用于增加白介素-2及其衍生蛋白质的分泌水平的方法与流程
本发明涉及生物技术领域,特别地涉及用于将突变引入到白介素-2(il-2)的基因中的方法,所述引入导致所述分子和由其衍生的免疫调节突变蛋白质家族的分泌水平增加而不影响其生物学功能。
现有技术
最初被描述为t细胞的生长因子(smith,k.a.immunol.rev.51:337-357,1980)的il-2后来作为在免疫应答内具有双重功能的调节剂而出现(malek,t.r.annu.rev.immunol.26:453-479,2008;hoyerk.k.等人,immunol.rev.226:19-28,2008),其展示出促进或负地调节免疫系统的效应功能的能力。目前认为其基本作用与免疫耐受的维持相关(malek,t.r.和bayer,a.l.nat.rev.immunol.4:665-674,2004),通过刺激以组成性方式表达高水平的il-2受体的α链的调节t细胞。虽然β和γ亚基形成组成性地存在于免疫系统的效应细胞中的具有中等亲和力的二聚体受体,但是高水平的α链的组成性存在赋予调节t细胞以具有高亲和力的三聚体受体,其允许该细胞群体优先使用所述细胞因子(malek,t.r.和castro,i.immunity.33:153-165,2010)。
il-2的功能二分型已被利用用于产生对于免疫系统的相反的治疗效果并且在各种不同情景下以所希望的方向调节免疫应答。其免疫增强能力已被用于刺激抗肿瘤应答(klapper,j.a.等人,cancer.113:293-301,2008)。另一方面,通过应用不足以刺激效应t细胞或产生毒性效应的低剂量,il-2优先刺激调节t细胞的能力已被利用用于控制自身免疫(hartemann,a.等人,lancetdiabetesendocrinol.1:295-305,2013)和炎性(saadoun,d.等人,n.engl.j.med.365:2067-2077,2011)病症,以及移植物抗宿主疾病(koreth,a.等人,n.engl.j.med.365:2055-2066,2011)。
通过将突变引入到不同的与受体的亚基的结合界面中来将il-2的相互作用隔离已被建议作为用于获得具有各种不同免疫调节特性的突变蛋白质的途径。通过定向诱变来选择性地扰乱与α链的界面已使得能够获得称为非-α的分子,其具有降低的刺激调节t细胞的能力但保留了其对于携带β/γ二聚体受体的效应细胞的激动剂作用(carmenate,t.等人,j.immunol.190:6230-6238,2013;us9,206,243b2)。该分子在小鼠中具有强的抗肿瘤效应。另一方面,通过诱变破坏il-2与β和/或γ亚基的界面可以产生il-2受体的拮抗剂,其选择性地调节各种不同细胞群体的刺激(shanafelt,a.b.等人,nat.biotechnol.18:1197-1202,2000;wo2011/063770)。该类型的分子的例子为在us8,759,486b2中所描述的突变蛋白质m1和m2。
除了丧失了其相互作用能力的突变蛋白质之外,还已描述了由于其与受体的一个或另一个亚基的结合能力的增加而具有超级激动剂特性的il-2的突变型变体。对于β亚基的亲和力的增加导致获得强有力地刺激效应细胞并具有强的抗肿瘤效应的分子(levin,a.m.等人,nature.484:529-533,2012)。另一方面,il-2对于受体的α亚基的亲和力的增加已导致其他具有出众的在体外刺激t细胞增殖应答的能力的超级激动剂变体(wo2005/007121)。
已经通过合理设计、计算机筛选和在酵母细胞表面上所展示的il-2的定向进化获得了上面所描述的衍生自il-2的突变蛋白质。虽然已在丝状噬菌体上取得了在生物学上有活性的il-2的展示(buchli,p.j.等人,arch.biochem.biophys.339:79-84,1997;vispo,n.s.等人,immunotechnology3:185-193,1997),但是该技术平台还未被利用用于选择具有经修饰的特性的该细胞因子的新变体。
除了il-2及其衍生突变蛋白质的免疫调节特性之外,关于其治疗性利用的一个必不可少的要素是开发使得能够以足够的量获得其的系统。特别地,以实验室规模、以工业规模或者通过正常和/或肿瘤细胞或组织的转染或转导。
用于通过重组途径来产生il-2和其他相关分子的主要途径是在大肠杆菌(e.coli)的细胞质中进行表达,从而形成内含体,随后为在体外进行复性的程序(devos,r.等人,nucl.acidsres.11:4307-4323,1983;weir,m.p.和sparks,j.,biochem.j.245:85-91,1987)。虽然对于该策略已经证实了实用性,但是从一开始就导致获得与天然il-2非常相似的经正确折叠的分子的其他表达系统的探索仍在继续。在这些表达系统的一些中的il-2分泌受到il-2的聚集倾向的限制(halfmann,g.等人,j.gen.microbiol.139:2465-2473,1993;cha,h.j.等人,biochem.eng.j.24:225-233,2005)。
令人惊讶地,本发明的发明人发现了几个以前未曾描述过并且从人il-2的晶体结构的分析中也不可预测的突变,所述突变的引入增加了各种不同类型的细胞分泌重组人il-2和多种由其衍生的具有特殊免疫调节特性的突变型变体的能力。该发现确立了用于以生产规模使用这些突变的基础。
发明简述
在一个实施方案中,本发明基于导致在各种不同宿主中重组人il-2的分泌水平增加而不影响其生物学功能的方法。所述方法基于将单一突变引入到编码人il-2和其他由其衍生的多肽(其中包括但不限于:被设计用于作为拮抗剂、超级激动剂或选择性激动剂起作用的衍生自人il-2的突变蛋白质)的基因中。当使用本发明的方法时所述蛋白质的分泌水平的增加是相关于未突变的对应物而言的至少三倍。在本发明中,衍生突变蛋白质是指与人il-2具有大于90%的同一性的那些。
本发明的方法涉及这样的突变,其导致占据一级蛋白质序列的位置35的氨基酸(在原始序列中为lys)的非保守变化,优选地k35e、k35d和k35q这些置换。
特别地,本发明涉及按照在本文中所描述的方法而获得的蛋白质,其选自包括seqidno.1至18的组。
本发明的目标还为遗传构建体,其包括与其他核苷酸序列相融合的上面所描述的突变型基因,并且编码由il-2或其他由其衍生的免疫调节多肽和另外的蛋白质序列所形成的融合蛋白的合成。所述另外的蛋白质序列包括但不限于:丝状噬菌体的衣壳蛋白、白蛋白、抗体的fc区、完整抗体或包括其可变结构域的抗体片段。
在一个特别的实施方案中,用于获得上面所描述的分子的宿主包括但不限于:大肠杆菌、酵母和哺乳动物细胞(例如尤其是hek-293、cho、nso)。本发明的方法可用于改善il-2和其他由其衍生的多肽的产生效率,无论是以实验室规模还是以工业规模。通过本发明的方法而获得的蛋白质可以以治疗目的进行使用。
在另一个实施方案中,本发明的目标为通过在体外以及在体内表达人il-2和/或由其衍生的免疫调节突变蛋白质家族(单独的或者与其他蛋白质相融合的)来修饰正常或肿瘤细胞和/或组织的生理学。例子为,用于过继转移疗法的t淋巴细胞、b淋巴细胞或nk细胞的转导;或者肿瘤组织的直接转导/转染。本发明的方法可用于在这些背景下增加目的分子的分泌。
发明详述
本发明基于导致在各种不同宿主中重组人il-2的分泌水平增加而不影响其生物学功能的方法。所述方法基于将单一突变引入到编码人il-2和其他由其衍生的多肽(其中包括但不限于:被设计用于作为拮抗剂、超级激动剂或选择性激动剂起作用的衍生自人il-2的突变蛋白质)的基因中。
鉴定增加衍生自人il-2的蛋白质在丝状噬菌体上进行展示的能力的突变。
具有导致其在丝状噬菌体上的展示水平增加的单一突变的衍生自人il-2的多肽的选择可以从在丝状噬菌体上所展示的具有多于108个分子的文库开始来进行。可以将相应于所述变体的基因插入在噬菌粒类型的表达载体之内(与编码丝状噬菌体的衣壳蛋白的基因之一相融合),并且用于产生在其表面上展现所述蛋白质变体的病毒颗粒。起始文库可以沿着整个序列或者在一组预先定义的位置中包括各种不同程度的多样化。在这些位置处的原始残基中的每一个可以被替换为20种氨基酸的混合物或一个亚组的所选择的残基。所述多样化可以通过随机或位点定向诱变来取得。
具有增加的人il-2的展示水平的噬菌体的选择可以基于:温育与固定化在固体表面上的选择分子相接触的来自文库的噬菌体混合物,通过洗涤来去除未结合的噬菌体,和在干扰蛋白质相互作用的条件下洗脱经结合的噬菌体。作为选择分子,可以使用通过重组途径获得的il-2受体的亚基之一或者针对il-2或针对与其以基因方式融合的肽的单克隆抗体。可以在相似的条件下进行几个连续的选择循环。插入到所选择的噬菌粒中的dna序列的分析可以揭示出导致鉴定出更丰富的和与在噬菌体上的展示能力的增加潜在地相关的那些置换的规律性。
可以通过在等价地识别各种不同突变型变体和天然参考il-2的经固定化的捕获分子上的结合测定法(例如elisa)来评价il-2的突变型变体的展示水平。作为用于该类型的评价的捕获分子,优选针对与il-2的变体以基因方式融合的标志肽序列(例如c-myc肽,其在基于噬菌粒载体pcsm的表达系统中与所有外来蛋白质相融合)的抗体。
可以通过将所述变化引入到对于人il-2所描述的各种不同突变型变体的序列中来证实在上面所描述的筛选中所鉴定的突变对于衍生自il-2的突变蛋白质家族的效应的普遍性,这包括沿着其序列的可变数目的各种各样性质的置换,其旨在选择性地影响与il-2受体的各种不同亚基的相互作用,随之修饰其免疫调节功能。所有这些经修饰的突变蛋白质都被展示在丝状噬菌体上,从将其编码基因插入到噬菌粒类型的载体中开始。每种突变蛋白质在丝状噬菌体上的展示水平的评价可以通过elisa来进行,如对于il-2所描述的。作为用于计算与被鉴定为本发明的一部分的变化的引入相关联的在噬菌体上的展示增加程度的参考,使用没有任何额外的变化并且展示在噬菌体上的原始突变蛋白质。
备选地,可以通过利用其他组合式生物学平台(例如在酵母或哺乳动物细胞中的展示)来施行本发明的方法,以便选择具有增加的在细胞膜上的展示水平的il-2变体和/或其衍生突变蛋白质。
上面所描述的选择流程可以使得在位置35处出现非保守类型的频发突变(特别是k35e、k35d和k35q)。
使用鉴定出的突变来增加人il-2及其衍生突变蛋白质的分泌水平(作为可溶性蛋白质)以及其从内含体开始的复性。
一旦鉴定出了导致人il-2及其衍生突变蛋白质在丝状噬菌体上的展示增加的突变组,就可以从将其引入到克隆在用于酵母或哺乳动物细胞的可溶性表达载体中的相应的编码基因中开始来证实这些相同的变化对于可溶性蛋白质的分泌的效应。对于由包含所述表达载体的宿主细胞分泌到上清液中的蛋白质的浓度进行的评价使得能够证明与使用本发明的方法的突变的引入相关联的il-2及其衍生突变蛋白质的分泌的增加,相比于不包括所述变化的其原始对应物而言。
备选地,人il-2及其衍生突变蛋白质的产生的增加可以从在体内或在体外转染和/或转导正常和/或肿瘤细胞或/和组织开始来进行验证。
采用本发明的方法的上面所描述的研究可以用单独的或者与另外的多肽序列(例如,白蛋白、人免疫球蛋白的fc区、完整抗体或基于其可变区的抗体片段)相融合的il-2及其衍生突变蛋白质来进行。
在本发明中所描述的突变还可以用于改善作为内含体在大肠杆菌的细胞质中获得的人il-2及其衍生突变蛋白质的体外复性过程。复性效率的增加可以通过以与未突变的变体相比较的方式测量“比生物学活性/蛋白质质量”来进行评价。
证实所使用的突变与il-2的生物学功能以及il-2与受体亚基的相互作用的选择性调节的相容性。
对于根据本发明的方法进行修饰的il-2变体的生物学活性的评价可以包括旨在证明其诱导各种不同细胞类型(例如,t淋巴细胞、nk细胞和依赖于il-2来进行其生长的淋巴起源的细胞系这些亚群体)的增殖、分化和激活的能力的保存的体外和体内技术。天然il-2对于表达三聚体受体的t淋巴细胞的增殖的效应可以通过采用alamar蓝还原比色技术的ctll-2细胞系体外增殖测定法或者通过流式细胞术来进行测定。天然il-2在体外对于cd4+t淋巴细胞向调节t淋巴细胞的分化的效应以及该分子在体外扩展和激活nk细胞的能力通过流式细胞术来进行测定。
本发明的方法中所使用的突变与il-2与其受体的相互作用的选择性调节的相容性可以从将所述变化引入到先前被设计和/或选择用于增加或减小其与il-2受体的亚基中的任一个的结合能力的突变蛋白质的框架上开始来进行证明。在结合特性中所希望的变化的出现可以通过在用受体亚基中的每一种包被的微量滴定板上在elisa类型的实验中直接测定它们来进行证实。作为另外的验证工具,可以采用先前被描述用于在体外和在体内表征各种不同突变蛋白质的免疫调节和/或抗肿瘤活性的测定法。对于非-α(no-alpha)突变蛋白质(carmenate,t.等人,j.immunol.190:6230-6238,2013),可以证实其保持了与天然il-2相等的在体外刺激cd8+t淋巴细胞的增殖的能力。在具有增加的与受体的β亚基的结合能力且具有超级激动剂活性的突变蛋白质(超-β(super-beta)突变蛋白质)的情况下,可以证实其保持了比天然il-2更大的在体外刺激nk细胞的增殖的能力。在这两种情况下,可以通过流式细胞术来测定增殖。在体内对于群体的增殖的差别效应可以通过溴脱氧尿苷掺入实验来进行测定。可以证实,在采用黑素瘤系mb16f0的实验性转移模型中,这两种突变蛋白质均诱导比天然il-2更大的体内抗肿瘤效应。
附图简述
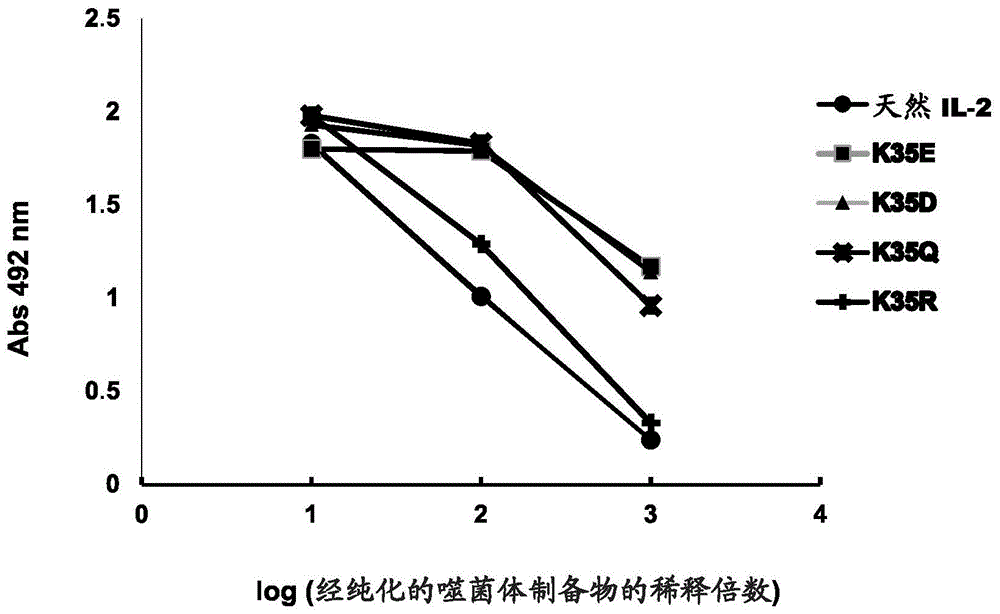
图1.通过elisa来评价突变型il-2在丝状噬菌体上的展示水平。将经纯化的各种不同噬菌体制备物调整至1013个病毒颗粒/ml的等价浓度。
图2.通过elisa来测定由处于悬浮液中的hek293-t细胞进行的由il-2及其衍生突变蛋白质和人igg1的fc区形成的融合蛋白的分泌水平。用聚乙烯亚胺和编码由下列各项形成的融合蛋白的遗传构建体转染细胞:
a.il-2和具有突变k35e的il-2,
b.非-α(no-alpha)il-2(il-2na)和具有额外突变k35e的非-αil-2,
c.超-β(super-beta)il-2(il-2sb)和具有额外突变k35e的超-βil-2,
d.非-γ(no-gamma)m1il-2(il-2ngm1)和具有额外突变k35e的非-γm1il-2,
e.非-γm2il-2(il-2ngm2)和具有额外突变k35e的非-γm2il-2。
图3.通过elisa来评价在k35e变体中天然il-2的分子相互作用的保存。
图4.使用ctll-2细胞系增殖技术,具有k35e这一变化的il-2的生物学活性的保存。
图5.il-2k35e在体内扩展依赖于il-2的细胞群体的能力。
5a.用il-2的变体k35e/fc2.2.2和pbs进行注射的小鼠的脾的照片。
5b.从在脾中的具有记忆表型(cd44hi)的cd3+cd8+细胞群体的通过流式细胞术的分析之中获得的直方图。
图6.通过elisa来证实k35e置换与对于衍生自il-2的突变蛋白质所描述的与受体的α亚基的结合能力的丧失具有相容性。用人(a)和小鼠(b)来源的重组α链包被微量滴定板。
图7.通过elisa来证实k35e置换与对于衍生自il-2的突变蛋白质所描述的与受体的β亚基的结合能力的增加具有相容性。
实施例
实施例1.展示功能性突变型人il-2的丝状噬菌体的选择和表征
构建人il-2的一组位置的轻度随机化文库。选择由其侧链位于与受体的α亚基的界面处的残基所占据的那些位置(k35、r38、t41、f42、k43、f44、y45、e61、e62、k64、p65、e68、v69、n71、l72、q74和y107)。为了使这些位置随机化,用倾斜的诱变寡核苷酸通过kunkel诱变来使人il-2基因多样化,所述诱变寡核苷酸在待修饰的三联体的每个位置处保存了85%的原始核苷酸,具有15%的其余三种核苷酸的等摩尔混合物,以在整个区域中引入中等程度的多样性。以这种方式,尽管由109个克隆组成的文库以其整体探查在界面的每个位置处20种氨基酸的存在,但是在文库内的每个分子相对于原始il-2而言仅包含几个置换,从而将新多肽的寻找限制于更接近起始分子的功能序列空间。
按照先前建立的程序通过用聚乙二醇进行沉淀来纯化从文库开始产生的噬菌体(marks,j.等人,j.mol.biol.222:581-597,1991)。为了依据其在噬菌体上展示的能力来分离人il-2的功能性突变型变体,在用il-2受体的重组α亚基(r&d)包被的immunotube(nunc,dinamarca)上温育所述病毒颗粒。在人和小鼠的重组α亚基上进行两个独立的选择程序。在进行洗涤以去除未结合的噬菌体后,通过与碱性三乙胺溶液一起进行温育来洗脱经结合的噬菌体。用所选择的噬菌体(其用辅助噬菌体m13ko7进行了扩增)感染tg1菌株的细菌,并且将其用作起始材料以用于新一轮选择。进行四个循环的噬菌体选择。对于插入在所选择的噬菌粒(来自第三个和第四个选择循环)中的基因所进行的测序揭示了在所获得的突变型变体之间的规律性。虽然原始il-2(在起始文库中是高占比的)的分离是占优势的,但是也观察到少数比例的具有突变k35e、k35d和k35q的变体,这证实了在位置35处的非保守变化对于功能性il-2在丝状噬菌体上的展示的正面影响。突变k35e是最常见的。该发现是令人惊讶的,因为“il-2/受体”复合物(pdb编号3b5i和2erj)的晶体结构的分析表明原始残基k35牵涉在与受体的α亚基的界面的极性周围区域中的离子相互作用,并且非保守变化(在这两种情况下都是电荷的反转)维持与选择分子相互作用的能力因此是出人意料的。
实施例2.证实在位置35处具有非保守变化的人il-2的分泌以及在丝状噬菌体上的展示增加
比较了从文库开始选择出的人il-2的各种不同突变型变体分泌到大肠杆菌的周质中和在丝状噬菌体的表面上展示的能力。使用天然il-2作为参考分子。此外,通过kunkel诱变构建了k35r变体(在位置35处的保守变化)作为额外的对照。所有这些蛋白质通过将其编码基因插入到噬菌粒载体pcsm(与噬菌体m13的基因3相融合)中并随后从用所述遗传构建体转染的大肠杆菌tg1菌株的细菌开始产生噬菌体来获得(rojas,g.等人,immunobiology.218:105-113,2013)。通过在用单克隆抗体9e10包被的微量滴定板上的elisa来评价所述变体中的每一种在噬菌体上的展示水平。检测与缀合至辣根过氧化物酶的抗-m13抗体相结合的噬菌体。证实了k35e、k35d和k35q变化引起人il-2在噬菌体上的展示的增加,相对于原始分子而言(图1)。该增加的大小程度在k35e和k35d的情况下(在这两者中都是电荷的反转)为10倍,和对于突变k35q,为7倍。相反地,保守变化k35r没有改变人il-2在噬菌体上展示的能力(图1)。选择k35e变化用于后续研究。
实施例3.突变k35e对于分泌和噬菌体展示的效应扩展至一组il-2的突变型变体
通过kunkel诱变将突变k35e引入到人il-2的几个突变型变体的编码基因中(以在丝状噬菌体上展示的形式)。衍生自il-2的分子的组包括先前所描述的具有各种不同免疫调节功能的四种突变蛋白质:非-α突变蛋白质,其具有对于效应t细胞的选择性激动剂活性(carmenate,t.等人,j.immunol.190:6230-6238,2013;us9,206,243b2);丧失了与受体的γ-亚基相互作用的能力的突变蛋白质(非-γ),其具有拮抗剂功能(us8,759,486b2);具有增加的与受体的β亚基的结合能力的突变蛋白质(超-β),其具有超级激动剂活性;和具有增加的与受体的α亚基的结合能力的突变蛋白质(超-α),其具有超级激动剂活性(levin,a.m.等人,nature.484:529-533,2012;wo2005/007121)。产生并纯化展示出这些变体中的每一种(还有在位置35处没有变化的原始分子)的噬菌体,并且通过用单克隆抗体9e10包被的微量滴定板上的elisa来评价外源蛋白质的展示水平。使用经纯化的展示出天然il-2的噬菌体制备物作为参考(采取100任意单位/ml的浓度)。从该制备物开始构建标准曲线并计算对于所述变体中的每一种的相对展示水平。表1显示了由k35e变化的引入而引起的每种突变蛋白质在噬菌体上的展示水平的增加。
表1.由k35e变化的引入而引起的所检定的突变蛋白质在噬菌体上的展示水平的增加。
实施例4.k35e置换增加基于il-2及其衍生突变蛋白质的融合蛋白从人宿主细胞开始的分泌
基于克隆在表达载体pcmx中的人il-2及其衍生突变蛋白质的基因(与人igg1的fc区的基因相融合),设计遗传构建体。构建另外一组其中引入了k35e置换的等价分子。用与聚乙烯亚胺相混合的前面的遗传构建体中的每一种转染人hek293-t细胞(其适应于悬浮生长)。每次转染的最终体积为50ml。在6天的培养后,收集经转染的细胞的上清液。通过在用单克隆抗体il-2.2(其识别在所分析的那组突变蛋白质中存在的线性表位)包被的微量滴定板上的elisa来评价在上清液中重组il-2及其衍生突变蛋白质的存在。用标记有辣根过氧化物酶的人抗-fc抗体检测所捕获的融合蛋白。在上清液中融合蛋白的水平对于包含突变k35e的分子来说是更高的,相比于其对应物而言(图2a-e)。通过与a蛋白基质的亲和性来纯化所述重组蛋白质。表2显示了在每个纯化过程后获得的量。
表2.从处于悬浮液中的经转染的hek-293t细胞开始来纯化与人免疫球蛋白的fc区相融合的il-2及其衍生突变蛋白质的收益。
实施例5.突变k35e与天然il-2的分子相互作用相容
通过elisa来评价重组突变型il-2(k35e)(以其与人免疫球蛋白的fc片段相融合的同二聚体形式)与一组能够与天然il-2相结合的分子相互作用的能力。用四种识别人il-2的各种不同区域的单克隆抗体中的每一种或者用人或小鼠来源的il-2受体的α亚基包被微量滴定板。用标记有辣根过氧化物酶的抗人fc抗体来检测所捕获的突变型il-2。作为对照,使用不具有k35e变化的由处于悬浮液中的hek293-t细胞产生的相似的重组同二聚体。突变型同二聚体与所述抗体和与所述α亚基的结合不受该突变的存在的影响。相反地,突变型变体对于所有其他分子的反应性高于其未突变的重组对应物的反应性(图3),这表明k35e变体的抗原性和功能性以比在相似条件下获得的未突变的重组蛋白质更大的程度再现了天然il-2的抗原性和功能性。
实施例6.与人免疫球蛋白的fc区相融合的il-2的k35e变体保存了刺激ctll-2细胞的增殖的能力
评价具有k35e变化的人il-2(其在处于悬浮液中的经转染的hek293-t细胞中作为与人igg1的fc区的融合蛋白而获得)诱导ctll-2细胞系的增殖的能力。作为对照,使用重组人il-2。在一系列浓度的经纯化的所述两种蛋白质中的每一种存在下培养细胞,并且在着色剂alamar蓝还原比色测定法中测定增殖(图4)。每种蛋白质的比活性的计算用graphpad程序从产生50%的最大增殖水平的分子剂量开始来进行。包含k35e变化的与fc相融合的il-2的比活性为4×106iu/mg,在与用作参考的重组il-2的比活性(2.3×106iu/mg)相同的范围内。该结果证明了在突变k35e存在下il-2的生物学活性的保存。
实施例7.与人免疫球蛋白的fc区相融合的il-2的k35e变体具有在体内刺激记忆cd8t细胞的扩展的能力
为了评价与fc相融合的重组突变型il-2(k35e)在体内刺激依赖于il-2的细胞群体的扩展的能力,用五个日剂量的4×104iu的该分子对c57bl/6小鼠进行注射。在处理结束时处死动物并评价其脾的大小。通过流式细胞术来测定在脾中具有记忆表型(cd44hi)的cd3+cd8+细胞群体的大小。作为对照,使用用磷酸盐缓冲盐水(pbs)进行注射的小鼠。与fc相融合的突变型il-2(k35e)具有所预期的刺激记忆cd8t细胞的扩展的效应,其中引起脾增大(图5a)和在它们中具有记忆表型的t细胞的比例增加(加倍)(图5b)。
实施例8.k35e置换与决定选择性激动剂的特性的与il-2受体的α亚基的结合能力的丧失相容
通过elisa来评价人il-2和先前由carmenate,t.等人,j.immunol.190:6230-6238,2013;us9,206,243所描述的非-α突变蛋白质(其特征在于具有突变r38a、f42a、y45a和e62a,并且丧失了与il-2受体的α亚基的结合能力,旨在减小对于调节t细胞的刺激,而不影响对于携带β/γ二聚体受体的效应细胞的激动剂作用)的结合能力。人il-2以及非-α突变蛋白质均具有额外的修饰k35e并且与人igg1的fc区相融合。用人(a)和小鼠(b)来源的重组α链包被微量滴定板。用标记有辣根过氧化物酶的抗人fc抗体来检测所捕获的融合蛋白。将k35e变化引入其中导致具有高于原始突变蛋白质的表达水平的表达水平的新分子(图2b),并且还显示出相对于也包含k35e变化的天然il-2而言其与人以及小鼠的受体的α链的结合能力的严重降低(图6)。这些结果提供了关于下述方面的初始证据:突变k35e与衍生自il-2的突变蛋白质的相互作用和免疫调节功能的选择性操纵的相容性。
实施例9.k35e置换与对于超级激动剂变体所描述的与il-2受体的β亚基的相互作用的增加相容
在用重组人β链包被的微量滴定板上通过elisa来评价il-2以及以突变l80f、r81d、l85v、i86v和i92f为特征的超-β突变蛋白质(两者均具有额外的修饰k35e并且与人igg1的fc区相融合)的结合能力。用标记有辣根过氧化物酶的抗人fc抗体来检测所捕获的融合蛋白。k35e变化的引入导致具有高于原始突变蛋白质的表达水平的表达水平(图2c)并且具有增加的与il-2受体的β亚基相结合的能力(其是原始分子的超级激动剂作用的基础)(图7)的新分子。该结果扩大了关于下述方面的证据:突变k35e与衍生自il-2的新分子的设计的相容性,所述新分子在其与受体的各种不同亚基的相互作用方面和在其免疫调节功能方面具有变化。
序列表
<110>centrodeinmunologiamolecular
<120>用于增加白介素-2及其衍生蛋白质的分泌水平的方法
<130>542016cu00
<160>18
<170>patentinversion3.5
<210>1
<211>133
<212>prt
<213>人工序列
<220>
<223>通过dna重组技术
<400>1
alaprothrserserserthrlyslysthrglnleuglnleugluhis
151015
leuleuleuaspleuglnmetileleuasnglyileasnasntyrlys
202530
asnprogluleuthrargmetleuthrphelysphetyrmetprolys
354045
lysalathrgluleulyshisleuglncysleugluglugluleulys
505560
proleuglugluvalleuasnleualaglnserlysasnphehisleu
65707580
argproargaspleuileserasnileasnvalilevalleugluleu
859095
lysglysergluthrthrphemetcysglutyralaaspgluthrala
100105110
thrilevalglupheleuasnargtrpilethrpheserglnserile
115120125
ileserthrleuthr
130
<210>2
<211>133
<212>prt
<213>人工序列
<220>
<223>通过dna重组技术
<400>2
alaprothrserserserthrlyslysthrglnleuglnleugluhis
151015
leuleuleuaspleuglnmetileleuasnglyileasnasntyrlys
202530
asnproaspleuthrargmetleuthrphelysphetyrmetprolys
354045
lysalathrgluleulyshisleuglncysleugluglugluleulys
505560
proleuglugluvalleuasnleualaglnserlysasnphehisleu
65707580
argproargaspleuileserasnileasnvalilevalleugluleu
859095
lysglysergluthrthrphemetcysglutyralaaspgluthrala
100105110
thrilevalglupheleuasnargtrpilethrpheserglnserile
115120125
ileserthrleuthr
130
<210>3
<211>133
<212>prt
<213>人工序列
<220>
<223>通过dna重组技术
<400>3
alaprothrserserserthrlyslysthrglnleuglnleugluhis
151015
leuleuleuaspleuglnmetileleuasnglyileasnasntyrlys
202530
asnproglnleuthrargmetleuthrphelysphetyrmetprolys
354045
lysalathrgluleulyshisleuglncysleugluglugluleulys
505560
proleuglugluvalleuasnleualaglnserlysasnphehisleu
65707580
argproargaspleuileserasnileasnvalilevalleugluleu
859095
lysglysergluthrthrphemetcysglutyralaaspgluthrala
100105110
thrilevalglupheleuasnargtrpilethrpheserglnserile
115120125
ileserthrleuthr
130
<210>4
<211>133
<212>prt
<213>人工序列
<220>
<223>通过dna重组技术
<400>4
alaprothrserserserthrlyslysthrglnleuglnleugluhis
151015
leuleuleuaspleuglnmetileleuasnglyileasnasntyrlys
202530
asnprogluleuthralametleuthralalysphealametprolys
354045
lysalathrgluleulyshisleuglncysleugluglualaleulys
505560
proleuglugluvalleuasnleualaglnserlysasnphehisleu
65707580
argproargaspleuileserasnileasnvalilevalleugluleu
859095
lysglysergluthrthrphemetcysglutyralaaspgluthrala
100105110
thrilevalglupheleuasnargtrpilethrpheserglnserile
115120125
ileserthrleuthr
130
<210>5
<211>133
<212>prt
<213>人工序列
<220>
<223>通过dna重组技术
<400>5
alaprothrserserserthrlyslysthrglnleuglnleugluhis
151015
leuleuleuaspleuglnmetileleuasnglyileasnasntyrlys
202530
asnproaspleuthralametleuthralalysphealametprolys
354045
lysalathrgluleulyshisleuglncysleugluglualaleulys
505560
proleuglugluvalleuasnleualaglnserlysasnphehisleu
65707580
argproargaspleuileserasnileasnvalilevalleugluleu
859095
lysglysergluthrthrphemetcysglutyralaaspgluthrala
100105110
thrilevalglupheleuasnargtrpilethrpheserglnserile
115120125
ileserthrleuthr
130
<210>6
<211>133
<212>prt
<213>人工序列
<220>
<223>通过dna重组技术
<400>6
alaprothrserserserthrlyslysthrglnleuglnleugluhis
151015
leuleuleuaspleuglnmetileleuasnglyileasnasntyrlys
202530
asnproglnleuthralametleuthralalysphealametprolys
354045
lysalathrgluleulyshisleuglncysleugluglualaleulys
505560
proleuglugluvalleuasnleualaglnserlysasnphehisleu
65707580
argproargaspleuileserasnileasnvalilevalleugluleu
859095
lysglysergluthrthrphemetcysglutyralaaspgluthrala
100105110
thrilevalglupheleuasnargtrpilethrpheserglnserile
115120125
ileserthrleuthr
130
<210>7
<400>7
000
<210>8
<211>133
<212>prt
<213>人工序列
<220>
<223>通过dna重组技术
<400>8
alaprothrserserserthrlyslysthrglnleuglnleugluhis
151015
leuleuleuaspleuvalmetileleuasnglyileasnasntyrlys
202530
asnproaspleuthrargmetleuthrphelysphetyrmetprolys
354045
lysalathrgluleulyshisleuglncysleugluglugluleulys
505560
proleuglugluvalleuasnleualaglnserlysasnphehisleu
65707580
argproargaspleuileserasnileasnvalilevalleugluleu
859095
lysglysergluthrthrphemetcysglutyralaaspgluthrala
100105110
thrilevalglupheleuasnargtrpilethrpheseralaserile
115120125
aspglythrleuthr
130
<210>9
<211>133
<212>prt
<213>人工序列
<220>
<223>通过dna重组技术
<400>9
alaprothrserserserthrlyslysthrglnleuglnleugluhis
151015
leuleuleuaspleuvalmetileleuasnglyileasnasntyrlys
202530
asnproglnleuthrargmetleuthrphelysphetyrmetprolys
354045
lysalathrgluleulyshisleuglncysleugluglugluleulys
505560
proleuglugluvalleuasnleualaglnserlysasnphehisleu
65707580
argproargaspleuileserasnileasnvalilevalleugluleu
859095
lysglysergluthrthrphemetcysglutyralaaspgluthrala
100105110
thrilevalglupheleuasnargtrpilethrpheseralaserile
115120125
aspglythrleuthr
130
<210>10
<211>133
<212>prt
<213>人工序列
<220>
<223>通过dna重组技术
<400>10
alaprothrserserserthrlyslysthrglnleuglnleugluhis
151015
leuasnleuaspleuglnmetileleuasnglyileasnasntyrlys
202530
asnprogluleuthrargmetleuthrphelysphetyrmetprolys
354045
lysalathrgluleulyshisleuglncysleugluglugluleulys
505560
proleuglugluvalleuasnleualaglnserlysasnphehisleu
65707580
argproargaspleuileserasnileasnvalilevalleugluleu
859095
lysglysergluthrthrphemetcysglutyralaaspgluthrala
100105110
thrilevalglupheleuasnargtrpilethrphesertyrserile
115120125
ileargthrleuthr
130
<210>11
<211>133
<212>prt
<213>人工序列
<220>
<223>通过dna重组技术
<400>11
alaprothrserserserthrlyslysthrglnleuglnleugluhis
151015
leuasnleuaspleuglnmetileleuasnglyileasnasntyrlys
202530
asnproaspleuthrargmetleuthrphelysphetyrmetprolys
354045
lysalathrgluleulyshisleuglncysleugluglugluleulys
505560
proleuglugluvalleuasnleualaglnserlysasnphehisleu
65707580
argproargaspleuileserasnileasnvalilevalleugluleu
859095
lysglysergluthrthrphemetcysglutyralaaspgluthrala
100105110
thrilevalglupheleuasnargtrpilethrphesertyrserile
115120125
ileargthrleuthr
130
<210>12
<211>133
<212>prt
<213>人工序列
<220>
<223>通过dna重组技术
<400>12
alaprothrserserserthrlyslysthrglnleuglnleugluhis
151015
leuasnleuaspleuglnmetileleuasnglyileasnasntyrlys
202530
asnproglnleuthrargmetleuthrphelysphetyrmetprolys
354045
lysalathrgluleulyshisleuglncysleugluglugluleulys
505560
proleuglugluvalleuasnleualaglnserlysasnphehisleu
65707580
argproargaspleuileserasnileasnvalilevalleugluleu
859095
lysglysergluthrthrphemetcysglutyralaaspgluthrala
100105110
thrilevalglupheleuasnargtrpilethrphesertyrserile
115120125
ileargthrleuthr
130
<210>13
<211>133
<212>prt
<213>人工序列
<220>
<223>通过dna重组技术
<400>13
alaprothrserserserthrlyslysthrglnleuglnleugluhis
151015
leuleuleuaspleuglnmetileleuasnglyileasnasntyrlys
202530
asnprogluleuthrargmetleuthrphelysphetyrmetprolys
354045
lysalathrgluleulyshisleuglncysleugluglugluleulys
505560
proleuglugluvalleuasnleualaglnserlysasnphehisphe
65707580
aspproargaspvalvalserasnileasnvalphevalleugluleu
859095
lysglysergluthrthrphemetcysglutyralaaspgluthrala
100105110
thrilevalglupheleuasnargtrpilethrpheserglnserile
115120125
ileserthrleuthr
130
<210>14
<211>133
<212>prt
<213>人工序列
<220>
<223>通过dna重组技术
<400>14
alaprothrserserserthrlyslysthrglnleuglnleugluhis
151015
leuleuleuaspleuglnmetileleuasnglyileasnasntyrlys
202530
asnproaspleuthrargmetleuthrphelysphetyrmetprolys
354045
lysalathrgluleulyshisleuglncysleugluglugluleulys
505560
proleuglugluvalleuasnleualaglnserlysasnphehisphe
65707580
aspproargaspvalvalserasnileasnvalphevalleugluleu
859095
lysglysergluthrthrphemetcysglutyralaaspgluthrala
100105110
thrilevalglupheleuasnargtrpilethrpheserglnserile
115120125
ileserthrleuthr
130
<210>15
<211>133
<212>prt
<213>人工序列
<220>
<223>通过dna重组技术
<400>15
alaprothrserserserthrlyslysthrglnleuglnleugluhis
151015
leuleuleuaspleuglnmetileleuasnglyileasnasntyrlys
202530
asnproglnleuthrargmetleuthrphelysphetyrmetprolys
354045
lysalathrgluleulyshisleuglncysleugluglugluleulys
505560
proleuglugluvalleuasnleualaglnserlysasnphehisphe
65707580
aspproargaspvalvalserasnileasnvalphevalleugluleu
859095
lysglysergluthrthrphemetcysglutyralaaspgluthrala
100105110
thrilevalglupheleuasnargtrpilethrpheserglnserile
115120125
ileserthrleuthr
130
<210>16
<211>133
<212>prt
<213>人工序列
<220>
<223>通过dna重组技术
<400>16
alaprothrserserserthrlyslysthrglnleuglnleugluhis
151015
leuleuleuaspleuglnmetileleuasnglyileasnasntyrlys
202530
asnprogluleuthrargmetleuthrphelysphetyrmetprolys
354045
lysalathrgluleulyshisleuglncysleugluglugluleulys
505560
proleugluglualaleuasnleualaproserlysasnphehisleu
65707580
argproargaspleuileserasnileasnvalilevalleugluleu
859095
lysglysergluthrthrphemetcysglutyralaaspgluthrala
100105110
thrilevalglupheleuasnargtrpilethrpheserglnserile
115120125
ileserthrleuthr
130
<210>17
<211>133
<212>prt
<213>人工序列
<220>
<223>通过dna重组技术
<400>17
alaprothrserserserthrlyslysthrglnleuglnleugluhis
151015
leuleuleuaspleuglnmetileleuasnglyileasnasntyrlys
202530
asnproaspleuthrargmetleuthrphelysphetyrmetprolys
354045
lysalathrgluleulyshisleuglncysleugluglugluleulys
505560
proleugluglualaleuasnleualaproserlysasnphehisleu
65707580
argproargaspleuileserasnileasnvalilevalleugluleu
859095
lysglysergluthrthrphemetcysglutyralaaspgluthrala
100105110
thrilevalglupheleuasnargtrpilethrpheserglnserile
115120125
ileserthrleuthr
130
<210>18
<211>133
<212>prt
<213>人工序列
<220>
<223>通过dna重组技术
<400>18
alaprothrserserserthrlyslysthrglnleuglnleugluhis
151015
leuleuleuaspleuglnmetileleuasnglyileasnasntyrlys
202530
asnproglnleuthrargmetleuthrphelysphetyrmetprolys
354045
lysalathrgluleulyshisleuglncysleugluglugluleulys
505560
proleugluglualaleuasnleualaproserlysasnphehisleu
65707580
argproargaspleuileserasnileasnvalilevalleugluleu
859095
lysglysergluthrthrphemetcysglutyralaaspgluthrala
100105110
thrilevalglupheleuasnargtrpilethrpheserglnserile
115120125
ileserthrleuthr
130
- 还没有人留言评论。精彩留言会获得点赞!