用于催化木质纤维素糖化的全菌酶制剂的制作方法
本发明属于生物技术领域,涉及一种酶制剂,具体涉及一种用于催化木质纤维素糖化的全菌酶制剂。
背景技术
木质纤维素是一类供应丰富且环境友好的可再生物质,是唯一可大规模再生且全面替代化石能源的资源,大力发展木质纤维素原料在能源等领域的利用,是加快发展循环经济,保障国家能源安全和碳减排的一项重要战略任务。然而,木质纤维素转化的最大瓶颈莫过于纤维素结晶区难降解、导致酶解效率低,成本高。因此,要实现木质纤维素的高效利用,就必须首先实现该不溶性复杂底物的高效水解糖化。目前,木质纤维素糖化主要是通过依赖于真菌游离酶纤维素降解酶系的工艺来实现。这些工艺中酶的成本占木质纤维素生物转化过程总成本的50%左右,且经过几十年的努力仍不能有效降低其用酶成本。此外,木质纤维素的水解效率也是重要的限制因素。因此,需要转换思路,开发不同于游离纤维素酶系的新型酶制剂实现木质纤维素的高效利用。
已知纤维小体是热纤梭菌等厌氧细菌生产的一种具有复杂结构和组分的多酶复合体,是自然界中已知的最高效的纤维素降解体系之一。纤维小体包括脚架蛋白等非催化单元以及具有不同催化活性的酶单元,并通过多级脚架蛋白和不同纤维小体酶类具有的多类型组装模块间特异性的非共价相互作用,将不同的功能组分组装成为分子量超过兆道尔顿的超分子多酶复合体。纤维小体组分和结构还具有时空调控特性,以适应木质纤维素的复杂成分,从而保证了其高效降解活力。此外,纤维小体还通过脚架蛋白或纤维素酶上所具有的纤维素结合模块和挂壁模块与底物及细胞组成三元复合体,纤维小体和细胞间的协同作用可以进一步提高了木质纤维素的糖化效率(专利文件ep2013355)。
尽管纤维小体及其生产菌株在木质纤维素糖化应用中具有巨大潜力,但其工业化应用仍受到多种因素的限制。例如,纤维二糖对纤维小体酶有严重的反馈抑制作用。针对这一问题,前人主要通过向水解体系里额外添加β-1,4-葡萄糖苷酶等非纤维小体蛋白来提高纤维小体的活力和对底物的适应力。然而,由于外源添加的蛋白的活性及稳定性会随着糖化过程的进行而降低,因此,酶的消耗需要提高酶添加量或添加次数。此外,这些非纤维小体蛋白作为游离酶添加到糖化体系中,不能与纤维小体相互作用,会显著降低与体系中其他酶的协同作用,不可避免造成大于需求量的酶的额外添加,导致酶制剂成本的居高不下。不仅如此,这种依赖于外源添加游离酶的糖化策略,工艺复杂,对设备要求高,转化效率还不能满足工业化生产要求。
研究人员此前的工作发现,通过在热纤梭菌中表达β-1,4葡萄糖苷酶可以提高热纤梭菌降解微晶纤维素的能力(zhang,j.,s.liu,r.li,w.hong,y.xiao,y.feng,q.cuiandy.-j.liu(2017)."efficientwhole-cell-catalyzingcellulosesaccharificationusingengineeredclostridiumthermocellum."biotechnolbiofuels10(1):124.),这一工作解决了纤维二糖对纤维小体酶有严重的反馈抑制作用的问题,同时解决了将非纤维小体蛋白作为游离酶添加到糖化体系中存在的成本高等问题。但是,该工作是以微晶纤维素这一纯纤维素为底物,所提供的酶制剂以及添加β-1,4葡萄糖苷酶的方法并不能解决木质纤维素糖化中遇到的其他问题。例如,由于木质纤维素的成分复杂(包括纤维素、半纤维素、果胶、淀粉、蛋白质、木质素等)非纤维素成分会与纤维素成分交联在一起,这一复杂结构会降低纤维素酶与纤维素底物的可及性,从而降低糖化效率。另外,纤维素酶会非特异性的吸附在木质纤维素的复杂成分中,从而大大降低糖化效率。此外,向纤维小体中加入其他酶仍然存在能否在细胞内表达,能否分泌到胞外,能否组装到纤维小体内,能否与纤维小体组分产生协同作用从而促进糖化等问题。
技术实现要素:
针对现有技术中纤维小体在木质纤维素糖化中所存在的问题,本发明提供了用于催化木质纤维素糖化的全菌酶制剂。所述全菌酶制剂解决了糖化过程中酶的消耗而导致成本高、工艺繁琐等问题。
本发明的技术方案:
用于催化木质纤维素糖化的全菌酶制剂,所述全菌酶制剂是通过非纤维小体蛋白与纤维小体中的组分相互作用,将非纤维小体蛋白结合在纤维小体复合体中,获得的纤维小体改造后的菌株;然后采用该菌株来糖化,即糖化过程有细菌细胞的参与。
所述非纤维小体蛋白为木聚糖酶、纤维素内切酶、纤维素外切酶、膨胀因子、蛋白酶、淀粉酶或果胶酶。所述纤维小体为由厌氧细菌生产且分泌在胞外的具有木质纤维素降解活性的多酶复合体。所述的纤维小体中的组分为脚架蛋白、有催化功能的酶、组装模块。
所述非纤维小体蛋白与纤维小体中的蛋白组分相互作用的方式为间接性连接或者直接性连接。所述间接性连接为:非纤维小体蛋白与纤维小体中的蛋白组分通过共价相互作用进行连接;所述直接性连接为:非纤维小体蛋白与纤维小体中的组分通过串联表达进行连接。
优选的是,所述非纤维小体蛋白具有序列表seqidno:1-6,18-21所示的氨基酸序列;以及与如seqidno:1-6,18-21所示的氨基酸序列的一致性在95%以上,且与如seqidno:1-6,18-21所示的氨基酸序列具有相同功能的氨基酸序列。其中:seqidno:18:genbank序列号crz35393.1;seqidno:19:由基因组cp001393.1中1968724至1973904核酸序列编码;seqidno:20:genbank序列号为kc763474.1;seqidno:21:由基因组cp001393.1中2531445至2532785核酸序列编码。
优选的是,所述厌氧细菌为热纤梭菌(ruminiclostridiumthermocellum或clostridiumthermocellum),黄色溶纤梭菌(clostridiumclariflavum),嗜纤维梭菌(clostridiumcellulovorans),解纤维梭菌(clostridiumcellulolyticum),解纤维醋弧菌(acetivibriocellulolyticus),溶纤维假拟杆菌(pseudobacteroidescellulosolvens),白色瘤胃球菌(ruminococcusalbus),黄化瘤胃球菌(ruminococcusflavefaciens)。其中,所述热纤梭菌为表达外泌的β1,4-葡萄糖苷酶的热纤梭菌。
其中,所述非纤维小体蛋白与纤维小体中的组分通过共价相互作用进行连接,具体为:非纤维小体蛋白和纤维小体组分分别与具有共价相互作用的多肽片段连接,利用多肽片段间特异性共价相互作用实现纤维小体组分与非纤维小体蛋白的共价交联,利用纤维小体组分所带的组装模块,使非纤维小体蛋白结合在纤维小体复合体中。所述与非纤维小体蛋白和纤维小体组分共价相互作用的多肽片段为seqidno:7中所示的碱基序列或seqidno:8中所示的氨基酸序列。具体实现步骤包括:
1)通过基因克隆的方法,连接非纤维小体蛋白与共价相互作用的多肽片段对中的一个片段(多肽片段i或ii,即seqidno:7或8)的编码基因;
2)根据1),通过基因克隆的方法,连接纤维小体组分蛋白与共价相互作用的多肽片段对中的另一个片段(多肽片段ii或i,即seqidno:8或7)的编码基因。
3)根据1),将非纤维小体蛋白与多肽片段的重组基因序列连接到产纤维小体细菌表达质粒(图1);
4)根据2),将纤维小体组分蛋白与多肽片段的重组基因序列连接到产纤维小体细菌同源重组质粒(图2),并根据基因组序列设计同源臂。
5)将4)中获得的质粒转化到产纤维小体细菌胞内,并通过同源重组筛选(图3)实现重组基因序列对基因组上原始纤维小体组分蛋白序列的替换,从而构建重组菌株,实现纤维小体组分蛋白与多肽片段在产纤维小体细菌胞内的融合表达。
6)将3)中获得的质粒转化到5)中获得的重组菌株中,实现非纤维小体蛋白与多肽片段在产纤维小体细菌胞内的融合表达。
最终获得的重组菌株中,基因组表达的纤维小体组分蛋白与质粒表达的非纤维小体蛋白通过所融合的多肽片段i及ii间的特异性共价相互作用实现共价交联,并组装在纤维小体复合体中。
其中,所述非纤维小体蛋白与纤维小体中的组分通过串联表达进行连接,具体为:非纤维小体蛋白与纤维小体的组装模块或具有组装模块的蛋白组分进行融合表达,通过组装模块间的特异性的非共价相互作用,实现将非纤维小体蛋白组装在纤维小体复合体中。所述融合表达为:非纤维小体蛋白的编码基因插入到编码纤维小体组分蛋白的序列的n端、c端或结构域序列中间的基因组上。
所述各自呼应的模块为粘连模块与对接模块。所述对接模块为seqidno:9、seqidno:12-16、seqidno:17中所示的碱基序列,粘连模块为seqidno:10中所示的氨基酸序列。具体实现步骤包括:
1)通过基因克隆的方法,连接非纤维小体蛋白与i型对接模块(seqidno:9)的编码基因,或
2)通过基因克隆的方法,连接非纤维小体蛋白与ii型粘连模块(seqidno:11)的编码基因
3)将1)或2)获得非纤维小体蛋白与组装模块的重组基因序列连接到产纤维小体细菌表达质粒(图1)
4)将3)中获得的质粒转化到产纤维小体细菌胞内,实现非纤维小体蛋白与i型对接模块或ii型粘连模块在产纤维小体细菌胞内的融合表达。
最终获得的重组菌株中,质粒表达的非纤维小体蛋白通过所融合的i型对接模块或ii型粘连模块实现与纤维小体脚架蛋白进行特异性非共价相互作用,从而组装在纤维小体复合体中。
所述直接融合表达的具体实现步骤包括:
1)通过基因克隆的方法,将非纤维小体蛋白的编码基因连接到产纤维小体细菌同源重组质粒(图2)中,并根据基因组序列设计同源臂。
2)将1)中获得的质粒转化到产纤维小体细菌胞内,并通过同源重组筛选(图3)实现非纤维小体蛋白的编码基因插入到基因组上纤维小体组分蛋白的序列的n端或c端或结构域序列中间,从而构建重组菌株,实现非纤维小体蛋白与纤维小体组分蛋白在产纤维小体细菌胞内的融合表达。
最终获得的重组菌株中,非纤维小体蛋白利用与其融合表达的纤维小体组分蛋白的组装模块结合到纤维小体复合体中。
本发明的有益效果:
(1)本发明提供了全菌酶制剂,所述全菌酶制剂通过非纤维小体蛋白(木聚糖酶、纤维素内切酶、纤维素外切酶、膨胀因子、蛋白酶、淀粉酶或果胶酶)与纤维小体中的组分相互作用,将非纤维小体蛋白结合在纤维小体复合体中,获得纤维小体改造后的菌株,然后采用该菌株来糖化,即糖化过程有细菌细胞的参与。
(2)与现有技术中采用的不能与纤维小体相互作用的游离酶相比,本发明所述的全菌酶制剂,不但实现了其稳定性和活性的维持,而且大大提升了其与体系中其他酶的协同作用,从而降低了酶的消耗量,降低了生产成本和工艺的繁琐性。
附图说明
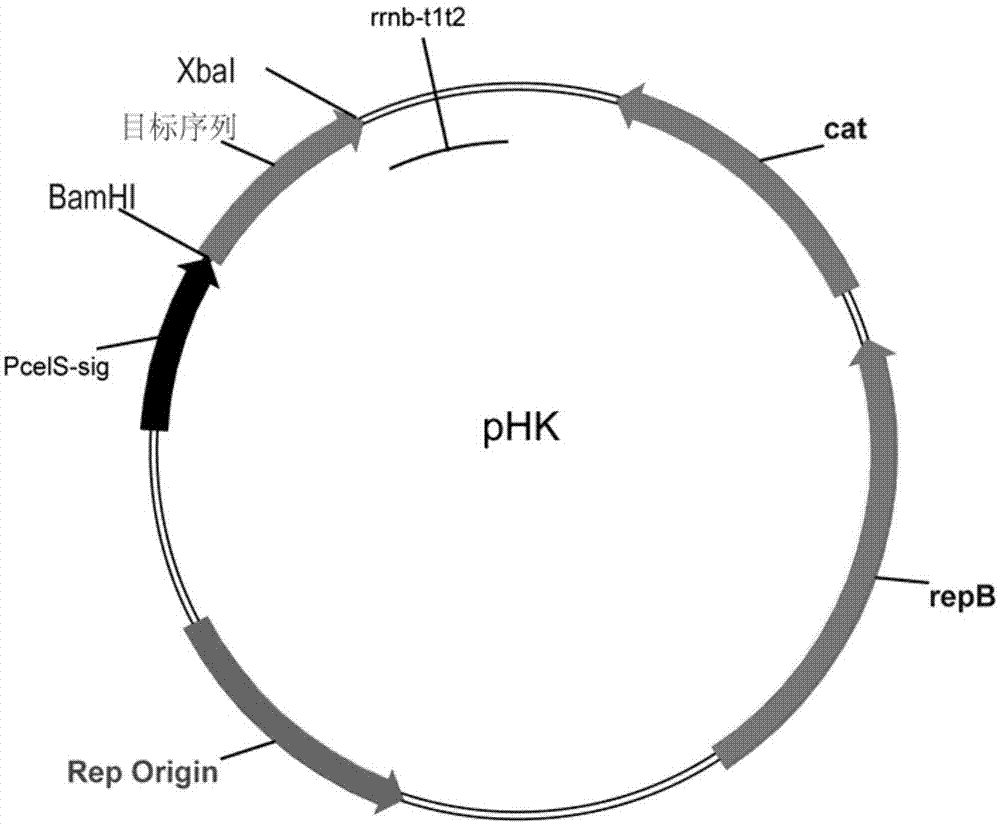
图1为本发明实施例提到的产纤维小体细菌表达质粒phk示意图。
图2为本发明实施例提到的产纤维小体细菌同源重组质粒phk-hr示意图。
图3为本发明实施例提到的产纤维小体细菌中进行同源重组筛选的过程示意图。
具体实施方式
下面结合实施例对本发明做进一步的说明。
实施例1:构建表达外泌的β-1,4葡萄糖苷酶的热纤梭菌
选择热纤梭菌纤维小体中的纤维素酶cel9k(外切纤维素酶,由基因组cp002416.1中2113813至2111293核酸序列编码)的酶催化结构域和对接模块之间作为靶向敲入位点。首先,将β-1,4-葡萄糖苷酶bgla(genbank序列号为afo70070.1)编码基因作为目标序列,利用mlui和eagi的酶切位点克隆到同源重组质粒phk-hr(图2)中,构建同源重组质粒phk-hr-bgla。上游同源臂hr-up序列为热纤梭菌dsm1313基因组(ncbi数据库中序列号为cp002416.1)中2111347到2112870核酸序列,下游同源臂hr-down为dsm1313基因组中2109848到2111354核酸序列,中间同源臂hr-short为dsm1313基因组中2111347到2111659核酸序列。其次,将构建好的质粒转化到δpyrf中,并按照三步筛选(图3)获得同源重组菌株1。具体步骤为:
1)将同源重组质粒phk-hr(图2)转化进入pyrf缺失的菌株中,利用含甲砜氯霉素的gs-2半固体培养基(kh2po41.5g/l,k2hpo4·3h2o3.8g/l,尿素2.1g/l,mgcl2·6h2o1.0g/l,cacl2·2h2o150mg/l,feso4·6h2o1.25mg/l,半胱氨酸盐酸1.0g/l,mops钠盐10g/l,酵母提取物6.0g/l,纤维二糖5.0g/l,二水柠檬酸三钠3.0g/l,刃天青0.1mg/l,ph7.4)平板进行筛选,以获得质粒转化子。
2)获得的转化子在mj液体培养基(kh2po41.5g/l,k2hpo4·3h2o3.8g/l,尿素2.1g/l,mgcl2·6h2o1.0g/l,cacl2·2h2o150mg/l,feso4·6h2o1.25mg/l,半胱氨酸盐酸1.0g/l,mops钠盐10g/l,纤维二糖5.0g/l,二水柠檬酸三钠3.0g/l,刃天青0.1mg/l,盐酸吡哆胺2mg/l,生物素0.2mg/l,对氨基苯甲酸0.4mg/l,维生素b120.2mg/l,ph7.4)中转接三代后,涂布含10μg/ml5-氟脱氧尿苷(fudr)的mj半固体培养基进行第一次同源重组筛选。在这一步骤中,由于tdk可以将fudr转化为对细胞有毒的f-dump,同时底盘细胞在mj培养基中必须依靠质粒上的pyrf基因合成尿嘧啶核苷酸才能生存,因此,这一筛选策略保证了质粒上的同源重组模块与基因组发生同源重组的同时,重组后的质粒也发生丢失。根据长同源臂优先发生同源重组的原理,前后两个长同源臂首先与基因组发生同源重组,此时获得的重组子由出发菌株的尿嘧啶营养缺陷型回复成原养型。
3)经第一次同源重组后获得的重组子先在gs-2液体培养基中传代3次,并用相同的培养基对菌液进行梯度稀释后涂布含有500μg/ml5-氟乳清酸(foa)的gs-2半固体培养基进行筛选,获得目的无疤基因敲除/敲入菌株。在这一步骤中,pyrf的反向筛选作用会促使上游长同源臂和短同源臂发生第二次同源重组将pyrf表达框从基因组中去除。第二次同源重组后的突变株又从原养型突变成尿嘧啶营养缺陷型。从而实现基因组上目标位点基因的敲除、敲入或者替换。
通过提取重组菌株1的纤维小体发现,表达的β-1,4-葡萄糖苷酶与cel9k的融合蛋白可以外泌到胞外,并组装到纤维小体复合体中。
实施例2:通过直接性连接的方法,构建基于热纤梭菌纤维小体的全菌酶制剂
利用重叠延伸聚合酶链式反应的方法,将木聚糖酶xync(seqidno:1)与热纤梭菌的ii型粘连模块cohiict的序列(seqidno:10)或i型对接模块docict的序列(seqidno:9)直接连接起来,其中cohiict或docict的序列连接至xync序列的3’端,从而获得xync-cohiict和xync-docict序列。再利用bamhi和xbai酶切位点,将连接起来的重组序列作为目标序列克隆到表达质粒phk(图1)上。由于phk上带有来源于热纤梭菌的纤维素酶cel48s的启动子及信号肽序列(seqidno:11),表达的目标基因可以外泌到细胞胞外。将构建好的质粒转化到实施例1构建的同源重组菌株1中,从而获得表达具有ii型粘连模块或i型对接模块的木聚糖酶的热纤梭菌重组菌株2或3。通过提取重组菌株的纤维小体发现,表达的具有docict和cohiict的xync都可以外泌到胞外,并组装到纤维小体复合体中。因此该重组菌株可以作为具有非纤维小体木聚糖酶的全菌酶制剂。
实施例3:通过直接性连接的方法,构建基于热纤梭菌纤维小体的全菌酶制剂
将木聚糖酶xynb(seqidno:2)编码基因作为目标序列,选择热纤梭菌纤维小体的纤维素外切酶cel48s(由基因组cp002416.1中3228088至3230229核酸序列编码)的5’端作为靶向敲入位点,利用mlui和eagi的酶切位点克隆到同源重组质粒phk-hr(图2)中,构建同源重组质粒phk-hr-xynb。上游同源臂hr-up为热纤梭菌dsm1313基因组(ncbi数据库中序列号为cp002416.1)中3230200到3230700核酸序列,下游同源臂hr-down为dsm1313基因组中3229699到3230199核酸序列,中间同源臂hr-short为dsm1313基因组中3230200到3230500核酸序列。将构建好的质粒再分别转化到实施例1构建的重组菌株1中,并按照实施例1及图3所示的三步筛选方法获得同源重组菌株4。
通过提取重组菌株的纤维小体发现,xynb与cel48s的融合蛋白可以外泌到胞外,并组装到纤维小体复合体中。因此该重组菌株可以作为具有非纤维小体木聚糖酶的全菌酶制剂。
实施例4:通过间接性连接的方法,构建基于热纤梭菌纤维小体的全菌酶制剂
选择热纤梭菌纤维小体的纤维素外切酶cel48s的5’端作为靶向敲入位点,将多肽片段i(seqidno:8)或多肽片段ii(seqidno:9)两个片段的编码序列作为目标序列,利用mlui和eagi的酶切位点分别克隆到同源重组质粒phk-hr(图2)中,分别构建获得同源重组质粒phk-hr-i或phk-hr-ii。同源臂核酸序列、转化及筛选方法与实施例1相同,分别获得同源重组菌株δpyrf::bgla-i或δpyrf::bgla-ii,分别表达cel48s与多肽片段i(seqidno:8)或多肽片段ii的融合蛋白。
利用重叠延伸聚合酶链式反应的方法,将多肽片段ii或多肽片段i连接到木聚糖酶xyna(seqidno:3)的3’端。利用bamhi和xbai酶切位点,将连接起来的重组序列作为目标序列克隆到表达质粒phk(图1)上。将含有xyna与多肽片段i的重组序列的表达质粒转化到δpyrf::bgla-ii,得到重组菌株5;将含有xyna与多肽片段ii的重组序列的表达质粒转化到δpyrf::bgla-i,,得到重组菌株6,从而实现cel48s和xyna能够通过多肽片段i和ii间特异性共价相互作用进行结合。通过提取获得热纤梭菌重组菌株的纤维小体发现,表达的具有共价结合模块的xyna可以外泌到胞外,并与具有共价结合模块的cel48s相互作用,组装到纤维小体复合体中。因此该重组菌株可以作为具有非纤维小体木聚糖酶的全菌酶制剂。
实施例5:通过间接性连接的方法,构建基于热纤梭菌纤维小体的全菌酶制剂
与实施例4不同的是,将多肽片段ii或多肽片段i连接到纤维素内切酶celz(seqidno:18)的3’端,得到重组菌株7。
实施例6:通过直接性连接的方法,构建基于热纤梭菌纤维小体的全菌酶制剂
利用重叠延伸聚合酶链式反应的方法,将纤维素外切酶cel9-48(seqidno:19)与热纤梭菌的ii型粘连模块cohiict的序列(seqidno:10)或i型对接模块docict的序列(seqidno:9)直接连接起来,其中cohiict或docict的序列连接至cel9-48序列的3’端,获得cel9-48-docict或cel9-48-cohiict序列。
以cel9-48-docict或cel9-48-cohiict序列为目标序列,利用mlui和eagi的酶切位点克隆到图2所示同源重组质粒phk-hr中,以乳酸脱氢酶基因clo1313_1160为靶向替换序列,构建同源重组质粒phk-hr-cel9-48。上游同源臂hr-up为热纤梭菌dsm1313基因组(ncbi数据库中序列号为cp002416.1)中1380180到1380679核酸序列,下游同源臂hr-down为dsm1313基因组中1380634到1381133核酸序列,中间同源臂hr-short为dsm1313基因组中1380833到1381133核酸序列。转化及筛选方法与实施例3相同获得获得表达具有i型对接模块或ii型粘连模块的纤维素内切酶的热纤梭菌重组菌株8或9。通过提取重组菌株的纤维小体发现,cel9-48与组装模块的融合蛋白可以外泌到胞外,并通过非共价交联的相互作用方式组装到纤维小体复合体中。因此该重组菌株可以作为具有非纤维小体木聚糖酶的全菌酶制剂。
实施例7:通过直接性连接的方法,构建基于黄色溶纤梭菌纤维小体的全菌酶制剂
与实施例2不同的是,将纤维素外切酶cel48s与黄色溶纤梭菌的i型对接模块序列dociccl(seqidno:12)直接连接起来,从而获得cel48s-dociccl序列。构建好的质粒转化到黄色溶纤梭菌(clostridiumclariflavumdsm19732)中进行表达。通过提取重组菌株的纤维小体发现,表达的具有dociccl的cel48s可以外泌到胞外,并组装到黄色溶纤梭菌纤维小体复合体中。
实施例8:通过直接性连接的方法,构建基于白色瘤胃球菌纤维小体的全菌酶制剂
与实施例7不同的是,将纤维素外切酶cel48s与白色瘤胃球菌的i型对接模块序列docira(seqidno:13)直接连接起来,从而获得cel48s-docira序列。构建好的质粒转化到白色瘤胃球菌(ruminococcusalbussy3)中进行表达。通过提取重组菌株的纤维小体发现,表达的具有docira的cel48s可以外泌到胞外,并组装到白色瘤胃球菌纤维小体复合体中。
实施例9:通过直接性连接的方法,构建基于黄化瘤胃球菌纤维小体的全菌酶制剂
与实施例7不同的是,将纤维素外切酶cel48s与黄化瘤胃球菌的i型对接模块序列docirf(seqidno:14)直接连接起来,从而获得cel48s-docirf序列。构建好的质粒转化到黄化瘤胃球菌(ruminococcusflavefaciens)中进行表达。通过提取重组菌株的纤维小体发现,表达的具有docirf的cel48s可以外泌到胞外,并组装到黄化瘤胃球菌纤维小体复合体中。
实施例10:通过直接性连接的方法,构建基于解纤维梭菌纤维小体的全菌酶制剂
与实施例7不同的是,将纤维素外切酶cel48s与解纤维梭菌的i型对接模块序列docicce(seqidno:15)直接连接起来,从而获得cel48s-docicce序列。构建好的质粒转化到解纤维梭菌(clostridiumcellulolyticumh10)中。通过提取重组菌株的纤维小体发现,表达的具有docicce的cel48s可以外泌到胞外,并组装到解纤维梭菌(纤维小体复合体中。
实施例11:通过直接性连接的方法,构建基于溶纤维假拟杆菌纤维小体的全菌酶制剂
与实施例7不同的是,将纤维素外切酶cel48s与溶纤维假拟杆菌的i型对接模块序列docipc(seqidno:16)直接连接起来,从而获得cel48s-docipc序列。构建好的质粒转化到溶纤维假拟杆菌(pseudobacteroidescellulosolvensdsm2933)中。通过提取重组菌株的纤维小体发现,表达的具有docipc的cel48s可以外泌到胞外,并组装到溶纤维假拟杆菌纤维小体复合体中。
实施例12:通过直接性连接的方法,构建基于溶纤维假拟杆菌纤维小体的全菌酶制剂
与实施例11不同的是,将木聚糖酶xync(seqidno:1)与溶纤维假拟杆菌的i型对接模块序列docipc(seqidno:16)直接连接起来,从而获得xync-docipc序列构建表达质粒,并转化溶纤维假拟杆菌(pseudobacteroidescellulosolvensdsm2933)。通过提取重组菌株的纤维小体发现,表达的具有docipc的xync可以外泌到胞外,并组装到溶纤维假拟杆菌纤维小体复合体中。
实施例13:通过直接性连接的方法,构建基于热纤梭菌纤维小体的的全菌酶制剂
利用重叠延伸聚合酶链式反应的方法,将木聚糖酶xyna(seqidno:3)与ii型粘连模块cohii的序列(seqidno:10)直接连接起来,其中cohii的序列连接至xyna序列的3’端,从而获得xyna-cohii的序列。以xyna-cohii序列为目标序列,利用mlui和eagi的酶切位点克隆到图2所示同源重组质粒phk-hr中,以乳酸脱氢酶基因clo1313_1878为靶向替换序列,构建同源重组质粒phk-hr-xyna。上游同源臂hr-up为热纤梭菌dsm1313基因组(ncbi数据库中序列号为cp002416.1)中2194853到2195353核酸序列,下游同源臂hr-down为dsm1313基因组中2196312到2196811核酸序列,中间同源臂hr-short为dsm1313基因组中2195053到2195353核酸序列。将构建好的质粒再转化到实施例6构建的重组菌株8中,并按照实施例1所述筛选方法获得同源重组菌株10。通过提取重组菌株的纤维小体发现,xyna与cel9-48都可以外泌到胞外,并通过各自携带的组装模块以非共价相互作用方式组装到纤维小体复合体中。因此该重组菌株可以作为具有非纤维小体木聚糖酶的全菌酶制剂。
实施例14:通过直接性连接的方法,构建基于热纤梭菌纤维小体的全菌酶制剂
将膨胀因子epn(seqidno:4或5)编码基因作为目标序列,选择热纤梭菌纤维小体的脚架蛋白sdba(由基因组cp002416.1中1108113至1109912核酸序列编码)的5’端作为靶向敲入位点构建同源重组质粒。上游同源臂hr-up为热纤梭菌dsm1313基因组(ncbi数据库中序列号为cp002416.1)中1107610到1108109核酸序列,下游同源臂hr-down为dsm1313基因组中1109916到1110415核酸序列,中间同源臂hr-short为dsm1313基因组中1107809到1108109核酸序列。同实施例1筛选方法,获得同源重组菌株11。通过提取重组菌株的纤维小体发现,epn与sdba的融合蛋白可以外泌到胞外,并组装到纤维小体复合体中。因此该重组菌株可以作为具有非纤维小体木聚糖酶的全菌酶制剂。
实施例15:通过直接性连接的方法,构建基于嗜纤维梭菌纤维小体的全菌酶制剂
与实施例3不同的是,将蛋白酶prol(seqidno:6)编码基因作为目标序列,选择嗜纤维梭菌纤维小体的纤维素酶clocel_2823(由基因组cp002160.1中3464080至3466140核酸序列编码)的5’端作为靶向敲入位点。同源臂hr-up、hr-down和hr-short分别为clostridiumcellulovorans743b基因组中(ncbi数据库中序列号为cp002160.1)中3466141到3466640、3465641到3466140以及3466141到3466441核酸序列。将构建好的质粒再分别转化到pyrf缺失的743b突变株中,筛选获得蛋白酶prol与纤维素酶clocel_2823融合表达的同源重组菌。通过提取重组菌株的纤维小体发现,该融合蛋白可以外泌到胞外,并组装到纤维小体复合体中。
实施例16:通过直接性连接的方法,构建基于解纤维梭菌纤维小体的全菌酶制剂
与实施例3不同的是,将淀粉酶amya(seqidno:20)编码基因作为目标序列,选择解纤维梭菌纤维小体的纤维素酶ccel_0729(由基因组cp001348.1中843122至845197核酸序列编码)的5’端作为靶向敲入位点。同源臂hr-up、hr-down和hr-short分别为clostridiumcellulolyticumh10基因组中(ncbi数据库中序列号为nc_011898)中842441到842941、842942到843441以及842641到842941核酸序列。将构建好的质粒再分别转化到pyrf缺失的h10突变株中,筛选获得淀粉酶amya与纤维素酶ccel_0729融合表达的同源重组菌。通过提取重组菌株的纤维小体发现,该融合蛋白可以外泌到胞外,并组装到纤维小体复合体中。
实施例17:通过直接性连接的方法,构建基于解纤维醋弧菌纤维小体的全菌酶制剂
与实施例7不同的是,将果胶酶pela(seqidno:21)编码基因与解纤维醋弧菌的i型对接模块序列dociac(seqidno:17)直接连接起来,从而获得pela-dociac序列。构建好的质粒转化到解纤维醋弧菌(acetivibriocellulolyticus)中。通过提取重组菌株的纤维小体发现,表达的pela可以外泌到胞外,并通过具有的dociac组装到解纤维醋弧菌纤维小体复合体中。
实施例18:通过直接性连接的方法,构建基于热纤梭菌纤维小体的全菌酶制剂
与实施例2不同的是,将果胶酶pela(seqidno:21)编码基因与热纤梭菌的i型对接模块docict的序列(seqidno:9)直接连接起来,获得pela-docict目标序列用于构建表达质粒phk(图1)。将构建好的质粒转化到在实施例13构建的热纤梭菌重组菌株10中,获得同时表达具有ii型粘连模块的木聚糖酶和具有i型对接模块的纤维素外切酶cel9-48和果胶酶pela的热纤梭菌重组菌株12。通过提取重组菌株的纤维小体发现,表达的具有docict或cohiict的xyna、cel9-48、pela都可以外泌到胞外,并组装到纤维小体复合体中。
实施例19:通过直接性连接的方法,构建基于热纤梭菌纤维小体的全菌酶制剂
将实施例14构建的含有膨胀因子epn序列seqidno:4的同源重组质粒,转化到实施例13构建的同源重组菌株10中,按照实施例3的筛选方法(图3),获得实现膨胀因子与脚架蛋白sdba融合表达的重组菌株13。随后,将实施例18中构建的表达果胶酶pela(seqidno:21)的phk质粒转化到该重组菌株中,从而获得同时表达具有ii型粘连模块的xyna、具有i型对接模块的cel9-48和pela以及融合蛋白epn-sdba的热纤梭菌重组菌株14。通过提取重组菌株的纤维小体发现,表达的木聚糖酶xyna、纤维素外切酶cel9-48、果胶酶pela都通过直接性连接的方式结合在热纤梭菌纤维小体上、膨胀因子epn通过与脚架蛋白sdba的融合表达也可以外泌并组装到纤维小体复合体中。
实施例20:通过直接性连接的方法,构建基于热纤梭菌纤维小体的全菌酶制剂
与实施例3不同的是,将纤维素内切酶celz(seqidno:18)编码基因作为目标序列构建同源重组质粒。采用实施例3的质粒转化及菌株筛选方法获得同源重组菌株15。通过提取重组菌株的纤维小体发现,celz与cel48s的融合蛋白可以外泌到胞外,并组装到纤维小体复合体中。因此该重组菌株可以作为具有非纤维小体木聚糖酶的全菌酶制剂。
与实施例5相比,该全菌催化剂所具有的非纤维小体组分是通过与纤维小体组分的直接融合表达实现与纤维小体的结合的。
实施例21:通过直接性连接的方法,构建基于热纤梭菌纤维小体的全菌酶制剂
与实施例2不同的是,将纤维素内切酶celz(seqidno:18)与热纤梭菌的i型对接模块docict的序列(seqidno:9)直接连接起来,从而获得celz-docict序列。采用实施例3的质粒转化及菌株筛选方法获得表达具有i型对接模块的纤维小体内切酶celz的热纤梭菌重组菌株16。通过提取重组菌株的纤维小体发现,表达的具有docict的celz都可以外泌到胞外,并组装到纤维小体复合体中。因此该重组菌株可以作为具有非纤维小体纤维素内切酶的全菌酶制剂。
与实施例5相比,该全菌催化剂所具有的非纤维小体组分是通过直接结合纤维小体组装模块以及组装模块间的非共价相互作用,实现与纤维小体的结合的。
实施例22:制备游离酶制剂
将膨胀因子epn(seqidno:4)、果胶酶pela(seqidno:21)、纤维素外切酶cel9-48(seqidno:19)、木聚糖酶xyna(seqidno:3)序列分别克隆到表达质粒pet28a中,并转化到大肠杆菌菌株bl21(de3),获得转化子后,经过测序验证后,在lb培养基中在37摄氏度条件下进行培养,当细胞生长到对数生长中期时,通过添加0.5mmiptg诱导蛋白的表达。随后通过基于组氨酸标签的亲和层析法纯化,分别获得游离的膨胀因子epn、果胶酶pela、纤维素外切酶cel9-48、木聚糖酶xyna。通过蛋白浓度测定,将四种游离酶按照1:1:1:1的蛋白摩尔数比例进行混合从而得到游离酶制剂。在60摄氏度条件下,每毫克该酶制剂具有400酶活单位的木聚糖酶活力。
实施例23:全菌酶制剂用于微晶纤维素糖化
采用实施例5-11、13-14、19构建的重组菌株作为酶制剂进行糖化反应,采用向培养基中直接添加100克每升微晶纤维素作为底物,所有操作都在厌氧条件下进行,分析不同酶制剂对纤维素底物水解的有益效果。
具体操作为:将实施例5-11、13-14、19构建的重组菌株预先在以5克每升微晶纤维素为碳源的gs-2培养基中培养至对数中期,然后按照1%(体积比)的接种量接种到以100克每升微晶纤维素为碳源的gs-2培养基中,在相应反应温度(表1),170r/min的摇床中进行培养。每12小时取样检测发酵液中还原糖的含量,当还原糖浓度高于20g/l时对培养基进行更换,减少发酵液中还原糖的累积,将获得的还原糖量进行累加,从而计算底物中纤维素到可溶性糖的转化率。转化率达到90%(质量比)的时间定义为木质纤维素底物的水解周期(天)。
与对照酶制剂(实施例1构建的热纤梭菌重组菌株或未经过改造的相应出发菌株)相比,水解周期大大缩短,说明全菌酶制剂在糖化微晶纤维素中具有有益效果(详见表1)。
实施例24:全菌酶制剂用于预处理底物糖化
与实施例23不同的是,采用实施例2-19构建的重组菌株作为酶制剂进行糖化反应,采用向培养基中直接添加40克每升干重的预处理秸秆作为底物。
同时,将未经过改造热纤梭菌在以5克每升微晶纤维素为碳源的gs-2培养基中培养至对数中期,然后按照1%(体积比)的接种量接种到40克每升干重的预处理秸秆作为碳源的gs-2培养基中,在接种的同时添加实施例21所构建的游离酶制剂。游离酶制剂是根据其所包含的木聚糖酶的酶活单位进行添加,即,每克预处理底物添加包含有50个酶活单位木聚糖酶的游离酶制剂。预处理秸秆采用文献(binli,etal.recentprogressonthepretreatmentandfractionationoflignocellulosesforbiorefineryatqibebt.journalofbioresourcesandbioproducts,2017,2(1),4-9)中的预处理技术获得,并采用木聚糖酶处理。处理后纤维素含量不小于45%(质量分数),半纤维素含量不大于15%(质量分数),木质素含量不大于12%(质量分数)。
与对照酶制剂(实施例1构建的热纤梭菌重组菌株或未经过改造的相应出发菌株)相比,实施例2-19构建的全菌酶制剂水解周期大大缩短,说明全菌酶制剂在糖化微晶纤维素中具有有益效果(详见表1)。
同时,与添加了实施例22所构建的游离酶制剂的对照酶制剂(未经过改造的相应出发菌株)相比,实施例19构建的全菌酶制剂水解周期更短,且不同非纤维小体组分之间的协同效应更加显著。
实施例25:全菌酶制剂用于预处理底物糖化
与实施例24不同的是,采用实施例2-6、11-12、18-19构建的重组菌株作为酶制剂进行糖化反应,预处理秸秆未经过木聚糖酶处理。处理后纤维素含量不小于35%(质量分数),半纤维素含量不大于21%(质量分数),木质素含量不大于15%(质量分数)。
与对照酶制剂(实施例1构建的热纤梭菌重组菌株1或未经过改造的相应出发菌株)相比,采用实施例2-6、11-12、18-19构建的重组菌株水解周期大大缩短,说明全菌酶制剂在糖化微晶纤维素中具有有益效果(详见表1)。
实施例26:全菌酶制剂用于预处理底物糖化
与实施例24不同的是,采用实施例19构建的重组菌株作为酶制剂进行糖化反应,采用向培养基中直接添加150克每升干重的预处理秸秆作为底物,在旋转混匀器中进行培养。
与对照酶制剂(实施例1构建的热纤梭菌重组菌株或未经过改造的相应出发菌株)相比,实施例19构建的重组菌株水解周期大大缩短,说明全菌酶制剂在糖化微晶纤维素中具有有益效果(详见表1)。
表1实施例23-26纤维素底物糖化的周期
由表1可知,
(1)来源于热纤梭菌的全菌酶制剂,根据实施例23可知,用于微晶纤维素糖化时,其反应周期为6.5-14天;与实施例1构建的只外泌表达β-1,4葡萄糖苷酶的对照酶制剂相比,缩短了6.7-56.7%;根据实施例24可知,用于预处理的秸秆(40克每升干重)糖化时,其反应周期为3.5-11.5天;与对照相比,缩短了4.2-70.8%;根据实施例22可知,用于未经预处理的秸秆糖化时,其反应周期为7.5-14天;与实施例1构建的只外泌表达β-1,4葡萄糖苷酶的对照酶制剂相比,缩短了6.7-50%;根据实施例26可知,用于高固含量的预处理秸秆(150克每升干重)糖化时,其反应周期为6.5-19天;与实施例1构建的只外泌表达β-1,4葡萄糖苷酶的对照酶制剂相比,缩短了13.6-70.5%。
来自黄色溶纤梭菌的全菌酶制剂,根据实施例23可知,用于微晶纤维素糖化时,其反应周期为15天;与对照相比,缩短了16.7%;根据实施例24可知,用于预处理的秸秆(40克每升干重)糖化时,其反应周期为13天;与对照相比,缩短了29.7%。
来自白色瘤胃球菌的全菌酶制剂,根据实施例23可知,用于微晶纤维素糖化时,其反应周期为21.5天;与对照相比,缩短了14%;根据实施例24可知,用于预处理的秸秆(40克每升干重)糖化时,其反应周期为15天;与对照相比,缩短了34.8%。
来自黄化瘤胃球菌的全菌酶制剂,根据实施例23可知,用于微晶纤维素糖化时,其反应周期为18.5天;与对照相比,缩短了26%;根据实施例24可知,用于预处理的秸秆(40克每升干重)糖化时,其反应周期为15天;与对照相比,缩短了40%。
来自解纤维梭菌的全菌酶制剂,根据实施例23可知,用于微晶纤维素糖化时,其反应周期为21.5天;与对照相比,缩短了14%;根据实施例24可知,用于预处理的秸秆(40克每升干重)糖化时,其反应周期为17.5-20.5天;与对照相比,缩短10.9-23.9%。
来自溶纤维假拟杆菌的全菌酶制剂,根据实施例23可知,用于微晶纤维素糖化时,其反应周期为14天;与对照相比,缩短了30%;根据实施例24可知,用于预处理的秸秆(40克每升干重)糖化时,其反应周期为12.5-14天;与对照相比,缩短了26.3-34.2%;根据实施例22可知,用于未经预处理的秸秆糖化时,其反应周期为14.5-15天;与对照相比,缩短了31.8-34.1%。
来自嗜纤维梭菌的全菌酶制剂,根据实施例24可知,用于预处理的秸秆(40克每升干重)糖化时,其反应周期为21天;与对照相比,缩短了12.5%。
来自解纤维醋弧菌的全菌酶制剂,根据实施例24可知,用于预处理的秸秆(40克每升干重)糖化时,其反应周期为20天;与对照相比,缩短了9.7%。
综上可知,实施例2-19制备的全菌酶制剂——即纤维小体改造后的菌株,与只外泌表达β-1,4葡萄糖苷酶的热纤梭菌菌株或者未经过改造的相应出发菌株相比,均显著缩短了糖化周期,说明所述全菌酶制剂大大提高了糖化效率,从而降低了生产成本,具备广阔的市场应用前景和巨大的经济价值。
(2)由实施例23可知,实施例5、20、21所制备的全菌酶制剂都是通过向纤维小体中添加celz这一非纤维小体组分获得的。其中,采用实施例5制备的全菌催化剂时反应周期为10.5天,与实施例1构建的只外泌表达β-1,4葡萄糖苷酶的对照酶制剂相比,缩短了30.0%;采用实施例20制备的全菌催化剂时反应周期为13.5天,与对照相比,缩短了10.0%;采用实施例21制备的全菌催化剂时反应周期为12天,与实施例1构建的只外泌表达β-1,4葡萄糖苷酶的对照酶制剂相比,缩短了20.0%。而上述三个实施例所制备的全菌酶制剂的区别在于,实施例5是基于共价交联的间接连接的方式添加非纤维小体组分,实施例20是通过与纤维小体组分的直接融合表达方式添加非纤维小体组分,实施例21是通过纤维小体组装模块间的非共价相互作用方式添加非纤维小体组分。
由此可见,采用共价交联的间接连接方式或基于纤维小体组装模块间非共价相互作用的直接连接方式,与融合表达的直接连接方式相比,更有利于糖化效率的提高。这主要是由于,融合表达的方式会降低所融合的纤维小体组分的表达量。
(3)由实施例24可知,单独使用实施例4采用纤维素外切酶9-48-doci改造的菌株时,反应周期为10天,与实施例1构建的只外泌表达β-1,4葡萄糖苷酶的对照酶制剂相比,缩短了16.7%。单独使用实施例6采用木聚糖酶xyna-cohii改造的菌株时,反应周期为9天,与对照相比,缩短了25%。单独使用实施例14采用膨胀因子改造的菌株时,反应周期为11.5天,与实施例1构建的只外泌表达β-1,4葡萄糖苷酶的对照酶制剂相比,缩短了4.2%。单独使用实施例17采用果胶酶pela-dociac改造的菌株时,反应周期为20天,与实施例1构建的只外泌表达β-1,4葡萄糖苷酶的对照酶制剂相比,缩短了9.1%。
而使用纤维素外切酶9-48-doci和木聚糖酶xyna-cohii改造的菌株时,反应周期为6天,与实施例1构建的只外泌表达β-1,4葡萄糖苷酶的对照酶制剂相比,缩短了50%。采用纤维素外切酶、木聚糖酶和果胶酶改造的菌株时,反应周期为4.5天,与实施例1构建的只外泌表达β-1,4葡萄糖苷酶的对照酶制剂相比,缩短了62.5%。采用纤维素外切酶、木聚糖酶、果胶酶和膨胀因子改造的菌株时,反应周期为3.5天,与实施例1构建的只外泌表达β-1,4葡萄糖苷酶的对照酶制剂相比,缩短了70.8%。
从而说明,不同非纤维小体组分之间、以及非纤维小体组分与纤维小体之间存在协同效应,因此不同非纤维小体组分同时组装到纤维小体中可以进一步缩短水解周期,这在产业上具有非常大的意义,将极大的推动木质纤维素糖化的产业化进程。
(4)由实施例24可知,采用实施例1构建的只外泌表达β-1,4葡萄糖苷酶的对照酶制剂添加实施例22所构建的游离酶制剂时,反应周期为9.5天,与实施例1构建的只外泌表达β-1,4葡萄糖苷酶的对照酶制剂相比,缩短了20.0%。而采用实施例19构建的全菌酶制剂,反应周期为3.5天,与实施例1构建的只外泌表达β-1,4葡萄糖苷酶的对照酶制剂相比,缩短了70.8%。
这说明,与添加游离酶制剂相比,所述全菌酶制剂显著缩短了水解周期;进一步说明,所述全菌酶制剂通过增强非纤维小体组分之间以及非纤维小体组分与纤维小体之间的协同作用关系,提高了糖化效率,降低了糖化过程中酶的消耗量,简化了工艺流程,降低了生产成本。
(5)与现有技术相比,本发明所述的全菌酶制剂,在纤维素糖化体系以及具有低固(4%质量比)和高固含量(15%质量比)的复杂底物(预处理底物)糖化体系中,其糖化周期的缩短和糖化效率的提高,具有更加明显的优势。
序列表
<110>中国科学院青岛生物能源与过程研究所
<120>用于催化木质纤维素糖化的全菌酶制剂
<160>17
<170>siposequencelisting1.0
<210>1
<211>394
<212>prt
<213>热纤梭菌(clostridiumthermocellum)
<400>1
gluphelysasnserproserproglnaspleumetaspphetyrile
151015
aspaspphethralathrproalaasnleuprogluileglulysasp
202530
ileproserleulysaspvalphealaglytyrphelysvalglygly
354045
alaalathrvalalagluleualaprolysproalalysgluleuphe
505560
leulyshistyrasnserleuthrpheglyasngluleulysproglu
65707580
servalleuasptyraspalathrilealatyrmetglualaasngly
859095
glyaspglnvalasnproglnilethrleuargalaalaargproleu
100105110
leugluphealalysgluhisasnileprovalargglyhisthrleu
115120125
valtrphisserglnthrproasptrpphephearggluasntyrser
130135140
glnaspgluasnalaprotrpalaserlysgluvalmetleuglnarg
145150155160
leugluasntyrilelysasnleumetglualaleualathrglutyr
165170175
prothrvallysphetyralatrpaspvalvalasnglualavalasp
180185190
proasnthrseraspglymetargthrproglyserasnasnlysasn
195200205
proglyserserleutrpmetglnthrvalglyargasppheileval
210215220
lysalapheglutyralaarglystyralaproalaaspcyslysleu
225230235240
phetyrasnasptyrasnglutyrgluasparglyscysasppheile
245250255
ilegluileleuthrgluleulysalalysglyleuvalaspglymet
260265270
glymetglnserhistrpvalmetasptyrproserilesermetphe
275280285
glulysserileargargtyralaalaleuglyleugluileglnleu
290295300
thrgluleuaspileargasnproaspasnserglntrpalaleuglu
305310315320
argglnalaasnargtyrlysgluleuvalthrlysleuvalaspleu
325330335
lyslysgluglyileasnilethralaleuvalphetrpglyilethr
340345350
aspalathrsertrpleuglyglytyrproleuleupheaspalaglu
355360365
tyrlysalalysproalaphetyralailevalasnservalpropro
370375380
leuprothrgluproprovalglnvalile
385390
<210>2
<211>356
<212>prt
<213>粪堆梭菌(clostridiumstercorarium)
<400>2
lyslysalapheasnaspglnthrseralagluaspileproserleu
151015
alaglualapheargasptyrpheproileglyalaalaileglupro
202530
glytyrthrthrglyglnilealagluleutyrlyslyshisvalasn
354045
metleuvalalagluasnalametlysproalaserleuglnprothr
505560
gluglyasnpheglntrpalaaspalaaspargilevalglnpheala
65707580
lysgluasnglymetgluleuargphehisthrleuvaltrphisasn
859095
glnthrprothrglypheserleuasplysgluglylysprometval
100105110
glugluthraspproglnlysargglugluasnarglysleuleuleu
115120125
glnargleugluasntyrileargalavalvalleuargtyrlysasp
130135140
aspilelyssertrpaspvalvalasngluvalilegluproasnasp
145150155160
proglyglymetargasnserprotrptyrglnilethrglythrglu
165170175
tyrilegluvalalapheargalaalaargglualaglyglyserasp
180185190
ilelysleutyrileasnasptyrasnthraspaspprovallysarg
195200205
aspileleutyrgluleuvallysasnleuleuglulysglyvalpro
210215220
ileaspglyvalglyhisglnthrhisileaspiletyrasnpropro
225230235240
valgluargileilegluserilelyslysphealaglyleuglyleu
245250255
aspasnileilethrgluleuaspmetseriletyrsertrpasnasp
260265270
argserasptyrglyaspserileproasptyrileleuthrleugln
275280285
alalysargtyrglngluleupheaspalaleulysgluasnlysasp
290295300
ilevalseralavalvalphetrpglyileserasplystyrsertrp
305310315320
leuasnglypheprovallysargthrasnalaproleuleupheasp
325330335
argasnphemetprolysproalaphetrpalailevalaspproser
340345350
argleuargglu
355
<210>3
<211>212
<212>prt
<213>热解纤维素果汁杆菌(caldicellulosiruptorsp.)
<400>3
alailethrleuthrserasnalaserglythrtyraspglytyrtyr
151015
tyrgluleutrplysaspserglyasnthrthrmetthrvalaspthr
202530
glyglyargphesercysglntrpserasnileasnasnalaleuphe
354045
argthrglylyslyspheasnthralatrpasnglnleuglythrval
505560
lysilethrtyrseralathrtyrasnproasnglyasnsertyrleu
65707580
cysiletyrglytrpserlysasnproleuvalgluphetyrileval
859095
glusertrpglysertrpargproproglyalathrserleuglythr
100105110
valthrileaspglyglythrtyraspiletyrlysthrthrargval
115120125
asnglnproserilegluglythrthrthrpheaspglntyrtrpser
130135140
valargthrserlysargthrserglythrvalthrvalthrasphis
145150155160
phelysalatrpalaalalysglyleuasnleuglythrileaspgln
165170175
ilethrleucysvalgluglytyrglnserserglyseralaasnile
180185190
thrglnasnthrpheserilethrseraspserserglyserthrthr
195200205
prothrthrthr
210
<210>4
<211>327
<212>prt
<213>黄色溶纤梭菌(clostridiumclariflavum)
<400>4
metasnphelyslysileargleuphethralaileleuileileala
151015
alaglnvalleusertyrasnpheileserseralaglnleuglnval
202530
glyaspvalasnglyaspasnasnvalaspserileaspphealaleu
354045
metlysserpheileleulysileileasnthrleuproalagluasp
505560
serleuleualaglyaspleuaspglyaspglyserileasnserile
65707580
aspcysalaleumetlysglntyrleuleuglymetilelysvalphe
859095
prolysthrglnserproalaprothrprothrasnthrproleupro
100105110
glutyrsergluprotyrproglytrpasplysileargserglytyr
115120125
alathrtyrthrglyserglytyrvalglyglyilealaleuleuasp
130135140
proileprogluaspmetgluilevalalavalasnlysproaspphe
145150155160
asncystyrglyvalglnalaalaleualaglyalatyrleugluval
165170175
thrglyprolysglythrthrvalvaltyrvalthraspcystyrthr
180185190
glualaprogluglyalaleuaspleucysglyilesercysasplys
195200205
ileglyaspthrasnvalproglyglylysileaspvalthrtrparg
210215220
ileileproalaproilethrglyasnpheiletyrargileleupro
225230235240
alaserserlystrptrpphealaileglnvalargasnhislystyr
245250255
provalmetlysmetglutyrphelysaspglyglutrpvalaspile
260265270
prolysaspargcysasntyrphevalileasnasnleuaspthrser
275280285
asnleulysileargilethraspileargglylysvalvalthrasp
290295300
ileileaspproileproaspasnleumetasnglycyspheilegln
305310315320
glyasnvalglnpheproasp
325
<210>5
<211>335
<212>prt
<213>黄色溶纤梭菌(clostridiumclariflavum)
<400>5
metlysarglysileilephecyscysalaileleumetthrleuleu
151015
glnpheserpheleuasnserpheserleugluglyglnvalthrval
202530
glyaspileasnglyaspglytyrvalaspserileasptyralaasn
354045
leulysmettyrvalleuglyleuilelysasppheprothralaglu
505560
glysertrpalaalaaspvalaspglyasnasnserileaspserile
65707580
aspcysalaleumetlyssertyrleuleuglyileilelyslysphe
859095
prolysseraspserleuproalaasnthrseralavalthrprothr
100105110
proglnilethrprolysproileprosertyrserprotyrproasp
115120125
trpasplysgluhisserglytyralathrphethrglyserglytyr
130135140
serglyglyalaalaleuleuaspproileaspproaspmetgluile
145150155160
thralaleuasnprotyrasptyrasnsertyrglyileaspalaala
165170175
leualaglyalatyrleugluvalthrglyglulysglyserthrile
180185190
valtyrvalthraspleutyrprogluglyalaproglyalaleuasp
195200205
leucysprolysserphealalysileaspaspileserlysglylys
210215220
ileaspilelystrpargvalvalalaalaprovalthrglyasnval
225230235240
sertyrargilelysgluglyserthrthrsertrpleualavalgln
245250255
valargasnhisargtyrprovalleulysmetglucystyrileasn
260265270
asnthrtrpvalaspmetlyslysmetphetrpasnhisphevalile
275280285
aspasnvalaspthrthrthrproargileargmetthraspilearg
290295300
glytyrvalleugluaspvalleuaspserileprovalserglyasp
305310315320
thrargproalatyrileileaspglyasnvalglnpheproglu
325330335
<210>6
<211>440
<212>prt
<213>溶蛋白杆菌(coprothermobacterproteolyticus)
<400>6
metlyslysileleuleuthrleuvalilealavalleuleuleuser
151015
glyphealaglyvallysseralagluleuleuphevalserasnser
202530
thrthrthrasnglngluaspprogluasngluileilevalglytyr
354045
lysgluasnthraspvalalavalleuserlysglnvalglulysthr
505560
thrglyalalysleuserarglysglyleulysasnphealavalphe
65707580
lysleuproglnglylysalaalaaspvalvalmetasnglnleulys
859095
asnaspproasnvalglutyrvalgluproasntyrilealahisala
100105110
pheaspvalproasnaspthrphepheasnprotyrglntrpasnphe
115120125
tyrasptyrglymetthrserasnglytyrvalserasntyrglyile
130135140
glnalavalseralatrpasnilethrlysglyalaglyvallysval
145150155160
alaileileaspthrglyvalalatyrgluasntyrglyalatyrthr
165170175
lysalaproaspleualaasnthrleupheaspthralaasnalatyr
180185190
aspphevalasnasnaspthrhisalaasnaspaspasnserhisgly
195200205
thrhisvalalaglythrilealaglnserthrasnasnglymetgly
210215220
alaalaglyilealatyrglnalathrileleuproilelysvalleu
225230235240
aspsergluglyserglythrtyraspalailealaasnglyileile
245250255
trpalaalaasplysglyalaargvalileasnmetserleuglygly
260265270
serserglyserthrthrleuglnasnalaileglntyralatyrasn
275280285
lysglyvalvalilevalcysalaserglyasnaspargargserthr
290295300
valsertyrproalaalatyrthrglncysilealavalglyserthr
305310315320
argpheaspglythrargalaargtyrserasntyrglyseralaleu
325330335
aspilevalalaproglyglyaspthrservalaspglnasnhisasp
340345350
glytyrglyaspglyileleuglnglnthrphealagluglyserpro
355360365
thraspphealatyrtyrphepheglnglythrsermetalaserpro
370375380
hisvalalaglyvalalaalaleuvalleuseralahisprothrtyr
385390395400
thrasngluglnvalargthralaleuglnserthralalysaspleu
405410415
glythralaglytrpasplystyrtyrglytyrglyleuvalasnala
420425430
tyralaalavalasntrpthrpro
435440
<210>7
<211>13
<212>prt
<213>酿脓链球菌(streptococcuspyogenes)
<400>7
alahisilevalmetvalaspalatyrlysprothrlys
1510
<210>8
<211>97
<212>prt
<213>酿脓链球菌(streptococcuspyogenes)
<400>8
sersergluglnglyglnserglyaspmetthrileglugluaspser
151015
alathrhisilelyspheserlysargaspgluaspglylysgluleu
202530
alaglyalathrmetgluleuargaspserserglylysthrileser
354045
thrtrpileseraspglyglnvallysaspphetyrleutyrprogly
505560
lystyrthrphevalgluthralaalaproaspglytyrgluvalala
65707580
thralailethrphethrvalasngluglnglyglnvalthrvalasn
859095
gly
<210>9
<211>55
<212>prt
<213>热纤梭菌(clostridiumthermocellum)
<400>9
tyrglyaspvalasnaspaspglylysvalasnserthraspalaval
151015
alaleulysargtyrvalleuargserglyileserileasnthrasp
202530
asnalaaspleuasngluaspglyargvalasnserthraspleugly
354045
ileleulysargtyrileleu
5055
<210>10
<211>160
<212>prt
<213>热纤梭菌(clostridiumthermocellum)
<400>10
serserilegluleulyspheaspargasnlysglygluvalglyasp
151015
ileleuileglythrvalargileasnasnilelysasnphealagly
202530
pheglnvalasnilevaltyraspprolysvalleumetalavalasp
354045
progluthrglylysgluphethrserserthrpheproproglyarg
505560
thrvalleulysasnasnalatyrglyproileglnilealaaspasn
65707580
aspproglulysglyileleuasnphealaleualatyrsertyrile
859095
alaglytyrlysgluthrglyvalthrglugluserglyileileala
100105110
lysileglyphelysileleuglnlyslysserthralavallysphe
115120125
glnaspthrleusermetproglyalaileleuglythrglnleuphe
130135140
asptrpaspglygluvalilethrglytyrgluvalileglnproasp
145150155160
<210>11
<211>891
<212>dna
<213>热纤梭菌(clostridiumthermocellum)
<400>11
tagtactattaaagtcagacttggttaaatataaattttatgacttgcatacaaacttga60
tgtgtattataataaaaatacaaaacaaaatagcaataatttactcagttatttttgaaa120
tgggggtagtattaatatcgtataatggggttgcatatctgctgtctttcgaaaaaagca180
caagaacttcaaatgtttccatagtgaaatttaaaaattggagatttctttgttgccccc240
tcaaaaagtatatttttttcgaagatatatatatggaatttattgattaatttaagttat300
taattttggccttttagggtcgttgaaaactgaatatgttaagttgttttgcgtgattca360
gctgcatttgacgtaagacttcgccggtctgtttaaattcccataataagatgtatttat420
tgtagtaataatctggcatctacaaatttcagtatttgcaatagtctctgttcaaaaaag480
caattgtcttttaaacctttcagtattgtcttcgtggcagtttcttttgttatacgtcgt540
tccgacaaaaaaatgtaaatttatgtcaaatgcgcggctgatttgataaaaaagtttgtt600
aacacaaatttattatgttaacacaagtattttttgggtccagcttagttttatgatgaa660
aataatgcgtaaaatttatccgcaaaaagggggaatgaatttattgcgggtaggttgcat720
tatttcatcatataacttaaaaagaataaaaaagtatatttgaaaggggaagatggagag780
atggtaaaaagcagaaagatttctattctgttggcagttgcaatgctggtatccataatg840
atacccacaactgcattcgcaggtcctacaaaggcacctacaaaagatggg891
<210>12
<211>59
<212>prt
<213>黄色溶纤梭菌(clostridiumclariflavum)
<400>12
tyrglyaspleuasnglyasplysleuvalasnserileasppheala
151015
leuleulysiletyrleuleuglytyrserlysglupheprotyrglu
202530
tyrglyilelysseralaaspleuasnargasnglygluvalaspser
354045
ileaspphealaileleuargserpheleuleu
5055
<210>13
<211>57
<212>prt
<213>白色瘤胃球菌(ruminococcusalbus)
<400>13
argglyaspvalasnglyaspglyvalvalasnvalthraspvalala
151015
lysilealaalahisvallysglylyslysileleuthrglyaspser
202530
leulysasnalaaspvalasnpheaspglyservalasnilethrasp
354045
ilethrargilealaalaphevallys
5055
<210>14
<211>56
<212>prt
<213>黄化瘤胃球菌(ruminococcusflavefaciens)
<400>14
tyrglyaspalaasncysaspglyasnvalserilealaaspalathr
151015
alaileleuglnhisleuglyasnargasplystyrglyleuargala
202530
glnglymetleuasnalaaspvalaspglyglnserglyvalthrala
354045
asnaspalaleuvalleuglnlys
5055
<210>15
<211>54
<212>prt
<213>解纤维梭菌(clostridiumcellulolyticum)
<400>15
tyrglyasptyrasnasnaspglyserileaspalaleuasppheser
151015
serphelysmettyrleumetasnprovalargthrtyrthrgluval
202530
leuaspleuasnseraspasnthrvalaspalaileaspphealaile
354045
metlysglntyrleuleu
50
<210>16
<211>57
<212>prt
<213>溶纤维假拟杆菌(pseudobacteroidescellulosolvens)
<400>16
tyrglyaspvalthrglyaspglnleuvalthraspalaasplysthr
151015
lysvalserasntyrileleuglyservaltyrleuthrserargglu
202530
phealaalaalaaspvalasnglyaspglnvalvalasnserglyasp
354045
leuthrleuileasnarghisileleu
5055
<210>17
<211>58
<212>prt
<213>解纤维醋弧菌(acetivibriocellulolyticus)
<400>17
lysglyaspvalaspleuaspglyalaalaasnserileaspphegly
151015
lysmetargleucysleuleuglylysserproalaphethrglygln
202530
alaleuaspasnalaaspleuasnaspaspglyalapheasnserile
354045
asppheglytyrmetarglyslysleuleu
5055
- 还没有人留言评论。精彩留言会获得点赞!