突变型AlkB基因的制作方法
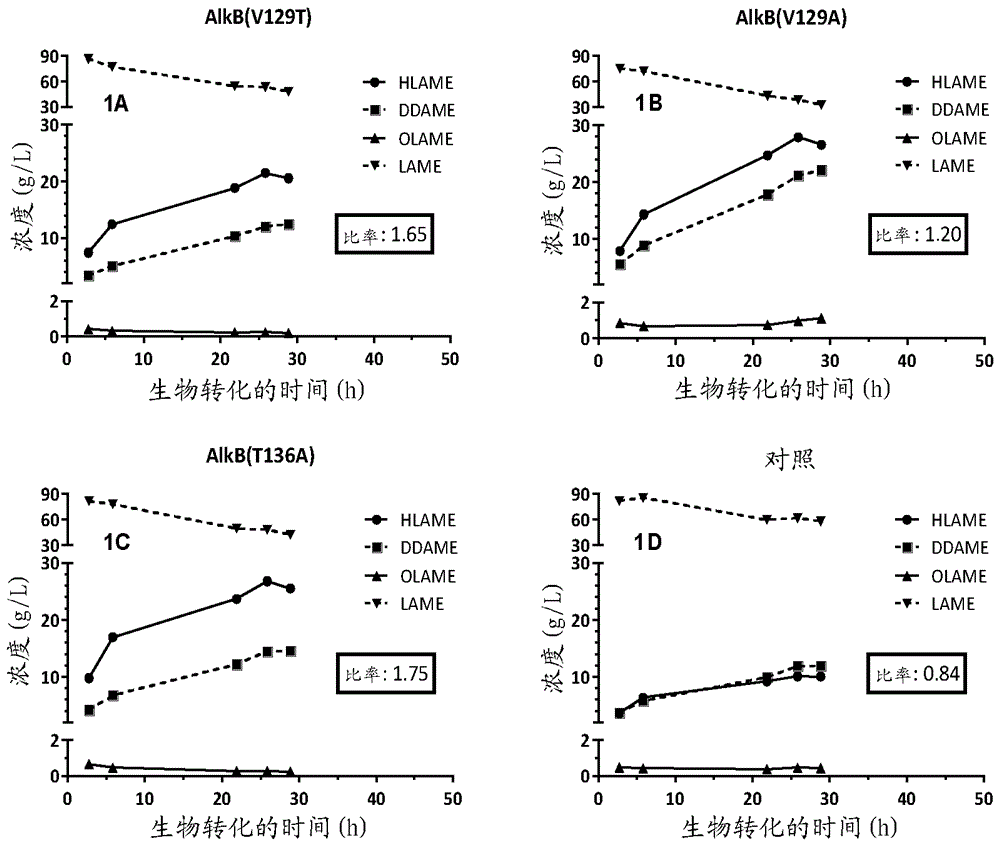
本发明涉及由至少一种c6-c14羧酸产生至少一种ω-羟基羧酸和/或其酯的生物技术手段。特别地,生物技术手段可以使用包含至少一个特异性点突变的突变型alkb基因,所述突变能够特异性地产生ω-羟基羧酸和/或其酯。发明背景ω-羟基化(ω-羟基)羧酸衍生物作为工业试剂的组分具有许多商业用途。可用于工业领域中的ω-羟基羧酸衍生物的一些实例包括ω-羟基羧酸;ω-羟基羧酸甲酯;ω-氧代羧酸;ω-氨基羧酸;ω-氨基羧酸甲酯;α-,ω-二酸(α,ω-二酸);ω-羧基羧酸甲酯(ω-羧基羧酸甲酯);α-,ω-二酯(α,ω-二酯);α-,ω-二醇(α,ω-二醇);等。这些化合物也可用作产生其它各种化合物的前体。例如,这些前体可用于产生α,ω-二羧酸,且α,ω-双官能分子是聚合物树脂、金属工作液、粘合剂、腐蚀抑制剂、电容器电解质、二酯合成润滑剂、纤维、粉末涂覆固化剂、增塑剂、聚酯涂料、环氧树脂、聚酰胺树脂、香料、芳香剂、表面活性剂、去垢剂、添加剂等的工业应用中特别重要的化学品。通常,ω-羟基羧酸衍生物主要由石油基材料或由石蜡和羧酸至ω-羟基羧酸和/或其衍生物的生物转化来产生。用于产生这些化合物的化学方法需要使用危险试剂。本领域中使用的方法也是能量密集的并且对环境造成很大的损害。替代性产生的发酵途径,尽管被认为是绿色工艺,但仍然过于昂贵,并且在可制备的产品的类型和数量方面受到限制。因此,本领域需要改进的产生ω-羟基羧酸及其衍生物的绿色方法。在产生ω-氨基月桂酸甲酯的一个实例中,前体可以是月桂酸甲酯(lame),其中中间体可以是ω-羟基月桂酸甲酯(hlame)。单加氧酶alkb是已知催化由lame形成hlame的反应的酶。alkb酶还能够催化hlame氧化为ω-氧代月桂酸甲酯(olame)。因此,单加氧酶alkb能够催化两个氧化步骤,这导致由lame经由hlame形成olame。然而,alkb酶还能够催化olame转化为十二烷二酸单甲酯(ddame),这导致olame的产物产率降低。因此,本领域需要改进的生物技术方法,其增加来自用作底物的相应的羧酸和/或其酯的特定ω-羟基化羧酸和/或其酯的产率,同时减少不必要副产物的产生。本领域还需要有效地增加所需产物诸如hlame和/或olame的产率。发明描述本发明试图通过提供相对于野生型酶包含至少一个点突变的突变型alkb酶来解决上述问题。突变型酶可以在至少一个细胞中转录以产生表达突变型alkb酶的微生物细胞。表达突变型alkb酶的该细胞能够由用作底物的相应羧酸和/或其酯产生至少一种ω-羟基化羧酸和/或其酯。特别地,羧酸和/或其酯可选自c6-c14羧酸和/或其酯。特别地,相对于表达野生型alkb酶的细胞,表达突变型alkb酶的该细胞能够产生更多的ω-羟基化羧酸和/或其酯。来自表达突变型alkb酶的细胞的ω-羟基化羧酸和/或其酯的产率相对于表达野生型alkb酶的细胞更高。在一个实例中,突变型alkb能够在其中突变型alkb相对于野生型细胞表达(即,其中不表达突变型alkb酶)的细胞中增加ω-羟基月桂酸甲酯(hlame)的产生和/或积累。根据本发明的一个方面,提供了至少一种表达突变型alkb酶的微生物细胞,所述突变型alkb酶在alkb的野生型序列中包含至少一个点突变,其中所述点突变在野生型alkb酶的氨基酸位置v129和/或t136处。特别地,突变型alkb酶中的点突变在野生型alkb酶的氨基酸位置v129和/或t136处。这类氧化还原酶alkb的原型是来自恶臭假单胞菌(pseudomonasputida)alkbgt系统的氧化还原蛋白,其依赖于两种辅助多肽alkg和alkt。alkt是依赖于fad的红素氧还蛋白还原酶,其将电子从nadh转移至alkg。alkg是一种红素氧还蛋白,一种含铁的氧化还原蛋白,其作为alkb的直接电子供体发挥功能。在一个实例中,如本文所用的术语“alkb型氧化还原酶”是指具有烃结合位点的红素氧还蛋白依赖性的氧化还原酶。特别地,如本文所用的术语“烃结合位点”是指能够结合烃、特别是烷烃和/或烯烃的蛋白表面的一段(stretch),更特别是在蛋白的催化中心的范围内。在一个实例中,如本文所用的术语“红素氧还蛋白依赖性的烷烃氧化酶”是指氧化还原酶,其识别作为其底物的经由红素氧还原蛋白接收电子的烷烃,所述红素氧还原蛋白在具体实例中具有铁-硫蛋白,其具有携带2个α螺旋和2至3个β-链的α+β类折叠,其将电子转移至烷烃氧化酶。更特别地,alkb型氧化还原酶可以是来自恶臭假单胞菌gpo1的alkb(访问代码:cab54050.1,本申请中使用的任何访问代码是指来自由ncbi运行的genbank数据库的相应序列,其中所提及的发布版本是指在2012年1月15日在线可得的发布版本)或其变体。特别地,野生型alkb可以包含与多肽seqidno:1具有至少50、60、65、70、75、80、85、90、91、94、95、98或100%的序列同一性的多肽。更特别地,野生型alkb具有包含seqidno:1的氨基酸的多肽序列,其对于功能,例如蛋白的催化活性或蛋白的折叠或结构而言是必需的。其它氨基酸可以缺失、取代或通过插入来替代,或者必需氨基酸以保守方式替代,以达到保持野生型alkb的生物活性的效果。特别地,根据本发明的任一方面的野生型alkb可以与seqidno:1具有70%序列同一性,其中野生型alkb酶在氨基酸位置129、132和136处不包含突变。特别地,野生型alkb酶包含氨基酸位置129处的氨基酸缬氨酸(v)、氨基酸位置132处的亮氨酸(l)和氨基酸位置136处的苏氨酸(t)。术语“酶”意指完全或主要由蛋白或多肽构成的任何物质,其或多或少特异性地催化或促进一种或多种化学或生物化学反应。根据本发明的任一方面使用的酶的实例可以是alkb。特别地,alkb烷烃羟化酶或ec1.14.15.3的烷烃单加氧酶。alkb烷烃羟化酶是反应体系的组分,该体系包含三种酶组分ec1.14.15.3的alkb烷烃羟化酶、ec1.18.1.1或ec1.18.1.4的alkt红素氧还蛋白nad(p)+还原酶以及红素氧还蛋白alkg。alkt是依赖于fad的红素氧还蛋白还原酶,其将电子从nadh转移至alkg。alkg是一种红素氧还蛋白,一种含铁的氧化还原蛋白,其作为alkb的直接电子供体发挥功能。在一个优选实施方案中,如本文所用的术语“alkb型氧化还原酶”是指具有烃结合位点的红素氧还蛋白依赖性的氧化还原酶。alkb酶可以与由alkbgt编码的恶臭假单胞菌gpo1的烷烃单加氧酶具有至少50%的序列同一性。特别地,alkb野生型酶可以与多肽cab54050.1(seqidno:1;本申请中使用的任何访问代码是指来自由ncbi运行的genbank数据库的相应序列,其中提及的发布版本是2016年4月28日在线可得的发布版本)具有至少70%的序列同一性。“多肽”(一种或多种肽)是通过称为肽键的化学键连接在一起的称为氨基酸的化学构建嵌段的链。蛋白或多肽,包括酶,可以是“天然的”或“野生型的”,意味着其分别在自然界中存在或具有天然蛋白的氨基酸序列。这些术语有时可互换使用。多肽可以是糖基化的,或可以不是糖基化的。如本文中与细胞或微生物结合使用的术语“野生型”可以表示具有呈如在自然环境中天然可见的形式的基因组组成的细胞。该术语可适用于全细胞和个别基因两者。因此,术语“野生型”还可以包括已在其它方面(即,关于一个或多个基因)、但不关于目标基因经遗传修饰的细胞。因此,术语“野生型”不包括如下这类细胞,其中已经使用重组方法人工至少部分改变特定目标基因的基因序列。因此,根据本发明的任一方面的野生型细胞是指相对于全基因组和/或特定基因没有遗传突变的细胞。因此,在一个实例中,野生型alkb可以是自然界中天然存在的酶,其没有根据本发明的任一方面的任何特定突变。在另一个实例中,相对于根据本发明的任一方面的突变型alkb而言的野生型细胞是指具有alkb的天然/未改变的表达的细胞,所述alkb具有可以天然存在、但不在氨基酸位置129、132和136处存在的突变。多肽也可以是“突变体”,意味着其已经从其野生型形式或另一种突变体制备、改变、衍生或以某种方式与其野生型形式或另一种突变体不同或由其改变。突变型蛋白通常在一个或多个位置处具有氨基酸取代。突变型dna分子通常在一个或多个位置处具有核苷酸取代。与野生型相比,蛋白或dna分子的突变型形式可以具有相同或改变的功能。为了简化,突变体可以通过它们与突变产生的单个氨基酸代码的变异来指代。例如,在一种形式中,突变体被称为xposy,其中“x”是指原始序列中氨基酸的单字母代码,“pos”是指序列中突变的位置,且y是指在突变的位置处出现的新氨基酸的单字母代码。例如,v129t意味着在原始蛋白中,位置129处的氨基酸是缬氨酸(“v”),但在突变体中,缬氨酸被苏氨酸(“t”)替代。特别地,根据本发明的任一方面的突变型alkb酶可以在alkb酶的氨基酸位置129、132和/或136处包含至少一个点突变。特别地,点突变可以在氨基酸位置129、132或136处。在另一个实例中,点突变可以在氨基酸位置129和132或129和136或132和136处。在一个进一步实例中,点突变可以在氨基酸位置129、136和136处。更特别地,位置129处的氨基酸缬氨酸可以被转化为选自以下的氨基酸:甘氨酸、丙氨酸、亮氨酸、异亮氨酸、脯氨酸、苯丙氨酸、酪氨酸、色氨酸、丝氨酸、苏氨酸、半胱氨酸、甲硫氨酸、天冬酰胺、谷氨酰胺、赖氨酸、精氨酸、组氨酸、天冬氨酸和谷氨酸。特别地,位置129处的氨基酸缬氨酸可以被转化为选自以下的氨基酸:苏氨酸、丙氨酸、丝氨酸、半胱氨酸、天冬酰胺和甘氨酸。更特别地,位置129处的氨基酸缬氨酸可以被转化为苏氨酸或丙氨酸。位置136处的氨基酸苏氨酸也可以被转化为选自以下的氨基酸:甘氨酸、丙氨酸、亮氨酸、异亮氨酸、脯氨酸、苯丙氨酸、酪氨酸、色氨酸、丝氨酸、缬氨酸、半胱氨酸、甲硫氨酸、天冬酰胺、谷氨酰胺、赖氨酸、精氨酸、组氨酸、天冬氨酸和谷氨酸。特别地,位置136处的氨基酸苏氨酸可以被转化为选自以下的氨基酸:甘氨酸、丙氨酸、亮氨酸、异亮氨酸、脯氨酸、苯丙氨酸、色氨酸、丝氨酸、缬氨酸、半胱氨酸和甲硫氨酸。根据本发明的任一方面的点突变可以在位置129和/或136处。在一个实例中,根据本发明的任一方面的突变型alkb中的点突变可以在位置129处。在另一个实例中,根据本发明的任一方面的突变型alkb中的点突变可以在位置136处。在一个进一步实例中,根据本发明的任一方面的突变型alkb中的点突变可以组合在位置129和136处。更特别地,相对于alkb的野生型序列,根据本发明的任一方面的细胞的点突变可以选自v129t、t136a、v129a及其组合。突变型alkb酶可以包含v129t、t136a或v129a点突变。特别地,根据本发明的任一方面的alkb酶可以包含点突变v129t。在一个实例中,alkb酶可以进一步包含点突变t136a。在另一个实例中,根据本发明的任一方面的任何细胞的突变型alkb酶可以包含点突变v129t和t136a、v129a和t136a。特别地,微生物细胞表达突变型alkb酶,所述突变型alkb酶在alkb的野生型序列中包含至少一个点突变,其中所述点突变在野生型alkb酶的氨基酸位置v129和/或t136处,其中氨基酸v129处的点突变选自v129t和v129a。更特别地,点突变可以在氨基酸位置t136处。甚至更特别地,氨基酸位置t136处的点突变可以是t136a。在一个实例中,当点突变在氨基酸位置v129处时,突变可以选自v129t和v129a。在另一个实例中,点突变可以在氨基酸位置v129和t136处。特别地,氨基酸位置v129处的点突变可以选自v129t和v129a,且氨基酸位置t136处的点突变是t136a。特别地,点突变可以是v129a。在一个实例中,野生型序列可以是seqidno:1或其变体。如本文所用的alkb或至少seqidno:1的“变体”是指被一个或多个氨基酸改变的氨基酸序列。所述变体可以具有“保守”变化,其中取代的氨基酸具有相似的结构或化学特性(例如,用异亮氨酸替代亮氨酸)。更罕见地,变体可以具有“非保守”变化(例如,用色氨酸替代甘氨酸)。类似的微小变异也可以包括氨基酸缺失或插入,或两者。可以使用本领域众所周知的计算机程序,例如dnastar软件,找到确定哪些氨基酸残基可以被取代、插入或缺失而不消除生物或免疫活性的指导。根据本发明的任一方面,用作野生型alkb的seqidno:1的变体可以不包含在氨基酸位置129、132和/或136处的突变。分子生物学中的任何技术都可用于产生具有特定点突变的alkb酶。术语“点突变”或单碱基修饰是一种类型的突变,其引起遗传物质dna或rna的单核苷酸碱基改变、插入或缺失。在一个实例中,点突变可以是指alkb基因的野生型序列中的氨基酸取代。可以使用的方法的一些实例包括易错聚合酶链反应、盒式诱变(其中优化的特定区域被合成诱变的寡核苷酸替代)、寡核苷酸定向诱变、平行pcr(其使用在相同容器中平行发生的大量不同的pcr反应,以使得一个反应的产物引发另一个反应的产物)、随机诱变(例如,通过随机片段化和通过相互引发的片段的重新组装);位点特异性突变(通过模板的随机片段化引入长序列中,随后在诱变寡核苷酸存在的情况下重新组装片段);平行pcr(例如,dna序列的库上的重组);有性pcr;和化学诱变(例如,通过亚硫酸氢钠、亚硝酸、羟胺、肼、甲酸或通过向pcr反应中添加亚硝基胍、5-溴尿嘧啶、2-氨基嘌呤和吖啶代替核苷酸前体;或通过添加插入剂诸如原黄素、吖啶黄素、奎纳克林);辐射(x-射线或紫外线,和/或使多核苷酸在缺乏正常dna损伤修复功能的宿主细胞中繁殖);或dna改组(例如,核酸片段或多核苷酸的合并物(pool)的体外或体内同源重组)。这些技术中的任一种也可以在低保真聚合条件下采用,以在长序列上随机引入低水平的点突变,或诱变未知序列的片段的混合物。根据本发明的任一方面使用的任何酶可以是分离的酶。特别地,根据本发明的任一方面使用的酶可以以活性状态和在对于其活性而言必需的所有辅因子、底物、辅助和/或活化多肽或因子存在下使用。如本文所用,术语“分离的”意指与其天然存在于其中的细胞相比,目标酶是富集的。所述酶可以通过sds聚丙烯酰胺电泳和/或活性测定来富集。例如,目标酶可以构成制备物中存在的所有多肽的多于5%、10%、20%、50%、75%、80%、85%、90%、95%或99%,如通过在用考马斯蓝染料染色后目视检查聚丙烯酰胺凝胶所判断。根据本发明的任一方面,所述微生物细胞可以选自以下的细菌物种:乏养菌属(abiotrophia)、acaryochloris、accumulibacter、醋弧菌属(acetivibrio)、醋酸杆菌属(acetobacter)、醋盐杆菌属(acetohalobium)、醋丝菌属(acetonema)、无色杆菌属(achromobacter)、氨基酸球菌属(acidaminococcus)、酸微菌属(acidimicrobium)、嗜酸菌属(acidiphilium)、酸硫杆状菌属(acidithiobacillus)、酸杆菌属(acidobacterium)、热酸菌属(acidothermus)、食酸菌属(acidovorax)、不动杆菌属(acinetobacter)、放线杆菌属(actinobacillus)、放线菌属(actinomyces)、束丝放线菌属(actinosynnema)、气球菌属(aerococcus)、气微菌属(aeromicrobium)、气单胞菌属(aeromonas)、阿菲波菌属(afipia)、凝聚杆菌属(aggregatibacter)、土壤杆菌属(agrobacterium)、ahrensia、akkermansia、食碱菌属(alcanivorax)、脂环酸芽胞杆菌属(alicycliphilus)、脂环酸杆菌属(alicyclobacillus)、aliivibrio、碱湖生菌属(alkalilimnicola)、alkaliphilus、异着色菌属(allochromatium)、交替单胞菌目(alteromonadales)、交替单胞菌属(alteromonas)、aminobacterium、氨基酸单胞菌属(aminomonas)、制氨菌属(ammonifex)、拟无枝酸菌属(amycolatopsis)、amycolicicoccus、鱼腥藻属(anabaena)、厌氧小杆菌属(anaerobaculum)、厌氧球菌属(anaerococcus)、anaerofustis、厌氧绳菌属(anaerolinea)、厌氧粘细菌属(anaeromyxobacter)、anaerostipes、anaerotruncus、无形体属(anaplasma)、厌氧芽孢杆菌属(anoxybacillus)、产液菌属(aquifex)、隐稳杆菌属(arcanobacterium)、弓形杆菌属(arcobacter)、aromatoleum、节杆菌属(arthrobacter)、arthrospira、不粘柄菌属(asticcacaulis)、陌生物菌属(atopobium)、橙单胞菌属(aurantimonas)、固氮弓菌属(azoarcus)、固氮根瘤菌属(azorhizobium)、固氮螺菌属(azospirillum)、固氮菌属(azotobacter)、杆菌属(bacillus)、巴通体属(bartonella)、basfia、baumannia、蛭弧菌属(bdellovibrio)、贝扎托菌属(beggiatoa)、拜叶林克氏菌属(beijerinckia)、bermanella、获山氏菌属(beutenbergia)、双岐杆菌属(bifidobacterium)、嗜胆菌属(bilophila)、blastopirellula、blautia、blochmannia、博德氏菌属(bordetella)、疏螺旋体属(borrelia)、短状杆菌属(brachybacterium)、短螺旋体属(brachyspira)、短根瘤菌属(bradyrhizobium)、短小芽孢杆菌属(brevibacillus)、短杆菌属(brevibacterium)、短波单胞菌属(brevundimonas)、布鲁菌属(brucella)、布赫纳氏菌属(buchnera)、bulleidia、伯克氏菌属(burkholderia)、丁酸弧菌属(butyrivibrio)、caldalkalibacillus、caldanaerobacter、热解纤维素菌属(caldicellulosiruptor)、calditerrivibrio、caminibacter、弯曲菌属(campylobacter)、carboxydibrachium、一氧化碳嗜热菌属(carboxydothermus)、心杆菌属(cardiobacterium)、肉杆菌属(carnobacterium)、carsonella、catenibacterium、catenulispora、卡托氏菌属(catonella)、柄杆菌属(caulobacter)、纤维单胞菌属(cellulomonas)、纤维弧菌属(cellvibrio)、蜈蚣菌属(centipeda)、根瘤菌属(chelativorans)、绿屈挠菌属(chloroflexus)、色杆菌属(chromobacterium)、色盐杆菌属(chromohalobacter)、chthoniobacter、citreicella、柠檬酸杆菌属(citrobacter)、柠檬酸微菌属(citromicrobium)、棍状杆菌属(clavibacter)、cloacamonas、梭状芽孢杆菌属(clostridium)、柯林斯氏菌属(collinsella)、科尔韦氏菌属(colwellia)、丛毛单胞菌属(comamonas)、conexibacter、聚集杆菌属(congregibacter)、coprobacillus、粪球菌属(coprococcus)、粪热杆菌属(coprothermobacter)、coraliomargarita、科里氏杆菌属(coriobacterium)、corrodens、棒状杆菌属(corynebacterium)、柯克斯体属(coxiella)、鳄球藻属(crocosphaera)、阪崎肠杆菌属(cronobacter)、短小杆菌属(cryptobacterium)、贪铜菌属(cupriavidus)、蓝菌属(cyanobium)、蓝丝菌属(cyanothece)、拟筒孢蓝细菌属(cylindrospermopsis)、dechloromonas、铁还原杆菌属(deferribacter)、dehalococcoides、dehalogenimonas、异常球菌属(deinococcus)、代尔夫特菌属(delftia)、氮还原弧菌属(denitrovibrio)、皮生球菌属(dermacoccus)、desmospora、脱硫盒菌属(desulfarculus)、desulphateibacillum、脱亚硫酸菌属(desulfitobacterium)、脱硫橄榄状菌属(desulfobacca)、脱硫杆菌属(desulfobacterium)、脱硫叶菌属(desulfobulbus)、脱硫球菌属(desulfococcus)、脱硫盐菌属(desulfohalobium)、脱硫微菌属(desulfomicrobium)、desulfonatronospira、desulforudis、脱硫小杆菌属(desulfotalea)、脱硫肠状菌属(desulfotomaculum)、脱硫弧菌属(desulfovibrio)、(desulfurispirillum)、脱硫杆菌属(desulfurobacterium)、除硫单胞菌属(desulfuromonas)、dethiobacter、脱硫代硫酸盐弧菌属(dethiosulfovibrio)、戴阿利斯特杆菌属(dialister)、偶蹄形菌属(dichelobacter)、dickeya、网球菌属(dictyoglomus)、迪茨氏菌属(dietzia)、dinoroseobacter、dorea、爱德华氏菌属(edwardsiella)、埃里希氏体属(ehrlichia)、艾肯氏菌属(eikenella)、elusimicrobium、endoriftia、水栖菌属(enhydrobacter)、肠杆菌属(enterobacter)、肠球菌属(enterococcus)、epulopiscium、欧文氏杆菌属(erwinia)、丹毒丝菌属(erysipelothrix)、赤细菌属(erythrobacter)、埃希氏杆菌属(escherichia)、产氢新菌属(ethanoligenens)、真杆菌属(eubacterium)、真杆菌属(eubacterium)、微小杆菌属(exiguobacterium)、faecalibacterium、铁还原单胞菌属(ferrimonas)、闪烁杆菌属(fervidobacterium)、丝状杆菌属(fibrobacter)、finegoldia、弯枝菌属(flexistipes)、弗朗西斯氏菌属(francisella)、弗兰克氏菌属(frankia)、fructobacillus、fulvimarina、梭杆菌属(fusobacterium)、gallibacterium、报毛菌属(gallionella)、加德纳氏菌属(gardnerella)、孪生球菌属(gemella)、出芽菌属(gemmata)、芽单胞菌属(gemmatimonas)、土壤芽孢杆菌属(geobacillus)、土杆菌属(geobacter)、地嗜皮菌属(geodermatophilus)、冰居菌属(glaciecola)、粘杆菌属(gloeobacter)、舌蝇属(glossina)、葡糖酸醋酸杆菌属(gluconacetobacter)、戈登氏菌属(gordonia)、granulibacter、颗粒链菌属(granulicatella)、格里蒙菌属(grimontia)、嗜血菌属(haemophilus)、河氏菌属(hahella)、halanaerobiumns、haliangium、盐单胞菌属(halomonas)、盐红螺旋菌属(halorhodospira)、盐热发菌属(halothermothrix)、盐硫杆状菌属(halothiobacillus)、hamiltonella、螺杆菌属(helicobacter)、阳光杆菌属(heliobacterium)、草螺菌属(herbaspirillum)、herminiimonas、滑柱菌属(herpetosiphon)、希普氏菌属(hippea)、海氏菌属(hirschia)、嗜组织菌属(histophilus)、hodgkinia、hoeflea、霍尔德曼氏菌属(holdemania)、hydrogenivirga、hydrogenobaculum、hylemonella、生丝微菌属(hyphomicrobium)、生丝单胞菌属(hyphomonas)、idiomarina、泥杆菌属(ilyobacter)、间孢囊菌属(intrasporangium)、白蚁菌属(isoptericola)、等球菌属(isosphaera)、两面神菌属(janibacter)、紫色杆菌属(janthinobacterium)、琼斯氏菌属(jonesia)、jonquetella、kangiella、ketogulonicigenium、动球菌属(kineococcus)、金氏菌属(kingella)、克雷伯氏菌属(klebsiella)、考克氏菌属(kocuria)、koribacter、kosmotoga、扑科研菌属(kribbella)、纤线杆菌属(ktedonobacter)、皮球菌属(kytococcus)、labrenzia、乳杆菌属(lactobacillus)、乳球菌属(lactococcus)、鸥杆菌属(laribacter)、劳特罗普氏菌属(lautropia)、劳森氏菌属(lawsonia)、军团菌属(legionella)、雷弗松氏菌属(leifsonia)、lentisphaera、纤发鞘丝蓝细菌属(leptolyngbya)、钩端螺旋体属(leptospira)、纤发菌属(leptothrix)、纤毛菌属(leptotrichia)、明串珠菌属(leuconostoc)、韧皮杆菌属(liberibacter)、limnobacter、李斯特氏菌属(listeria)、loktanella、lutiella、鞘丝蓝细菌属(lyngbya)、lysinibacillus、巨大球菌属(macrococcus)、磁球菌属(magnetococcus)、磁螺菌属(magnetospirillum)、mahella、默罕氏菌属(mannheimia)、海洋杆状菌属(maricaulis)、海栖热菌属(marinithermus)、海杆菌属(marinobacter)、海单胞菌属(marinomonas)、深海菌属(mariprofundus)、maritimibacter、marvinbryantia、巨球形菌属(megasphaera)、稍热菌属(meiothermus)、蜜蜂球菌属(melissococcus)、中间根瘤菌属(mesorhizobium)、methylacidiphilum、methylibium、甲基小杆菌属(methylobacillus)、甲基杆菌属(methylobacter)、甲基杆菌属(methylobacterium)、甲基球菌属(methylococcus)、甲基孢囊菌属(methylocystis)、甲基微菌属(methylomicrobium)、噬甲基菌属(methylophaga)、嗜甲基菌目(methylophilales)、甲基曲菌属(methylosinus)、methyloversatilis、食甲基菌属(methylovorus)、微杆菌属(microbacterium)、微球菌属(micrococcus)、微鞘蓝细菌属(microcoleus)、微囊蓝细菌属(microcystis)、小月菌属(microlunatus)、小单孢菌属(micromonospora)、光岗菌属(mitsuokella)、动弯杆菌属(mobiluncus)、穆尔氏菌属(moorella)、莫拉氏菌属(moraxella)、南极嗜冷菌属(moritella)、分枝杆菌属(mycobacterium)、粘球菌属(myxococcus)、nakamurella、盐碱厌氧菌属(natranaerobius)、奈瑟氏球菌属(neisseria)、新立克次氏体属(neorickettsia)、neptuniibacter、nitratifractor、nitratiruptor、硝化杆菌属(nitrobacter)、硝化球菌属(nitrococcus)、亚硝化单胞菌属(nitrosomonas)、亚硝化螺菌属(nitrosospira)、硝化螺菌属(nitrospira)、诺卡氏菌属(nocardia)、类诺卡氏菌属(nocardioides)、拟诺卡氏菌属(nocardiopsis)、节球蓝细菌属(nodularia)、念珠蓝细菌属(nostoc)、新鞘脂菌属(novosphingobium)、oceanibulbus、oceanicaulis、海洋生菌属(oceanicola)、海洋栖热菌属(oceanithermus)、海洋芽胞杆菌属(oceanobacillus)、苍白杆菌属(ochrobactrum)、十八杆菌属(octadecabacter)、odyssella、寡养菌属(oligotropha)、olsenella、丰佑菌属(opitutus)、oribacterium、东方体属(orientia)、ornithinibacillus、颤蓝细菌属(oscillatoria)、绿颤细菌属(oscillochloris)、草酸杆菌属(oxalobacter)、类芽孢杆菌属(paenibacillus)、泛菌属(pantoea)、副球菌属(paracoccus)、parascardovia、parasutterella、parvibaculum、微单胞菌属(parvimonas)、短小盒菌属(parvularcula)、巴斯德氏菌属(pasteurella)、巴斯德氏芽菌属(pasteuria)、果胶杆菌属(pectobacterium)、片球菌属(pediococcus)、pedosphaera、pelagibaca、远洋杆菌属(pelagibacter)、暗杆菌属(pelobacter)、pelotomaculum、peptoniphilus、消化链球菌属(peptostreptococcus)、persephonella、石袍菌属(petrotoga)、褐杆菌属(phaeobacter)、考拉杆菌属(phascolarctobacterium)、苯基杆菌属(phenylobacterium)、发光杆菌属(photobacterium)、小小梨形菌属(pirellula)、浮霉状菌属(planctomyces)、动性球菌属(planococcus)、plesiocystis、极胞菌属(polaromonas)、极胞菌属(polaromonas)、polymorphum、多核杆菌属(polynucleobacter)、海绵杆菌门(poribacteria)、原绿球菌属(prochlorococcus)、丙酸杆菌属(propionibacterium)、变形菌属(proteus)、普罗威登斯菌属(providencia)、假交替单胞菌属(pseudoalteromonas)、pseudoflavonifractor、假单胞菌属(pseudomonas)、假诺卡氏菌属(pseudonocardia)、假枝杆菌属(pseudoramibacter)、pseudovibrio、假黄色单胞菌属(pseudoxanthomonas)、嗜冷杆菌属(psychrobacter)、冷单胞菌属(psychromonas)、puniceispirillum、pusillimonas、维形杆菌属(pyramidobacter)、拉恩氏菌属(rahnella)、罗尔斯通氏菌属(ralstonia)、尖头藻属(raphidiopsis)、regiella、reinekea、肾杆菌属(renibacterium)、根瘤菌属(rhizobium)、红细菌属(rhodobacter)、红球菌属(rhodococcus)、红育菌属(rhodoferax)、红微菌属(rhodomicrobium)、红小梨形菌属(rhodopirellula)、红假单胞菌属(rhodopseudomonas)、红螺菌属(rhodospirillum)、立克次氏体属(rickettsia)、立克次氏小体属(rickettsiella)、riesia、罗斯伯里氏菌属(roseburia)、玫瑰菌属(roseibium)、玫瑰弯菌属(roseiflexus)、玫瑰杆菌属(roseobacter)、玫瑰单胞菌属(roseomonas)、玫瑰变色菌属(roseovarius)、罗斯氏菌属(rothia)、红长命菌属(rubrivivax)、红色杆菌属(rubrobacter)、鲁杰氏菌属(ruegeria)、瘤胃球菌属(ruminococcus)、ruthia、糖单孢菌属(saccharomonospora)、saccharophagus、糖多孢菌属(saccharopolyspora)、箭头菌属(sagittula)、salinispora、沙门氏菌属(salmonella)、血杆菌属(sanguibacte)、scardovia、塞巴鲁德菌属(sebaldella)、segniliparus、月形单胞菌属(selenomonas)、沙雷氏菌属(serratia)、希瓦氏菌属(shewanella)、志贺氏菌属(shigella)、shuttleworthia、sideroxydans、silicibacter、西蒙斯氏菌属(simonsiella)、中华根瘤菌属(sinorhizobium)、史雷克氏菌属(slackia)、伴蝇菌属(sodalis)、solibacter、solobacterium、堆囊菌属(sorangium)、球杆菌属(sphaerobacter)、鞘脂菌属(sphingobium)、鞘氨醇单胞菌属(sphingomonas)、sphingopyxis、螺旋体属(spirochaeta)、芽孢八叠球菌属(sporosarcina)、stackebrandtia、葡萄球菌属(staphylococcus)、starkeya、寡养单胞菌属(stenotrophomonas)、标桩菌属(stigmatella)、链杆菌属(streptobacillus)、链球菌属(streptococcus)、链霉菌属(streptomyces)、链孢囊菌属(streptosporangium)、subdoligranulum、subvibrioides、succinatimonas、亚硫酸盐杆菌属(sulfitobacter)、硫化杆菌属(sulfobacillus)、sulfuricurvum、sulfurihydrogenibium、sulfurimonas、硫化螺旋菌属(sulfurospirillum)、sulfurovum、萨特氏菌属(sutterella)、symbiobacterium、集胞蓝细菌属(synechocystis)、互营杆菌属(syntrophobacter)、共养香肠样杆菌属(syntrophobotulus)、共养单胞菌属(syntrophomonas)、互营热菌属(syntrophothermus)、互养菌属(syntrophus)、taiwanensis、泰勒氏菌属(taylorella)、teredinibacter、terriglobus、thalassiobium、索氏菌属(thauera)、嗜热好氧杆菌属(thermaerobacter)、热厌氧弧菌属(thermanaerovibrio)、thermincola、热厌氧杆菌属(thermoanaerobacter)、热厌氧杆菌属(thermoanaerobacterium)、thermobaculum、喜热裂孢菌属(thermobifida)、嗜热双孢菌属(thermobispora)、热发状菌属(thermocrinis)、thermodesulphateator、热脱硫杆菌属(thermodesulfobacterium)、热脱硫菌属(thermodesulfobium)、热脱硫弧菌属(thermodesulfovibrio)、热微菌属(thermomicrobium)、高温单孢菌属(thermomonospora)、thermosediminibacter、thermosinus、栖热腔菌属(thermosipho)、thermosynechococcus、栖热孢菌属(thermotoga)、热弧菌属(thermovibrio)、栖热菌属(thermus)、thioalkalimicrobium、thioalkalivibrio、硫杆菌属(thiobacillus)、硫微螺菌属(thiomicrospira)、硫单胞菌属(thiomonas)、甲苯单胞菌属(tolumonas)、密螺旋体属(treponema)、tribocorum、束毛蓝细菌属(trichodesmium)、tropheryma、特吕珀菌属(truepera)、冢村氏菌属(tsukamurella)、turicibacter、贪噬菌属(variovorax)、韦荣氏菌属(veillonella)、verminephrobacter、疣微菌属(verrucomicrobium)、疣孢菌属(verrucosispora)、vesicomyosocius、弧菌属(vibrio)、弧菌目(vibrionales)、食物谷菌属(victivallis)、魏斯氏菌属(weissella)、威格尔斯沃思氏菌属(wigglesworthia)、沃尔巴克氏体属(wolbachia)、沃林氏菌属(wolinella)、黄色杆菌属(xanthobacter)、黄单胞菌属(xanthomonas)、致病杆菌属(xenorhabdus)、xylanimonas、木杆菌属(xylella)、耶尔森氏菌属(yersinia)、zinderia和发酵单孢菌属(zymomonas)。特别地,所述微生物细胞可以来自大肠杆菌(e.coli)、假单胞菌属种(pseudomonassp.)、荧光假单胞菌(pseudomonasfluorescens)、恶臭假单胞菌(pseudomonasputida)、施氏假单胞菌(pseudomonasstutzeri)、不动杆菌属种(acinetobactersp.)、伯克氏菌属种(burkholderiasp.)、burkholderiathailandensis、cyanobakterien、克雷伯氏菌属种(klebsiellasp.)、产酸克雷伯氏菌(klebsiellaoxytoca)、沙门氏菌属种(salmonellasp.)、根瘤菌属种(rhizobiumsp.)和苜蓿根瘤菌(rhizobiummeliloti)、芽孢杆菌属种(bacillussp.)、枯草芽孢杆菌(bacillussubtilis)、梭状芽胞杆菌属种(clostridiumsp.)、棒状杆菌属种(corynebacteriumsp.)、谷氨酸棒状杆菌(corynebacteriumglutamicum)、短杆菌属种(brevibacteriumsp.)、小球藻属种(chlorellasp.)和念珠藻属种(nostocsp.)。更特别地,所述微生物细胞可以是大肠杆菌。在另一个实例中,微生物细胞可以是酵母细胞。根据本发明的任一方面的合适酵母可以属于在http://www.dsmz.de/species/yeasts.htm列出的属。特别地,所述酵母可以选自粟酒裂殖酵母、酿酒酵母和巴斯德毕赤酵母。根据本发明的任一方面使用的酶可以是重组的。如本文所用,术语“重组体”是指分子或由此类分子编码,特别是多肽或核酸,其本身不是天然存在的,而是遗传工程改造的结果;或者是指包含重组分子的细胞。例如,如果核酸分子包含与编码催化活性多肽的序列功能性连接的启动子且该启动子已经工程改造、以使得相对于包含原始的未改变的核酸分子的相应野生型细胞中的多肽的水平而言过表达该催化活性多肽,则所述核酸分子是重组的。技术人员将能够使用本领域已知的任何方法来遗传修饰细胞或微生物。根据本发明的任一方面,经遗传修饰的细胞可以进行遗传修饰,以使得在限定时间间隔内,在2小时内,特别是在8小时或24小时内,其形成野生型细胞的至少一倍或两倍、特别是至少10倍、至少100倍、至少1000倍或至少10000倍的ω-官能化羧酸酯。产物形成的增加可以例如通过以下来测定:在相同条件(相同细胞密度、相同营养培养基、相同培养条件)下在合适的营养培养基中将根据本发明的任一方面的细胞和野生型细胞各自分别培养特定时间间隔,然后测定营养培养基中目标产物(ω-官能化羧酸酯)的量。经遗传修饰的细胞或微生物可以在遗传上不同于野生型细胞或微生物。根据本发明的任一方面的经遗传修饰的微生物与野生型微生物之间的遗传差异可以是经遗传修饰的微生物中存在完整基因、氨基酸、核苷酸等,其可能在野生型微生物中不存在。在一个实例中,根据本发明的任一方面的经遗传修饰的微生物可以包含使得微生物能够产生相对于具有野生型或无alkb表达的野生型细胞更多的hlame的酶,特别是突变型alkb。野生型微生物相对于本发明的经遗传修饰的微生物可以没有酶或没有酶的可检测活性。如本文所用,术语“经遗传修饰的微生物”可以与术语“经遗传修饰的细胞”互换使用。根据本发明的任一方面的经遗传修饰在微生物的细胞上实施。根据本发明的任一方面的细胞根据本领域已知的任何方法进行遗传转化。特别地,所述细胞可以根据wo2013024114中公开的方法制备。如本文所用,短语“与其野生型相比经遗传修饰的细胞的酶活性增加”是指相应的酶活性增加到至少2倍、特别是至少10倍、更特别是至少100倍、还更特别是至少1000倍、甚至更特别是至少10000倍。根据本发明的任一方面的突变型alkb酶能够(a)催化作为底物的c6-c14羧酸和/或其酯转化为相应的ω-羟基羧酸和/或其酯;或(b)催化作为底物的c6-c14羧酸和/或其酯转化为相应的ω-氧代羧酸和/或其酯。术语“底物”意指通过酶催化剂的作用转化或意图转化为另一化合物的任何物质或化合物。该术语包括芳族和脂族化合物,并且不仅包括单一化合物,而且包括化合物的组合,诸如含有至少一种底物的溶液、混合物和其它材料。根据本发明的任一方面,所述底物可以是至少一种c6-c14羧酸和/或其酯。因此,底物可以选自c6、c7、c8、c9、c10、c11、c12、c13、c14及其酯。所述底物可以是c6-c14羧酸和/或其酯的组合。特别地,所述底物可以是至少一种c8-c12羧酸和/或其酯、至少一种c6-c10羧酸和/或其酯、或至少一种c8-c10羧酸和/或其酯。在一个实例中,所述底物可以是月桂酸和/或其酯。特别地,所述酯可以是羧酸的乙基或甲酯。在一个实例中,根据本发明的任一方面的作为底物的酯可以是甲基月桂酸甲酯(lame)。在该实例中,突变型alkb能够分别催化月桂酸和/或月桂酸甲酯(lame)转化为ω-羟基月桂酸和/或ω-羟基月桂酸甲酯(hlame)的氧化反应。在另一个实例中,突变型alkb能够分别催化月桂酸和/或月桂酸甲酯(lame)转化为ω-氧代月桂酸和/或ω-氧代月桂酸甲酯(olame)。在又一个进一步实例中,突变型alkb能够同时催化月桂酸和/或月桂酸甲酯(lame)转化为ω-羟基月桂酸和/或ω-羟基月桂酸甲酯(hlame)和ω-氧代月桂酸和/或ω-氧代月桂酸甲酯(olame)。如本文所用,“氧化反应”或“加氧反应”是化学或生物化学反应,其涉及将氧添加至底物,以形成加氧或氧化的底物或产物。氧化反应通常伴随还原反应(因此为术语“氧化还原”反应,用于氧化和还原)。当化合物接收氧或失去电子时,化合物被“氧化”。当化合物失去氧或获得电子时,化合物被“还原”。氧化反应的一些实例包括但不限于将lame转化为hlame、将hlame转化为olame、将olame转化为ddame等。特别地,突变型alkb酶能够比野生型alkb酶由羧酸产生更多的ω-羟基-和/或ω-氧代羧酸和/或其酯。形成的产品也将更少。在一个实例中,可以使用大肠杆菌作为宿主细胞来分析根据本发明的任一方面的alkb突变体的异源表达的效果。这些突变型alkb酶可以被转化至大肠杆菌中,并且这些转化的细胞在生物反应器中在底物诸如lame存在的情况下在工艺相关条件下生长。与表达野生型alkb的任何参考菌株相比,突变型菌株可显示lame对hlame的生物转化率显著增加以及hlame:ddame的比率改善至最高两倍。该比率可以表明hlame的底物特异性降低,因为hlame是alkb反应级联中产生ddame的前体。因此,根据本发明的任一方面的细胞可以产生更节能的方法,其中可以增加时空产率以及产物产率,其有助于更经济的聚酰胺12前体生产方法。特别地,根据本发明的任一方面的细胞和/或方法能够相对于野生型细胞中的hlame/ddame比率增加ω-羟基月桂酸甲酯(hlame)/十二烷二酸单甲酯(ddame)(hlame/ddame)比率。特别地,该方法包括使根据本发明的任一方面的细胞与底物lame接触。通过在给定时间点将反应混合物中产生的hlame的总量除以反应中产生的ddame的总量来计算该比率。特别地,hlame和/或ddame的量可以通过本领域任一已知方法在任何给定时间点测量。更特别地,hlame和/或ddame的浓度可以使用气相或液相色谱联用质谱来测量。hlame的浓度越高,比率越高。特别地,hlame/ddame比率可以至少为1:1。这本身可以高于野生型酶和/或细胞。更特别地,hlame/ddame比率可以至少为约1.1:1、1.2:1、1.3:1、1.4:1、1.5:1、1.6:1、1.7:1、1.8:1、1.9:1、2:1、2.1:1、2.2:1、2.3:1、2.4:1、2.5:1、2.6:1、2.7:1、2.8:1、2.9:1、3:1等。甚至更特别地,hlame/ddame比率可以至少为1.5:1。在另一个实例中,根据本发明的任一方面的细胞和/或方法能够相对于野生型细胞中的alame/ddame比率增加ω-氨基月桂酸甲酯(alame)/十二烷二酸单甲酯(ddame)(alame/ddame)比率。特别地,该方法包括使根据本发明的任一方面的细胞与底物lame接触。通过在给定时间点将反应混合物中产生的alame的总量除以反应中产生的ddame的总量来计算该比率。特别地,alame和/或ddame的量可以通过本领域任一已知方法在任何给定时间点测量。更特别地,alame和/或ddame的浓度可以使用气相或液相色谱联用质谱来测量。alame的浓度越高,比率越高。特别地,alame/ddame比率可以至少为1:1。这本身可以高于野生型酶和/或细胞。更特别地,alame/ddame比率可以至少为约1.1:1、1.2:1、1.3:1、1.4:1、1.5:1、1.6:1、1.7:1、1.8:1、1.9:1、2:1、2.1:1、2.2:1、2.3:1、2.4:1、2.5:1、2.6:1、2.7:1、2.8:1、2.9:1、3:1等。甚至更特别地,alame/ddame比率可以至少为1.5:1。根据本发明的一个进一步方面,提供了根据本发明任一方面的突变型alkb酶和/或包含和表达突变型alkb酶的细胞用于催化月桂酸甲酯(lame)转化为ω-羟基月桂酸甲酯(hlame)的用途。如本文所用,“约”或“近似”应表示在给定值或范围的20%内,特别是10%内,且更特别是5%内。特别地,根据本发明的任一方面的突变型alkb酶就底物具有非官能化的末端甲基键的c6-c14的羧酸和/或其酯,诸如羧酸甲酯、乙酯或丙酯或烷烃而言相对于野生型alkb酶可具有降低的km值或增加的vmax值,因此导致突变型alkb的活性改善。例如,根据本发明的任一方面的突变型alkb酶能够识别lame,且因此催化底物lame氧化转化为hlame。在另一个实例中,根据本发明的任一方面的突变型alkb酶就底物olame或具有氧代官能化末端键的其它底物,诸如ω-氧代羧酸甲酯、乙酯或丙酯或链烷醛而言相对于野生型alkb酶可具有增加的km值或降低的vmax值,因此导致活性降低。在一个进一步实例中,根据本发明的任一方面的突变型alkb酶就底物hlame或具有羟基官能化末端键的其它底物,诸如ω-羟基脂肪酸甲酯、乙酯或丙酯或链烷醇而言相对于野生型alkb酶可具有增加的km值或降低的vmax值,因此导致活性降低。因此,根据本发明的任一方面的突变型alkb酶可用于产生和/或积累hlame。特别地,根据本发明的任一方面的突变型alkb酶相对于野生型alkb酶能够产生更多的hlame和/或该hlame可以不转化为其它下游产物,诸如olame和/或ddame。或者,根据本发明的任一方面的突变型alkb酶可用于相对于同时作为不希望的副产物产生的ddame产生和/或积累更多的hlame。特别地,与野生型alkb酶相比,根据本发明的任一方面的突变型alkb酶能够相对于同时作为不希望的副产物产生的ddame产生更多的hlame。因此,浪费的资源更少,因为相对于野生型alkb酶,少量底物lame可以足以产生大量hlame。特别地,可以证明,与野生型alkb相比,根据本发明的任一方面的alkb突变体证明具有更高的hlame生产率并且另外对hlame和/或olame的底物特异性降低(导致hlame/ddame产物比增加)。类似的解释可能与可以看到的增加的alame/ddame比率相关。在一个实例中,在根据本发明的任一方面的细胞中发现的内源性醇脱氢酶可以参与alame的形成,因此显示olame没有增加。根据本发明的任一方面的细胞可以进一步进行遗传修饰,以包含相对于野生型细胞增加的酶e1、ω-转氨酶的表达。特别地,酶e1能够催化ω-氧代羧酸和/或其酯转化为相应的ω-氨基羧酸和/或其酯。如本文所用,短语“酶活性增加”应理解为细胞内活性增加。基本上,酶促活性的增加可以通过以下实现:增加编码所述酶的一个基因序列或多个基因序列的拷贝数,使用强启动子或采用编码具有增加活性的相应酶的基因或等位基因,改变基因的密码子利用,以各种方式增加mrna或酶的半衰期,修饰基因表达的调节和任选通过组合这些措施。根据本发明的任一方面使用的经遗传修饰的细胞例如通过用含有期望基因、该基因的等位基因或其部分的载体和使得基因表达可能的载体转化、转导、缀合或这些方法的组合来产生。异源表达特别地通过在细胞染色体或染色体外复制载体中整合基因或等位基因来实现。在一个实例中,根据本发明的任一方面的细胞可以包含相对于野生型细胞增加的ω-转氨酶(e1)的表达。特别地,ω-转氨酶(e1)来自ec2.6.1。特别地,所述酶e1可以是选自以下的转氨酶(eh):恶臭假单胞菌(pseudomonasputida)(wp_016502144;wp_016500675.1)、紫色色杆菌(chromobacteriumviolaceum)(np_901695.1)、类球红细菌(rhodobactersphaeroides)2.4.1(yp_353455.1)和3hmu_a、aad41041.1、aak15486.1、abe03917.1、adr60699.1、adr61066.1、adr62525.1、ael07495.1、caz86955.1、efw82310.1、efw87681.1、egc99983.1、egd03176.1、ege58369.1、egh06681.1、egh08331.1、egh24301.1、egh32343.1、egh46412.1、egh55033.1、egh62152.1、egh67339.1、egh70821.1、egh71404.1、egh78772.1、egh85312.1、egh97105.1、egp57596.1、np_102850.1、np_106560.1、np_248912.1、np_248990.1、np_354026.2、np_421926.1、np_637699.1、np_642792.1、np_744329.1、np_744732.1、np_747283.1、np_795039.1、xp_002943905.1、yp_001021095.1、yp_001059677.1、yp_001061726.1、yp_001066961.1、yp_001074671.1、yp_001120907.1、yp_001140117.1、yp_001170616.1、yp_001185848.1、yp_001188121.1、yp_001233688.1、yp_001268866.1、yp_001270391.1、yp_001345703.1、yp_001412573.1、yp_001417624.1、yp_001526058.1、yp_001579295.1、yp_001581170.1、yp_001668026.1、yp_001669478.1、yp_001671460.1、yp_001685569.1、yp_001747156.1、yp_001749732.1、yp_001765463.1、yp_001766294.1、yp_001790770.1、yp_001808775.1、yp_001809596.1、yp_001859758.1、yp_001888405.1、yp_001903233.1、yp_001977571.1、yp_002229759.1、yp_002231363.1、yp_002280472.1、yp_002297678.1、yp_002543874.1、yp_002549011.1、yp_002796201.1、yp_002801960.1、yp_002875335.1、yp_002897523.1、yp_002912290.1、yp_002974935.1、yp_003060891.1、yp_003264235.1、yp_003552364.1、yp_003578319.1、yp_003591946.1、yp_003607814.1、yp_003641922.1、yp_003674025.1、yp_003692877.1、yp_003755112.1、yp_003896973.1、yp_003907026.1、yp_003912421.1、yp_004086766.1、yp_004142571.1、yp_004147141.1、yp_004228105.1、yp_004278247.1、yp_004305252.1、yp_004356916.1、yp_004361407.1、yp_004378186.1、yp_004379856.1、yp_004390782.1、yp_004472442.1、yp_004590892.1、yp_004612414.1、yp_004676537.1、yp_004693233.1、yp_004701580.1、yp_004701637.1、yp_004704442.1、yp_108931.1、yp_110490.1、yp_168667.1、yp_237931.1、yp_260624.1、yp_262985.1、yp_271307.1、yp_276987.1、yp_334171.1、yp_337172.1、yp_350660.1、yp_351134.1、yp_364386.1、yp_366340.1、yp_369710.1、yp_370582.1、yp_426342.1、yp_440141.1、yp_442361.1、yp_468848.1、yp_521636.1、yp_554363.1、yp_608454.1、yp_610700.1、yp_614980.1、yp_622254.1、yp_625753.1、yp_680590.1、yp_751687.1、yp_767071.1、yp_774090.1、yp_774932.1、yp_788372.1、yp_858562.1、yp_928515.1、yp_983084.1、yp_995622.1、zp_00948889.1、zp_00954344.1、zp_00959736.1、zp_00998881.1、zp_01011725.1、zp_01037109.1、zp_01058030.1、zp_01076707.1、zp_01103959.1、zp_01167926.1、zp_01224713.1、zp_01442907.1、zp_01446892.1、zp_01550953.1、zp_01625518.1、zp_01745731.1、zp_01750280.1、zp_01754305.1、zp_01763880.1、zp_01769626.1、zp_01865961.1、zp_01881393.1、zp_01901558.1、zp_02145337.1、zp_02151268.1、zp_02152332.1、zp_02167267.1、zp_02190082.1、zp_02242934.1、zp_02360937.1、zp_02367056.1、zp_02385477.1、zp_02456487.1、zp_02883670.1、zp_03263915.1、zp_03263990.1、zp_03400081.1、zp_03452573.1、zp_03456092.1、zp_03517291.1、zp_03529055.1、zp_03571515.1、zp_03572809.1、zp_03587785.1、zp_03588560.1、zp_03697266.1、zp_03697962.1、zp_04521092.1、zp_04590693.1、zp_04890914.1、zp_04891982.1、zp_04893793.1、zp_04902131.1、zp_04905327.1、zp_04941068.1、zp_04944536.1、zp_04945255.1、zp_04959332.1、zp_04964181.1、zp_05053721.1、zp_05063588.1、zp_05073059.1、zp_05077806.1、zp_05082750.1、zp_05091128.1、zp_05095488.1、zp_05101701.1、zp_05116783.1、zp_05121836.1、zp_05127756.1、zp_05637806.1、zp_05742087.1、zp_05783548.1、zp_05786246.1、zp_05843149.1、zp_05945960.1、zp_06459045.1、zp_06487195.1、zp_06492453.1、zp_06493162.1、zp_06703644.1、zp_06731146.1、zp_06839371.1、zp_07007312.1、zp_07266194.1、zp_07374050.1、zp_07662787.1、zp_07778196.1、zp_07797983.1、zp_08099459.1、zp_08138203.1、zp_08141719.1、zp_08142973.1、zp_08177102.1、zp_08185821.1、zp_08186468.1、zp_08208888.1、zp_08266590.1、zp_08402041.1、zp_08406891.1、zp_08522175.1、zp_08527488.1、zp_08631252.1和zp_08636687.1。特别地,所述酶e1可以是选自以下的转氨酶:np_901695.1、zp_03697266.1、aad41041.1、yp_002796201.1、zp_03697962.1、yp_001859758.1、yp_002229759.1、yp_001120907.1、yp_110490.1、zp_04964181.1、yp_774932.1、yp_001766294.1、yp_001581170.1、yp_622254.1、zp_03588560.1、yp_001809596.1、yp_370582.1、zp_03572809.1、np_248990.1、yp_001888405.1、zp_04905327.1、yp_001061726.1、yp_001668026.1、zp_01750280.1、zp_07778196.1、egh71404.1、np_744329.1、yp_004147141.1、adr61066.1、zp_05783548.1、yp_004701637.1、yp_366340.1、yp_003264235.1、egd03176.1、yp_001268866.1、zp_01901558.1、zp_05121836.1、yp_003692877.1、zp_03517291.1、yp_002974935.1、yp_001668026.1、adr61066.1、np_744329.1、yp_001268866.1、yp_004701637.1、zp_08142973.1、adr62525.1、yp_610700.1、np_747283.1、adr62525.1、yp_001270391.1、yp_004704442.1、yp_610700.1、yp_001747156.1、zp_08138203.1、zp_07266194.1、egh70821.1、yp_351134.1、egh32343.1、egh08331.1、egh67339.1、yp_001668026.1、yp_004701637.1、yp_237931.1、zp_03400081.1、zp_05116783.1、zp_01550953.1、zp_07662787.1、yp_928515.1、yp_788372.1、yp_001021095.1、zp_07797983.1、yp_003578319.1、yp_004305252.1、np_248912.1、zp_08636687.1、yp_003912421.1、yp_751687.1、zp_08142973.1、yp_271307.1、zp_05082750.1、yp_001417624.1和yp_353455.1。甚至更特别地,酶e1可以是来自紫色色杆菌dsm30191的转氨酶cv2025。根据本发明的任一方面的酶e1可以相对于seqidno:2具有至少50%序列同一性。更特别地,e1可以包含与多肽seqidno:2具有至少50、60、65、70、75、80、85、90、91、94、95、98或100%的序列同一性的多肽。酶(例如e1)可以包含这样的多肽序列,其中与以下参考序列(登录号)相比最多60%、优选最多25%、特别是最多15%、特别是最多10、9、8、7、6、5、4、3、2或1%的氨基酸残基通过缺失、插入、取代或其组合而被修饰,并且其仍拥有具有相应的以下参考序列的蛋白的活性的至少50%、优选65%、特别是优选80%、特别是大于90%,其中参考蛋白的100%活性被理解为意指用作生物催化剂的细胞的活性(即基于使用的细胞量的每单位时间转化的物质的量(单位/克细胞干重[u/gcdw]))与在参考蛋白不存在下的生物催化剂的活性相比的增加。给定多肽序列的氨基酸残基的修饰(其导致给定多肽的特性和功能没有显著改变)是本领域技术人员已知的。因此,例如许多氨基酸经常可以互相交换而没有问题;此类合适的氨基酸取代的实例是:ala取代为ser;arg取代为lys;asn取代为gln或his;asp取代为glu;cys取代为ser;gln取代为asn;glu取代为asp;gly取代为pro;his取代为asn或gln;ile取代为leu或val;leu取代为met或val;lys取代为arg或gln或glu;met取代为leu或ile;phe取代为met或leu或tyr;ser取代为thr;thr取代为ser;trp取代为tyr;tyr取代为trp或phe;val取代为ile或leu。还已知的是,修饰,特别是以例如氨基酸插入或缺失形式的多肽的n-或c-末端处的修饰,经常对多肽的功能没有显著影响。借助1向和2向蛋白凝胶分离、随后用适当的评估软件光学鉴定凝胶中的蛋白浓度,可测定上述和下述所有的酶和基因的表达。如果酶活性的增加仅仅基于相应基因表达的增加,则可以通过比较野生型和经遗传修饰的细胞之间的1向或2向蛋白分离来简单地确定酶活性增加的定量。用细菌制备蛋白凝胶和鉴定蛋白的常用方法是hermann等人(electrophoresis,22:1712-23(2001))描述的方法。蛋白浓度也可以通过以下分析:用对于待测定的蛋白而言特异性的抗体的western印迹杂交(sambrook等人,molecularcloning:alaboratorymanual,第2版.coldspringharborlaboratorypress,coldspringharbor,n.y.usa,1989),随后用适当软件进行光学评估用于测定浓度(lohaus和meyer(1989)biospektrum,5:32-39;lottspeich(1999),angewandtechemie111:2630-2647)。当由待测定的酶活性催化的反应的可能产物可以在微生物中快速代谢或者野生型中的活性本身对于其太低而不可能足以基于产生形成而测定待测定的酶活性时,该方法也总是一种选项。根据本发明的另一个方面,提供了由作为底物的至少一种羧酸和/或其酯产生至少一种ω-羟基羧酸和/或其酯的方法,所述方法包括-使至少一种表达突变型alkb酶的经遗传修饰的细胞与底物羧酸和/或其酯接触,其中底物羧酸和/或其酯是c6-c14羧酸和/或其酯;且所述突变型alkb酶在alkb的野生型序列中包含至少一个点突变,其中所述点突变在野生型alkb酶的选自v129、l132和t136的氨基酸位置处。特别地,所述点突变可以选自v129t、t136a、v129a和l132v。根据本发明的另一个方面,提供了由作为底物的至少一种羧酸和/或其酯产生至少一种ω-羟基羧酸和/或其酯的方法,所述方法包括-使根据本发明的任一方面的细胞与底物羧酸和/或其酯接触。根据本发明的一个进一步方面,提供了由作为底物的至少一种羧酸和/或其酯产生至少一种ω-氨基羧酸和/或其酯的方法,所述方法包括-使根据本发明的任一方面的细胞与底物羧酸和/或其酯接触。根据本发明的又一个进一步方面,提供了根据本发明的任一方面的细胞用于催化羧酸和/或其酯转化为相应的ω-羟基羧酸和/或其酯和/或ω-氨基羧酸和/或其酯的用途。如本文所用,术语“接触”意指在水溶液中使底物羧酸和/或其酯与根据本发明的任一方面的细胞之间直接接触。例如,所述细胞和羧酸和/或其酯可以不在通过屏障诸如无机膜隔开的不同隔室中。如果所述底物羧酸和/或其酯是可溶的或制备成可溶的并且可以被细胞吸收或可以扩散穿过生物膜,则其可以简单地添加至在水溶液中的根据本发明的任一方面的细胞。在其不充分溶解的情况下,其可以在添加至水溶液中之前溶解于合适的有机溶剂中。本领域技术人员能够通过添加合适的有机和/或极性溶剂来制备溶解性不足的羧酸和/或其酯的水溶液。此类溶剂可以以包含液体有机溶剂的有机相的形式提供。在一个实例中,当在25℃和标准大气压下为液体时,有机溶剂或相可以被认为是液体。在另一个实例中,化合物和催化剂可以在体外,即以或多或少富集或甚至纯化的状态接触,或者可以原位接触,即它们作为细胞代谢的一部分制备,且随后在细胞内反应。术语“水溶液”与术语“水性介质”可互换使用,并且是指包含水的任何溶液,主要是水作为溶剂,其可以用于至少暂时地将根据本发明的任一方面的细胞保持在代谢活性和/或活力状态,并且包含(如果此为必要的)任何额外底物。本领域技术人员熟悉制备许多水溶液,通常称为可用于保持本发明细胞的培养基,例如在大肠杆菌的情况下为lb培养基。有利的是使用作为水溶液的基本培养基,即合理简单组成的培养基,其与复合培养基相比仅包含用于将细胞保持在代谢活性和/或活力状态必不可少的盐和营养物的最小集合,以避免产物被不想要的副产物不必要地污染。例如,m9培养基可以用作基本培养基。附图简述图1a-1d是具有生物转化29小时后hlame和ddame浓度的图。hlame:ω-羟基月桂酸甲酯;ddame:十二烷二酸单甲酯;olame:ω-氧代月桂酸甲酯;lame:月桂酸甲酯。实施例前述描述了优选实施方案,如本领域技术人员将理解,在不背离权利要求的范围的情况下,其可以进行设计、构建或操作的变化或修改。这些变化例如意欲被权利要求的范围所覆盖。实施例1定点诱变。alkb的氨基酸位置v129、l132和t136被认为参与底物识别和底物定位。为了交换alkb酶中的氨基酸,将编码期望氨基酸的相应碱基三联体引入合成的寡核苷酸中。用野生型alkb基因序列作为pcr模板,经由pcr通过使用合成的寡核苷酸来扩增突变型alkb基因(分别编码alkb突变体v129t、v129a和t136a)。pcr产物用作插入dna,用于体外构建表达质粒。核苷酸交换根据本领域技术人员已知的方法经由定点诱变引入,并显示于表1的右栏中。这些交换产生表1的左栏中显示的相应alkb变体。alkb酶变体alkb编码序列中的核苷酸交换野生型alkbseqidno:3v129tg385a,t386c,g387cv129at386c,g387ct136aa406g,a408t表1.改变为产生特定alkb突变体的核苷酸序列。实施例2表达质粒的体外构建作为用于质粒构建的骨架载体,使用质粒pbt10_alkl[对于序列和构建,参见wo/2011/131420的实施例1(wo/2011/131420的seqidno:8)]。在pbt10_alkl中,通过使用来自neb(newenglandbiolabs,inc.)的nebuilder®hifidna组装克隆试剂盒根据制造商的说明书,使用含有突变型alkb基因的pcr产物替代野生型alkb序列。质粒alkb变体转氨酶seqidno:pbt10_alkl野生型-5pbt10_alkl_v129tv129t-6pbt10_alkl_t136at136a-7pbt10_alkl_v129av129a-8pj294_aladh_b.s._ta_c.v.(ct)-cv_20254表2.用于构建大肠杆菌菌株的质粒。对于质粒pj294_aladh_b.s._ta_c.v.(ct)的构建,参见wo/2013/024114的实施例1。实施例3大肠杆菌菌株的构建使用大肠杆菌w3110δfade(无b-氧化)δbioh(无甲酯水解)。使用ep2730655的实施例1中公开的方法制备敲除bioh的表达的大肠杆菌w3110δbioh菌株。然后,随后使用该菌株并使用ep2607490的实施例1中公开的方法来敲除fade的表达。所得菌株被称为大肠杆菌w3110δfadeδbioh。为了构建表达野生型alkb、alkb突变基因(alkb变体)和/或转氨酶的大肠杆菌菌株,经由电穿孔根据本领域技术人员已知的方法,将大肠杆菌菌株w3110δfadeδbioh用作用于转化的宿主。在含有卡那霉素(50μg/ml)和/或氨苄青霉素(100μg/ml)的lb琼脂板上选择转化体。转化体的列表提供于表3中。大肠杆菌菌株质粒1质粒2ecw3110δfadeδbioh_pbt10_alklpbt10_alkl-ecw3110δfadeδbioh_pbt10_alkl_v129tpbt10_alkl_v129t-ecw3110δfadeδbioh_pbt10_alkl_t136apbt10_alkl_t136a-ecw3110δfadeδbioh_pbt10_alkl_v129apbt10_alkl_v129a-ecw3110δfadeδbioh_pbt10_alkl_tapbt10_alklpj294_aladh_b.s._ta_c.v.(ct)ecw3110δfadeδbioh_pbt10_alkl_v129t_tapbt10_alkl_v129tpj294_aladh_b.s._ta_c.v.(ct)ecw3110δfadeδbioh_pbt10_alkl_t136a_tapbt10_alkl_t136apj294_aladh_b.s._ta_c.v.(ct)ecw3110δfadeδbioh_pbt10_alkl_v129a_tapbt10_alkl_v129apj294_aladh_b.s._ta_c.v.(ct)表3.通过用质粒1转化大肠杆菌w3110δfadeδbioh获得的大肠杆菌菌株。经由两种质粒的转化生成具有两种质粒(质粒1和质粒2)的大肠杆菌菌株。实施例4alkb突变体的活性对表3的菌株进行补料分批发酵,以研究它们由月桂酸甲酯(月桂酸甲基酯)产生ω-羟基月桂酸甲酯、ω-氧代月桂酸甲酯和十二烷二酸单甲酯的能力。这在来自dasgip®的8重平行发酵系统中实施。对于发酵,使用1l反应器,其装备有顶置式搅拌器和叶轮涡轮。为了监测过程,在过程期间使用本领域已知的方法直接且连续地测量ph和po2。otr/ctr测量尤其用于估计细胞的代谢活性和适合性。ph探针根据dasgip的技术参考文献用ph4.0和ph7.0的测量溶液借助两点校准来进行校准。根据技术参考文献提供具有所需传感器和连接的反应器,并且安装搅拌器轴。然后用300ml水装填反应器,并在121℃下高压灭菌20min以确保无菌。连接至测量放大器后,将po2探针极化过夜(至少6h)。然后在洁净台下去除水,并通过高细胞密度培养基替换,所述高细胞密度培养基由以下组成:(nh4)2so41.76g/l、k2hpo419.08g/l、kh2po412.5g/l、酵母提取物6.66g/l、柠檬酸三钠二水合物11.2g/l、17ml/l的滤器灭菌的1%强度的柠檬酸铁铵溶液和5ml/l的滤器灭菌的痕量元素储备溶液(其由hcl(37%)36.50g/l、mncl2*4h2o1.91g/l、znso4*7h2o1.87g/l、乙二胺四乙酸二水合物0.84g/l、h3bo30.30g/l、na2moo4*2h2o0.25g/l、cacl2*2h2o4.70g/l、feso4*7h2o17.80g/l、cucl2*2h2o0.15g/l组成),具有15g/l葡萄糖作为碳源(通过计量添加30ml/l无菌补料溶液来添加,所述补料溶液由500g/l葡萄糖、1%(w/v)mgso4*7h2o和2.2%(w/v)nh4cl组成)以及50mg/l卡那霉素。随后,根据技术参考文献,将po2探针使用单点校准(搅拌器:600rpm/通气:10sl/h空气)进行校准至100%,并且借助原地清洁(cleaning-in-place)清洁补料、校正剂(correctionagent)和诱导剂部分(stretches)。为此,将管首先用70%乙醇、随后用1mnaoh、然后用无菌的去矿物质水冲洗,并最终用各种培养基装填。所有前述的大肠杆菌菌株首先从冷冻培养物在具有50mg/l卡那霉素的lb培养基(25ml,在100ml带有挡板的摇瓶中)中在37℃和200rpm下过夜培养约18h。然后,将2ml的该培养物转移至100ml摇瓶中的25ml的高细胞密度培养基中并在37℃/200rpm下再孵育6h以进行第二预培养阶段,所述高细胞密度培养基由以下组成:(nh4)2so41.76g/l、k2hpo419.08g/l、kh2po412.5g/l、酵母提取物6.66g/l、柠檬酸三钠二水合物11.2g/l、17ml/l的滤器灭菌的1%强度的柠檬酸铁铵溶液和5ml/l的滤器灭菌的痕量元素储备溶液(其由hcl(37%)36.50g/l、mncl2*4h2o1.91g/l、znso4*7h2o1.87g/l、乙二胺四乙酸二水合物0.84g/l、h3bo30.30g/l、na2moo4*2h2o0.25g/l、cacl2*2h2o4.70g/l、feso4*7h2o17.80g/l、cucl2*2h2o0.15g/l组成),具有15g/l葡萄糖作为碳源(通过计量添加30ml/l无菌补料溶液来添加,所述补料溶液由500g/l葡萄糖、1%(w/v)mgso4*7h2o和2.2%(w/v)nh4cl组成)以及已描述的抗生素。为了以0.1的光密度接种反应器,测量第二预培养阶段的od600,并计算对于接种所需的培养物的量。借助于5ml注射器将所需量的培养物经隔膜(septum)添加入经热处理且通气的反应器中。使用的标准程序显示于表4中:描述引发物尖峰(triggersharp)31%do(1/60h)诱导dcpk补料开始后10h补料引发物50%do补料速率3[ml/h]表4.用于测定alkb突变体的活性的标准程序。用12.5%强度的氨溶液在一侧将ph调节至ph6.8。在培养和生物转化期间,借助搅拌器补料和通气速率将培养物中的溶解氧(po2或do)调节至至少30%。接种后,do从100%降至该30%,其中其对于发酵的剩余过程中保持稳定。发酵作为补料分批实施,其中经由诱导分批期结束的do峰,用由500g/l葡萄糖、1%(w/v)mgso4*7h2o和2.2%(w/v)nh4cl组成的5g/l*h葡萄糖补料,引发补料开始为进入补料期。随着补料开始,37℃的温度降低至30℃。补料开始后10h,用0.025%(v/v)dcpk诱导氧化基因的表达。补料开始后14h,实施羟基月桂酸甲酯产生的开始(=生物转化的开始)。为此目的,将150ml月桂酸甲酯和油酸的混合物(工业级90%)作为批次添加至发酵液中。结果显示于图1a-1d中。所有突变体都显示与对照相比更快的hlame产生(2-3倍),与对照相比更快的总体氧化(1.5-2倍)和与对照相比所有突变体中的改善的化学选择性(hlame/ddame比)(1.20-1.75相比于0.84)。olame产量在所有菌株中都低(v129a突变体中1g/l,在所有其它菌株中<0.6g/l)。实施例5产物的基于lc-esi/ms2的定量为了定量发酵样品中的lame和hla,在生物转化开始后1/2/4/20/22h取样。制备这些样品用于分析。发酵样品中反应物(educts)、中间体和产物的定量借助lc-esi/ms2参考所有分析物的外部校准(0.1–50mg/l)并使用内标氨基十一烷酸(aud,对于hlame)和d3-lame(对于lame)来实施。使用以下仪器:●具有自动取样器(g1367e)、二元泵(g1312b)和柱温箱(g1316a)的hplc系统1260(agilent;böblingen)●具有esi源的质谱仪tripelquad6410(agilent;böblingen)●hplc柱:kinetexc18,100x2.1mm,粒径:2.6µm,孔径100å(phenomenex;aschaffenburg)●预柱:krudkatcherultrahplc串联滤器;0.5µm滤器深度和0.004mm内径(phenomenex;aschaffenburg)。通过移取1900µl溶剂(80%(v/v)乙腈、20%双蒸h2o(v/v)和0.1%甲酸)和100µl样品至2ml反应容器中来制备样品。将混合物涡旋约10秒,然后以约13000rpm离心5min。使用移液器移取澄清的上清液,并在适当稀释后,用稀释液(80%(v/v)acn、20%双蒸h2o(v/v)和0.1%甲酸)进行分析。将100µl的istd移取至每一900µl样品中(对于90µl的样品体积,10µl)。用上述柱或预柱实施hplc分离。注射体积为0.7µl,柱温为50℃,流速为0.6ml/min。流动相由洗脱液a(0.1%强度(v/v)甲酸水溶液)和洗脱液b(含0.1%(v/v)甲酸的乙腈)组成。使用如表5中所示的梯度概况:表5.hplc分离中使用的梯度概况。用esi源的以下参数以正模式实施esi-ms2分析:●气体温度280℃●气体流速11l/min●雾化压力50psi●毛细管电压4000v。化合物alame、ddame、hlame、olame、lame、di-olame的检测和定量用以下mrm参数进行,在每种情况下一种产物离子用作定性物(qualifier),且一种用作定量物(quantifier)。实施例6具有转氨酶的alkb突变体的活性如实施例5中所述对菌株进行补料分批发酵,以研究它们由月桂酸甲酯(月桂酸甲基酯)产生ω-羟基月桂酸甲酯、ω-氧代月桂酸甲酯和十二烷二酸单甲酯的能力。如实施例4中所述,这在来自dasgip的8倍平行发酵系统中实施。实施例7具有转氨酶的alkb突变体的产物的基于lc-esi/ms2的定量如实施例5中所述实施反应物、中间体和产物的定量的程序。菌株ecw3110δfadeδbioh_pbt10_alkl_v129t_ta、ecw3110δfadeδbioh_pbt10_alkl_t136a_ta和ecw3110δfadeδbioh_pbt10_alkl_v129a_ta比参考菌株ecw3110δfadeδbioh_pbt10_alkl_ta产生相对于ddame更多的alame。当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1