国兰花器官发育调控基因及其编码蛋白与应用的制作方法
本发明涉及植物基因工程技术领域,具体涉及国兰花器官发育调控基因及其编码蛋白与应用。
背景技术
兰科植物拥有高度特化的花器官和丰富的生物多样性,是珍贵的园艺观赏花卉。全世界已发现801属3万余种,按生态习性可分为地生兰、气生兰、腐生兰三大类。其中兰属地生种大多原产中国,因此又称国兰,被列为中国十大名花之一,主要包括建兰、墨兰、春兰、蕙兰、莲瓣兰、豆瓣兰、寒兰和春剑等8大种类,具有很高经济价值。植物分子生物学研究发现,b类mads-box基因表达区域扩张展第1轮花器官,是造成多数单子叶植物第1、2轮花器官发育成类似花被片结构的分子基础。然而兰花与其他一些单子叶植物如百合(liliumbrownii)、郁金香(tulipagesneriana)、百子莲(agapanthuspraecox)等不同,最外层三个花萼通常可分为一背侧(dorsal)及二个侧生(lateral)花萼,其外形相近,而内层三个花瓣分为二个侧生花瓣和另一个特化卷曲的唇瓣(liporlabellum)。mondragon-palomino等(2011)通过对蝴蝶兰、香荚兰和美洲长叶兜兰的研究,提出花器官发育“兰花密码(orchidcode)”,认为b类mads-box基因中的ap3类基因在早期经过两轮复制,形成4个亚族,分别促进花萼与花瓣的分离,侧瓣与唇瓣的分离,从而特异性调控不同花器官形态建成。然而有研究表明,兰科植物不同属种的b类mads-box基因家族成员数量各不相同,不同亚族成员的表达模式和功能也有较大差异。
国兰的花瓣变异类型远比蝴蝶兰、文心兰等热带气生兰丰富,在人工驯化选育过程中,已培育出了梅瓣、荷瓣、水仙瓣、蝴蝶瓣等多种萼片及花瓣形态变异,以及少瓣或无瓣化、多瓣化、树形花等多种变异类型,显著提高其观赏价值和经济价值,但目前国兰品种主要依靠野生资源驯化选育,适宜规模化、产业化的品种不多,杂交育种周期长、难度大且子代性状难以预期,现代分子生物技术育种手段严重滞后。如何通过分子生物技术手段进行观赏性状改良,缩短育种周期,高效培育新品种,成为目前亟待解决的科学问题。
技术实现要素:
本发明的目的在于:克服传统国兰杂交育种存在周期长、性状难以预期等问题,本发明通过运用最新的分子生物学手段,挖掘可运用于植物花型改良的基因资源,实现植物花朵瓣型的性状改良。
为了实现上述发明目的,本发明提供了一种国兰花器官发育调控基因,它是从中国兰花的墨兰品种“达摩”(cymbidiumsinense‘dharma’)的花器官cdna中分离得到,其核苷酸序列由669个碱基组成,如seqidno:1所示。
本发明的第二个目的是提供由上述国兰花器官发育调控基因编码的蛋白,该蛋白由222个氨基酸残基组成,如seqidno:2所示。
为了实现上述发明目的,本发明还提供了包含所述国兰花器官发育调控基因的表达载体、转基因细胞系、宿主菌和转基因材料。
本发明通过基因工程技术发现,所述国兰花器官发育调控基因在拟南芥和兰花中过量表达,能改变植物花型,造成花瓣卷曲,兰花花瓣向唇瓣转变,唇瓣性状加强,因此,可将该基因及其编码的蛋白用于兰科植物花器官发育调控途径的研究,以及植物开花性状花朵瓣型的改良。
与现有技术相比,本发明具有以下有益效果:
本发明从中国兰花的墨兰品种“达摩”(cymbidiumsinense‘dharma’)的花器官cdna中分离得到兰花的花器官发育调控基因,通过提高其表达量后可造成拟南芥四轮花器官由辐射对称变成两侧对称,花瓣向上卷曲类似唇瓣结构;在国兰中高表达上述兰花花器官发育调控基因后,使花瓣向唇瓣转变,唇瓣增多。本发明提供的兰花花器官发育调控基因及其编码的调控蛋白可用于兰科植物花器官发育分子机制的研究,以及植物开花性状花朵瓣型的改良。
附图说明
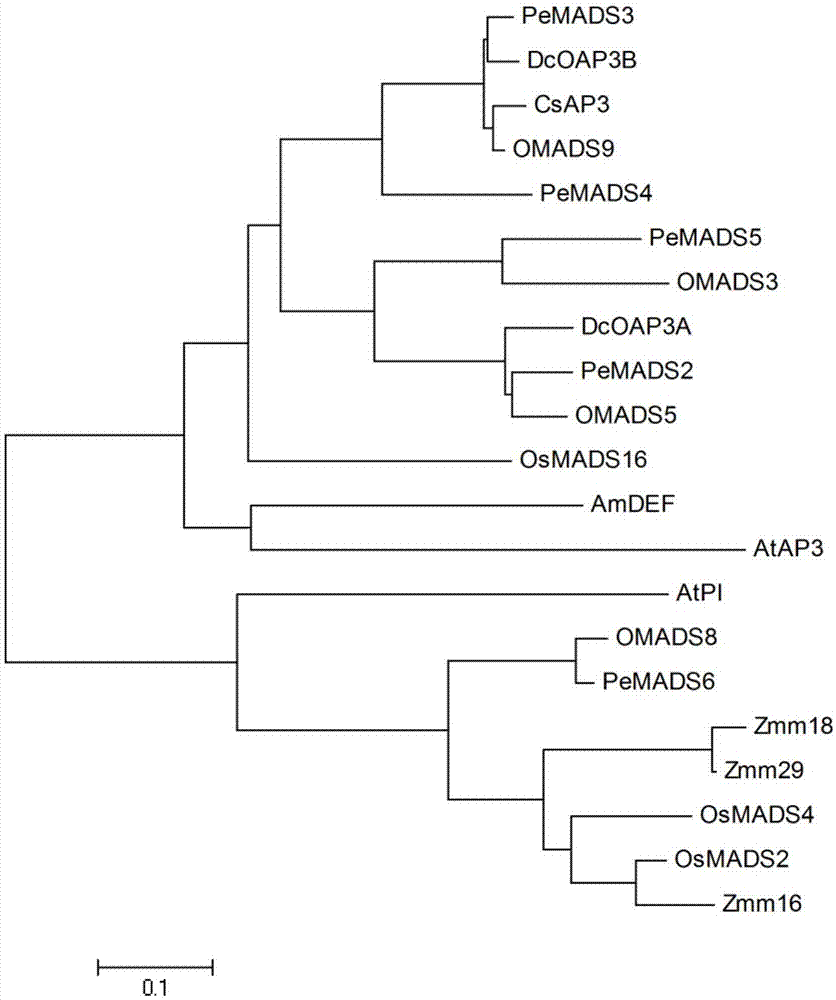
图1为本发明实施例1中,csap3与其他物种来源的ap3进化树分析及多重序列比较。
图2为本发明实施例2中,csap3在墨兰中的表达模式分析图。
图3为本发明实施例3中,转基因拟南芥植株csap3表达量分析,其中oe1-6分别表示转基因拟南芥植株。
图4为本发明实施例3中,转基因拟南芥表型分析:花瓣卷曲,其中wt表示野生型,csap3-oe1、csap3-oe2、csap3-oe3表示转基因植株。
图5为本发明实施例4中,国兰体内csap3基因瞬时高表达及表型分析,ck表示含有空载体的农杆菌液注射植株,csap3-oe表示含有pcam1301-csap3质粒的农杆菌液注射植株。
具体实施方式
为了使本发明的目的、技术方案和有益技术效果更加清晰,以下结合实施例,对本发明进行进一步详细说明。应当理解的是,本说明书中描述的实施例仅仅是为了解释本发明,并非为了限定本发明,实施例的参数、比例等可因地制宜做出选择而对结果并无实质性影响。
实施例1csap3基因的克隆以及序列分析
1.rna的提取
取墨兰栽培品种达摩花器官组织2g,利用植物trizol(invitrogen)试剂抽提其总rna,并反转录成cdna(thermoscientificrevertaidfirststrandcdnasynthesiskit)。
2.目的基因csap3的获得
以csap3-f1:atggggagagggaagatagagat(seqidno:3)和csap3-r1:tcaagcgagacgcagatcat(seqidno:4)为引物,以上述步骤1所得cdna为模板,利用ex-taq酶(takarabiotechnologyco.)进行pcr,按照下列条件:94℃4min,然后34cycles(94℃40s,59.5℃40s,72℃1.5min,72℃10min)。从琼脂糖凝胶中回收pcr产物,然后连接pmd19-t载体(takarabiotechnologyco.)并送华大基因研究院进行测序。测序结果分析发现扩增片段含有目的基因csap3的完整cds序列,由669个碱基组成,核苷酸序列如seqidno:1所示,命名为墨兰花器官发育调控基因(csap3基因),其编码的蛋白的氨基酸序列由222个氨基酸残基组成,如seqidno:2所示,命名为墨兰花器官发育调控蛋白(csap3蛋白)。获得的含有csap3-pmd19-t的大肠杆菌目前保存于广东省农业科学院环境园艺研究所。
3.csap3基因序列分析
选取兰科植物文心兰,姬蝴蝶兰,鸽石斛等物种中的ap3类基因与墨兰中获得ap3进行同源比对,各蛋白质氨基酸序列对应的genbank登录号分别为:omads5(hm140840)、omads3(ay196350)、omads9(hm140841)、pemads2(ay378149)、pemads3(ay378150)、pemads4(ay378147)、pemads5(ay378151)、dcoap3a(dq119838)和dcoap3b(dq119839),经同源比对结果显示csap3与omads3、omads5、omads9、pemads2、pemads3、pemads4、pemads5、dcoap3a和dcoap3b的同源性分别为51%、64%、95%、62%、93%、77%、56%、62%和92%,主要为5’端保守,而3’端保守性比较低,且均具有mads-box基因保守的mads结构域和k结构域,以及ap3类基因典型paleo-ap3基序。
用软件mega将墨兰csap3基因与拟南芥、水稻、玉米、文心兰、蝴蝶兰、姬蝴蝶兰、鸽石斛中的b类mads-box基因编码氨基酸进行比对并构建进化树(图1)。结果显示,单子叶植物和双子叶植物又分别聚类成亚支,不同物种类ap3类基因编码氨基酸序列差异相对较大,分散在4个小亚支clade1/2/3/4中,csap3属于clade3亚支。
实施例2csap3在兰花中的表达模式
1.rna的提取
取墨兰栽培品种达摩幼嫩叶片、老叶、根、假鳞茎、花不同组织以及不同花朵瓣型变异品种的萼片、花瓣、唇瓣和合蕊柱四轮花器官各2g,利用植物trizol(invitrogen)试剂抽提其总rna,取其中2μl采用反转录试剂盒thermoscientificrevertaidfirststrandcdnasynthesiskit反转录成cdna。
2.定量pcr
利用引物csap3qrt-f:aaggcaagcgagctgactgt(seqidno:5)和csap3qrt-r:ccatcctctgcctgatctcc(seqidno:6),对兰花不同组织中csap3基因表达量进行实时定量pcr检测。采用ubiquitinqrt-f:caaagaaggcattccaccagat(seqidno:7)和ubiquitinqrt-r:ccgagtccccaactttgtagaa(seqidno:8)作为引物,扩增ubiquitin作为内参。按照下列程序:95℃预变性30s,然后经40个循环(95℃10s、59.5℃10s、72℃30s),72℃延伸10min。采用icycleriqreal-timepcrdetectionsystem(bio-rad,usa)进行扩增,操作按照hiscriptⅱqrtsupermixforqpcr(+gdnawiper)(vazymebiotechco.,ltd)试剂盒说明书进行。
3.表达分析
用icyclerrealtimedetectionsystemsoftware(version5.0)对pcr结果进行分析。发现csap3基因在根中表达量最低,在嫩叶和假鳞茎中的表达量相对较低,而在花中的表达量最高。进一步将国兰花发育过程从茎尖分生组织转变为花分生组织开始到完全开花分为花分生组织时期、花器官原基初期、花器官原基末期、花蕾期和开花期5个阶段,通过实时荧光定量pcr检测csap3基因在花芽不同发育阶段的表达量,发现其在花芽开始发育到完全开花的五个阶段中各阶段表达量差异在0.5-1倍间,表达量差异相对较小,不明显。
在不同瓣型变异体中的表达分析显示(图2),csap3在普通花型唇瓣中强烈优势表达,其次是花瓣和合蕊柱,在萼片中几乎不表达;在花瓣向唇瓣转化的“三星蝶”中,csap3同时在花瓣和唇瓣内强烈优势表达,整体表达量均达到萼片中两千倍以上,且唇瓣的表达量约为花瓣中表达量的1.5倍,即当三星蝶花瓣向唇瓣转化时,csap3在花瓣中表达量也随之相对增多,而在荷瓣花型,即唇瓣退化的品种“粉荷”中,csap3表现为花瓣中表达量最高,其次是唇瓣,但整体表达量较蝶瓣花显著降低;在花瓣蕊柱化的梅瓣型品种“常乐梅”唇瓣中也同样缺失表达优势,表现为花瓣中表达量较高,可见,csap3在唇瓣中优势高表达,可能与唇瓣特异分化相关。
实施例3拟南芥中csap3基因的功能分析
1.转化拟南芥高表达载体构建
利用引物csap3-f1:atggggagagggaagatagagat和csap3-r1:tcaagcgagacgcagatcat扩增出墨兰csap3基因的cds序列(即csap3基因,核苷酸序列如seqidno:1所示),并克隆至pmd19-tvector(takara),送华大基因测序。测序无误之后,使用ecori和saci双酶切(thermofisherscientific),回收纯化目的片断,并连接到经过同样两个酶的酶切,纯化的pbi121载体上命名为pbi-ap3。
2.转化拟南芥植物
2.1农杆菌gv3101的转化
1)将-80℃保存的感受态细胞于冰上化冻,待细胞完全溶解后,加入约1μg质粒(pbi-ap3),轻柔混匀,冰上放置30min。
2)液氮速冻1min后37℃,5min化冻,重复一次,加入1mllb于28℃恒温振荡培养4~6hr,200rpm。
3)菌液摇浓后,4000rpm室温下离心5min,弃上清,用100μl的lb培养液重悬农杆菌沉淀。
4)把所有混合物涂板到含50μg/ml卡那霉素和25μg/ml庆大霉素的lb板上,置于28℃恒温培养箱中培养48~72h,待长出群落后再挑菌重新划线以确保是单克隆,由此得到转化有pbi-csap3的农杆菌。
2.2花序侵染法转化拟南芥。
用od=0.3~0.5的转化有pbi-csap3的农杆菌液浸泡拟南芥花序2~3s后取出,避光密封16h后,移至生长室中继续生长。大约3~4周后,收集种子贮藏于4℃,黑暗条件。
2.3拟南芥转化子筛选
1)配制0.1%的氯化汞溶液。
2)将收集的种子放到1.5ml的eppendorf管中,加1ml的75%的酒精摇匀,在bloodtuberotator上旋转5min,用0.1%的氯化汞消毒5min。
3)离心,6000rpm,2min,弃上清液,加入1ml的灭菌水,混匀,在bloodtuberotator上旋转2min,重复3次。
4)去上清液,用1ml的无菌水悬浮种子,平铺于含50μg/ml卡那霉素的1/2ms培养基表面,封板,于4℃黑暗春化3天后移入组织培养室在(16hl/8hd)光照,23℃的条件下育苗。
5)待苗长出绿色子叶后观察统计绿色子叶幼苗数目,并去除粘附在根上的培养基,移至基质中培养。
6)经60d左右的生活周期,得到转基因拟南芥第一代种子t1,收获第二代纯合种子t2(转基因植株)用于后续试验。
3.转基因拟南芥csap3表达量分析
为了确定csap3基因在拟南芥中的生物学功能,分别以拟南芥野生型和上述步骤2所得转化子(t2纯合种子生长后的转基因植株)cdna为模板,采用引物csap3qrt-f:aaggcaagcgagctgactgt和csap3qrt-r:ccatcctctgcctgatctcc检测转基因拟南芥中csap3基因的表达量,按照下列程序:95℃预变性30s,然后经40个循环(95℃10s、59.5℃10s、72℃30s),72℃延伸10min。采用icycleriqreal-timepcrdetectionsystem(bio-rad,usa)进行扩增,操作按照hiscriptⅱqrtsupermixforqpcr(+gdnawiper)(vazymebiotechco.,ltd)试剂盒说明书进行。结果发现在获得的转基因植株中csap3的表达量均提高,选取其中三个株系用于后续表型分析(图3)。
4.转基因拟南芥表型分析
收获的t3代纯合转基因植株种子经表面灭菌后铺在含50μg/ml卡那霉素的ms培养基上,4℃黑暗培养2天,转移到(16hl/8hd)光照,23℃条件下培养。在拟南芥开花期,发现转基因植株花器官形态结构异常,表现为每朵花有一个花瓣卷曲,且野生型柱头笔直向上生长,而转基因植株的花内部柱头弯曲生长,并随着柱头伸长弯曲越来越明显,最后形成的果荚弯曲程度各不一样,将果荚进行剖切,发现果荚两心室发育程度不一样,果荚弯曲外侧发育较完整饱满,而弯曲内侧则发育数量少并且大小也相对较小(图4)。
实施例4兰花中csap3基因的功能分析
1.高表达载体构建
利用引物csap3-f1:atggggagagggaagatagagat和csap3-r1:tcaagcgagacgcagatcat扩增出墨兰csap3的cds序列,并克隆至pmd19-tvector(takara),测序无误之后。使用ecori和saci双酶切,回收纯化目的片断,并连接到经过同样两个酶的酶切,纯化的pcam1301载体上,得到pcam1301-csap3。
2.转化兰花叶片
2.1农杆菌gv3101的转化
1)将-80℃保存的感受态细胞于冰上化冻,待细胞完全溶解后,加入约1μg质粒(pcam1301-csap3),轻柔混匀,冰上放置30min。
2)液氮速冻1min后37℃,5min化冻,重复一次,加入1mllb于28℃恒温振荡培养4~6h,200rpm。
3)菌液摇浓后,4000rpm室温下离心5min,弃上清,用100μl的lb培养液重悬农杆菌沉淀。
4)把所有混合物涂板到含50μg/ml卡那霉素和32μg/ml庆大霉素的lb板上,置于28℃恒温培养箱中培养48~72h,待长出群落后再挑菌重新划线以确保是单克隆,得到含有pcam1301-csap3质粒的农杆菌。
2.2农杆菌直接注射法转化国兰花芽
用od=0.3~0.5,含有pcam1301-csap3质粒的农杆菌液直接注射国兰品种“银针”幼嫩叶芽和花芽,以pcam1301空载体作为对照。每周注射一次,连续三周,置于培养室中继续生长。大约8~12周后,收集接种后花器官进行基因表达量检测。
3.接种农杆菌后兰花csap3基因表达量分析
为了确定csap3基因在国兰中的生物学功能,分别以国兰品种“银针”未接种的野生型、空载体和接种后叶片的cdna为模板,采用引物csap3qrt-f:aaggcaagcgagctgactgt和csap3qrt-r:ccatcctctgcctgatctcc,检测注射农杆菌后兰花中csap3基因的表达量,按照下列程序:95℃预变性30s,然后经40个循环(95℃10s、57℃10s、72℃26s)。以同样的cdna作为模板,使用ubiquitinqrt-f:caaagaaggcattccaccagat和ubiquitinqrt-r:ccgagtccccaactttgtagaa,扩增ubiquitin作为内参,按照下列程序:95℃预变性30s,然后经40个循环(95℃10s、59.5℃10s、72℃30s),72℃延伸10min。采用icycleriqreal-timepcrdetectionsystem(bio-rad,usa)进行扩增,操作按照hiscriptⅱqrtsupermixforqpcr(+gdnawiper)(vazymebiotechco.,ltd)试剂盒说明书进行。结果表明在含有pcam1301-csap3质粒的农杆菌液注射植株中csap3基因的表达量均提高。
4.高表达csap3兰花表型分析
接种含有pcam1301-csap3质粒的农杆菌液注射植株的花芽生长3周后,花朵开放,跟未接种的野生型或空载体相比,含有pcam1301-csap3质粒的农杆菌液注射植株花瓣向唇瓣转变,唇瓣性状加强,产生更多的唇瓣结构(图5)。
以上仅是本发明的优选实施方式,应当指出的是,上述优选实施方式不应视为对本发明的限制,本发明的保护范围应当以权利要求所限定的范围为准。对于本技术领域的普通技术人员来说,在不脱离本发明的精神和范围内,还可以做出若干改进和润饰,这些改进和润饰也应视为本发明的保护范围。
序列表
<110>广东省农业科学院环境园艺研究所
<120>国兰花器官发育调控基因及其编码蛋白与应用
<160>8
<170>siposequencelisting1.0
<210>1
<211>669
<212>dna
<213>墨兰达摩(cymbidiumsinense‘dharma’)
<400>1
atggggagagggaagatagagataaagaagatagagaaccctactaacaggcaagtcact60
tactctaagaggagagctgggatcatgaagaaggcaagcgagctgactgttctctgcgac120
gctcagctctctcttgtaacgttctcgagcactggcaagttctctgagtattgtagtcca180
accactgacaccaagagcatatatgatcgttaccagcaggtgtccggcataaatctatgg240
agctcgcagtacgagaagatgcagaatacgttgaatcatttaaaggagataaaccaaacc300
ctgagaagggagatcaggcagaggatgggggaggaccttgatgggctggaaatcaaggaa360
ctgcgtggtcttgagcaaaatatggatgagtccctgaagcttgtaaggaatcggaagtat420
catgtcatcagtacccagacagatacctacaaaaagaagctgaagaactctcaagaaacc480
cacaggaacttactgcgggagctggaagcggagcacgcagtctattatgtggatgatgat540
ccgaacagctatgatggtgcacttgcactaggaaatgggccttcctacctgtactcattc600
cgtagccaaccaagccagccaaaccttcaaggaatgggatatggccctcatgatctgcgt660
ctcgcttga669
<210>2
<211>222
<212>prt
<213>墨兰达摩(cymbidiumsinense‘dharma’)
<400>2
metglyargglylysilegluilelyslysilegluasnprothrasn
151015
argglnvalthrtyrserlysargargalaglyilemetlyslysala
202530
sergluleuthrvalleucysaspalaglnleuserleuvalthrphe
354045
serserthrglylyspheserglutyrcysserprothrthraspthr
505560
lysseriletyraspargtyrglnglnvalserglyileasnleutrp
65707580
serserglntyrglulysmetglnasnthrleuasnhisleulysglu
859095
ileasnglnthrleuargarggluileargglnargmetglygluasp
100105110
leuaspglyleugluilelysgluleuargglyleugluglnasnmet
115120125
aspgluserleulysleuvalargasnarglystyrhisvalileser
130135140
thrglnthraspthrtyrlyslyslysleulysasnserglngluthr
145150155160
hisargasnleuleuarggluleuglualagluhisalavaltyrtyr
165170175
valaspaspaspproasnsertyraspglyalaleualaleuglyasn
180185190
glyprosertyrleutyrserpheargserglnproserglnproasn
195200205
leuglnglymetglytyrglyprohisaspleuargleuala
210215220
<210>3
<211>23
<212>dna
<213>人工序列csap3-f1(artificialsequencecsap3-f1)
<400>3
atggggagagggaagatagagat23
<210>4
<211>20
<212>dna
<213>人工序列csap3-r1(artificialsequencecsap3-r1)
<400>4
tcaagcgagacgcagatcat20
<210>5
<211>20
<212>dna
<213>人工序列csap3qrt-f(artificialsequencecsap3qrt-f)
<400>5
aaggcaagcgagctgactgt20
<210>6
<211>20
<212>dna
<213>人工序列csap3qrt-r(artificialsequencecsap3qrt-r)
<400>6
ccatcctctgcctgatctcc20
<210>7
<211>22
<212>dna
<213>人工序列ubiquitinactinqrt-f(artificialsequenceubiquitinactinqrt-f)
<400>7
caaagaaggcattccaccagat22
<210>8
<211>22
<212>dna
<213>人工序列ubiquitinactinqrt-r(artificialsequenceubiquitinactinqrt-r)
<400>8
ccgagtccccaactttgtagaa22
- 还没有人留言评论。精彩留言会获得点赞!