检测AAV的方法与流程
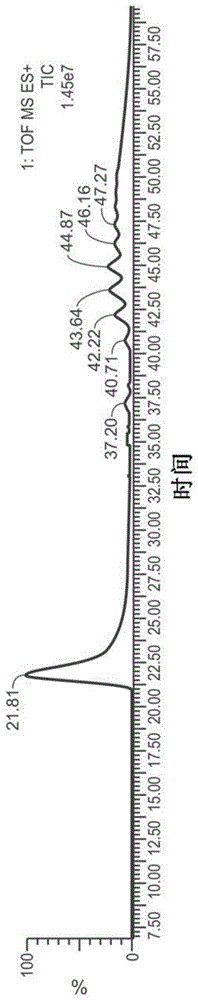
本申请要求2016年8月15日提交的美国临时申请序列号62/375,314的优先权权益,将其通过引用以其整体特此并入。序列表将以下提交的ascii文本文件的内容通过引用以其整体并入本文:计算机可读形式(crf)的序列表(文件名:159792014140seqlist.txt,记录日期:2017年8月14日,大小:51kb)。本发明涉及用于使用质量测定(例如,通过采用液相色谱/质谱(lc/ms)或液相色谱/质谱-质谱(lc/ms/ms))对病毒颗粒(例如,腺相关病毒(aav)颗粒)进行血清分型和/或确定其异质性的方法。在一些方面,本发明涉及改善aav颗粒的稳定性的方法。
背景技术:
:由于病毒衣壳蛋白(vp)对病毒感染性至关重要,因此在基因治疗研究和开发中需要病毒载体(例如,aav载体)的病毒衣壳蛋白的完整表征,包括它们的序列和翻译后修饰。通常使用靶向核酸转基因的分子工具鉴定病毒载体产物,如重组腺相关病毒(raav)产物。这些方法可以包括靶向转基因特异性序列的聚合酶链式反应(pcr)和限制性片段长度多态性(rflp)技术。随着raav技术的发展,许多研究机构开始研究编码其治疗性转基因的多种aav衣壳血清型,以努力改善靶向组织嗜性。传统的分子鉴定方法鉴定含有独特转基因的产物,但不能辨别具有不同aav衣壳血清型的那些产物。目前,大多数aav血清型身份测试基于sds-page显带模式(sds-pagebandingpattern)、基于抗体的elisa或western印迹测定。然而,显带模式和抗体不足以特异性地区分不同的aav血清型。已经报道凝胶-lc/ms/ms作为一种衣壳血清型鉴定方法。然而,这种方法涉及多个步骤,包括sds-page、凝胶内消化和lc/ms/ms,并且因此需要多天进行分析,同时提供有限的序列覆盖。用于鉴定诸如raav载体等载体的方法是基因治疗载体感兴趣的(参见例如,美国专利授予前公开号us20110275529)。因此,具有改善的表征病毒颗粒的方法将是有用的。本文引用的所有参考文献(包括专利申请和出版物)均通过引用以其整体而并入。技术实现要素:使用raav作为例子,本文描述了使用lc/ms作为分析工具以特异性地鉴定不同的病毒衣壳血清型(例如,raav衣壳血清型)。作为病毒表征的一部分,lc/ms可以用于增强分子鉴定方法。这种分析组合可以通过辨别产物的治疗性转基因的身份和衣壳血清型的身份两者来满足监管要求。这种方法可以例如用作aav血清型身份测试或用于监测重组aav基因治疗开发中的病毒衣壳蛋白异质性。它还可以用于确认衣壳工程研究中的vp序列。另外,这种技术可以用于研究翻译后修饰(如病毒衣壳蛋白的n末端乙酰化)对转染效力和细胞内蛋白质运输的影响。本文所述的方法还可以用于设计aav颗粒以获得更高的稳定性和/或改善的转导效率;例如,通过改变aav衣壳的vp1和/或vp3的2位的氨基酸残基,使得与野生型aav衣壳相比,2位的氨基酸被乙酰化至更高的程度。在一些实施方案中,该方法可以用于设计具有降低的转导效率的aav颗粒;例如,通过改变aav衣壳的vp1和/或vp3的2位的氨基酸残基,使得与野生型aav衣壳相比,2位的氨基酸被脱乙酰化至更高或更低的程度。在一些方面,本发明提供了一种确定病毒颗粒的血清型的方法,其包括a)使该病毒颗粒变性,b)使该变性的病毒颗粒经历液相色谱/质谱(lc/ms),并且c)确定该病毒颗粒的一种或多种衣壳蛋白的质量;其中该一种或多种衣壳蛋白的质量的特定组合指示该病毒血清型。在一些实施方案中,将该一种或多种衣壳蛋白的计算质量与一种或多种病毒血清型的该一种或多种衣壳蛋白的理论质量进行比较。在一些方面,本发明提供了一种确定病毒颗粒的异质性的方法,其包括a)使该病毒颗粒变性,b)使该变性的病毒颗粒经历液相色谱/质谱/质谱(lc/ms/ms),c)确定该病毒颗粒的一种或多种衣壳蛋白的质量,并且d)将步骤c)的质量与该病毒血清型的该一种或多种衣壳蛋白的理论质量进行比较;其中该一种或多种衣壳蛋白的质量中的一个或多个的偏差指示病毒衣壳异质性。在一些实施方案中,该异质性包括混合血清型、变体衣壳、衣壳氨基酸取代、截短衣壳或修饰衣壳中的一种或多种。在以上方面的一些实施方案中,该液相色谱是反相液相色谱、尺寸排阻色谱、亲水相互作用液相色谱或阳离子交换色谱。在一些实施方案中,该病毒颗粒包含编码异源转基因的病毒载体。在一些方面,本发明提供了一种确定病毒颗粒的血清型的方法,其包括a)使该病毒颗粒变性,b)使该变性的病毒颗粒经历还原和/或烷基化,c)使该变性的病毒颗粒经历消化以产生该病毒颗粒的一种或多种衣壳蛋白的片段,d)使该一种或多种衣壳蛋白的片段经历液相色谱/质谱-质谱(lc/ms/ms),并且e)确定该病毒颗粒的该一种或多种衣壳蛋白的片段的质量;其中该一种或多种衣壳蛋白的片段的质量的特定组合指示该病毒血清型。在一些实施方案中,将该一种或多种衣壳蛋白的片段的计算质量与一种或多种病毒血清型的该一种或多种衣壳蛋白的片段的理论质量进行比较。在一些方面,本发明提供了一种确定病毒颗粒的血清型的异质性的方法,其包括a)使该病毒颗粒变性,b)使该变性的病毒颗粒经历还原和/或烷基化,c)使该变性的病毒颗粒经历消化以产生该病毒颗粒的一种或多种衣壳蛋白的片段,d)使该一种或多种衣壳蛋白的片段经历液相色谱/质谱-质谱(lc/ms/ms),e)确定该病毒颗粒的该一种或多种衣壳蛋白的片段的质量,并且f)将步骤e)的质量与该病毒血清型的该一种或多种衣壳蛋白的片段的理论质量进行比较;其中该一种或多种衣壳蛋白的质量中的一个或多个的偏差指示病毒衣壳异质性。在一些实施方案中,该异质性包括混合血清型、变体衣壳、衣壳氨基酸取代、截短衣壳或修饰衣壳中的一种或多种。在一些实施方案中,该液相色谱是反相液相色谱、尺寸排阻色谱、亲水相互作用液相色谱或阳离子交换色谱。如本文所示,该方法可以在不存在凝胶分离步骤(例如,十二烷基硫酸钠聚丙烯酰胺凝胶电泳(sds-page))的情况下进行。在以上方面和实施方案的一些实施方案中,该病毒颗粒包含编码异源转基因的病毒载体。在一些实施方案中,该病毒颗粒属于选自以下的病毒科:腺病毒科(adenoviridae)、细小病毒科(parvoviridae)、逆转录病毒科(retroviridae)、杆状病毒科(baculoviridae)和疱疹病毒科(herpesviridae)。在一些实施方案中,该病毒颗粒属于选自以下的病毒属:腺胸腺病毒属(atadenovirus)、禽腺病毒属(aviadenovirus)、美洲白鲟腺病毒属(ichtadenovirus)、哺乳动物腺病毒属(mastadenovirus)、唾液酸酶腺病毒属(siadenovirus)、双义浓核病毒属(ambidensovirus)、短浓核病毒属(brevidensovirus)、虾肝胰腺浓核病毒属(hepandensovirus)、重复浓核病毒属(iteradensovirus)、对虾浓核病毒属(penstyldensovirus)、水貂阿留申细小病毒属(amdoparvovirus)、aveparvovirus、博卡细小病毒属(bocaparvovirus)、copiparvovirus、腺病毒伴随细小病毒属(dependoparvovirus)、嗜红细胞细小病毒属(erythroparvovirus)、protoparvovirus、tetraparvovirus、α逆转录病毒属(alpharetrovirus)、β逆转录病毒属(betaretrovirus)、δ逆转录病毒属(deltaretrovirus)、ε逆转录病毒属(epsilonretrovirus)、γ逆转录病毒属(gammaretrovirus)、慢病毒属(lentivirus)、泡沫病毒属(spumavirus)、α杆状病毒属(alphabaculovirus)、β杆状病毒属(betabaculovirus)、δ杆状病毒属(deltabaculovirus)、γ杆状病毒属(gammabaculovirus)、传喉炎病毒属(iltovirus)、马立克病毒属(mardivirus)、单纯疱疹病毒属(simplexvirus)、水痘病毒属(varicellovirus)、巨细胞病毒属(cytomegalovirus)、鼠巨细胞病毒属(muromegalovirus)、长鼻动物病毒属(proboscivirus)、玫瑰疱疹病毒属(roseolovirus)、淋巴滤泡病毒属(lymphocryptovirus)、玛卡病毒属(macavirus)、马疱疹病毒属(percavirus)和猴病毒属(rhadinovirus)。在一些方面,本发明提供了一种确定腺相关病毒(aav)颗粒的血清型的方法,其包括a)使该aav颗粒变性,b)使该变性的aav颗粒经历液相色谱/质谱(lc/ms),并且c)确定该aav颗粒的vp1、vp2和vp3的质量;其中vp1、vp2和vp3的质量的特定组合指示该aav血清型。在一些实施方案中,将vp1、vp2和vp3的计算质量与一种或多种aav血清型的vp1、vp2和vp3的理论质量进行比较。在一些方面,本发明提供了一种确定aav颗粒的异质性的方法,其包括a)使该aav颗粒变性,b)使该变性的aav颗粒经历液相色谱/质谱/质谱(lc/ms/ms),c)确定该aav颗粒的vp1、vp2和vp3的质量,并且d)将步骤c)的质量与该aav血清型的vp1、vp2和vp3的理论质量进行比较;其中vp1、vp2或vp3的质量中的一个或多个的偏差指示aav衣壳异质性。在一些实施方案中,该异质性包括混合血清型、变体衣壳、衣壳氨基酸取代、截短衣壳或修饰衣壳中的一种或多种。在以上方面和实施方案的一些实施方案中,用乙酸、盐酸胍和/或有机溶剂使该aav颗粒变性。在一些实施方案中,该液相色谱是反相液相色谱、尺寸排阻色谱、亲水相互作用液相色谱或阳离子交换色谱。在一些实施方案中,该液相色谱是反相液相色谱。在一些实施方案中,该反相色谱是c4或c8反向色谱。在一些实施方案中,该色谱使用包含甲酸水溶液的流动相a。在一些实施方案中,该流动相a包含约0.1%甲酸。在一些实施方案中,该色谱包含流动相b,其包含甲酸乙腈溶液。在一些实施方案中,该流动相b包含约0.1%甲酸。在一些实施方案中,该色谱中流动相b的比例随时间增加。在一些实施方案中,该色谱中流动相b的比例以逐步方式增加。在一些实施方案中,流动相b从约10%增加到约20%,从约20%增加到约30%,并且从约30%增加到约38%。在一些实施方案中,流动相b在约6分钟内从约10%增加到约20%,在约10分钟内从约20%增加到约30%,并且在约40分钟内从约30%增加到约38%。在一些实施方案中,该液相色谱是超高效液相色谱(uplc)。在以上方面和实施方案的一些实施方案中,该质谱包括约3.5kv的毛细管电压。在一些实施方案中,该质谱包括约45v的采样锥电压。在一些实施方案中,该质谱包括辅助校准。在一些实施方案中,将碘化钠用作校准物。在以上方面和实施方案的一些实施方案中,vp1和/或vp3的n末端被乙酰化。在一些实施方案中,该aav颗粒是重组aav(raav)颗粒。在一些实施方案中,该aav颗粒包含aav1衣壳、aav2衣壳、aav3衣壳、aav4衣壳、aav5衣壳、aav6衣壳、aav7衣壳、aav8衣壳、aavrh8衣壳、aav9衣壳、aav10衣壳、aavrh10衣壳、aav11衣壳、aav12衣壳、aavlk03衣壳、aav2r471a衣壳、aav2/2-7m8衣壳、aavdj衣壳、aavdj8衣壳、aav2n587a衣壳、aav2e548a衣壳、aav2n708a衣壳、aavv708k衣壳、山羊aav衣壳、aav1/aav2嵌合衣壳、牛aav衣壳或小鼠aav衣壳raav2/hbov1(嵌合aav/人博卡病毒1)。在一些实施方案中,该aav衣壳包含酪氨酸突变或肝素结合突变。在一些实施方案中,将vp1、vp2和vp3的质量与aav1衣壳、aav2衣壳、aav3衣壳、aav4衣壳、aav5衣壳、aav6衣壳、aav7衣壳、aav8衣壳、aavrh8衣壳、aav9衣壳、aav10衣壳、aavrh10衣壳、aav11衣壳、aav12衣壳、aavlk03衣壳、aav2r471a衣壳、aav2/2-7m8衣壳、aavdj衣壳、aavdj8衣壳、aav2n587a衣壳、aav2e548a衣壳、aav2n708a衣壳、aavv708k衣壳、山羊aav衣壳、aav1/aav2嵌合衣壳、牛aav衣壳或小鼠aav衣壳raav2/hbov1(嵌合aav/人博卡病毒1)、aav2hbko衣壳、aavphp.b衣壳或aavphp.eb衣壳中的一种或多种的理论质量进行比较。在以上方面和实施方案的一些实施方案中,该病毒颗粒包含aav1itr、aav2itr、aav3itr、aav4itr、aav5itr、aav6itr、aav7itr、aav8itr、aavrh8itr、aav9itr、aav10itr、aavrh10itr、aav11itr或aav12itr。在一些实施方案中,该aav颗粒包含编码异源转基因的aav载体。在一些方面,本发明提供了一种确定腺相关病毒(aav)颗粒的血清型的方法,其包括a)使该aav颗粒变性,b)使该变性的aav颗粒经历还原和/或烷基化,c)使该变性的aav颗粒经历消化以产生该aav颗粒的vp1、vp2和/或vp3的片段,d)使vp1、vp2和/或vp3的片段经历液相色谱/质谱-质谱(lc/ms/ms),并且e)确定该aav颗粒的vp1、vp2和vp3的片段的质量;其中vp1、vp2和vp3的片段的质量的特定组合指示该aav血清型。在一些实施方案中,将vp1、vp2和/或vp3的片段的计算质量与一种或多种aav血清型的vp1、vp2和/或vp3的片段的理论质量进行比较。在一些方面,本发明提供了一种确定aav颗粒的血清型的异质性的方法,其包括a)使该aav颗粒变性,b)使该变性的aav颗粒经历还原和/或烷基化,c)使该变性的aav颗粒经历消化以产生该aav颗粒的vp1、vp2和/或vp3的片段,d)使vp1、vp2和/或vp3的片段经历液相色谱/质谱-质谱(lc/ms/ms),e)确定该aav颗粒的vp1、vp2和vp3的片段的质量,并且f)将步骤e)的质量与该aav血清型的vp1、vp2和vp3的片段的理论质量进行比较;其中vp1、vp2或vp3的质量中的一个或多个的偏差指示aav衣壳异质性。在一些实施方案中,该异质性包括混合血清型、变体衣壳、衣壳氨基酸取代、截短衣壳或修饰衣壳中的一种或多种。在一些实施方案中,该还原是通过使该aav颗粒经历二硫苏糖醇、β-巯基乙醇或三(2-羧乙基)膦(tcep)来进行。在一些实施方案中,该烷基化是通过使该aav颗粒经历碘乙酸、碘乙酰胺或4-乙烯基吡啶来进行。在一些实施方案中,该消化是酶消化或化学消化。在一些实施方案中,该酶消化是内肽酶消化。在一些实施方案中,该酶消化是胰蛋白酶消化、lysc消化、asp-n消化或glu-c消化。在一些实施方案中,该化学消化是溴化氰消化或酸消化。在一些实施方案中,用乙酸、盐酸胍和/或有机溶剂使该aav颗粒变性。在以上方面和实施方案的一些实施方案中,该液相色谱是反相液相色谱、尺寸排阻色谱、亲水相互作用液相色谱或阳离子交换色谱。在一些实施方案中,该液相色谱是反相液相色谱。在一些实施方案中,该反相色谱是c18反向色谱。在一些实施方案中,该色谱使用包含甲酸水溶液的流动相a。在一些实施方案中,该流动相a包含约0.1%甲酸。在一些实施方案中,该色谱包含流动相b,其包含甲酸乙腈溶液。在一些实施方案中,该流动相b包含约0.1%甲酸。在一些实施方案中,该色谱中流动相b的比例随时间增加。在一些实施方案中,流动相b从约2%增加到约60%。在一些实施方案中,流动相b在约121分钟内从约2%增加到约60%。在一些实施方案中,该液相色谱是高效液相色谱(hplc)。在以上方面和实施方案的一些实施方案中,该质谱包括约3.5kv的毛细管电压。在一些实施方案中,该质谱包括约45v的采样锥电压。在一些实施方案中,该质谱包括辅助校准。在一些实施方案中,将碘化钠用作校准物。在以上方面和实施方案的一些实施方案中,vp1和/或vp3的n末端被乙酰化。在一些实施方案中,该aav颗粒是重组aav(raav)颗粒。在一些实施方案中,该aav颗粒包含aav1衣壳、aav2衣壳、aav3衣壳、aav4衣壳、aav5衣壳、aav6衣壳、aav7衣壳、aav8衣壳、aavrh8衣壳、aav9衣壳、aav10衣壳、aavrh10衣壳、aav11衣壳、aav12衣壳、aavlk03衣壳、aav2r471a衣壳、aav2/2-7m8衣壳、aavdj衣壳、aavdj8衣壳、aav2n587a衣壳、aav2e548a衣壳、aav2n708a衣壳、aavv708k衣壳、山羊aav衣壳、aav1/aav2嵌合衣壳、牛aav衣壳或小鼠aav衣壳raav2/hbov1(嵌合aav/人博卡病毒1)。在一些实施方案中,该aav衣壳包含酪氨酸突变或肝素结合突变。在一些实施方案中,将vp1、vp2和vp3的质量与aav1衣壳、aav2衣壳、aav3衣壳、aav4衣壳、aav5衣壳、aav6衣壳、aav7衣壳、aav8衣壳、aavrh8衣壳、aav9衣壳、aav10衣壳、aavrh10衣壳、aav11衣壳、aav12衣壳、aavlk03衣壳、aav2r471a衣壳、aav2/2-7m8衣壳、aavdj衣壳、aavdj8衣壳、aav2n587a衣壳、aav2e548a衣壳、aav2n708a衣壳、aavv708k衣壳、山羊aav衣壳、aav1/aav2嵌合衣壳、牛aav衣壳或小鼠aav衣壳raav2/hbov1(嵌合aav/人博卡病毒1)中的一种或多种的理论质量进行比较。在以上方面和实施方案的一些实施方案中,该病毒颗粒包含aav1itr、aav2itr、aav3itr、aav4itr、aav5itr、aav6itr、aav7itr、aav8itr、aavrh8itr、aav9itr、aav10itr、aavrh10itr、aav11itr或aav12itr。在一些实施方案中,该aav颗粒包含编码异源转基因的aav载体。在一些实施方案中,本发明提供了一种重组aav(raav)颗粒,其包含在vp1和/或vp3的氨基酸残基2处的氨基酸取代;其中与该亲本aav颗粒的vp1和/或vp3的氨基酸残基2处的n末端乙酰化相比,vp1和/或vp3的氨基酸残基2处的该氨基酸取代改变了n末端乙酰化。在一些实施方案中,该取代导致更高频率的n末端乙酰化或更低频率的n末端乙酰化。在一些实施方案中,该raav颗粒包含在vp1的氨基酸残基2处的氨基酸取代;其中与该亲本aav颗粒的vp1的氨基酸残基2处的n末端乙酰化相比,vp1的氨基酸残基2处的该氨基酸取代改变了n末端乙酰化。在一些实施方案中,该raav颗粒包含在vp3的氨基酸残基2处的氨基酸取代;其中与该亲本aav颗粒的vp3的氨基酸残基2处的n末端乙酰化相比,vp3的氨基酸残基2处的该氨基酸取代改变了n末端乙酰化。在一些实施方案中,氨基酸残基2被cys、ser、thr、val、gly、asn、asp、glu、ile、leu、phe、gln、lys、met、pro或tyr取代。在一些实施方案中,氨基酸残基2被ser、asp或glu取代。在以上方面和实施方案的一些实施方案中,该aav颗粒包含aav1、aav2、aav3、aav4、aav5、aav6、aav7、aav8、aavrh8、aavrh8r、aav9、aav10、aavrh10、aav11、aav12、aavlk03、aav2r471a、aav2/2-7m8、aavdj、aavdj8衣壳、aav2n587a、aav2e548a、aav2n708a、aavv708k、山羊aav、aav1/aav2嵌合体、牛aav、小鼠aav、raav2/hbov1、aav2hbko、aavphp.b或aavphp.eb血清型衣壳。在一些实施方案中,该aav衣壳还包含酪氨酸突变或肝素结合突变。在一些实施方案中,该raav颗粒包含raav载体。在一些实施方案中,该raav载体包含一个或多个aavitr。在一些实施方案中,该raav载体包含aav1itr、aav2itr、aav3itr、aav4itr、aav5itr、aav6itr、aav7itr、aav8itr、aavrh8itr、aav9itr、aav10itr、aavrh10itr、aav11itr或aav12itr。在一些实施方案中,该aav衣壳和该aavitr衍生自相同的血清型。在一些实施方案中,该aav衣壳和该aavitr衍生自不同的血清型。在一些实施方案中,该aav颗粒包含编码侧接一个或多个aavitr的异源转基因的aav载体。在以上方面和实施方案的一些实施方案中,该raav载体是自身互补载体。在一些实施方案中,该raav载体包含编码该转基因的第一核酸序列和编码该转基因的互补序列的第二核酸序列,其中该第一核酸序列可以与该第二核酸序列沿着其大部分或所有长度形成链内碱基对。在一些实施方案中,该第一核酸序列和该第二核酸序列通过突变的aavitr连接,其中该突变的aavitr包含d区的缺失并且包含末端解析(resolution)序列的突变。在以上方面和实施方案的一些实施方案中,该raav颗粒是通过以下方式产生的:用编码该raav载体的核酸和编码aavrep和cap功能的核酸转染宿主细胞,并且提供编码aav辅助功能的核酸。在一些实施方案中,通过用编码该aav辅助功能的核酸转染该宿主细胞来提供该aav辅助功能。在一些实施方案中,通过用提供该aav辅助功能的aav辅助病毒感染该宿主细胞来提供该aav辅助功能。在一些实施方案中,该aav辅助病毒是腺病毒、单纯疱疹病毒或杆状病毒。在一些实施方案中,该raav颗粒由aav生产细胞产生,该aav生产细胞包含编码该raav载体的核酸和编码aavrep和cap功能的核酸并且提供编码aav辅助功能的核酸。在一些实施方案中,该aav生产细胞包含编码aav辅助功能的核酸。在一些实施方案中,通过用提供该aav辅助功能的aav辅助病毒感染该aav生产细胞来提供该aav辅助功能。在一些实施方案中,该aav辅助病毒是腺病毒、单纯疱疹病毒或杆状病毒。在一些实施方案中,该aavcap功能提供了在vp1和/或vp3的氨基酸残基2处的氨基酸取代,其中与该亲本aav颗粒的vp1和/或vp3的氨基酸残基2处的n末端乙酰化相比,vp1和/或vp3的氨基酸残基2处的该氨基酸取代改变了n末端乙酰化。在一些方面,本发明提供了一种包含如本文所述的raav颗粒的药物组合物。在一些方面,本发明提供了一种包含如本文所述的raav颗粒或药物组合物的试剂盒。在一些方面,本发明提供了一种包含如本文所述的raav颗粒或药物组合物的制品。在一些方面,本发明提供了一种aav衣壳蛋白,其包含在亲本aav衣壳蛋白的氨基酸残基2处的氨基酸取代;其中与该亲本aav衣壳蛋白的氨基酸残基2处的n末端乙酰化相比,氨基酸残基2处的该氨基酸取代改变了n末端乙酰化。在一些实施方案中,该取代导致更高频率的n末端乙酰化或更低频率的n末端乙酰化。在一些实施方案中,该aav衣壳蛋白是vp1或vp3。在一些实施方案中,该aav衣壳蛋白的氨基酸残基2被cys、ser、thr、val、gly、asn、asp、glu、ile、leu、phe、gln、lys、met、pro或tyr取代。在一些实施方案中,该aav衣壳蛋白的氨基酸残基2被ser、asp或glu取代。在一些实施方案中,该氨基酸取代导致该aav衣壳的更少脱酰胺化。在以上方面和实施方案的一些实施方案中,该aav颗粒包含aav1、aav2、aav3、aav4、aav5、aav6、aav7、aav8、aavrh8、aavrh8r、aav9、aav10、aavrh10、aav11、aav12、aavlk03、aav2r471a、aav2/2-7m8、aavdj、aavdj8衣壳、aav2n587a、aav2e548a、aav2n708a、aavv708k、山羊aav、aav1/aav2嵌合体、牛aav、小鼠aav、raav2/hbov1、aav2hbko、aavphp.b或aavphp.eb血清型衣壳。在一些实施方案中,该aav衣壳还包含酪氨酸突变或肝素结合突变。在一些方面,本发明提供了一种改善raav颗粒的稳定性的方法,其包括取代亲本vp1和/或vp3的vp1和/或vp3的氨基酸残基2;其中与该亲本vp1和/或vp3的氨基酸残基2相比,该取代氨基酸残基2改变了vp1和/或vp3的n末端乙酰化。在一些方面,本发明提供了一种改善raav颗粒在细胞中的组装的方法,其包括取代vp1和/或vp3或亲本vp1和/或vp3的氨基酸残基2;其中与该亲本vp1和/或vp3的氨基酸残基2相比,取代2位的氨基酸改变了vp1和/或vp3的n末端乙酰化。在一些方面,本发明提供了一种改善raav颗粒在细胞中的转导的方法,其包括取代vp1和/或vp3或亲本vp1和/或vp3的氨基酸残基2;其中与该亲本vp1和/或vp3的氨基酸残基2相比,取代氨基酸残基2改变了vp1和/或vp3的n末端乙酰化。在一些实施方案中,该取代的氨基酸导致更高频率的n末端乙酰化或更低频率的n末端乙酰化。在一些实施方案中,vp1的氨基酸残基2处的该氨基酸取代被取代。在一些实施方案中,vp3的氨基酸残基2处的该氨基酸取代被取代。在一些实施方案中,氨基酸残基2被cys、ser、thr、val、gly、asn、asp、glu、ile、leu、phe、gln、lys、met、pro或tyr取代。在一些实施方案中,氨基酸残基2被ser、asp或glu取代。在一些方面,本发明提供了一种降低raav颗粒在细胞中的转导的方法,其包括取代vp1和/或vp3的氨基酸残基2;其中与该亲本vp1和/或vp3的氨基酸残基2相比,2位的该取代的氨基酸改变了vp1和/或vp3的n末端乙酰化。在以上方面和实施方案的一些实施方案中,该aav颗粒包含aav1、aav2、aav3、aav4、aav5、aav6、aav7、aav8、aavrh8、aavrh8r、aav9、aav10、aavrh10、aav11、aav12、aavlk03、aav2r471a、aav2/2-7m8、aavdj、aavdj8衣壳、aav2n587a、aav2e548a、aav2n708a、aavv708k、山羊aav、aav1/aav2嵌合体、牛aav、小鼠aav、raav2/hbov1、aav2hbko、aavphp.b或aavphp.eb血清型衣壳。在一些实施方案中,该aav衣壳还包含酪氨酸突变或肝素结合突变。在一些实施方案中,该raav颗粒包含raav载体。在一些实施方案中,该raav载体包含一个或多个aavitr。在一些实施方案中,该raav载体包含aav1itr、aav2itr、aav3itr、aav4itr、aav5itr、aav6itr、aav7itr、aav8itr、aavrh8itr、aav9itr、aav10itr、aavrh10itr、aav11itr或aav12itr。在一些实施方案中,该aav衣壳和该aavitr衍生自相同的血清型。在一些实施方案中,该aav衣壳和该aavitr衍生自不同的血清型。在一些实施方案中,该aav颗粒包含编码侧接一个或多个aavitr的异源转基因的aav载体。在以上方面和实施方案的一些实施方案中,该raav载体是自身互补载体。在一些实施方案中,该raav载体包含编码该转基因的第一核酸序列和编码该转基因的互补序列的第二核酸序列,其中该第一核酸序列可以与该第二核酸序列沿着其大部分或所有长度形成链内碱基对。在一些实施方案中,该第一核酸序列和该第二核酸序列通过突变的aavitr连接,其中该突变的aavitr包含d区的缺失并且包含末端解析序列的突变。在以上方面和实施方案的一些实施方案中,该raav颗粒是通过以下方式产生的:用编码该raav载体的核酸和编码aavrep和cap功能的核酸转染宿主细胞,并且提供编码aav辅助功能的核酸。在一些实施方案中,通过用编码该aav辅助功能的核酸转染该宿主细胞来提供该aav辅助功能。在一些实施方案中,通过用提供该aav辅助功能的aav辅助病毒感染该宿主细胞来提供该aav辅助功能。在一些实施方案中,该aav辅助病毒是腺病毒、单纯疱疹病毒或杆状病毒。在一些实施方案中,该raav颗粒由aav生产细胞产生,该aav生产细胞包含编码该raav载体的核酸和编码aavrep和cap功能的核酸并且提供编码aav辅助功能的核酸。在一些实施方案中,该aav生产细胞包含编码aav辅助功能的核酸。在一些实施方案中,通过用提供该aav辅助功能的aav辅助病毒感染该aav生产细胞来提供该aav辅助功能。在一些实施方案中,该aav辅助病毒是腺病毒、单纯疱疹病毒或杆状病毒。在一些实施方案中,该aavcap功能提供了在vp1和/或vp3的氨基酸残基2处的氨基酸取代,其中与该亲本aav颗粒的vp1和/或vp3的氨基酸残基2处的n末端乙酰化相比,vp1和/或vp3的氨基酸残基2处的该氨基酸取代改变了n末端乙酰化。在一些方面,本发明提供了一种重组aav(raav)颗粒,其包含在亲本颗粒的vp1或vp3的氨基酸残基a35、n57、g58、n382、g383、n511、g512、n715或g716处的一个或多个氨基酸取代,残基编号基于aav2的vp1;其中与该亲本aav颗粒的vp1和/或vp3的脱酰胺化相比,该一个或多个氨基酸取代改变了脱酰胺化。在一些实施方案中,该一个或多个氨基酸取代在如下氨基酸残基处:a35、vp1的n57、vp1的g58、vp3的n382、vp3的g383、vp3的n511、vp3的g512、vp3的n715或vp3的g716,并且与该亲本aav颗粒的vp1和/或vp3的脱酰胺化相比,其改变了脱酰胺化。在一些实施方案中,该一个或多个氨基酸取代包含在vp1的n57、vp3的n382、vp3的n511或vp3的n715处用asp取代;并且与该亲本aav颗粒的vp1和/或vp3的脱酰胺化相比,其导致更高频率的脱酰胺化。在一些实施方案中,该一个或多个氨基酸取代包含n57k或n57q取代,并且与该亲本aav颗粒的vp1和/或vp3的脱酰胺化相比,其导致更低频率的脱酰胺化。在一些实施方案中,该一个或多个氨基酸取代包含在vp1的a35处用asp取代,并且与该亲本aav颗粒的vp1的脱酰胺化相比,其导致更高频率的脱酰胺化。在一些实施方案中,该一个或多个氨基酸取代在vp1的g58、vp3的g383、vp3的g512或vp3的g716处,并且与该亲本aav颗粒的vp1和/或vp3的脱酰胺化相比,其导致更低频率的脱酰胺化。在一些实施方案中,vp1的g58被asp取代。在一些实施方案中,该raav颗粒是aav1颗粒或aav2颗粒。在一些方面,本发明提供了包含aav颗粒的药物组合物,该aav颗粒包含在vp1或vp3的氨基酸残基a35、n57、g58、n382、g383、n511、g512、n715或g716处的一个或多个氨基酸取代,残基编号基于aav2的vp1;其中与该亲本aav颗粒的vp1和/或vp3的脱酰胺化相比,该氨基酸取代改变了脱酰胺化。在一些方面,本发明提供了包含aav颗粒或包含aav颗粒的组合物的试剂盒,其中该aav颗粒包含在vp1或vp3的氨基酸残基a35、n57、g58、n382、g383、n511、g512、n715或g716处的一个或多个氨基酸取代,残基编号基于aav2的vp1;其中与该亲本aav颗粒的vp1和/或vp3的脱酰胺化相比,该氨基酸取代改变了脱酰胺化。在一些方面,本发明提供了包含aav颗粒或包含aav颗粒的组合物的制品,其中该aav颗粒包含在vp1或vp3的氨基酸残基a35、n57、g58、n382、g383、n511、g512、n715或g716处的一个或多个氨基酸取代,残基编号基于aav2的vp1;其中与该亲本aav颗粒的vp1和/或vp3的脱酰胺化相比,该氨基酸取代改变了脱酰胺化。在一些方面,本发明提供了一种aav衣壳蛋白,其包含亲本aav衣壳蛋白的氨基酸取代;其中与该亲本aav衣壳蛋白相比,该氨基酸取代改变了该衣壳的脱酰胺化。在一些方面,本发明提供了一种改善raav颗粒的稳定性的方法,其包括取代一个或多个氨基酸残基,其中该一个或多个氨基酸残基是a35、n57、g58、n382、g383、n511、g512、n715或g716,残基编号基于aav2的vp1;其中与该亲本aav颗粒的vp1和/或vp3的脱酰胺化相比,该氨基酸取代改变了脱酰胺化。在一些方面,本发明提供了一种改善raav颗粒在细胞中的组装的方法,其包括取代一个或多个氨基酸残基,其中该一个或多个氨基酸残基是a35、n57、g58、n382、g383、n511、g512、n715或g716,残基编号基于aav2的vp1;其中与该亲本aav颗粒的vp1和/或vp3的脱酰胺化相比,该氨基酸取代改变了脱酰胺化。在一些方面,本发明提供了一种改善raav颗粒在细胞中的转导的方法,其包括取代一个或多个氨基酸残基,其中该一个或多个氨基酸残基是a35、n57、g58、n382、g383、n511、g512、n715或g716,残基编号基于aav2的vp1;其中与该亲本aav颗粒的vp1和/或vp3的脱酰胺化相比,该氨基酸取代改变了脱酰胺化。在一些实施方案中,该一个或多个氨基酸取代在vp1的a35、vp1的n57、vp1的g58、vp3的n382、vp3的g383、vp3的n511、vp3的g512、vp3的n715或vp3的g716处;其中与该亲本aav颗粒的vp1和/或vp3的脱酰胺化相比,该氨基酸取代改变了脱酰胺化。在一些实施方案中,vp1的35位的亲本ala残基被asn取代。在一些实施方案中,vp1的58位的亲本gly残基被asp取代。在一些实施方案中,该raav颗粒是aav1颗粒或aav2颗粒。在一些实施方案中,本发明提供了一种改善raav颗粒的稳定性、组装和/或转导效率的方法,其包括取代一个或多个氨基酸残基,其中该一个或多个氨基酸残基是a35、n57、g58、n382、g383、n511、g512、n715或g716,残基编号基于aav2的vp1;其中如上所述,与该亲本aav颗粒的vp1和/或vp3的脱酰胺化相比,该氨基酸取代改变了脱酰胺化,其中该aav颗粒包含aav1、aav2、aav3、aav4、aav5、aav6、aav7、aav8、aavrh8、aavrh8r、aav9、aav10、aavrh10、aav11、aav12、aavlk03、aav2r471a、aav2/2-7m8、aavdj、aavdj8衣壳、aav2n587a、aav2e548a、aav2n708a、aavv708k、山羊aav、aav1/aav2嵌合体、牛aav、小鼠aav或raav2/hbov1血清型衣壳。在一些实施方案中,该aav衣壳还包含酪氨酸突变或肝素结合突变。在一些实施方案中,该raav颗粒包含raav载体。在一些实施方案中,该raav载体包含一个或多个aavitr。在一些实施方案中,该raav载体包含aav1itr、aav2itr、aav3itr、aav4itr、aav5itr、aav6itr、aav7itr、aav8itr、aavrh8itr、aav9itr、aav10itr、aavrh10itr、aav11itr或aav12itr。在一些实施方案中,该aav衣壳和该aavitr衍生自相同的血清型。在一些实施方案中,该aav衣壳和该aavitr衍生自不同的血清型。在一些实施方案中,该aav颗粒包含编码侧接一个或多个aavitr的异源转基因的aav载体。在以上方面和实施方案的一些实施方案中,该raav载体是自身互补载体。在一些实施方案中,该raav载体包含编码该转基因的第一核酸序列和编码该转基因的互补序列的第二核酸序列,其中该第一核酸序列可以与该第二核酸序列沿着其大部分或所有长度形成链内碱基对。在一些实施方案中,该第一核酸序列和该第二核酸序列通过突变的aavitr连接,其中该突变的aavitr包含d区的缺失并且包含末端解析序列的突变。在以上方面和实施方案的一些实施方案中,该raav颗粒是通过以下方式产生的:用编码该raav载体的核酸和编码aavrep和cap功能的核酸转染宿主细胞,并且提供编码aav辅助功能的核酸。在一些实施方案中,通过用编码该aav辅助功能的核酸转染该宿主细胞来提供该aav辅助功能。在一些实施方案中,通过用提供该aav辅助功能的aav辅助病毒感染该宿主细胞来提供该aav辅助功能。在一些实施方案中,该aav辅助病毒是腺病毒、单纯疱疹病毒或杆状病毒。在一些实施方案中,该raav颗粒由aav生产细胞产生,该aav生产细胞包含编码该raav载体的核酸和编码aavrep和cap功能的核酸并且提供编码aav辅助功能的核酸。在一些实施方案中,该aav生产细胞包含编码aav辅助功能的核酸。在一些实施方案中,通过用提供该aav辅助功能的aav辅助病毒感染该aav生产细胞来提供该aav辅助功能。在一些实施方案中,该aav辅助病毒是腺病毒、单纯疱疹病毒或杆状病毒。在一些实施方案中,该aavcap功能提供了vp1和/或vp3的氨基酸取代,其中与该亲本aav颗粒相比,该氨基酸取代调节该衣壳的脱酰胺化。附图说明图1a-d提供了aav2vp的lc/ms的总离子色谱图。图1a:梯度为1.7%/min的10cm长的behc4柱;图1b:梯度为0.5%/min的10cm长的behc4柱;图1c:梯度为0.5%/min的15cm长的behc4柱;图1d:梯度为0.5%/min的15cm长的behc8柱。图2a和b提供了来自图1d峰1(图2a)和图1d峰2(图2b)的解卷积质谱。图3提供了aav2vp1(seqidno:3)的序列覆盖:绿色,胰蛋白酶肽;蓝色,lys-c肽;粉红色,asp-n肽。图4a-4c提供了aav2vpn末端肽的ms/ms谱。图4a:vp1n末端胰蛋白酶肽a(ac)adgylpdwledtlsegir(seqidno:4);图4b:vp2n末端asp-n肽apgkkrpvehspvep(seqidno:15)。图4c:vp-3n末端asp-n衍生肽a(ac)tgsgapm(seqidno:5)。图5提供了13种aav血清型的序列比对:黑色字母/白色背景:不相似;蓝色字母/蓝色背景:保守;黑色字母/绿色背景:相似块;红色字母/黄色背景:相同;绿色字母/白色背景:弱相似。aavrh10(seqidno:17);aav10(seqidno:18);aav8(seqidno:19);aav7(seqidno:20);aav1(seqidno:21);aav6(seqidno:22);aav2(seqidno:23);aav3(seqidno:24);aav11(seqidno:25);aav12(seqidno:26);aav4(seqidno:27);aav5(seqidno:28);aav9(seqidno:29);共有序列(seqidno:30)。图6a和6b显示了比较通过ttx和pcl方法产生的aav1和aav2颗粒中脱酰胺化的百分比的lc/ms/ms分析的结果。将t9肽ylgpfldk(seqidno:9)用于监测aav1和aav2两者中潜在的脱酰胺化位点n57。图7a和7b显示了比较通过ttx和pcl方法产生的aav1和aav2颗粒中脱酰胺化的百分比的lc/ms/ms分析的结果。将t49肽ynlr(seqidno:11)和yhlr(seqidno:12)分别用于监测aav1和aav2中潜在的脱酰胺化位点n511。图8a和8b显示了比较通过ttx和pcl方法产生的aav1和aav2颗粒中脱酰胺化的百分比的lc/ms/ms分析的结果。将t67肽sanvdftvdnlytepr(seqidno:13)和svnvdftvdtvysepr(seqidno:14)分别用于监测aav1和aav2中潜在的脱酰胺化位点n715。图9显示了aav5脱乙酰化突变型变体的产生和vp1:vp2:vp3比率的sypro蛋白质凝胶分析的结果。图10说明了用于测试aav5脱乙酰化变体的转导效率的体外转导测定。图11显示了如通过载体基因组拷贝/μg蛋白质测量的指示的aav5脱乙酰化变体或亲本未修饰的aav5的细胞进入效率。使用了三种细胞系:293、hela和huh7。图12显示了与用亲本未修饰的aav5转导相比,用指示的aav5脱乙酰化变体转导的细胞的egfp表达(如通过elisa测量的)。使用了三种细胞系:293、hela和huh7。图13提供了13种aav血清型的序列比对,突出显示了aav2中保守的n57g58脱酰胺化位点和a35残基。aavrh10(seqidno:31);aav10(seqidno:31);aav8(seqidno:32);aav7(seqidno:33);aav1(seqidno:31);aav6(seqidno:31);aav2(seqidno:34);aav3(seqidno:35);aav11(seqidno:31);aav12(seqidno:36);aav4(seqidno:37);aav5(seqidno:38);aav9(seqidno:39);共有序列(seqidno:40)。图14显示了来自通过pcl或ttx方法产生的aav1或aav2颗粒的vp1、vp2和vp3衣壳蛋白的蛋白质凝胶。*突出显示了截短的vp1(tvp1)蛋白。图15显示了与对照aav2衣壳相比,指示的aav2突变体的脱酰胺化的lc/ms分析的结果。图16显示了aav2脱酰胺化突变型变体的产生和vp1:vp2:vp3比率的sypro蛋白质凝胶分析的结果。图17说明了用于测试aav2脱酰胺化变体的转导效率的体外转导测定。图18显示了如通过载体基因组拷贝/μg蛋白质测量的指示的aav2脱酰胺化变体或亲本未修饰的aav2的细胞进入效率。使用了三种细胞系:293、hela和huh7。图19显示了与用亲本未修饰的aav2转导相比,用指示的aav2脱酰胺化变体转导的细胞的egfp表达(如通过elisa测量的)。使用了三种细胞系:293、hela和huh7。具体实施方式在一些方面,本发明提供了确定一个或多个腺相关病毒(aav)颗粒的血清型的方法,其包括:a)使aav颗粒变性,b)使变性的aav颗粒经历液相色谱/质谱(lc/ms),并且c)确定aav颗粒的vp1、vp2和vp3的质量;其中vp1、vp2和vp3的质量的特定组合指示aav血清型。在其他方面,本发明提供了确定aav颗粒的异质性的方法,其包括:a)使aav颗粒变性,b)使变性的aav颗粒经历液相色谱/质谱(lc/ms),并且c)确定aav颗粒的vp1、vp2和vp3的质量,并将步骤c)的质量与aav血清型的vp1、vp2和vp3的理论质量进行比较;其中vp1、vp2或vp3的质量中的一个或多个的偏差指示aav衣壳异质性。在其他方面,本发明提供了确定腺相关病毒(aav)颗粒的血清型的方法,其包括a)使aav颗粒变性,b)使变性的aav颗粒经历还原和/或烷基化,c)使变性的aav颗粒经历消化以产生aav颗粒的vp1、vp2和/或vp3的片段,d)使vp1、vp2和/或vp3的片段经历液相色谱/质谱-质谱(lc/ms/ms),并且e)确定aav颗粒的vp1、vp2和vp3的片段的质量;其中vp1、vp2和vp3的片段的质量的特定组合指示aav血清型。在其他方面,本发明提供了确定某一血清型的aav颗粒的异质性的方法,其包括:a)使aav颗粒变性,b)使变性的aav颗粒经历还原和/或烷基化,c)使变性的aav颗粒经历消化以产生aav颗粒的vp1、vp2和/或vp3的片段,d)使vp1、vp2和/或vp3的片段经历液相色谱/质谱-质谱(lc/ms/ms),e)确定aav颗粒的vp1、vp2和vp3的片段的质量,并且f)将步骤e)的质量与aav血清型的vp1、vp2和vp3的片段的理论质量进行比较;其中vp1、vp2或vp3的质量中的一个或多个的偏差指示aav衣壳异质性。在一些方面,本发明提供了重组aav(raav)颗粒,其包含在vp1和/或vp3的氨基酸残基2处的氨基酸取代;其中与亲本aav颗粒的vp1和/或vp3的氨基酸残基2处的n末端乙酰化相比,vp1和/或vp3的氨基酸残基2处的氨基酸取代改变了n末端乙酰化。在一些方面,本发明提供了改善raav颗粒在细胞中的组装的方法,其包括取代vp1和/或vp3的氨基酸残基2;其中2位的取代的氨基酸以比亲本vp1和/或vp3的氨基酸残基2更高的频率被n-乙酰化。在一些方面,本发明提供了改善raav颗粒在细胞中的转导的方法,其包括取代vp1和/或vp3的氨基酸残基2;其中2位的取代的氨基酸以比亲本vp1和/或vp3的氨基酸残基2更高的频率被n-乙酰化。i.通用技术本领域技术人员通常很好地理解并且通常使用常规方法来采用本文描述或引用的技术和程序,例如像描述在以下文献中的广泛使用的方法:molecularcloning:alaboratorymanual(sambrook等人,第4版,coldspringharborlaboratorypress,coldspringharbor,n.y.,2012);currentprotocolsinmolecularbiology(f.m.ausubel等人编辑,2003);系列丛书methodsinenzymology(academicpress,inc.);pcr2:apracticalapproach(m.j.macpherson,b.d.hames和g.r.taylor编辑,1995);antibodies,alaboratorymanual(harlow和lane编辑,1988);cultureofanimalcells:amanualofbasictechniqueandspecializedapplications(r.i.freshney,第6版,j.wileyandsons,2010);oligonucleotidesynthesis(m.j.gait编辑,1984);methodsinmolecularbiology,humanapress;cellbiology:alaboratorynotebook(j.e.cellis编辑,academicpress,1998);introductiontocellandtissueculture(j.p.mather和p.e.roberts,plenumpress,1998);cellandtissueculture:laboratoryprocedures(a.doyle,j.b.griffiths和d.g.newell编辑,j.wileyandsons,1993-8);handbookofexperimentalimmunology(d.m.weir和c.c.blackwell编辑,1996);genetransfervectorsformammaliancells(j.m.miller和m.p.calos编辑,1987);pcr:thepolymerasechainreaction(mullis等人编辑,1994);currentprotocolsinimmunology(j.e.coligan等人编辑,1991);shortprotocolsinmolecularbiology(ausubel等人编辑,j.wileyandsons,2002);immunobiology(c.a.janeway等人,2004);antibodies(p.finch,1997);antibodies:apracticalapproach(d.catty.编辑,irlpress,1988-1989);monoclonalantibodies:apracticalapproach(p.shepherd和c.dean编辑,oxforduniversitypress,2000);usingantibodies:alaboratorymanual(e.harlow和d.lane,coldspringharborlaboratorypress,1999);theantibodies(m.zanetti和j.d.capra编辑,harwoodacademicpublishers,1995);和cancer:principlesandpracticeofoncology(v.t.devita等人编辑,j.b.lippincottcompany,2011)。ii.定义如本文所用的“载体”是指包含有待在体外或在体内递送至宿主细胞中的核酸的重组质粒或病毒。如本文所用的术语“多核苷酸”或“核酸”是指任何长度的核苷酸(核糖核苷酸或脱氧核糖核苷酸)的聚合形式。因此,此术语包括但不限于单链、双链或多链dna或rna,基因组dna,cdna,dna-rna杂合体,或包含嘌呤和嘧啶碱基或其他天然的、化学或生物化学修饰的、非天然的或衍生的核苷酸碱基的聚合物。核酸的骨架可以包含糖和磷酸基团(如通常可以在rna或dna中发现的)或者修饰或取代的糖或磷酸基团。可替代地,核酸的骨架可以包含合成亚基(如氨基磷酸酯)的聚合物,并且因此可以是寡脱氧核苷氨基磷酸酯(p-nh2)或混合的氨基磷酸酯-磷酸二酯寡聚物。另外,双链核酸可以从化学合成的单链多核苷酸产物,通过合成互补链并在适当的条件下使链退火或者通过使用dna聚合酶用适当的引物从头合成互补链来获得。术语“多肽”和“蛋白质”可互换地使用以指代氨基酸残基的聚合物,并且不限于最小长度。氨基酸残基的此类聚合物可以含有天然或非天然氨基酸残基,并且包括但不限于氨基酸残基的肽、寡肽、二聚体、三聚体和多聚体。该定义涵盖了全长蛋白质及其片段两者。该术语还包括多肽的翻译后修饰,例如糖基化、唾液酸化、乙酰化、磷酸化等。此外,用于本发明的目的,“多肽”是指相对于天然序列包括修饰(如缺失、添加和取代)的蛋白质(通常在本质上是保守的),只要该蛋白质保持所需活性即可。这些修饰可能是故意的(如通过定点诱变),或者可能是偶然的(如通过产生蛋白质的宿主的突变或由于pcr扩增引起的错误)。“重组病毒载体”是指包含一个或多个异源序列(即,不是病毒来源的核酸序列)的重组多核苷酸载体。在重组aav载体的情况下,重组核酸侧接至少一个(例如,两个)反向末端重复序列(itr)。“重组aav载体(raav载体)”是指包含侧接至少一个(例如,两个)aav反向末端重复序列(itr)的一个或多个异源序列(即,不是aav来源的核酸序列)的多核苷酸载体。当此类raav载体存在于已感染合适的辅助病毒(或表达合适的辅助功能)并且表达aavrep和cap基因产物(即aavrep和cap蛋白)的宿主细胞中时,它们可以被复制并包装在感染性病毒颗粒中。当将raav载体掺入较大多核苷酸(例如,在用于克隆或感染的染色体或另一种载体如质粒中)中时,则raav载体可以被称为“前载体”,其可以通过在aav包装功能和合适辅助功能的存在下复制和衣壳化被“挽救”。raav载体可以呈多种形式中的任何一种,包括但不限于质粒、线性人工染色体、与脂质复合、包封在脂质体内和(在多个实施方案中)衣壳化于病毒颗粒(特别是aav颗粒)中。raav载体可以被包装在aav病毒衣壳中,以产生“重组腺相关病毒颗粒(raav颗粒)”。“raav病毒”或“raav病毒颗粒”是指由至少一种aav衣壳蛋白和衣壳化的raav载体基因组组成的病毒颗粒。在比较n-乙酰化和/或脱酰胺化的背景下,如本文所用的“亲本aav颗粒”和“亲本aav衣壳蛋白”是指向其中引入氨基酸修饰以调节n-乙酰化和/或脱酰胺化的aav颗粒或衣壳蛋白(例如,与本发明的aav颗粒/衣壳相同或相似,但不包含如本文所述的调节/改变n-乙酰化和/或脱酰胺化的突变的aav颗粒/衣壳蛋白)。在一些实施方案中,亲本aav颗粒是包含重组aav基因组的重组aav颗粒。在一些实施方案中,亲本aav衣壳颗粒或亲本aav衣壳蛋白包含影响aav颗粒的其他方面的氨基酸取代。例如,亲本aav颗粒可以包含影响aav与其受体的结合(如影响aav2与硫酸肝素蛋白多糖的结合(例如aav2hbko颗粒))的氨基酸取代。可以突变aav2hbko颗粒以引入调节n-乙酰化和/或脱酰胺化的氨基酸取代。然后可以将这样的突变的aav颗粒与如本文所述的本发明方面的亲本aav2hbko颗粒进行比较。亲本aav衣壳蛋白可以包括亲本vp1衣壳蛋白、亲本vp2衣壳蛋白或vp3衣壳蛋白。如本文所用,关于亲本分子的术语“调节”或“改变”意指改变亲本分子的特征。例如,与亲本aav颗粒相比,具有改变的n-乙酰化的aav颗粒可以显示出增加或减少的n-乙酰化;并且与亲本aav颗粒相比,具有改变的脱酰胺化的aav颗粒可以显示增加或减少的脱酰胺化。“异源的”意指衍生自基因型不同于其所比较或其所引入或掺入的实体的剩余部分的实体。例如,通过基因工程技术引入不同细胞类型中的核酸是异源核酸(并且在表达时可以编码异源多肽)。类似地,掺入病毒载体中的细胞序列(例如,基因或其部分)就该载体而言是异源核苷酸序列。术语“转基因”是指引入细胞中并且能够被转录成rna并且任选地在适当条件下翻译和/或表达的核酸。在多个方面,它赋予其所引入的细胞所需的特性,或以其他方式产生所需的治疗或诊断结果。在另一个方面,它可以被转录成介导rna干扰的分子,如sirna。如关于病毒滴度使用的术语“基因组颗粒(gp)”、“基因组等同物”或“基因组拷贝”是指含重组aavdna基因组的病毒粒子的数量,与感染性或功能性无关。特定载体制剂中的基因组颗粒的数量可以通过诸如本文实施例中或例如clark等人(1999)hum.genether.,10:1031-1039;veldwijk等人(2002)mol.ther.,6:272-278中所述的程序测量。如关于病毒滴度使用的术语“感染单位(iu)”、“感染性颗粒”或“复制单位”是指感染性和复制型重组aav载体颗粒的数量,如通过感染中心测定(也称为复制中心测定)测量的,如例如mclaughlin等人(1988)j.virol.,62:1963-1973中所述。如关于病毒滴度使用的术语“转导单位(tu)”是指导致产生功能转基因产物的感染性重组aav载体颗粒的数量,如在功能测定中测量的,如本文实施例中或例如xiao等人(1997)exp.neurobiol.,144:113-124中;或fisher等人(1996)j.virol.,70:520-532(lfu测定)中所述。“反向末端重复”或“itr”序列是本领域中熟知的术语,并且是指在病毒基因组末端发现的相对较短的序列,它们的方向相反。“aav反向末端重复(itr)”序列为本领域中熟知的术语,是存在于天然单链aav基因组的两端的大约145个核苷酸的序列。itr的最外侧的125个核苷酸能以两个替代方向中的任何一个存在,导致不同aav基因组之间以及单个aav基因组两端之间的异质性。最外侧的125个核苷酸也含有几个较短的自身互补的区域(指定为a、a'、b、b'、c、c’和d区),允许在itr的这个部分内发生链内碱基配对。“末端解析序列”或“trs”是aavitr的d区中的序列,其在病毒dna复制期间被aavrep蛋白切割。突变型末端解析序列难以被aavrep蛋白切割。“aav辅助功能”是指允许aav被宿主细胞复制和包装的功能。aav辅助功能可以按多种形式中的任何一种提供,包括但不限于协助aav复制和包装的辅助病毒或辅助病毒基因。其他aav辅助功能在本领域中是已知的,如基因毒性剂。“aav辅助功能”是指允许aav被宿主细胞复制和包装的功能。aav辅助功能可以按多种形式中的任何一种提供,包括但不限于协助aav复制和包装的辅助病毒或辅助病毒基因。其他aav辅助功能在本领域中是已知的,如基因毒性剂。aav的“辅助病毒”是指允许aav(其是缺陷型细小病毒)被宿主细胞复制和包装的病毒。已经鉴定了多种此类辅助病毒,包括腺病毒、疱疹病毒、痘病毒(如牛痘)和杆状病毒。腺病毒涵盖多种不同子群,但子群c的5型腺病毒(ad5)是最常用的。人类、非人类哺乳动物和鸟类来源的许多腺病毒是已知的,并且可从诸如atcc等保藏机构获得。也可从诸如atcc等保藏机构获得的疱疹家族病毒包括例如单纯疱疹病毒(hsv)、爱泼斯坦-巴尔(epstein-barr)病毒(ebv)、巨细胞病毒(cmv)和假狂犬病病毒(prv)。可从保藏机构获得的杆状病毒包括苜蓿银纹夜蛾(autographacalifornica)核型多角体病毒。将就参考多肽或核酸序列而言的“序列同一性百分比(%)”定义为在用以实现最大百分比序列同一性并且不将任何保守取代视为序列同一性的一部分而比对序列和引入缺口(如果需要)后,候选序列中与参考多肽或核酸序列中的氨基酸残基或核苷酸相同的氨基酸残基或核苷酸的百分比。用于确定氨基酸或核酸序列同一性百分比的目的的比对可以按在本领域技术范围内的多种方式实现,例如使用可公开获得的计算机软件程序,例如currentprotocolsinmolecularbiology(ausubel等人编辑,1987),增刊30,第7.7.18章,表7.7.1中描述的那些,并且包括blast、blast-2、align或megalign(dnastar)软件。潜在的比对程序是alignplus(scientificandeducationalsoftware,宾夕法尼亚州)。本领域技术人员可以确定用于测量比对的适当参数,包括为了在被比较的序列的全长上实现最大比对所需要的任何算法。用于本文的目的,给定氨基酸序列a对、与或相对于给定氨基酸序列b的氨基酸序列同一性%(或者这可以用短语表示为对、与或相对于给定氨基酸序列b具有或包含某一氨基酸序列同一性%的给定氨基酸序列a)计算如下:100x分数x/y,其中x是在a和b的程序比对中通过序列比对程序评定为完全匹配的氨基酸残基的数量,并且其中y是b中氨基酸残基的总数量。应理解,在氨基酸序列a的长度不等于氨基酸序列b的长度的情况下,a与b的氨基酸序列同一性%将不等于b与a的氨基酸序列同一性%。用于本文的目的,给定核酸序列c对、与或相对于给定核酸序列d的核酸序列同一性%(或者这可以用短语表示为对、与或相对于给定核酸序列d具有或包含某一核酸序列同一性%的给定核酸序列c)计算如下:100x分数w/z,其中w是在c和d的程序比对中通过序列比对程序评定为完全匹配的核苷酸的数量,并且其中z是d中核苷酸的总数量。应理解,在核酸序列c的长度不等于核酸序列d的长度的情况下,c与d的核酸序列同一性%将不等于d与c的核酸序列同一性%。“分离的”分子(例如,核酸或蛋白质)或细胞意指已经从其天然环境的组分中鉴别并分离和/或回收。“质谱”是指通过测量气相离子的质荷比和丰度来鉴定化合物(例如,多肽)的量和/或类型的分析化学技术。术语“质谱”在本文中可互换地使用。当关于aav衣壳使用时,“异质性”是指如下aav衣壳,其特征在于观察到一种或多种衣壳多肽偏离vp1、vp2和/或vp3多肽或其片段的参考质量。参考质量可以包括但不限于例如已知aav血清型的vp1、vp2和/或vp3多肽的理论、预测或预期质量。例如,如果aav衣壳表现出一种或多种以下特性(无限制),则可以说它显示异质性:混合血清型、变体衣壳、衣壳氨基酸取代、截短衣壳或修饰衣壳。本文对“约”某一值或参数的提及包括(并描述)针对该值或参数本身的实施方案。例如,涉及“约x”的描述包括“x”的描述。除非另外说明,否则如本文所用,冠词的单数形式“一个”、“一种”和“该”包括复数指示物。应理解,本文所述的本发明的方面和实施方案包括“包含多个方面和实施方案”、“由多个方面和实施方案组成”和/或“基本上由多个方面和实施方案组成”。iii.方法本公开文本的某些方面涉及确定病毒颗粒的血清型的方法。本公开文本的其他方面涉及确定病毒颗粒的异质性的方法。如下所述,每种aav血清型的vp1、vp2和vp3的精确质量是独特的,并且可以用于鉴定或区分aav衣壳血清型。这些方法部分地基于本文所述的发现,即在变性后不同类型的aav的直接lc/ms可以用于用完整蛋白质水平的精确质量测量来监测蛋白质序列和翻译后修饰。此外,还可以在不同的aav血清型中鉴定和/或监测vp1和vp3的n末端的乙酰化。基于这些aav结果和本文提供的指导,预期此类方法可以容易地应用于分析(profile)多种病毒,例如本公开文本的病毒家族、亚家族和属。本公开文本的方法可以例如用于分析vp以监测vp表达、翻译后修饰和截短,并且以确保vlp生产期间的产物一致性,以确认定点诱变或结构表征用于衣壳蛋白工程应用,和/或以监测或检测病毒颗粒或制剂的异质性。在一些实施方案中,该方法包括使病毒颗粒变性。在一些实施方案中,可以使用洗涤剂、热、高盐或具有低或高ph的缓冲使病毒颗粒(如aav颗粒)变性。在某些实施方案中,可以使用乙酸或盐酸胍使aav颗粒变性。技术人员应认识到,可用于促进和/或监测蛋白质变性的多种方法在本领域中是可获得的,并且可以适当地选择与液相色谱/质谱相容的变性方法。例如,如果使用热变性,可以小心避免蛋白质沉淀和反相柱堵塞。类似地,高盐变性可以在lc/ms或lc/ms/ms之前与脱盐步骤结合。在其他实施方案中,使用高ph变性、低ph变性或使用有机溶剂变性。在一些实施方案中,该方法包括使本公开文本的变性的病毒颗粒经历液相色谱/质谱(lc/ms)。如在本领域中已知的,lc/ms利用液相色谱进行离子的物理分离,并且利用质谱从离子产生质谱数据。此类质谱数据可以用于确定例如分子量或结构,按质量、数量、纯度等鉴定颗粒。这些数据可以表示检测到的离子的特性,如随时间(例如,保留时间)的信号强度(例如,丰度)或随质荷比的相对丰度。在一些实施方案中,液相色谱(例如,如本文所述的lc/ms中使用的)是超高效液相色谱(uplc;术语“超高效液相色谱”或uhplc在本文中可互换地使用)。uplc在本领域中作为如下lc技术而已知,其依赖于具有减小的粒度(例如,小于2μm)和增加的流速的柱以改善色谱分辨率、效率、峰容量和灵敏度(参见例如,plumb,r.等人(2004)rapidcommun.massspectrom.18:2331-2337)。在一些实施方案中,uplc是指在液相色谱中使用粒度小于2μm的柱。在一些实施方案中,uplc是指在液相色谱中使用高线性溶剂速度(例如,如当在6000psi或更高压力下操作时观察到的)。示例性uplc机器是可商购的(例如,来自马萨诸塞州米尔福德的waters的acquity)。在一些实施方案中,质谱(例如,如本文所述的lc/ms中使用的)可以指电喷雾电离质谱(esi-ms)。esi-ms在本领域中作为如下技术而已知,其使用电能来使用质谱分析来源于溶液的离子(参见例如,yamashita,m.和fenn,j.b.(1984)j.phys.chem.88:4451-4459)。离子物质(或在溶液中或在气相中电离的中性物质)通过以下方式从溶液转移到气相:在带电液滴的气溶胶中扩散,然后进行减小带电液滴的尺寸的溶剂蒸发,并且当溶液藉由相对于地面(例如,周围室的壁)的电压通过小毛细管时从电荷液滴进行样品离子喷射。在一些实施方案中,毛细管电压从约2kv至约10kv或约2.5kv至约6.0kv。在某些实施方案中,液相色谱(例如,如本文所述的lc/ms中使用的)使用约3.5kv的毛细管电压。在一些实施方案中,毛细管电压的范围从约1kv至约10kv。在其他实施方案中,质谱(例如,如本文所述的lc/ms中使用的)可以指基质辅助激光解吸/电离(maldi)。在一些实施方案中,质谱(例如,如本文所述的lc/ms中使用的)使用采样锥和/或撇浮器,离子可以在进入分析器之前通过该采样锥和/或撇浮器。在一些实施方案中,例如当如上所述向毛细管施加电压时,样品锥保持在比毛细管电压低的电压。在某些实施方案中,液相色谱(例如,如本文所述的lc/ms中使用的)使用约45v的采样锥电压。在一些实施方案中,采样锥电压的范围从约0v至约200v。在一些实施方案中,质谱(例如,如本文所述的lc/ms中使用的)使用辅助校准。当关于质谱使用时,校准可以包括引入一种或多种具有已知质量的化合物(例如,标准物),用于就质量检测(例如,m/z测量)而言校准仪器的目的。在一些实施方案中,辅助校准可以指使用软件将已知标准物(例如,校准物)的峰和/或位置与特定质荷(m/z)比相关联。一旦校准,使用者然后可以在一定的精确度或误差度和/或所需的再现性水平(例如,与先前或已知的实验条件相比)内对具有一种或多种未知化合物或化合物以未知浓度存在的样品进行质谱。多种校准物在本领域中是已知的,包括但不限于碘化钠、碘化钠铯、聚乙二醇和全氟三丁胺。在某些实施方案中,将碘化钠用作校准物。在一些实施方案中,校准物是glu-1-纤维蛋白肽b和亮氨酸脑啡肽,以在lc/ms操作期间锁定质量。在一些实施方案中,该方法包括使本公开文本的变性的病毒颗粒经历或使本公开文本的变性的病毒颗粒的消化片段经历液相色谱/质谱-质谱(lc/ms/ms)。如在本领域中已知的,lc/ms/ms(术语“液相色谱-串联质谱”在本文中可互换地使用)利用液相色谱进行离子的物理分离,并且利用质谱从离子产生质谱数据,其中质谱使用多个质量(例如,m/z)分离阶段,通常通过片段化步骤来分离。例如,一定m/z范围内的感兴趣的离子可以在第一轮ms中分离出来,片段化,并且然后在第二轮ms中基于个体m/z进一步分离。离子片段化可以包括但不限于诸如碰撞诱导解离(cid)、高能碰撞解离(hcd)、电子捕获解离(ecd)或电子转移解离(etd)等技术。在一些实施方案中,该方法包括使本公开文本的变性的病毒颗粒经历还原和/或烷基化。减少病毒颗粒的手段包括但不限于用二硫苏糖醇、β-巯基乙醇或三(2-羧乙基)膦(tcep)处理。烷基化病毒颗粒的手段包括但不限于用碘乙酸、碘乙酰胺或4-乙烯基吡啶处理aav颗粒。在一些实施方案中,该方法包括使本公开文本的变性的病毒颗粒经历消化,例如以产生aav颗粒的vp1、vp2和/或vp3的片段。例如,可以使变性的aav颗粒经历消化以产生肽片段,其可以例如使用lc(用于分离)和ms/ms(用于分析)来分析(关于更多描述参见下文)。在一些实施方案中,消化是酶消化。在一些实施方案中,消化使用化学消化,如仪器片段化的cnbr处理(例如,自上而下)。在一些实施方案中,消化使用化学消化,如酸消化。在一些实施方案中,酶消化是内肽酶消化。内肽酶可以包括催化多肽的非末端氨基酸的肽键的蛋白水解的任何肽酶。已知的内肽酶可以包括但不限于胰蛋白酶、胰凝乳蛋白酶、aspn、glu-c、lysc、胃蛋白酶、嗜热菌蛋白酶、谷氨酰内肽酶、弹性蛋白酶和脑啡肽酶。在某些实施方案中,酶消化是胰蛋白酶消化或lysc消化。在一些实施方案中,液相色谱(例如,在本文所述的lc/ms或lc/ms/ms中使用的)是反相液相色谱(术语“反相液相色谱(reversedphaseliquidchromatography)”或rplc在本文中可与反相液相色谱互换使用)。如在本领域中已知的,反相液相色可以指使用疏水性固定相(例如,支持体或基体如柱)的色谱分离,以吸附极性流动相中的疏水分子。通过降低流动相的极性(例如,通过添加有机溶剂),可以通过疏水性实现分子的梯度分离,因为由于与柱的更强的疏水相互作用,更具疏水性的分子将在更高浓度的有机溶剂中留在柱上。在一些实施方案中,分离是通过以下来进行:毛细管电泳(ce)、尺寸排阻色谱(sec)、离子交换色谱(iec)(如阳离子交换色谱)、疏水相互作用色谱(hic)、亲水相互作用液相色谱(hilic)、但不限于在线lc/ms(如ms之前的离线分离);例如,尖端、柱、板或筒。通常,适用于反相液相色谱的固定相(例如,疏水部分)可以与支持体偶联,该支持体包括但不限于填充有颗粒或珠粒(例如,多孔二氧化硅颗粒或聚苯乙烯)的柱或树脂。多种疏水性固定相在本领域中是已知的,包括但不限于疏水烷基链、辛基或十八烷基甲硅烷基部分、氰基部分和氨基部分。在一些实施方案中,固定相可以包括特定长度(如c4、c8或c18)的疏水烷基链。在某些实施方案中,反相色谱是c4或c8反相色谱(例如,利用c4或c8固定相的反相色谱)。本领域技术人员可以基于感兴趣的分子(例如,变性的aav颗粒或其片段)适当地选择固定相。适用于反相液相色谱的多种流动相在本领域中是已知的。如上所述,反相液相色谱流动相可以包括有机(例如,疏水)和水性(例如,极性)溶剂的混合物。增加有机溶剂的比例会增加其从固定相中洗脱疏水化合物的能力。可以例如通过改变固定相的类型或暴露、添加极性试剂(如封端试剂)、改变温度和/或改变流动相特征(如有机溶剂的比例、ph、缓冲液和所用有机溶剂的类型)来改变化合物保留和/或选择性。在一些实施方案中,流动相的极性组分可以包括但不限于水或水性缓冲液。在一些实施方案中,流动相的极性组分可以包括但不限于乙腈、甲醇、乙醇、异丙醇、四氢呋喃(thf)和甲酸。在一些实施方案中,可以按感兴趣的梯度或比例使用两种或更多种流动相(例如,流动相a、流动相b等)。在某些实施方案中,色谱使用包含甲酸水溶液的流动相a。在某些实施方案中,流动相a包含约0.1%甲酸。在某些实施方案中,流动相a包含约0.1%至约5%甲酸。在某些实施方案中,色谱使用包含甲酸乙腈溶液的流动相b。在某些实施方案中,流动相b包含约0.1%甲酸。在一些实施方案中,色谱中流动相b的比例随时间增加。例如,色谱中流动相b的比例能以逐步方式增加。在某些实施方案中,流动相b从约10%增加到约20%,从约20%增加到约30%,并且从约30%增加到约38%。在其他实施方案中,流动相b从约2%增加到约60%。在其他实施方案中,流动相b从约1min至约200min从约2%增加到约100%。在一些实施方案中,流动相的其余部分是本公开文本的第二流动相,例如流动相a。在某些实施方案中,流动相b在约6分钟内从约10%增加到约20%,在约10分钟内从约20%增加到约30%,并且在约40分钟内从约30%增加到约38%。在其他实施方案中,流动相b在约121分钟内从约2%增加到约60%。本领域技术人员可以基于所需的色谱分离和/或感兴趣的分析物适当地调整感兴趣的流动相和所用的梯度定时。在一些实施方案中,液相色谱是高效液相色谱(hplc)。hplc在本领域中作为如下形式的液相色谱而已知,其中含有样品的液体溶剂在通过含有固相的柱时被加压。虽然传统或低压lc可以使用重力使流动相通过固相,但hplc使用泵向流动相施加压力,并且通常使用具有较小颗粒的固相来提高分辨率。在一些实施方案中,hplc使用在约50巴与约350巴之间的压力。在一些实施方案中,反相hplc可以用于浓缩和/或脱盐蛋白质(例如,aav衣壳蛋白)用于ms分析。在一些实施方案中,可以基于本文所述的发现,例如在如本文所用的lc/ms/ms中调整一个或多个参数,包括但不限于源电压、毛细管温度、esi电压(如果使用esi-ms)、cid能量和ms/ms事件的数量。在一些实施方案中,质谱(例如,如本文所述的lc/ms/ms中使用的)使用约2.5kv的源电压(例如,毛细管电压)。在一些实施方案中,质谱(例如,如本文所述的lc/ms/ms中使用的)使用约275℃的毛细管温度。在一些实施方案中,毛细管温度的范围从约20℃至约400℃。适用于lc/ms和/或lc/ms/ms的多种质量分析器在本领域中是已知的,包括但不限于飞行时间(tof)分析器、四极杆滤质器、四极杆tof(qtof)和离子阱(例如,基于傅立叶变换的质谱仪或orbitrap)。在orbitrap中,使用处于地电位的桶状外电极和心轴状中心电极将离子捕获在轨迹中,围绕中心电极以椭圆旋转,沿中心轴振荡,受到离心力和静电力平衡的限制。此类仪器的使用采用傅里叶变换操作将时域信号(例如,频率)从图像电流的检测转换成高分辨率质量测量(例如,纳米lc/ms/ms)。进一步的描述和细节可见于例如scheltema,r.a.等人(2014)mol.cellproteomics13:3698-3708;perry,r.h.等人(2008)mass.spectrom.rev.27:661-699;和scigelova,m.等人(2011)mol.cellproteomics10:m111.009431。如上所述,在一些实施方案中,ms包括例如使用orbitrap质量分析器的纳米lc/ms/ms。在一些实施方案中,离子源可以包括堆叠环离子导向器或s透镜。如在本领域中已知的,可以采用s透镜来使用射频(rf)聚焦离子束,从而增加离子到仪器中的传输。这可以改善灵敏度(例如,对于低强度离子)和/或提高扫描速率。在某些实施方案中,质谱的s透镜rf电平为约55%。在某些实施方案中,质谱的s透镜rf电平为约20%至约100%。在一些实施方案中,可以例如基于lc/ms和/或lc/ms/ms数据确定病毒衣壳蛋白的质量。在一些实施方案中,可以例如基于lc/ms和/或lc/ms/ms数据确定aav颗粒的vp1、vp2和vp3或aav颗粒的vp1、vp2和vp3的片段的质量。从ms数据确定蛋白质质量和/或身份的多种方法在本领域中是已知的。例如,肽质量指纹图谱可以用于基于ms数据确定蛋白质序列,或者可以基于与一种或多种组成肽相关的ms/ms数据鉴定蛋白质。当使用串联ms时,产物离子扫描可以用于分析与感兴趣的蛋白质的一种或多种肽相关的m/z数据。然后可以使用本领域中已知的软件,例如以将所鉴定的峰与参考或已知峰匹配,以将峰分组为同位素异构体包络(isotopomerenvelope)等。可以将肽质量值与已知肽序列的数据库进行比较。例如,mascot可以用于将观察到的肽与理论数据库肽匹配,例如通过将特定消化模式应用于计算机(insilico)蛋白质数据库而来。其他合适的软件可以包括但不限于proteomediscoverer、proteinprospector、x!tandem、pepfinder、bonics或masslynxtm(waters)。适用于ms数据分析的各个步骤的其他软件可见于例如www.ms-utils.org/wiki/pmwiki.php/main/softwarelist。在一些实施方案中,可以将本公开文本的确定或计算质量(例如,aav颗粒的vp1、vp2和/或vp3的确定或计算质量)与参考(例如,一种或多种aav血清型的vp1、vp2和/或vp3的理论质量)进行比较。本公开文本的参考可以包括本文所述的任何aav血清型中的一种或多种的vp1、vp2和/或vp3的理论质量。例如,在一些实施方案中,将vp1、vp2和/或vp3的质量与aav1衣壳、aav2衣壳、aav3衣壳、aav4衣壳、aav5衣壳、aav6衣壳、aav7衣壳、aav8衣壳、aavrh8衣壳、aav9衣壳、aav10衣壳、aavrh10衣壳、aav11衣壳、aav12衣壳、aavlk03衣壳(参见美国专利号9,169,299)、aav2r471a衣壳、aav2/2-7m8衣壳、aavdj衣壳(参见美国专利号7,588,772)、aavdj8衣壳、aav2n587a衣壳、aav2e548a衣壳、aav2n708a衣壳、aavv708k衣壳、山羊aav衣壳、aav1/aav2嵌合衣壳、牛aav衣壳或小鼠aav衣壳raav2/hbov1(嵌合aav/人博卡病毒1)、aav2hbko衣壳、aavphp.b衣壳或aavphp.eb衣壳中的一种或多种的理论质量进行比较。在一些实施方案中,可以将本公开文本的确定或计算质量(例如,aav颗粒的vp1、vp2和/或vp3的确定或计算质量)与对应aav血清型的vp1、vp2和/或vp3的理论质量进行比较。在一些实施方案中,本公开文本的方法可以包括确定aav颗粒的异质性。在一些实施方案中,vp1、vp2和/或vp3的质量中的一个或多个(例如,与参考质量,如理论、预测或预期质量)的偏差指示aav衣壳异质性。在一些实施方案中,异质性可以包括但不限于以下一种或多种:混合血清型、变体衣壳、衣壳氨基酸取代、截短衣壳或修饰衣壳。在一些实施方案中,可以组合如本文所述的lc/ms和lc/ms/ms的使用。在一些实施方案中,确定aav颗粒的血清型的方法可以包括使变性的aav颗粒经历lc/ms(例如,如本文所述的)并确定aav颗粒的vp1、vp2和vp3的质量;以及使vp1、vp2和/或vp3的片段经历lc/ms/ms并确定aav颗粒的vp1、vp2和vp3的片段的质量(vp1、vp2和vp3的片段的质量的特定组合指示aav血清型)。在一些实施方案中,确定aav颗粒的异质性的方法可以包括使变性的aav颗粒经历lc/ms(例如,如本文所述的),确定aav颗粒的vp1、vp2和vp3的质量,并将这些质量与aav血清型的vp1、vp2和vp3的理论质量进行比较;以及使vp1、vp2和/或vp3的片段经历lc/ms/ms,确定aav颗粒的vp1、vp2和vp3的片段的质量,并将这些质量与aav血清型的vp1、vp2和vp3的理论质量进行比较(vp1、vp2或vp3的质量中的一个或多个的偏差指示aav衣壳异质性)。在一些实施方案中,本公开文本的aav颗粒可以被乙酰化。例如,在一些实施方案中,vp1和/或vp3的n末端被乙酰化。如下文更详细描述的,可以突变aav衣壳蛋白的起始甲硫氨酸的第2位(imetx)的氨基酸,以便确定其对n末端(nt-)乙酰化的影响,以及影响nt-乙酰化对aav颗粒运输、转导和/或翻译后修饰(例如,糖基化、泛素化等)的功能性结果。在一些实施方案中,aav衣壳蛋白(例如,vp1或vp3)的n末端可以指起始甲硫氨酸之后的第一个氨基酸,其在一些情况下可以通过例如met-氨肽酶去除。在一些实施方案中,本公开文本的aav颗粒(例如,重组aav或raav颗粒)包含在vp1和/或vp3的氨基酸残基2处的氨基酸取代。在一些实施方案中,与参考(例如,氨基酸取代前的亲本aav颗粒或具有vp1和/或vp3的不同氨基酸残基2的aav颗粒)相比,vp1和/或vp3的氨基酸残基2处的氨基酸取代导致具有不同频率或比例的n末端乙酰化的vp1和/或vp3。在一些实施方案中,与亲本aav颗粒的vp1和/或vp3的氨基酸残基2处的n末端乙酰化相比,vp1和/或vp3的氨基酸残基2处的氨基酸取代改变了n末端乙酰化。例如,在某些实施方案中,与亲本aav颗粒的vp1的氨基酸残基2处的n末端乙酰化相比,vp1的氨基酸残基2处的氨基酸取代改变了n末端乙酰化。在某些实施方案中,与亲本aav颗粒的vp3的氨基酸残基2处的n末端乙酰化相比,vp3的氨基酸残基2处的氨基酸取代改变了n末端乙酰化。在一些实施方案中,“改变”n末端乙酰化的氨基酸取代(例如,vp1或vp3的氨基酸残基2处的氨基酸取代)导致更高频率的n末端乙酰化或更低频率的n末端乙酰化,例如与没有取代的vp1或vp3(如亲本vp1或vp3)相比。vp1和/或vp3可以属于本文所述的任何示例性aav血清型,包括其变体或杂合体(例如,带有酪氨酸突变或肝素结合突变)。用于n末端乙酰化的示例性测定包括但不限于质谱、同位素标记(例如,用同位素标记的乙酰基或其前体)、用乙酰化特异性抗体进行western印迹等。在一些实施方案中,aav衣壳蛋白(例如,vp1或vp3)的氨基酸残基2被cys、ser、thr、val、gly、asn、asp、glu、ile、leu、phe、gln、lys、met、pro或tyr取代。在一些实施方案中,氨基酸取代导致aav衣壳的更少脱酰胺化。在一些实施方案中,本公开文本的aav颗粒可以被脱酰胺化。例如,在一些实施方案中,vp1的n57和/或vp3的n382、n511和/或n715被脱酰胺化。如下文更详细描述的,可以突变选自aav衣壳蛋白(例如,vp1或vp3)的vp1的a35、vp1的n57、vp1的g58、vp3的n382、vp3的g383、vp3的n511、vp3的g512、vp3的n715或vp3的g716的氨基酸,以便确定其对脱酰胺化的影响,以及影响脱酰胺化对aav颗粒运输、转导和/或翻译后修饰(例如,糖基化、泛素化等)的功能性结果。在一些实施方案中,本公开文本的aav颗粒(例如,重组aav或raav颗粒)包含选自vp1的a35、vp1的n57、vp1的g58、vp3的n382、vp3的g383、vp3的n511、vp3的g512、vp3的n715和vp3的g716的一个或多个氨基酸残基处的氨基酸取代。在一些实施方案中,与参考(例如,氨基酸取代前的亲本aav颗粒或具有不同的对应氨基酸残基2的aav颗粒)相比,vp1的a35、vp1的n57、vp1的g58、vp3的n382、vp3的g383、vp3的n511、vp3的g512、vp3的n715和/或vp3的g716处的氨基酸取代导致具有不同频率或比例的脱酰胺化的vp1和/或vp3。在一些实施方案中,“改变”脱酰胺化的氨基酸取代(例如,vp1的a35、vp1的n57、vp1的g58、vp3的n382、vp3的g383、vp3的n511、vp3的g512、vp3的n715或vp3的g716处的氨基酸取代)导致更高频率的脱酰胺化或更低频率的脱酰胺化,例如与没有取代的vp1或vp3(如亲本vp1或vp3)相比。vp1和/或vp3可以属于本文所述的任何示例性aav血清型,包括其变体或杂合体(例如,带有酪氨酸突变或肝素结合突变)。用于脱酰胺化的示例性测定包括但不限于质谱、hplc(参见例如,来自promega的异天冬氨酸检测试剂盒)等。在一些实施方案中,vp1的n57、vp3的n382、vp3的n511和/或vp3的n715被asp取代,并且与亲本aav颗粒的vp1和/或vp3的脱酰胺化相比,氨基酸取代导致更高频率的脱酰胺化。在其他实施方案中,氨基酸取代是n57k或n57q,并且与亲本aav颗粒的vp1和/或vp3的脱酰胺化相比,氨基酸取代导致更低频率的脱酰胺化。在又其他实施方案中,vp1的g58、vp3的g383、vp3的g512和/或vp3的g716被不是gly的氨基酸(例如,ala、arg、asn、asp、cys、glu、gln、his、ile、leu、lys、met、phe、pro、ser、thr、trp、tyr或val)取代,并且与亲本aav颗粒的vp1和/或vp3的脱酰胺化相比,氨基酸取代导致更低频率的脱酰胺化。在又其他实施方案中,vp1的a35被asn取代,并且与亲本颗粒的vp1的脱酰胺化相比,导致更高频率的脱酰胺化。如本文所用,“n-乙酰化”是指借以将乙酰基共价添加到蛋白质的n末端氨基酸的氨基上的过程。通常,n末端乙酰转移酶(nat)将乙酰基从乙酰辅酶a(ac-coa)转移到蛋白质的第一个氨基酸残基的α-氨基。如本文所用,“脱酰胺化”是指其中天冬酰胺或谷氨酰胺侧链中的酰胺官能团被除去或转化为另一种官能团的化学反应。例如,天冬酰胺可以被转化为天冬氨酸或异天冬氨酸。在其他例子中,谷氨酰胺被转化为谷氨酸或焦谷氨酸(5-氧代脯氨酸)。在一些实施方案中,与亲本aav衣壳蛋白相比,aav颗粒被n-乙酰化到更高的程度。在一些实施方案中,与亲本aav颗粒相比,aav颗粒包含多于约5%、10%、15%、20%、25%、30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、95%或100%中的任何一个多的n-乙酰基。在一些实施方案中,与亲本aav颗粒相比,aav颗粒包含约5%-10%、10%-15%、15%-20%、20%-25%、25%-30%、30%-35%、35%-40%、40%-55%、45%-50%、50%-55%、55%-60%、60%-65%、65%-70%、70%-75%、75%-80%、80%-85%、85%-90%、90%-95%、95%-100%、5%-25%、25%-50%、50%-75%、75%-100%、5%-50%或50%-100%之间中的任何一个多的n-乙酰基。在一些实施方案中,与亲本aav颗粒相比,aav颗粒包含多于约2倍、3倍、4倍、5倍、10倍、25倍、50倍、100倍、500倍或1000倍中的任何一个多的n-乙酰基。在一些实施方案中,与亲本aav颗粒相比,aav颗粒包含约2倍至3倍、3倍至4倍、4倍至5倍、5倍至10倍、10倍至25倍、25倍至50倍、50倍至100倍、100倍至500倍、500倍至1000倍、2倍至10倍、10倍至100倍或100倍至1000倍之间中的任何一个多的n-乙酰基。在一些实施方案中,与亲本aav衣壳蛋白相比,aav颗粒被n-乙酰化到更低的程度。在一些实施方案中,与亲本aav颗粒相比,aav颗粒包含多于约5%、10%、15%、20%、25%、30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、95%或100%中的任何一个少的n-乙酰基。在一些实施方案中,与亲本aav颗粒相比,aav颗粒包含约5%-10%、10%-15%、15%-20%、20%-25%、25%-30%、30%-35%、35%-40%、40%-55%、45%-50%、50%-55%、55%-60%、60%-65%、65%-70%、70%-75%、75%-80%、80%-85%、85%-90%、90%-95%、95%-100%、5%-25%、25%-50%、50%-75%、75%-100%、5%-50%或50%-100%之间中的任何一个少的n-乙酰基。在一些实施方案中,与亲本aav颗粒相比,aav颗粒包含多于约2倍、3倍、4倍、5倍、10倍、25倍、50倍、100倍、500倍或1000倍中的任何一个少的n-乙酰基。在一些实施方案中,与亲本aav颗粒相比,aav颗粒包含约2倍至3倍、3倍至4倍、4倍至5倍、5倍至10倍、10倍至25倍、25倍至50倍、50倍至100倍、100倍至500倍、500倍至1000倍、2倍至10倍、10倍至100倍或100倍至1000倍之间中的任何一个少的n-乙酰基。在一些实施方案中,与亲本aav颗粒相比,aav颗粒被脱酰胺化到更高的程度。在一些实施方案中,与亲本aav颗粒相比,aav颗粒被多于约5%、10%、15%、20%、25%、30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、95%或100%中的任何一个多地脱酰胺化。在一些实施方案中,aav颗粒比亲本aav颗粒被约5%-10%、10%-15%、15%-20%、20%-25%、25%-30%、30%-35%、35%-40%、40%-55%、45%-50%、50%-55%、55%-60%、60%-65%、65%-70%、70%-75%、75%-80%、80%-85%、85%-90%、90%-95%、95%-100%、5%-25%、25%-50%、50%-75%、75%-100%、5%-50%或50%-100%之间中的任何一个多地脱酰胺化。在一些实施方案中,与亲本aav颗粒相比,aav颗粒被脱酰胺化多于约2倍、3倍、4倍、5倍、10倍、25倍、50倍、100倍、500倍或1000倍中的任何一个。在一些实施方案中,aav颗粒比亲本aav颗粒被约2倍至3倍、3倍至4倍、4倍至5倍、5倍至10倍、10倍至25倍、25倍至50倍、50倍至100倍、100倍至500倍、500倍至1000倍、2倍至10倍、10倍至100倍或100倍至1000倍之间中的任何一个多地脱酰胺化。在一些实施方案中,与亲本aav衣壳蛋白相比,aav的衣壳蛋白被脱酰胺化到更低的程度。在一些实施方案中,与亲本aav颗粒相比,aav颗粒被多于约5%、10%、15%、20%、25%、30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、95%或100%中的任何一个少地脱酰胺化。在一些实施方案中,aav颗粒比亲本aav颗粒被约5%-10%、10%-15%、15%-20%、20%-25%、25%-30%、30%-35%、35%-40%、40%-55%、45%-50%、50%-55%、55%-60%、60%-65%、65%-70%、70%-75%、75%-80%、80%-85%、85%-90%、90%-95%、95%-100%、5%-25%、25%-50%、50%-75%、75%-100%、5%-50%或50%-100%之间中的任何一个少地脱酰胺化。在一些实施方案中,aav颗粒比亲本aav颗粒被多于约2倍、3倍、4倍、5倍、10倍、25倍、50倍、100倍、500倍或1000倍中的任何一个少地脱酰胺化。在一些实施方案中,aav颗粒比亲本aav颗粒被约2倍至3倍、3倍至4倍、4倍至5倍、5倍至10倍、10倍至25倍、25倍至50倍、50倍至100倍、100倍至500倍、500倍至1000倍、2倍至10倍、10倍至100倍或100倍至1000倍之间中的任何一个少地脱酰胺化。本发明提供了n-乙酰化和脱酰胺化的任何组合。例如,aav衣壳蛋白可以被n-乙酰化到比亲本aav衣壳蛋白高的程度并且被脱酰胺化到比亲本aav衣壳蛋白高的程度,aav衣壳蛋白可以被n-乙酰化到比亲本aav衣壳蛋白高的程度并且被脱酰胺化到与亲本aav衣壳蛋白相同的程度,aav衣壳蛋白可以被n-乙酰化到比亲本aav衣壳蛋白高的程度并且被脱酰胺化到比亲本aav衣壳蛋白低的程度,aav衣壳蛋白可以被n-乙酰化到与亲本aav衣壳蛋白相同的程度并且被脱酰胺化到比亲本aav衣壳蛋白高的程度,aav衣壳蛋白可以被n-乙酰化到与亲本aav衣壳蛋白相同的程度并且被脱酰胺化到与亲本aav衣壳蛋白相同的程度,aav衣壳蛋白可以被n-乙酰化到与亲本aav衣壳蛋白相同的程度并且被脱酰胺化到比亲本aav衣壳蛋白低的程度,aav衣壳蛋白可以被n-乙酰化到比亲本aav衣壳蛋白低的程度并且被脱酰胺化到比亲本aav衣壳蛋白高的程度,aav衣壳蛋白可以被n-乙酰化到比亲本aav衣壳蛋白低的程度并且被脱酰胺化到与亲本aav衣壳蛋白相同的程度,或者aav衣壳蛋白可以被n-乙酰化到比亲本aav衣壳蛋白低的程度并且被脱酰胺化到比亲本aav衣壳蛋白低的程度。iv.载体在某些方面,本发明涉及适用于本文所述的任何方法的病毒颗粒,其可以包含aav载体(例如,raav载体)或衍生自另一种病毒的载体。在一些实施方案中,病毒颗粒包含编码异源核酸(例如,异源转基因)的载体。在一些实施方案中,aav颗粒包含编码异源核酸(例如,异源转基因)的aav载体基因组。本发明考虑了使用重组病毒基因组引入编码治疗性多肽和/或核酸的一个或多个核酸序列以用于包装到raav病毒颗粒中。重组病毒基因组可以包括建立治疗性多肽和/或核酸的表达的任何元件,例如启动子、本公开文本的itr、核糖体结合元件、终止子、增强子、选择标记、内含子、polya信号和/或复制起点。在一些实施方案中,异源核酸编码治疗性多肽。治疗性多肽可以例如提供在细胞或生物体中不存在或以降低的水平存在的多肽和/或酶活性。可替代地,治疗性多肽可以提供间接抵消细胞或生物体中的失衡的多肽和/或酶活性。例如,用于与代谢酶或活性缺陷引起的代谢物累积相关的障碍的治疗性多肽可以提供缺失的代谢酶或活性,或者它可以提供导致代谢物减少的替代代谢酶或活性。治疗性多肽还可以通过例如作为显性失活多肽起作用而用于降低多肽(例如,过表达、通过功能获得性突变激活或其活性以其他方式被错误调节的多肽)的活性。本发明的核酸可以编码作为细胞内蛋白质、锚定在细胞膜内、保留在细胞内或由用本发明的载体转导的细胞分泌的多肽。对于由接受载体的细胞分泌的多肽,多肽可以是可溶的(即,不附着于细胞)。例如,可溶性多肽缺乏跨膜区并且从细胞分泌。鉴定和去除编码跨膜结构域的核酸序列的技术在本领域中是已知的。本发明的核酸(例如aav载体基因组)可以包含转基因,其是编码调节或治疗cns相关障碍的蛋白质或功能性rna的核酸。以下是与cns相关障碍相关的基因的非限制性列表:神经元凋亡抑制蛋白(naip)、神经生长因子(ngf)、胶质细胞衍生生长因子(glial-derivedgrowthfactor,gdnf)、脑源性生长因子(bdnf)、睫状神经营养因子(cntf)、酪氨酸羟化酶(tm、gtp环水解酶(gtpch)、天冬氨酸酰酶(aspartoacylase,aspa)、超氧化物歧化酶(sod1)、抗氧化剂、抗血管生成多肽、抗炎多肽和氨基酸脱羧酶(aadc)。例如,在帕金森病的治疗中有用的转基因编码th,其是多巴胺合成中的限速酶。编码产生tii辅因子四氢生物蝶呤的gtpcii的转基因也可以用于帕金森病的治疗。编码促进左旋多巴转化为da的gdnf或bdnf或aadc的转基因也可以用于帕金森病的治疗。对于als的治疗,有用的转基因可以编码:gdnf、bdnf或cntf。同样对于als的治疗,有用的转基因可以编码抑制sod1表达的功能性rna,例如shrna、mirna。对于局部缺血的治疗,有用的转基因可以编码naip或ngf。编码β-葡糖醛酸糖苷酶(gus)的转基因可用于某些溶酶体贮积病(例如,vii型粘多糖病(mpsvii))的治疗。编码前药激活基因(例如,将更昔洛韦转化为破坏dna合成并导致细胞死亡的毒性核苷酸的hsv-胸苷激酶)的转基因可用于治疗某些癌症,例如当与前药联合使用时。编码内源性阿片样物质(如β-内啡肽)的转基因可用于治疗疼痛。抗氧化剂的例子包括但不限于sod1;sod2;过氧化氢酶;去乙酰化酶1、3、4,或5;nrf2;pgc1a;gcl(催化亚基);gcl(修饰物亚基);脂联素;谷胱甘肽过氧化物酶1;和脑红蛋白。抗血管生成多肽的例子包括但不限于血管抑素、内皮抑素、pedf、可溶性vegf受体和可溶性pdgf受体。抗炎多肽的例子包括但不限于il-10、可溶性il17r、可溶性tnf-r、tnf-r-ig、il-1抑制剂和il18抑制剂。可以用于本发明的raav载体的转基因的其他例子对于技术人员而言是清楚的(参见例如,costantinilc等人,genetherapy(2000)7,93-109)。在一些实施方案中,异源核酸编码治疗性核酸。在一些实施方案中,治疗性核酸可以包括但不限于sirna、shrna、rnai、mirna、反义rna、核酶或dna核酶(dnazyme)。因此,治疗性核酸可以编码如下rna,当从载体的核酸转录时,其可以通过干扰与本发明的障碍相关的异常或过量蛋白质的翻译或转录来治疗障碍。例如,本发明的核酸可以编码通过高度特异性消除或减少编码异常和/或过量蛋白质的mrna来治疗障碍的rna。治疗性rna序列包括rnai、小抑制性rna(sirna)、微小rna(mirna)和/或核酶(如锤头和发夹核酶),其可以通过高度特异性消除或减少编码异常和/或过量蛋白质的mrna来治疗障碍。在一些实施方案中,治疗性多肽或治疗性核酸用于治疗cns障碍。不希望受理论束缚,据信治疗性多肽或治疗性核酸可以用于降低或消除其功能获得与障碍相关的多肽的表达和/或活性,或者增强多肽的表达和/或活性以补充与障碍相关的缺陷(例如,其表达显示相似或相关活性的基因的突变)。可以通过本发明的治疗性多肽或治疗性核酸治疗的本发明的障碍的非限制性例子(可以被靶向或提供的示例性基因提供在每种障碍的圆括号中)包括中风(例如,胱天蛋白酶-3、beclin1、ask1、par1、hif1α、puma和/或fukuda,a.m.和badaut,j.(2013)genes(basel)4:435-456中描述的任何基因)、亨廷顿病(突变型htt)、癫痫(例如,scn1a、nmdar、adk和/或boison,d.(2010)epilepsia51:1659-1668中描述的任何基因)、帕金森病(α-突触核蛋白)、葛雷克病(也称为肌萎缩侧索硬化症;sod1)、阿尔茨海默病(tau,淀粉样前体蛋白)、皮质基底节变性或cbd(tau)、皮质基底节神经节变性或cbgd(tau)、额颞叶痴呆或ftd(tau)、进行性核上性麻痹或psp(tau)、多系统萎缩或msa(α-突触核蛋白)、脑癌(例如,脑癌中牵涉的突变型或过表达的致癌基因)和溶酶体贮积病(lsd)。本发明的障碍可以包括涉及大面积皮质(例如,皮质的多于一个功能区域、皮质的多于一个叶和/或整个皮质)的那些。可以通过本发明的治疗性多肽或治疗性核酸治疗的本发明的障碍的其他非限制性例子包括创伤性脑损伤、酶功能障碍症、精神障碍(包括创伤后应激综合征)、神经变性疾病和认知障碍(包括痴呆、自闭症和抑郁症)。酶功能障碍症包括但不限于脑白质营养不良(包括卡纳万病(canavan’sdisease))和下文描述的任何溶酶体贮积病。在一些实施方案中,治疗性多肽或治疗性核酸用于治疗溶酶体贮积病。如在本领域中通常已知的,溶酶体贮积病是罕见的遗传性代谢障碍,其特征在于溶酶体功能的缺陷。此类障碍通常由适当的粘多糖、糖蛋白和/或脂质代谢所需的酶缺陷引起,导致溶酶体储存的细胞材料的病理性积累。可以通过本发明的治疗性多肽或治疗性核酸治疗的本发明的溶酶体贮积病的非限制性例子(可以被靶向或提供的示例性基因提供在每种障碍的圆括号中)包括2型或3型戈谢病(酸性β-葡糖苷酶,gba)、gm1神经节苷脂沉积症(β-半乳糖苷酶-1,glb1)、亨特病(艾杜糖醛酸2-硫酸酯酶,ids)、克拉伯病(半乳糖神经鞘氨醇酶,galc)、甘露糖苷贮积病(甘露糖苷酶,如α-d-甘露糖苷酶,man2b1)、β甘露糖苷贮积病(β-甘露糖苷酶,manba)、异染性脑白质营养不良病(假芳基硫酸酯酶a,arsa)、ii/iii型粘脂糖症(n-乙酰氨基葡萄糖-1-磷酸转移酶,gnptab)、a型尼曼-皮克病(niemann-pickadisease)(酸性鞘磷脂酶,asm)、c型尼曼-皮克病(尼曼-皮克c蛋白,npc1)、蓬佩病(酸性α-1,4-葡糖苷酶,gaa)、山德霍夫病(sandhoffdisease)(氨基己糖苷酶β亚基,hexb)、a型圣菲利波病(sanfilippoadisease)(n-磺基葡糖胺磺基水解酶,mps3a)、b型圣菲利波病(n-α-乙酰氨基葡糖苷酶,naglu)、c型圣菲利波病(肝素乙酰辅酶a:α-氨基葡糖苷酶n-乙酰转移酶,mps3c)、d型圣菲利波病(n-乙酰氨基葡萄糖-6-硫酸酯酶,gns)、辛德勒病(α-n-乙酰氨基半乳糖苷酶,naga)、斯莱病(slydisease)(β-葡糖醛酸酶,gusb)、泰-萨二氏病(氨基己糖苷酶α亚基,hexa)和沃尔曼病(溶酶体酸性脂肪酶,lipa)。另外的溶酶体贮积病以及与每种疾病相关的缺陷酶列于下表1中。在一些实施方案中,下表中列出的疾病通过本发明的治疗性多肽或治疗性核酸进行治疗,该治疗性多肽或治疗性核酸补充或以其他方式补偿对应的酶缺陷。表1.溶酶体贮积症和相关的缺陷酶。在一些实施方案中,异源核酸与启动子可操作地连接。示例性启动子包括但不限于巨细胞病毒(cmv)即时早期启动子、rsvltr、momlvltr、磷酸甘油酸激酶-1(pgk)启动子、猿猴病毒40(sv40)启动子和ck6启动子、转甲状腺素蛋白启动子(ttr)、tk启动子、四环素应答性启动子(tre)、hbv启动子、haat启动子、lsp启动子、嵌合肝脏特异性启动子(lsp)、e2f启动子、端粒酶(htert)启动子、巨细胞病毒增强子/鸡β-肌动蛋白/兔β-球蛋白启动子(cag启动子;niwa等人,gene,1991,108(2):193-9)和延长因子1-α启动子(efl-α)启动子(kim等人,gene,1990,91(2):217-23和guo等人,genether.,1996,3(9):802-10)。在一些实施方案中,启动子包含人β-葡糖醛酸酶启动子或连接至鸡β-肌动蛋白(cba)启动子的巨细胞病毒增强子。启动子可以是组成型、诱导型或阻抑型启动子。在一些实施方案中,本发明提供了包含编码与cba启动子可操作地连接的本公开文本的异源转基因的核酸的重组载体。示例性启动子和描述可见于例如美国专利授予前公开案20140335054。组成型启动子的例子包括但不限于逆转录劳斯肉瘤病毒(rsv)ltr启动子(任选地具有rsv增强子)、巨细胞病毒(cmv)启动子(任选地具有cmv增强子)[参见例如,boshart等人,cell,41:521-530(1985)]、sv40启动子、二氢叶酸还原酶启动子、13-肌动蛋白启动子、磷酸甘油激酶(pgk)启动子和efla启动子[invitrogen]。诱导型启动子允许调节基因表达,并且可以通过外源提供的化合物、环境因子(如温度)或存在特定生理状态(例如,急性期)、细胞的特定分化状态或在仅复制细胞时进行调节。诱导型启动子和诱导型系统可从多种商业来源获得,包括但不限于invitrogen、clontech和ariad。已经描述了许多其他系统,并且可以由本领域技术人员容易地选择。通过外源提供的启动子调节的诱导型启动子的例子包括锌诱导型绵羊金属硫蛋白(mt)启动子、地塞米松(dex)诱导型小鼠乳腺肿瘤病毒(mmtv)启动子、t7聚合酶启动子系统(wo98/10088)、蜕皮激素昆虫启动子(no等人,proc.natl.acad.sci.usa,93:3346-3351(1996))、四环素阻抑型系统(gossen等人,proc.natl.acad.sci.usa,89:5547-5551(1992))、四环素诱导型系统(gossen等人,science,268:1766-1769(1995),还参见harvey等人,curr.opin.chem.biol.,2:512-518(1998))、ru486诱导型系统(wang等人,nat.biotech.,15:239-243(1997)和wang等人,genether.,4:432-441(1997))和雷帕霉素诱导型系统(magari等人,j.clin.invest.,100:2865-2872(1997))。在这种背景下可使用的仍其他类型的诱导型启动子是通过特定生理状态(例如,温度、急性期)、细胞的特定分化状态或在仅复制细胞时调节的那些。在另一个实施方案中,将使用用于转基因的天然启动子或其片段。当希望转基因的表达应模拟天然表达时,可以使用天然启动子。当必须暂时或发展地或以组织特异性方式或响应于特定转录刺激物调节转基因的表达时,可以使用天然启动子。在另一实施方案中,也可以使用其他天然表达控制元件(如增强子元件、多腺苷酸化位点或kozak共有序列)模拟天然表达。在一些实施方案中,调节序列赋予组织特异性基因表达能力。在一些情况下,组织特异性调节序列结合以组织特异性方式诱导转录的组织特异性转录因子。此类组织特异性调节序列(例如,启动子、增强子等)在本领域中是众所周知的。在一些实施方案中,载体包含内含子。例如,在一些实施方案中,内含子是衍生自鸡β-肌动蛋白和兔β-球蛋白的嵌合内含子。在一些实施方案中,内含子是小鼠微小病毒(mvm)内含子。在一些实施方案中,载体包含多腺苷酸化(polya)序列。多腺苷酸化序列的许多例子在本领域中是已知的,如牛生长激素(bgh)poly(a)序列(参见例如,登录号ef592533)、sv40多腺苷酸化序列和hsvtkpa多腺苷酸化序列。v.病毒颗粒和产生病毒颗粒的方法本公开文本的某些方面涉及重组病毒颗粒(例如,raav颗粒)。基于本文提供的指导,本公开文本的技术可以由本领域技术人员适当地适用于各种不同的病毒。在一些实施方案中,病毒属于腺病毒科,其包括通常称为腺病毒的无包膜病毒。在一些实施方案中,病毒属于腺胸腺病毒属、禽腺病毒属、美洲白鲟腺病毒属、哺乳动物腺病毒属或唾液酸酶腺病毒属。在一些实施方案中,病毒属于细小病毒科,其包括诸如aav和博卡细小病毒(bocaparvovirus)等无包膜病毒。在一些实施方案中,病毒属于浓核病毒亚科(densovirinae)。在一些实施方案中,病毒属于双义浓核病毒属、短浓核病毒属、虾肝胰腺浓核病毒属、重复浓核病毒属或对虾浓核病毒属。在一些实施方案中,病毒属于细小病毒亚科(parvovirinae)。在一些实施方案中,病毒属于水貂阿留申细小病毒属、aveparvovirus、博卡细小病毒属、copiparvovirus、腺病毒伴随细小病毒属、嗜红细胞细小病毒属、protoparvovirus或tetraparvovirus。在一些实施方案中,病毒属于逆转录病毒科,其包括包膜病毒(包括慢病毒)。在一些实施方案中,病毒属于正逆转录病毒亚科(orthoretrovirinae)。在一些实施方案中,病毒属于α逆转录病毒属、β逆转录病毒属、δ逆转录病毒属、ε逆转录病毒属、γ逆转录病毒属或慢病毒属。在一些实施方案中,病毒属于泡沫病毒亚科(spumaretrovirinae)。在一些实施方案中,病毒属于泡沫病毒属。在一些实施方案中,病毒属于杆状病毒科,其包括包膜病毒(包括α杆状病毒)。在一些实施方案中,病毒属于α杆状病毒属、β杆状病毒属、δ杆状病毒属或γ杆状病毒属。在一些实施方案中,病毒属于疱疹病毒科,其包括诸如单纯疱疹病毒hsv-1和hsv-2等包膜病毒。在一些实施方案中,病毒属于α疱疹病毒亚科(alphaherpesvirinae)。在一些实施方案中,病毒属于传喉炎病毒属、马立克病毒属、单纯疱疹病毒属或水痘病毒属。在一些实施方案中,病毒属于β疱疹病毒亚科(betaherpesvirinae)。在一些实施方案中,病毒属于巨细胞病毒属、鼠巨细胞病毒属、长鼻动物病毒属或玫瑰疱疹病毒属。在一些实施方案中,病毒属于γ疱疹病毒亚科(gammaherpesvirinae)。在一些实施方案中,病毒属于淋巴滤泡病毒属、玛卡病毒属、马疱疹病毒属或猴病毒属。在一些实施方案中,病毒是aav病毒。在aav颗粒中,核酸衣壳化于aav颗粒中。aav颗粒还包含衣壳蛋白。在一些实施方案中,核酸包含在转录方向上可操作地连接的异源核酸和/或一种或多种以下组分:控制序列(包括转录起始序列和终止序列),从而形成表达盒。在一些实施方案中,病毒颗粒包含aavitr序列。例如,表达盒可以在5'和3'端侧接至少一个功能性aavitr序列。“功能性aavitr序列”意指旨在用于挽救、复制和包装aav病毒粒子的itr序列功能。参见davidson等人,pnas,2000,97(7)3428-32;passini等人,j.virol.,2003,77(12):7034-40;和pechan等人,genether.,2009,16:10-16,将其全部通过引用以其整体并入本文。为了实施本发明的一些方面,重组载体包含至少所有为衣壳化所必需的aav序列和用于由raav感染的物理结构。用于本发明的载体的aavitr无需具有野生型核苷酸序列(例如,如kotin,hum.genether.,1994,5:793-801中所述),并且可以通过插入、缺失或取代核苷酸而改变,或者aavitr可以衍生自几种aav血清型中的任何一种。目前已知超过40种aav血清型,并且不断鉴定出新的血清型和现有血清型的变体。参见gao等人,pnas,2002,99(18):11854-6;gao等人,pnas,2003,100(10):6081-6;和bossis等人,j.virol.,2003,77(12):6799-810。使用任何aav血清型都被认为在本发明的范围之内。在一些实施方案中,raav载体是衍生自aav血清型的载体,包括但不限于aav1、aav2、aav3、aav4、aav5、aav6、aav7、aav8、aavrh8、aavrh8r、aav9、aav10、aavrh10、aav11、aav12、aavlk03、aav2r471a、aavdj、aavdj8、山羊aav、牛aav或小鼠aavitr等。在一些实施方案中,aav(例如,raav载体)中的核酸包含aav1、aav2、aav3、aav4、aav5、aav6、aav7、aav8、aavrh8、aavrh8r、aav9、aav10、aavrh10、aav11、aav12、aavlk03、aav2r471a、aavdj、aavdj8、山羊aav、牛aav或小鼠aavitr等的itr。在一些实施方案中,aav颗粒包含编码侧接一个或多个aavitr的异源转基因的aav载体。在一些实施方案中,raav颗粒包含选自以下的衣壳化蛋白:aav1、aav2、aav3、aav4、aav5、aav6(例如,野生型aav6衣壳或变体aav6衣壳如shh10,如美国专利授予前公开案2012/0164106中所述)、aav7、aav8、aavrh8、aavrh8r、aav9(例如,野生型aav9衣壳或修饰的aav9衣壳,如美国专利授予前公开案2013/0323226中所述)、aav10、aavrh10、aav11、aav12、酪氨酸衣壳突变体、肝素结合衣壳突变体、aav2r471a衣壳、aavaav2/2-7m8衣壳、aavlk03衣壳、aavdj衣壳(例如,aav-dj/8衣壳、aav-dj/9衣壳或美国专利授予前公开案2012/0066783中所述的任何其他衣壳)、aav2n587a衣壳、aav2e548a衣壳、aav2n708a衣壳、aavv708k衣壳、山羊aav衣壳、aav1/aav2嵌合衣壳、牛aav衣壳、小鼠aav衣壳、raav2/hbov1衣壳、aav2hbko衣壳、aavphp.b衣壳或aavphp.eb衣壳或者美国专利号8,283,151或国际公开号wo/2003/042397中所述的aav衣壳。在进一步的实施方案中,raav颗粒包含来自进化枝a-f的aav血清型的衣壳蛋白。本公开文本的某些方面涉及包含氨基酸残基2处的氨基酸取代的aav(例如,raav)衣壳蛋白。在一些实施方案中,与亲本aav衣壳蛋白的氨基酸残基2处的n末端乙酰化相比,氨基酸残基2处的氨基酸取代改变了n末端乙酰化。如本文所述的,可以检查aav衣壳蛋白的起始甲硫氨酸的第2位(imetx)的氨基酸对n末端乙酰化、运输、转导和/或其他一种或多种翻译后修饰(例如,糖基化、泛素化等)的影响。本文所述的用于检查乙酰化或其与aav颗粒相关的功能性结果的任何测定都可以用于评估n末端乙酰化。在一些实施方案中,aav衣壳蛋白(例如,vp1或vp3)的氨基酸残基2被cys、ser、thr、val、gly、asn、asp、glu、ile、leu、phe、gln、lys、met、pro或tyr取代。在一些实施方案中,氨基酸取代导致aav衣壳的更少脱酰胺化。本公开文本的其他方面涉及包含改变脱酰胺化的氨基酸取代的aav(例如,raav)衣壳蛋白。在一些实施方案中,“改变”脱酰胺化的氨基酸取代(例如,vp1的a35、vp1的n57、vp1的g58、vp3的n382、vp3的g383、vp3的n511、vp3的g512、vp3的n715或vp3的g716处的氨基酸取代)导致更高频率的脱酰胺化或更低频率的脱酰胺化,例如与没有取代的vp1或vp3(如亲本vp1或vp3)相比。如本文所述的,可以检查aav衣壳蛋白(例如,vp1或vp3)的潜在脱酰胺位点对脱酰胺化、运输、转导和/或其他一种或多种翻译后修饰(例如,糖基化、泛素化等)的影响。本文所述的用于检查脱酰胺化或其与aav颗粒相关的功能性结果的任何测定都可以用于评估脱酰胺化。本文描述了几个潜在的脱酰胺化位点。在一些实施方案中,改变脱酰胺化的氨基酸取代选自vp1的a35、vp1的n57、vp1的g58、vp3的n382、vp3的g383、vp3的n511、vp3的g512、vp3的n715或vp3的g716。例如,在一些实施方案中,vp1的n57、vp3的n382、vp3的n511和/或vp3的n715被asp取代,并且与亲本aav颗粒的vp1和/或vp3的脱酰胺化相比,氨基酸取代导致更高频率的脱酰胺化。在其他实施方案中,氨基酸取代是n57k或n57q,并且与亲本aav颗粒的vp1和/或vp3的脱酰胺化相比,氨基酸取代导致更低频率的脱酰胺化。在又其他实施方案中,vp1的g58、vp3的g383、vp3的g512和/或vp3的g716被不是gly的氨基酸(例如,ala、arg、asn、asp、cys、glu、gln、his、ile、leu、lys、met、phe、pro、ser、thr、trp、tyr或val)取代,并且与亲本aav颗粒的vp1和/或vp3的脱酰胺化相比,氨基酸取代导致更低频率的脱酰胺化。在一些实施方案中,aav衣壳蛋白是vp1、vp2或vp3。aav颗粒可以包含本文所述的任何示例性aav衣壳血清型,如aav1、aav2、aav3、aav4、aav5、aav6、aav7、aav8、aavrh8、aavrh8r、aav9、aav10、aavrh10、aav11、aav12、aavlk03、aav2r471a、aav2/2-7m8、aavdj、aavdj8、aav2n587a、aav2e548a、aav2n708a、aavv708k、山羊aav、aav1/aav2嵌合体、牛aav、小鼠aav或raav2/hbov1。aav衣壳蛋白还可以包含本文所述的任何衣壳蛋白突变,如酪氨酸和/或肝素结合突变。本公开文本的其他方面涉及改善raav颗粒的稳定性的方法。在一些实施方案中,该方法包括取代vp1和/或vp3的氨基酸残基2,例如如本文所述。例如,在一些实施方案中,vp1的氨基酸残基2被取代。在其他实施方案中,vp3的氨基酸残基2被取代。在一些实施方案中,2位的取代的氨基酸以比亲本vp1和/或vp3的氨基酸残基2更高的频率被n-乙酰化,例如如本文所述。在一些实施方案中,取代vp1和/或vp3的氨基酸残基2将raav颗粒的稳定性改善了至少约5%、至少约10%、至少约15%、至少约20%、至少约25%、至少约30%、至少约35%、至少约40%、至少约45%、至少约50%、至少约55%、至少约60%、至少约65%、至少约70%、至少约75%、至少约80%、至少约85%、至少约90%、至少约95%或至少约100%。在一些实施方案中,可以将在2位具有取代的氨基酸的raav颗粒的稳定性与例如相同血清型的野生型或亲本aav衣壳进行比较。例如,在一些实施方案中,例如与包含野生型衣壳的raav颗粒的稳定性相比,取代vp1和/或vp3的氨基酸残基2将raav颗粒的稳定性改善了约10%至约100%、约20%至约100%、约30%至约100%、约40%至约100%、约50%至约100%、约60%至约100%、约70%至约100%、约80%至约100%、约90%至约100%、约10%至约90%、约20%至约90%、约30%至约90%、约40%至约90%、约50%至约90%、约60%至约90%、约70%至约90%、约80%至约90%、约10%至约80%、约20%至约80%、约30%至约80%、约40%至约80%、约50%至约80%、约60%至约80%、约70%至约80%、约10%至约70%、约20%至约70%、约30%至约70%、约40%至约70%、约50%至约70%、约60%至约70%、约10%至约60%、约20%至约60%、约30%至约60%、约40%至约60%、约50%至约60%、约10%至约50%、约20%至约50%、约30%至约50%、约40%至约50%、约10%至约40%、约20%至约40%、约30%至约40%、约10%至约30%、约20%至约30%或约10%至约20%中的任何一个。aav颗粒稳定性可以使用本领域中已知的多种测定来测量,包括但不限于差示扫描荧光(dsf)、差示扫描量热法(dsc)、其他热变性测定、对蛋白水解的敏感性、观察变性的成像或结构分析(例如,使用电子显微术)、转导效率或在特定温度(例如,室温或4℃,用于热稳定性)下保持指定的时间间隔或在特定ph(例如,ph稳定性)下处理的aav颗粒组合物的另一种功能测定等。本公开文本的其他方面涉及改善raav颗粒的组装的方法。在一些实施方案中,该方法包括取代vp1和/或vp3的氨基酸残基2,例如如本文所述。例如,在一些实施方案中,vp1的氨基酸残基2被取代。在其他实施方案中,vp3的氨基酸残基2被取代。在一些实施方案中,2位的取代的氨基酸以比亲本vp1和/或vp3的氨基酸残基2更高的频率被n-乙酰化,例如如本文所述。在一些实施方案中,取代vp1和/或vp3的氨基酸残基2将raav颗粒的组装改善了至少约5%、至少约10%、至少约15%、至少约20%、至少约25%、至少约30%、至少约35%、至少约40%、至少约45%、至少约50%、至少约55%、至少约60%、至少约65%、至少约70%、至少约75%、至少约80%、至少约85%、至少约90%、至少约95%或至少约100%。在一些实施方案中,可以将在2位具有取代的氨基酸的raav颗粒的组装与例如相同血清型的野生型或亲本aav衣壳进行比较。例如,在一些实施方案中,例如与包含野生型衣壳的raav颗粒的组装相比,取代vp1和/或vp3的氨基酸残基2将raav颗粒的组装改善了约10%至约100%、约20%至约100%、约30%至约100%、约40%至约100%、约50%至约100%、约60%至约100%、约70%至约100%、约80%至约100%、约90%至约100%、约10%至约90%、约20%至约90%、约30%至约90%、约40%至约90%、约50%至约90%、约60%至约90%、约70%至约90%、约80%至约90%、约10%至约80%、约20%至约80%、约30%至约80%、约40%至约80%、约50%至约80%、约60%至约80%、约70%至约80%、约10%至约70%、约20%至约70%、约30%至约70%、约40%至约70%、约50%至约70%、约60%至约70%、约10%至约60%、约20%至约60%、约30%至约60%、约40%至约60%、约50%至约60%、约10%至约50%、约20%至约50%、约30%至约50%、约40%至约50%、约10%至约40%、约20%至约40%、约30%至约40%、约10%至约30%、约20%至约30%或约10%至约20%中的任何一个。aav颗粒组装可以使用本领域中已知的多种测定来测量,包括但不限于测量颗粒产生量和/或速率、定量衣壳产生(例如,使用本文所述的任何方法纯化后)、测定完全型载体与空衣壳的产生、测量转导效率、观察颗粒形成的成像或结构分析(例如,使用电子显微术)、aav衣壳蛋白的产生(例如,如通过western印迹测定)等。本公开文本的其他方面涉及改善raav颗粒的转导的方法。在一些实施方案中,该方法包括取代vp1和/或vp3的氨基酸残基2,例如如本文所述。例如,在一些实施方案中,vp1的氨基酸残基2被取代。在其他实施方案中,vp3的氨基酸残基2被取代。在一些实施方案中,2位的取代的氨基酸以比亲本vp1和/或vp3的氨基酸残基2更高的频率被n-乙酰化,例如如本文所述。在一些实施方案中,取代vp1和/或vp3的氨基酸残基2将raav颗粒的转导改善了至少约5%、至少约10%、至少约15%、至少约20%、至少约25%、至少约30%、至少约35%、至少约40%、至少约45%、至少约50%、至少约55%、至少约60%、至少约65%、至少约70%、至少约75%、至少约80%、至少约85%、至少约90%、至少约95%或至少约100%。在一些实施方案中,可以将在2位具有取代的氨基酸的raav颗粒的转导与例如相同血清型的野生型或亲本aav衣壳进行比较。例如,在一些实施方案中,例如与包含野生型衣壳的raav颗粒的转导相比,取代vp1和/或vp3的氨基酸残基2将raav颗粒的转导改善了约10%至约100%、约20%至约100%、约30%至约100%、约40%至约100%、约50%至约100%、约60%至约100%、约70%至约100%、约80%至约100%、约90%至约100%、约10%至约90%、约20%至约90%、约30%至约90%、约40%至约90%、约50%至约90%、约60%至约90%、约70%至约90%、约80%至约90%、约10%至约80%、约20%至约80%、约30%至约80%、约40%至约80%、约50%至约80%、约60%至约80%、约70%至约80%、约10%至约70%、约20%至约70%、约30%至约70%、约40%至约70%、约50%至约70%、约60%至约70%、约10%至约60%、约20%至约60%、约30%至约60%、约40%至约60%、约50%至约60%、约10%至约50%、约20%至约50%、约30%至约50%、约40%至约50%、约10%至约40%、约20%至约40%、约30%至约40%、约10%至约30%、约20%至约30%或约10%至约20%中的任何一个。aav颗粒转导可以使用本领域中已知的多种测定来测量,包括但不限于本文所述的转导效率测定。在一些实施方案中,本发明提供了降低raav颗粒的转导的方法;例如,通过取代vp1和/或vp3的氨基酸残基2。本公开文本的其他方面涉及改善raav颗粒的稳定性的方法。在一些实施方案中,该方法包括取代vp1和/或vp3的改变脱酰胺化的氨基酸,例如如本文所述。例如,在一些实施方案中,vp1的a35、vp1的n57、vp1的g58、vp3的n382、vp3的g383、vp3的n511、vp3的g512、vp3的n715或vp3的g716被取代。在一些实施方案中,取代的氨基酸以比亲本vp1和/或vp3的氨基酸残基更高的频率被脱酰胺化,例如如本文所述。在一些实施方案中,取代vp1的a35、vp1的n57、vp1的g58、vp3的n382、vp3的g383、vp3的n511、vp3的g512、vp3的n715和/或vp3的g716将raav颗粒的稳定性改善了至少约5%、至少约10%、至少约15%、至少约20%、至少约25%、至少约30%、至少约35%、至少约40%、至少约45%、至少约50%、至少约55%、至少约60%、至少约65%、至少约70%、至少约75%、至少约80%、至少约85%、至少约90%、至少约95%或至少约100%。在一些实施方案中,可以将具有取代的vp1的a35、vp1的n57、vp1的g58、vp3的n382、vp3的g383、vp3的n511、vp3的g512、vp3的n715或vp3的g716的raav颗粒的稳定性与例如相同血清型的野生型或亲本aav衣壳进行比较。例如,在一些实施方案中,例如与包含野生型衣壳的raav颗粒的稳定性相比,取代vp1的a35、vp1的n57、vp1的g58、vp3的n382、vp3的g383、vp3的n511、vp3的g512、vp3的n715和/或vp3的g716将raav颗粒的稳定性改善了约10%至约100%、约20%至约100%、约30%至约100%、约40%至约100%、约50%至约100%、约60%至约100%、约70%至约100%、约80%至约100%、约90%至约100%、约10%至约90%、约20%至约90%、约30%至约90%、约40%至约90%、约50%至约90%、约60%至约90%、约70%至约90%、约80%至约90%、约10%至约80%、约20%至约80%、约30%至约80%、约40%至约80%、约50%至约80%、约60%至约80%、约70%至约80%、约10%至约70%、约20%至约70%、约30%至约70%、约40%至约70%、约50%至约70%、约60%至约70%、约10%至约60%、约20%至约60%、约30%至约60%、约40%至约60%、约50%至约60%、约10%至约50%、约20%至约50%、约30%至约50%、约40%至约50%、约10%至约40%、约20%至约40%、约30%至约40%、约10%至约30%、约20%至约30%或约10%至约20%中的任何一个。aav颗粒稳定性可以使用本领域中已知的多种测定来测量,包括但不限于差示扫描荧光(dsf)、差示扫描量热法(dsc)、其他热变性测定、对蛋白水解的敏感性、观察变性的成像或结构分析(例如,使用电子显微术)、转导效率或在特定温度(例如,室温或4℃,用于热稳定性)下保持指定的时间间隔或在特定ph(例如,ph稳定性)下处理的aav颗粒组合物的另一种功能测定等。本公开文本的其他方面涉及改善raav颗粒的组装的方法。在一些实施方案中,该方法包括取代vp1和/或vp3的改变脱酰胺化的氨基酸,例如如本文所述。例如,在一些实施方案中,vp1的a35、vp1的n57、vp1的g58、vp3的n382、vp3的g383、vp3的n511、vp3的g512、vp3的n715或vp3的g716被取代。在一些实施方案中,取代的氨基酸以比亲本vp1和/或vp3的氨基酸残基更高的频率被脱酰胺化,例如如本文所述。在一些实施方案中,取代vp1的a35、vp1的n57、vp1的g58、vp3的n382、vp3的g383、vp3的n511、vp3的g512、vp3的n715和/或vp3的g716将raav颗粒的组装改善了至少约5%、至少约10%、至少约15%、至少约20%、至少约25%、至少约30%、至少约35%、至少约40%、至少约45%、至少约50%、至少约55%、至少约60%、至少约65%、至少约70%、至少约75%、至少约80%、至少约85%、至少约90%、至少约95%或至少约100%。在一些实施方案中,可以将具有取代的vp1的a35、vp1的n57、vp1的g58、vp3的n382、vp3的g383、vp3的n511、vp3的g512、vp3的n715或vp3的g716的raav颗粒的稳定性与例如相同血清型的野生型或亲本aav衣壳进行比较。例如,在一些实施方案中,例如与包含野生型衣壳的raav颗粒的组装相比,取代vp1的a35、vp1的n57、vp1的g58、vp3的n382、vp3的g383、vp3的n511、vp3的g512、vp3的n715和/或vp3的g716将raav颗粒的组装改善了约10%至约100%、约20%至约100%、约30%至约100%、约40%至约100%、约50%至约100%、约60%至约100%、约70%至约100%、约80%至约100%、约90%至约100%、约10%至约90%、约20%至约90%、约30%至约90%、约40%至约90%、约50%至约90%、约60%至约90%、约70%至约90%、约80%至约90%、约10%至约80%、约20%至约80%、约30%至约80%、约40%至约80%、约50%至约80%、约60%至约80%、约70%至约80%、约10%至约70%、约20%至约70%、约30%至约70%、约40%至约70%、约50%至约70%、约60%至约70%、约10%至约60%、约20%至约60%、约30%至约60%、约40%至约60%、约50%至约60%、约10%至约50%、约20%至约50%、约30%至约50%、约40%至约50%、约10%至约40%、约20%至约40%、约30%至约40%、约10%至约30%、约20%至约30%或约10%至约20%中的任何一个。aav颗粒组装可以使用本领域中已知的多种测定来测量,包括但不限于测量颗粒产生量和/或速率、定量衣壳产生(例如,使用本文所述的任何方法纯化后)、测定完全型载体与空衣壳的产生、测量转导效率、观察颗粒形成的成像或结构分析(例如,使用电子显微术)、aav衣壳蛋白的产生(例如,如通过western印迹测定)等。本公开文本的其他方面涉及改善raav颗粒的转导的方法。在一些实施方案中,该方法包括取代vp1和/或vp3的改变脱酰胺化的氨基酸,例如如本文所述。例如,在一些实施方案中,vp1的a35、vp1的n57、vp1的g58、vp3的n382、vp3的g383、vp3的n511、vp3的g512、vp3的n715或vp3的g716被取代。在一些实施方案中,取代的氨基酸以比亲本vp1和/或vp3的氨基酸残基更高的频率被脱酰胺化,例如如本文所述。在一些实施方案中,取代vp1的a35、vp1的n57、vp1的g58、vp3的n382、vp3的g383、vp3的n511、vp3的g512、vp3的n715和/或vp3的g716将raav颗粒的转导改善了至少约5%、至少约10%、至少约15%、至少约20%、至少约25%、至少约30%、至少约35%、至少约40%、至少约45%、至少约50%、至少约55%、至少约60%、至少约65%、至少约70%、至少约75%、至少约80%、至少约85%、至少约90%、至少约95%或至少约100%。在一些实施方案中,可以将具有取代的vp1的a35、vp1的n57、vp1的g58、vp3的n382、vp3的g383、vp3的n511、vp3的g512、vp3的n715或vp3的g716的raav颗粒的稳定性与例如相同血清型的野生型或亲本aav衣壳进行比较。例如,在一些实施方案中,例如与包含野生型衣壳的raav颗粒的转导相比,取代vp1的a35、vp1的n57、vp1的g58、vp3的n382、vp3的g383、vp3的n511、vp3的g512、vp3的n715和/或vp3的g716将raav颗粒的转导改善了约10%至约100%、约20%至约100%、约30%至约100%、约40%至约100%、约50%至约100%、约60%至约100%、约70%至约100%、约80%至约100%、约90%至约100%、约10%至约90%、约20%至约90%、约30%至约90%、约40%至约90%、约50%至约90%、约60%至约90%、约70%至约90%、约80%至约90%、约10%至约80%、约20%至约80%、约30%至约80%、约40%至约80%、约50%至约80%、约60%至约80%、约70%至约80%、约10%至约70%、约20%至约70%、约30%至约70%、约40%至约70%、约50%至约70%、约60%至约70%、约10%至约60%、约20%至约60%、约30%至约60%、约40%至约60%、约50%至约60%、约10%至约50%、约20%至约50%、约30%至约50%、约40%至约50%、约10%至约40%、约20%至约40%、约30%至约40%、约10%至约30%、约20%至约30%或约10%至约20%中的任何一个。aav颗粒转导可以使用本领域中已知的多种测定来测量,包括但不限于本文所述的转导效率测定。在一些方面,本发明提供了包含重组自身互补的基因组(例如,自身互补的(self-complementary或self-complementing)raav载体)的病毒颗粒。具有自身互补的载体基因组的aav病毒颗粒和使用自身互补的aav基因组的方法描述在美国专利号6,596,535;7,125,717;7,465,583;7,785,888;7,790,154;7,846,729;8,093,054;和8,361,457;以及wangz.等人,(2003)genether10:2105-2111中,将其各自通过引用以其整体并入本文。包含自身互补的基因组的raav将借助其部分互补的序列(例如,异源核酸的互补编码链和非编码链)迅速形成双链dna。在一些实施方案中,载体包含编码异源核酸的第一核酸序列和编码该核酸的互补序列的第二核酸序列,其中第一核酸序列可以与第二核酸序列沿着其大部分或所有长度形成链内碱基对。在一些实施方案中,第一异源核酸序列和第二异源核酸序列通过突变的itr(例如,正确的itr)连接。在一些实施方案中,itr包含多核苷酸序列5'-cactccctctctgcgcgctcgctcgctcactgaggccgggcgaccaaaggtcgcccacgcccgggctttgcccgggcg–3'(seqidno:8)。突变的itr包含含末端解析序列的d区的缺失。因此,在复制aav病毒基因组时,rep蛋白将不会在突变的itr处切割病毒基因组,并且因此,以5'至3'顺序包含以下的重组病毒基因组将被包装在病毒衣壳中:aavitr、包括调节序列的第一异源多核苷酸序列、突变的aavitr、与第一异源多核苷酸反向的第二异源多核苷酸和第三aavitr。使用不同的aav血清型优化特定靶细胞的转导或靶向特定靶组织(例如,病变组织)内的特定细胞类型。raav颗粒可以包含相同血清型或混合血清型的病毒蛋白和病毒核酸。例如,raav颗粒可以含有衍生自相同aav血清型的一个或多个itr和衣壳,或者raav颗粒可以含有衍生自与raav颗粒衣壳不同的aav血清型的一个或多个itr。在一些实施方案中,aav衣壳包含突变,例如衣壳包含突变型衣壳蛋白。在一些实施方案中,突变是酪氨酸突变或肝素结合突变。在一些实施方案中,突变型衣壳蛋白保留形成aav衣壳的能力。在一些实施方案中,raav颗粒包含aav2或aav5酪氨酸突变型衣壳(参见例如,zhongl.等人,(2008)procnatlacadsciusa105(22):7827-7832),如y444或y730中的突变(根据aav2编号)。在进一步的实施方案中,raav颗粒包含来自进化枝a-f的aav血清型的衣壳蛋白(gao等人,j.virol.2004,78(12):6381)。在一些实施方案中,衣壳蛋白包含在与硫酸肝素蛋白多糖相互作用的一个或多个位置处或在对应于氨基酸484、487、527、532、585或588的一个或多个位置处的一个或多个氨基酸取代,编号基于aav2的vp1编号。在本领域中已知硫酸乙酰肝素蛋白多糖(hspg)充当aav2颗粒的细胞受体(summerford,c.和samulski,r.j.(1998)j.virol.72(2):1438-45)。在细胞膜处aav2颗粒与hspg之间的结合用于将颗粒附着到细胞上。其他细胞表面蛋白(如成纤维细胞生长因子受体和αvβ5整联蛋白)也可以促进细胞感染。结合后,aav2颗粒可以通过包括经由网格蛋白包被的凹陷进行的受体介导的内吞作用在内的机制进入细胞。在内体酸化后,aav2颗粒可以从内吞囊泡释放。这允许aav2颗粒行进到核周区域,并且然后行进到细胞核。还已知aav3颗粒结合肝素(rabinowitz,j.e.等人(2002)j.virol.76(2):791-801)。已知aav2衣壳蛋白与hspg之间的结合通过碱性aav2衣壳蛋白残基与带负电荷的糖胺聚糖残基之间的静电相互作用发生(opie,sr等人,(2003)j.virol.77:6995-7006;kern,a等人,(2003)j.virol.77:11072-11081)。这些相互作用中牵涉的特定衣壳残基包括r484、r487、k527、k532、r585和r588。已经显示这些残基中的突变降低aav2与hela细胞和肝素本身的结合(opie,sr等人,(2003)j.virol.77:6995-7006;kern,a等人,(2003)j.virol.77:11072-11081;wo2004/027019a2,美国专利号7,629,322)。此外,不希望受理论束缚,据信在对应于氨基酸484、487、527、532、585或588的一个或多个残基处的一个或多个氨基酸取代(编号基于aav2的vp1编号)可以调节不与hspg结合的aav衣壳类型的转导特性,或者可以独立于它们结合hspg的能力来调节aav衣壳类型的转导特性。在一些实施方案中,该一个或多个氨基酸取代包含vp1、vp2和/或vp3的r484、r487、k527、k532、r585和/或r588位的取代,编号基于aav2的vp1。在一些实施方案中,该一个或多个氨基酸取代将raav颗粒与硫酸肝素蛋白多糖的结合降低约至少10%、约至少25%、约至少50%、约至少75%或约至少100%。在一些实施方案中,该一个或多个氨基酸取代将raav颗粒与硫酸肝素蛋白多糖的结合降低约至少10%、约至少15%、约至少20%、约至少25%、约至少30%、约至少35%、约至少40%、约至少45%、约至少50%、约至少55%、约至少60%、约至少65%、约至少70%、约至少75%、约至少80%、约至少85%、约至少90%、约至少95%或约至少100%(与包含野生型衣壳的raav颗粒的结合相比)。在一些实施方案中,该一个或多个氨基酸取代将raav颗粒与硫酸肝素蛋白多糖的结合降低约10%至约100%、约20%至约100%、约30%至约100%、约40%至约100%、约50%至约100%、约60%至约100%、约70%至约100%、约80%至约100%、约90%至约100%、约10%至约90%、约20%至约90%、约30%至约90%、约40%至约90%、约50%至约90%、约60%至约90%、约70%至约90%、约80%至约90%、约10%至约80%、约20%至约80%、约30%至约80%、约40%至约80%、约50%至约80%、约60%至约80%、约70%至约80%、约10%至约70%、约20%至约70%、约30%至约70%、约40%至约70%、约50%至约70%、约60%至约70%、约10%至约60%、约20%至约60%、约30%至约60%、约40%至约60%、约50%至约60%、约10%至约50%、约20%至约50%、约30%至约50%、约40%至约50%、约10%至约40%、约20%至约40%、约30%至约40%、约10%至约30%、约20%至约30%或约10%至约20%中的任何一个(与包含野生型衣壳的raav颗粒的结合相比)。在一些实施方案中,与野生型raav颗粒的结合相比,该一个或多个氨基酸取代导致raav颗粒与硫酸肝素蛋白多糖没有可检测的结合。测量aav颗粒与hspg结合的手段在本领域中是已知的;例如,与硫酸肝素色谱介质结合或与已知在其表面表达hspg的细胞结合。例如,参见opie,sr等人,(2003)j.virol.77:6995-7006和kern,a等人,(2003)j.virol.77:11072-11081。在一些实施方案中,该一个或多个氨基酸取代将raav颗粒到细胞(例如,眼睛或cns中的细胞)的转导效率改善了约至少10%、约至少15%、约至少20%、约至少25%、约至少30%、约至少35%、约至少40%、约至少45%、约至少50%、约至少55%、约至少60%、约至少65%、约至少70%、约至少75%、约至少80%、约至少85%、约至少90%、约至少95%或约至少100%(与包含野生型衣壳的raav颗粒的转导效率相比)。在一些实施方案中,该一个或多个氨基酸取代将raav颗粒到细胞(例如,眼睛或cns中的细胞)的转导效率改善了约10%至约100%、约20%至约100%、约30%至约100%、约40%至约100%、约50%至约100%、约60%至约100%、约70%至约100%、约80%至约100%、约90%至约100%、约10%至约90%、约20%至约90%、约30%至约90%、约40%至约90%、约50%至约90%、约60%至约90%、约70%至约90%、约80%至约90%、约10%至约80%、约20%至约80%、约30%至约80%、约40%至约80%、约50%至约80%、约60%至约80%、约70%至约80%、约10%至约70%、约20%至约70%、约30%至约70%、约40%至约70%、约50%至约70%、约60%至约70%、约10%至约60%、约20%至约60%、约30%至约60%、约40%至约60%、约50%至约60%、约10%至约50%、约20%至约50%、约30%至约50%、约40%至约50%、约10%至约40%、约20%至约40%、约30%至约40%、约10%至约30%、约20%至约30%或约10%至约20%中的任何一个(与包含野生型衣壳的raav颗粒的转导效率相比)。测量aav颗粒到细胞(例如,培养物或组织的一部分中的细胞)的转导效率的手段在本领域中是已知的。例如,可以用一定浓度的含有载体的raav颗粒感染细胞群(例如,在培养物或组织的一部分中),该载体在细胞中表达时产生可测定的报道分子(例如,gfp荧光、sflt产生等)。aav衣壳蛋白在一些方面,本发明提供了aav衣壳蛋白,其包含在氨基酸残基2处的氨基酸取代;其中与亲本aav衣壳蛋白的氨基酸残基2处的n末端乙酰化相比,氨基酸残基2处的氨基酸取代改变了n末端乙酰化。在一些实施方案中,aav衣壳蛋白是vp1或vp3。在一些实施方案中,aav衣壳蛋白(例如,vp1或vp3)的氨基酸残基2被cys、ser、thr、val、gly、asn、asp、glu、ile、leu、phe、gln、lys、met、pro或tyr取代。在一些实施方案中,氨基酸取代导致aav衣壳蛋白的更少脱酰胺化。本发明的aav衣壳蛋白的非限制性例子包括以下任何aav血清型的vp1和/或vp3:aav1、aav2、aav3、aav4、aav5、aav6、aav7、aav8、aavrh8、aavrh8r、aav9、aav10、aavrh10、aav11、aav12、aavlk03、aav2r471a、aav2/2-7m8、aavdj、aavdj8、aav2n587a、aav2e548a、aav2n708a、aavv708k、山羊aav、aav1/aav2嵌合体、牛aav、小鼠aav或raav2/hbov1血清型衣壳。在一些实施方案中,aav衣壳还包含酪氨酸突变或肝素结合突变。aav颗粒的产生在本领域中已知许多方法用于产生raav载体,包括转染、稳定细胞系生产和感染性杂交病毒生产系统,该系统包括腺病毒-aav杂合体、疱疹病毒-aav杂合体(conway,je等人,(1997)j.virology71(11):8780-8789)和杆状病毒-aav杂合体(urabe,m.等人,(2002)humangenetherapy13(16):1935-1943;kotin,r.(2011)hummolgenet.20(r1):r2–r6)。用于产生raav病毒颗粒的raav生产培养物都需要:1)合适的宿主细胞;2)合适的辅助病毒功能;3)aavrep和cap基因和基因产物;4)侧接至少一个aavitr序列(例如,编码gnptab的aav基因组)的核酸(如治疗性核酸);以及5)支持raav产生的合适的培养基和培养基组分。在一些实施方案中,合适的宿主细胞是灵长类动物宿主细胞。在一些实施方案中,合适的宿主细胞是人源细胞系,如hela、a549、293或perc.6细胞。在一些实施方案中,合适的辅助病毒功能由野生型或突变型腺病毒(如温度敏感性腺病毒)、疱疹病毒(hsv)、杆状病毒或提供辅助功能的质粒构建体提供。在一些实施方案中,aavrep和cap基因产物可以来自任何aav血清型。通常但不是必须的,aavrep基因产物与raav载体基因组的itr具有相同血清型,只要rep基因产物可以发挥复制和包装raav基因组的作用即可。本领域中已知的合适的培养基可以用于产生raav载体。这些培养基包括但不限于hyclonelaboratories和jrh生产的培养基,包括改良伊格尔培养基(mem);达尔伯克改良伊格尔培养基(dmem);定制制剂,如美国专利号6,566,118描述的那些;以及如美国专利号6,723,551中描述的sf-900iisfm培养基,将每个专利(特别是关于用于产生重组aav载体的定制培养基制剂)均通过引用以其整体并入本文。在一些实施方案中,aav辅助功能由腺病毒或hsv提供。在一些实施方案中,aav辅助功能由杆状病毒提供,并且宿主细胞是昆虫细胞(例如,草地贪夜蛾(spodopterafrugiperda)(sf9)细胞)。在一些实施方案中,aavcap功能提供了在vp1和/或vp3的氨基酸残基2处的氨基酸取代,其中与亲本aav颗粒的vp1和/或vp3的氨基酸残基2处的n末端乙酰化相比,vp1和/或vp3的氨基酸残基2处的氨基酸取代改变了n末端乙酰化。在一些实施方案中,aav衣壳蛋白(例如,vp1或vp3)的氨基酸残基2被cys、ser、thr、val、gly、asn、asp、glu、ile、leu、phe、gln、lys、met、pro或tyr取代。在一些实施方案中,氨基酸取代导致aav衣壳的更少脱酰胺化。用于产生raav颗粒的一种方法是三重转染方法。简而言之,可以将含rep基因和衣壳基因的质粒连同辅助腺病毒质粒转染(例如,利用磷酸钙法)到细胞系(例如,hek-293细胞)中,并且可以收集并任选地纯化病毒。因此,在一些实施方案中,通过将编码raav载体的核酸、编码aavrep和cap的核酸以及编码aav辅助病毒功能的核酸三重转染到宿主细胞中来产生raav颗粒,其中核酸向宿主细胞的转染产生能够产生raav颗粒的宿主细胞。在一些实施方案中,raav颗粒可以通过生产细胞系方法产生(参见martin等人,(2013)humangenetherapymethods24:253-269;美国专利授予前公开号us2004/0224411;和liu,x.l.等人(1999)genether.6:293-299)。简而言之,可以用含有rep基因、衣壳基因和包含启动子-异源核酸序列(例如,gnptab)的载体基因组的质粒稳定地转染细胞系(例如,hela、293、a549偶perc.6细胞系)。可以筛选细胞系,以选择用于raav产生的前导克隆(leadclone),然后可以将其扩增至生产反应器,并且用辅助病毒(例如,腺病毒或hsv)转染,以启动raav产生。随后可以收获病毒,可以使腺病毒失活(例如,通过加热)和/或去除,并且可以纯化raav颗粒。因此,在一些实施方案中,通过包含编码raav载体的核酸、编码aavrep和cap的核酸和编码aav辅助病毒功能的核酸中的一种或多种的生产细胞系产生raav颗粒。如本文所述的,与三重转染方法相比,生产细胞系方法对于产生具有过大基因组的raav颗粒而言可能是有利的。在一些实施方案中,编码aavrep和cap基因和/或raav基因组的核酸稳定地维持在生产细胞系中。在一些实施方案中,将编码aavrep和cap基因和/或raav基因组的核酸在一种或多种质粒上引入细胞系中以产生生产细胞系。在一些实施方案中,将aavrep、aavcap和raav基因组在相同质粒上引入细胞中。在其他实施方案中,将aavrep、aavcap和raav基因组在不同质粒上引入细胞中。在一些实施方案中,用质粒稳定地转染的细胞系维持质粒用于细胞系的多次传代(例如,5、10、20、30、40、50或超过50次细胞传代)。例如,该一种或多种质粒可以在细胞复制时复制,或者该一种或多种质粒可以整合到细胞基因组中。已经鉴定了使质粒能够在细胞(例如,人细胞)中自主复制的多种序列(参见例如,krysan,p.j.等人(1989)mol.cellbiol.9:1026-1033)。在一些实施方案中,该一种或多种质粒可以含有允许选择维持质粒的细胞的选择标记(例如,抗生素抗性标记)。通常用于哺乳动物细胞的选择标记包括但不限于杀稻瘟素、g418、潮霉素b、博莱霉素、嘌呤霉素及其衍生物。用于将核酸引入细胞中的方法在本领域中是已知的,并且包括但不限于病毒转导、阳离子转染(例如,使用阳离子聚合物如deae-葡聚糖或阳离子脂质如lipofectamine)、磷酸钙转染、显微注射、粒子轰击、电穿孔和纳米颗粒转染(关于更多细节,参见例如,kim,t.k.和eberwine,j.h.(2010)anal.bioanal.chem.397:3173-3178)。在一些实施方案中,编码aavrep和cap基因和/或raav基因组的核酸稳定地整合到生产细胞系的基因组中。在一些实施方案中,将编码aavrep和cap基因和/或raav基因组的核酸在一种或多种质粒上引入细胞系中以产生生产细胞系。在一些实施方案中,将aavrep、aavcap和raav基因组在相同质粒上引入细胞中。在其他实施方案中,将aavrep、aavcap和raav基因组在不同质粒上引入细胞中。在一些实施方案中,该一种或多种质粒可以含有允许选择维持质粒的细胞的选择标记(例如,抗生素抗性标记)。用于将核酸稳定地整合到多种宿主细胞系中的方法在本领域中是已知的。例如,重复选择(例如,通过使用选择标记)可以用于选择已整合含有选择标记(和aavcap和rep基因和/或raav基因组)的核酸的细胞。在其他实施方案中,核酸能以位点特异性方式整合到细胞系中以产生生产细胞系。在本领域中已知几种位点特异性重组系统,如flp/frt(参见例如,o'gorman,s.等人(1991)science251:1351-1355)、cre/loxp(参见例如,sauer,b.和henderson,n.(1988)proc.natl.acad.sci.85:5166-5170)和phic31-att(参见例如,groth,a.c.等人(2000)proc.natl.acad.sci.97:5995-6000)。在一些实施方案中,生产细胞系衍生自灵长类动物细胞系(例如,非人灵长类动物细胞系,如vero或frhl-2细胞系)。在一些实施方案中,细胞系衍生自人细胞系。在一些实施方案中,生产细胞系衍生自hela、293、a549或(crucell)细胞。例如,在将编码aavrep和cap基因和/或过大的raav基因组的核酸引入和/或稳定维持/整合到细胞系中以产生生产细胞系之前,细胞系是hela、293、a549或(crucell)细胞系或其衍生物。在一些实施方案中,生产细胞系适于在悬浮液中生长。如在本领域中已知的,锚定依赖性细胞通常不能在没有基质(如微载体珠)的情况下在悬浮液中生长。使细胞系适于在悬浮液中生长可以包括例如使用搅拌桨在旋转培养基中培养细胞系,使用缺少钙和镁离子的培养基以防止结块(和任选地消泡剂),使用包被有渗硅化合物的培养容器,并且在每次传代时选择培养物中(而不是在大块中或在容器的侧面)的细胞。关于进一步的描述,参见例如,atcc常见问题文件(可在万维网的atcc.org/global/faqs/9/1/adapting%20a%20monolayer%20cell%20line%20to%20suspension-40.aspx处获得)和其中引用的参考文献。本发明的合适的aav生产培养基可以补充有0.5%-20%(v/v或w/v)水平的血清或血清衍生的重组蛋白。可替代地,如在本领域中已知的,aav载体可以在无血清条件下产生,该条件也可以称为不含动物源产品的培养基。本领域普通技术人员可以理解,设计用于支持aav载体产生的商业或定制培养基还可以补充有本领域中已知的一种或多种细胞培养组分,包括但不限于葡萄糖、维生素、氨基酸和/或生长因子,以便提高生产培养物中aav的滴度。aav生产培养物可以在适合于所用特定宿主细胞的多种条件下(在很宽的温度范围内、持续不同的时间长度等)生长。如在本领域中已知的,aav生产培养物包括附着依赖性培养物,其可以在合适的附着依赖性容器(例如像,滚瓶、中空纤维过滤器、微载体和填充床或流化床生物反应器)中培养。aav载体生产培养物还可以包括悬浮适应的宿主细胞(如hela、293和sf-9细胞),其能以多种方式(包括例如旋转瓶、搅拌槽生物反应器和一次性系统如wave袋系统)培养。本发明的aav载体颗粒可以通过裂解生产培养物的宿主细胞或通过从生产培养物中收获用过的培养基而从aav生产培养物中收获,条件是细胞在本领域中已知的引起aav颗粒从完整细胞释放到培养基中的条件下培养,如美国专利号6,566,118中更全面地描述的。裂解细胞的合适方法在本领域中也是已知的,并且包括例如多次冷冻/解冻循环、超声处理、微流化和用化学品(如洗涤剂和/或蛋白酶)处理。在一个进一步的实施方案中,纯化aav颗粒。如本文所用的术语“纯化的”包括aav颗粒的如下制剂,其缺乏至少一些也可以存在于aav颗粒天然存在或最初所制备的地方的其他组分。因此,例如,分离的aav颗粒可以使用纯化技术使其从源混合物(如培养裂解物或生产培养上清液)富集而制备。能以多种方式测量富集情况,例如像根据溶液中存在的dna酶抗性颗粒(drp)或基因组拷贝(gc)的比例、或根据感染性,或者可以根据源混合物中存在的第二潜在干扰物质(如污染物,包括生产培养污染物或进程内污染物,包括辅助病毒、培养基组分等)来测量。在一些实施方案中,澄清aav生产培养收获物以除去宿主细胞碎片。在一些实施方案中,通过经由一系列深度过滤器过滤来澄清生产培养收获物,该深度过滤器包括例如dohcmilliporemillistak+hcpod级过滤器、a1hcmilliporemillistak+hcpod级过滤器和0.2μmfilteropticapxl1omilliporeexpressshchydrophilicmembrane过滤器。澄清也可以通过本领域中已知的多种其他标准技术来实现,如离心或通过本领域中已知的0.2μm或更大孔径的任何醋酸纤维素过滤器过滤。在一些实施方案中,用进一步处理aav生产培养收获物以消化生产培养物中存在的任何高分子量dna。在一些实施方案中,消化在本领域中已知的标准条件下进行,包括例如1-2.5单位/ml的的终浓度,在范围从环境温度至37℃的温度下持续30分钟至几小时。可以使用以下一个或多个纯化步骤分离或纯化aav颗粒:平衡离心;流过式阴离子交换过滤;用于浓缩aav颗粒的切向流过滤(tff);通过磷灰石色谱捕获aav;辅助病毒的热灭活;通过疏水相互作用色谱捕获aav;通过尺寸排阻色谱(sec)进行缓冲液交换;纳滤;以及通过阴离子交换色谱、阳离子交换色谱或亲和色谱捕获aav。这些步骤可以单独使用,以各种组合使用,或者以不同顺序使用。在一些实施方案中,该方法以如下所述的顺序包括所有步骤。纯化aav颗粒的方法见于例如xiao等人,(1998)journalofvirology72:2224-2232;美国专利号6,989,264和8,137,948;以及wo2010/148143。药物组合物在一些实施方案中,本公开文本的aav颗粒(例如,raav颗粒)在药物组合物中。药物组合物可以适用于本文所述的或在本领域中已知的任何给药方式。在一些实施方案中,药物组合物包含经修饰以改善raav颗粒的稳定性和/或改善转导效率的raav颗粒;例如,用于取代vp1和/或vp3的2位的氨基酸残基以改善raav衣壳蛋白的乙酰化。在一些实施方案中,药物组合物包含经修饰以调节raav颗粒的稳定性和/或转导效率(例如,提高稳定性和/或转导效率或降低稳定性和/或转导效率)的raav颗粒;例如,用于取代调节脱酰胺化(例如,增强脱酰胺化或降低脱酰胺化)的氨基酸残基。在一些实施方案中,raav颗粒在包含药学上可接受的赋形剂的药物组合物中。如在本领域中众所周知的,药学上可接受的赋形剂是相对惰性的物质,其促进药理学有效物质的给药,并且可以作为液体溶液或悬浮液、作为乳液或作为适用于在使用前溶解或悬浮在液体中的固体形式提供。例如,赋形剂可以赋予形式或稠度,或充当稀释剂。合适的赋形剂包括但不限于稳定剂、润湿剂和乳化剂、用于改变渗透压的盐、包封剂、ph缓冲物质和缓冲剂。此类赋形剂包括适用于直接递送至眼睛的任何药剂,其可以在没有过度毒性的情况下给予。药学上可接受的赋形剂包括但不限于山梨糖醇、多种tween化合物中的任何一种、以及诸如水、盐水、甘油和乙醇等液体。其中可以包括药学上可接受的盐,例如矿物酸盐,如盐酸盐、氢溴酸盐、磷酸盐、硫酸盐等;和有机酸盐,如乙酸盐、丙酸盐、丙二酸盐、苯甲酸盐等。药学上可接受的赋形剂的透彻讨论可在remington’spharmaceuticalsciences(mackpub.co.,新泽西州1991)中获得。在一些实施方案中,包含本文所述的raav颗粒和药学上可接受的载体的药物组合物适用于给予给人。此类载体在本领域中是众所周知的(参见例如,remington'spharmaceuticalsciences,第15版,第1035-1038页和第1570-1580页)。此类药学上可接受的载体可以是无菌液体,如水和油,该油包括石油、动物、植物或合成源的那些油,如花生油、大豆油、矿物油等。盐水溶液和右旋糖水溶液、聚乙二醇(peg)和甘油溶液也可以用作液体载体,特别是用于可注射溶液。药物组合物还可以包含另外的成分,例如防腐剂、缓冲剂、张力剂、抗氧化剂和稳定剂、非离子润湿剂或澄清剂、增粘剂等。本文所述的药物组合物可以按单一单位剂量或按多剂量形式包装。通常将组合物配制成无菌且基本上等渗的溶液。试剂盒和制品本发明还提供了包含本公开文本的任何raav颗粒和/或药物组合物的试剂盒或制品。试剂盒或制品可以包含本发明的任何raav颗粒或raav颗粒组合物。在一些实施方案中,使用试剂盒改善raav颗粒的稳定性和/或改善转导效率;例如,用于取代vp1和/或vp3的2位的氨基酸残基以改善raav衣壳蛋白的乙酰化。在一些实施方案中,使用试剂盒调节raav颗粒的稳定性和/或转导效率(例如,提高稳定性和/或转导效率或降低稳定性和/或转导效率);例如,用于取代调节脱酰胺化(例如,增强脱酰胺化或降低脱酰胺化)的氨基酸残基。在一些实施方案中,试剂盒或制品还包括用于给予raav颗粒组合物的说明书。本文所述的试剂盒或制品还可以包括从商业和使用者角度所需的其他材料,包括其他缓冲剂、稀释剂、过滤器、针头、注射器和具有用于实施本文所述的任何方法的说明的包装说明书。还可以包括合适的包装材料,并且其可以是本领域中已知的任何包装材料,包括例如小瓶(如密封小瓶)、器皿、安瓿、瓶子、罐子、软包装(例如,密封的聚酯薄膜(mylar)或塑料袋)等。可以进一步将这些制品灭菌和/或密封。在一些实施方案中,试剂盒或制品还含有一种或多种本文所述的缓冲剂和/或药学上可接受的赋形剂(例如,如remington’spharmaceuticalsciences(mackpub.co.,新泽西州1991)中所述)。在一些实施方案中,试剂盒或制品包括一种或多种本文所述的药学上可接受的赋形剂、载体、溶液和/或另外的成分。本文所述的试剂盒或制品可以按单一单位剂量或按多剂量形式包装。试剂盒或制品的内容物通常配制成无菌的并且可以冻干或作为基本上等渗的溶液提供。实施例通过参考以下实施例将更全面地理解本发明。然而,它们不应被解读为限制本发明的范围。应当理解,本文所述的实施例和实施方案仅用于说明目的,并且根据它们进行的各种修改或改变将为本领域技术人员知晓,并且应包括在本申请的精神和范围内以及所附权利要求的范围内。实施例1:用于重组aav病毒衣壳蛋白的完整表征的直接lc/ms和lc/ms/ms由于其非致病性、感染分裂和非分裂细胞两者的能力以及长期基因表达,重组腺相关病毒(raav)已经成为受欢迎的基因治疗载体。目前,基于aav的基因治疗用于许多疾病靶标(如肌营养不良、血友病、帕金森病、利伯氏先天性黑蒙和黄斑变性)的临床试验。aav是一种小且无包膜的细小病毒,其单链dna基因组衣壳化于二十面体壳中。每个衣壳包括六十个拷贝的三种病毒衣壳蛋白vp1(87kda)、vp2(73kda)和vp3(62kda),比率大约为1:1:10。通过使用选择性剪接和非典型起始密码子从相同的开放阅读框表达三种病毒衣壳蛋白,并且因此其具有重叠序列。与vp3相比,vp1具有约137个另外的n末端氨基酸残基;而与vp3相比,vp2具有约65个另外的n末端氨基酸残基。已经从人类和非人类灵长类动物组织中分离出至少13种aav血清型和约150个基因序列;aav血清型在病毒衣壳蛋白的氨基酸序列及其用于靶向的对应细胞受体和共受体上不同。除了保护内部基因组外,aav衣壳还在介导受体结合、病毒从内体逃逸以及病毒dna在病毒感染周期中转运到细胞核中起重要作用,从而直接影响病毒感染性。已经显示vp1n末端含有磷脂酶pla2结构域(a.a.52-97),其在病毒的内体逃逸中是至关重要的[1-3]。vp1和vp2的n末端还含有三个碱性氨基酸簇作为核定位信号。这些序列在不同的aav血清型中是高度保守的。已经显示这些氨基酸的突变降低或完全消除感染性[4]。另外,每种aav血清型具有对应的序列特异性受体和共受体。例如,硫酸肝素蛋白多糖被鉴定为aav2的主要受体,并且已经鉴定了几种其他共受体,包括αvβ5整联蛋白、成纤维细胞生长因子受体1和肝细胞生长受体[5-8]。aav2衣壳蛋白的突变分析已经鉴定出一组碱性氨基酸(精氨酸484、487、585和赖氨酸532)作为肝素结合基序,其有助于肝素和hela细胞结合[9]。aav2中的ngr结构域被鉴定为整联蛋白α5β1结合结构域,其对于病毒细胞进入而言是必需的[10]。总之,病毒衣壳蛋白序列在病毒感染周期中的细胞靶向和运输中是重要的。由于不同的生产条件可能导致病毒衣壳蛋白的不同表达水平、翻译后修饰和截短,因此需要对病毒衣壳蛋白进行表征和监测,以确保基因治疗开发计划中的产品一致性。传统上,sds-page已经用于表征aav病毒衣壳蛋白,提供粗略的分子量信息,如87kda、73kda和62kda。没有从edman测序获得序列信息,可能是由于病毒衣壳蛋白的n末端被阻断,vp2除外。虽然已经解决了多种aav的x射线结构,但在晶体结构中仅观察到vp3区序列。vp3的十五个n末端氨基酸残基在x射线结构中仍缺失,可能是由于其内在无序[11-13]。有可能在原子结构中缺乏vp1和vp2n末端区域的信息可能是由于衣壳中vp1和vp2的化学计量低。另外,如一些文献中所报道的,vp1和vp2的n末端被埋在衣壳内并且不能被天然状态的抗体接近[3,14,15]。常规地,将凝胶-lc/ms方法(sds-page、凝胶内胰蛋白酶消化和lc/ms/ms)用于表征vp[16-18]。然而,使用这种方法尚未确认vp1、vp2和vp3的n末端,因为这种方法由于肽从凝胶中的有限回收而未能获得vp的100%序列覆盖率。使用maldi-tofms的直接分析报道了几种病毒衣壳蛋白,包括用有机酸解离后的烟草花叶病毒u2[19]。在酰胺氢交换和质谱后的直接肽谱分析(peptidemapping)已经用于研究ph诱导的雀麦花叶病毒(bmv)衣壳的结构变化[20]。由于aav是仅含有衣壳蛋白和基因组的无包膜病毒,因此可以通过蛋白质的rp-lc/ms和肽谱分析的lc/ms/ms直接分析aav衣壳,以在衣壳解离后实现100%序列覆盖率而无需sds-page分离。dna片段可以在空隙体积中洗脱,并且因此对lc/ms进行的蛋白质/肽检测没有干扰。为了研究这些方法,使用变性后的不同类型的aav的直接lc/ms来监测aav衣壳蛋白的蛋白质序列和翻译后修饰。如本文所述的,aav的vp1、vp2和vp3的n末端已经通过质谱得到确认。在aav的不同血清型中也鉴定了vp1和vp3的n末端的乙酰化。还开发了aav的直接lc/ms/ms肽谱分析,以提供vp1、vp2和vp3的序列覆盖并确认vp1和vp3的n末端乙酰化。方法材料和试剂二硫苏糖醇(dtt)、4-乙烯基吡啶、超纯甲酸、乙酸、盐酸胍、tris-hcl和tris碱购自sigmachemicals(密苏里州圣路易斯)。amiconultra-4过滤器购自millipore(马萨诸塞州比勒利卡)。猪测序级胰蛋白酶购自promega(威斯康星州密尔沃基)。内切蛋白酶lys-c和asp-n购自roche(德国)。具有10,000mwco的slide-a-lyzer盒购自pierce(伊利诺伊州罗克福德)。载体产生和纯化使用如前所述的瞬时三重转染方法(xiao,1998#123)产生aav载体。简而言之,使用聚乙烯亚胺、pei和1:1:1比率的三种质粒(itr载体、aavrep/cap和ad辅助质粒)转染hek293细胞。载体质粒含有载体基因组cba-egfp和来自aav2的itr序列。egfp表达由如(miyazaki,1989#124)所述的cmv增强子鸡β肌动蛋白杂合启动子(cba)驱动(niwa,1991#125)。aavrep/cap辅助质粒含有来自aav2的rep序列和血清型特异性衣壳序列,具有名称rep2/cap2、rep2/cap5、rep2/cap7等。所用的pad辅助质粒是phelper(stratagene/agilenttechnologies,加利福尼亚州圣克拉拉)。如qu等人(2007,j.virol.methods140:183-192)所述的进行aav的纯化。lc/ms完整蛋白质分析用amiconultra-4过滤器(10kdamwco)浓缩aav病毒粒子,并且用10%乙酸使其变性,然后在acquityuplc–qtofms仪器(waters,马萨诸塞州米尔福德)中直接分析。用uplcbehc4或c8柱(1.7μm,2.1mmi.d.)以0.25ml/min的流速进行分离。流动相a是0.1%甲酸水溶液,而流动相b是0.1%甲酸乙腈溶液。最终梯度如下:从10%b到20%b持续6分钟,在10min内从20%b到30%b,然后从30%到38%b持续40分钟。对于ms,毛细管电压和采样锥电压分别设定为3.5kv和45v。质谱以灵敏度模式获得,m/z范围为500-4000。用碘化钠作为校准物进行辅助校准以进行质量校准。将masslynx软件中的maxent1用于蛋白质解卷积。aav2vp的酶消化用6m盐酸胍、0.1mtris(ph8.5)使浓缩的aav2病毒粒子变性。将蛋白质用30mmdtt在55℃下在黑暗中还原1小时,并且在室温下用0.07%4-乙烯基吡啶烷基化2小时。通过添加1mdtt淬灭反应。将样品用slide-a-lyzer盒(10,000mwco)对ph8.5的25mmtris缓冲液透析约18小时。透析后,将样品分成三个等分试样。将每个等分试样分别用1:25酶:蛋白质比率(wt/wt)的胰蛋白酶或1:50酶:蛋白质比率(wt/wt)的lys-c或1:100酶:蛋白质比率(wt/wt)的asp-n在37℃下消化18小时。lc/ms/ms肽谱分析使用nanoacquityhplc系统(waters,马萨诸塞州米尔福德)结合orbitrapvelos质谱仪(thermo-fisherscientific,马萨诸塞州沃尔瑟姆)进行纳米lc/ms/ms,使用家庭包装的nanolc柱(75μmx10mm)和具有包装材料的magicc18(5μm,bruker,马萨诸塞州比勒利卡),流速为300nl/min。流动相a和b分别为0.1%甲酸水溶液和乙腈溶液。梯度在121min内从2%b到60%b。velos的源参数如下:源电压:2.5kv;毛细管温度275℃;s透镜rf电平:55%。使用前十种数据依赖性方法以精确的ms在离子阱中以60,000分辨率和10ms/ms获得数据。将mascot用于针对aav2病毒衣壳蛋白序列的数据库搜索。数据库搜索使用10ppm的ms容差和0.8da的ms/ms容差。uplc/ms/ms肽谱分析还通过uplc/ms/ms在acquityuplc-xevoqtofms中分析了蛋白质消化物。使用beh300c18柱(2.1x150mm)在流动相中以0.1%甲酸水溶液/乙腈溶液的梯度以0.25ml/min的流速进行分离。质谱以正mse模式获得,质量范围为200-2000。结果aav变性方法aav可以通过多种方法使用洗涤剂、热、高盐或具有低或高ph的缓冲液变性。热变性可以导致蛋白质沉淀,并且因此反相柱易被堵塞和过度加压。用高盐变性需要在lc/ms分析之前进行另外的脱盐步骤。将用10%乙酸变性用于lc/ms完整蛋白质分析,因为它允许干净的质谱。对于肽谱分析,可以使用0.1%rapigest或6m盐酸胍作为变性试剂。完整蛋白质分析方法开发使用uplcbehc4柱以快速梯度进行aav2的初始完整蛋白质分析。在这种条件下,仅在总离子色谱图观察到一个单峰,质量对应于vp3(图1a)。不希望受理论束缚,据信不存在vp1和vp2可能是由于vp1和vp2的化学计量低或vp3对vp1和vp2信号的抑制,如果所有vp共洗脱的话。为了检测vp1和vp2,已经尝试增加注射或柱长度、使用较浅的梯度和使用替代柱。0.5%b/min的较浅梯度的较高负载(1.7μg)导致左侧的肩峰(图1b)。柱长从10cm增加到15cm不会增强肩峰的分离(图1c)。然而,使用behc8柱将肩峰与主峰进一步分离,观察到改善的信号强度(图1d)。结果,在这个肩峰中获得了vp1和vp2质量,其信号强度如图2a所示。vp1和vp3的质量分别对应于a.a.2-735(乙酰化)和a.a.204-735(乙酰化)(图2a和2b)。在vp2中没有观察到乙酰化(a.a.139-735)。另外,观察到质量小于vp3的次峰,其质量对应于具有一个乙酰化的氨基酸序列212-735(图2b)。这些数据与dna序列一致,因为vp3在aav2中含有两个atg起始密码子:(seqidno:1),产生两个可能的n末端(加下划线):matgsgapmad(seqidno:2)。如通过完整蛋白质分析测量的,vp1和vp3两者中均不存在n末端甲硫氨酸残基。vp1和vp3的乙酰化不是方法诱导的伪像(通过10%乙酸使aav变性),因为在使用不含乙酸的替代变性方法的aav制剂中也观察到vp1和vp3的乙酰化。完整蛋白质的数据也确认病毒衣壳蛋白中不存在糖基化,即使存在几个n连接的共有序列[16]。lc/ms/ms肽谱分析为了进一步确认在完整蛋白质分析中观察到的n末端和乙酰化,使用多种酶进行肽谱分析并使用多种仪器进行分析。已经评估了多种样品制备方法,包括变性方法和脱盐步骤。最终的消化方法(包括用6m盐酸胍变性,还原和用4-乙烯基吡啶烷基化,和使用slide-a-lyzer进行透析,然后酶消化)在消化过程中产生具有低人工修饰的干净的肽谱分析。测试低至5μg的起始材料,使用纳米lc/ms/ms和uplc/ms/ms产生完整的序列覆盖。仅来自纳米lc/ms/ms的胰蛋白酶消化物的mascot搜索得到78%的序列覆盖率,其离子得分截留为13,如图3所示。在具有behc18柱的xevotofms中的lc/ms中发现了来自纳米lc/ms/ms的两种大的缺失的胰蛋白酶肽t27和t38(加框)(图3)。另外,大多数t27和t38肽序列通过asp-n消化物的纳米lc/ms/ms得到进一步确认,如图3中的斜体所示。完整的n末端和c末端肽被lys-c消化物覆盖,如图3中的下划线所示。因此,通过多种酶消化和两种lc/ms/ms方法实现了vp1的100%序列覆盖率。lc/ms/ms确认了在完整蛋白质分析中观察到的vp1、vp2和vp3的n末端和c末端以及vp1和vp3的n末端乙酰化。图4a-4c显示了vp1n末端胰蛋白酶肽a(ac)adgylpdwledtlsegir(seqidno:4)(图4a)、vp2n末端asp-n衍生肽(apgkkrpvehspvep)(seqidno:15)(图4b)和vp3n末端asp-n肽a(ac)tgsgapm(seqidno:5)(图4c)的ms/ms谱。ms/ms已经确认了vp1和vp3肽两者中n末端丙氨酸残基处的乙酰化位置。在图4a中存在未修饰的y18和y17离子以及所有检测到的具有42da质量偏移的b离子指示42da修饰位于vp1的n末端。类似地,在图4c中存在未修饰的y3至y8离子确认了n末端丙氨酸残基处的乙酰化位置。aavvpn末端的比较除了aav2外,还通过完整蛋白质分析对aav1、aav5、aav7、aav9和aavrh10进行了分析。aav中vp的理论和预测质量示于表2中。表2:aavvp的理论质量与实验质量n末端以及它们的翻译后修饰在所分析的aav血清型中是高度保守的,即使aav5被报道为最多样化的aav血清型序列,如图5中的序列比对所示。在13种aav血清型中的11种中,vp1的n末端共享相同的13个氨基酸残基的序列(maadgylpdwled)(seqidno:6),而所有13种aav血清型在vp2中具有相同的tap…n末端序列(图5)。aav2的lc/ms指示在vp2中在蛋白质水平上缺失t。vp3的n末端在三种病毒衣壳蛋白中是最多样的,其中13种aav血清型中的8种共享ma…n末端序列。与aav2类似,基于lc/ms完整蛋白质分析,aav1和aavrh10也具有两个atg起始密码子,其中第一个是主要的n末端。有趣的是,尽管aav7具有两个潜在的起始密码子(gctgcaggcggtggcgcaccagcagacaataac…)(seqidno:7),但基于完整蛋白质分析,第二个起始密码子(atg)是有利的:具有213(ac)-737的vp3`是主要峰,而具有203(ac)-737的vp3是次要峰。结论aavvp的lc/ms完整蛋白质分析和lc/ms/ms肽谱分析在基因治疗研究和开发中的应用这些结果表明,变性后的不同类型的aav的直接lc/ms被证明是一种简单有效的监测蛋白质序列和翻译后修饰的方式,并在完整蛋白质水平上进行精确的质量测量。通过质谱确认了aav的vp1、vp2和vp3的n末端。在aav的不同血清型中也鉴定了vp1和vp3的n末端的乙酰化。开发了aav的直接lc/ms/ms肽谱分析,提供了vp1、vp2和vp3的100%序列覆盖率,并且确认了vp的n末端乙酰化。基于几种aav血清型的序列比对和完整蛋白质分析的13种aav血清型的预测序列的理论质量示于表3中。表3:预测序列和质量每种血清型的vp1、vp2和vp3的精确质量是独特的,并且因此完整蛋白质分析可以用作区分aav衣壳血清型的身份测试。表4-6显示了13种常见aav血清型中vp的质量差异。以常规字体显示的是大于10的δ质量,δ质量小于10以粗体显示。表4:13种aav同种型中vp1的质量差异表5:13种aav同种型中vp2的质量差异表6:13种aav同种型中vp3的质量差异在两种同种型之间的所有三种vp中没有观察到10da内的质量。尽管vp2和vp3在aav1与aav6之间仅具有2da的差异,但aav1与aav6之间的vp1的质量差异为36,足以通过精确的质量测量来区别。因此,vp1、vp2和vp3的完整蛋白质测量作为身份测试是高度特异性的。这些结果表明,完整蛋白质分析和lc/ms/ms可以用于分析vp以监测vp表达、翻译后修饰和截短,并且确保vlp生产期间的产品一致性。这两种分析也可以用于确认衣壳蛋白工程应用的定点诱变或结构表征。实施例2:aav衣壳蛋白的n末端乙酰化的作用细胞蛋白质的化学修饰是控制其功能的常用手段(arnesen,t.(2006)virology353(2):283–293)。n末端乙酰化(nt-乙酰化)涉及将乙酰基从乙酰辅酶a转移到蛋白质的第一个氨基酸残基的α-氨基(brown,j.l.和roberts,w.k.(1976)jbiolchem251:1009–1014;arnesen,t.等人(2009)procnatlacadsciusa106:8157–8162),是最丰富的蛋白质修饰之一。与大多数其他蛋白质修饰不同,nt-乙酰化是不可逆的;它主要发生在蛋白质的合成过程中,由与核糖体相关的n末端乙酰转移酶(nat)催化(gautschi,m.等人(2003)molcellbiol23:7403–7414;pestana,a.和pitot,h.c.(1975)biochemistry14:1404–1412;polevoda,b.等人(2003)jbiolchem278:30686–97)。在真核生物中有几种不同的nat,即nata-natf,每种由一个或多个亚基组成,并且每种取决于前几个氨基酸的氨基酸序列乙酰化一个子组特定的n末端(jornvall,h.(1975)jtheorbiol55:1–12;persson,b.等人(1985)eurjbiochem152:523–527)。实验数据指示,具有乙酰化n末端的蛋白质在体内比非乙酰化蛋白质更稳定;即,nt-乙酰化保护蛋白质免于降解(hershko,a.等人(1984)procnatlacadsciusa81:7021–7025)。对此的一种解释可能是2004年发现另一种n末端修饰泛素化(其涉及将小蛋白质泛素直接连接到n末端氨基酸残基上)促进蛋白质的后续降解(bensaadon,r.等人(2004)jbiolchem279:41414–41421)。相反,nt-乙酰化信号也可以是质量控制机制的一部分,以降解未折叠或错误折叠的蛋白质并调节体内蛋白质化学计量(hwang,c.s.等人(2010)science327:973–977)。与那些注定要分选到分泌途径的蛋白质相比,预测的胞质蛋白的n末端加工的系统分析揭示,胞质蛋白深深地偏向于支持加工,但对分泌蛋白的这种修饰有相等且相反的偏向(forte,g.m.a.等人(2011)plosbiology,2011年5月4日第9卷)。导致其乙酰化的分泌信号序列中的突变以依赖于n末端加工机器的方式导致错误分选到胞质溶胶中。因此,n末端乙酰化代表新生多肽的细胞分选中的早期确定步骤,其代表额外的严格层,以便确保注定保留在胞质溶胶中的蛋白质实际上留于胞质溶胶中。真核细胞包含执行特定功能所需的几个不同的区室,称为细胞器。这些区室中的蛋白质在细胞质中合成,并且因此需要复杂的分选机制以确保其递送到适当的细胞器。蛋白质在其合成的非常早期阶段通过其氨基末端的乙酰化来修饰。对细胞质蛋白的这种修饰的可能性与那些去往主要细胞器之一内质网(er)的蛋白质的这种修饰的可能性之间存在明显差异;而细胞质蛋白通常被乙酰化,那些与er结合的蛋白质大部分未被修饰。此外,当工程化特定er蛋白以诱导其乙酰化时,其对er的靶向被抑制(forte,g.m.a.等人(2011)plosbiology,2011年5月4日第9卷)。已经显示收缩蛋白肌动蛋白和原肌球蛋白需要natb介导的nt-乙酰化才能发挥正常功能,特别是涉及肌动蛋白-原肌球蛋白结合和肌动球蛋白调节(coulton,a.t.等人(2010)jcellsci123:3235–3243;polevoda,b.等人(2003)jbiolchem278:30686–97)。因此,aav衣壳蛋白的nt乙酰化可能在raav载体的转导潜力中具有重要性。如果aav载体未能进入细胞核中,则它们因此不能转导细胞。肌动蛋白丝和fkbp52(fk506结合蛋白p52)在将aav衣壳从内体易位到细胞核中的作用是明确定义的(zhao,w.等人(2006)virology353(2):283–293)。重要的是,通过调节蛋白质-蛋白质相互作用,nt-乙酰化对于肌动蛋白丝的功能而言是必需的(coulton,a.t.等人(2010)jcellsci123:3235–3243;polevoda,b.等人(2003)jbiolchem278:30686–97)。虽然蛋白质的n末端乙酰化是一种众所周知的现象,但是对aav衣壳蛋白的nt-乙酰化的生物学意义尚不理解。基于dna测序预测的vp1和vp3的n末端都是甲硫氨酸,然后是丙氨酸。据报道,通过met-氨肽酶去除n末端甲硫氨酸经常导致所得n末端丙氨酸、缬氨酸、丝氨酸、苏氨酸和半胱氨酸残基的nt-乙酰化,并且n末端的乙酰化充当潜在的降解信号[21]。病毒衣壳蛋白的泛素化被认为是在病毒粒子解体时加工衣壳的潜在信号[22]。进一步研究vp1和vp3的n乙酰化与病毒衣壳在细胞核进入前的降解和脱壳之间的联系。为了理解关于aav衣壳蛋白的n末端乙酰化的功能意义,使用vp3n末端起始密码子的定点诱变来产生aav突变体。方法产生在起始甲硫氨酸的第2位(imetx)具有不同氨基酸的aav衣壳蛋白,以确定nt-乙酰化是否被抑制或降低,并且然后测量功能结果。评估衣壳蛋白在细胞内被运输和/或获得翻译后修饰(如糖基化)的能力,并且随后确定这种能力是否影响组装的aav颗粒的感染性。另外,确定乙酰化对泛素化/降解和靶向溶酶体、er、高尔基体或内核膜的影响。例如,为了测定运输或靶向,对具有突变的第2位(例如,imetx)的衣壳蛋白的aav颗粒进行荧光标记并用于感染细胞(例如,hela细胞)。测定这些aav颗粒的以下中的一种或多种:病毒颗粒摄取的时间,aav颗粒与特定区室标记(例如,高尔基体、er或溶酶体蛋白或其他标记)的共定位,核积累(例如,如通过与核标记或染色剂共定位测定的),和/或运输对早期内体逃逸的特定抑制剂(如巴伐洛霉素a(bafilomycina)或氯化铵)的敏感性,与用于感染相同细胞系的荧光标记的野生型aav颗粒相比(关于此类测定的描述,参见例如bartlett,j.s.等人(2000)j.virol.74:2777-2785)。为了测定感染性,使用具有突变的第2位(例如,imetx)的衣壳蛋白的aav颗粒感染细胞(例如,hela细胞),并且将它们的转导效率与野生型aav颗粒(例如,具有相同的aav血清型并感染相同类型的细胞)进行比较。为了测定糖基化,使用具有突变的第2位(例如,imetx)的衣壳蛋白的aav颗粒感染细胞(例如,hela细胞)。使来自感染细胞的aav颗粒经历一种或多种测定,包括但不限于糖基化的化学检测(例如,在变性和电泳分离的衣壳蛋白上应用市售的地高辛(dig)聚糖检测和/或荧光糖蛋白检测试剂盒)和质谱(例如,ft-icrms),与用于感染相同细胞系的野生型aav颗粒相比(关于此类测定的描述,参见例如murray,s.等人(2006)j.virol.80:6171-6176)。为了测定泛素化,使用具有突变的第2位的衣壳蛋白的aav颗粒感染细胞(例如,hela细胞)。用抗衣壳抗体从感染的细胞中免疫沉淀aav颗粒,然后使其经历用抗泛素抗体进行的western印迹,并且与用于以相同方式感染细胞的野生型aav颗粒进行比较。与野生型aav颗粒相比,突变型aav颗粒也可以用于体外泛素化测定(参见例如,yan,z.等人(2002)j.virol.76:2043-2053)。实施例3:aav2衣壳蛋白的脱酰胺化的作用aav2衣壳蛋白的序列分析揭示了潜在的脱酰胺化位点,如下面的氨基酸序列中加下划线的:maadgylpdwledtlsegirqwwklkpgppppkpaerhkddsrglvlpgykefqerlkedtsfggnlgravfqakkrvleplglveepvktapgkkrpvehspvepdsssgtgkagqqparkrlnfgqtgdadsvpdpqplgqppaapsglgtntmatgsgapmadnnegadgvgnssgnwhcdstwmgdrvittstrtwalptynnhlykqissqsgasndnhyfgystpwgyfdfnrfhchfsprdwqrlinnnwgfrpkrlnfklfniqvkevtqndgtttiannltstvqvftdseyqlpyvlgsahqgclppfpadvfmvpqygyltlnngsqavgrssfycleyfpsqmlrtgnnftfsytfedvpfhssyahsqsldrlmnplidqylyylsrtntpsgtttqsrlqfsqagasdirdqsrnwlpgpcyrqqrvsktsadnnnseyswtgatkyhlngrdslvnpgpamashkddeekffpqsgvlifgkqgsektnvdiekvmitdeeeirttnpvateqygsvstnlqrgnrqaatadvntqgvlpgmvwqdrdvylqgpiwakiphtdghfhpsplmggfglkhpppqilikntpvpanpsttfsaakfasfitqystgqvsveiewelqkenskrwnpeiqytsnynksvnvdftvdtngvyseprpigtryltrnl(seqidno:3)。特别地,在磷脂酶a2结构域(ca++结合位点)的n57/g58处发现了潜在的脱酰胺化位点,在上述序列中以粗体和斜体显示。以下实验旨在探索n57处的脱酰胺化是否会导致aav2的效力降低和/或截短,以及不同的aav生产方法是否可能对脱酰胺化具有不同的影响。例如,与三重转染方法相比,生产细胞系方法(参见martin等人,(2013)humangenetherapymethods24:253-269;美国专利授予前公开号us2004/0224411;和liu,x.l.等人(1999)genether.6:293-299)可以在n57处诱导更高水平的脱酰胺化。根据aav2的晶体结构,未显示n57;然而,n382和n511部分暴露,并且n715完全暴露。方法aav1和aav2vp的酶消化使用amicon过滤器(10kdamwco)浓缩10μg每种aav1-egfp或aav2-egfp材料(由三重转染以及生产细胞系过程产生),用6m盐酸胍、50mmtris(ph8.5)变性。将蛋白质用5mmdtt在60℃下在黑暗中还原30分钟,用15mm碘乙酰胺在室温下烷基化30分钟,并且然后使用bio-6tris微柱缓冲液交换到25mmtrisph7.1中进行消化。缓冲液交换后,将样品分成两个等分试样。将每个等分试样分别用1:25酶:蛋白质比率(wt/wt)的胰蛋白酶或1:50酶:蛋白质比率(wt/wt)的asp-n在37℃下消化2小时。uplc/ms/ms肽谱分析还通过uplc/ms/ms在acquityuplc-xevoqtofms中分析了蛋白质消化物。使用beh300c18柱(2.1×150mm)在流动相中以0.1%甲酸水溶液/乙腈溶液的梯度以0.25ml/min的流速进行分离。质谱以正mse分辨率模式获得,质量范围为50-2000。确定aavvp中的脱酰胺化水平将含有ng位点(aa1和aav2vp中的t9、t49和t67)的肽及其对应的脱酰胺化物质的提取离子色谱图(xic)用于计算脱酰胺化水平。为了比较通过三重转染(ttx)和生产细胞系(pcl)方法产生的aav载体,使用ttx或pcl方法产生用egfp标记的aav1或aav2。发现截短的vp1(tvp1)存在于通过pcl产生的aav2-egfp中,但不存在于通过ttx产生的aav2-egfp中。无论生产方法如何,均未发现aav1-egfp具有tvp1。与通过ttx产生的aav2相比,还发现通过pcl方法产生的aav2的体外效力降低。还发现突变型n57k和n57qaav2颗粒具有降低的效力并破坏ca++结合。下表提供了分析以检查每个潜在的脱酰胺化位点的胰蛋白酶肽以及对应的残基。表7:含有ng位点的胰蛋白酶肽如表7所示,将t9肽ylgpfldk(seqidno:9)用于监测n57,将t38肽evtqndgtttiannltstvqvftdseyqlpyvlgsahqgclppfpadvfmvpqygyltlnsqavgrssfycleyfpsqmlr(seqidno:10)用于监测n382,将t49肽ynlr(seqidno:11)和yhlr(seqidno:12)用于(分别)监测aav1或aav2中的n511,并且将t67肽sanvdftvdnlytepr(seqidno:13)和svnvdftvdtvysepr(seqidno:14)用于(分别)监测aav1或aav2中的n715。使用lc/ms/ms分析来比较通过ttx和pcl方法产生的aav1和aav2颗粒中脱酰胺化的百分比。来自t9肽的结果示于图6a和6b中。来自t49肽的结果示于图7a和7b中。来自t67肽的结果示于图8a和8b中。将这些结果汇总于表8中。由于其大小,未检测到t38肽。表8:lc/ms/ms结果的汇总具体而言,与通过ttx产生的aav2相比,通过pcl产生的aav2显示出近3倍的脱酰胺化增加。这些结果表明,脱酰胺化降低了aav效力,因为通过pcl产生的aav2的体外效力是降低的。结论总之,实施例1-3说明了用于使用lc/ms分析病毒颗粒(例如,aav衣壳蛋白)的完整蛋白质的方法。精确地测量了分子量,并且这些技术也可以用于评估病毒衣壳蛋白的n末端和/或修饰。此外,这些方法适合作为可用于基因治疗的衣壳血清型身份测定,例如作为分析平台。这些结果进一步确定了衣壳蛋白结构(例如,截短、脱酰胺化等)与效力之间的相关性,表明关键位点的点突变可以用于工程化更有效的载体。实施例4:阐明aav衣壳蛋白的n末端乙酰化的作用如上文所讨论的,aav衣壳蛋白的n末端在血清型中是高度保守的(图5)。实施例1中描述的技术允许探询vp表达和翻译后修饰。接下来检查aav衣壳蛋白的n末端乙酰化的作用和生物学意义。结果为了阐明aav衣壳蛋白脱乙酰化的潜在作用,测试了aav5脱乙酰化变体。将在cba启动子下表达egfp的aav5颗粒(aav5-cba-egfp)与具有对vp1和vp3突变的起始甲硫氨酸(imet)相邻的氨基酸的aav5变体(deac-aav5-cba-egfp)进行比较。选择预测被nata、natc或natd乙酰化可能性低的三个氨基酸用于产生变体:gly、leu和pro,如下表9中所示。表9:n末端乙酰化频率产生了以下aav5脱乙酰化(deac)突变体:s2gvp1–aav5vp1中的2位由ser变为glys2lvp1–aav5vp1中的2位由ser变为leus2pvp1–aav5vp1中的2位由ser变为pros2gvp3–aav5vp3中的2位由ser变为glys2lvp3–aav5vp3中的2位由ser变为leus2pvp3–aav5vp3中的2位由ser变为pros2pvp1/vp3–aav5vp1和vp3两者中的2位都由ser变为pros2gvp1/vp3–aav5vp1和vp3两者中的2位都由ser变为glys2lvp1/vp3–aav5vp1和vp3两者中的2位都由ser变为leu使用如上所述的ttx方法产生这些变体。所有aav5变体都显示出良好的生产力,产率大于1013总vg。通过sypro蛋白质凝胶分析,所有aav5变体还都显示出预期的vp1:vp2:vp3蛋白质比率(图9)。接下来,使用lc/ms确认所有aav5变体具有降低的乙酰化,如表10所示。表10:aav5变体乙酰化的lc/ms分析nd=未确定这些lc/ms分析确认aav5变体被脱乙酰化。变体s2lvp1、s2lvp3和s2lvp1/vp3均显示出vp1和vp3蛋白的质量增加(从173增加到182),表明将vp1或vp3中的第二个n末端氨基酸变为亮氨酸会改变蛋白质,导致质量增加。接下来,使用egfp作为报告基因在体外转导测定中测定aav5变体(图10)。设计该测定以评估在106感染复数(moi)下aav5脱乙酰化突变型变体的转导,将每种变体与亲本未修饰的aav5颗粒进行比较。使用了三种细胞系:293、huh7和hela细胞。感染后,测定细胞以确定载体基因组拷贝数(vg/μg细胞蛋白)和egfp表达(通过elisa)。载体基因组拷贝数(vg/μg蛋白质)代表aav5变体进入细胞的效率,并且egfp代表衣壳细胞内运输的效率,因为转基因表达需要衣壳/载体dna有效地进入细胞核(图10)。通过taqman分析定量载体基因组。图11显示,基于载体基因组分析,与亲本未修饰的aav5颗粒相比,aav5脱乙酰化的突变型载体以相似但降低的水平感染所有三种测试细胞系。图12显示与用亲本未修饰的aav5转导相比,aav5脱乙酰化突变型体载体均导致所有三种细胞系中egfp表达降低。结论如所预测的,当通过lc/ms检查时,在n末端ser至pro/leu/gly突变型变体中未观察到乙酰化。aav5deac变体显示稳健的载体产生,并且aav5deac变体以与亲本aav5相当的水平感染细胞。然而,与亲本aav5相比,用deac变体感染的细胞中的功能性蛋白质水平大大降低。这些数据表明,在vp1/vp3中缺乏n末端脱乙酰化对嗜性的影响最小,但下游加工(例如,运输和/或降解)受到显著影响。由于测试的变体展示出降低的体外活性,本领域技术人员可以理解,当需要降低转导水平时,可以尤其使用由降低或消除的乙酰化表征的变体。实施例5:aav衣壳蛋白脱酰胺化的评估实施例1和3展示了允许探询aav衣壳蛋白的翻译后修饰并探索aav2衣壳脱酰胺化的作用的技术。以下实施例测试了脱酰胺化是否降低衣壳蛋白的效力和/或诱导其截短,以及不同的制造过程是否可以诱导不同水平的脱酰胺化。方法如实施例3中所述产生aav颗粒并测定其脱酰胺化状态。结果如实施例3中所述,在aav2衣壳的vp1中的磷脂酶a2结构域(ca++结合位点)的n57/g58处发现了潜在的脱酰胺化位点。n57/g58基序在aav血清型中是保守的(图13)。实施例3显示,与通过ttx产生的aav2相比,通过pcl产生的aav2表现出近3倍的脱酰胺化增加(参见图6a和6b和表8)。在通过蛋白质凝胶检测vp1、vp2和vp3产生时,仅在通过pcl方法产生的aav2衣壳蛋白中检测到截短的vp1蛋白(tvp1)(图14)。接下来产生一系列aav2脱酰胺化突变体。这些突变体靶向经典ng序列中的gly残基。还产生了靶向a35残基(tvp1的n末端氨基酸)的突变(参见图13),如表11所示。携带多个突变的paf277和paf279突变体未包装。表11:脱酰胺化突变体名称突变平均drp/细胞paf274g58k4.54e+03paf275g58d5.00e+03paf276g58q5.41e+03paf277g58,383,512,716k1.2paf278a35n6.89e+03paf279a35n,g58,383,512,765k2.2293-0.9pim45对照6.28e+03k=正电荷(碱性)d=负电荷(酸性)q=极性接下来如上文实施例3中所述的通过lc/ms分析变体的脱酰胺化。与亲本aav2相比,aav2a35n和aav2g58d变体具有改变的脱酰胺化(图15)。具体而言,与亲本aav2(5.7%)相比,aav2a35n突变体具有增加的脱酰胺化(17.8%)。与亲本aav2相比,aav2g58d变体具有降低的脱酰胺化(1.1%)。sypro蛋白质凝胶分析证明,aav2脱酰胺化突变体表现出正确的vp1:vp2:vp2比率(图16)。接下来,使用egfp作为报告基因在体外转导测定中测定aav2脱酰胺化变体(图17)。设计该测定以评估在106感染复数(moi)下aav2脱酰胺化突变型变体的转导,将每种变体与亲本未修饰的aav2颗粒进行比较。使用了三种细胞系:293、huh7和hela细胞。感染后,测定细胞以确定载体基因组拷贝数(vg/μg细胞蛋白)和egfp表达(通过elisa)。载体基因组拷贝数(vg/μg蛋白质)代表aav2变体进入细胞的效率,并且egfp代表衣壳细胞内运输的效率,因为转基因表达需要衣壳/载体dna有效地进入细胞核(图17)。通过taqman分析定量载体基因组。载体基因组分析指示,aav2脱酰胺化突变型变体以与亲本aav2载体相当的水平感染所有测试的细胞系(图18)。重要的是,发现aav2a35n变体比亲本aav2载体在所有三种细胞系中更有效地进行转导(图19)。发现aav2g58d变体比亲本aav2载体在huh7细胞中更有效(图19)。结论总之,aav2脱酰胺化突变型载体以与亲本aav2颗粒相当的水平(例如,相当的vg/μg细胞蛋白)感染细胞。然而,基于转导细胞中egfp水平的分析,aav2a35n变体比亲本aav2在测试的所有细胞系中具有更高的效力,并且aav2g58d变体比亲本aav2在huh7细胞(肝源细胞系)中具有更高的效力。这些结果表明,a35n突变可以有效地提高转导许多细胞类型的载体效力,并且g58d突变也可以有效地提高某些细胞类型(例如,肝细胞)的效力。参考文献1.girod,a.,etal.,thevp1capsidproteinofadeno-associatedvirustype2iscarryingaphospholipasea2domainrequiredforvirusinfectivity.jgenvirol,2002.83(pt5):p.973-8.2.stahnke,s.,etal.,intrinsicphospholipasea2activityofadeno-associatedvirusisinvolvedinendosomalescapeofincomingparticles.virology,2011.409(1):p.77-83.3.bleker,s.,f.sonntag,andj.a.kleinschmidt,mutationalanalysisofnarrowporesatthefivefoldsymmetryaxesofadeno-associatedvirustype2capsidsrevealsadualroleingenomepackagingandactivationofphospholipasea2activity.jvirol,2005.79(4):p.2528-40.4.popa-wagner,r.,etal.,impactofvp1-specificproteinsequencemotifsonadeno-associatedvirustype2intracellulartraffickingandnuclearentry.jvirol,2012.86(17):p.9163-74.5.kashiwakura,y.,etal.,hepatocytegrowthfactorreceptorisacoreceptorforadeno-associatedvirustype2infection.jvirol,2005.79(1):p.609-14.6.qing,k.,etal.,humanfibroblastgrowthfactorreceptor1isaco-receptorforinfectionbyadeno-associatedvirus2.natmed,1999.5(1):p.71-7.7.sanlioglu,s.,etal.,endocytosisandnucleartraffickingofadeno-associatedvirustype2arecontrolledbyrac1andphosphatidylinositol-3kinaseactivation.jvirol,2000.74(19):p.9184-96.8.summerford,c.,j.s.bartlett,andr.j.samulski,alphavbeta5integrin:aco-receptorforadeno-associatedvirustype2infection.natmed,1999.5(1):p.78-82.9.kern,a.,etal.,identificationofaheparin-bindingmotifonadeno-associatedvirustype2capsids.jvirol,2003.77(20):p.11072-11081.10.asokan,a.,etal.,adeno-associatedvirustype2containsanintegrinalpha5beta1bindingdomainessentialforviralcellentry.jvirol,2006.80(18):p.8961-9.11.xie,q.,etal.,theatomicstructureofadeno-associatedvirus(aav-2),avectorforhumangenetherapy.procnatlacadsciusa,2002.99(16):p.10405-10.12.dimattia,m.a.,etal.,structuralinsightintotheuniquepropertiesofadeno-associatedvirusserotype9.jvirol,2012.86(12):p.6947-58.13.nam,h.j.,etal.,structureofadeno-associatedvirusserotype8,agenetherapyvector.jvirol,2007.81(22):p.12260-71.14.kronenberg,s.,etal.,aconformationalchangeintheadeno-associatedvirustype2capsidleadstotheexposureofhiddenvp1ntermini.jvirol,2005.79(9):p.5296-303.15.sonntag,f.,etal.,adeno-associatedvirustype2capsidswithexternalizedvp1/vp2traffickingdomainsaregeneratedpriortopassagethroughthecytoplasmandaremaintaineduntiluncoatingoccursinthenucleus.jvirol,2006.80(22):p.11040-54.16.murray,s.,etal.,characterizationofthecapsidproteinglycosylationofadeno-associatedvirustype2byhigh-resolutionmassspectrometry.jvirol,2006.80(12):p.6171-6.17.salganik,m.,etal.,evidenceforph-dependentproteaseactivityintheadeno-associatedviruscapsid.jvirol,2012.86(21):p.11877-85.18.vanvliet,k.,etal.,adeno-associatedviruscapsidserotypeidentification:analyticalmethodsdevelopmentandapplication.jvirolmethods,2009.159(2):p.167-77.19.thomas,j.j.,etal.,viralcharacterizationbydirectanalysisofcapsidproteins.analchem,1998.70(18):p.3863-7.20.wang,l.,l.c.lane,andd.l.smith,detectingstructuralchangesinviralcapsidsbyhydrogenexchangeandmassspectrometry.proteinsci,2001.10(6):p.1234-43.21.hwang,c.s.,a.shemorry,anda.varshavsky,n-terminalacetylationofcellularproteinscreatesspecificdegradationsignals.science,2010.327(5968):p.973-7.22.yan,z.,etal.,ubiquitinationofbothadeno-associatedvirustype2and5capsidproteinsaffectsthetransductionefficiencyofrecombinantvectors.jvirol,2002.76(5):p.2043-2053.序列除非另有说明,否则所有多肽序列均以n末端到c末端呈现。除非另有说明,否则所有核酸序列均以5'到3'呈现。潜在aav2vp3起始密码子的核苷酸序列(atg密码子加下划线)atggctacaggcagtggcgcaccaatggcagac(seqidno:1)对应于潜在aav2vp3起始密码子的多肽序列(甲硫氨酸加下划线)matgsgapmad(seqidno:2)aav2vp1多肽序列maadgylpdwledtlsegirqwwklkpgppppkpaerhkddsrglvlpgykylgpfngldkgepvneadaaalehdkaydrqldsgdnpylkynhadaefqerlkedtsfggnlgravfqakkrvleplglveepvktapgkkrpvehspvepdsssgtgkagqqparkrlnfgqtgdadsvpdpqplgqppaapsglgtntmatgsgapmadnnegadgvgnssgnwhcdstwmgdrvittstrtwalptynnhlykqissqsgasndnhyfgystpwgyfdfnrfhchfsprdwqrlinnnwgfrpkrlnfklfniqvkevtqndgtttiannltstvqvftdseyqlpyvlgsahqgclppfpadvfmvpqygyltlnngsqavgrssfycleyfpsqmlrtgnnftfsytfedvpfhssyahsqsldrlmnplidqylyylsrtntpsgtttqsrlqfsqagasdirdqsrnwlpgpcyrqqrvsktsadnnnseyswtgatkyhlngrdslvnpgpamashkddeekffpqsgvlifgkqgsektnvdiekvmitdeeeirttnpvateqygsvstnlqrgnrqaatadvntqgvlpgmvwqdrdvylqgpiwakiphtdghfhpsplmggfglkhpppqilikntpvpanpsttfsaakfasfitqystgqvsveiewelqkenskrwnpeiqytsnynksvnvdftvdtngvyseprpigtryltrnl(seqidno:3)vp1n末端胰蛋白酶肽(n末端丙氨酸被乙酰化)aadgylpdwledtlsegir(seqidno:4)vp3n末端asp-n肽(n末端丙氨酸被乙酰化)atgsgapm(seqidno:5)常见的vp1n末端序列maadgylpdwled(seqidno:6)潜在aav7vp3起始密码子的核苷酸序列(起始密码子加下划线)gtggctgcaggcggtggcgcaccaatggcagacaataac(seqidno:7)突变的itr的核苷酸序列cactccctctctgcgcgctcgctcgctcactgaggccgggcgaccaaaggtcgcccacgcccgggctttgcccgggcg(seqidno:8)序列表<110>jin,xiaoyingo'riordan,catherineliu,lingzhang,kate<120>检测aav的方法<130>159792014140<140>尚未分配<141>与此同时<150>62/375,314<151>2016-08-15<160>40<170>用于windows版本4.0的fastseq<210>1<211>33<212>prt<213>人工序列<220><223>合成构建体<400>1alathrglyglycysthralacysalaglyglycysalaglythrgly151015glycysglycysalacyscysalaalathrglyglycysalaglyala202530cys<210>2<211>11<212>prt<213>人工序列<220><223>合成构建体<400>2metalathrglyserglyalaprometalaasp1510<210>3<211>735<212>prt<213>腺相关病毒2<400>3metalaalaaspglytyrleuproasptrpleugluaspthrleuser151015gluglyileargglntrptrplysleulysproglypropropropro202530lysproalagluarghislysaspaspserargglyleuvalleupro354045glytyrlystyrleuglypropheasnglyleuasplysglyglupro505560valasnglualaaspalaalaalaleugluhisasplysalatyrasp65707580argglnleuaspserglyaspasnprotyrleulystyrasnhisala859095aspalaglupheglngluargleulysgluaspthrserpheglygly100105110asnleuglyargalavalpheglnalalyslysargvalleuglupro115120125leuglyleuvalglugluprovallysthralaproglylyslysarg130135140provalgluhisserprovalgluproaspserserserglythrgly145150155160lysalaglyglnglnproalaarglysargleuasnpheglyglnthr165170175glyaspalaaspservalproaspproglnproleuglyglnpropro180185190alaalaproserglyleuglythrasnthrmetalathrglysergly195200205alaprometalaaspasnasngluglyalaaspglyvalglyasnser210215220serglyasntrphiscysaspserthrtrpmetglyaspargvalile225230235240thrthrserthrargthrtrpalaleuprothrtyrasnasnhisleu245250255tyrlysglnileserserglnserglyalaserasnaspasnhistyr260265270pheglytyrserthrprotrpglytyrpheasppheasnargphehis275280285cyshispheserproargasptrpglnargleuileasnasnasntrp290295300glypheargprolysargleuasnphelysleupheasnileglnval305310315320lysgluvalthrglnasnaspglythrthrthrilealaasnasnleu325330335thrserthrvalglnvalphethraspserglutyrglnleuprotyr340345350valleuglyseralahisglnglycysleupropropheproalaasp355360365valphemetvalproglntyrglytyrleuthrleuasnasnglyser370375380glnalavalglyargserserphetyrcysleuglutyrpheproser385390395400glnmetleuargthrglyasnasnphethrphesertyrthrpheglu405410415aspvalprophehissersertyralahisserglnserleuasparg420425430leumetasnproleuileaspglntyrleutyrtyrleuserargthr435440445asnthrproserglythrthrthrglnserargleuglnphesergln450455460alaglyalaseraspileargaspglnserargasntrpleuprogly465470475480procystyrargglnglnargvalserlysthrseralaaspasnasn485490495asnserglutyrsertrpthrglyalathrlystyrhisleuasngly500505510argaspserleuvalasnproglyproalametalaserhislysasp515520525aspgluglulysphepheproglnserglyvalleuilepheglylys530535540glnglyserglulysthrasnvalaspileglulysvalmetilethr545550555560aspgluglugluileargthrthrasnprovalalathrgluglntyr565570575glyservalserthrasnleuglnargglyasnargglnalaalathr580585590alaaspvalasnthrglnglyvalleuproglymetvaltrpglnasp595600605argaspvaltyrleuglnglyproiletrpalalysileprohisthr610615620aspglyhisphehisproserproleumetglyglypheglyleulys625630635640hisproproproglnileleuilelysasnthrprovalproalaasn645650655proserthrthrpheseralaalalysphealaserpheilethrgln660665670tyrserthrglyglnvalservalgluileglutrpgluleuglnlys675680685gluasnserlysargtrpasnprogluileglntyrthrserasntyr690695700asnlysservalasnvalaspphethrvalaspthrasnglyvaltyr705710715720sergluproargproileglythrargtyrleuthrargasnleu725730735<210>4<211>19<212>prt<213>人工序列<220><223>合成构建体<220><221>乙酰化<222>1<400>4alaalaaspglytyrleuproasptrpleugluaspthrleuserglu151015glyilearg<210>5<211>8<212>prt<213>人工序列<220><223>合成构建体<220><221>乙酰化<222>1<400>5alathrglyserglyalapromet15<210>6<211>13<212>prt<213>人工序列<220><223>合成构建体<400>6metalaalaaspglytyrleuproasptrpleugluasp1510<210>7<211>39<212>prt<213>人工序列<220><223>合成构建体<400>7glythrglyglycysthrglycysalaglyglycysglyglythrgly151015glycysglycysalacyscysalaalathrglyglycysalaglyala202530cysalaalathralaalacys35<210>8<211>78<212>prt<213>人工序列<220><223>合成构建体<400>8cysalacysthrcyscyscysthrcysthrcysthrglycysglycys151015glycysthrcysglycysthrcysglycysthrcysalacysthrgly202530alaglyglycyscysglyglyglycysglyalacyscysalaalaala354045glyglythrcysglycyscyscysalacysglycyscyscysglygly505560glycysthrthrthrglycyscyscysglyglyglycysgly657075<210>9<211>10<212>prt<213>人工序列<220><223>合成构建体<400>9tyrleuglypropheasnglyleuasplys1510<210>10<211>83<212>prt<213>人工序列<220><223>合成构建体<400>10gluvalthrglnasnaspglythrthrthrilealaasnasnleuthr151015serthrvalglnvalphethraspserglutyrglnleuprotyrval202530leuglyseralahisglnglycysleupropropheproalaaspval354045phemetvalproglntyrglytyrleuthrleuasnasnglysergln505560alavalglyargserserphetyrcysleuglutyrpheprosergln65707580metleuarg<210>11<211>6<212>prt<213>人工序列<220><223>合成构建体<400>11tyrasnleuasnglyarg15<210>12<211>6<212>prt<213>人工序列<220><223>合成构建体<400>12tyrhisleuasnglyarg15<210>13<211>18<212>prt<213>人工序列<220><223>合成构建体<400>13seralaasnvalaspphethrvalaspasnasnglyleutyrthrglu151015proarg<210>14<211>18<212>prt<213>人工序列<220><223>合成构建体<400>14servalasnvalaspphethrvalaspthrasnglyvaltyrserglu151015proarg<210>15<211>15<212>prt<213>人工序列<220><223>合成构建体<400>15alaproglylyslysargprovalgluhisserprovalglupro151015<210>16<211>735<212>prt<213>腺相关病毒2<400>16metalaalaaspglytyrleuproasptrpleugluaspthrleuser151015gluglyileargglntrptrplysleulysproglypropropropro202530lysproalagluarghislysaspaspserargglyleuvalleupro354045glytyrlystyrleuglypropheasnglyleuasplysglyglupro505560valasnglualaaspalaalaalaleugluhisasplysalatyrasp65707580argglnleuaspserglyaspasnprotyrleulystyrasnhisala859095aspalaglupheglngluargleulysgluaspthrserpheglygly100105110asnleuglyargalavalpheglnalalyslysargvalleuglupro115120125leuglyleuvalglugluprovallysthralaproglylyslysarg130135140provalgluhisserprovalgluproaspserserserglythrgly145150155160lysalaglyglnglnproalaarglysargleuasnpheglyglnthr165170175glyaspalaaspservalproaspproglnproleuglyglnpropro180185190alaalaproserglyleuglythrasnthrmetalathrglysergly195200205alaprometalaaspasnasngluglyalaaspglyvalglyasnser210215220serglyasntrphiscysaspserthrtrpmetglyaspargvalile225230235240thrthrserthrargthrtrpalaleuprothrtyrasnasnhisleu245250255tyrlysglnileserserglnserglyalaserasnaspasnhistyr260265270pheglytyrserthrprotrpglytyrpheasppheasnargphehis275280285cyshispheserproargasptrpglnargleuileasnasnasntrp290295300glypheargprolysargleuasnphelysleupheasnileglnval305310315320lysgluvalthrglnasnaspglythrthrthrilealaasnasnleu325330335thrserthrvalglnvalphethraspserglutyrglnleuprotyr340345350valleuglyseralahisglnglycysleupropropheproalaasp355360365valphemetvalproglntyrglytyrleuthrleuasnasnglyser370375380glnalavalglyargserserphetyrcysleuglutyrpheproser385390395400glnmetleuargthrglyasnasnphethrphesertyrthrpheglu405410415aspvalprophehissersertyralahisserglnserleuasparg420425430leumetasnproleuileaspglntyrleutyrtyrleuserargthr435440445asnthrproserglythrthrthrglnserargleuglnphesergln450455460alaglyalaseraspileargaspglnserargasntrpleuprogly465470475480procystyrargglnglnargvalserlysthrseralaaspasnasn485490495asnserglutyrsertrpthrglyalathrlystyrhisleuasngly500505510argaspserleuvalasnproglyproalametalaserhislysasp515520525aspgluglulysphepheproglnserglyvalleuilepheglylys530535540glnglyserglulysthrasnvalaspileglulysvalmetilethr545550555560aspgluglugluileargthrthrasnprovalalathrgluglntyr565570575glyservalserthrasnleuglnargglyasnargglnalaalathr580585590alaaspvalasnthrglnglyvalleuproglymetvaltrpglnasp595600605argaspvaltyrleuglnglyproiletrpalalysileprohisthr610615620aspglyhisphehisproserproleumetglyglypheglyleulys625630635640hisproproproglnileleuilelysasnthrprovalproalaasn645650655proserthrthrpheseralaalalysphealaserpheilethrgln660665670tyrserthrglyglnvalservalgluileglutrpgluleuglnlys675680685gluasnserlysargtrpasnprogluileglntyrthrserasntyr690695700asnlysservalasnvalaspphethrvalaspthrasnglyvaltyr705710715720sergluproargproileglythrargtyrleuthrargasnleu725730735<210>17<211>239<212>prt<213>人工序列<220><223>合成构建体<400>17metalaalaaspglytyrleuproasptrpleugluaspasnleuser151015gluglyileargglutrptrpaspleulysproglyalaprolyspro202530lysalaasnglnglnlysglnaspaspglyargglyleuvalleupro354045glytyrlystyrleuglypropheasnglyleuasplysglyglupro505560valasnalaalaaspalaalaalaleugluhisasplysalatyrasp65707580glnglnleulysalaglyaspasnprotyrleuargtyrasnhisala859095aspalaglupheglngluargleuglngluaspthrserpheglygly100105110asnleuglyargalavalpheglnalalyslysargvalleuglupro115120125leuglyleuvalglugluglyalalysthralaproglylyslysarg130135140provalgluproserproglnargserproaspserserthrglyile145150155160glylyslysglyglnglnproalalyslysargleuasnpheglygln165170175thrglyaspsergluservalproaspproglnproileglyglupro180185190proalaglyproserglyleuglyserglythrmetalaalaglygly195200205glyalaprometalaaspasnasngluglyalaaspglyvalglyser210215220serserglyasntrphiscysaspserthrtrpleuglyasparg225230235<210>18<211>239<212>prt<213>人工序列<220><223>合成构建体<400>18metalaalaaspglytyrleuproasptrpleugluaspasnleuser151015gluglyileargglutrptrpaspleulysproglyalaprolyspro202530lysalaasnglnglnlysglnaspaspglyargglyleuvalleupro354045glytyrlystyrleuglypropheasnglyleuasplysglyglupro505560valasnalaalaaspalaalaalaleugluhisasplysalatyrasp65707580glnglnleulysalaglyaspasnprotyrleuargtyrasnhisala859095aspalaglupheglngluargleuglngluaspthrserpheglygly100105110asnleuglyargalavalpheglnalalyslysargvalleuglupro115120125leuglyleuvalgluglualaalalysthralaproglylyslysarg130135140provalgluproserproglnargserproaspserserthrglyile145150155160glylyslysglyglnglnproalalyslysargleuasnpheglygln165170175thrglyglusergluservalproaspproglnproileglyglupro180185190proalaglyproserglyleuglyserglythrmetalaalaglygly195200205glyalaprometalaaspasnasngluglyalaaspglyvalglyser210215220serserglyasntrphiscysaspserthrtrpleuglyasparg225230235<210>19<211>239<212>prt<213>人工序列<220><223>合成构建体<400>19metalaalaaspglytyrleuproasptrpleugluaspasnleuser151015gluglyileargglutrptrpalaleulysproglyalaprolyspro202530lysalaasnglnglnlysglnaspaspglyargglyleuvalleupro354045glytyrlystyrleuglypropheasnglyleuasplysglyglupro505560valasnalaalaaspalaalaalaleugluhisasplysalatyrasp65707580glnglnleuglnalaglyaspasnprotyrleuargtyrasnhisala859095aspalaglupheglngluargleuglngluaspthrserpheglygly100105110asnleuglyargalavalpheglnalalyslysargvalleuglupro115120125leuglyleuvalglugluglyalalysthralaproglylyslysarg130135140provalgluproserproglnargserproaspserserthrglyile145150155160glylyslysglyglnglnproalaarglysargleuasnpheglygln165170175thrglyaspsergluservalproaspproglnproleuglyglupro180185190proalaalaproserglyvalglyproasnthrmetalaalaglygly195200205glyalaprometalaaspasnasngluglyalaaspglyvalglyser210215220serserglyasntrphiscysaspserthrtrpleuglyasparg225230235<210>20<211>239<212>prt<213>人工序列<220><223>合成构建体<400>20metalaalaaspglytyrleuproasptrpleugluaspasnleuser151015gluglyileargglutrptrpaspleulysproglyalaprolyspro202530lysalaasnglnglnlysglnaspasnglyargglyleuvalleupro354045glytyrlystyrleuglypropheasnglyleuasplysglyglupro505560valasnalaalaaspalaalaalaleugluhisasplysalatyrasp65707580glnglnleulysalaglyaspasnprotyrleuargtyrasnhisala859095aspalaglupheglngluargleuglngluaspthrserpheglygly100105110asnleuglyargalavalpheglnalalyslysargvalleuglupro115120125leuglyleuvalglugluglyalalysthralaproalalyslysarg130135140provalgluproserproglnargserproaspserserthrglyile145150155160glylyslysglyglnglnproalaarglysargleuasnpheglygln165170175thrglyaspsergluservalproaspproglnproleuglyglupro180185190proalaalaproserservalglyserglythrvalalaalaglygly195200205glyalaprometalaaspasnasngluglyalaaspglyvalglyasn210215220alaserglyasntrphiscysaspserthrtrpleuglyasparg225230235<210>21<211>238<212>prt<213>人工序列<220><223>合成构建体<400>21metalaalaaspglytyrleuproasptrpleugluaspasnleuser151015gluglyileargglutrptrpaspleulysproglyalaprolyspro202530lysalaasnglnglnlysglnaspaspglyargglyleuvalleupro354045glytyrlystyrleuglypropheasnglyleuasplysglyglupro505560valasnalaalaaspalaalaalaleugluhisasplysalatyrasp65707580glnglnleulysalaglyaspasnprotyrleuargtyrasnhisala859095aspalaglupheglngluargleuglngluaspthrserpheglygly100105110asnleuglyargalavalpheglnalalyslysargvalleuglupro115120125leuglyleuvalglugluglyalalysthralaproglylyslysarg130135140provalgluglnserproglngluproaspserserserglyilegly145150155160lysthrglyglnglnproalalyslysargleuasnpheglyglnthr165170175glyaspsergluservalproaspproglnproleuglyglupropro180185190alathrproalaalavalglyprothrthrmetalaserglyglygly195200205alaprometalaaspasnasngluglyalaaspglyvalglyasnala210215220serglyasntrphiscysaspserthrtrpleuglyasparg225230235<210>22<211>238<212>prt<213>人工序列<220><223>合成构建体<400>22metalaalaaspglytyrleuproasptrpleugluaspasnleuser151015gluglyileargglutrptrpaspleulysproglyalaprolyspro202530lysalaasnglnglnlysglnaspaspglyargglyleuvalleupro354045glytyrlystyrleuglypropheasnglyleuasplysglyglupro505560valasnalaalaaspalaalaalaleugluhisasplysalatyrasp65707580glnglnleulysalaglyaspasnprotyrleuargtyrasnhisala859095aspalaglupheglngluargleuglngluaspthrserpheglygly100105110asnleuglyargalavalpheglnalalyslysargvalleuglupro115120125pheglyleuvalglugluglyalalysthralaproglylyslysarg130135140provalgluglnserproglngluproaspserserserglyilegly145150155160lysthrglyglnglnproalalyslysargleuasnpheglyglnthr165170175glyaspsergluservalproaspproglnproleuglyglupropro180185190alathrproalaalavalglyprothrthrmetalaserglyglygly195200205alaprometalaaspasnasngluglyalaaspglyvalglyasnala210215220serglyasntrphiscysaspserthrtrpleuglyasparg225230235<210>23<211>238<212>prt<213>人工序列<220><223>合成构建体<400>23metalaalaaspglytyrleuproasptrpleugluaspthrleuser151015gluglyileargglntrptrplysleulysproglypropropropro202530lysproalagluarghislysaspaspserargglyleuvalleupro354045glytyrlystyrleuglypropheasnglyleuasplysglyglupro505560valasnglualaaspalaalaalaleugluhisasplysalatyrasp65707580argglnleuaspserglyaspasnprotyrleulystyrasnhisala859095aspalaglupheglngluargleulysgluaspthrserpheglygly100105110asnleuglyargalavalpheglnalalyslysargvalleuglupro115120125leuglyleuvalglugluprovallysthralaproglylyslysarg130135140provalgluhisserprovalgluproaspserserserglythrgly145150155160lysalaglyglnglnproalaarglysargleuasnpheglyglnthr165170175glyaspalaaspservalproaspproglnproleuglyglnpropro180185190alaalaproserglyleuglythrasnthrmetalathrglysergly195200205alaprometalaaspasnasngluglyalaaspglyvalglyasnser210215220serglyasntrphiscysaspserthrtrpmetglyasparg225230235<210>24<211>238<212>prt<213>人工序列<220><223>合成构建体<400>24metalaalaaspglytyrleuproasptrpleugluaspasnleuser151015gluglyileargglutrptrpalaleulysproglyvalproglnpro202530lysalaasnglnglnhisglnaspasnargargglyleuvalleupro354045glytyrlystyrleuglyproglyasnglyleuasplysglyglupro505560valasnglualaaspalaalaalaleugluhisasplysalatyrasp65707580glnglnleulysalaglyaspasnprotyrleulystyrasnhisala859095aspalaglupheglngluargleuglngluaspthrserpheglygly100105110asnleuglyargalavalpheglnalalyslysargileleuglupro115120125leuglyleuvalgluglualaalalysthralaproglylyslysgly130135140alavalaspglnserproglngluproaspserserserglyvalgly145150155160lysserglylysglnproalaarglysargleuasnpheglyglnthr165170175glyaspsergluservalproaspproglnproleuglyglupropro180185190alaalaprothrserleuglyserasnthrmetalaserglyglygly195200205alaprometalaaspasnasngluglyalaaspglyvalglyasnser210215220serglyasntrphiscysaspserglntrpleuglyasparg225230235<210>25<211>232<212>prt<213>人工序列<220><223>合成构建体<400>25metalaalaaspglytyrleuproasptrpleugluaspasnleuser151015gluglyileargglutrptrpaspleulysproglyalaprolyspro202530lysalaasnglnglnlysglnaspaspglyargglyleuvalleupro354045glytyrlystyrleuglypropheasnglyleuasplysglyglupro505560valasnalaalaaspalaalaalaleugluhisasplysalatyrasp65707580glnglnleulysalaglyaspasnprotyrleuargtyrasnhisala859095aspalaglupheglngluargleuglngluaspthrserpheglygly100105110asnleuglyargalavalpheglnalalyslysargvalleuglupro115120125leuglyleuvalglugluglyalalysthralaproglylyslysarg130135140proleugluserproglngluproaspserserserglyileglylys145150155160lysglylysglnproalaarglysargleuasnpheglugluaspthr165170175glyalaglyaspglyproprogluglyseraspthrseralametser180185190seraspileglumetargalaalaproglyglyasnalavalaspala195200205glyglnglyseraspglyvalglyasnalaserglyasptrphiscys210215220aspserthrtrpsergluglylys225230<210>26<211>241<212>prt<213>人工序列<220><223>合成构建体<400>26metalaalaaspglytyrleuproasptrpleugluaspasnleuser151015gluglyileargglutrptrpalaleulysproglyalaproglnpro202530lysalaasnglnglnhisglnaspasnglyargglyleuvalleupro354045glytyrlystyrleuglypropheasnglyleuasplysglyglupro505560valasnglualaaspalaalaalaleugluhisasplysalatyrasp65707580lysglnleugluglnglyaspasnprotyrleulystyrasnhisala859095aspalaglupheglnglnargleualathraspthrserpheglygly100105110asnleuglyargalavalpheglnalalyslysargileleuglupro115120125leuglyleuvalglugluglyvallysthralaproglylyslysarg130135140proleuglulysthrproasnargprothrasnproaspserglylys145150155160alaproalalyslyslysglnlysaspglygluproalaaspserala165170175argargthrleuaspphegluaspserglyalaglyaspglypropro180185190gluglyserserserglyglumetserhisaspalaglumetargala195200205alaproglyglyasnalavalglualaglyglnglyalaaspglyval210215220glyasnalaserglyasptrphiscysaspserthrtrpserglugly225230235240arg<210>27<211>232<212>prt<213>人工序列<220><223>合成构建体<400>27metthraspglytyrleuproasptrpleugluaspasnleuserglu151015glyvalargglutrptrpalaleuglnproglyalaprolysprolys202530alaasnglnglnhisglnaspasnalaargglyleuvalleuprogly354045tyrlystyrleuglyproglyasnglyleuasplysglygluproval505560asnalaalaaspalaalaalaleugluhisasplysalatyraspgln65707580glnleulysalaglyaspasnprotyrleulystyrasnhisalaasp859095alaglupheglnglnargleuglnglyaspthrserpheglyglyasn100105110leuglyargalavalpheglnalalyslysargvalleugluproleu115120125glyleuvalgluglnalaglygluthralaproglylyslysargpro130135140leuilegluserproglnglnproaspserserthrglyileglylys145150155160lysglylysglnproalalyslyslysleuvalphegluaspgluthr165170175glyalaglyaspglyproprogluglyserthrserglyalametser180185190aspaspserglumetargalaalaalaglyglyalaalavalglugly195200205glyglnglyalaaspglyvalglyasnalaserglyasptrphiscys210215220aspserthrtrpsergluglyhis225230<210>28<211>228<212>prt<213>人工序列<220><223>合成构建体<400>28metserphevalasphisproproasptrpleuglugluvalglyglu151015glyleuargglupheleuglyleuglualaglyproprolysprolys202530proasnglnglnhisglnaspglnalaargglyleuvalleuprogly354045tyrasntyrleuglyproglyasnglyleuaspargglygluproval505560asnargalaaspgluvalalaarggluhisaspilesertyrasnglu65707580glnleuglualaglyaspasnprotyrleulystyrasnhisalaasp859095alaglupheglnglulysleualaaspaspthrserpheglyglyasn100105110leuglylysalavalpheglnalalyslysargvalleugluprophe115120125glyleuvalglugluglyalalysthralaprothrglylysargile130135140aspasphispheprolysarglyslysalaargthrglugluaspser145150155160lysproserthrserseraspalaglualaglyproserglysergln165170175glnleuglnileproalaglnproalaserserleuglyalaaspthr180185190metseralaglyglyglyglyproleuglyaspasnasnglnglyala195200205aspglyvalglyasnalaserglyasptrphiscysaspserthrtrp210215220metglyasparg225<210>29<211>238<212>prt<213>人工序列<220><223>合成构建体<400>29metalaalaaspglytyrleuproasptrpleugluaspasnleuser151015gluglyileargglutrptrpalaleulysproglyalaproglnpro202530lysalaasnglnglnhisglnaspasnalaargglyleuvalleupro354045glytyrlystyrleuglyproglyasnglyleuasplysglyglupro505560valasnalaalaaspalaalaalaleugluhisasplysalatyrasp65707580glnglnleulysalaglyaspasnprotyrleulystyrasnhisala859095aspalaglupheglngluargleulysgluaspthrserpheglygly100105110asnleuglyargalavalpheglnalalyslysargleuleuglupro115120125leuglyleuvalgluglualaalalysthralaproglylyslysarg130135140provalgluglnserproglngluproaspserseralaglyilegly145150155160lysserglyalaglnproalalyslysargleuasnpheglyglnthr165170175glyaspthrgluservalproaspproglnproileglyglupropro180185190alaalaproserglyvalglyserleuthrmetalaserglyglygly195200205alaprovalalaaspasnasngluglyalaaspglyvalglyserser210215220serglyasntrphiscysaspserglntrpleuglyasparg225230235<210>30<211>246<212>prt<213>人工序列<220><223>合成构建体<220><221>变体<222>24,148,151,152,154,155,168,169,170,171,172,208,209<223>xaa=任何氨基酸<400>30metalaalaaspglytyrleuproasptrpleugluaspasnleuser151015gluglyileargglutrptrpxaaleulysproglyalaprolyspro202530lysalaasnglnglnlysglnaspaspglyargglyleuvalleupro354045glytyrlystyrleuglypropheasnglyleuasplysglyglupro505560valasnalaalaaspalaalaalaleugluhisasplysalatyrasp65707580glnglnleulysalaglyaspasnprotyrleuargtyrasnhisala859095aspalaglupheglngluargleuglngluaspthrserpheglygly100105110asnleuglyargalavalpheglnalalyslysargvalleuglupro115120125leuglyleuvalglugluglyalalysthralaproglylyslysarg130135140provalgluxaaserproxaaxaaglnxaaxaaproaspserserser145150155160glyileglylyslysglyglnxaaxaaxaaxaaxaaglnproalalys165170175lysargleuasnpheglyglnthrglyaspsergluservalproasp180185190proglnproleuglygluproproalaalaproserglyleuglyxaa195200205xaathrmetalaalaglyglyglyalaprometalaaspasnasnglu210215220glyalaaspglyvalglyasnalaserglyasntrphiscysaspser225230235240thrtrpleuglyasparg245<210>31<211>61<212>prt<213>人工序列<220><223>合成构建体<400>31metalaalaaspglytyrleuproasptrpleugluaspasnleuser151015gluglyileargglutrptrpaspleulysproglyalaprolyspro202530lysalaasnglnglnlysglnaspaspglyargglyleuvalleupro354045glytyrlystyrleuglypropheasnglyleuasplys505560<210>32<211>61<212>prt<213>人工序列<220><223>合成构建体<400>32metalaalaaspglytyrleuproasptrpleugluaspasnleuser151015gluglyileargglutrptrpalaleulysproglyalaprolyspro202530lysalaasnglnglnlysglnaspaspglyargglyleuvalleupro354045glytyrlystyrleuglypropheasnglyleuasplys505560<210>33<211>61<212>prt<213>人工序列<220><223>合成构建体<400>33metalaalaaspglytyrleuproasptrpleugluaspasnleuser151015gluglyileargglutrptrpaspleulysproglyalaprolyspro202530lysalaasnglnglnlysglnaspasnglyargglyleuvalleupro354045glytyrlystyrleuglypropheasnglyleuasplys505560<210>34<211>61<212>prt<213>人工序列<220><223>合成构建体<400>34metalaalaaspglytyrleuproasptrpleugluaspthrleuser151015gluglyileargglntrptrplysleulysproglypropropropro202530lysproalagluarghislysaspaspserargglyleuvalleupro354045glytyrlystyrleuglypropheasnglyleuasplys505560<210>35<211>61<212>prt<213>人工序列<220><223>合成构建体<400>35metalaalaaspglytyrleuproasptrpleugluaspasnleuser151015gluglyileargglutrptrpalaleulysproglyvalproglnpro202530lysalaasnglnglnhisglnaspasnargargglyleuvalleupro354045glytyrlystyrleuglyproglyasnglyleuasplys505560<210>36<211>61<212>prt<213>人工序列<220><223>合成构建体<400>36metalaalaaspglytyrleuproasptrpleugluaspasnleuser151015gluglyileargglutrptrpalaleulysproglyalaproglnpro202530lyslysalaasnglnglnhisglnaspasnglyargglyleuvalleu354045proglylystyrleuglypropheasnglyleuasplys505560<210>37<211>60<212>prt<213>人工序列<220><223>合成构建体<400>37metthraspglytyrleuproasptrpleugluaspasnleuserglu151015glyvalargglutrptrpalaleuglnproglyalaprolysprolys202530alaasnglnglnhisglnaspasnalaargglyleuvalleuprogly354045tyrlystyrleuglyproglyasnglyleuasplys505560<210>38<211>60<212>prt<213>人工序列<220><223>合成构建体<400>38metserphevalasphisproproasptrpleuglugluvalglyglu151015glyleuargglupheleuglyleuglualaglyproprolysprolys202530proasnglnglnhisglnaspglnalaargglyleuvalleuprogly354045tyrasntyrleuglyproglyasnglyleuasparg505560<210>39<211>61<212>prt<213>人工序列<220><223>合成构建体<400>39metalaalaaspglytyrleuproasptrpleugluaspasnleuser151015gluglyileargglutrptrpalaleulysproglyalaproglnpro202530lysalaasnglnglnhisglnaspasnalaargglyleuvalleupro354045glytyrlystyrleuglyproglyasnglyleuasplys505560<210>40<211>61<212>prt<213>人工序列<220><223>合成构建体<220><221>变体<222>24<223>xaa=任何氨基酸<400>40metalaalaaspglytyrleuproasptrpleugluaspasnleuser151015gluglyileargglutrptrpxaaleulysproglyalaprolyspro202530lysalaasnglnglnlysglnaspaspglyargglyleuvalleupro354045glytyrlystyrleuglypropheasnglyleuasplys505560当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1