多肽LDHA237-244及其作为参与肥胖相关慢性炎症的CD8+T细胞识别的靶点的制作方法
属于免疫识别和治疗领域,涉及来源于乳酸脱氢酶a链的多肽ldha237-244及其作为参与肥胖相关慢性炎症的cd8+t细胞识别的靶点。
背景技术:
随着全球经济的快速发展,生活水平的日益提高、饮食结构不断变化,以及体力劳动的减少,肥胖症的发病率与日俱增,已成为全球首要健康问题。目前肥胖与艾滋病、吸毒、酗酒并列为世界性四大医学社会问题。肥胖常伴随着一系列的健康问题,包括胰岛素抵抗、2型糖尿病、脂肪肝、动脉粥样硬化及退行性疾病(如痴呆)、呼吸道疾病、癌症等,而这些疾病的发生与存在于内脏脂肪组织中的慢性炎症密切相关。
肥胖诱导的慢性炎症与多种代谢性疾病密切相关,肥胖促发慢性炎症的机制研究成为肥胖及相关代谢疾病研究的前沿和热点,近年来研究证实:在肥胖相关慢性炎症中,特异性免疫细胞t细胞是内脏脂肪组织定居巨噬细胞(adiposetissueresidentmacrophages:atms)极化的关键调控者。内脏脂肪组织(vat)浸润的效应性cd8+t细胞参与招募atm及启动脂肪组织慢性炎症,但这些cd8+t细胞的活化受到何种靶抗原驱动尚未见报道。我们推测在营养和能量过剩所致脂肪细胞代谢紊乱状态下,小鼠vat中可能有一些新生自身抗原肽产生,并通过激活特异性cd8+t细胞免疫反应而参与肥胖相关慢性炎症的发生及维持。对于肥胖慢性炎症相关cd8+t细胞免疫识别及活化机制的研究,有助于从全新角度深入理解肥胖诱导慢性炎症发生机制,并可能发现全新的免疫治疗靶点。
技术实现要素:
有鉴于此,本发明的目的在于提供一种来源于乳酸脱氢酶a链的多肽ldha237-244及其作为参与肥胖相关慢性炎症的cd8+t细胞识别的靶点。
为达到上述目的,本发明提供如下技术方案:
利用高分辨率高通量质谱分析技术发现来源于乳酸脱氢酶a链的多肽ldha237-244,其氨基酸序列为sayevikl,如seqidno.1所示;
多肽ldha237-244,是一条特异性存在于肥胖内脏脂肪组织中被mhc-i类分子提呈的新生肽;
多肽ldha237-244具有免疫原性可被cd8+t细胞识别与活化,并产生促炎反应,也就是说,ldha237-244可有效激活cd8+t细胞炎症反应,是参与肥胖相关慢性炎症的cd8+t细胞识别的靶点,可能作为预防肥胖相关慢性炎症的潜在疫苗靶标。本发明的有益效果在于:
本发明通过建立高脂饮食诱导的肥胖(dio)小鼠模型,获取dio小鼠与正常对照小鼠附睾脂肪组织(内脏脂肪组织,vat)mhc-i类分子相关免疫肽组(mips);首次利用高分辨率高通量质谱分析技术,对比正常与肥胖小鼠vat相关mip差异,揭示高脂饮食产生脂肪细胞应激及压力可能导致mhc-i类免疫肽组产生变化,而一些新生的抗原肽如ldha237-244则成为早期驱动cd8+t细胞激活的关键因素。申请人发现这些抗原肽均来源于正常表达于脂肪细胞中的蛋白质,前期研究发现口服这些来源于脂肪组织的蛋白质混合物可诱导特异性免疫耐受性,可有效保护hfd诱导的肥胖及肥胖相关代谢紊乱,故这些具有免疫原性的内脏脂肪组织新生抗原肽可以作为负调疫苗候选靶标,未来预防肥胖可能通过口服某蛋白抗原或肽疫苗就能轻松实现。
附图说明
为了使本发明的目的、技术方案和有益效果更加清楚,本发明提供如下附图进行说明:
图1为高脂饮食诱导的肥胖模型建立;
图2为高脂和正常饮食小鼠脾脏及内脏脂肪组织mips特征分析;
图3为仅存在于肥胖脂肪组织中的新生抗原肽诱导cd8+t细胞促炎免疫应答;
图4为肥胖脂肪组织新生抗原ldha237-244诱导cd8+t细胞促炎免疫应答;
图5为洗脱下来的ldha237-244和合成的ldha237-244二级质谱图对比;
图6为ldha234-244特异性t细胞在hfd喂养的肥胖小鼠体内表达增加。
具体实施方式
下面将结合附图,对本发明的优选实施例进行详细的描述。
实施例:
1、建立高脂饮食诱导的肥胖(dio)小鼠模型
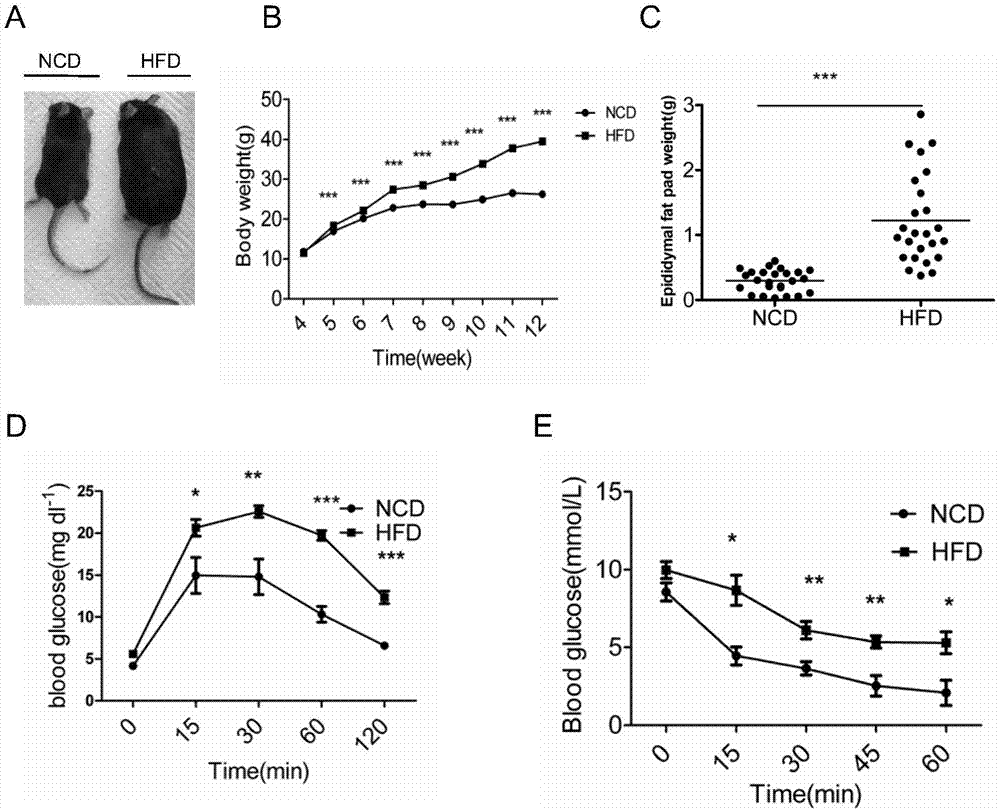
采用d12492(含有60%卡路里饲料)作为高脂饲料(hfd组),同时采用d12450b(含有10%卡路里饲料)作为正常对照(ncd组),动态监测小鼠体重变化情况,hfd组比ncd组体重显著增加(图1中a,b)、附睾脂肪组织重量也显著增加(图1中c)(第12周);分别检测第12周和第13周的各组小鼠葡萄糖耐量及胰岛素敏感性,hfd组出现明显的葡萄糖耐受不良及胰岛素抵抗(图1中d-e),表明dio小鼠及代谢紊乱模型成功建立。
2、dio小鼠与正常对照小鼠附睾脂肪组织mhc-i类分子相关免疫肽组(mips)的获取及质谱分析
从12周龄正常小鼠或者肥胖c57小鼠内脏脂肪组织(vat)28g,溶解在40ml含完全蛋白酶抑制剂的冰冷的蛋白裂解缓冲液中(包含50mmtrisph8.0,150mmnacl,和体积百分数1%的chaps),裂解液经过3次离心,收集上清。hitrapnhs-activatedhp(gehealthcare)偶联上anti-h2-kbmaby-3,y-3抗体(bioxcell公司)(1mg/ml,10ml)4℃环境下循环4小时,蛋白裂解液4度环境下柱子过夜,亲和柱子依次用下列缓冲液清洗:50mmtris(ph8.0),150mmnacl,和体积百分数1%的chaps,50mmtris-hcl(ph8.0),150mmnacl,5mmedta;50mmtris-hcl(ph8.0),150mmnacl;50mmtris-hcl(ph8.0),450mmnacl;50mmtris-hcl(ph8.0),得到hla-肽组复合物。随后hla-肽组复合物用6ml10%的乙酸水溶液洗脱,过3kd超滤管分离hla和肽组复合物,分离出来的肽组复合物过c18柱子进行纯化,真空浓缩至20μl后上液质联用检测仪进行肽段序列检测。
分别从ncd、hfd小鼠的脾组织(spl)中获得总肽段数量为392条及583条,从ncd、hfd小鼠内脏脂肪组织(vat)中分别获得总肽段数量为932及813条。根据三次质谱结果重现性,并比对uniprot数据库中小鼠蛋白质组,对三次质谱结果重现性最好的多肽进行多肽来源分析,发现mips中约有三分之一的高重现性多肽为剪切多肽,无法追索其蛋白来源,因此在后续分析时被忽略。后续分析仅保留来源蛋白属于小鼠蛋白质组,且同时满足与h2-kb结合亲和力ic50<1000nm的多肽,最终从ncd、hfd小鼠vat中筛到的肽段分别为125条及71条,其中在ncd、hfd组vatmips同时存在的肽段有39条;从ncd、hfd小鼠的spl中获得肽段数量分别为84条及124条,其中在ncd、hfd组splmips同时存在的肽段有45条。进一步将筛选出的多肽的肽段长度分布、与h2-kb结合亲和力大小以及多肽序列特征进行分析,发现鉴定到的肽长度分布集中在8-mer最多,9-mer长度次之(图2中a),绝大多数肽段显示出与h2-kb较强的结合能力(图2中b、c),这些肽均具有典型的h2-kb结合的特征基序(图2中d),并对肽段来源的蛋白定位进行了分析(图2中e)。
3、对比正常与肥胖小鼠vat相关mip差异,筛选肥胖小鼠新生肽
进一步筛选出部分在ncd及hfdvatmips中出现差异的多肽,并根据不同差异进行了分组,各肽的蛋白来源、多肽序列、组织分布、表达丰度等如表1-a和表1-b所示。其中包括只在hfdvat肽组中出现,而没有在ncdvat肽组中出现的多肽。
表1-a.用来进行免疫原性鉴定的肽段
表1-b.用来进行免疫原性鉴定的肽段
注:表1-b中,“-”表示未检测到,“+”表示质谱检测到的相对定量,一个“+”对应表示1份。
4、明确肥胖小鼠vat特有的新生抗原肽激活cd8+t细胞反应的能力
将上述各组肽池、阳性对照肽gp33(杭州中肽生化有限公司),(终浓度100μg/ml)分别与tap缺陷的rma-s细胞系(atcc公司)孵育2小时,通过流式细胞术检测rma-s细胞表面h2-kb分子荧光强度,h2-kb分子荧光强度与外源性肽-h2kb分子亲和力呈正相关。结果如图3中a所示,三个混合肽池均表现出一定的h2-kb结合能力,其中g3肽池与h2-kb亲和力相对较高,其次是g1和g2结合力相同,但均低于病毒来源的h2-kb结合的阳性对照肽gp33。
分离c57bl/6小鼠脾细胞,调节细胞浓度为106/ml,然后分别给予各肽池(终浓度20μg/ml)刺激,il-2浓度50iu/ml,第3.5天半量换液,同时补充mil-2,第7天用rma-s作为抗原递呈细胞,负载肽池(终浓度10μg/ml)再次刺激小鼠脾细胞,用高尔基阻断剂阻断,胞内因子染色检测cd8+t细胞分泌ifn-γ和tnf-α的能力。结果如图4中b-e所示,经g1肽池诱导出的效应细胞能对负载相应肽池的rma-s细胞产生特异性反应,明显诱导cd8+t细胞分泌ifn-γ和tnf-α,这一反应能被h2-kb抗体y-3完全阻断,但对于不负载任何肽的rma-s细胞则无应答;而g2、g3肽池则不具有明显的免疫原性。结果如图3中b,c,d,e所示。
分别检测g1肽池中各肽刺激cd8+t细胞反应的能力,具体方案同前,结果如图4中a,b所示,ldha237-244能刺激显著的肽特异性cd8+t细胞促炎细胞因子分泌,其中刺激能力最强。设计不同浓度ldha237-244刺激cd8+t细胞反应的能力。发现能够呈剂量依赖关系刺激cd8+t细胞促炎因子ifn-γ和tnf-α的表达(图4中c)。
5、合成ldha237-244肽二级质谱图与原始肽组中相应肽段二级质谱对比
将洗脱下来检测到得ldha237-244肽二级质谱图和在杭州中肽生化有限公司合成的ldha237-244肽二级质谱图做对比,发现质谱峰图几乎一致(图5),证实质谱鉴定结果的可靠性。
6、在dio小鼠体内证实存在ldha237-244特异性cd8+t细胞
采用流式细胞技术对正常及肥胖小鼠肠系膜淋巴结检测ldha237-244特异性cd8+t细胞进行检测,分离正常及肥胖小鼠c57bl/6小鼠肠系膜淋巴结,调节细胞浓度为106/ml,加入10μlh2-kb-ldha237-244五聚体(美国proimmune公司制备合成),充分混匀,室温黑暗处孵育10分钟。加入合适体积的连接有荧光染料的cd8抗体,加入其它荧光染料的其它抗体(cd3或者cd4抗体)。2-8℃避光孵育20分钟。发现ldha237-244特异性cd8+t细胞在hfd小鼠中的表达量和ncd相比有显著差异,在hfd小鼠中表达增加(图6)。
最后说明的是,以上优选实施例仅用以说明本发明的技术方案而非限制,尽管通过上述优选实施例已经对本发明进行了详细的描述,但本领域技术人员应当理解,可以在形式上和细节上对其作出各种各样的改变,而不偏离本发明权利要求书所限定的范围。
序列表
<110>中国人民解放军陆军军医大学
<120>多肽ldha237-244及其作为促进肥胖相关慢性炎症的cd8+t细胞识别的靶点
<160>11
<170>siposequencelisting1.0
<210>1
<211>8
<212>prt
<213>人工序列(artificialsequence)
<400>1
seralatyrgluvalilelysleu
15
<210>2
<211>8
<212>prt
<213>人工序列(artificialsequence)
<400>2
alathrleuargtyralaserleu
15
<210>3
<211>8
<212>prt
<213>人工序列(artificialsequence)
<400>3
glyglntyrgluphehisserleu
15
<210>4
<211>8
<212>prt
<213>人工序列(artificialsequence)
<400>4
lyssertyraspmetserglnval
15
<210>5
<211>9
<212>prt
<213>人工序列(artificialsequence)
<400>5
valphetyrgluarggluvalglnmet
15
<210>6
<211>8
<212>prt
<213>人工序列(artificialsequence)
<400>6
aspasnvalilephegluargleu
15
<210>7
<211>9
<212>prt
<213>人工序列(artificialsequence)
<400>7
seralatyrglnargglygluserleu
15
<210>8
<211>8
<212>prt
<213>人工序列(artificialsequence)
<400>8
serthrphevaltyrasnthrmet
15
<210>9
<211>9
<212>prt
<213>人工序列(artificialsequence)
<400>9
alaasntyrglnargaspglypromet
15
<210>10
<211>9
<212>prt
<213>人工序列(artificialsequence)
<400>10
serasntyrgluargleugluserleu
15
<210>11
<211>8
<212>prt
<213>人工序列(artificialsequence)
<400>11
servalvalalaphehisasnleu
15
- 还没有人留言评论。精彩留言会获得点赞!