III)高、中、低剂量MPS干预组:分别灌胃MPS(60、120和240mg/kg/d),同时腹腔注射D-半乳糖(150mg/kg/d)。(IV,V,VI)高、中、低剂量MZPS干预组:分别灌胃MZPS(60、120和240mg/kg/d),同时腹腔注射D-半乳糖(150mg/kg/d)。实验一直持续30d。(2)样品制备小鼠禁食12h后,摘眼球取血,然后颈椎脱臼处死小鼠。将血液离心(12000r/min,10min,4℃)分离血清。(3)指标检测SOD活性测定:用SOD测定试剂盒进行测定,试剂盒采用黄嘌呤氧化酶法(羟胺法)测定SOD活力。T-AOC测定:用T-AOC测定试剂盒进行测定。测定原理为抗氧化物质,能使Fe3+还原成Fe2+,后者可与菲啉类物质形成稳固的络和物,通过比色可测出其抗氧化能力的高低。LPO含量测定:取0.5mL样品于离心管中,并加入2.5mL硫代巴比妥酸乙酸溶液(2.68g/L),混匀后置于沸水浴中30min。冷却后加入7mL正丁醇,离心(10000r/min,10min),取上清液于520nm处测定吸光度。同时用40nmol/mL1,1,3,3-四乙氧基丙烷(1,1,3,3-tetraethoxy-propane,TEP)作为对照,按照以下公式计算LPO浓度(C):C(nmol/mL)=A/A0×40。A为样品的吸光度,A0为TEP的吸光度。MDA含量测定:将0.5mL样品、1.5mL去离子水和2mL硫代巴比妥酸溶液(6g/L)混合均匀,置于沸水浴中15min,迅速冷却后,离心(10000r/min,10min),上清液于450、532和600nm处测定吸光度,按照以下公式计算MDA浓度(C):C(μmol/L)=6.45×(A532nm–A600nm)–0.56×A450nm。MPS和MZPS对衰老小鼠SOD活性的影响见图6。机体抵抗自由基伤害的防御体系有酶促与非酶促两个体系组成,SOD是酶促防御体系的关键酶,它可以有效的清除自由基反应的启动因子超氧阴离子自由基,从而抑制自由基反应,降低自由基反应的毒害作用。因此,SOD活性反映了机体清除自由基的能力,是机体抗氧化能力的重要指标。由图6可知,与空白对照组(NC)小鼠相比,衰老模型组(MC)小鼠体内的SOD活性显著降低(P<0.01)。而在MPS(I,II,III)和MZPS(IV,V,VI)干预下,小鼠体内的SOD活性显著上升,并呈现出一定的量效关系。在低、中、高剂量MPS(I,II,III)干预下,小鼠血清中SOD活性较D-半乳糖致衰老组小鼠分别提高了40.12%,86.89%和109.81%。在低、中、高剂量MZPS(IV,V,VI)干预下,小鼠血清中SOD活性分别提高了36.56%,100.97%和122.92%。MPS和MZPS对衰老小鼠T-AOC水平的影响见图7。非酶促防御体系主要包含维生素E、胡萝卜素、维生素C、半胱氨酸、蛋氨酸、色氨酸、转铁蛋白和乳铁蛋白等维生素、氨基酸和金属蛋白质,它们主要通过以下途径发挥抗氧化作用:清除体内的自由基;阻断过氧化链;分解过氧化物;除去起催化作用的金属离子。T-AOC水平反映了机体抵抗自由基伤害的非酶促防御能力,研究表明衰老时各组织中的T-AOC水平明显降低。由图7可以看出,腹腔注射D-半乳糖30d后,衰老模型组(MC)小鼠血清中的T-AOC水平显著低于空白对照组(P<0.01)。与衰老模型组(MC)小鼠相比较,MPS(I,II,III)和MZPS(IV,V,VI)干预组T-AOC水平随着多糖剂量的增加而升高。当MPS灌胃剂量为80,160,240mg/kg/d時(III),小鼠血清中T-AOC水平分别升高了21.10%,29.22%和98.26%。当MZPS灌胃剂量为80,160,240mg/kg/d時(VI),T-AOC水平分别升高了14.65%,21.75%和107.76%。MPS和MZPS对衰老小鼠LPO含量的影响见图8。脂质过氧化反应的产物LPO具有一定的毒性,可以影响细胞膜的流动性和细胞的代谢,进而导致细胞损伤和凋亡。LPO的积累与机体衰老和某些疾病的发生、发展过程有着密切的联系。因此,LPO的含量作为组织损伤的一项指标,被广泛应用于抗氧化的研究。如图8所示,衰老模型组(MC)小鼠各组织中LPO含量显著升高(P<0.01)。而MPS(I,II,III)和MZPS(IV,V,VI)显著降低了D-半乳糖致衰老小鼠血清中的LPO含量,并且表现出了明显的量效关系。在低、中、高剂量MPS干预组(III)中,小鼠血清LPO抑制率分别为12.34%,40.73%和56.67%。在低、中、高剂量MZPS干预组(VI)中,小鼠血清LPO抑制率分别为20.33%,57.08%和61.64%。MPS和MZPS对衰老小鼠MDA含量的影响见图9。脂质过氧化反应的产物LPO是不稳定的,它会进一步分解成为一系列复杂的化合物,其中,MDA是最主要的分解终产物,它可以导致DNA的损伤。因此,MDA的含量可以间接反映机体脂质过氧化的程度,是评价氧化损伤和衰老的重要指标之一。如图9所示,MDA含量在衰老模型组(MC)小鼠各组织中显著升高,与空白对照组(NC)小鼠相比存在显著差异性(P<0.01)。而在MPS(I,II,III)和MZPS(IV,V,VI)干预下,D-半乳糖致衰老小鼠血清中的MDA含量显著降低,并且呈现出了一定的量效关系。在低、中、高剂量MPS干预组(I,II,III)中,小鼠血清的MDA抑制率分别为14.84%,35.10%和48.29%。在低、中、高剂量MZPS干预组(VI,V,VI)中,小鼠血清的MDA抑制率分别为24.26%,65.96%和72.20%。上述实施例为本发明较佳的实施方式,但本发明的实施方式并不受实施例的限制,其它任何未背离本发明的精神实质与原理下所做的改变、修饰、组合、替代、简化均应为等效替换方式,都包含在本发明的保护范围之内。当前第1页1 2 3
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1