用于治疗HBV感染和相关病症的三特异性结合分子的制作方法
本发明涉及包含至少三种结合特异性的结合分子,其中(a)第一特异性是针对选自hbv小表面抗原、hbv中表面抗原和hbv大表面抗原的乙型肝炎病毒(hbv)表面抗原;(b)(i)第二和第三特异性分别针对cd3和cd28;或(ii)第二和第三特异性选自针对cd16、cd56、nkp30、nkp46、4-1bb和nkg2d的特异性;(c)每种结合特异性由一个或更多个结合位点提供,每个结合位点由以下独立地提供:(i)六个互补决定区(cdr)的组,其中所述六个cdr的组由三个cdr的第一组和三个cdr的第二组构成,其中所述第一组和所述第二组各自被包含在免疫球蛋白结构域中;或(ii)三个cdr的组,其中所述三个cdr的组被包含在免疫球蛋白结构域中。
在本说明书中,引用了包括专利申请和制造商手册在内的许多文件。虽然不认为这些文件的公开内容与本发明的可专利性相关,但是在此通过引用将其整体地并入。更具体地,所有引用的文献均通过引用并入,就像每个单独的文献被具体和单独地指出要通过引用并入一样。
约有3.5亿人慢性地感染有乙型肝炎病毒(hbv)。hbv感染可能引起肝硬化和肝细胞癌(hcc),其每年造成约一百万人伤亡(ganem等人,hepatitisbvirusinfection—naturalhistoryandclinicalconsequences.nengljmed;350:1118-29(2004))。hbv感染在约5%的成人患者和约90%的新生儿中目前无法控制。在这种情况下,hbv感染变为慢性的。可能的原因是细胞免疫反应不足。目前可用的用于治疗hbv感染的抗病毒药物抑制病毒复制。然而,共价闭合的环状dna(cccdna)保留在受感染的肝细胞的核中,并且一旦患者停止服用药物,可能导致hbv感染的再激活。因此,如果要完全治愈感染,消除携带所述cccdna的hbv感染细胞是必不可少的(protzer等人,natimmunolrev12:2013-213(2012))。
然而,hbv感染细胞的这种细胞毒性消除(通过细胞毒性t淋巴细胞或自然杀伤(nk)细胞)不发生或发生的程度不足。
携带hbvcccdna的感染细胞在它们的表面上展示病毒表面蛋白。尽管病毒被释放到细胞内囊泡中,但由于许多hbv表面蛋白仍然整合到内质网的细胞内膜中,所以推测发生这种情况。在囊泡运输过程中,所述细胞内膜可能与细胞膜融合,其结果是hbv表面蛋白在被感染细胞的表面上展示。
bohne等人(tcellsredirectedagainsthepatitisbvirussurfaceproteinseliminateinfectedhepatocytes.gastroenterology;134:239-247(2008))和krebs等人(tcellsexpressingachimericantigenreceptorthatbindshepatitisbvirusenvelopeproteinscontrolvirusreplicationinmice.gastroenterology(2013))描述了嵌合抗原受体,当其被逆转录病毒递送并在t细胞表面上表达时,使原代人类和小鼠t细胞能够识别展示hbv小表面抗原的肝细胞并裂解hbv复制的细胞。
ep2524699a1描述了三功能抗体。这些抗体“具有功能性fc部分”并且“必须由不同亚类的免疫球蛋白重链组成”。另一方面,hornigund
liao等人(oncologyreports3,637-644(1996))描述了用于裂解裸鼠中的人类肝癌异种移植物的再靶向效应细胞的双特异性单克隆抗体。所描述的双特异性抗体通过两个杂交瘤的融合产生,得到表达两种不同抗体的重/轻链组合的杂交瘤细胞系。这可能导致两种不同重链的配对,也可能导致相同重链的配对,引起单特异性亲本抗体和双特异性抗体的随机混合物。双特异性抗体含有重链和轻链并二聚以形成ig分子。
鉴于现有技术,在提供治疗hbv感染以及由hbv感染引起的病症如肝硬化或肝细胞癌的替代或改进的手段和方法中可以看到技术问题。在提供治愈hbv感染的方法中也可以看到待解决的技术问题。从细胞生物学的角度来说,在提供根除携带有hbvcccdna的细胞的手段和方法中可以看到技术问题。该技术问题由所附的权利要求解决。
因此,本发明在第一方面涉及包含至少三种结合特异性的结合分子,其中(a)第一特异性是针对选自hbv小表面抗原、hbv中表面抗原和hbv大表面抗原的乙型肝炎病毒(hbv)表面抗原;(b)(i)第二和第三特异性分别针对cd3和cd28;或(ii)第二和第三特异性是选自针对cd16、cd56、nkp30、nkp46、4-1bb和nkg2d的特异性;(c)每种结合特异性由一个或更多个结合位点提供,每个结合位点由以下独立地提供:(i)六个互补决定区(cdr)的组,其中所述六个cdr的组由三个cdr的第一组和三个cdr的第二组构成,其中所述第一组和所述第二组各自被包含在免疫球蛋白结构域中;或(ii)三个cdr的组,其中所述三个cdr的组被包含在免疫球蛋白结构域中。
“hbv表面抗原”也缩写为“hbs”或“hbsag”。
根据本发明的结合分子的结合能力通过使用术语“结合特异性”在功能上限定。提供所述结合特异性的结构要素被称为“结合位点”。关于每个所提及的特异性设想的是在根据本发明的结合分子中存在提供给定特异性的一个或多个结合位点。如果根据本发明的结合分子中通过多于一个结合位点实现给定结合特异性,则应理解,优选的是实现给定结合特异性的结合位点在结构和功能方面上是相同的,功能是识别待结合的抗原的给定表位。然而这并不排除其中给定结合特异性由多个结合位点实现的实施方案,所述结合位点虽然与相同的抗原结合,但经由抗原的不同表位来结合。这通常还需要结合位点之间的结构差异,尽管如此,总体而言,它们都具有结合相同抗原的能力。
根据提供给定特异性的结合位点的精确数目,示例性数目是1、2、3和4个,优选2个,可以很好地调整或优化整体结合性质。通常,通过增加提供给定特异性的结合位点的数目,可以改善结合动力学以及结合热力学。
结合位点的总数在本领域中也称为“价”。因此,虽然对于三特异性的结合分子需要至少是三价的,但应理解根据本发明的结合分子也可以是四价、五价、六价、七价或八价的。还设想更高的价。预期还通过使用九价、十价的结合分子或具有更高价数的结合分子实现携带hbvcccdna的细胞的根除。
对于将是至少三特异性的结合分子,每种结合特异性恰好由一个结合位点表示就足够了。因此,根据本发明的三特异性结合分子可以是三价分子。一般而言,优选的是其中一个、两个或三个结合特异性各自用两个结合位点实现的那些三特异性分子。还优选的是其中一个、两个或三个结合特异性各自用三个结合位点实现的那些三特异性分子。
三种结合特异性也可以各自由不同数目的结合位点实现。可以优选的是一种结合特异性由比另外两种结合特异性更多的结合位点表示,或者两种结合特异性可以由比第三种结合特异性更多的结合位点表示。
根据第一方面的(c)(i)项明显的是,六个cdr的组是给定结合位点的优选实施方案。如本领域众所周知的,六个cdr的组包含第一组和第二组,每组由三个cdr构成。三个cdr的这两组可以位于同一多肽链上或不同的多肽链上,从而产生不同的分子结构。另外,设想在根据本发明的给定结合分子内,部分结合位点是这样的:构成给定结合位点的全部六个cdr位于单个多肽链上,而其他结合位点是这样的:构成给定结合位点的第一组的和第二组的三个cdr位于不同的多肽上。不同的多肽可以仅仅是形式上不同的,在其他方面是相同的,尤其是对于氨基酸序列。或者,不同的多肽在其氨基酸序列方面可以是彼此不同的。
一般而言,根据以下优选实施方案的讨论也会明显的是,根据本发明的结合分子可以仅表现出由一条多肽形成的结合位点。然而,应理解这并不等于要求结合分子作为整体由单个多肽链构成。另一方面,本发明扩展至其中全部结合位点由两条多肽链形成的结合分子。类似地,后者的叙述并不等于要求结合分子作为整体仅由两条多肽链构成。相反,结合分子也可以由多于两条多肽链,例如三条或四条或多于四条多肽链构成。最后,如上所述,并且如由以下结构(iii)举例说明的,本发明的结合分子包含的结合位点的一部分可以是这样的:它们中的每一个由两条多肽链形成,而结合位点的其余部分是这样的:它由单个多肽链形成。
关于项(b)(ii),应理解第二和第三特异性彼此不同,从而提供至少三特异性的结合分子。
如果考虑到(c)(i)项,根据六个cdr的要求可知,需要两个免疫球蛋白结构域以形成单个结合位点。这不同于(c)(ii)项,其中单个免疫球蛋白结构域足以提供结合位点,在这种情况下结合位点由三个cdr构成。这也被称为单域结合位点。
术语“互补决定区”,缩写为“cdr”,具有如本领域所确定的含义。这些是短的子序列,通常约3至约25个氨基酸,其赋予抗体特异性识别抗原表位的能力。通常,抗体轻链可变区提供三个cdr,抗体重链可变区提供三个cdr。如果没有不同规定,限定结合位点的六个cdr通常按以下顺序存在:重链cdr1、重链cdr2、重链cdr3、轻链cdr1、轻链cdr2、轻链cdr3。虽然cdr通常是免疫球蛋白结构域的一部分,但根据本发明在这方面无要求。包含所述cdr的氨基酸序列就足够了,条件是所述氨基酸序列在生理条件下折叠时以空间接近的方式呈现所述cdr并保持其识别同源抗原的能力。提及的空间接近和抗原结合的能力也可以被称为“配置为结合抗原”。术语“免疫球蛋白结构域”是本领域已知的,并且是指具有7至9个反平行β-链的2层夹心三维结构的通常为70至100个氨基酸的序列。在限定一些实施方案的结构以及图中,单个免疫球蛋白结构域用椭圆形描绘。
六个cdr的组通常限定结合位点。在替代方案中,如从骆驼科和软骨鱼类抗体中已知的,三个cdr的组(cdr1、cdr2、cdr3)可以限定结合位点。如本领域已知的,尽管没有要求,但是优选三个cdr的这种组,其中cdr3由比在天然存在的免疫球蛋白可变区中更突出的肽环组成。
术语“抗原”具有其本领域确定的含义。它是指被通常由免疫球蛋白结构域呈现的六个cdr的组特异性地识别和结合的分子。被所述cdr识别和结合的抗原的特定部分也称为表位。
hbv小/中/大表面蛋白是hbv外包膜中的小表面蛋白、中表面蛋白、大表面蛋白(stibbe,w.,和w.h.gerlich,structuralrelationshipsbetweenminorandmajorproteinsofhepatitisbsurfaceantigen.j.virol.198346:626-628)。
三种hbv表面抗原从一个开放阅读框转录和翻译,并且彼此在n末端部分的长度上不同。因此,大表面抗原包含在中表面抗原和小表面抗原中都不存在的部分,而中表面抗原包含大抗原中包含但小抗原中不包含的部分。小抗原由包含在中抗原和大抗原的c末端部分的序列组成。
大hbv表面抗原可以以两种方式插入细胞质膜中。n末端或c末端可以位于细胞外侧。两种构型都在hbv感染的细胞中发现。换言之,大hbv表面抗原的n末端可以位于er膜的细胞质侧或腔侧,最终也可以以这些构型存在于hbv颗粒的外包膜和受感染肝细胞的质膜中。
出人意料地,将根据(b)的第二和第三特异性组合到三特异性分子中提供比两种双特异性分子的组合显著更强的免疫效应细胞的活化。参考实施例。
在进一步优选的实施方案中,提供对hbv表面抗原的结合特异性的结合位点结合位于(a)所述hbv小表面抗原中的表位;或(b)所述hbv小表面抗原中不包含的所述hbv大表面抗原的那部分中的表位;或(c)部分所述hbv大表面抗原中的表位,该部分与所述hbv小表面抗原在结构上不同。
(a)项是指存在于hbv小表面抗原中的表位。由于上述hbv小表面抗原、hbv中表面抗原、hbv大表面抗原之间的关系,小抗原的整个序列被包含在中抗原和大抗原中。通常,但并非必然地,由小表面抗原呈现的三维表位也会由中表面抗原和/或大表面抗原呈现。
根据(b)项,优选所述hbv大表面抗原的所述部分也不被所述hbv中表面抗原所包含。关于(c)项,应理解,“结构上不同”包括所述hbv大表面抗原的表位,其包含或由所述hbv小表面抗原序列的一部分组成,其中所述表位不存在于所述hbv小表面抗原上。根据(c)项,更优选的是所述表位是在所述hbv大表面抗原的一部分中,该部分与所述hbv中表面抗原在结构上也不同。
所述(a)项,结合本发明的所有方面和实施方案,特别优选的是所述第一抗原为所述hbv小表面抗原。
根据(b)和(c)项,多肽或结合分子会分别特异性识别hbv的大表面抗原。
第二和第三特异性针对由免疫效应细胞呈递、优选由nk细胞和/或ctl特异性地呈递的表面抗原。免疫效应细胞是对hbv感染细胞重定向的细胞,所述hbv感染细胞在其表面上呈现所述hbv表面抗原。
特别优选的是第二和第三特异性都针对ctl,或都针对nk细胞。
cd3(分化群3)t细胞共受体是蛋白质复合物,其由四条不同的链组成。在哺乳动物中,复合物含有cd3γ链、cd3δ链和两个cd3ε链。结合特异性可以针对这些链中的任何一个。这些链与被称为t细胞受体(tcr)和δ链的分子缔合以在t淋巴细胞中产生活化信号。tcr、δ链和cd3分子一起构成了tcr复合物。
在优选的实施方案中,cd3代表cd3ε链,其为cd3-t细胞受体复合物的一部分。(borst,j.等人,thedelta-andepsilon-chainsofthehumant3/t-cellreceptorcomplexaredistinctpolypeptides.nature.1984.312:455-458)。优选地,根据本发明的cd3的结合特异性是针对所述cd3ε链的。
cd28是主要的t细胞共刺激受体。(lesslauer,w.等人,t90/44(9.3抗原)。acellsurfacemoleculewithafunctioninhumantcellactivation.eur.j.immunol.1986.16:1289-1296)。
4-1bb(cd137)是活化的t细胞和nk细胞的共刺激受体。(kwon,b.s.等人,cdnasequencesoftwoinduciblet-cellgenes.proc.natl.acad.sci.u.s.a.1989.86:1963-1967)。
cd16(fcγriiia)是低亲和力fc受体,其由nk细胞、活化的细胞毒性t细胞的亚群以及来自髓单核细胞系的细胞类型表达,与igg分子的fc结构域结合。(lanier,l.l.等人,functionalpropertiesofauniquesubsetofcytotoxiccd3+tlymphocytesthatexpressfcreceptorsforigg(cd16/leu-11antigen).j.exp.med.1985,162:2089-2106)。
cd56(ncam)是由nk细胞表达的细胞黏附分子。(lanier,l.l.等人,identityofleu-19(cd56)leukocytedifferentiationantigenandneuralcelladhesionmolecule.j.exp.med.1989.169:2233-2238)。
nkg2d是由nk细胞表达的活化性受体(houchins,j.等人,dnasequenceanalysisofnkg2,afamilyofrelatedcdnaclonesencodingtypeiiintegralmembraneproteinsonhumannaturalkillercells.1991.j.exp.med.173:1017-1020)。
nkp30(ncr3)是由nk细胞表达的活化性受体(pende,d.等人,identificationandmolecularcharacterizationofnkp30,anoveltriggeringreceptorinvolvedinnaturalcytotoxicitymediatedbyhumannaturalkillercells.2000.j.exp.med.192:337-346)。
nkp46(ncr1)是由nk细胞表达的活化性受体(pessino等人,molecularcloningofnkp46:anovelmemberoftheimmunoglobulinsuperfamilyinvolvedintriggeringofnaturalcytotoxicity.1998.j.exp.med.188:953-960)。
cd3、cd28和4-1bb存在于ctl的表面上。本发明的多肽与这些表面抗原中任何一种的结合引起ctl的刺激或共刺激。
cd16、cd56、nkg2d、nkp30、nkp46和4-1bb存在于nk细胞的表面上。本发明的多肽与这些表面抗原中任何一种的结合引起nk细胞的刺激或共刺激。
对于人类ctls,优选cd3和cd28。对于人类nk细胞,优选cd16和cd56。
所提及的表面抗原以本领域确定的名称命名(还参见kennethmurphy,janeway'simmunobiology,第7版,garlandscience;williame.paul,fundamentalimmunology,第7版,lippincottwilliams&wilkins)。
特别优选根据本发明的结合、特别是cdr和抗原之间的结合是特异性的。如根据本发明所使用的术语“特异性结合”(具有与“特异性相互作用”相同的含义)指这些结合部分不与或基本上不与跟目标抗原结构相似的表位或结构交叉反应。正在研究的一组分子的交叉反应性可以例如通过评估所述分子组在常规条件下与感兴趣的表位以及一些或多或少(结构和/或功能上)密切相关的表位的结合来测试。只有与感兴趣的表位在其相关情况下(例如蛋白质结构中的特定模序)结合,但与任何其他表位不结合或基本上不结合的那些分子才被认为对感兴趣的表位是特异性的。
慢性hbv感染的特征在于免疫耐受状态。更具体地,患者的ctl和nk细胞是这样运行的:不发生受感染细胞的完全根除或hbv复制的完全控制或hbv的完全消除。根据本发明的结合分子一方面特异性地识别hbv表面抗原,另一方面特异性地识别免疫效应细胞表面抗原。本发明的结合分子可被认为赋予免疫效应细胞人工特异性。事实上,本发明的结合分子使ctl和nk细胞重新靶向,如此使得它们被募集至hbv感染细胞并杀伤它们。
一方面,本发明的结合分子与hbv感染细胞的结合以及另一方面免疫效应细胞的募集可以以任何顺序发生或同时发生。
特别地,旨在通过注射或作为口服施用形式全身地施用本发明的结合分子,并允许它们结合受hbv感染或表达hbv抗原的靶细胞并将所述免疫效应细胞募集至所述靶细胞。
虽然这样说,但还设想使本发明的结合分子与免疫效应细胞(或包含所述效应细胞的外周血单核细胞群)接触,如此使得所述效应细胞负载有所述结合分子。已经在体外或离体进行了负载的这种效应细胞(或包含这种经负载的效应细胞的pbmc群)然后可以施用于患有hbv感染或与之相关的和以下定义的病症的患者。这种施用可以是血管内施用,例如施用至肝动脉。具有根据本发明的结合分子的免疫效应细胞与所述免疫效应细胞的表面抗原结合也是本发明的一个方面。下文进一步公开该方面。
这种杀伤,特别是与免疫细胞分泌的抗病毒免疫介质(例如细胞因子)结合的杀伤提供hbv感染的根除或hbv感染的持续控制或表达hbv表面抗原的肿瘤细胞的消除。
本发明的结合分子性能的优选测量包括细胞培养物中t细胞介导的细胞毒性、t细胞至肝脏的募集、肝脏的炎症反应、血清中病毒滴度的降低、血清中病毒抗原hbsag和/或hbeag的减少、肝脏中病毒复制的减少、受感染细胞的数量的减少。
这些性质中的每一个可以通过本领域确定的流程来测量。
为了进一步解释,t细胞至肝脏的募集可以通过经由rt-pcr确定在肝脏中一种或更多种t细胞标记基因mrna,特别是cd3、cd4、cd8、ifnγ和ip-10(cxcl10)的水平来定量。根据本发明,这些标记基因的诱导与阴性对照相比被显著地诱导,并且可以达到2倍至1000倍或以上的诱导,优选大于2倍、优选大于5倍、优选大于10倍、优选大于25倍、优选大于50倍、优选大于100倍、优选大于500倍、优选大于750倍、优选大于1000倍。
根据本发明,细胞培养物中t细胞介导的细胞毒性为5%至100%、优选大于5%、优选大于10%、优选大于20%、优选大于30%、优选大于40%、优选大于50%、优选大于75%、优选100%。
体内t细胞介导的细胞毒性可以例如通过测量被消除的肝细胞释放的肝转氨酶(优选丙氨酸转氨酶(alt)和/或天冬氨酸转氨酶(ast))来评估。个体中正常的alt水平低于40国际单位每升(u/l),并且通过免疫效应细胞的重新靶向在有效的细胞毒性时可以达到50u/l至5000u/l的水平。优选地,水平会是大于50u/l、优选大于100u/l、优选大于200u/l、优选大于300u/l、优选大于400u/l、优选大于500u/l、优选大于750u/l、优选大于1000u/l。
根据本发明,可以通过he染色的方式确定肝脏的炎症反应。he代表苏木精和曙红,并且是病理学的标准染色技术,其使得细胞结构以蓝色、紫色、粉红色和橙色的阴影显示。利用该方法,与阴性对照比较可以看到浸润的免疫效应细胞并进行定量,并且根据本发明,与未经处理的对照相比数量会增加5倍至5000倍、优选大于5倍、优选大于10倍、优选大于20倍、优选大于50倍、优选大于100倍、优选大于250倍、优选大于500倍、优选大于1000倍。
从肝脏采集的活组织检查的cd3染色和/或本发明的三特异性结合分子的染色分别显示t细胞和重定向t细胞的特异性浸润。利用该方法,与阴性对照相比,可以看到重定向的cd3+t细胞并进行定量,并且根据本发明,与未经处理的对照相比数量会增加5倍至5000倍、优选大于5倍、优选大于10倍、优选大于20倍、优选大于50倍、优选大于100倍、优选大于250倍、优选大于500倍、优选大于1000倍。
在血清或肝组织样品上通过使用pcr和/或southern印迹法可以定量病毒复制。本发明的结合分子应用于hbv感染的对象引起血清中病毒滴度的降低。根据本发明,设想在施用有效量的结合分子后,实现血清中病毒滴度减少大于10%、优选大于20%、优选大于30%、优选大于40%、优选大于50%、优选大于60%、优选大于70%、优选大于80%、优选大于90%,优选低于检测水平(每毫升50个hbvdna拷贝)。例如,血清hbvdna在hbv转基因小鼠中达到最高每毫升109个拷贝的水平,并且在用重定向的t细胞处理后可以降低至每毫升107个拷贝(krebs等人,tcellsexpressingachimericantigenreceptorthatbindshepatitisbvirusenvelopeproteinscontrolvirusreplicationinmice.gastroenterology(2013))。
通过针对hbeag、hbcag或hbsag的染色肝切片,可以确定仍受感染的细胞数量。与未经处理的对照相比,根据本发明的hbv复制细胞的减少为最小5%至最大100%、优选大于5%、优选大于10%、优选大于20%、优选大于30%、优选大于50%、优选大于70%、优选大于90%、优选100%。
“病毒滴度”(也称为病毒载量、病毒负载量)是本领域众所周知的术语,其指示给定生物样品中病毒的量。病毒的量由各种量度指示,包括但不限于病毒核酸的量、病毒颗粒(如通过沉降梯度离心确定的完整病毒体)和亚病毒颗粒(例如通过免疫测定如elisa确定的hbsag或乙型肝炎表面抗原颗粒)的存在。通常,对于液体样品例如血液和尿液,确定每单位液体,例如每毫升的病毒量。对于固体样品例如组织样品,确定每重量单位、例如每克的病毒量。用于确定病毒的量的方法是本领域已知的。目前,例如可用于乙型肝炎病毒的常规测试是例如病毒载量、定性和定量hbsag和hbeag测量。
“有效量”是指足以实现上述目的的试剂的量。具体地,根据本发明的结合分子的量在待治疗的受乙型肝炎病毒感染的对象中减少病毒滴度水平是有效的。有效量可以以一次或更多次施用来施用。有效量可以根据预期的应用(体外或体内)或被治疗的对象和疾病病症,例如对象的体重和年龄、疾病病症的严重程度、施用方式等变化,这可以由本领域普通技术人员容易地确定。
例如有许多不同的使用核酸测试(nat)来定量病毒载量的基于分子的测试方法。用于扩增的起始材料可以用于将这些分子方法分成组。体外dna合成的聚合酶链反应(pcr)方法使用dna模板、聚合酶、缓冲液、引物和核苷酸来使血液样品中的hiv扩增。然后化学反应标记病毒。测量标记物并用于计算病毒的量。pcr用于定量整合的dna。
逆转录聚合酶链反应(rt-pcr)是可用于定量病毒转录物的pcr的变化方案。使用rna作为该方法的起始物质,并使用酶逆转录酶(rt)将其转化为双链dna。
慢性乙型肝炎的治疗目标是hbv感染的“功能性”治疗,这是利用目前可用的抗病毒剂没有实现的。如上进一步指出的,功能性治疗的主要障碍是hbvcccdna的复杂的稳定性和功能障碍的抗hbv免疫应答。在本发明的情况下,hbv的功能性治疗意味着每毫升的hbv基因组的数量低于100000、优选低于10000、更优选低于1000、100或10。特别优选的是完全根除或减少到低于检测极限。
鉴于根据本发明的结合分子为免疫效应细胞提供定制的特异性,免疫效应细胞的天然固有特异性或抗原呈递对于它们变得不相关。因此,大量候选效应细胞可以被重新靶向。此外,本发明的结合分子具有至少与单克隆抗体的生物利用度和半衰期相当的生物利用度和半衰期。
在第一方面的优选实施方案中,所述至少三种结合特异性为(a)三种特异性;或(b)四种特异性,其中第四特异性针对肝细胞。
优选地,结合分子是三特异性结合分子。换言之,存在恰好三种结合特异性。存在第四特异性的替代方案对于靶向某些细胞,特别是靶向肝细胞可以是有利的。在这方面,肝细胞上的优选标记物是ntcp。ntcp在本领域中称为hbv进入介导受体。
在进一步优选的实施方案中,所述分子是(a)多肽的多聚体;或(b)多肽。应理解,该优选实施方案的(b)项限定了由单个多肽构成的结合分子。
术语“多肽”定义了为氨基酸缩聚物的分子,所述氨基酸形成一条具有一个n末端和一个c末端的单链。组成氨基酸包括20种天然存在的蛋白氨基酸。优选地,所述多肽仅由所述天然存在的蛋白氨基酸组成。话虽如此,该术语扩展至除了所述天然存在的蛋白氨基酸之外还含有最高20%、10%、5%、2%或1%的氨基酸所述氨基酸选自非天然存在的α-氨基酸、β-氨基酸、d-氨基酸、硒代半胱氨酸、硒代蛋氨酸、羟脯氨酸、吡咯赖氨酸和鸟氨酸。还应理解,可以磷酸化一个或更多个例如2、3、4、5、6、7、8、9或10个氨基酸。磷酸化特别适用于丝氨酸、苏氨酸和酪氨酸。还可以存在本领域已知的其它翻译后修饰,包括糖基化。糖基化包括n-连接的糖基化,典型的为在天冬酰胺上的n-连接的糖基化,和o-连接的糖基化,典型的为在丝氨酸或苏氨酸残基上的o-连接的糖基化。n-末端和/或c-末端可以被保护,保护基团包括用于n-末端的乙酰基和用于c-末端的胺。所述多肽中包含的氨基酸之间的连接类型限于酰胺(conh)键。术语“酰胺键”包括将给定氨基酸的α-羧酸酯与后续氨基酸的α-氨基连接的肽键。“酰胺键”也扩展至异肽键,其是不存在于多肽主链上的酰胺键。例如,代替α-氨基,可以涉及赖氨酸的侧链氨基。类似地,代替α-羧基,可以涉及谷氨酸或天冬氨酸的侧链羧酸酯。设想出现一个或更多个例如2、3、4、5、6、7、8、9或10个异肽键。然而,优选其中组成氨基酸仅通过肽键彼此连接的多肽。
通常,多肽中的氨基酸数量没有上限。从序列表中包含的示例性多肽序列可以看出,本发明的多肽通常含有数百个氨基酸,优选250至1000个、400至900个,或700至800个氨基酸。区分一方面的肽和另一方面的多肽是常见的,其中肽具有30个或更少的氨基酸,多肽具有多于30个氨基酸。
根据本发明的术语“多聚体”包括二聚体、三聚体、四聚体、五聚物、六聚体、七聚体和八聚体。
在最广泛的形式中,关于至少三种所需特异性在多聚体的各个组成多肽上的位置没有限制。而可以想到的是使用组成多肽,其中每个多肽提供恰好一种结合特异性-由一个、两个、三个、四个或更多个结合位点实现-优选多聚体,其中每个组成多肽具有两种或三种特异性,尤其优选具有两种特异性的组成多肽。
就优选为三特异性的结合分子的本发明的结合分子而言,其为多肽的多聚体,优选二聚体或四聚体,优选的是(a)每个多肽是双特异性多肽;或(b)每个多肽独自地选自单特异性多肽、双特异性多肽和三特异性多肽。
毫无疑问,该优选实施方案仅涵盖也符合本发明的第一方面的最宽限定的构造。例如,两个单特异性多肽的二聚体不符合“三特异性”的要求,因此不被该优选实施方案所包含。
根据以上解释的在一方面的结合特异性和另一方面的结合位点之间的区别,应理解,单特异性多肽可以是一价的、二价的、三价的、四价的或可以含有甚至大于四价。类似地,双特异性多肽可以是二价的、三价的、四价的或含有大于四价。最后,三特异性多肽可以是三价的、四价的或含有大于四价。如上所指出的,结合位点可以由位于同一多肽上的两个免疫球蛋白结构域提供,也可以由位于不同多肽上的两个免疫球蛋白结构域提供。因此,在根据本发明的结合分子是多肽的多聚体的那些情况下,结合分子的价数可以超过所有组成多肽合起来的价数。结合分子的总价数与由所有组成多肽提供的累积价数之间的差异是由位于不同多肽链上的两个免疫球蛋白结构域所形成的那些结合位点的数目。
使用以下进一步公开的结构(iii)作为实例,该分子中的结合位点总数为5,其中4个是由成对的免疫球蛋白结构域形成,其中每一对位于单个多肽链上,一个结合位点是由位于不同多肽链上的一对免疫球蛋白结构域形成。结构(iii)中多肽的总数为2。
就多聚化或寡聚化的机理而言,应注意多肽可以共价地和/或非共价地连接在一起。例如,如果结合分子是两个多肽的二聚体,则所述多肽可以仅通过一个或更多个共价键连接在一起,所述共价键优选为二硫键。或者,两个多肽可以仅通过非共价相互作用而连接。同样可想到的是,两个多肽通过共价和非共价相互作用彼此结合。类似地,并且考虑到更高阶的多聚体,应理解仅共价相互作用,仅非共价相互作用,或共价和非共价相互作用的组合都可以使多聚体结合在一起。共价和非共价相互作用的组合可以是这样的:对于多聚体内给定的二元相互作用,仅存在共价相互作用或仅存在非共价相互作用,或共价和非共价相互作用的组合。
术语“多聚化”和“寡聚化”(等同使用)包括二聚化。
如以下进一步详述的,在某些情况下,对于本发明某些优选的结合分子,相对于非共价相互作用可以优选共价相互作用,反之亦然。
就本发明涉及多聚体而言,组成多肽还包含二聚化或多聚化区域。所述二聚化或多聚化区域可以提供共价和/或非共价的二聚化或多聚化。
在特别优选的实施方案中,连接本发明的两个多肽的所述二聚化区域由igg重链的铰链区组成或包含负责抗体重链二聚化的半胱氨酸残基。可以用于二聚化和多聚化的其他天然存在的二硫键是重链的第一恒定区与轻链的恒定区的各个c末端之间的二硫键。
优选地,所述二聚化区域由长度为32个氨基酸的子序列,即所谓的重链的铰链区(epkssdkthtcppcpapefegapsvflfppkp,参见seqidno:43至46)组成,并且包含负责重链二聚化的两个半胱氨酸残基(以上序列中加下划线的)。优选地,将介导天然抗体中igg重链恒定区和轻链恒定区之间的分子间二硫键的igg重链铰链区内的单个半胱氨酸残基突变成丝氨酸以防止异常的二硫键。
适合于非共价二聚化的二聚化结构域是本领域已知的,并且包括亮氨酸拉链(参见例如,dekruif和logtenberg,leucinezipperdimerizedbivalentandbispecificscfvantibodiesfromasemi-syntheticantibodyphagedisplaylibrary.j.biol.chem.1996,271:7630-7634)。触发二聚化的另一个选择是免疫球蛋白结构域之间天然存在的相互作用。这些相互作用例如在igg分子中在重链和轻链的可变区之间、在重链的第一恒定区和轻链的恒定区之间、在重链的各个第二恒定区和重链各个第三恒定区之间发生。
在进一步优选的实施方案中,组成多肽还包含间隔区,所述间隔区优选包含ch2区和ch3区。在包含scfv片段的组成多肽的情况下,所述间隔区可以位于第一scfv片段和第二scfv片段之间。
包含或由ch2区和ch3区组成的间隔区,特别是来自igg的间隔区是有利的。优点包括从产生的细胞有效分泌。此外,这种间隔区结合蛋白a的能力提供用于之后从试剂中纯化的手段。
一方面所述ch2和ch3区和另一方面所述二聚化区都可以由igg,优选igg1或igg2分子,甚至更优选人类igg1或igg2(higg1,higg2)分子的相应区提供。提供ch2区和ch3区的higg1分子的优选子序列可见于seqidno:43至46中。seqidno:50和51的序列实现非共价二聚化的优选实施方案,更具体地通过亮氨酸拉链结构域来实施。优选地—并且这适用于所提及的序列—higg1的部分,特别是所述ch2区在多个位置被诱变处理以减少与fc受体的结合(以下进一步给出的序列中以加粗斜体指示)。更一般地,fc区,特别是ch2区和/或ch3区可以在一个或更多个位置突变以减少与fc受体的结合。这种方法是本领域已知的,并且例如在armour等人,recombinanthumaniggmoleculeslackingfcgammareceptoribindingandmonocytetriggeringactivities.eur.j.immunol.1999.29:2613-2624和shields等人,highresolutionmappingofthebindingsiteonhumanigg1forfcgammari,fcgammarii,fcgammariii,andfcrnanddesignofigg1variantswithimprovedbindingtothefcgammar.j.biol.chem.2001.276:6591-6604.)。这是有利的,因为根据本发明,抗体依赖性细胞介导的细胞毒性(adcc)的触发是不优选的。
换言之,抗体fc片段,优选如上定义的经修饰的抗体fc片段可以用于实现间隔区和二聚化区。术语“fc片段”是本领域技术人员已知的,并且定义为通过用木瓜蛋白酶切割获得的igg片段并包含ch2和ch3区。
在所述第一scfv片段和所述间隔区之间和/或所述间隔区和所述第二scfv片段(a)之间可以存在连接序列。优选的连接序列在上文中公开。从序列表中包含的优选序列,特别是seqidno:43至46的序列可以看出,这样的连接序列可以由甘氨酸或者甘氨酸和丝氨酸组成。
术语“ch2区”和“ch3区”具有其本领域确定的含义。它们指抗体重链的第二恒定区和第三恒定区。
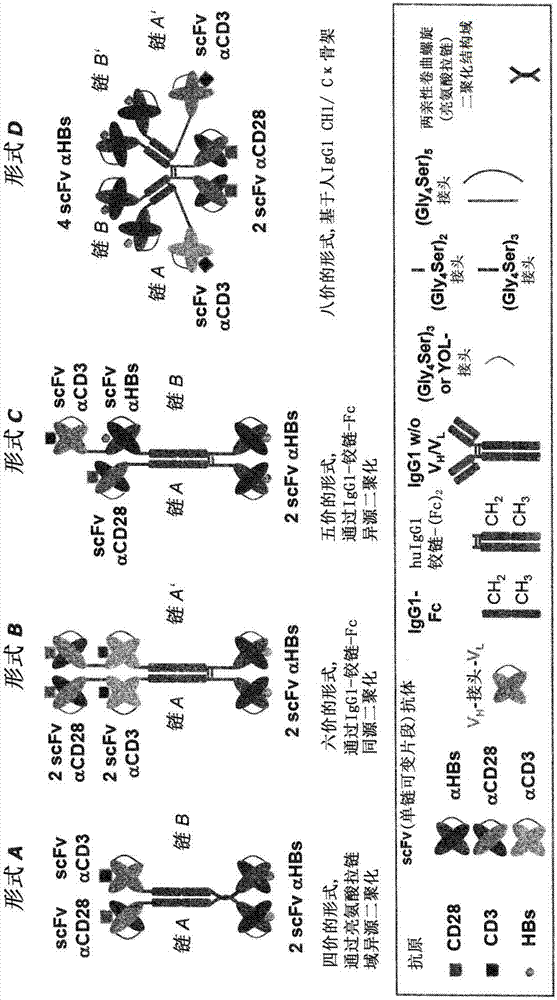
转向如所附的图1和2中所示的优选分子结构,分别是二聚化和多聚化,优选如下进行。相同多肽链的同源二聚化,例如形式b、d、g、h、l、m和o,优选通过形成人类igg1的铰链域的分子间二硫键来实现。同样适用于如形式c中不同氨基酸序列的多肽链的异源二聚化。在形式b和c中,ch2和ch3区通过非共价相互作用有助于二聚体的稳定性。在形式g、h、n和o中cκ和ch1区之间天然存在的分子间二硫键作为共价机制与ch2和ch3区之间的非共价相互作用一样有助于二聚化。这两种机制的前者也有助于形式d、l和m中的二聚化。
为了具有不同氨基酸序列的多肽的异源二聚化,例如在形式a、e、f和k中,优选上述亮氨酸拉链结构域,其也被称为两亲性卷曲螺旋二聚化结构域。这种二聚化结构域的实例在dekruif和logtenberg,j.biol.chem.271,7630-7634(1996)和arndt等人,j.mol.biol.312,221-228(2001)中描述。在形式a和e中,ch2和ch3区有助于二聚化。在形式f中,人类igg1的ch2和ch3区之间的非共价相互作用以及cκ和ch1区之间的分子间二硫键有助于二聚化。
在另一个优选的实施方案中,所述多肽中的一个、更多个或所有由一种或更多种scfv分子组成或包含一种或更多种scfv分子。该实施方案扩展至串联scfv分子,也指向(scfv)2以及(scfv)3分子,例如结构(vii)。
该优选的实施方案限定本发明结合分子中所包含的多肽的优选组成要素。多肽可以由那些优选的组成分子组成或包含那些优选的组成分子。例如,当多肽会被用于提供单一结合特异性时,这可以通过使用由scfv分子组成的多肽来完成。在另一方面,当要使用提供两种结合特异性的多肽时,可以通过包含两个scfv分子的多肽或由双抗体来完成。
术语“scfv”在本领域是已确定的。该缩写代表抗体的“单链可变片段”,并且定义能够特异性识别和结合抗原表位的多肽。如上所指出的,三个cdr由抗体轻链可变区(vl)呈现,三个cdr由抗体重链可变区(vh)呈现。就vl和vh不共价连接来说,它们构成fv片段。在另一方面,在scfv中,两个可变区通过肽接头彼此连接。获得的融合构建体是单个多肽链。这提供了scfv分子的容易表达。
术语“vh区”和“vl区”是本领域确定的。因此,它们分别指免疫球蛋白的重链可变区(vh)和免疫球蛋白的轻链可变区(vl)。通常,vh和vl区各自包含三个互补决定区(cdr)。
术语“双抗体”、“三抗体”和“四抗体”在本领域中是已知的,并涉及含有两个、三个或四个fv片段的单链构建体。一般而言,双抗体、三抗体和四抗体包含2条、3条或4条多肽链。这些可以彼此连接,如此获得单个多肽链。重组抗体构建体的综述是little等人,immunologytoday,21,364-370(2000)。
肽接头优选用于连接scfv的可变区或将scfv连接至二聚化区/多聚化区和/或间隔区或以下定义的fc。类似地,短的肽接头可以用于双抗体、三抗体和四体抗体的构建,如上述综述中所阐述的。
通常,肽接头具有3至30个氨基酸,优选5至25个或10至20个氨基酸的长度。优选的是不干扰或基本不干扰它们所连接的结构域或多肽(连接产生单个连续的多肽链)的结构和/或功能的那些接头。接头包括富gly的接头,例如在本发明的优选多肽中用于连接ctl或nk特异性scfv的vh/vl区的(gly4ser)3(seqidno:47)接头和yol接头(seqidno:48;akttpkleegefsearv,如sellrie等人,journalofbiochemistryandmolecularbiology,第40卷,第6期,2007年11月,875-880页所述的),其在本发明的优选多肽中用于连接对hbv表面抗原有特异性的scfv的vh/vl区。此外,(gly4ser)4接头(seqidno:49)可以用于连接对hbv表面抗原有特异性的scfv的vh/vl区或本发明的结合分子的其他结构单元。
在涉及为多肽的多聚体的那些结合分子的进一步优选的实施方案中,所述多聚体优选为二聚体或四聚体,所述结合分子优选为三特异性结合分子,一个、更多个或所有结合位点由两个所述多肽一起提供。
以上已经解释了该优选实施方案的结构背景。特别地,该优选实施方案提供了两个免疫球蛋白结构域,其提供构成位于不同多肽链上的结合位点的两组三个cdr。
在特别优选的实施方案中,所述结合分子包含fv片段;fab片段;二抗体;三抗体;四抗体;或免疫球蛋白g分子,其任选地具有经修饰的fc部分。
使用的与该特别优选的实施方案相关的术语具有其在本领域确定的意义。特别地,术语“fv片段”表示两个免疫球蛋白结构域的非共价复合物,所述免疫球蛋白结构域是轻链的可变区和重链的可变区。
从结构的角度来看,fv片段包含在fab片段中,其中fab片段还包含抗体的轻链恒定区cl和重链第一恒定ch1区。总而言之,fab片段包含四个免疫球蛋白结构域,其中所述四个免疫球蛋白结构域中的两个,即vl区和vh区一起形成结合位点。可以通过用木瓜蛋白酶切割从抗体获得fab片段。
当根据本发明的结合分子在某一位置包含fv片段时,应理解在该特定位置处,它没有相邻的轻链恒定区并且没有相邻的重链恒定区1,否则要注意的是在考虑的位置处会是fab片段。类似地,fab片段的要求排除了在所考虑的位置处存在ch2和ch3区。在本文中“相邻的”是指在天然存在的抗体中发现的邻近。
所述免疫球蛋白g分子的fc部分的修饰包括减少或消除与fc受体结合的修饰;见下文。
对于根据本发明的结合分子,优选的是完全没有fc部分的结合分子。还优选存在fc部分,其中所述fc部分是如上所述经修饰的,或者另外地被修饰以使得fc部分不被n-糖基化。最后,也可以使用天然fc部分。
提供降低fc结合和/或减少补体活化的修饰是利用来自hlgg2或hlgg4的fc替代higg1衍生的fc。
本文所用的术语“抗体”具有其在本领域确定的含义。优选地,抗体指单克隆抗体。单克隆抗体可以例如通过
在涉及为多肽的多聚体并且其中每个多肽是双特异性多肽的本发明的那些结合分子的优选实施方案中,每个双特异性多肽的第一特异性是根据第一方面所限定的第一特异性,每个多肽的第二特异性是针对cd3或cd28的。
因此,应理解优选的是,具有结合hbv表面抗原能力并且它们对ctl表面标记物的特异性彼此不同的组成双特异性多肽。
在这方面尤其优选的结合分子是多肽的二聚体和四聚体。在由双特异性二价多肽的二聚体组成的二聚体的组内,优选非共价二聚体。
尤其优选的二聚体是结构(i)、(ii)和(iii)的那些二聚体,尤其优选的四聚体是结构(iv)的四聚体:
“链a”和“链b”表示具有不同氨基酸序列的多肽。“链a”和“链a'”表示具有相同氨基酸序列的不同多肽。在靠近每个结合位点处指示特异性。如本领域常见的,术语“抗”缩写为“α”。
结构(i)是由两个双特异性二价多肽组成的非共价二聚体。结构(i)在图1的“形式a”中示出。
结构(ii)是两个多肽的共价二聚体,其中表示为“链a”的多肽是双特异性二价多肽,表示为“链b”的多肽是双特异性三价多肽。结构(ii)在图1的“形式c”中示出。
我们注意到,结构(iii)具有每个结合位点由两个免疫球蛋白结构域提供的四个结合位点,所述两个免疫球蛋白结构域位于相同的多肽链上,而第五个结合位点由位于不同多肽链上的两个免疫球蛋白结构域形成。实施例包括本发明的结构(iii)的结合分子的数据。
结构(iv)在图1的“形式d”中示出。实施例包括本发明的结构(iv)的结合分子的数据。
在本发明的结合分子的进一步优选实施方案中,所述结合分子优选为三特异性结合分子,每个多肽是三特异性结合分子。
换言之,每个组成多肽已经包括根据本发明的第一方面的三种特异性。使用三特异性多肽的多聚体、优选二聚体,可以微调或优化结合特性。
在这方面尤其优选的是相同多肽的二聚体,其最优选具有结构(v):
结构(v)在图1的“形式b”中示出。
本发明还提供了其中结合位点仅由位于不同多肽链上的免疫球蛋白结构域形成的结合分子。实例是具有结构(vi)的结合分子:
结构(vi)是三抗体。结构(vi)在图2的“形式j”中示出。
为了提供具有结构(vi)的结合分子的功能性,也可以采用不同的拓扑学。因此,本发明在进一步优选的实施方案中扩展至具有结构(vii)的结合分子:
结构(vii)由一个多肽构成。它实现了以上公开的(scfv)3结构。结构(vii)在图2的“形式i”中示出。
图1中描述了本发明的优选实施方案。为了举例说明,指示了抗原。应理解,抗原不是根据本发明的结合分子的一部分。根据本发明的其他分子结构在图2中示出。
上述第一特异性优选由六个cdr的组提供,其中六个cdr的组优选为seqidno:1至6、7至12或13至18。
针对cd3的本发明的结合分子的上文限定的第二特异性优选由具有seqidno:19至24的序列的六个cdr的组提供。
针对cd28的本发明的结合分子的上文限定的第三特异性优选由具有seqidno:25至30的序列的六个cdr的组提供。
就第二和第三特异性选自针对cd16、cd56、nkp30、nkp46、4-1bb和nkg2d的特异性而言,优选的是第二特异性是针对cd16而第三特异性是针对cd56。优选的相应的6个cdr的组分别是seqidno:31至36和37至42的那些。
序列表中使用的c8、5f9、5a19、okt3、9.3、a9和ncam29.2表示各个cdr所来自的抗体,并且分别指优选的抗hbs抗体、第二种不同的抗hbs抗体、针对hbv大表面抗原的抗体、针对人cd3的抗体、针对人cd28的抗体、针对人cd16的抗体和针对人cd56的抗体。“hbs”表示hbv小表面抗原。
就使用双特异性二价多肽而言,特别优选的是所述多肽包含seqidno:43至46中任一项的氨基酸序列或由其组成或者包含显示与seqidno:43至46中任一项至少80%一致性的氨基酸序列,条件是显示80%一致性的所述氨基酸序列的cdr分别与包含在seqidno:43至46中任一项的那些一致。在seqidno:43中,最后三个残基“gns”是非必要的。
序列一致性的优选水平包括至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%和至少99%。用于确定序列一致性的手段和方法是本领域众所周知的。用于确定成对序列一致性的优选算法是例如在mcginnis和madden(nucleicacidresearch32,w20-w25(2004))中所描述的基本局部比对搜索工具(blast)。
在给定序列中,在当前情况下在seqidno:43至46的序列中,所述cdr的位置可以用本领域确定的方法确定,已知的本领域确定的方法分别包括chothia,kabat和lefranc/imgt的系统。在没有任何相反指示的情况下,应理解根据上文限定的特别优选的实施方案的cdr是上文限定的那些,即具有seqidno:1至6、7至12或13至18的序列的第一组和具有seqidno:19至24、25至30、31至36或37至42的序列的第二组。从包含在所附序列表中的序列可以看出,这些特异性cdr序列(在下文还出现的序列中加下划线)实际被包含在seqidno:43至46的序列中。
seqidno:1至6的序列限定cdr,seqidno:37至40限定能够结合hbv的小表面抗原中的特异性表位的双特异性二价多肽。该表位位于a-决定簇中,其分别暴露于受感染的细胞和病毒体的表面。术语“a-决定簇”用于表示hbv的小表面抗原中用于诱导保护性体液免疫应答的主要表位所位于的区域。这些cdr以及seqidno:43至46的多肽具有可以用于所有hbv血清型的优点。
在第二方面,本发明提供编码构成结合分子的多肽的一组核酸,该结合分子是多肽的多聚体。在替代方案中,所述第二方面涉及编码为单个多肽的结合分子的核酸。多肽的优选实施方案导致相应的所述核酸的优选实施方案。
术语“核酸”具有其在本领域确定的含义,并且没有特别限制。优选的是dna,例如基因组dna或cdna,以及rna,例如mrna。虽然不是优选的,但还设想使用核苷酸衍生物,核苷酸衍生物包括2'衍生化的核苷酸,例如2'甲基核苷酸;如在肽核酸等中出现的肽核苷酸等。
在另一个方面,本发明提供了包含或由第一方面的一种或更多种结合分子和/或第二方面的一种或更多种核酸组成的药物组合物。
药物组合物还可以包含药学上可接受的载体、赋形剂和/或稀释剂。合适的药物载体、赋形剂和/或稀释剂的实例是本领域众所周知的,并且包括磷酸盐缓冲盐溶液、水、乳液,例如油/水乳液,各种类型的润湿剂、无菌溶液等。包含这类载体的组合物可以用众所周知的常规方法配制。这些药物组合物可以以合适的剂量向对象施用。合适的组合物的施用可以通过不同的方式实现,例如通过静脉内、皮下或口服施用,这三种选择是优选的,此外可以通过腹膜内、肌肉内、局部、皮内、鼻内或支气管内施用。用于口服施用的制剂包括片剂和糖浆剂。特别优选所述施用通过注射进行。也可以将组合物直接施用于靶位点,例如通过向外部或内部靶位点的基因枪递送。剂量方案会由主治医师和临床因素确定。如在医学领域中众所周知的,对于任何一个患者的剂量取决于许多因素,包括患者的体型、体表面积、年龄、待施用的具体化合物、性别、施用的时间和途径、一般健康状况和同时施用的其他药物。蛋白质药学上的活性物质可以以每剂量1ng/kg体重至10mg/kg体重的量存在;然而,预期低于或高于该示例性范围的剂量,尤其是考虑到上述因素。如果方案是连续输注,它应该是每分钟每公斤体重1μg至10mg单位。
特别优选静脉内施用。
在另一个方面,本发明提供在治疗或预防hbv感染和/或由所述hbv感染引起的病症的方法中使用的本发明的一种或更多种结合分子、核酸和/或药物组合物,由所述hbv感染引起的所述病症选自肝硬化、肝细胞癌和肝癌,所述肝癌的特征在于一种或更多种hbv表面抗原的表达。优选的是,所述肝细胞癌的特征在于一种或更多种上文限定的hbv表面抗原的表达。
在另一个方面,本发明提供治疗或预防hbv感染和/或由所述hbv感染引起的病症的方法,由所述hbv感染引起的所述病症选自肝硬化和肝细胞癌,所述方法包括向需要其的患者分别施用有效治疗量或预防量的本发明的一种或更多种结合分子、核酸和/或药物组合物。
在一个实施方案中,用于治疗方法的所述药物组合物、所述结合分子和/或核酸和所述治疗方法分别是这样的:所列举的结合分子和/或核酸是所包含或使用的唯一药物活性剂。
话虽如此,也慎重地设想在联合治疗中并入一种或更多种另外的药物活性剂。这种另外的药物活性剂可以选自干扰素或其他免疫调节剂(例如干扰素α2a或2b、干扰素λ)、直接起作用的抗病毒剂如核苷类似物(例如拉米夫定(epivir-hbv、zeffix或heptodin)、阿德福韦酯(hepsera、preveon)、恩替卡韦(baraclude、entaliv)、替比夫定(tyzeka、sebivo)、替诺福韦(viread))、进入抑制剂(例如myrcludex-b)、其他抗病毒剂、或细胞因子如白介素-2。
因此,设想在所述药物组合物中和根据用于治疗方法的所述结合分子和所述治疗方法,所列举的结合分子和此外一种或更多种上文限定的特异性药物活性剂是所包含或使用的唯一药物活性剂。
或者,可以存在又一种药物活性剂。可以添加增强结合分子的稳定性的另外成分或药物活性剂,例如蛋白质如牛奶蛋白、营养物质或盐溶液以增加生物利用度和/或半衰期。
在另一个方面,本发明提供一种杀伤被hbv感染的细胞的体外方法,所述方法包括用(i)免疫效应细胞和(ii)本发明的一种或更多种结合分子培养所述被hbv感染的细胞。
在体外方法的优选实施方案中,所述免疫效应细胞(i)被包含在外周血单核细胞中;或(ii)是或包含nk细胞和/或ctl。
在另一个方面,本发明提供体外或离体免疫效应细胞,其(a)具有与所述免疫效应细胞的表面抗原结合的第一方面的结合分子;和/或(b)已经用根据第二方面的一种或更多种核酸转化、转染或转导。优选免疫效应细胞和优选的由免疫效应细胞呈递的表面抗原如上所定义。这种免疫效应细胞可用于向患有hbv感染、肝硬化或肝细胞癌的患者施用。因此,还提供一种药物组合物,其包含或由其表面抗原已经与本发明的结合分子结合的免疫效应细胞组成。还提供其表面抗原已经与本发明的结合分子结合的免疫效应细胞,其用于治疗或预防hbv感染、肝硬化或肝细胞癌的方法。
本发明扩展至次优选的某些实施方案,因此可以通过附带条件的方式从较宽的实施方案中排除。
根据本发明次优选的是由第一和第二多肽组成的三特异性四价结合分子,所述第一和第二多肽各自是双特异性且二价的,其中任一多肽的第一特异性针对选自hbv小表面抗原、hbv中表面抗原和hbv大表面抗原的hbv表面抗原,第二特异性分别针对cd3和cd28。
在这方面更加次优选的是由第一和第二多肽组成的三特异性四价结合分子,所述第一和第二多肽各自是双特异性且二价的,其中所述第一多肽和所述第二多肽彼此非共价地结合,并且其中任一多肽的第一特异性针对选自hbv小表面抗原、hbv中表面抗原和hbv大表面抗原的hbv表面抗原;第二特异性分别针对cd3和cd28。
特别次优选的是由第一和第二多肽组成的三特异性四价结合分子,所述第一多肽和第二多肽各自是双特异性且二价的,其中所述第一多肽和所述第二多肽之间有至少一个共价键,优选在所述第一多肽的cys残基和所述第二多肽的cys残基之间的至少一个二硫键,并且其中任一多肽的第一特异性针对选自hbv小表面抗原、hbv中表面抗原和hbv大抗原的hbv表面抗原;第二特异性分别针对cd3和cd28。
尤其次优选的是由第一和第二多肽组成的三特异性四价结合分子,所述第一和第二多肽各自是双特异性且二价的,其中所述第一多肽和所述第二多肽之间有两个二硫键,并且其中任一多肽的第一特异性针对选自hbv小表面抗原、hbv中表面抗原和hbv大表面抗原的hbv表面抗原;第二特异性分别针对cd3和cd28。
关于每个以上限定的次优选的实施方案,特别次优选的是在第一和第二多肽上的结合位点由scfv片段提供。此外,关于任何前述次优选的实施方案,特别次优选的是在所述第一和第二多肽两者中,在提供结合位点的两个scfv片段之间有ch2区和ch3区。
因此,尤其次优选的是由第一和第二多肽组成的三特异性四价结合分子,所述第一和第二多肽各自是双特异性且二价的,其中所述第一多肽和所述第二多肽之间有两个二硫键,其中任一多肽的第一特异性针对选自hbv小表面抗原、hbv中表面抗原和hbv大表面抗原的hbv表面抗原;第二特异性分别针对cd3和cd28,其中在第一和第二多肽上的结合位点由scfv片段提供,并且其中在所述第一和第二多肽两者中,在提供结合位点的两个scfv片段之间有ch2区和ch3区。
因此,例如,本发明还提供了包含至少三种结合特异性的结合分子,其中(a)第一特异性针对选自hbv小表面抗原、hbv中表面抗原和hbv大表面抗原的乙型肝炎病毒(hbv)表面抗原;(b)(i)第二和第三特异性分别针对cd3和cd28;或(ii)第二和第三特异性选自针对cd16、cd56、nkp30、nkp46、4-1bb和nkg2d的特异性;(c)每种结合特异性由一个或更多个结合位点提供,每个结合位点由以下独立地提供:(i)六个互补决定区(cdr)的组,其中所述六个cdr的组由三个cdr的第一组和三个cdr的第二组组成,其中所述第一和第二组各自被包含在免疫球蛋白结构域中;或(ii)三个cdr的组,其中所述三个cdr的组被包含在免疫球蛋白结构域中;条件是三特异性四价结合分子由第一和第二多肽组成,所述第一和第二多肽各自是双特异性且二价的,其中所述第一多肽和所述第二多肽之间有两个二硫键,其中任一多肽的第一特异性针对选自hbv小表面抗原、hbv中表面抗原和hbv大表面抗原的hbv表面抗原,第二特异性分别针对cd3和cd28,其中第一和第二多肽上的结合位点由scfv片段提供,并且其中在所述第一和第二多肽中的两者中,排除了提供结合位点的两个scfv片段之间有ch2区和ch3区。
关于本说明书、特别是权利要求中描述的实施方案,其旨在将从属权利要求中提及的每个实施方案与所述从属权利要求引用的每个(独立或从属)权利要求的各个实施方案组合。例如,在独立权利要求1描述3个替代方案a、b和c,从属权利要求2描述3个替代方案d、e和f和从属于权利要求1和2的权利要求3描述3个任选方案g、h和i的情况下,应理解,该说明书明确地公开了对应于以下组合的实施方案:a、d、g;a、d、h;a、d、i;a、e、g;a、e、h;a、e、i;a、f、g;a、f、h;a、f、i;b、d、g;b、d、h;b、d、i;b、e、g;b、e、h;b、e、i;b、f、g;b、f、h;b、f、i;c、d、g;c、d、h;c、d、i;c、e、g;c、e、h;c、e、i;c、f、g;c、f、h;c、f、i,除非另有明确说明。
类似地,并且在独立权利要求和/或从属权利要求中不描述替代方案的那些情况下,应理解,如果从属权利要求引用多个在前的权利要求,则认为明确地公开了主题由此覆盖的任何组合。例如,在独立权利要求1、从属权利要求2引用权利要求1,从属权利要求3引用权利要求2和1两者的情况下,由此认为权利要求3和1的主题的组合被清楚地和明确地地公开,如权利要求3、2和1的主题组合一样。在还存在引用权利要求1至3中任一项的从属权利要求4的情况下,由此认为权利要求4和1、权利要求4、2和1、权利要求4、3和1以及权利要求4、3、2和1的主题组合被清楚地和明确地公开。
在本申请中公开了序列:
seqidno1
c8hccdr1
glyphethrpheserglytyrala
seqidno2
c8hccdr2
ileserglyserglyglyserthr
seqidno3
c8hccdr3
alalysproproglyargglnglutyrtyrglyserseriletyrtyrpheproleuglyasn
seqidno4
c8lccdr1
asnileglyserlysser
seqidno5
c8lccdr2
aspaspser
seqidno6
c8lccdr3
glnvaltrpaspserserseraspleuvalval
seqidno7
5f9hccdr1
glyphethrpheasnasntyrala
seqidno8
5f9hccdr2
ileasnseraspglyargserthr
seqidno9
5f9hccdr3
alaargthrphetyralaasptyr
seqidno10
5f9lccdr1
glnasnvalaspthrthr
seqidno11
5f9lccdr2
trpalaser
seqidno12
5f9lccdr3
glnglntyrserilepheprotyrthr
seqidno13
5a19hccdr1
glyphethrphesersertyrala
seqidno14
5a19hccdr2
valserseraspglysertyrala
seqidno15
5a19hccdr3
alaserpheashtrpaspvalalatyr
seqidno16
5a19lccdr1
glnserleuleuasnthrargthrarglyssertyr
seqidno17
5a19lccdr2
trpalaser
seqidno18
5a19lccdr3
lysglnsertyrserleutyrthr
seqidno19
okt3hccdr1
glytyrthrphethrargtyrthr
seqidno20
okt3hccdr2
ileasnproserargglytyrthr
seqidno21
okt3hccdr3
alaargtyrtyraspasphistyrcysleuasptyr
seqidno22
okt3lccdr1
serservalsertyr
seqidno23
okt3lccdr2
aspthrser
seqidno24
okt3lccdr3
glnglntrpserserasnprophethr
seqidno25
9.3hccdr1
glypheserleuserasptyrgly
seqidno26
9.3hccdr2
iletrpalaglyglyglythr
seqidno27
9.3hccdr3
alaargasplysglytyrsertyrtyrtyrsermetasptyr
seqidno28
9.3lccdr1
gluservalglutyrtyrvalthrserleu
seqidno29
9.3lccdr2
alaalaser
seqidno30
9.3lccdr3
glnglnserarglysvalprotyrthr
seqidno31
a9hccdr1
glytyrthrphethrasntyrtrp
seqidno32
a9hccdr2
iletyrproglyglyglytyrthr
seqidno33
a9hccdr3
alaargseralasertrptyrpheaspval
seqidno34
a9lccdr1
thrglythrvalthrthrserasntyr
seqidno35
a9lccdr2
histhrasn
seqidno36
a9lccdr3
alaleutrptyrasnasnhistrpval
seqidno37
ncam29.2hccdr1
glyphethrpheserserphegly
seqidno38
ncam29.2hccdr2
ileserserglysertyralaile
seqidno39
ncam29.2hccdr3
valargglyargargleuglygluglytyralametasptyr
seqidno40
ncam29.2lccdr1
glnserleuleutyrserserashglnlysashtyr
seqidno41
ncam29.2lccdr2
trpalaser
seqidno42
ncam29.2lccdr3
glnglntyrsersertrpthr
seqidno43
c8-higg1fcmut-okt3
mdfevqifsfllisasvimsrmaevqlvesgggllqpggslrlscaasgftfsgyamswvrqapgkglewvssisgsggstyyadsvkgrftisrdnskntlylqmnslraedtalyycakppgrqeyygssiyyfplgnwgqgtlvtvssastkgpkleegefsearvqsaltqpasvsvapgqtaritcggnnigsksvhwyqqkpgqapvlvvyddsdrpsgiperfsgsnsgntatltisrveagdeadyycqvwdsssdlvvfgggtkltvlgnsggggsggggsggggsasepkssdkthtcppcpappaagpsvflfppkpkdtlmisrtpevtcvvvdvshedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykcavsnkglassiektiskakgqprepqvytlppsrdeltknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnhytqkslslspgkdpgwshpqfeksrggggqvqlqqsgaelarpgasvkmsckasgytftrytmhwvkqrpgqglewigyinpsrgytnynqkfkdkatlttdkssstaymqlssltsedsavyycaryyddhycldywgqgttltvssgnsggggsggggsggggsasqivltqspaimsaspgekvtmtcsasssvsymnwyqqksgtspkrwiydtsklasgvpahfrgsgsgtsysltisgmeaedaatyycqqwssnpftfgsgtkleingns
seqidno44
c8-hlgg1fcmut-9.3
mdfevqifsfllisasvimsrmaevqlvesgggllqpggslrlscaasgftfsgyamswvrqapgkglewvssisgsggstyyadsvkgrftisrdnskntlylqmnslraedtalyycakppgrqeyygssiyyfplgnwgqgtlvtvssastkgpkleegefsearvqsaltqpasvsvapgqtaritcggnnigsksvhwyqqkpgqapvlvvyddsdrpsgiperfsgsnsgntatltisrveagdeadyycqvwdsssdlvvfgggtkltvlgnsggggsggggsggggsasepkssdkthtcppcpappaagpsvflfppkpkdtlmisrtpevtcvvvdvshedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykcavsnkglassiektiskakgqprepqvytlppsrdeltknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnhytqkslslspgkdpgwshpqfekssggggqvqlqesgpglvtpsqslsitctvsgfslsdygvhwvrqspgqglewlgviwagggtnynsalmsrksiskdnsksqvflkmnslqaddtavyycardkgysyyysmdywgqgttvtvssrgggsggggsggggsdieltqspaslavslgqratiscrasesveyyvtslmqwyqqkpgqppkllifaasnvesgvparfsgsgsgtnfslnihpvdeddvamyfcqqsrkvpytfgggtkleikr
seqidno45
c8-hlgg1fcmut-a9
mdfevqifsfllisasvimsrmaevqlvesgggllqpggslrlscaasgftfsgyamswvrqapgkglewvssisgsggstyyadsvkgrftisrdnskntlylqmnslraedtalyycakppgrqeyygssiyyfplgnwgqgtlvtvssastkgpkleegefsearvqsaltqpasvsvapgqtaritcggnnigsksvhwyqqkpgqapvlvvyddsdrpsgiperfsgsnsgntatltisrveagdeadyycqvwdsssdlvvfgggtkltvlgnsggggsggggsggggsasepkssdkthtcppcpappaagpsvflfppkpkdtlmisrtpevtcvvvdvshedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykcavsnkglassiektiskakgqprepqvytlppsrdeltknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnhytqkslslspgkdpgwshpqfeksrggggqvqlqqsgaelvrpgtsvkisckasgytftnywlgwvkqrpghglewigdiypgggytnynekfkgkatvtadtssrtayvqvrsltsedsavyfcarsaswyfdvwgagttvtvssgnsggggsggggsggggsasqavvtqesalttspgetvtltcrsntgtvttsnyanwvqekpdhlftglightnnrapgvparfsgsligdkaaltitgaqtedeaiyfcalwynnhwvfgggtkltvl
seqidno46
c8-hlgg1fcmut-ncam29.2
mdfevqifsfllisasvimsrmaevqlvesgggllqpggslrlscaasgftfsgyamswvrqapgkglewvssisgsggstyyadsvkgrftisrdnskntlylqmnslraedtalyycakppgrqeyygssiyyfplgnwgqgtlvtvssastkgpkleegefsearvqsaltqpasvsvapgqtaritcggnnigsksvhwyqqkpgqapvlvvyddsdrpsgiperfsgsnsgntatltisrveagdeadyycqvwdsssdlvvfgggtkltvlgnsggggsggggsggggsasepkssdkthtcppcpappaagpsvflfppkpkdtlmisrtpevtcvvvdvshedpevkfnwryvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykcavsnkglassiektiskakgqprepqvytlppsrdeltknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnhytqkslslspgkdpgwshpqfekssgggdvqlvesggglvqpggsrklscaasgftfssfgmhwvrqapekglewvayissgsyaiyyadtvkgrftisrdnpentlflqmtslrsedsamyycvrgrrlgegyamdywgqgtsvtvssgnsggggsggggsggggsasdivmsqspsslavsvgekvtmsckssqsllyssnqknylawyqqkpgqspklliywastrksgvpdrftgsgsgtdftltissvkaedlavyycqqysswtfgggtkleikr
seqidno47
glyglyglyglyserglyglyglyglyserglyglyglyglyser
seqidno48
alalysthrthrprolysleuglugluglyglupheserglualaargval
seqidno49
glyglyglyglyserglyglyglyglyserglyglyglyglyserglyglyglyglyser
seqidno50
具有jun亮氨酸拉链二聚化结构域(黑体斜体下划线)的c8-higg1fcmut-okt3
mdfevqifsfllisasvimsrmaevqlvesgggllqpggslrlscaasgftfsgyamswvrqapgkglewvssisgsggstyyadsvkgrftisrdnskntlylqmnslraedtalyycakppgrqeyygssiyyfplgnwgqgtlvtvssastkgpkleegefsearvqsaltqpasvsvapgqtaritcggnnigsksvhwyqqkpgqapvlvvyddsdrpsgiperfsgsnsgntatltisrveagdeadyycqvwdsssdlvvfgggtkltvlgnsggggsggggsggggsasriarleekvktlkaqnselastanmlreqvaqlkqkvmnyssepkssdkthtcppcpappaagpsvflfppkpkdtlmisrtpevtcvvvdvshedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykcavsnkglassiektiskakgqprepqvytlppsrdeltknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnhytqkslslspgkdpgwshpqfeksrggggqvqlqqsgaelarpgasvkmsckasgytftrytmhwvkqrpgqglewigyinpsrgytnynqkfkdkatlttdkssstaymqlssltsedsavyycaryyddhycldywgqgttltvssgnsggggsggggsggggsasqivltqspaimsaspgekvtmtcsasssvsymnwyqqksgtspkrwiydtsklasgvpahfrgsgsgtsysltisgmeaedaatyycqqwssnpftfgsgtkleingns
seqidno51
具有fos亮氨酸拉链二聚化结构域(黑体斜体下划线)的c8-hlgglfcmut-9.3
mdfevqifsfllisasvimsrmaevqlvesgggllqpggslrlscaasgftfsgyamswvrqapgkglewvssisgsggstyyadsvkgrftisrdnskntlylqmnslraedtalyycakppgrqeyygssiyyfplgnwgqgtlvtvssastkgpkleegefsearvqsaltqpasvsvapgqtaritcggnnigsksvhwyqqkpgqapvlvvyddsdrpsgiperfsgsnsgntatltisrveagdeadyycqvwdsssdlvvfgggtkltvlgnsggggsggggsggggsasltdtlqaetdqledkksalqteianllkekeklefilaayssepkssdkthtcppcpappaagpsvflfppkpkdtlmisrtpevtcvvvdvshedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykcavsnkglassiektiskakgqprepqvytlppsrdeltknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnhytqkslslspgkdpgwshpqfekssggggqvqlqesgpglvtpsqslsitctvsgfslsdygvhwvrqspgqglewlgviwagggtnynsalmsrksiskdnsksqvflkmnslqaddtavyycardkgysyyysmdywgqgttvtvssrgggsggggsggggsdieltqspaslavslgqratiscrasesveyyvtslmqwyqqkpgqppkllifaasnvesgvparfsgsgsgtnfslnihpvdeddvamyfcqqsrkvpytfgggtkleikr
附图示出:
图1和图2举例说明了本发明的优选三特异性结合分子。
图3:t细胞被五价和八价的三特异性抗体活化。确定在双特异性和三特异性抗体存在下在固定的hbsag上培养48小时的t细胞的ifnγ分泌。当与用cd3活化的双特异性抗体进行单一处理相比,cd3活化的双特异性抗体(bsabαcd3)和cd28活化的双特异性抗体(bsabαcd28)的组合应用导致在抗原存在下ifnγ的分泌增加。五价(trimab)和八价(okta)的三特异性抗体构建体的应用诱导显著更高的干扰素分泌,其中trimab是最有效的构建体。用cd28活化抗体以及未经处理的对照pbmc的单独处理未显示升高的ifnγ水平。
图4:t细胞被五价和八价的三特异性抗体活化变成细胞毒性。(a)确定指示了在双特异性和三特异性的抗体存在下在固定的hbsag上培养48小时的t细胞的细胞毒性活化的lamp-1易位。当与用cd3活化的双特异性抗体进行单一处理相比,cd3活化的双特异性抗体(bsabαcd3)和cd28活化的双特异性抗体(bsabαcd28)的组合应用导致在抗原存在下脱粒标记物lamp-1的易位增强。五价(trimab)和八价(okta)的三特异性抗体构建体的应用诱导显著更高水平的细胞毒性活化,其中trimab是在83%的cd8+t细胞中诱导脱粒的最有效的构建体。用cd28活化抗体和未经处理的对照pbmc的单独处理未显示lamp-1的易位。
(b)指示细胞毒性活化的颗粒酶b的表达。trimab和okta三特异性抗体构建体诱导显著更高水平的细胞毒性活化,其中trimab是在95%的cd8+t细胞中诱导颗粒酶b表达的最有效的构建体。
图5:来自健康供体的初始t细胞被trimab活化。在trimab存在下,当与hbsag阳性(huh7s)或hbsag阴性(huh7)的靶细胞共培养时,在来自健康供体的pbmc中的ifnγ分泌。只有当添加trimab并且靶细胞表达hbsag时,检测到通过ifnγ的分泌确定的t细胞的活化。
图6:来自健康供体的初始t细胞被活化以通过trimab杀伤hbsag阳性靶细胞。来自健康供体的pbmc与hbsag阴性(huh7)和hbsag阳性靶细胞(huh7s)的共培养。添加trimab后,通过xcelligence分析确定随时间的细胞活力,三次重复。在pbmc对照中,细胞与huh7s细胞共培养,但不添加抗体。只有当添加trimab并且靶细胞表达hbsag时,才在添加抗体后24小时开始观察到特异性杀伤。
图7:中和抗hbs抗体与通过trimab活化t细胞竞争。在含有中和αhbs抗体的人血清存在下,经trimab处理的pbmc与hbsag阳性huh7(huh7s)细胞共培养。0%:没有含抗体血清的共培养;50%:一半的fcs被含抗体的人血清代替,得到1175iu/l的αhbs浓度;100%:用含抗体的血清代替fcs,得到235iu/l的αhbs浓度。(a)不添加中和抗体,t细胞被活化并分泌ifnγ和其他细胞因子,(b)t细胞被活化以在典型的时间过程中杀伤靶细胞,导致在72小时内细胞活力50%的损失。中和抗体与trimab活性竞争,导致t细胞活化和细胞毒性的部分或甚至完全损失。
图8:当被trimab活化时,来自患有慢性乙型肝炎患者的pbmc杀伤hbsag阳性靶细胞。来自患有慢性乙型肝炎(chb)患者的pbmc与hbsag阴性(huh7)和hbsag阳性靶细胞(huh7s)的共培养。添加trimab后,通过xcelligence分析确定随时间的细胞活力,三次重复。在pbmc对照中,细胞与huh7s细胞共培养,但不添加抗体。只有当添加trimab并且靶细胞表达hbsag时,才在添加抗体后24至30小时开始观察到特异性杀伤。
图9:来自患有慢性乙型肝炎患者的pbmc被trimab激活。在trimab存在下与hbsag阳性(huh7s)或hbsag阴性(huh7)靶细胞共培养,来自患有慢性乙型肝炎(chb)患者的pbmcs的ifnγ分泌。只有当添加trimab并且靶细胞表达hbsag时,才检测到通过ifnγ的分泌确定的t细胞的活化。
实施例举例说明本发明。
实施例1
材料和方法
三特异性抗体的克隆和制备
使用覆盖所有vh和vκ/vλ亚型的一组引物通过来自各个杂交瘤的逆转录mrna的pcr扩增获得编码抗cd3(okt3)和抗cd28(9.3)的可变重链和可变轻链的互补dna。将pcr产物连接到pcr2.1-topo(invitrogen,lifetechnologies)中并测序。使用噬菌体展示技术从乙型肝炎疫苗接种者的b细胞的cdna库选择抗hbsagscfvc8并进行密码子优化。使用含有在5'和3'两侧中合适的限制性位点的引物,将编码上述抗体的可变重链和可变轻链cdna用甘氨酸-丝氨酸接头组装到scfv中。为了获得五价分子,将okt3和9.3scfv融合到抗体5f9的fab片段的c末端,同时将scfvc8的两个实体添加至其n-末端。为了获得八价分子,将四个scfvc8结构域和两个结构域okt3和9.3连接到igg样分子。
在所提及的5'甘氨酸-丝氨酸接头的5'末端克隆c8scfv编码序列。将完整的scfv-接头-higg1fc-接头-scfv序列亚克隆到哺乳动物表达载体pcdna3.1(-)(invitrogen)中。使用peqfect转染试剂(peqlab)用maxi-prep质粒dna转染hek293细胞。使用0.8-1.0mg/mlg418选择稳定的转染子并扩增。收集来自hek转染子的上清液,并通过elisa分析分泌的三特异性抗体的浓度,并使用山羊抗人igg-fc特异性的过氧化物酶标记抗体通过蛋白印迹分析所分泌抗体的完整性。
细胞培养条件和huh7s细胞的产生
将huh7肝癌细胞(nakabayaski等人,1982.growthofhumanhepatomacelllineswithdifferentiatedfunctionsinchemicallydefinedmedium.cancerres.42:3858-3863)和hek293细胞保存在补充有10%的牛胎儿血清(fbs)、青霉素(100u/ml)、链霉素(100μg/ml)和l-谷氨酰胺(2毫摩尔/l)的dulbecco改良eagle培养基(dmem)中(均来自gibco,lifetechnologies)。
为了产生huh7s细胞,使用fugene转染试剂(promega)利用编码hbv基因型d亚型ayw1的小hbv表面抗原的质粒和抗生素抗性基因转染huh-7细胞。抗生素选择后,分离单细胞克隆并在24孔板中生长。选择具有最高hbsag分泌的克隆,用于建立用作靶细胞的细胞系huh7-s。
pbmc的分离
使用lsm1077淋巴细胞分离培养基(paa)通过密度梯度离心从肝素化全血分离外周血单核细胞(pbmc)。使25ml血液在13ml的lsm1077上分层。在室温下以2000rpm离心20分钟(无间歇)后收获pbmc,并在补充有10%牛胎儿血清(fbs)、青霉素(100u/ml)、链霉素(100μg/ml)和l-谷氨酰胺(2毫摩尔/l)(均来自gibco)的rpmi1640培养基中培养。在37℃下在5%co2中静置过夜步骤后,将pbmc用于共培养实验。
pbmc在固定的hbsag上的培养
将在pbs(5μg/ml)中的100μl重组体hbsag转移到96孔板中,并在4℃下包被过夜。第二天将hbsag溶液轻轻倒出,并用磷酸盐缓冲液洗涤板。将1×105个pbmc/孔转移到hbsag包被的板上并与100μl含有三特异性抗体的上清液混合。没有固定的hbsag的孔作为阴性对照。将pbmc在结合了hbsag的板上培养48小时,然后收获上清液和细胞用于进一步分析。使用人ifn-γelisamaxtm(biolegend)通过elisa在上清液中检测由效应t细胞活化导致的细胞因子分泌。用针对lamp-1和颗粒酶b的抗体染色细胞,并通过流式细胞术进行分析。所有实验重复三次地进行。
hbv阳性靶细胞和重定向效应细胞的共培养
将huh7或huh7s靶细胞在96孔板中培养融合。在每孔100μl体积的培养基中添加1×105个pbmc细胞。每孔应用100μl含有三特异性抗体的hek上清液。为了确定cd3和cd28活化的协同作用,每孔添加50μl的每种双特异性抗体上清液。将仅用效应细胞(未添加抗体)孵育的未经处理的靶细胞作为阴性对照。在96或120小时后,收集上清液,并通过elisa分析ifnγ分泌(见上文)。
为了确定细胞毒性,将表达hbsag的huh7s细胞在与xcellingenceratc系统(acea,bioscience,inc.)兼容的96孔e板中培养,直到它们实现融合。将新鲜分离的pbmc和100μl含抗体的上清液添加在每组三个复孔中。未转基因的huh7细胞作为特异性对照。没有抗体的与huh7s细胞共培养的pbmc作为阴性对照。使用xcellingenceratc系统实时监控靶细胞的细胞活力120小时。
为了确定三特异性构建体活化来自慢性乙型肝炎患者ctl的能力,我们分离来自慢性携带者的pbmc,并将其用于实验。
实施例2
通过针对hbsag的五价和八价的三特异性抗体构建体活化ctl
在第一组实验中,已经评估了针对ctl表面抗原cd3和cd28和hbv的小表面抗原的五价和八价的三特异性抗体构建体的活性。我们将重组体hbsag包被到细胞培养皿的塑料表面上。向这些经包被的孔添加免疫效应细胞、即从健康供体分离的pbmc,和双特异性以及三特异性的抗体构建体。作为阴性对照,在双特异性或三特异性抗体构建体不存在的情况下在hbsag包被的孔上培养pbmc以评估非特异性的免疫细胞活化。
这些实验显示出在cd3活化的双特异性构建体存在下共培养后细胞毒性t淋巴细胞(ctl)的特异性活化,如通过最高5000pg/ml的促炎细胞因子干扰素γ(ifnγ)的分泌确定的(图3)。共同施用cd3特异性和cd28特异性的构建体后,该效应进一步得到增强,这证明了协同效应,而cd28活化的构建体靠其自身不能激活t细胞。
出人意料地,将cd3和cd28刺激和共刺激结构域结合到三特异性分子中显示出比两种双特异性构建体的组合显著更强的ctl活化(图3),如通过ifn分泌确定的。当在染色后通过流式细胞术来分析免疫细胞的细胞毒性潜力时,确认了这一点,因为lamp-1易位指示在由三特异性抗体构建体活化的绝大多数细胞中细胞毒性囊泡脱粒和颗粒酶b的表达(图4a和图b))。
因此,五价的构建体被证明是最有效的,并且能够在80%至95%的免疫细胞中诱导细胞毒性功能(图4)。由于pbmc含有约70%的t细胞,但它们中大多数是初始t细胞,ctl的活化需要tcr复合物的强烈活化。
实施例3
通过三特异性抗体构建体活化ctl后hbsag阳性肝细胞的特异性杀伤
在第二组实验中,已经评估了针对ctl表面抗原cd3和cd28和hbv小表面抗原的三特异性抗体构建体特异性杀伤表达hbsag的细胞的能力。将产生hbv表面抗原的稳定转染的huh7肝癌细胞系(huh7s细胞)用作靶细胞。在确定hbv蛋白表达后,将这些靶细胞与免疫效应细胞、即从健康供体分离的pbmc和五价的三特异性抗体构建体共培养。作为阴性对照,分析了与hbv阴性靶细胞的共培养物,其已经用含有hbsag的上清液进行了预培养。该对照用于排除由于hbv颗粒在hbv-阴性靶细胞表面上的非特异性结合的效应细胞活化。此外,在不存在三特异性抗体构建体的情况下,将hbv阳性靶细胞与免疫效应细胞共培养以评估非特异性背景细胞毒性。
这些实验显示出在cd3活化和cd28活化的五价的三特异性构建体存在下,与huh7s细胞而非huh7细胞共培养后的ctl的特异性活化。通过促炎细胞因子干扰素γ(ifn-γ)的分泌确定ctl活化最高为2ng/ml。在不存在三特异性分子的情况下,没有观察到ctl活化(图5)。
此外,三特异性构建体介导了产生hbsag的huh7肝癌细胞系的特异性细胞毒性消除(图6),如通过与huh7细胞或在不添加抗体构建体下孵育的对照相比,huh7s靶细胞活力>90%的降低所证明的。
为了进一步证明ctl活化的hbsag特异性,进行了含有中和抗hbs抗体的人血清的竞争试验(图7)。实验显示当添加117.5iu/l抗hbs时,只有ctl活化以及靶细胞杀伤的部分竞争,而杀伤效果在235iu/l抗hbs的浓度下完全消除。
实施例4
来自慢性乙型肝炎患者的t细胞的重定向和活化
为了分析三特异性抗体构建体在慢性乙型肝炎患者中活化ctl功能的能力,从高病毒血症乙型肝炎病毒携带者的患者分离pbmc。将产生hbv表面抗原的稳定转染的huh7肝癌细胞系(huh7s细胞)用作靶细胞。在存在和不存在五价的三特异性抗体构建体的情况下,将这些靶细胞与从乙型肝炎患者分离的pbmc共培养。分析阴性对照与hbv阴性靶细胞的共培养物。
这些实验表明,通过cd3活化和cd28活化五价的三特异性构建体在慢性乙型肝炎患者的pbmc中活化的ctl特异性杀伤huh7s细胞,而不是huh7细胞(图8)。在没有三特异性抗体构建体的情况下,没有观察到杀伤,这证明该方法的特异性和在患者的pbmc中缺少自发的ctl活性。促炎细胞因子干扰素γ(ifn-γ)的分泌证实特异性ctl活化(图9)。
序列表
<110>亥姆霍兹慕尼黑中心-德国环境健康研究中心等
<120>用于治疗hbv感染和相关病状病症的三特异性结合分子
<130>y1213pct
<150>ep15159274.8
<151>2015-03-16
<160>51
<170>bissap1.2
<210>1
<211>8
<212>prt
<213>人工序列
<220>
<223>scfvc8重链cdr1
<400>1
glyphethrpheserglytyrala
15
<210>2
<211>8
<212>prt
<213>人工序列
<220>
<223>scfvc8重链cdr2
<400>2
ileserglyserglyglyserthr
15
<210>3
<211>21
<212>prt
<213>人工序列
<220>
<223>scfvc8重链cdr3
<400>3
alalysproproglyargglnglutyrtyrglyserseriletyrtyr
151015
pheproleuglyasn
20
<210>4
<211>6
<212>prt
<213>人工序列
<220>
<223>scfvc8轻链cdr1
<400>4
asnileglyserlysser
15
<210>5
<211>3
<212>prt
<213>人工序列
<220>
<223>scfvc8轻链cdr2
<400>5
aspaspser
1
<210>6
<211>11
<212>prt
<213>人工序列
<220>
<223>scfvc8轻链cdr3
<400>6
glnvaltrpaspserserseraspleuvalval
1510
<210>7
<211>8
<212>prt
<213>人工序列
<220>
<223>scfv5f9重链cdr1
<400>7
glyphethrpheasnasntyrala
15
<210>8
<211>8
<212>prt
<213>人工序列
<220>
<223>scfv5f9重链cdr2
<400>8
ileasnseraspglyargserthr
15
<210>9
<211>8
<212>prt
<213>人工序列
<220>
<223>scfv5f9重链cdr3
<400>9
alaargthrphetyralaasptyr
15
<210>10
<211>6
<212>prt
<213>人工序列
<220>
<223>scfv5f9轻链cdr1
<400>10
glnasnvalaspthrthr
15
<210>11
<211>3
<212>prt
<213>人工序列
<220>
<223>scfv5f9轻链cdr2
<400>11
trpalaser
1
<210>12
<211>9
<212>prt
<213>人工序列
<220>
<223>scfv5f9轻链cdr3
<400>12
glnglntyrserilepheprotyrthr
15
<210>13
<211>8
<212>prt
<213>人工序列
<220>
<223>scfv5a19重链cdr1
<400>13
glyphethrphesersertyrala
15
<210>14
<211>8
<212>prt
<213>人工序列
<220>
<223>scfv5a19重链cdr2
<400>14
valserseraspglysertyrala
15
<210>15
<211>9
<212>prt
<213>人工序列
<220>
<223>scfv5a19重链cdr3
<400>15
alaserpheasntrpaspvalalatyr
15
<210>16
<211>12
<212>prt
<213>人工序列
<220>
<223>scfv5a19轻链cdr1
<400>16
glnserleuleuasnthrargthrarglyssertyr
1510
<210>17
<211>3
<212>prt
<213>人工序列
<220>
<223>scfv5a19轻链cdr2
<400>17
trpalaser
1
<210>18
<211>8
<212>prt
<213>人工序列
<220>
<223>scfv5a19轻链cdr3
<400>18
lysglnsertyrserleutyrthr
15
<210>19
<211>8
<212>prt
<213>人工序列
<220>
<223>scfvokt3重链cdr1
<400>19
glytyrthrphethrargtyrthr
15
<210>20
<211>8
<212>prt
<213>人工序列
<220>
<223>scfvokt3重链cdr2
<400>20
ileasnproserargglytyrthr
15
<210>21
<211>12
<212>prt
<213>人工序列
<220>
<223>scfvokt3重链cdr3
<400>21
alaargtyrtyraspasphistyrcysleuasptyr
1510
<210>22
<211>5
<212>prt
<213>人工序列
<220>
<223>scfvokt3轻链cdr1
<400>22
serservalsertyr
15
<210>23
<211>3
<212>prt
<213>人工序列
<220>
<223>scfvokt3轻链cdr2
<400>23
aspthrser
1
<210>24
<211>9
<212>prt
<213>人工序列
<220>
<223>scfvokt3轻链cdr3
<400>24
glnglntrpserserasnprophethr
15
<210>25
<211>8
<212>prt
<213>人工序列
<220>
<223>scfv9.3重链cdr1
<400>25
glypheserleuserasptyrgly
15
<210>26
<211>7
<212>prt
<213>人工序列
<220>
<223>scfv9.3重链cdr2
<400>26
iletrpalaglyglyglythr
15
<210>27
<211>14
<212>prt
<213>人工序列
<220>
<223>scfv9.3重链cdr3
<400>27
alaargasplysglytyrsertyrtyrtyrsermetasptyr
1510
<210>28
<211>10
<212>prt
<213>人工序列
<220>
<223>scfv9.3轻链cdr1
<400>28
gluservalglutyrtyrvalthrserleu
1510
<210>29
<211>3
<212>prt
<213>人工序列
<220>
<223>scfv9.3轻链cdr2
<400>29
alaalaser
1
<210>30
<211>9
<212>prt
<213>人工序列
<220>
<223>scfv9.3轻链cdr3
<400>30
glnglnserarglysvalprotyrthr
15
<210>31
<211>8
<212>prt
<213>人工序列
<220>
<223>scfva9重链cdr1
<400>31
glytyrthrphethrasntyrtrp
15
<210>32
<211>8
<212>prt
<213>人工序列
<220>
<223>scfva9重链cdr2
<400>32
iletyrproglyglyglytyrthr
15
<210>33
<211>10
<212>prt
<213>人工序列
<220>
<223>scfva9重链cdr3
<400>33
alaargseralasertrptyrpheaspval
1510
<210>34
<211>9
<212>prt
<213>人工序列
<220>
<223>scfva9轻链cdr1
<400>34
thrglythrvalthrthrserasntyr
15
<210>35
<211>3
<212>prt
<213>人工序列
<220>
<223>scfva9轻链cdr2
<400>35
histhrasn
1
<210>36
<211>9
<212>prt
<213>人工序列
<220>
<223>scfva9轻链cdr3
<400>36
alaleutrptyrasnasnhistrpval
15
<210>37
<211>8
<212>prt
<213>人工序列
<220>
<223>scfvncam29.2重链cdr1
<400>37
glyphethrpheserserphegly
15
<210>38
<211>8
<212>prt
<213>人工序列
<220>
<223>scfvncam29.2重链cdr2
<400>38
ileserserglysertyralaile
15
<210>39
<211>14
<212>prt
<213>人工序列
<220>
<223>scfvncam29.2重链cdr3
<400>39
valargglyargargleuglygluglytyralametasptyr
1510
<210>40
<211>12
<212>prt
<213>人工序列
<220>
<223>scfvncam29.2轻链cdr1
<400>40
glnserleuleutyrserserasnglnlysasntyr
1510
<210>41
<211>3
<212>prt
<213>人工序列
<220>
<223>scfvncam29.2轻链cdr2
<400>41
trpalaser
1
<210>42
<211>7
<212>prt
<213>人工序列
<220>
<223>scfvncam29.2轻链cdr3
<400>42
glnglntyrsersertrpthr
15
<210>43
<211>794
<212>prt
<213>人工序列
<220>
<223>抗hbsagscfv[c8]^甘氨酸-丝氨酸-连接子^higg1-fc[c245s,l259f,l26
0e,g262a,a355t]^链霉素标记^甘氨酸-连接子^抗-cd3scfv[okt3]
<400>43
metaspphegluvalglnilepheserpheleuleuileseralaser
151015
valilemetserargmetalagluvalglnleuvalgluserglygly
202530
glyleuleuglnproglyglyserleuargleusercysalaalaser
354045
glyphethrpheserglytyralametsertrpvalargglnalapro
505560
glylysglyleuglutrpvalserserileserglyserglyglyser
65707580
thrtyrtyralaaspservallysglyargphethrileserargasp
859095
asnserlysasnthrleutyrleuglnmetasnserleuargalaglu
100105110
aspthralaleutyrtyrcysalalysproproglyargglnglutyr
115120125
tyrglyserseriletyrtyrpheproleuglyasntrpglyglngly
130135140
thrleuvalthrvalserseralaserthrlysglyprolysleuglu
145150155160
gluglyglupheserglualaargvalglnseralaleuthrglnpro
165170175
alaservalservalalaproglyglnthralaargilethrcysgly
180185190
glyasnasnileglyserlysservalhistrptyrglnglnlyspro
195200205
glyglnalaprovalleuvalvaltyraspaspseraspargproser
210215220
glyileprogluargpheserglyserasnserglyasnthralathr
225230235240
leuthrileserargvalglualaglyaspglualaasptyrtyrcys
245250255
glnvaltrpaspserserseraspleuvalvalpheglyglyglythr
260265270
lysleuthrvalleuglyasnserglyglyglyglyserglyglygly
275280285
glyserglyglyglyglyseralasergluprolysserserasplys
290295300
thrhisthrcysproprocysproalaprogluphegluglyalapro
305310315320
servalpheleupheproprolysprolysaspthrleumetileser
325330335
argthrprogluvalthrcysvalvalvalaspvalserhisgluasp
340345350
progluvallyspheasntrptyrvalaspglyvalgluvalhisasn
355360365
alalysthrlysproargglugluglntyrasnserthrtyrargval
370375380
valservalleuthrvalleuhisglnasptrpleuasnglylysglu
385390395400
tyrlyscyslysvalserasnlysalaleuprothrproileglulys
405410415
thrileserlysalalysglyglnproarggluproglnvaltyrthr
420425430
leuproproserargaspgluleuthrlysasnglnvalserleuthr
435440445
cysleuvallysglyphetyrproseraspilealavalglutrpglu
450455460
serasnglyglnprogluasnasntyrlysthrthrproprovalleu
465470475480
aspseraspglyserphepheleutyrserlysleuthrvalasplys
485490495
serargtrpglnglnglyasnvalphesercysservalmethisglu
500505510
alaleuhisasnhistyrthrglnlysserleuserleuserprogly
515520525
lysaspproglytrpserhisproglnpheglulysserargglygly
530535540
glyglyglnvalglnleuglnglnserglyalagluleualaargpro
545550555560
glyalaservallysmetsercyslysalaserglytyrthrphethr
565570575
argtyrthrmethistrpvallysglnargproglyglnglyleuglu
580585590
trpileglytyrileasnproserargglytyrthrasntyrasngln
595600605
lysphelysasplysalathrleuthrthrasplysserserserthr
610615620
alatyrmetglnleuserserleuthrsergluaspseralavaltyr
625630635640
tyrcysalaargtyrtyraspasphistyrcysleuasptyrtrpgly
645650655
glnglythrthrleuthrvalserserglyasnserglyglyglygly
660665670
serglyglyglyglyserglyglyglyglyseralaserglnileval
675680685
leuthrglnserproalailemetseralaserproglyglulysval
690695700
thrmetthrcysseralaserserservalsertyrmetasntrptyr
705710715720
glnglnlysserglythrserprolysargtrpiletyraspthrser
725730735
lysleualaserglyvalproalahispheargglyserglysergly
740745750
thrsertyrserleuthrileserglymetglualagluaspalaala
755760765
thrtyrtyrcysglnglntrpserserasnprophethrpheglyser
770775780
glythrlysleugluileasnglyasnser
785790
<210>44
<211>793
<212>prt
<213>人工序列
<220>
<223>抗-hbsagscfv[c8]^甘氨酸-丝氨酸-连接子^higg1-fc[c245s,l259f,l26
0e,g262a,a355t]^链霉素标记^甘氨酸-连接子^抗-cd28scfv[9.3]
<400>44
metaspphegluvalglnilepheserpheleuleuileseralaser
151015
valilemetserargmetalagluvalglnleuvalgluserglygly
202530
glyleuleuglnproglyglyserleuargleusercysalaalaser
354045
glyphethrpheserglytyralametsertrpvalargglnalapro
505560
glylysglyleuglutrpvalserserileserglyserglyglyser
65707580
thrtyrtyralaaspservallysglyargphethrileserargasp
859095
asnserlysasnthrleutyrleuglnmetasnserleuargalaglu
100105110
aspthralaleutyrtyrcysalalysproproglyargglnglutyr
115120125
tyrglyserseriletyrtyrpheproleuglyasntrpglyglngly
130135140
thrleuvalthrvalserseralaserthrlysglyprolysleuglu
145150155160
gluglyglupheserglualaargvalglnseralaleuthrglnpro
165170175
alaservalservalalaproglyglnthralaargilethrcysgly
180185190
glyasnasnileglyserlysservalhistrptyrglnglnlyspro
195200205
glyglnalaprovalleuvalvaltyraspaspseraspargproser
210215220
glyileprogluargpheserglyserasnserglyasnthralathr
225230235240
leuthrileserargvalglualaglyaspglualaasptyrtyrcys
245250255
glnvaltrpaspserserseraspleuvalvalpheglyglyglythr
260265270
lysleuthrvalleuglyasnserglyglyglyglyserglyglygly
275280285
glyserglyglyglyglyseralasergluprolysserserasplys
290295300
thrhisthrcysproprocysproalaprogluphegluglyalapro
305310315320
servalpheleupheproprolysprolysaspthrleumetileser
325330335
argthrprogluvalthrcysvalvalvalaspvalserhisgluasp
340345350
progluvallyspheasntrptyrvalaspglyvalgluvalhisasn
355360365
alalysthrlysproargglugluglntyrasnserthrtyrargval
370375380
valservalleuthrvalleuhisglnasptrpleuasnglylysglu
385390395400
tyrlyscyslysvalserasnlysalaleuprothrproileglulys
405410415
thrileserlysalalysglyglnproarggluproglnvaltyrthr
420425430
leuproproserargaspgluleuthrlysasnglnvalserleuthr
435440445
cysleuvallysglyphetyrproseraspilealavalglutrpglu
450455460
serasnglyglnprogluasnasntyrlysthrthrproprovalleu
465470475480
aspseraspglyserphepheleutyrserlysleuthrvalasplys
485490495
serargtrpglnglnglyasnvalphesercysservalmethisglu
500505510
alaleuhisasnhistyrthrglnlysserleuserleuserprogly
515520525
lysaspproglytrpserhisproglnpheglulysserserglygly
530535540
glyglyglnvalglnleuglngluserglyproglyleuvalthrpro
545550555560
serglnserleuserilethrcysthrvalserglypheserleuser
565570575
asptyrglyvalhistrpvalargglnserproglyglnglyleuglu
580585590
trpleuglyvaliletrpalaglyglyglythrasntyrasnserala
595600605
leumetserarglysserileserlysaspasnserlysserglnval
610615620
pheleulysmetasnserleuglnalaaspaspthralavaltyrtyr
625630635640
cysalaargasplysglytyrsertyrtyrtyrsermetasptyrtrp
645650655
glyglnglythrthrvalthrvalserserargglyglyglysergly
660665670
glyglyglyserglyglyglyglyseraspilegluleuthrglnser
675680685
proalaserleualavalserleuglyglnargalathrilesercys
690695700
argalasergluservalglutyrtyrvalthrserleumetglntrp
705710715720
tyrglnglnlysproglyglnproprolysleuleuilephealaala
725730735
serasnvalgluserglyvalproalaargpheserglyserglyser
740745750
glythrasnpheserleuasnilehisprovalaspgluaspaspval
755760765
alamettyrphecysglnglnserarglysvalprotyrthrphegly
770775780
glyglythrlysleugluilelysarg
785790
<210>45
<211>792
<212>prt
<213>人工序列
<220>
<223>抗-hbsagscfv[c8]^甘氨酸-丝氨酸-连接子^higg1-fc[c245s,l259f,l26
0e,g262a,a355t]^链霉素标记^甘氨酸-连接子^链霉素标记^抗-cd16scfv
[a9]
<400>45
metaspphegluvalglnilepheserpheleuleuileseralaser
151015
valilemetserargmetalagluvalglnleuvalgluserglygly
202530
glyleuleuglnproglyglyserleuargleusercysalaalaser
354045
glyphethrpheserglytyralametsertrpvalargglnalapro
505560
glylysglyleuglutrpvalserserileserglyserglyglyser
65707580
thrtyrtyralaaspservallysglyargphethrileserargasp
859095
asnserlysasnthrleutyrleuglnmetasnserleuargalaglu
100105110
aspthralaleutyrtyrcysalalysproproglyargglnglutyr
115120125
tyrglyserseriletyrtyrpheproleuglyasntrpglyglngly
130135140
thrleuvalthrvalserseralaserthrlysglyprolysleuglu
145150155160
gluglyglupheserglualaargvalglnseralaleuthrglnpro
165170175
alaservalservalalaproglyglnthralaargilethrcysgly
180185190
glyasnasnileglyserlysservalhistrptyrglnglnlyspro
195200205
glyglnalaprovalleuvalvaltyraspaspseraspargproser
210215220
glyileprogluargpheserglyserasnserglyasnthralathr
225230235240
leuthrileserargvalglualaglyaspglualaasptyrtyrcys
245250255
glnvaltrpaspserserseraspleuvalvalpheglyglyglythr
260265270
lysleuthrvalleuglyasnserglyglyglyglyserglyglygly
275280285
glyserglyglyglyglyseralasergluprolysserserasplys
290295300
thrhisthrcysproprocysproalaprogluphegluglyalapro
305310315320
servalpheleupheproprolysprolysaspthrleumetileser
325330335
argthrprogluvalthrcysvalvalvalaspvalserhisgluasp
340345350
progluvallyspheasntrptyrvalaspglyvalgluvalhisasn
355360365
alalysthrlysproargglugluglntyrasnserthrtyrargval
370375380
valservalleuthrvalleuhisglnasptrpleuasnglylysglu
385390395400
tyrlyscyslysvalserasnlysalaleuprothrproileglulys
405410415
thrileserlysalalysglyglnproarggluproglnvaltyrthr
420425430
leuproproserargaspgluleuthrlysasnglnvalserleuthr
435440445
cysleuvallysglyphetyrproseraspilealavalglutrpglu
450455460
serasnglyglnprogluasnasntyrlysthrthrproprovalleu
465470475480
aspseraspglyserphepheleutyrserlysleuthrvalasplys
485490495
serargtrpglnglnglyasnvalphesercysservalmethisglu
500505510
alaleuhisasnhistyrthrglnlysserleuserleuserprogly
515520525
lysaspproglytrpserhisproglnpheglulysserargglygly
530535540
glyglyglnvalglnleuglnglnserglyalagluleuvalargpro
545550555560
glythrservallysilesercyslysalaserglytyrthrphethr
565570575
asntyrtrpleuglytrpvallysglnargproglyhisglyleuglu
580585590
trpileglyaspiletyrproglyglyglytyrthrasntyrasnglu
595600605
lysphelysglylysalathrvalthralaaspthrserserargthr
610615620
alatyrvalglnvalargserleuthrsergluaspseralavaltyr
625630635640
phecysalaargseralasertrptyrpheaspvaltrpglyalagly
645650655
thrthrvalthrvalserserglyasnserglyglyglyglysergly
660665670
glyglyglyserglyglyglyglyseralaserglnalavalvalthr
675680685
glngluseralaleuthrthrserproglygluthrvalthrleuthr
690695700
cysargserasnthrglythrvalthrthrserasntyralaasntrp
705710715720
valglnglulysproasphisleuphethrglyleuileglyhisthr
725730735
asnasnargalaproglyvalproalaargpheserglyserleuile
740745750
glyasplysalaalaleuthrilethrglyalaglnthrgluaspglu
755760765
alailetyrphecysalaleutrptyrasnasnhistrpvalphegly
770775780
glyglythrlysleuthrvalleu
785790
<210>46
<211>799
<212>prt
<213>人工序列
<220>
<223>抗-hbsagscfv[c8]^甘氨酸-丝氨酸-连接子^higg1-fc[c245s,l259f,l26
0e,g262a,a355t]^链霉素标记^甘氨酸-连接子^链霉素标记^抗-cd56scfv
[ncam29.2]
<400>46
metaspphegluvalglnilepheserpheleuleuileseralaser
151015
valilemetserargmetalagluvalglnleuvalgluserglygly
202530
glyleuleuglnproglyglyserleuargleusercysalaalaser
354045
glyphethrpheserglytyralametsertrpvalargglnalapro
505560
glylysglyleuglutrpvalserserileserglyserglyglyser
65707580
thrtyrtyralaaspservallysglyargphethrileserargasp
859095
asnserlysasnthrleutyrleuglnmetasnserleuargalaglu
100105110
aspthralaleutyrtyrcysalalysproproglyargglnglutyr
115120125
tyrglyserseriletyrtyrpheproleuglyasntrpglyglngly
130135140
thrleuvalthrvalserseralaserthrlysglyprolysleuglu
145150155160
gluglyglupheserglualaargvalglnseralaleuthrglnpro
165170175
alaservalservalalaproglyglnthralaargilethrcysgly
180185190
glyasnasnileglyserlysservalhistrptyrglnglnlyspro
195200205
glyglnalaprovalleuvalvaltyraspaspseraspargproser
210215220
glyileprogluargpheserglyserasnserglyasnthralathr
225230235240
leuthrileserargvalglualaglyaspglualaasptyrtyrcys
245250255
glnvaltrpaspserserseraspleuvalvalpheglyglyglythr
260265270
lysleuthrvalleuglyasnserglyglyglyglyserglyglygly
275280285
glyserglyglyglyglyseralasergluprolysserserasplys
290295300
thrhisthrcysproprocysproalaprogluphegluglyalapro
305310315320
servalpheleupheproprolysprolysaspthrleumetileser
325330335
argthrprogluvalthrcysvalvalvalaspvalserhisgluasp
340345350
progluvallyspheasntrptyrvalaspglyvalgluvalhisasn
355360365
alalysthrlysproargglugluglntyrasnserthrtyrargval
370375380
valservalleuthrvalleuhisglnasptrpleuasnglylysglu
385390395400
tyrlyscyslysvalserasnlysalaleuprothrproileglulys
405410415
thrileserlysalalysglyglnproarggluproglnvaltyrthr
420425430
leuproproserargaspgluleuthrlysasnglnvalserleuthr
435440445
cysleuvallysglyphetyrproseraspilealavalglutrpglu
450455460
serasnglyglnprogluasnasntyrlysthrthrproprovalleu
465470475480
aspseraspglyserphepheleutyrserlysleuthrvalasplys
485490495
serargtrpglnglnglyasnvalphesercysservalmethisglu
500505510
alaleuhisasnhistyrthrglnlysserleuserleuserprogly
515520525
lysaspproglytrpserhisproglnpheglulysserserglygly
530535540
glyglyaspvalglnleuvalgluserglyglyglyleuvalglnpro
545550555560
glyglyserarglysleusercysalaalaserglyphethrpheser
565570575
serpheglymethistrpvalargglnalaproglulysglyleuglu
580585590
trpvalalatyrileserserglysertyralailetyrtyralaasp
595600605
thrvallysglyargphethrileserargaspasnprogluasnthr
610615620
leupheleuglnmetthrserleuargsergluaspseralamettyr
625630635640
tyrcysvalargglyargargleuglygluglytyralametasptyr
645650655
trpglyglnglythrservalthrvalserserglyasnserglygly
660665670
glyglyserglyglyglyglyserglyglyglyglyseralaserasp
675680685
ilevalmetserglnserproserserleualavalservalglyglu
690695700
lysvalthrmetsercyslysserserglnserleuleutyrserser
705710715720
asnglnlysasntyrleualatrptyrglnglnlysproglyglnser
725730735
prolysleuleuiletyrtrpalaserthrarglysserglyvalpro
740745750
aspargphethrglyserglyserglythraspphethrleuthrile
755760765
serservallysalagluaspleualavaltyrtyrcysglnglntyr
770775780
sersertrpthrpheglyglyglythrlysleugluilelysarg
785790795
<210>47
<211>15
<212>prt
<213>人工序列
<220>
<223>连接子
<400>47
glyglyglyglyserglyglyglyglyserglyglyglyglyser
151015
<210>48
<211>17
<212>prt
<213>人工序列
<220>
<223>连接子
<400>48
alalysthrthrprolysleuglugluglyglupheserglualaarg
151015
val
<210>49
<211>20
<212>prt
<213>人工序列
<220>
<223>连接子
<400>49
glyglyglyglyserglyglyglyglyserglyglyglyglysergly
151015
glyglyglyser
20
<210>50
<211>835
<212>prt
<213>人工序列
<220>
<223>具有jun亮氨酸拉链二聚体结构域的c8-higg1fcmut-okt3
<400>50
metaspphegluvalglnilepheserpheleuleuileseralaser
151015
valilemetserargmetalagluvalglnleuvalgluserglygly
202530
glyleuleuglnproglyglyserleuargleusercysalaalaser
354045
glyphethrpheserglytyralametsertrpvalargglnalapro
505560
glylysglyleuglutrpvalserserileserglyserglyglyser
65707580
thrtyrtyralaaspservallysglyargphethrileserargasp
859095
asnserlysasnthrleutyrleuglnmetasnserleuargalaglu
100105110
aspthralaleutyrtyrcysalalysproproglyargglnglutyr
115120125
tyrglyserseriletyrtyrpheproleuglyasntrpglyglngly
130135140
thrleuvalthrvalserseralaserthrlysglyprolysleuglu
145150155160
gluglyglupheserglualaargvalglnseralaleuthrglnpro
165170175
alaservalservalalaproglyglnthralaargilethrcysgly
180185190
glyasnasnileglyserlysservalhistrptyrglnglnlyspro
195200205
glyglnalaprovalleuvalvaltyraspaspseraspargproser
210215220
glyileprogluargpheserglyserasnserglyasnthralathr
225230235240
leuthrileserargvalglualaglyaspglualaasptyrtyrcys
245250255
glnvaltrpaspserserseraspleuvalvalpheglyglyglythr
260265270
lysleuthrvalleuglyasnserglyglyglyglyserglyglygly
275280285
glyserglyglyglyglyseralaserargilealaargleugluglu
290295300
lysvallysthrleulysalaglnasnsergluleualaserthrala
305310315320
asnmetleuarggluglnvalalaglnleulysglnlysvalmetasn
325330335
tyrsersergluprolysserserasplysthrhisthrcyspropro
340345350
cysproalaproproalaalaglyproservalpheleuphepropro
355360365
lysprolysaspthrleumetileserargthrprogluvalthrcys
370375380
valvalvalaspvalserhisgluaspprogluvallyspheasntrp
385390395400
tyrvalaspglyvalgluvalhisasnalalysthrlysproargglu
405410415
gluglntyrasnserthrtyrargvalvalservalleuthrvalleu
420425430
hisglnasptrpleuasnglylysglutyrlyscysalavalserasn
435440445
lysglyleualaserserileglulysthrileserlysalalysgly
450455460
glnproarggluproglnvaltyrthrleuproproserargaspglu
465470475480
leuthrlysasnglnvalserleuthrcysleuvallysglyphetyr
485490495
proseraspilealavalglutrpgluserasnglyglnprogluasn
500505510
asntyrlysthrthrproprovalleuaspseraspglyserphephe
515520525
leutyrserlysleuthrvalasplysserargtrpglnglnglyasn
530535540
valphesercysservalmethisglualaleuhisasnhistyrthr
545550555560
glnlysserleuserleuserproglylysaspproglytrpserhis
565570575
proglnpheglulysserargglyglyglyglyglnvalglnleugln
580585590
glnserglyalagluleualaargproglyalaservallysmetser
595600605
cyslysalaserglytyrthrphethrargtyrthrmethistrpval
610615620
lysglnargproglyglnglyleuglutrpileglytyrileasnpro
625630635640
serargglytyrthrasntyrasnglnlysphelysasplysalathr
645650655
leuthrthrasplysserserserthralatyrmetglnleuserser
660665670
leuthrsergluaspseralavaltyrtyrcysalaargtyrtyrasp
675680685
asphistyrcysleuasptyrtrpglyglnglythrthrleuthrval
690695700
serserglyasnserglyglyglyglyserglyglyglyglysergly
705710715720
glyglyglyseralaserglnilevalleuthrglnserproalaile
725730735
metseralaserproglyglulysvalthrmetthrcysseralaser
740745750
serservalsertyrmetasntrptyrglnglnlysserglythrser
755760765
prolysargtrpiletyraspthrserlysleualaserglyvalpro
770775780
alahispheargglyserglyserglythrsertyrserleuthrile
785790795800
serglymetglualagluaspalaalathrtyrtyrcysglnglntrp
805810815
serserasnprophethrpheglyserglythrlysleugluileasn
820825830
glyasnser
835
<210>51
<211>834
<212>prt
<213>人工序列
<220>
<223>具有fos亮氨酸拉链二聚体结构域的c8-higg1fcmut-9.3
<400>51
metaspphegluvalglnilepheserpheleuleuileseralaser
151015
valilemetserargmetalagluvalglnleuvalgluserglygly
202530
glyleuleuglnproglyglyserleuargleusercysalaalaser
354045
glyphethrpheserglytyralametsertrpvalargglnalapro
505560
glylysglyleuglutrpvalserserileserglyserglyglyser
65707580
thrtyrtyralaaspservallysglyargphethrileserargasp
859095
asnserlysasnthrleutyrleuglnmetasnserleuargalaglu
100105110
aspthralaleutyrtyrcysalalysproproglyargglnglutyr
115120125
tyrglyserseriletyrtyrpheproleuglyasntrpglyglngly
130135140
thrleuvalthrvalserseralaserthrlysglyprolysleuglu
145150155160
gluglyglupheserglualaargvalglnseralaleuthrglnpro
165170175
alaservalservalalaproglyglnthralaargilethrcysgly
180185190
glyasnasnileglyserlysservalhistrptyrglnglnlyspro
195200205
glyglnalaprovalleuvalvaltyraspaspseraspargproser
210215220
glyileprogluargpheserglyserasnserglyasnthralathr
225230235240
leuthrileserargvalglualaglyaspglualaasptyrtyrcys
245250255
glnvaltrpaspserserseraspleuvalvalpheglyglyglythr
260265270
lysleuthrvalleuglyasnserglyglyglyglyserglyglygly
275280285
glyserglyglyglyglyseralaserleuthraspthrleuglnala
290295300
gluthraspglnleugluasplyslysseralaleuglnthrgluile
305310315320
alaasnleuleulysglulysglulysleuglupheileleualaala
325330335
tyrsersergluprolysserserasplysthrhisthrcyspropro
340345350
cysproalaproproalaalaglyproservalpheleuphepropro
355360365
lysprolysaspthrleumetileserargthrprogluvalthrcys
370375380
valvalvalaspvalserhisgluaspprogluvallyspheasntrp
385390395400
tyrvalaspglyvalgluvalhisasnalalysthrlysproargglu
405410415
gluglntyrasnserthrtyrargvalvalservalleuthrvalleu
420425430
hisglnasptrpleuasnglylysglutyrlyscysalavalserasn
435440445
lysglyleualaserserileglulysthrileserlysalalysgly
450455460
glnproarggluproglnvaltyrthrleuproproserargaspglu
465470475480
leuthrlysasnglnvalserleuthrcysleuvallysglyphetyr
485490495
proseraspilealavalglutrpgluserasnglyglnprogluasn
500505510
asntyrlysthrthrproprovalleuaspseraspglyserphephe
515520525
leutyrserlysleuthrvalasplysserargtrpglnglnglyasn
530535540
valphesercysservalmethisglualaleuhisasnhistyrthr
545550555560
glnlysserleuserleuserproglylysaspproglytrpserhis
565570575
proglnpheglulysserserglyglyglyglyglnvalglnleugln
580585590
gluserglyproglyleuvalthrproserglnserleuserilethr
595600605
cysthrvalserglypheserleuserasptyrglyvalhistrpval
610615620
argglnserproglyglnglyleuglutrpleuglyvaliletrpala
625630635640
glyglyglythrasntyrasnseralaleumetserarglysserile
645650655
serlysaspasnserlysserglnvalpheleulysmetasnserleu
660665670
glnalaaspaspthralavaltyrtyrcysalaargasplysglytyr
675680685
sertyrtyrtyrsermetasptyrtrpglyglnglythrthrvalthr
690695700
valserserargglyglyglyserglyglyglyglyserglyglygly
705710715720
glyseraspilegluleuthrglnserproalaserleualavalser
725730735
leuglyglnargalathrilesercysargalasergluservalglu
740745750
tyrtyrvalthrserleumetglntrptyrglnglnlysproglygln
755760765
proprolysleuleuilephealaalaserasnvalgluserglyval
770775780
proalaargpheserglyserglyserglythrasnpheserleuasn
785790795800
ilehisprovalaspgluaspaspvalalamettyrphecysglngln
805810815
serarglysvalprotyrthrpheglyglyglythrlysleugluile
820825830
lysarg
- 还没有人留言评论。精彩留言会获得点赞!
- 用于治疗HBV感染和相关病症的三特异性结合分子的制造方法与工艺
- 对磺胺类药物具有特异性的固相微萃取纤维的制备方法与流程
- 一种检测猪初乳中抗PEDV特异性IgA抗体的夹心ELISA试剂盒的制造方法与工艺
- 用于施用CD19xCD3双特异性抗体的给药方案的制造方法与工艺
- 双特异性抗‑VEGF/抗‑ANG‑2抗体及其在治疗眼血管疾病中的应用的制造方法与工艺
- 特异性人EWS‑FLI1融合蛋白抗体的制备方法及在制备尤文肉瘤诊断抗体试剂的应用与制造工艺
- 双特异性四价抗体及其制造和使用方法与制造工艺
- 利用特异性挥发物筛选昆虫相关嗅觉蛋白的方法与制造工艺
- 使用与表面偶联的单特异性四聚抗体复合物组合物从样品中分离出靶实体的方法与制造工艺
- 抗-WT1/HLA双特异性抗体的制造方法与工艺