生长生物体的方法和系统与流程
本申请是原申请的申请日为2013年7月10日、申请号为201380040913.x、发明名称为《用于抑制和/或控制生物系统生长的含有寄生性、致病性或杂草生物系统的核酸的组合物》的中国专利申请的分案申请。
本发明涉及用于抑制和/或控制生物系统生长的含有寄生性、致病性或侵染性的生物系统的核酸的组合物及其制备方法。更具体地说,本发明涉及一种方法和相关的系统,通过分离由所述生物体产生的核酸,允许改进在生物反应器或光生物反应器中高产率的微生物的生产/生长、或改进在多种培养系统中的植物的生产/生长。
背景技术:
多年以来,对抑制生物系统的新产品的研究需要科学上、经济上和工业上的巨大努力,其包括体内和体外的对天然来源或通过合成方式获得的活性成分的检测以及连续的生物和药理试验,以及现场和临床应用测试(morgan等,2011.卫生政策(healthpolicy),100:4-17)。
用于抑制生物系统的药物用途(灭害剂、除草剂、抗生素)涉及各个应用领域,其中主要有农业领域和医药领域,它最终完成了对有致病性、寄生性或者是侵染性的有害生物的控制、抑制或消灭,并且对抗微生物生物膜的形成进行处理。
尽管这样的应用范围是广阔的、多样化的,但是,药物的用途显示了共同的问题(i)作用的特异性,(ii)对人类和其它物种的可能的毒性,(iii)对环境的污染,(iv)靶标生物体种群抗药性的出现。
通常,药物(灭害剂、除草剂、抗生素)由小型和中型大小的有机分子组成,并在化学成分和作用机制方面显示出很大的多样化。这样的药物已被普遍基于“随机筛选”方法检测出,随机筛选是从来自自然产生的主链的各种基本结构开始以随机方式进行合成,对合成得到的化学化合物的活性和使用潜力进行估计。因此,在过去的50年中,数量非常庞大的产品在市场上销售,其中最近还包括多核苷酸。
用在农业和医药中的当前活性成分的一个主要问题是,对于寄生性、致病性或侵染性的生物体来说,其作用的特异性不够,进而对宿主生物体带来副作用和毒性。例如,已知的百草枯与帕金森氏病的发病是关联的,该产品已被证明对人类胚胎是致命的。除了急性毒性的问题以外,长期暴露于一些产品中显示出了确认的致癌风险,为此,强制规范要求生态毒理学的监测,以及与环境动力学和各种产品的持久性有关的使用模式。
最后,由药物(灭害剂、除草剂、抗生素)引起的环境污染问题是不可忽视的。环保领域法规的最新运用已经大大减少了对植物用药和灭害剂的使用的认证,因为更严格的法规的生效,许多产品在不久的将来只能被弃用。例如,甲基溴,其作为熏蒸剂广泛用于控制土壤中的包括线虫在内的病原体,由于甲基溴被归入于被认为是破坏臭氧层的化合物因而已被禁用。考虑到这些问题,多数更现代的活性成分被选定为在土壤或农作物中快速分解。无论如何,在环境污染方面的这个可能的优点会限制所使用的活性成分的经时有效性。
无论是在农业领域还是医学领域的处理应用中,对于目前使用的产品来说,另一个共有的基本问题是,在开始处理一段时间后,出现了药理学抗性。
例如,就农业来说,在美国,在六十年代初期,2,4-d已经是第一个除草剂,被大规模地使用,对于2,4-d,报道了胡萝卜寡毛实蝇(dacuscarota)种群出现抗药性(switzer,1957年,proc.northeasternweedcont.conf.,11:315;whitehead和switzer,1963年,can.j.plantsci.,43:255)。接着,从70年代初期开始,大量报道了侵染性的物种对敌菌灵(triazines)有抗药性的案例(ryan,1970年,除草科学(weedsci.),18:614;lebaron和gressel,eds.,植物的除草剂抗药性(herbicideresistanceinplants),wiley,纽约,1982年)。然后,伴随着用于生化战的新产品的扩散和它们的密集使用,该记录在随后的几十年中呈指数增长(powles,shaner,2001年,除草剂抗药性和世界粮食(herbicideresistanceandworldgrains),crc出版社,纽约)。
对除草剂处理的侵染性抗药性的各种出现机制是已知的。例如,在文献中报道的各种机制是:选择靶标基因的多个拷贝而导致抗药性(gaines等,2010年,proc.natl.acad.sci.107(3):1029-1034);靶标基因的突变(wakelin等,2006年,weedres.(牛津),46(5):432-440);空泡封存(ge等,2010年,虫害管理科学(pestmanagementsci.)66:576);使除草剂代谢的酶的表达(hidayat等,1997年,病虫害生化生理学(pest.biochem.physiol.)57(2):137-146)。
目前,在农业领域中,超过200种侵染性的物种的抗药性出现已被检测、选择,首先是因为使用了以相同作用机理为特征的活性成分。在意大利的波河平原,侵染性草本植物种群对莠去津的抗药性案例是已知的,比如龙葵(solanumnigrum)、白藜(chenopodiumalbum)和繁穗苋(amaranthuscruentus);或在其他地区,那些对不同的其它侵染性物种(黑麦草属(loliumspp.)、奇异虉草(plalarisparadoxa)、虞美人(papaverrhoeas)、新疆白芥(sinapisarvensis)、稗草(echinocloacrusgalliums)、假高粱(echinocloacrusgalliums)、异型莎草(cyperusdifformis))中对酶抑制剂产生抗药性的案例。
此外,在医疗领域中,对药物的抗药性的发展使药物的使用不再有效,这构成了与抗生素疗法有关的最大问题之一(clatworthy等,2007年,自然化学生物学(nat.chem.biol.),3(9):541-548)。尽管为了发现新的分子和研究抗药性机理,投入了巨大的人力和经济资源,但是,抗药性的出现被证明比新药的发现更快。
药物抗药性的问题已成为卫生领域全球范围内的主要问题,因为它会导致由抗生素疗法产生的临床并发症(疾病持续时间的增加、并发症的增加、流行病爆发的可能性),以及处理具有抗生素抗性的细菌的侵染而需要的额外费用(通过使用更多药物,从而延长了住院时间)。越来越多地出现并对公共健康造成危害的并且代表了医院内感染的主要原因之一的抗生素抗药性的案例是,葡萄球菌(staphylococci)、首先是各种金黄色葡萄球菌(staphylococcusaureus)菌株对甲氧苯青霉素(meticilline)的抗药性。这些细菌,通过降低靶标的亲合力的机制,都能表达不与β-内酰胺抗生素相互作用的修饰的蛋白质。欧盟已经面临这个问题,提出了“针对抗菌剂抗药性的社群战略”,其中,建议在医药中谨慎使用抗菌剂药物,以遏制抗药性的发展。
自zamecnik和stephenson首次公开的内容开始,抑制特定基因的表达的可能性引起了极大的兴趣,其中,其公开了使用基于反义寡核苷酸(aso)的组合物能抑制病毒蛋白的转译(zemecnik和stephenson,1978年,proc.natl.acad.ski.75:285-288)。随着这一发现,在关于打击寄生性、致病性和侵染性的媒介体的技术的最新实验进一步研究了各种rna和/或dna小尺寸分子(大约几十个不同的碱基)的使用,选择性地将其结合到显示互补序列(例如,mrna、微小rna或线粒体rna)的靶标,从而抑制其转译,进而实现控制靶标物种的活性。
这些现代化方法找到了在针对寄生性物种(如wo2005/049841a1)、侵染性植物(如wo20111257a1)和抗菌、肿瘤治疗方面的应用。
目前使用的方法有多种,其中包括rna的修饰,从而控制特定rna信使的水平,进而限制编码基因的表达。这样的方法的特征在于施用,通过适当构建rna或dna序列,该rna或dna序列瞄准特定的靶标,例如它的一个基因或一部分基因。与这样的处理方法的使用相关的问题包括:需要表征特定的序列作为处理靶标。还有,获得在靶标器官结构的有效最小浓度存在难度。此外,反义技术的近期历史显示,尽管结合特定rna的aso的识别现在来说是比较简单的,但是,在其显示有效应用潜能的组合物中用于特定基因表达抑制的剂型仍有问题。最近进行解决前述问题的各种研究,提出了更稳定和更有效的结构和组合物(例如,wo2011/0315202a1)。在作用位点的特异性方面,该类型的技术显示提高的选择性是已知的。迄今为止,对aso在除草剂和灭害剂处理中的使用出现抗药性仍没有报道过,因为,它们的应用仍没有达到商业化以及随后的产品的扩散的阶段。但是,考虑到aso的特定作用的特异性,比较接近实际情况的假设是,由于对侵染性、寄生性和/或致病性种群的大量的使用和选择压力,aso的应用将导致在处理种群时出现抗药性,从而使该处理无效,在同样的方式下,它过去发生于以作用特异性提高为特点的其他类型药物上。
在生物反应器和光生物反应器的批量培养领域,进而在密闭的控制系统中,在微生物的批量培养中还没有解决的主要问题之一是出现所述生物体的生长曲线的饱和现象(静止阶段),随之而来的是不可能在单位体积超过一定浓度。这样的微生物(细菌、光合细菌、酵母、微藻类和蓝藻)的典型生长曲线的特征在于起始的潜伏阶段、连续的指数阶段、以及发生生长曲线的饱和的稳态阶段,随后有时还有细胞死亡阶段。
在反应器和批量培养系统中的大规模培养过程中,不可能超过微生物所能达到的单位体积最大浓度阈值,这明显地限制了这样的生物体的发展潜力,不允许从出稳定阶段超出的产量的显著增量。
提出了各种推动方法,以降低微生物在饱和阶段细胞生长速率,主要都涉及培养基中营养元素的限制,或者对于光合生物体来说,穿透性辐射培养的自动限制(自遮蔽现象)。各种实验研究表明,在缺少明显的限制因素时也出现生长曲线饱和现象,这种现象与培养基中抑制物质的积累有关。
根据上面的披露,显然需要新方法来实现有害生物的抑制或在微生物生成过程中产量的增加,从而克服现有方法的缺点。
在对于土壤中有机物质的循环的生态研究过程出现了重要的观测结果,本发明就是源自该观测结果。在这些研究中,可以观察到,由土壤中积累的有机材料的分解而释放的dna,被证明能抑制产生所述有机材料的植物物种,同时,所述dna不抑制其它物种,相反地,所述dna对其它物种来说是营养来源。
在森林生态系统中,这种特殊的胞外dna的抑制作用仅对具有同源dna的植物物种有效,这代表了在各种物种中自然的共存调节机制,并且有利于植物群落的生物多样性。事实上,当一个单一物种因为某种原因而处在适于增加自己相对于其他物种的优势度的情况中,此时,必要地,其会积累大量的自身有机物质的残渣,它的分解以及随之而来的dna的释放将对同一物种产生抑制作用,从而降低了竞争力,进而降低了优势度。基于同样的现象,在农业中即是所谓的“土壤疲劳”(或称为“土壤病”),就是说,重复单一作物的种植造成的随时间的生产率损失,不是因为营养性质的问题,而是因为相同作物的植物残渣的积累。观察到在有机物质的循环过程中,由于自身dna的积累而造成的物种特异性抑制,这解释了上述生产率损失的现象,并且所述发现从未在科学文献中报道过。
在这些研究的基础上,获得了第二个重要的结果,包括以下观察:微生物种群(例如细菌,藻类和真菌)在生长过程中产生和分泌大小不同的片段的自身dna分子,通过在种群生长基质中的积累,其对所述种群的生长施加抑制作用。
众所周知,微生物群体开始以指数方式生长,然后进入所谓的静止阶段,随后,可能的话,进入死亡阶段。这样的微生物种群的各种生长限制因素是已知的,其中,最根本是营养限制和/或各种性质的有毒物质的积累。在文献中,已经报道了各种生物体生产和分泌胞外dna(peters和pretorius,2011年,临床化学学报(chemicalclinicacta),412:806-81);但到目前为止,还未报道过在微生物种群的生长过程中的胞外dna的释放对该种群的生长产生抑制作用,该抑制作用的方式与观察到的土壤中自身dna的积累对植物产生的影响完全等同。本发明的观察基于揭示一个一般的生物学规律,根据该规律,生物体释放的自身dna被证明对该生物体的生长和同一物种种群生物体的生长的调节作用。这一结果引起了在农业医药领域的新的可观的应用前景。具体地讲,通过暴露于总的随机的片段化dna而有可能控制任何有害的物种,以此复制在有机物质的分解的自然循环中或者在微生物种群生长过程中的细胞外分泌的现象中所观察到的情形。因此,可以推测如下的组合物的可能的用途,该组合物的有效成分包括想要控制的物种的总的随机的片段化dna。
下面显示的测试将证明这样的方法对任何被处理的物种都是有效的。那些被报道的都是容易复制的实验,并不需要对想要处理的物种的基因组在特定靶标基因的测序和/或检测方面有任何先有的知识。为了准备抑制组合物,仅有从靶标物种的有机材料得到的总dna的提取物(例如,从植物的叶子、真菌菌丝体、微生物生物质中)以及该提取的dna的连续随机片段就足够了。本发明设置的程序模拟了有机物质在土壤中自然分解的过程,产生了各种尺寸的包括从几十至几千个碱基的多核苷酸的混合物。这样的片段代表了要控制的物种的总dna,在不同类型的反义寡核苷酸的情况中,其对具有同源dna的生物体产生高度选择性作用,而对其产生的单一靶标基因不起作用。这方面可能构成了明显的优势,因为它消除了在被处理种群中出现抗药性的可能性,因为据推测没有生物体可能同时发展对所有自身功能抑制的抗性。实际上,根据科学文献已知,在寄生性、致病性和侵染性的种群中,抗药性现象的出现速度与负责此抗药性的基因的数量密切相关。更准确地说,用于抗药性出现的必要基因的数量越低,在要控制的种群内主动选择该抗性的速度越快(prather等,2000年,除草剂的抗药性:确定与管理策略(herbicideresistance:definitionsandmanagementstrategies),加利福尼亚大学农业与自然资源部(universityofcalifornia,divisionofagriculturesandnaturalresources);oakland–brent和hollomon,2007年,作物病原体的杀真菌剂的抗药性:如何管理它?(fungicideresistanceincroppathogens:howcanitbemanaged?),国际作物生命协会(croplifeinternational),布鲁塞尔(brussel))。
技术实现要素:
因此,本发明涉及组合物在抑制有寄生性、致病性和/或侵染性的物种中的用途,其中,与要抑制的物种的同一基因组相同或相似的总的自身dna的随机片段如活性成分那样被使用。此外,本发明涉及包括总dna的随机片段的组合物。
另外,本发明的发明人开发了一种方法和相关的系统,用于在生物反应器或光生物中微生物的生产/生长、或者在用于水培的系统中植物的生产/生长,其在土壤外或在土壤中的培养基上、通过分离微生物或植物的核酸来进行。
下面,在本专利申请中,术语“饱和度”是在微生物生长的过程中形成的阶段,当在培养基中的核酸或其片段的浓度引起微生物生长过程中的稳态阶段时,其生长曲线饱和。同样地,术语“耗尽培养基”(exhaustmedium)指的是培养基中核酸及其片段的浓度达到引起在微生物的生长过程中的饱和水平的浓度。
至于术语“回收”是指适合于从耗尽培养基中清除核酸和其片段的操作。“再生培养基”是指在耗尽培养基回收后获得的培养基。
具体实施方式
本发明基于的实验证据为,生物体暴露与总自身dna的提取物导致对该生物体的功能的抑制作用,从而限制生长或在细胞水平上以基础方式干扰其生理作用。具体来说,不选择特异性片段或使用特定构建体而被提取和施用的有寄生性、致病性和/或侵染性的生物体的总dna,可被用作相同生物体或具有相同或相似基因组的其它生物体的抑制产品。当有寄生性、致病性和/或侵染性的物种在它的生长培养基或在系统层面上与随机片段化的总自身dna接触,则发挥该抑制作用。一旦被吸收,这样的dna对生长产生限制作用,并对生物体的各种功能产生综合抑制作用,而这种作用在其他情况中在以“基因沉默”或“rna干扰”作用为目的而应用特定核酸序列时发生。吸附和抑制的活性取决于系统所暴露的核酸的浓度和片段化水平。更大的作用被指出取决于所用分子的片段化等级(例如在超声处理或随后的自然分解以后)。使用各种平均长度的dna片段进行了实验,并且证明了对于具有约200个碱基的较高长度频率的片段混合物,抑制作用更有效。
具体而言,根据示例的实验从不同物种提取的总dna的随机片段的生产以各种不同的方法实现,以证明片段化技术不影响抑制效果的有效性。所述片段化可以通过自然降解(暴露于环境因素中)、人工降解(通过燃烧、酶化或机械方法)或通过合成手段(随机扩增方法,如dop,即通过对退变寡核苷酸进行pcr反应获得)实现。
本发明的作者对植物、真菌、昆虫、酵母、藻类和原生生物进行了研究。这样的实验的结果清楚地表明了所描述的对任何被处理物种的抑制效果是普遍存在的。
对于植物来说,在通过简单的提取和随机片段化获得的低浓度的物种总dna的水溶液中,与同物种的根部相接触时,迅速地产生根部坏死、叶子枯萎、分生组织的活性中断,并且当与种子接触时,抑制发芽能力。与此相反,物种的根和/或种子暴露于不同物种的随机片段化总dna的溶液中不会产生明显的抑制,当暴露于高浓度中也是一样,也就是说,呈现出高度物种特异性的作用。因此,显而易见的是,随机片段化总dna在生长环境中的存在可以对具有相同或相似的基因组的生物体实现选择性抑制功能,而不影响其他物种。当总dna被人工片段化并被插入到生物系统的生长基质中时以及当dna被所述生物系统在其生长过程中产生和分泌时,都有可能都发现该抑制作用。
因此,通过用含有对该物种有特异性的dna的制剂进行处理,有可能使得作用只针对对有寄生性、致病性和/或侵染性的物种,这具有极大的优势:该分子将只对靶标物种显示抑制作用,但对要保护的物种或其它物种不起作用,在可能发生的重复暴露的情况下也不起作用。
本发明的发明人观察到的相同的原理可以有利地采用,以增加在生物反应器和光生物反应器、水培罐以及土壤外培养系统中的生产微生物的方法的产量。事实上,本发明的发明人,通过使用用于生长藻类、酵母和细菌的生物反应器进行的实验测试,在生长阶段观察到属于同一被生产微生物的各种大小的dna片段(大约50-800bp)的上清液积累,其浓度不断增加。据观察,这种积累出现在生长阶段且浓度增加,其导致了当在生物反应器中到达稳态阶段和后续的死亡阶段时,对所述培养物的生长具有抑制作用(图11a和11b)。
该实验测试表明了存在物种特异性的自抑制作用,因为给定物种的培养物显示仅被相同物种的培养物的上清液所抑制。此外,可以观察到,从培养基中清除dna适用于清除抑制作用,使生长阶段被恢复,并在生物反应器和光生物反应器中到达更高的细胞密度(图11c)。类似地,在植物的水培培养罐中,在没有完全替换循环营养溶液的情况下出现产量的降低时,可以观察到培养物种的dna在培养基的积累。此外,在这种情况下,从生长培养基清除积累的dna使得生产能力被恢复。
考虑到这些实验结果,因此,为了优化生产,有可能开发在细胞培养物(生物反应器和光生物反应器)、水培罐以及在土壤外培养系统中清除dna的技术系统。
因此,本发明的特定目的是一种包括总dna片段的混合物或由总dna片段的混合物组成的组合物的用途,其中,所述总dna被随机片段化,用于针对至少一种对植物或环境有致病性、寄生性或侵染性的物种进行预防和处理,其中,所述总dna是所述预防和处理所针对的有致病性、寄生性或侵染性的物种和/或至少一种系统发育上类似的物种的总dna。如上述所述,根据本发明的片段混合物是通过总dna的片段化获得的,总dna是从所述预防和处理所针对的有致病性、寄生性或侵染性的物种中提取的,不进行片段的任何选择,即使用从核酸中获得的全部片段。因此,根据本发明,使用随机片段化总dna,该随机片段化总dna不进行特定的和/或特定大小的序列的任何选择。所述总dna的片段可通过对提取的总dna的随机片段化或通过从总dna开始的随机片段合成而获得。本发明的技术与现有技术“基因沉默”和“rna干扰”相比,实质差异在于:在本领域的当前状态中,必须主动对要使用核酸的特定区域进行选择;而在本发明的技术的情况下,不需要先有的知识也不需要对要使用的特定序列进行选择。所述总dna可以从所述至少一种有致病性、寄生性或侵染性的物种中提取或人工合成。所述核酸可以通过化学和/或物理类过程被扩增和/或片段化,并且获得的片段可能可以进一步扩增。总dna的随机片段的生产方法包括,例如,化学、生物化学和分子方法,超声技术,热处理和高温分解过程。例如,在从拟南芥(arabidopsisthaliana)的叶片组织中提取总dna后,将来自相同物种的种子暴露于这样的包括未片段化形式的dna的提取物中,不显示明显的抑制作用。与之不同地,用超声处理提取液,其使用浸渍超声波仪在80%的功率下进行3或4个周期,持续3分钟,生成各种大小的随机片段混合物,增加了对拟南芥种子的发芽的抑制活性,并且使所达到的片段化程度的函数。从拟南芥中提取的总dna的随机片段的相同混合物对其它被处理的物种未显示抑制作用,如番茄(lycopersiconaesculentum)和燕麦(avenasative)(在此实验中用作对照生物)。当总dna的提取物通过上面列举的其它化学-物理方法被随机片段化时,可以观察到类似的结果。最后,通过将拟南芥种子暴露于总dna随机片段混合物中可获得相同结果,其中,所述片段通过使用dop进行总dna的扩增获得。该组合物可以用作生物灭杀剂、除草剂、杀真菌剂、杀昆虫剂、杀螨剂、杀线虫剂、抗原生生物剂、除藻剂、杀细菌剂。该组合物可以通过表面接触、亲细胞性施药、通过例如注射、摄入或吸入方式的全身施药、或吸附来施用到至少一种有致病性、寄生性或侵染性的物种。该组合物可以配制成干燥或液体处理剂的形式,可选自包括以下形式的组:分散剂(例如气雾剂)、悬浮液、可湿性或可溶性粉剂、在水或其它溶剂中的乳剂、可分散的颗粒、微胶囊的悬浮液、乳油、流体糊剂、粗滴乳状液、油分散剂、饵剂。根据一个特定的实施方案,核酸随机片段可构建在载体中。根据本发明的组合物可进一步包括灭害剂,其选自包括由以下所组成的组:真菌剂、杀昆虫剂、杀线虫剂、杀螨药、抗原生生物剂、杀细菌剂、除藻剂。
本发明进一步涉及一种方法,用于针对至少一种对植物或环境有致病性、寄生性或侵染性的物种进行预防或处理,所述方法包括或由下列步骤组成:a)提取所述预防和处理所针对的至少一种有致病性、寄生性或侵染性的物种的总dna;b)产生所述总dna的随机片段,从而获得dna随机片段的混合物;和c)使所述随机片段的混合物与所述有致病性、寄生性或侵染性的物种接触。
一种包括总dna片段的混合物或由总dna片段的混合物组成的组合物,用于针对至少一种对人类或动物有致病性、寄生性或侵染性的物种进行预防和处理,其中,所述总dna是所述预防和处理所针对的至少一种有致病性、寄生性或侵染性的物种的总dna,这是本发明的另一个目的。所述总dna的随机片段可通过总dna的降解而获得,总dna的降解可通过化学和/或物理方法、或通过所述总dna的随机片段的合成获得。根据本发明的组合物可以配制为干燥或液体处理剂或气雾分散剂,有以下形式:悬浮液、可湿性或可溶性粉剂、在水或其它溶剂中的乳剂、可分散的颗粒、微胶囊的悬浮液、乳油、流体糊剂和粗滴乳状液、油分散剂、乳脂、胶囊剂和片剂。施药途径包括接触或通过注射、吸收、摄入和吸入进行的表面、全身或亲细胞性处理,还包括其他任何对于最后应用有用的施药途径。根据本发明的组合物可包括与其它化合物的混合物,其它化合物作为增粘剂、润湿剂、助悬剂、赋形剂、修正剂和溶剂、表面活性剂,以及对最后应用有用的其它物质。最后,根据本发明的组合物可在不抵制而有可能起协同作用的媒介物的制剂和混合物中包括其它药物化合物。在这种情况下,化合物的组合通常将对应于约0%至100%的重量比。
本发明还有一个特定目的是提供一种在生物反应器或光生物反应器或土壤中和土壤外的植物培养系统中以高产率生产微生物的方法,其特征在于:将来自通过所述方法生产的相同生物体的核酸从培养基中清除,并可将被清除了所述核酸的培养基再次用于所述方法。
根据本发明的方法,使用分离技术将核酸从培养基或生长基质中清除,所述分离技术选自由以下所组成的组:外部清除单元,优选由外部容器、以及用于生成电场、磁场或电磁场的系统、以及用于从前述系统中提取浓缩核酸的至少一个管道和至少一个输送装置组成;集成清除单元,优选由生成电场、磁场或电磁场的系统以及用于从前述系统中提取浓缩核酸的至少一个管道和至少一个输送装置组成,所述集成清除单元整体集成在相同生物反应器内;可在原位和外部的单元应用的技术,优选离心技术;过滤技术;用dna酶和用于核酸的其它降解酶进行的处理;热处理;酸化处理。
优选地,根据本发明的方法,利用至少一个外部的或生物反应器或光生物反应器集成的清除单元或用于水培的系统将所述核酸从培养基中清除,使用选自由以下所组成的组的技术进行清除:施加静态电场、施加静态磁场、施加动态磁场和/或电场、离心、过滤、dna酶处理、热处理、酸化处理。
本发明的额外方面在于,提供一种用于生产微生物的系统,其包括至少一个生物反应器或光生物反应器或用于水培的系统,以及位于所述至少一个生物反应器或光生物反应器或用于水培的系统之外的至少一个核酸清除单元,所述核酸清除单元通过至少一个第一管道和至少一个第一输送装置连接到所述至少一个生物反应器或光生物反应器或用于水培的系统,以回收包含核酸的培养基;所述核酸清除单元通过至少一个第二管道和至少一个第二输送装置连接到至少一个生物反应器或光生物反应器或用于水培的系统,以将被清除了核酸的培养基再次引入到所述至少一个生物反应器或光生物反应器或用于水培的系统中;所述系统还进一步还包括至少一个第三输送装置,用于回收已被分离的核酸。优选地,所述管道可以是管道。所述输送装置可以优选是泵。
培养基的回收可通过如下两者来实现:连续进行用于在生长阶段从培养基中清除dna的处理;当细胞生长饱和阶段出现时,实施耗尽培养基的处理。
培养基的回收可以直接在生物反应器或光生物反应器中直接原位进行,其是通过利用对在细胞培养物的上清液中发生的dna的降解有效的酶或其它化合物进行处理,或者通过施加电场、磁场或电磁场的技术。利用这些技术的回收证明是可行的,考虑到利用核酸分子的极性特征从而可被电场分离;或者利用dna分子的磁化技术:通过将其与磁性分子结合并对其施加磁场以将其分离。或者,该回收的实施可以利用放置在生物反应器(图12a)或水培(图13a)之外、具有再生培养基再循环的清除系统、或者生物反应器(图12b)或水培(图13b)集成式的清除系统。
从培养基中清除dna的技术可以是:施加静态电场的技术(图14)、施加静态磁场的技术(图15)、施加动态磁场和/或电场的技术、离心、过滤技术、用dna酶进行的处理、热处理、酸化处理。
在本发明的优选实施方案(图12a)中,所述至少一个生物反应器(1)可以以这样的方式连接到至少一个外部清除单元(4):通过至少一个第一管道(2)和至少一个第一输送装置(3),将耗尽培养基输送进至少一个所述清除单元(4),以回收包含核酸的培养基。在这样的单元(4)中,可利用上述从培养基中清除dna的技术之中的一种来实现回收。在这一点上,分离出的核酸会被所述清除单元(4)收集,优选通过至少一个第三管道(5)和至少一个第三输送装置(6)回收分离出的核酸,而再生的培养基将通过至少一个第二管道(7)和至少一个第二输送装置(8)被再次引入到所述至少一个生物反应器(1)中。
此外,将野外或保护培养的农业灌溉装置与用于清除核酸的系统集成,代表了本发明进一步的应用。图16给出了这种应用的一种可能的示意图,其中,具有包括核酸的溶液的罐被集成到农业灌溉设备上。
与现有技术和产品相比,在农业和医药领域使用用于控制和抑制有寄生性、侵染性和/或致病性的物种的总dna随机片段,结果是具有明显的优势。
第一个优点源自用寄生性、侵染性和/或致病性的物种的总dna随机片段进行处理所带来的绝对选择性,因为所述总dna随机片段对与被处理的物种不同的物种不显示作用。
使用寄生性、侵染性和/或致病性的物种的总dna随机片段不会在同一物种种群中诱发抗药性,因为其作用机理不同于当前市售的或在正在实验的产品(包括rna干扰方法和aso),其是基于用整个基因组对整个转录和蛋白合成系统的多重的、同期的干扰,因而不允许产生特异性抗药性和抗性种群的选择性。
另一个优点源自由核酸的应用引起的毒性的消失,即,作为自然状态下普遍存在的初级代谢产物,其不会导致环境污染。事实上,非特异性核酸片段的施用对被处理物种以外的物种不显示毒性。
本发明进一步对研究新药带来显著的简易化,进而产生了巨大的经济效益。事实上,对有寄生性、致病性和/或侵染性的物种的检测构成了用于药物处理的特定药物的制备的唯一条件。与此相反,与基于如rna干扰或使用反义寡核苷酸的技术的当前对产品的药理研究,不仅需要靶标生物体的全基因序列的知识,还需要基因功能的详细知识,以检测带来抑制活性的作为靶标的可能的特定序列。
另一方面,传统药理研究是基于合成化合物或天然分子的“随机筛选”的研究。这种物质一旦被检测到并测试它们的活性,在大多数的情况下,其特点也是对其它生物体和人类具有普适毒性。实际上,这样的产品的应用由于其毒性而经常与环境污染的考虑相关。
另一个优点源自满足本申请要求的核酸相对容易获得和批量生产。实际上,靶标生物体的总dna的随机片段可通过目前在生物分子领域极其普及的方式和方法获得。该技术可基于从靶标生物体中提取遗传物质或基于合成手段。
此外,dna片段的稳定性可以在不利的环境条件下(例如在土壤中)也能保证经时持久性,因此,在耐久性应用中在环境和经济方面都具有益处。
另一优点源自以下事实:用于所提出的用途的dna能基于目前在生物分子学中已被广泛普及和确定的合成和提取技术以及扩增技术,从天然材料中提取或人工合成获得。可能的生产技术的显著但不是限制性的实例包括细胞组织的溶解,细胞核酸酶的失活和从含生物裂解物的溶液中回收核酸。
根据不同的方法,核酸进一步可从模板分子开始或者从头合成。
所考虑的分子可以被扩增,从而产生与所考虑的模板相同或相似的多个拷贝,例如通过克隆或基于pcr的技术。
pcr突变体是有用的,以产生活性成分,用于确保源自模板样品的核酸片段随机扩增。例如,像随机扩增多态性dna(rapd)的方法非常适合于我们的目标,与传统的pcr不同,不要求起始dna模板序列的知识。
乳化pcr代表了创新方法的另一实施例,适于以非常快速的方式确保从基因组dna样品开始扩增各种dna片段,基因组dna样品进行裂化以获得300-800个核苷酸长度的片段,或者从由其它方法获得的扩增子开始扩增。
另外,至于在生物反应器和光生物反应器以及水培罐中的微生物生产方法,在相同的单位体积细胞浓度和相同的轮廓条件下,耗尽培养基的清除和在“再生”培养基中细胞的再悬浮,允许恢复培养物的生长阶段。
除了由抑制因素的清除引起的产品增加的优点,另一个明显的优点源自耗尽培养基在其再生之后再利用于其它后续的培养的可能性,该可能性在当前生产系统是不可行的。
附图说明
现在将通过说明性但非限制性的方式,特别参考实施例和附图,来说明性地描述本发明,其中:
图1显示了本发明对象的概念图;
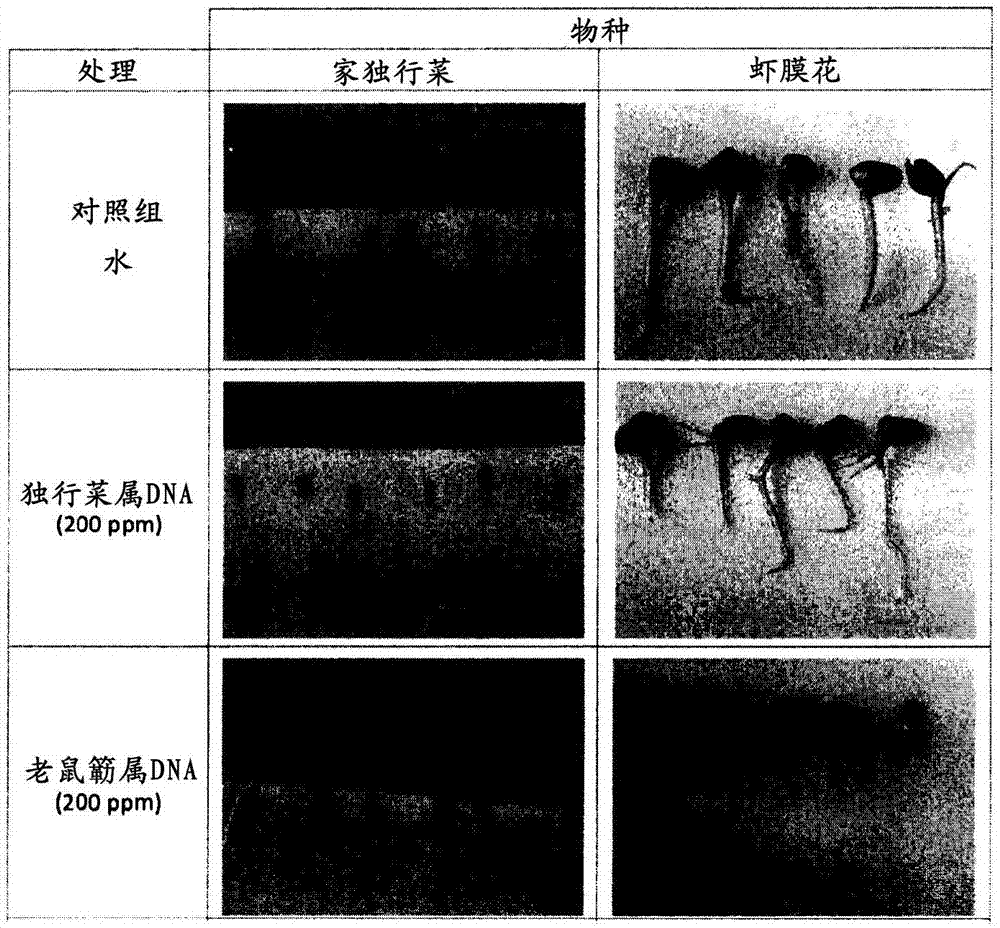
图2显示了暴露于最佳浓度为200ppm的自身dna中对独行菜属(lepidium)和老鼠簕属(acanthus)的种子发芽的抑制;
图3显示了暴露于自身dna中对植物冬青栎(quercusilex)、栎毛竹(quercuspubescens)、洋常春藤(hederaelix)、毛里塔尼亚大戟(ampelodesmamauritanica)、羊茅(festucadrimeja)、蝎子旃那(coronillaemerus)、滨海苜蓿(medicagomarina)、意大利赤杨(alnuscordata)、刺槐(robiniapseudoacacia)、地中海松(pinushalepensis)的抑制;
图4显示了暴露于自身dna和番茄(lycopersiconesculentum)dna中对植物拟南芥(arabidopsisthaliana)的抑制;
图5显示了当黑曲霉菌(aspergillusniger)暴露于自身或其它真菌物种(哈茨木霉菌(trichodermahartianum))的dna中时,黑曲霉菌的发芽和菌丝生长的抑制;
图6显示了暴露于自身dna中对麻蝇(sarcophagacarnaria)昆虫的抑制;
图7显示了在三种不同浓度的同源dna中,或真菌(产黄青霉菌(penicilliumchrysogenum))或细菌(枯草杆菌(bacillussubtilis))提取的异源dna中暴露4周的双翅目麻蝇幼虫中,完全变态与非暴露对照组的百分比;
图8显示了在三种不同浓度的同源dna中暴露24小时和在非暴露对照组中,枯草杆菌细菌培养物中活细胞的数量;
图9显示了在暴露于三种不同浓度的同源dna中,或从其它真菌(黑曲霉菌)、昆虫(麻蝇)或细菌(枯草杆菌)提取的异源dna中的培养物中,哈茨木霉菌的萌芽与非暴露对照组的百分比;
图10显示了在暴露于不同浓度的同源dna中的两种培养物中和非暴露对照组中,斜生栅藻(scenedesmusobliquus)微藻的生长动力学;
图11显示了在两个不同的生物反应器的液态培养基中,胞外dna的积累以及相关的对生长的抑制作用的试验证据;
图12显示:(a)在生物反应器中生产微生物的系统的框图,特征在于用于dna清除的外部单元;(b)在生物反应器中生产微生物的系统的框图,其中,通过集成于微生物反应器的清除单元,实现从培养基中清除dna;其中,1表示生物反应器,2表示用于饱和装置输送的管道,3表示用于饱和装置输送的泵,4表示外部清除单元,5表示用于饱和核酸的管道,6表示用于饱和核酸的泵,7表示用于洗净装置循环的管道,8表示用于洗净装置循环的泵,9表示补给,10表示内部清除单元;
图13显示:(a)水培罐,其特征在于,外部清除dna单元的存在和在水培系统中培养基的再循环;(b)水培罐,其中,通过集成到同一个罐的清除单元,实现了从培养基中清除dna;其中,2表示用于饱和装置输送的管道,3表示用于饱和装置输送的泵,4表示外部清除单元,5表示用于饱和核酸的管道,6表示用于饱和核酸的泵,7表示用于洗净装置循环的管道,8表示用于洗净装置循环的泵,9表示补给,10表示内部清除单元,11表示水培罐;
图14显示了用于图12所示的系统中的外部清除单元的具体框图,其中,核酸的分离通过施加电场来实现;其中,2表示用于饱和装置输送的管道,3表示用于饱和装置输送的泵,4表示外部清除单元,5表示用于饱和核酸的管道,6表示用于饱和核酸的泵,7表示用于洗净装置循环的管道,8表示用于洗净装置循环的泵,12表示场的电子检测,13表示金属板,14表示电场;
图15显示了用于图12所示的系统中的外部清除单元的具体框图,其中,核酸的分离通过施加磁场来实现;其中,2表示用于饱和装置输送的管道,3表示用于饱和装置输送的泵,4表示外部清除单元,5表示用于饱和核酸的管道,6表示用于饱和核酸的泵,7表示用于洗净装置循环的管道,8表示用于洗净装置循环的泵,15表示磁场,16表示永久磁铁/电磁铁;
图16显示了植物培养物的框图,其中,通过与土培系统的集成,用核酸酶处理培养基;其中,17表示水泵,18表示肥料溶液罐,19表示核酸酶溶液罐,20表示主流管道,21表示肥料溶液喷射路线,22表示核酸酶喷射路线,23表示头管,24表示两翼滴灌或线状滴灌,25表示滴灌或微滴灌,26表示灌溉区域。
在报道的实验的所有例子中,用于不同处理的核酸组合物都是根据图1所概述的方法制备的。尤其是,用标准方法从有机材料(叶子、真菌菌丝体、微生物生物质)中提取的总dna被浸渍超声波仪以最大功率持续三分钟的至少三个循环超声处理,直到获得随机片段的组合物的产品,其中随机片段落到50-1000bp的大小范围内。片段级别的验证通过利用琼脂糖或聚丙烯酰胺凝胶电泳和染色技术、sybrsafe型和uv可视化的标准过程实现。
实施例1:暴露于自身dna对植物虾膜花(acanthusmollis)和家独行菜(lepidiumsativum)的抑制
第一个实验由植物虾膜花和家独行菜完成,后一个物种被选中的原因是它对毒素特别敏感。老鼠簕属(acanthus)虾膜花和豆瓣菜(watercress)家独行菜的dna通过从这两个物种的叶子中直接提取并存储在蒸馏水中获得。皮氏培养皿(直径9厘米)中的10粒预先灭菌的虾膜花和家独行菜种子被依次放置在无菌滤纸上。每个物种的种子分别被两个物种的2、20、200ppm的浓度的dna处理,而无菌水被加入到对照组中。在24℃经过7天的培育,两个物种的发芽和根系总长通过观察和测量根部被量化。每个处理都重复三次。
用提取自这两个物种的植物的dna的处理这两个物种的种子,分别应用,其允许估计dna对根部生长的影响和最佳活性浓度。图2所报告的实验结果显示,家独行菜的种子和虾膜花的种子在发芽过程中,都被暴露于最佳浓度200ppm的自身dna中所抑制。与此相反,种子暴露于其它物种的dna中,对其种子的发芽并不显示明显的作用。
实施例2:暴露于自身dna中对植物冬青栎、栎毛竹、洋常春藤、毛里塔尼亚大戟、羊茅、蝎子旃那、滨海苜蓿、意大利赤杨、刺槐、地中海松的抑制。
植物冬青栎(quercusilex)、栎毛竹(quercuspubescens)、洋常春藤(hederaelix)、毛里塔尼亚大戟(ampelodesmamauritanica)、羊茅(festucadrimeja)、蝎子旃那(coronillaemerus)、滨海苜蓿(medicagomarina)、意大利赤杨(alnuscordata)、刺槐(robiniapseudoacacia)、地中海松(pinushalepensis)的
第二个实验涉及10种自然环境植物物种的发芽和根系生长的分析。表面灭菌的植物冬青栎、栎毛竹、洋常春藤、毛里塔尼亚大戟、羊茅、蝎子旃那、滨海苜蓿、意大利赤杨、刺槐、地中海松的种子分别被浓度为500ppm的所有物种的dna处理。随即,将每个物种的10粒种子放置在皮氏培养皿(直径9厘米)中的无菌滤纸上。浓度为500ppm的不同dna被加到培养皿中,而只有无菌水被加到对照组中。在24℃经过7天的培育,物种种子的发芽和根系总长通过观察和测量根部被量化。每个处理都重复三次。图3所报告的结果(上面报告的不同物种完成的实验的平均值)显示,暴露于自身dna产生了对发芽的抑制作用,在存在异源dna时无抑制作用。
实施例3:暴露于自身dna中对植物拟南芥、番茄、家独行菜和兵豆(lensesculentum)的抑制
第三个实验涉及再次通过发芽和根系生长实验,对提取自同一物种的核酸对各种植物物种的毒性的评价。拟南芥、番茄、家独行菜和兵豆的dna通过从各种植物中直接提取并存储在蒸馏水中获得。皮氏培养皿(直径9厘米)中的10粒预先灭菌的每种物种的种子被依次放置在无菌滤纸上。每种植物的种子分别被四个物种的2、20、200ppm的浓度的dna处理,而无菌水被加入到对照组中。该实验在处在受控条件下完全无菌的生长室中进行。在24℃经过7天的培育,四个物种的发芽和根系总长通过观察和测量根部被量化。每个处理都重复三次。这四个物种显示了类似的行为,在存在自身dna时有明显的抑制作用,在存在其它物种dna时无抑制作用。该抑制作用被证明与dna浓度正相关。为了示例性的目的,报告了分别存在自身dna和番茄dna时拟南芥的情况。当拟南芥种子暴露在通过利用pcr技术对相同dna片段的扩增获得的同一物种的dna中时,可以在拟南芥中观察到类似结果。
实施例4:暴露于自身dna中对黑曲霉菌和哈茨木霉菌的抑制
第四个实验在真菌黑曲霉菌上完成,以评估自身dna和从另一种真菌即哈茨木霉菌分离的dna对于细胞生长的影响。黑曲霉菌孢子通过实验室纯培养在琼脂处理的培养基(pda,马铃薯葡萄糖琼脂)上获得。孢子在无菌条件下被取出,并被稀释到1×106孢子/ml的浓度。发芽实验在96孔elisa板中的液体培养基(pdb10%)上完成。对比处理由作为异源的哈茨木霉菌的dna完成,而对照组不被处理。提取自两个物种的dna以100和1000ppm的浓度被应用。随即,分别具有两种不同浓度的所述两种dna,连同10μl的液体营养培养基(pdb,马铃薯葡萄糖培养基)、无菌水和黑曲霉菌孢子被加入到总容量为100μl的各孔中。在24℃经过20小时的培育,孢子的发芽和芽管的长度被分光读数仪和光学显微镜量化。图5所报告的结果显示,只有黑曲霉菌暴露于自身dna中时,才会对该真菌的孢子的发芽和菌丝的生长产生明显的抑制效果。
实施例5:暴露于自身dna对麻蝇昆虫的抑制
第五个实验在麻蝇昆虫上完成,以评估自身dna对生命周期的影响。双翅目麻蝇幼虫是在10℃温度的实验室纯培养中以肉末喂养使其生长。dna毒性试验在方形塑料、皿(尺寸12×12厘米,高2厘米)中完成。对比处理由作为异源dna的枯草杆菌和家独行菜的dna完成。作为对照组,仅使用肉末而不附加其他处理。从双翅目和其它两个物种提取的dna以2、20和200ppm的浓度被添加到肉末中,用混合器搅拌。随即,不同浓度的dna被添加到每个培养皿中,与1g肉末搅拌混合。该培养皿在10℃温度在黑暗中温育21天。蛹的发育、生存和形成所需要的时间在21天的孵化期中,每三天进行监控。对照条件下的幼虫,以及用那些异源dna处理过的幼虫,都显示了规律的生命周期。反之,在自身dna中的暴露抑制了生命周期,造成幼虫与处理浓度成比例地死亡。图6和图7报告了上述结果。
实施例6:暴露于自dna中对微生物枯草杆菌的抑制
为了证明核酸可能可以作为抗生素使用,对用自身dna以不同浓度处理过的枯草杆菌进行毒性评估。该试验使用注入了10μl枯草杆菌预培养物的4mllb培养基(luriabroth)作为生长培养基来进行。该处理包括培养物的准备,其中,存在最终浓度为4、40和400ppm的枯草杆菌dna。将培养物在搅拌下35℃温度温育24小时,重复处理3次。在温育24小时后,从每个试管中取出0.5毫升并连续稀释到lb培养基中,涂覆在皮氏培养皿中100微升琼脂处理的lb培养基。培养皿在28℃温度温育,直到菌落出现(cfu-菌落形成单元)。图8所报告的结果显示,作为对处理的响应,发生cfu依赖于浓度的明显减少。
实施例7:暴露于自dna中对真菌哈茨木霉菌的抑制
为了证明核酸可能可以作为杀细菌剂使用和具有特异性作用,对真菌哈茨木霉菌的孢子的发芽进行试验。哈茨木霉菌的孢子通过实验室纯培养在琼脂处理的培养基(pda,马铃薯葡萄糖琼脂)上获得。孢子在无菌条件下被取出,并被稀释到1×106孢子/ml的浓度。发芽试验在96孔elisa板中的液体培养基(pdb10%)完成。该处理由同源或异源dna完成,同源或异源dna提取自相同物种的哈茨木霉菌、或不同物种的真菌(黑曲霉菌)、昆虫(麻蝇)或细菌(枯草杆菌)。提取自不同物种的dna在8、80和800ppm的浓度被应用。随即,分别具有不同浓度的dna,连同10μl的液体营养培养基(pdb,马铃薯葡萄糖培养基)、无菌水和黑曲霉菌孢子被加入到总容量为100μl的各孔中。经过在24℃的20小时的培育后,孢子的发芽和芽管的长度通过分光读数仪和光学显微镜被量化。
图9所报告的结果显示,只有哈茨木霉菌的dna才能对该物种的孢子的发芽产生明显的依赖于浓度的抑制作用。反之,各种物种dna的处理对发芽显示刺激作用(与不被暴露的对照组相比,百分比值高于100%)。
实施例8:暴露于自dna中对斜生栅藻微藻的抑制
为了证明dna可能可以作为除藻剂使用,绿藻即斜生栅藻在最佳对照条件下和存在自身dna的情况下的生长测试在培养基(chu#10)中被完成。该处理在两个不同的浓度(50和500ppm)重复两次完成。图10显示了藻类的生长动力学,还显示了与不被暴露的对照组相比,同源dna的依赖于浓度的明显的抑制作用。
实施例9:暴露于自dna中对原生生物即黄色粘液菌(physariumpolycephalum)的抑制
为了证明dna可能可以作为抗原生生物产品使用,在原生生物黄色粘液菌上完成了试验。作为试验材料,使用“卡罗莱纳生物供应(carolinabiologicalsupply)”生产的培养试剂盒。培养从具有水-琼脂的皮氏培养皿上开始,以利于生物体的移动。燕麦片(oatflake)被作为营养物质使用。第一批次培养在20个培养皿上完成,15天后收集产生的原生生物生物质,并被用于利用quiagen试剂盒提取dna。后续试验包括:准备三个加入了水琼脂培养基的皮氏培养皿,将两小部分燕麦片添加到培养皿中,一个用5ml蒸馏水润湿用于对照组,另一个添加有5ml水且添加有浓度为200ppm的原生生物dna。试验另外再重复两次,其具有如下变化,所加的dna来自细菌(枯草杆菌)和昆虫(麻蝇)。试验结果显示,在被同一原生生物dna处理过的培养基上黄色粘液菌的没有生长;而在对照条件下或存在异源dna时,生物体没有显示出生长差异。
实施例10:酵母、细菌和藻类在生物反应器和光生物反应器中生产过程的研究
考虑到上述报道论证了关于当暴露于自身dna中时对不同物种的抑制作用,检测分析使用高密度细胞培养物在生物反应器和光生物反应器的生长培养基中的胞外dna上完成,此时,出现生长条件放缓的情况,即使是在最佳营养培养基存在的情况。这项研究涉及在生物反应器中在指数增长阶段、减缓和稳态阶段的不同培养物的液体上清液取样。该分析涉及酵母类的酵母酿酒(saccharomycescerevisiae)、细菌类的枯草杆菌和微藻类的三角褐指藻(phaeodactylumtricornutum)、斜生栅藻的培养物。通过在3000rpm下15分钟的两个离心周期获得的细胞培养上清液的样品被分析,以分离可能的细胞残留物,然后,在用syber-safe处理后进行电泳,并进行荧光评估。图11显示了这些分析的一部分的结果,从中可以明显看出胞外dna在生物反应器中的液体培养基中的累积。这种积累显然与不同细胞培养物的生长下降有关,并与达到稳态阶段有关(图11a、11b)。图11c清楚地显示,通过化学-物理方法,胞外dna从培养基中的清除,以及随后将再生培养基引入到反应器中,导致抑制作用的消除和随之细胞培养物生长的恢复。
- 还没有人留言评论。精彩留言会获得点赞!