一种双特异性抗体及其应用的制作方法
本发明涉及生物制药技术领域,特别是涉及一种双特异性抗体及其应用。
背景技术:
癌症是21世纪人类健康面临的最大挑战之一。抗体药物在癌症的治疗中发挥着重要作用,随着临床数据的不断反馈,人们发现传统的单抗药物由于单一的表位容易造成耐药或者由于抗体药效不佳容易造成复发等限制了单抗药物在临床的应用上。双特异性抗体可以同时识别和结合两种不同的抗原表位,根据发挥作用的途径一般可分为两种,一种是通过协同作用同时阻断两种不同的信号通路而发挥作用,另一种是通过激活效应细胞(t细胞,nk细胞等)来发挥药效。首先针对多表位的双特异性抗体可以解决单抗单一表位造成的耐药;此外近些年来免疫治疗在肿瘤治疗中获得了巨大的成功,有研究者提出利用人体自身的免疫系统是是治疗肿瘤最为有效方式,效应细胞依赖的双特异性抗体可以利用人体自身的免疫细胞高效的杀伤肿瘤细胞。因此双特异性抗体以其独特的优势,在肿瘤的治疗中占据着举足轻重的地位。
目前研究最为广泛的为t细胞依赖的双特异性抗体,该抗体一条臂可以结合肿瘤相关抗原,同时另一条臂可以和效应t细胞表面的cd3分子结合,形成免疫突触从而激活t细胞,激活的t细胞可以释放穿孔素和颗粒酶通过裂解或者凋亡作用杀伤肿瘤细胞。
双特异性抗体早期主要的制备方法包括双杂交瘤细胞法和化学偶联方法。双杂交瘤细胞法产生的双特异性抗体一般为鼠源性抗体,具有较强的免疫原性,在临床上应用有限。化学偶联方法反应条件不佳且容易对抗体活性产生影响,因此应用范围不广。随着基因工程技术的发展,越来越多的研究者尝试用基因工程的方法制备双特异性抗体,例如目前应用最为广泛的基因泰克开发的knobintohole技术以及在其基础上罗氏开发的crossmab技术,虽然这些方法显著的改善了早期方法的缺陷,但是两组抗体之间仍然存在着一定的错配概率以及抗体的表达水平较低的缺陷。
传统的双特异性抗体形式为igg型,具有较大的分子结构,难以透过血管内皮质及细胞间隙从而到达实体瘤深部的肿瘤细胞,因此在实体瘤的治疗效果不如血液瘤。因此研究者尝试利用基因工程技术表达小型化的双特异性抗体,其中应用最为广泛的bite(双特异性t细胞衔接器),安进公司的blinatumomab(cd19×cd3)是fda第一个批准上市用于费城染色体阴性的复发性难治性前体b细胞急性淋巴细胞白血病(gerhardzugmaier,molecularimmunology,clinicaloverviewofanti-cd19
目前处于临床或者临床前研究的cd20×cd3双特异性抗多数是以利妥昔为靶抗体的igg型,如由trionpharma公司开发的处于临床ⅱ期的fbta-05,genentech公司开发的处于临床ⅰ期的cd20-tdb等。此外lu等人尝试将igg-cd20通过一段柔性连接结构与scfv-cd3抗体进行融合表达(chia-yenlu,biochemicalandbiophysicalresearchcommunications,tetravalentanti-cd20/cd3bispecificantibodyforthetreatmentofbcelllymphoma,2016,(473)808-813),制备得到的抗体分子量约200kda。目前尚无以ofatumumab抗体作为靶向cd20的双特异性抗体的研究,而且目前没有关于小型化的cd20×cd3双特异性抗体形式被报道,所以一种有适宜药动学性质的小型化的cd20×cd3双特异性抗体有待开发。此外对于t细胞依赖的双特异性抗体,肿瘤细胞激活t细胞后产生的较强的免疫级联反应,脱靶效应是造成其毒性的关键因素。2f(ab’)形式的双特异性抗由于缺乏fc段,不仅避免了fc片段与体内表达fcrn细胞相互作用带来的毒副反应,也可避免由于fc段引起的全身性的免疫副作用,进一步保证了抗体的特异性。此2f(ab’)形式的双特异性抗体保留了bite形式易于穿透肿瘤的性质,且有着较好的药动学性质,是双特异性抗体表达形式的首选。
技术实现要素:
本发明针对现有技术中存在的不足,提供了一种以cd20为靶抗原,以t细胞为效应细胞的基于抗体的抗原结合片段为基础的bi-2f(ab’)形式的双特异性抗体的制备方法和应用。
一种双特异性抗体,制备方法包括以下步骤:
(1)分别提供抗cd20和抗cd3的单克隆抗体的fab结构域,且两者fab结构域的c端带有lpxtg序列,分别命名为fab-cd20和fab-cd3,
(2)分别提供第一连接臂和第二连接臂,所述第一连接臂的两端分别具有寡甘氨酸接头和炔基接头,所述第二连接臂的两端分别具有寡甘氨酸接头和叠氮基接头,
(3)在sortasea酶催化下,lpxtg序列与寡甘氨酸接头发生转肽反应,使fab-cd20连接第一连接臂、fab-cd3连接第二连接臂,或者fab-cd20连接第二连接臂、fab-cd3连接第一连接臂,
(4)所述叠氮基接头与炔基接头发生环加成反应,使第一连接臂与第二连接臂相连,从而将fab-cd20和fab-cd3相连,获得所述双特异性抗体。
寡甘氨酸接头一般为1~5个甘氨酸残基,寡甘氨酸残基与lpxtg序列在sortasea酶催化下偶联。两种抗体的fab结构域分别偶联第一连接臂和第二连接臂后,再通过两个连接臂之间的叠氮基接头与炔基接头发生叠氮-炔基环加成反应(strain-promotedalkyne-azidecycloadditions,spaac),从而两种抗体的fab偶联获得双特异性抗体。spaac反应属于点击化学(clickchemistry)中最经典的反应。该反应在水溶液中就可以进行,且操作简单,不需要外源的催化剂,反应条件温和,反应速率快。最突出的优点就是反应收率高,节省成本投入,同时在生产过程中减少了易产生危险的有毒废物,为实现工业上大规模的生产创造了可能。
优选的,两者fab结构域仅在重链c端带有lpxtg序列。
所述fab-cd20的重链氨基酸序列如seqidno.1所示,轻链氨基酸序列如seqidno.2所示。
所述fab-cd3的重链氨基酸序列如seqidno.5所示,轻链氨基酸序列如seqidno.6所示。
所述lpxtg序列为lpetg,fab-cd20及fab-cd3与lpxtg序列之间具有柔性接头,所述柔性接头由若干甘氨酸残基和/或丝氨酸残基组成。
优选的,所述柔性接头为ggggs、(ggggs)2或(ggggs)3。
步骤(1)提供的fab-cd20和fab-cd3的lpxtg序列末端还连接有用于蛋白纯化的组氨酸标签。组氨酸标签用于对表达得到的抗体进行纯化,且该组氨酸标签在第一步与连接臂偶联时会被去除,不影响后续双特异性抗体的结构。
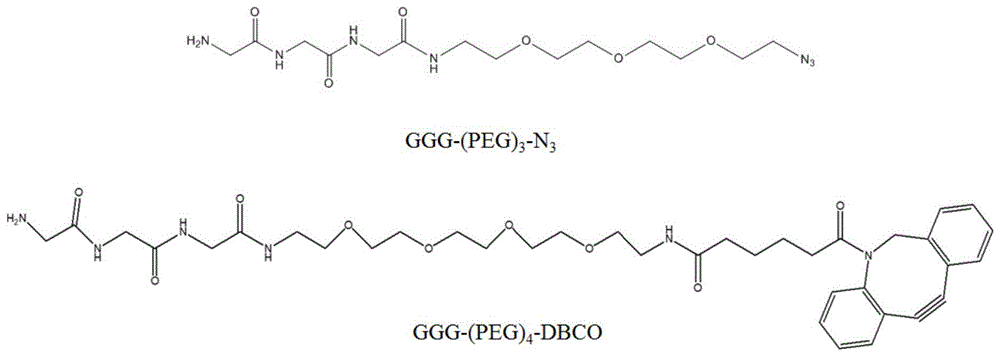
所述第一连接臂为ggg-(peg)m-dbco,第二连接臂为ggg-(peg)n-n3,其中m≤4,n≤4。peg为聚乙二醇,dbco为二苯基环辛炔接头。
本发明还提供了所述的双特异性抗体在制备抗肿瘤药物中的应用。
优选的,肿瘤类型为cd20阳性的b淋巴瘤。
本发明双特异性抗体的制备方法较传统的双特异性抗体的制备方法有着表达水平高,偶联效率高,组装正确率高的优点,该形式的双特异性抗体具有更高的亲和性和特异性,在体内外均能有效激活淋巴细胞并产生对cd20阳性的肿瘤细胞的特异性杀伤,而且由于缺乏抗体的fc段,不仅更易于穿透实体瘤,而且能够避免fc造成的非特异性杀伤的毒副作用具有更高的特异性。
附图说明
图1为连接臂小分子ggg-(peg)3-n3和ggg-(peg)4-dbco的化学结构式示意图。
图2为酶促-化学偶联制备bi-2f(ab’)-cd20-cd3双特异性抗体的原理图。
图3为分子筛纯化点击化学反应产物bi-2f(ab’)-cd20-cd3的结果图。
图4为sds-page分析分子筛纯化偶联产物bi-2f(ab’)-cd20-cd3的结果图,其中,泳道1和2为还原状态结果,泳道3和4位非还原状态结果,泳道1和3为偶联产物2f(ab’)-cd20-cd3样品,泳道2和4为未偶联产物fab-cd20-n3和fab-cd3-dbco混合样品。
图5为hplc分析偶联产物bi-2f(ab’)-cd20-cd3结果图。
图6为ramos细胞检测bi-2f(ab’)-cd20-cd3亲和力检测结果图,其中图a~e加入抗体情况分别为:阴性对照、fab-cd201μg/ml、fab-cd205μg/ml、bi-2f(ab’)-cd20-cd31μg/ml、bi-2f(ab’)-cd20-cd35μg/ml。
图7为pbmc细胞检测bi-2f(ab’)-cd20-cd3亲和力检测结果图,其中图a~e加入抗体情况分别为:阴性对照、fab-cd31μg/ml、fab-cd35μg/ml、bi-2f(ab’)-cd20-cd31μg/ml、bi-2f(ab’)-cd20-cd35μg/ml。
图8为bi-2f(ab’)-cd20-cd3体外杀伤实验结果图。
图9为bi-2f(ab’)-cd20-cd3和igg-cd20-cd3特异性验证结果图。
图10为流式分析bi-2f(ab’)-cd20-cd3对pbmc的特异性早期激活标志cd69流式图,其中,图a为阴性对照、图b为2000ng/ml的cd3抗体、图c~g分别为浓度为16、80、400、2000、10000ng/ml的bi-2f(ab’)-cd20-cd3抗体。
图11为流式分析bi-2f(ab’)-cd20-cd3对pbmc的特异性晚期激活标志cd25流式图,其中,图a为阴性对照、图b为2000ng/ml的cd3抗体、图c~g分别是浓度梯度依次为16、80、400、2000、10000ng/ml的bi-2f(ab’)-cd20-cd3抗体。
图12为流式分析bi-2f(ab’)-cd20-cd3对pbmc与cd20阴性细胞k562共孵育后t细胞增殖图。
图13为流式分析bi-2f(ab’)-cd20-cd3对pbmc与cd20阳性细胞ramos共孵育后t细胞增殖图,其中a~h分别是pbmc空白、pbmc-cfse空白、0、16、80、400、2000、10000ng/mlbi-2f(ab’)-cd20-cd3抗体。
图14为bi-2f(ab’)-cd20-cd3体外抗肿瘤活性实验结果图。
具体实施方式
志愿者新鲜外周血样本:与浙江大学校医院合作获得,经浙江大学药学院医学伦理委员会审批通过,编号:(2018)伦研批第(003)号。
实施例1
fab-cd20或fab-cd3的制备。
fab表示抗原结合片段,包括一条重链(h)和一条轻链(l),fab-cd20重链氨基酸序列如seqidno.1所示,轻链氨基酸序列如seqidno.2所示,重链编码基因序列如seqidno.3所示,轻链编码基因序列如seqidno.4所示。fab-cd3重链氨基酸序列如seqidno.5所示,轻链氨基酸序列如seqidno.6所示,重链编码基因序列如seqidno.7所示,轻链编码基因序列如seqidno.8所示。在本发明中,fab-cd20还在原ofa单抗的抗原结合片段的基础上做了改造,用于与药物进行偶联,fab-cd3还在原okt3单抗的抗原结合片段的基础上做了改造,用于与药物进行偶联,改造方式为在重链的ch1末端(c端)带有(ggggs)3-lpetgghhhhhh的多肽序列,(ggggs)3为柔性连接肽,lpetg为sortasea酶识别序列,6×his标签用于蛋白纯化。
ofa单抗和okt3单抗的抗原结合片段的轻链和重链编码基因序列由生工生物工程(上海)股份有限公司代为合成,然后用pcr方法完成上述改造。然后分别连接pmh3表达载体并瞬转至人胚肾细胞(293f)中,并用histraphp亲和柱对抗体进行纯化,并获得fab-cd20或fab-cd3单抗。
实施例2
双特异性抗体bi-2f(ab’)-cd20-cd3的制备,制备流程如图2所示。
1、fab-cd20-n3,fab-cd3-dbco偶联物的制备
(1)在10ml反应体系中,分别加入6μmfab单抗(溶于50mmtris-hcl,150mmnacl,ph7.4),用缓冲液(50mmtris-hcl,150mmnacl,ph7.4)溶解的150μmggg-peg4-dbco/ggg-peg3-n3(图1,由南京联宁生物技术有限公司合成),50mμsortasea,5mm的cacl2,于37℃水浴反应12h。
(2)用proteinl(1ml,ge)柱用来纯化反应液,洗脱液洗脱得到抗体偶联物,用1mtris调ph至7.0左右,10kdamillipore超滤管超滤浓缩并用pbs缓冲液置换,-20℃保存备用。fab-cd20与ggg-peg3-n3反应获得fab-cd20-n3,fab-cd3与ggg-peg4-dbco反应获得fab-cd3-dbco。
2、fab-cd20-n3,fab-cd3-dbco点击化学偶联反应及纯化
(1)在1ml的反应体系中(50mmtris-hcl,150mmnacl,ph7.4),fab-cd20-n3和fab-cd3-dbco按照摩尔比1∶1在4℃震荡反应过夜。
(2)用分子筛(superdex200increase10/300gl,ge)柱来纯化反应液,收集流出的第一个峰的的流出液如图3所示,为两者偶联后的产物,命名为bi-2f(ab’)-cd20-cd3,第二个峰为未偶联的fab-cd20-n3和fab-cd3-dbco(两者分子量接近),根据投料量(3mgfab-cd20-n3和3mgfab-cd20-n3)和回收到蛋白量(~2.7mg)可以计算得到回收率约为45%。由于蛋白分子之间空间位阻较大,使得偶联效率未能到达100%。目的产物用30kdamillipore超滤管超滤浓缩并用pbs缓冲液置换,-20℃保存备用。
(3)sds-page分析纯化产物如图4所示,在还原条件下有一条约50kda的条带(两条重链点击化学连在一起)和一条25kda的轻链的条带(两条轻链大小相近难以分开);在非还原条件下有一条约100kda的条带与理论值相符。
(4)hplc分析纯化产物的纯度(非还原条件下),如图5所示,蛋白纯度可以达到95%左右。
实施例3
流式亲和力测定。
(1)收集1×106个ramos细胞分别fab-cd20、bi-2f(ab’)-cd20-cd3(1μg/ml和5μg/ml)在4℃共孵育30min;同样的收集1×106个pbmc分别和fab-cd3和bi-2f(ab’)-cd20-cd3(1μg/ml和5μg/ml)在4℃共孵育30min;
(2)pbs洗涤三次后,每管加入抗人的鼠单抗fab(1∶1500稀释)200μl,并于4℃孵育30min;
(3)pbs洗涤三次后,每管加入fitc标记的山羊抗鼠的igg(h+l)(1∶250稀释)200μl,4℃孵育30min后,pbs洗涤3次,用流式细胞仪检测。
(4)流式细胞仪检测抗体与cd20阳性/cd3阳性的细胞的亲和力主要通过二抗标记后的fitc的平均荧光强度来表示。
(5)如图6和7所示酶促-化学偶联产物bi-2f(ab’)-cd20-cd3的亲和力明显高于单独的fab-cd20和fab-cd3,说明二价的2f(ab’)双特异性抗体亲和力和稳定性高于单价的fab抗体。
实施例4
体外活性测定。
(1)收集cd20阳性的ramos,daudi,raji细胞和cd20阴性的k562细胞,计数后用rpmi-1640(10%fbs)培养基重悬成细胞密度为1×106细胞悬液,分别以100μl/孔铺于12孔板中;
(2)抽取志愿者新鲜外周血,用人淋巴细胞分离液分离出其中的淋巴细胞pbmc后,pbs缓冲液洗两遍并计数后,用pbs重悬成1×107细胞悬液,并加入适量的cfse至5μm的终浓度。在37℃避光水浴15min后,pbs洗两遍并计数后用rpmi-1640(10%fbs)培养基重悬成细胞密度为1.25×106细胞悬液,分别以400μl/孔铺于(1)中的12孔板中;
(3)加入不同浓度(10000、2000、400、200、80、16、8ng/ml)的bi-2f(ab’)-cd20-cd3抗体,共孵育48h,收集细胞用pbs洗涤后,用100μl含有5μlannexinv、5μlpi重悬,室温孵育15min,补加400μl的结合液,流式细胞仪检测细胞凋亡百分比;
(4)从图8中的结果可以看出,bi-2f(ab’)-cd20-cd3抗体对cd20阴性的k562细胞没有明显的杀伤作用,而对cd20阳性的肿瘤细胞ramos、daudi和raji有浓度梯度依赖的特异性杀伤作用,且在8ng/ml的浓度就可以对细胞产生杀伤,体外杀伤肿瘤效果非常显著。
实施例5
体外特异性验证实验。
(1)选择表达fcrn受体的k562细胞作为研究对象,收集细胞后计数,用rpmi-1640(10%fbs)培养基重悬成细胞密度为1×106细胞悬液,分别以100μl/孔铺于12孔板中;
(2)抽取志愿者新鲜外周血,用人淋巴细胞分离液分离出其中的淋巴细胞pbmc后,pbs缓冲液洗两遍并计数后,用pbs重悬成1×107细胞悬液,并加入适量的cfse至5μm的终浓度。在37℃避光水浴15min后,pbs洗两遍并计数后用rpmi-1640(10%fbs)培养基重悬成细胞密度为1.25×106细胞悬液,分别以400μl/孔铺于(1)中的12孔板中;
(3)分别加入不同浓度(10000、100、10ng/ml)的bi-2f(ab’)-cd20-cd3抗体和对照抗体igg型的igg-cd20-cd3(由申请人根据knobintohole和crossmab技术构建得到的igg形式的抗体,该抗体的一条臂为抗cd20抗体,另一条臂为抗cd3抗体),共孵育48h,收集细胞用pbs洗涤后,用100μl含有5μlannexinv、5μlpi重悬,室温孵育15min,补加400μl的结合液,流式细胞仪检测细胞凋亡百分比;
(4)结果如图9所示,bi-2f(ab’)-cd20-cd3抗体在三个浓度下对表达fcrn的k562没有明显的杀伤作用,而igg-cd20-cd3在1000ng/ml的浓度下对表达fcrn的k562有着明显的杀伤作用,说明缺乏fc段的bi-2f(ab’)-cd20-cd3抗体特异性更好。
实施例6
pbmc特异性激活检测。
(1)收集cd20阳性的ramos细胞,计数后用rpmi-1640(10%fbs)培养基重悬成细胞密度为1×106细胞悬液,分别以100μl/孔铺于12孔板中;
(2)抽取志愿者新鲜外周血,用人淋巴细胞分离液分离出其中的淋巴细胞pbmc后,pbs缓冲液洗两遍并计数后,用rpmi-1640(10%fbs)培养基重悬成细胞密度为1.25×106细胞悬液,分别以400μl/孔铺于(1)中的12孔板中;
(3)加入不同浓度(10000、2000、400、80、16ng/ml)的bi-2f(ab’)-cd20-cd3抗体,选择2000ng/ml的cd3抗体作为对照组共孵育48h,收集细胞用pbs洗涤后,加入适当浓度的抗cd4-fitc,cd8-fitc,cd69-pe,cd25-apc抗体(抗体购自美国bd公司)4℃孵育30min。pbs洗涤两遍重悬后进行流式检测。
(4)结果如图10和11所示,cd25和cd69是t细胞激活的表面标志,单独的fab-cd3抗体在2000ng/ml的浓度下对t(cd4+cd8+)细胞有微弱的激活作用(较低的cd4+cd8+cd69+或者cd4+cd8+cd25+阳性比例),但是明显低于同浓度下的bi-2f(ab’)-cd20-cd3抗体激活t细胞的能力。说明bi-2f(ab’)-cd20-cd3抗体能够特异性的激活t细胞,且激活比例与给药浓度呈正相关。
实施例7
pbmc特异性增殖检测。
(1)收集cd20阳性的ramos细胞,计数后用rpmi-1640(10%fbs)培养基重悬成细胞密度为1×106细胞悬液,分别以100μl/孔铺于12孔板中;
(2)抽取志愿者新鲜外周血,用人淋巴细胞分离液分离出其中的淋巴细胞pbmc后,pbs缓冲液洗两遍并计数后,用pbs重悬成1×107细胞悬液,并加入适量的cfse至5μm的终浓度。在37℃避光水浴15min后,pbs洗两遍并计数后用rpmi-1640(10%fbs)培养基重悬成细胞密度为1.25×106细胞悬液,分别以400μl/孔铺于(1)中的12孔板中;
(3)加入不同浓度(10000、2000、400、200、80、16、8ng/ml)的bi-2f(ab’)-cd20-cd3抗体,共孵育48h,收集细胞用pbs洗涤并重悬后,流式细胞仪检细胞增殖;
(4)结果如图12和13所示,bi-2f(ab’)-cd20-cd3抗体与cd20阴性细胞系k562的共孵育时不能引起pbmc的增殖,但是cd20阳性的ramos细胞共孵育可以特异性的引起pbmc的增殖,且增殖的比例与给药浓度成正比。
实施例8
体内抗肿瘤活性实验。
(1)抽取志愿者新鲜外周血,用人淋巴细胞分离液分离出其中的淋巴细胞pbmc后,pbs缓冲液洗两遍并计数后,用pbs重悬成2×108细胞悬液;
(2)体外培养cd20阳性细胞ramos,离心收集,并用pbs洗涤两次,计数后用一定体积的pbs重悬配置成5×107cell/ml的细胞悬液;
(3)效应细胞pbmc悬液与靶细胞ramos悬液按照1∶1(v/v)的比例混合(effect:target4∶1),接种于scidbeige小鼠的右侧腋下。并将小鼠随机分为四组:saline对照组,cd3抗体对照组(3mg/kg),bi-2f(ab’)-cd20-cd3低剂量组(1mg/kg),bi-2f(ab’)-cd20-cd3高剂量组(3mg/kg)。接瘤24h后,开始给药每隔两天给药一次,共给药五次。
(4)结果如图14所示,saline对照组和和cd3抗体对照组中小鼠的肿瘤在逐渐长大,bi-2f(ab’)-cd20-cd3高剂量给药组(3mg/kg)肿瘤全部消失,bi-2f(ab’)-cd20-cd3低剂量给药组(1mg/kg)中有三只肿瘤全部消失,只有一只在缓慢生长。实验结果说明bi-2f(ab’)-cd20-cd3在低浓度的给药剂量下便能很好的抑制肿瘤的生长,具有较强的抗肿瘤活性。
序列表
<110>浙江大学
<120>一种双特异性抗体及其应用
<160>14
<170>siposequencelisting1.0
<210>1
<211>257
<212>prt
<213>人工序列(artificialsequence)
<400>1
gluvalglnleuvalgluserglyglyglyleuvalglnproglyarg
151015
serleuargleusercysalaalaserglyphethrpheasnasptyr
202530
alamethistrpvalargglnalaproglylysglyleuglutrpval
354045
serthrilesertrpasnserglyserileglytyralaaspserval
505560
lysglyargphethrileserargaspasnalalyslysserleutyr
65707580
leuglnmetasnserleuargalagluaspthralaleutyrtyrcys
859095
alalysaspileglntyrglyasntyrtyrtyrglymetaspvaltrp
100105110
glyglnglythrthrvalthrvalserseralaserthrlysglypro
115120125
servalpheproleualaproserserlysserthrserglyglythr
130135140
alaalaleuglycysleuvallysasptyrpheprogluprovalthr
145150155160
valsertrpasnserglyalaleuthrserglyvalhisthrphepro
165170175
alavalleuglnserserglyleutyrserleuserservalvalthr
180185190
valproserserserleuglythrglnthrtyrilecysasnvalasn
195200205
hislysproserasnthrlysvalasplyslysvalgluprolysser
210215220
cysasplysthrhisthrglyglyglyglyserglyglyglyglyser
225230235240
glyglyglyglyserleuprogluthrglyglyhishishishishis
245250255
his
<210>2
<211>214
<212>prt
<213>人工序列(artificialsequence)
<400>2
gluilevalleuthrglnserproalathrleuserleuserprogly
151015
gluargalathrleusercysargalaserglnservalsersertyr
202530
leualatrptyrglnglnlysproglyglnalaproargleuleuile
354045
tyraspalaserasnargalathrglyileproalaargphesergly
505560
serglyserglythraspphethrleuthrileserserleuglupro
65707580
gluaspphealavaltyrtyrcysglnglnargserasntrpproile
859095
thrpheglyglnglythrargleugluilelysargthrvalalaala
100105110
proservalpheilepheproproseraspgluglnleulyssergly
115120125
thralaservalvalcysleuleuasnasnphetyrproarggluala
130135140
lysvalglntrplysvalaspasnalaleuglnserglyasnsergln
145150155160
gluservalthrgluglnaspserlysaspserthrtyrserleuser
165170175
serthrleuthrleuserlysalaasptyrglulyshislysvaltyr
180185190
alacysgluvalthrhisglnglyleuserserprovalthrlysser
195200205
pheasnargglyglucys
210
<210>3
<211>774
<212>dna
<213>人工序列(artificialsequence)
<400>3
gaagtgcagctggtggagtctgggggaggcttggtacagcctggcaggtccctgagactc60
tcctgtgcagcctctggattcacctttaatgattatgccatgcactgggtccggcaagct120
ccagggaagggcctggagtgggtctcaactattagttggaatagtggttccataggctat180
gcggactctgtgaagggccgattcaccatctccagagacaacgccaagaagtccctgtat240
ctgcaaatgaacagtctgagagctgaggacacggccttgtattactgtgcaaaagatata300
cagtacggcaactactactacggtatggacgtctggggccaagggaccacggtcaccgtc360
tcctcagctagcaccaagggcccatcggtcttccccctggcaccctcctccaagagcacc420
tctgggggcacagcggccctgggctgcctggtcaaggactacttccccgaaccggtgacg480
gtgtcgtggaactcaggcgccctgaccagcggcgtgcacaccttcccggctgtcctacag540
tcctcaggactctactccctcagcagcgtggtgaccgtgccctccagcagcttgggcacc600
cagacctacatctgcaacgtgaatcacaagcccagcaacaccaaggtggacaagaaagtt660
gagcccaaatcttgtgacaaaactcacacaggcggtggaggctctggaggcgggggttcc720
ggtggcggaggttcactgccggagactggtggtcatcaccaccatcaccattaa774
<210>4
<211>645
<212>dna
<213>人工序列(artificialsequence)
<400>4
gaaattgtgttgacacagtctccagccaccctgtctttgtctccaggggaaagagccacc60
ctctcctgcagggccagtcagagtgttagcagctacttagcctggtaccaacagaaacct120
ggccaggctcccaggctcctcatctatgatgcatccaacagggccactggcatcccagcc180
aggttcagtggcagtgggtctgggacagacttcactctcaccatcagcagcctagagcct240
gaagattttgcagtttattactgtcagcagcgtagcaactggccgatcaccttcggccaa300
gggacacgactggagattaaacgtacggtggctgcaccatctgtcttcatcttcccgcca360
tctgatgagcagttgaaatctggaactgcctctgttgtgtgcctgctgaataacttctat420
cccagagaggccaaagtacagtggaaggtggataacgccctccaatcgggtaactcccag480
gagagtgtcacagagcaggacagcaaggacagcacctacagcctcagcagcaccctgacg540
ctgagcaaagcagactacgagaaacacaaagtctacgcctgcgaagtcacccatcagggc600
ctgagctcgcccgtcacaaagagcttcaacaggggagagtgttag645
<210>5
<211>254
<212>prt
<213>人工序列(artificialsequence)
<400>5
glnvalglnleuglnglnserglyalagluleualaargproglyala
151015
servallysmetsercyslysalaserglytyrthrphethrargtyr
202530
thrmethistrpvallysglnargproglyglnglyleuglutrpile
354045
glytyrileasnproserargglytyrthrasntyrasnglnlysphe
505560
lysasplysalathrleuthrthrasplysserserserthralatyr
65707580
metglnleuserserleuthrsergluaspseralavaltyrtyrcys
859095
alaargtyrtyraspasphistyrcysleuasptyrtrpglyglngly
100105110
thrthrleuthrvalserseralaserthrlysglyproservalphe
115120125
proleualaproserserlysserthrserglyglythralaalaleu
130135140
glycysleuvallysasptyrpheprogluprovalthrvalsertrp
145150155160
asnserglyalaleuthrserglyvalhisthrpheproalavalleu
165170175
glnserserglyleutyrserleuserservalvalthrvalproser
180185190
serserleuglythrglnthrtyrilecysasnvalasnhislyspro
195200205
serasnthrlysvalasplyslysvalgluprolyssercysasplys
210215220
thrhisthrglyglyglyglyserglyglyglyglyserglyglygly
225230235240
glyserleuprogluthrglyglyhishishishishishis
245250
<210>6
<211>214
<212>prt
<213>人工序列(artificialsequence)
<400>6
glnilevalleuthrglnserproalailemetseralaserprogly
151015
glulysvalthrmetthrcysseralaserserservalsertyrmet
202530
asntrptyrglnglnlysserglythrserprolysargtrpiletyr
354045
aspthrserlysleualaserglyvalproalahispheargglyser
505560
glyserglythrsertyrserleuthrileserglymetglualaglu
65707580
aspalaalathrtyrtyrcysglnglntrpserserasnprophethr
859095
pheglyserglythrlysleugluileasnargargthrvalalaala
100105110
proservalpheilepheproproseraspgluglnleulyssergly
115120125
thralaservalvalcysleuleuasnasnphetyrproarggluala
130135140
lysvalglntrplysvalaspasnalaleuglnserglyasnsergln
145150155160
gluservalthrgluglnaspserlysaspserthrtyrserleuser
165170175
serthrleuthrleuserlysalaasptyrglulyshislysvaltyr
180185190
alacysgluvalthrhisglnglyleuserserprovalthrlysser
195200205
pheasnargglyglucys
210
<210>7
<211>765
<212>dna
<213>人工序列(artificialsequence)
<400>7
caggtccagctgcagcagtctggggctgaactggcaagacctggggcctcagtgaagatg60
tcctgcaaggcttctggctacacctttactaggtacacgatgcactgggtaaaacagagg120
cctggacagggtctggaatggattggatacattaatcctagccgtggttatactaattac180
aatcagaagttcaaggacaaggccacattgactacagacaaatcctccagcacagcctac240
atgcaactgagcagcctgacatctgaggactctgcagtctattactgtgcaagatattat300
gatgatcattactgccttgactactggggccaaggcaccactctcacagtctcctcagct360
agcaccaagggcccatcggtcttccccctggcaccctcctccaagagcacctctgggggc420
acagcggccctgggctgcctggtcaaggactacttccccgaaccggtgacggtgtcgtgg480
aactcaggcgccctgaccagcggcgtgcacaccttcccggctgtcctacagtcctcagga540
ctctactccctcagcagcgtggtgaccgtgccctccagcagcttgggcacccagacctac600
atctgcaacgtgaatcacaagcccagcaacaccaaggtggacaagaaagttgagcccaaa660
tcttgtgacaaaactcacacaggcggtggaggctctggaggcgggggttccggtggcgga720
ggttcactgccggagactggtggtcatcaccaccatcaccattaa765
<210>8
<211>645
<212>dna
<213>人工序列(artificialsequence)
<400>8
caaattgttctcacccagtctccagcaatcatgtctgcatctccaggggagaaggtcacc60
atgacctgcagtgccagctcaagtgtaagttacatgaactggtaccagcagaagtcaggc120
acctcccccaaaagatggatttatgacacatccaaactggcttctggagtccctgctcac180
ttcaggggcagtgggtctgggacctcttactctctcacaatcagcggcatggaggctgaa240
gatgctgccacttattactgccagcagtggagtagtaacccattcacgttcggctcgggg300
acaaagttggaaataaaccggcgtacggtggctgcaccatctgtcttcatcttcccgcca360
tctgatgagcagttgaaatctggaactgcctctgttgtgtgcctgctgaataacttctat420
cccagagaggccaaagtacagtggaaggtggataacgccctccaatcgggtaactcccag480
gagagtgtcacagagcaggacagcaaggacagcacctacagcctcagcagcaccctgacg540
ctgagcaaagcagactacgagaaacacaaagtctacgcctgcgaagtcacccatcagggc600
ctgagctcgcccgtcacaaagagcttcaacaggggagagtgttag645
<210>9
<211>5
<212>prt
<213>人工序列(artificialsequence)
<400>9
leuprogluthrgly
15
<210>10
<211>5
<212>prt
<213>人工序列(artificialsequence)
<400>10
glyglyglyglyser
15
<210>11
<211>10
<212>prt
<213>人工序列(artificialsequence)
<400>11
glyglyglyglyserglyglyglyglyser
1510
<210>12
<211>15
<212>prt
<213>人工序列(artificialsequence)
<400>12
glyglyglyglyserglyglyglyglyserglyglyglyglyser
151015
<210>13
<211>3
<212>prt
<213>人工序列(artificialsequence)
<400>13
glyglygly
1
<210>14
<211>27
<212>prt
<213>人工序列(artificialsequence)
<400>14
glyglyglyglyserglyglyglyglyserglyglyglyglyserleu
151015
progluthrglyglyhishishishishishis
2025
- 还没有人留言评论。精彩留言会获得点赞!
- 用于治疗HBV感染和相关病症的三特异性结合分子的制造方法与工艺
- 对磺胺类药物具有特异性的固相微萃取纤维的制备方法与流程
- 一种检测猪初乳中抗PEDV特异性IgA抗体的夹心ELISA试剂盒的制造方法与工艺
- 用于施用CD19xCD3双特异性抗体的给药方案的制造方法与工艺
- 双特异性抗‑VEGF/抗‑ANG‑2抗体及其在治疗眼血管疾病中的应用的制造方法与工艺
- 特异性人EWS‑FLI1融合蛋白抗体的制备方法及在制备尤文肉瘤诊断抗体试剂的应用与制造工艺
- 双特异性四价抗体及其制造和使用方法与制造工艺
- 利用特异性挥发物筛选昆虫相关嗅觉蛋白的方法与制造工艺
- 使用与表面偶联的单特异性四聚抗体复合物组合物从样品中分离出靶实体的方法与制造工艺
- 抗-WT1/HLA双特异性抗体的制造方法与工艺