可以有益地调节针对呼吸道病毒感染的免疫应答的长双歧杆菌的制作方法
本发明涉及双歧杆菌,其是存在于人类结肠微生物区系(microflora)中的几种主要可培养的细菌之一。
发明背景
双歧杆菌(bifidobacteria)被认为是益生菌,因为当摄入足够数量时,它们是发挥超出基础营养的健康作用的生物体。高水平的摄入的双歧杆菌必须达到其作用部位才能发挥益生菌作用。已经提出了每克肠内容物的最低水平约为106-107个活双歧杆菌(bouhnik,y.,lait1993)。文献中有报道显示,在成人和婴儿中完成的体内研究表明,一些双歧杆菌菌株能够在通过胃肠道后存活。观察到不同双歧杆菌菌株之间耐受酸和胆盐的能力的显著差异,表明存活是选择潜在益生菌菌株的重要标准。
摄入双歧杆菌可改善胃肠通过,并可预防或协助治疗可能由微生物区系缺陷或受损造成的疾病,如胃肠道(git)感染、便秘、肠易激综合征(ibs)、炎性肠病(ibd)-克罗恩病和溃疡性结肠炎、食物过敏、抗生素诱导的腹泻、心血管疾病和某些癌症(如结肠直肠癌)。
病毒感染是发病率和死亡率的主要原因。流感病毒、鼻病毒(普通感冒)和呼吸道合胞病毒(rsv)是高度传染性的病毒,通常在健康个体中通过适当的宿主免疫应答成功清除。然而,这些感染的许多问题不在于最初的病毒,而是通常伴随它的继发感染,如导致肺炎、窦组织感染等的细菌感染。易感个体如患有copd或哮喘或肥胖的那些,对病毒感染的应答非常差,并且随后的继发感染是严重的并且可能对这些个体造成生命威胁。
这些病毒诱导的这些病症的恶化与巨额的医疗费用和显著的痛苦相关。因为存在大量的未满足的临床需求,为这些恶化开发新的和有效的疗法将是有益的。对受流感病毒(特别是大流行性禽流感病毒)影响的患者施用皮质类固醇虽然相对常见,但仍存在争议。常规使用类固醇不是流感病毒感染的理想选择,因为它不减少病毒负荷,也不抑制之后继发细菌感染的清除(yangetal,2015)。
在感染的早期阶段,流感病毒、鼻病毒和rsv的复制发生在肺上皮细胞中,导致病毒传感器的活化和抗病毒i型和iii型干扰素(ifn)以及趋化因子和细胞因子的释放。这些促炎介质有助于清除原发感染,但是i型干扰素也可导致后续损伤(免疫病理学)。已经表明,i型ifn应答如ifn-α和ifn-β分子的增加与流感感染模型中发病率和死亡率的增加直接相关(davidsonetal,2014)。抗病毒i型干扰素和相关的ip-10趋化因子的过量产生,抑制了清除继发感染的适当的免疫应答(nakamuraetal,2011;shahangianetal,2009;lietal,2012),所述继发感染由例如细菌物质(如肺炎链球菌(streptococcuspneumonia)、卡他莫拉菌(moraxellacatarrhalis)和流感嗜血杆菌(haemophilusinfluenzae),甚至金黄色葡萄球菌(staphylococcusaureus))引起(hewittetal,2016)。这些继发感染导致肺内过多的细胞死亡。在易感个体中,这会导致肺组织损伤和肺功能降低,在某些情况下,例如在copd患者中,会导致严重的并发症和死亡率。重要的是,促炎活化仅发生在引起嗜中性粒细胞募集的i型ifn应答中,而不发生在iii型ifn应答中(galanietal,2017)。
然而,iii型ifn如ifn-lambda(ifn-λ)可以限制病毒复制而不诱导促炎应答或免疫病理学(davidsonetal,2016;galanietal,2017)。
因此,可以限制针对病毒感染的i型ifn应答和伴随的促炎和组织损伤应答,同时保持适当的iii型应答和抗病毒防御的治疗剂,将是呼吸道病毒感染管理的显著进步。免疫病理学的减少对宿主存活和解决疾病至关重要。
其他固有的传感器或免疫介质也可在早期抗病毒防御中发挥重要作用。表面活性蛋白d(sp-d)可阻止流感病毒进入上皮细胞,这是感染早期阶段的一部分(thieletal,1989;sastryetal,1993)。同样,可与病毒结合并阻止其感染支气管上皮细胞的可诱导这种固有传感器治疗剂,将是治疗病毒如流感病毒、鼻病毒和rsv的显著进步。此外,抑制肿瘤坏死因子-α(tnf-α)(一种重要的炎性细胞因子)对肺免疫病理学的程度和抑制的炎性细胞浸润和细胞因子应答有显著影响。已表明,tnf-α与感染有高毒力流感病毒的猕猴和人类中的发病率和死亡率相关。当tnf-α受到抑制时,观察到流感病毒复制减少和流感病毒感染的小鼠的存活增加(shietal,2013)。用依那西普抑制这种炎性细胞因子在小鼠中提供了针对致死的h1n1流感感染的保护(shietal,2013)。
可诱导iii型ifn-λ和/或sp-d的治疗剂可在早期清除原发病毒感染中发挥作用,同时降低继发细菌感染的可能性,并将成为ards、哮喘、肥胖和copd管理中的显著进步。
技术实现要素:
根据本发明,提供了具有保藏号ncimb42020的长双歧杆菌(bifidobacteriumlongum)菌株。所述菌株可以是生物纯培养物的形式。
本发明还提供了长双歧杆菌ncimb42020的分离菌株。
该菌株可是活细胞的形式和/或非活细胞的形式。
该菌株可分离自健康人类对象的粪便或胃肠道。
该菌株可以是细菌培养液的形式。或者,该菌株是冻干粉末的形式。
该菌株减弱ip-10对病毒的应答。这很重要,因为ip-10是由几种细胞类型应答ifn-γ分泌的。这些细胞类型包括单核细胞和内皮细胞。ip-10被认为有几个角色,包括对单核细胞/巨噬细胞、t细胞、nk细胞和树突状细胞的趋化作用以及对t细胞粘附内皮细胞的促进作用,并且是病毒诱导的宿主系统活化的标志物。
该菌株可增强iii型干扰素对病毒的应答,如干扰素λ对病毒的应答。
该菌株可抑制i型干扰素对病毒的应答,如干扰素α对病毒的应答,和/或干扰素β对病毒的应答。
该菌株可增强表面活性蛋白d对病毒的应答。
本发明还提供了包含本发明的双歧杆菌菌株的制剂。
该制剂可进一步包含另外的益生菌材料和/或益生元材料。
该制剂可进一步包含可摄入的载体。该可摄入的载体可以是药学上可接受的载体,如胶囊、片剂或粉末。该可摄入的载体可以是食品,如酸化乳、酸奶、冷冻酸奶、冰淇淋、乳粉、浓缩乳、冰淇淋、奶酪酱、调味品或饮料。
该制剂可进一步包含蛋白质和/或肽(特别是富含谷氨酰胺/谷氨酸的蛋白质和/或肽)、脂质、碳水化合物、维生素、矿物质和/或微量元素。
双歧杆菌菌株可以多于106cfu每克制剂的量存在。
该制剂还可包含佐剂、药物实体和/或生物化合物。
该制剂可适合于施用至肺部或鼻部。例如,该制剂可是鼻喷雾剂的形式。该制剂的粘度可为1cps至2000cps。
双歧杆菌菌株或其制剂可用于食物或用于药物。
双歧杆菌菌株或制剂可用于预防和/或治疗不期望的炎性活性。
双歧杆菌菌株或其活性衍生物或片段或突变体或变体可用于预防和/或治疗不期望的炎性活性。
双歧杆菌菌株或其制剂可用于制备治疗哮喘和/或过敏的药物,所述药物可以是适合于吸入的形式。
双歧杆菌菌株或其制剂可用于预防或治疗对象的病毒感染。
双歧杆菌菌株或其制剂可用于预防与对象的呼吸道病毒感染相关的继发细菌感染。
在某些情况下,所述病毒是可选自流感病毒、鼻病毒和呼吸道合胞病毒的呼吸道病毒。
对象可能已被诊断患有炎性肺病。
在某些情况下,对象对呼吸道感染的易感性增加。
对象可以是肥胖的、急性呼吸窘迫综合征(ards)患者、哮喘患者或慢性阻塞性肺病(copd)患者中的一种或多种。
在一种情况下,对象是小于5岁的儿童。
在另一种情况下,对象是年龄超过60岁的老年人。
本发明还提供了预防或治疗对象的病毒感染的方法,包括将有效量的长双歧杆菌ncimb42020菌株或其活性衍生物或片段或突变体或变体施用给有需要的对象。
长双歧杆菌菌株ncimb42020或其活性衍生物或片段或突变体或变体可:
减弱ip-10对病毒的应答;
增强iii型干扰素对病毒的应答,如干扰素λ对病毒的应答;
抑制i型干扰素对病毒的应答,如干扰素α对病毒的应答,和/或干扰素β对病毒的应答;和/或
增强表面活性蛋白d对病毒的应答。
本发明还提供了分离自长双歧杆菌菌株ncimb42020的细胞壁组分。
该细胞壁组分可:
减弱ip-10对病毒的应答;
增强iii型干扰素对病毒的应答,如干扰素λ对病毒的应答;
抑制i型干扰素对病毒的应答,如干扰素α对病毒的应答,和/或干扰素β对病毒的应答;和/或
增强表面活性蛋白d对病毒的应答。
在某些情况下,所述病毒是可选自流感病毒、鼻病毒和呼吸道合胞病毒的呼吸道病毒。
对象可能已被诊断患有炎性肺病。
在某些情况下,对象对呼吸道感染的易感性增加。
对象可以是肥胖的、急性呼吸窘迫综合征(ards)患者、哮喘患者或慢性阻塞性肺病(copd)患者中的一种或多种。
在一种情况下,对象是小于5岁的儿童。
在另一种情况下,对象是年龄超过60岁的老年人。
该细胞壁组分的分子量可大于100kda。该细胞壁组分的尺寸可小于0.45μm。
在一个实施方案中,通过打开长双歧杆菌并将细胞壁组分与细胞质组分分开来分离所述组分。
本发明还提供了分离自长双歧杆菌菌株的细胞壁组分用于预防或治疗对象的呼吸道病毒感染。
本发明还提供了分离自长双歧杆菌菌株的细胞壁组分用于预防与对象的呼吸道病毒感染相关的继发细菌感染。
本发明还提供了分离长双歧杆菌细胞壁组分的方法,包括以下步骤:
打开长双歧杆菌以形成细胞壁组分和细胞质组分;和
将细胞壁组分与细胞质组分分开。
打开长双歧杆菌包括以下的至少一种:
用螯合剂处理;
用酶处理;和
施加剪切力。
螯合剂可以是钙螯合剂,如乙二胺四乙酸(edta)。酶可以是糖苷水解酶,如溶菌酶。可选地或另选地,酶是溶胞壁(muralytic)酶,如变溶菌素(mutanolysin)。
剪切力可以通过超声处理和/或通过压力(如通过弗氏压碎器)施加。
分开可包括离心。
在某些情况下,分开后,过滤细胞壁组分以提供尺寸小于0.45μm的组分。
本发明还提供了一种制剂,其包含细胞壁组分。
该制剂还可包含另外的益生菌材料和/或益生元材料。
该制剂还可包含可摄入的载体。可摄入的载体可以是药学上可接受的载体,如胶囊、片剂或粉末。可摄入的载体可以是食品,如酸化乳、酸奶、冷冻酸奶、冰淇淋、乳粉、浓缩乳、冰淇淋、奶酪酱、调味品或饮料。
该制剂还可包含蛋白质和/或肽(特别是富含谷氨酰胺/谷氨酸的蛋白质和/或肽)、脂质、碳水化合物、维生素、矿物质和/或微量元素。
该制剂还可包含佐剂、药物实体和/或生物化合物。
该制剂可适合于施用至肺部或鼻部。例如该制剂可以以鼻喷雾剂的形式。该制剂的粘度可为1cps至2000cps。
本发明提供了长双歧杆菌ncimb42020(ah0106)的分离菌株。
本发明还提供了长双歧杆菌ncimb42020分离菌株的突变体或变体。
分离菌株可以是活细胞形式。
分离菌株可以是非活细胞形式。
本发明还提供了包含长双歧杆菌ncimb42020的分离菌株的制剂。
该制剂可包含可摄入的载体。可摄入的载体可以是药学上可接受的载体,如胶囊、片剂或粉末。可摄入的载体可以是食品,如酸化乳、酸奶、冷冻酸奶、乳粉、浓缩乳、奶酪酱、调味品或饮料。
在一个实施方案中,该菌株减弱ip-10对病毒的应答。
在一个实施方案中,该菌株增强iii型干扰素对病毒的应答。
在一个实施方案中,该菌株抑制干扰素α对病毒的应答。
在一个实施方案中,该菌株抑制干扰素β对病毒的应答。
在一种情况下,该菌株:
减弱ip-10对病毒的应答;
增强iii型干扰素对病毒的应答;
抑制干扰素α对病毒的应答;和
抑制干扰素β对病毒的应答。
该菌株可以多于106cfu每克可摄入的载体存在。
本发明进一步提供了一种组合物,其包含长双歧杆菌ncimb42020的分离菌株和药学上可接受的载体。
本发明还提供了长双歧杆菌菌株ncimb42020作为益生菌菌株的用途。
在一个实施方案中,该制剂适合于施用至肺部或鼻部。
应当理解的是,本发明的特定菌株可以以常规制备物(如胶囊、微胶囊、片剂、颗粒、粉末、锭剂、丸剂、栓剂、悬浮剂和糖浆)以口服可摄入形式施用于动物(包括人)。合适的制剂可以通过通常采用的使用常规有机和无机添加剂的方法制备。医药组合物中活性成分的量可以是行使期望的治疗效果的水平。
制剂还可以包括细菌成分、药物实体或生物化合物。
此外,包含本发明菌株的疫苗可以使用任何合适的已知方法制备,并且可以包括药学上可接受的载体或佐剂。
本发明还提供了本发明的双歧杆菌菌株或本发明的制剂,其用于预防或治疗对象的病毒感染。
在一种情况下,对象是急性呼吸窘迫综合征(ards)患者。
在另一种情况下,对象是哮喘患者。
在再另一种情况下,对象是慢性阻塞性肺病(copd)患者。
本发明还提供了分离自长双歧杆菌ncimb42020的细胞壁组分,其中所述细胞壁组分减弱ip-10对病毒的应答。
在一种情况下,该组分增强iii型干扰素对病毒的应答。
在一种情况下,该组分抑制干扰素α对病毒的应答。
在一种情况下,该组分抑制干扰素β对病毒的应答。
本发明还提供了分离自长双歧杆菌ncimb42020的细胞壁组分,其中所述细胞壁组分:
减弱ip-10对病毒的应答;
增强iii型干扰素对病毒的应答;
抑制干扰素α对病毒的应答;和
抑制干扰素β对病毒的应答。
在一种情况下,细胞壁组分的分子量大于100kda。
在一种情况下,细胞壁组分的尺寸小于0.45μm。
在一种情况下,该细胞壁组分的分子量大于100kda且尺寸小于0.45μm。
本发明还提供了预防或治疗对象的病毒感染的方法,包括施用分离自长双歧杆菌ncimb42020的细胞壁组分的步骤。
在一种情况下,该细胞壁组分减弱ip-10对病毒的应答。
在一种情况下,该组分增强iii型干扰素对病毒的应答。
在一种情况下,该组分抑制干扰素α对病毒的应答。
在一种情况下,该组分抑制干扰素β对病毒的应答。
还提供了预防或治疗对象的病毒感染的方法,包括施用分离自长双歧杆菌ncimb42020的细胞壁组分,其中该组分:
减弱ip-10对病毒的应答;
增强iii型干扰素对病毒的应答;
抑制干扰素α对病毒的应答;和
抑制干扰素β对病毒的应答。
在一种情况下,细胞壁组分的分子量大于100kda。
在一种情况下,细胞壁组分的尺寸小于0.45μm。
在一种情况下,细胞壁组分以适合于施用至肺部或鼻部的制剂施用。
在一种情况下,对象是急性呼吸窘迫综合征(ards)患者。
在一种情况下,对象是哮喘患者。
在一种情况下,对象是慢性阻塞性肺病(copd)患者。
本发明还包括保藏菌株的突变体和变体。在整个说明书中,术语突变体、变体和遗传修饰突变体,包括与亲本菌株相比,其基因和/或表型特性改变的菌株。天然存在的变体包括选择性分离的靶特性的自发改变。通过常规(体外)遗传操作技术,如基因破坏、接合转移等,来完成亲本菌株特性的有意改变。遗传修饰包括将外源和/或内源dna序列引入菌株的基因组中,如通过载体(包括质粒dna或噬菌体)插入细菌菌株的基因组中。
天然或诱导型突变包括至少单碱基改变,如缺失、插入、颠换或其他dna修饰,其可导致由dna序列编码的氨基酸序列的改变。
术语突变体、变体和遗传修饰的突变体还包括已经历遗传改变的菌株,所述遗传改变以与自然界中的所有微生物一致的速率在基因组上累积,和/或所述遗传改变通过不是由有意(体外)操作基因组实现而是通过自然选择在暴露于环境压力如抗生素时提供支持细菌存活的选择性优势的变体和/或突变体来实现的自发突变和/或基因获得和/或基因丧失发生。可以通过将特定基因有意(体外)插入基因组中来创建突变体,其不会从根本上改变生物体的生物化学功能,但其产物可用于鉴定或选择细菌,如抗生素抗性。
本领域技术人员应当理解,可以通过与亲本菌株的dna序列同源性分析来鉴定突变体或变体菌株。与亲本菌株具有接近序列相同性而没有明显的表型或可测量的功能差异的菌株被认为是突变体或变体菌株。与亲本dna序列具有99.5%或更高序列相同性(同源性)的菌株可以被认为是突变体或变体。序列同源性可以使用在线同源性算法“blast”程序确定,该程序可在http://www.ncbi.nlm.nih,gov/blast/公开获得。
附图说明
从以下仅以实例方式给出的描述并参考所附附图,将更清楚地理解本发明。
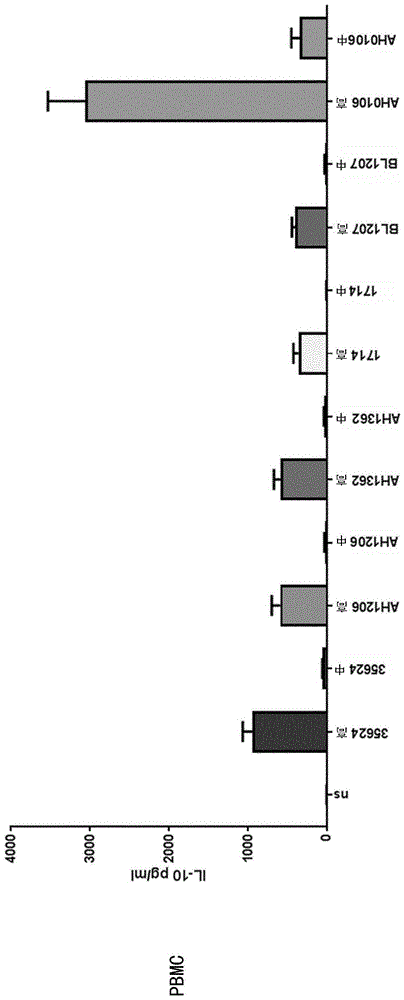
图1是显示在体外用浓度递增(中度或高度)的长双歧杆菌菌株35624、ah1206、ah1362、1714、bl1207和ah0106刺激后,pbmc和mddc中il-10的诱导谱的条形图;
图2是显示在体外用浓度递增(中度或高度)的长双歧杆菌菌株35624、ah1206、ah1362、1714、bl1207和ah0106刺激后,pbmc和mddc中il-12p40的诱导谱的条形图;
图3是显示在体外用浓度递增(中度或高度)的长双歧杆菌菌株35624、ah1206、ah1362、1714、bl1207和ah0106刺激后,pbmc和mddc中tnf-α的诱导谱的条形图;
图4是显示在体外用浓度递增(中度或高度)的长双歧杆菌菌株35624、ah1206、ah1362、1714、bl1207和ah0106刺激后,pbmc和mddc中il-1β的诱导谱的条形图;
图5是在长双歧杆菌ah0106存在下,ip-10对鼻病毒应答的条形图;
图6(a)到图6(c)是在长双歧杆菌ah0106存在下,干扰素λ(iii型干扰素)、干扰素α和干扰素β(i型干扰素)对鼻病毒应答的一系列条形图;
图7是比较在长双歧杆菌ah0106和金黄色葡萄球菌(staphylococcusaureus)菌株存在下,ip-10对鼻病毒16应答的条形图;
图8是在长双歧杆菌菌株ah0106和35624存在下,ip-10对lps应答的条形图;
图9是在长双歧杆菌菌株ah0106和35624存在下,tnf-α对lps应答的条形图;
图10是病毒感染后,肺中的病毒复制对菌株长双歧杆菌ah0106、长双歧杆菌35624和安慰剂应答的图;
图11是用长双歧杆菌菌株ah0106、35624和安慰剂感染后的一段时间内的存活的图;
图12是病毒感染后,支气管肺泡灌洗(bal)液中的细胞因子和血清中的表面活性蛋白d对菌株长双歧杆菌菌株ah0106、35624和安慰剂应答的一系列图;
图13显示了病毒感染后,bal中测量的不同生物标志物对菌株长双歧杆菌菌株ah0106、35624和安慰剂应答的热图;条带强度越高,诱导越大;
图14是病毒感染后,bal巨噬细胞计数对菌株长双歧杆菌菌株ah0106、35624和安慰剂应答的图;
图15是用于从长双歧杆菌ah0106分离细胞壁组分的方法的流程图。
图16(a)是在单核细胞衍生的树突状细胞中,在来自菌株ah0106的细胞壁组分(bifah0106沉淀物)存在下,ip-10对人鼻病毒(hrv16)应答的图;
图16(b)是在单核细胞衍生的树突状细胞中,在来自菌株ah0106的细胞壁组分(bifah0106沉淀物)存在下,ifn-α对人鼻病毒(hrv16)应答的图;
图16(c)是在单核细胞衍生的树突状细胞中,在来自菌株ah0106的细胞壁组分(bifah0106沉淀物)存在下,ifn-β对人鼻病毒(hrv16)应答的图;
图17(a)是在单核细胞衍生的树突状细胞中,相较于来自菌株ah0106的细胞壁组分,在来自菌株ucc2003的细胞壁组分存在下,ip-10对人鼻病(hrv16)应答的条形图;
图17(b)是在单核细胞衍生的树突状细胞中,相较于来自菌株ah0106的细胞壁组分,在来自菌株ucc2003的细胞壁组分存在下,ip-10对干扰素β(ifn-β)应答的条形图;
图18(a)是在单核细胞衍生的树突状细胞中,相较于来自长双歧杆菌菌株ah0106和35624的细胞壁组分,在来自菌株ucc2003的细胞壁组分存在下,tnf-α对lps应答的条形图;
图18(b)是在单核细胞衍生的树突状细胞中,相较于来自长双歧杆菌菌株ah0106和35624的细胞壁组分,在来自菌株ucc2003的细胞壁组分存在下,tnf-α对poly:ic应答的条形图;
图19(a)是对病毒感染的异常级联应答的说明;
图19(b)是对由长双歧杆菌ah106的菌株或细胞壁组分介导的病毒感染的级联应答的说明;
具体实施方式
长双歧杆菌(bifidobacteriumlongum)菌株ah0106于2012年8月2日以保藏号ncimb42020保藏在英国国家工业和海洋细菌保藏中心(nationalcollectionsofindustrialandmarinebacterialimited(ncimb)fergusonbuilding,craibstoneestate,bucksburn,aberdeen,ab219ya,苏格兰,英国)。
本说明书也提到长双歧杆菌(bifidobacteriumlongum)ncimb41003(
我们已经发现,特定细菌菌株促进抗病毒防御并抑制破坏性促炎应答,并且特别用于预防和/或治疗病毒诱导的ards、或哮喘和copd患者中病毒诱导的恶化。
实施例
以下实施例进一步描述和说明了本发明范围内的实施方案。给出的实施例仅用于说明的目的,不应解释为对本发明的限制,因为在不脱离本发明的精神和范围的情况下,可以对其进行许多变化。
实施例1:长双歧杆菌ncimb42020的分离
从健康人对象的粪便样品中分离长双歧杆菌菌株ncimb42020。
筛选粪便样品以得到益生菌菌株。将样品转移到含有磷酸盐缓冲盐水(pbs)、补充有0.05%半胱氨酸-hcl)的收集管中。然后将溶液孵育10分钟。将样品涡旋并铺板在选择性琼脂(deman,rogosaandsharpe(mrs)琼脂+10%蔗糖+莫匹罗星和刚果红+半胱氨酸+莫匹罗星)上。使用刚果红琼脂筛选来表型筛选eps表达细菌菌株。简言之,用新鲜生长的细菌菌株菌落无菌接种至10ml改良rogosa培养液培养基中(+0.05%半胱氨酸),然后在37℃厌氧孵育直至混浊(约16到24小时)。将培养液培养物无菌划线到刚果红琼脂平板上,并在37℃厌氧孵育48小时。据信,作为某些菌株的生长和/或代谢的副产物产生的eps,阻止了刚果红染色的摄取,导致米色/白色菌落形态。产生较少eps的菌株容易吸收刚果红染色,导致粉色/红色菌落形态。不产生eps的菌株染成红色,并在红色琼脂背景下看起来几乎透明。
从平板中挑选分离的菌落,并重新划线三次以确保纯度。显微镜检查、革兰氏染色、过氧化氢酶测试、果糖-6-磷酸磷酸酮醇酶评估用于确定假定的双歧杆菌物种,并将分离物贮存在40%甘油中并储存在-80℃中。使用16s基因间隔区测序(igs)来确认新分离菌株的身份。
在分离纯双歧杆菌菌株后,指定名称为ah0106,随后将其保藏在ncimb且保藏号为42020。评估微生物特征并总结于下表1中。长双歧杆菌ncimb42020是一种革兰氏阳性、过氧化物酶阴性的多形性型细菌,其为果糖-6-磷酸磷酸酮醇酶阳性,证实其身份为一种双歧杆菌。
表1-长双歧杆菌ncimb42020的理化特征
进行16s-23s基因间隔区(igs)测序以鉴定分离的双歧杆菌的物种。简言之,使用100μl提取溶液和25μl组织制备溶液(sigma,xnat2kit)从ncimb42020中分离dna。将样品在室温孵育2小时,随后在95℃2小时,然后加入100μl的中和溶液(sigma,xnat2kit)。利用nanodrop分光光度计定量基因组dna溶液并储存于4℃。使用igs引物进行pcr。使用的引物对是igsr5’-ctggtgccaaggcatcca-3’(seqidno.1)和igsl5’-gctggatcacctcctttct-3’(seqidno.2)。循环条件是94℃4分钟(1个循环),94℃45秒,53℃45秒,72℃45秒(28个循环)。该pcr反应含有2μl(100ng)的dna、pcr混合物(sigma,redtaq)、0.025nmigsl和r引物(mwgbiotech,germany)。pcr反应在biotherma热循环仪中进行。将pcr产物(10μl)与分子量标志物(100bpladder,roche)一起在tae中的2%琼脂糖etbr染色凝胶上跑,以确定igs谱。使用promegawizardpcr纯化试剂盒纯化双歧杆菌的pcr产物(单一条带)。使用针对基因间隔区的引物序列(上文)对纯化的pcr产物进行测序。然后针对ncbi核苷酸数据库检索序列数据,以通过核苷酸同源性确定菌株的身份。将得到的dna序列数据置于ncbi标准核苷酸-核苷酸同源性blast检索引擎中(http://www.ncbi.nlm.nih.gov/blast/)。鉴定与序列最相近的匹配,然后使用dnastarmegalignra软件比对序列用于比较。序列可见于序列表(表2)。检索ncimb数据库揭示了ncimb42020具有独特的igs(表2)序列,其与长双歧杆菌具有最接近的序列同源性。
表2长双歧杆菌ncimb42020的igs序列(seqidno.3)
ttgctgggatcacctcctttttacggagaattcagtcggatgttcgtccgacggtgtgcgccccgcgcgtcgcatggtgcgatggcggcggggttgctggtgtggaaaacgtcgttggctttgccctgccggtcgtgcggtgggtgcggggtggtatggatgcgcttttgggctcccggatcgccaccccaggctttttgcctggcgcgattcgatgcccgtcgtgcctgggggccggccgtgtgccggcgcgatggcgtggcggtgcgtggtggcttgagaactggatagtggacgcgagcaaaacaagggtttttgaatctttgttttgctgttgatttcgaatcgaactctattgttcgtttcgatcgttttgtgatcatttttagtgtgatgatttgtcgtcctgggaatttgctagaggaatacttgcgggccatgcactttcgtggtgtgtgttgcttgcaagggcgtatggtggaggccttggcaccagaa
实施例2:ah106的细胞因子谱和与其他具有潜在健康益处的长双歧杆菌菌株的谱的比较
使用密度梯度离心从健康供体中分离外周血单核细胞(pbmc)。将pbmc洗涤并重悬于dulbecco'smodifiedeaglemedium-glutamax(dmem)tm(glutamax(谷氨酰胺替代品)+丙酮酸盐+4.5g/l葡萄糖(gibcocatalog10569-010)10%胎牛血清(sigmacatalogf4135),和1%青霉素/链霉素(sigmacatalogp0781)。在37℃、5%co2的补充dmem中,用不同剂量的长双歧杆菌((总细菌:pbmc)高度(100:1)和中度(50:1))刺激pbmc。
对于单核细胞衍生的树突状细胞的产生:使用密度梯度离心从健康供体分离外周血单核细胞(pbmc)。利用macs系统(miltenyibiotec,130-050-201)使用cd14阳性分离来分离人外周血单核细胞。将细胞在含有白细胞介素41000u/ml(novartis)和粒细胞巨噬细胞集落刺激因子(peprotech,300-03)1000u/ml的crpmi培养基(lifetechnologies,21875-091)中培养6天,以将它们分化成单核细胞衍生的树突状细胞(mddc)。在37℃、5%co2的crpmi中,用不同剂量的长双歧杆菌((总细菌:mddc)高度(100:1)和中度(50:1))刺激mddc。使用商业细胞因子试剂盒(mesoscalediscovery)平台按照制造商的说明测量细胞因子的分泌。
与彼此具有相似谱的所有其他细菌菌株(35624、ah1206、ah1362、1714、bl1207)相比,ah0106在pbmc和mddc中诱导更多的il-10、il-12p40、tnf-α、il-1β(图1-4)。ah106是比其他长双歧杆菌(特别是已知的抗炎菌株如35624和1714)更有效的细胞因子刺激物,并且证明ah0106用与其他长双歧杆菌菌株不同的方式(更加免疫刺激的方式)参与人类免疫系统。
实施例3:单核细胞衍生的树突状细胞中的ah0106菌株(有益地阻断i型干扰素和由此产生的ip-10(一种促炎趋化因子)诱导并增加了iii型干扰素ifn-λ)
树突状细胞对病毒感染的过度免疫应答引起肺中的促炎应答。i型干扰素可以刺激ip-10的产生,其结合的趋化因子是th1细胞的化学引诱物。为了确定长双歧杆菌ah0106是否可能对人类单核细胞衍生的树突状细胞具有有益的抗病毒作用,在暴露于人鼻病毒16(hrv16)之前用长双歧杆菌ah0106刺激mddc,并监测干扰素和ip-10对hrv16的应答。用多剂量的长双歧杆菌ah0106(100:1、50:1、25:1、10:1、1:1)预处理1小时后,用hrv16(moi)25:1)刺激37℃、5%co2的crpmi中的mddc24小时,或仅用hrv16单独刺激。通过bio-plex多重悬浮阵列(bio-radlaboratories)检测细胞因子分泌,测量ifn-α、ifn-β、ifn-λ和ip-10。令人惊讶的是,ip-10对rv的应答被长双歧杆菌ah0106菌株以剂量依赖性方式减弱(图5)。同样令人惊讶的是,与较低剂量的长双歧杆菌35624共孵育,导致iii型干扰素应答(干扰素λ)的增强,而在较高剂量干扰素α和β应答被抑制,类似于ip-10应答(图6)。该数据显示,长双歧杆菌ah0106改变了对rv的免疫应答,支持保护性免疫应答,同时抑制了破坏性应答。
实施例4:不是所有的革兰氏阳性细菌菌株具有该作用
为了确定另外的革兰氏阳性细菌菌株是否可能对人类单核细胞衍生的树突状细胞具有有益的抗病毒作用,用金黄色葡萄球菌刺激的mddc暴露于人鼻病毒16(hrv16),并监测ip-10的应答。简言之,与长双歧杆菌ah0106(100:1)相比,用多剂量金黄色葡萄球菌(100:1、50:1)预处理1小时后,用hrv16(moi)25:1)刺激37℃、5%co2的crpmi中的mddc24小时,或仅用hrv16单独刺激。金黄色葡萄球菌菌株不减弱dc对hrv16应答的ip-10分泌(图7)。
实施例5:ah0106菌株有益地阻断单核细胞衍生的树突状细胞中的tnf-α和ip-10,但并非所有长双歧杆菌菌株具有相同的作用
在发炎的粘膜内,不仅病毒本身诱导ip-10和tnf-α分泌,而且其他(toll样受体)tlr配体也可以诱导其产生。因此,在暴露于lps之前,用长双歧杆菌ah0106和长双歧杆菌35624预处理mddc。监测ip-10和tnf对lps(一种tlr-4激动剂)的应答。简言之,用剂量为(100:1)的长双歧杆菌ah0106或35624预处理1小时后,用lps(50ng/ml)刺激37℃、5%co2的crpmi中的mddc24小时,或仅用lps单独刺激。令人惊讶的是,长双歧杆菌ah106减弱了ip-10和tnf-α对lps的应答,而暴露于长双歧杆菌35624后,ip-10对lps的应答仅略有下降(图8),并且在相同剂量下,长双歧杆菌35624增加了tnf-α(图9)。这是一个非常令人惊讶的结果,因为长双歧杆菌35624是一种熟知的抗炎菌株,并且是假定的最佳候选菌株。长双歧杆菌ah0106的一些细胞因子数据表明,它会引起广泛的免疫刺激反应,但在实践中,长双歧杆菌ah0106在该细胞系统中具有优异的和特异性的免疫应答。
实施例6:长双歧杆菌ah106和长双歧杆菌35624在呼吸道感染临床前体内模型中的比较
我们测试了长双歧杆菌ah0106菌株在旨在显示治疗益处的临床前模型中的功效。鼻病毒小鼠模型不被认为是人类感染的良好替代品,因为小鼠不具有icam,icam是主要群体鼻病毒进入细胞的受体,因此,我们在小鼠中使用了致死流感模型,这被认为是更好的模型(bartlettetal2015)。使用h1n1流感株a/pr8/34病毒株(100pfu/50ul)感染小鼠。
在病毒感染后-2小时、+1天、+3天和+5天鼻内施用长双歧杆菌ah0106菌株、长双歧杆菌35624菌株或安慰剂。以1x109个总细胞的剂量施用这些菌株。
第2小时、第1天和第3天经鼻部施用载体对照(第1组)、长双歧杆菌ah0106(第2组)和长双歧杆菌35624细胞壁组分(第3组)(体积50μl)。
第0天经鼻部施用一剂致死流感(pr8)(第1-3组)。
第0-10天监测动物的发病率(体重、体温和临床评分,第1-3组)。
第5天:每组处死5只动物进行末端出血、器官移除和分析。第5天(第1-3组)。
第5天:分离bal液以测量细胞因子和细胞浸润。收集肺组织,通过定量pcr定量肺中的病毒滴度(所有肺叶的一半)。
测量肺组织中的病毒滴度。
准备分离的肺叶用于通过定量pcr定量肺组织中的病毒载量。用trireagent(molecularresearchcenter)制备rna,然后用dna酶(invitrogen)处理以避免基因组dna污染,然后使用superscriptiii(invitrogen)通过逆转录将rna转化为cdna。使用sybrgreen(stratagene)通过实时pcr(icycler;bio-rad)定量cdna,并用gapdh表达水平标准化样品。使用的引物序列(正向和反向分别)是流感pr8m蛋白,5’-ggactgcagcgtagacgctt-3’(seqidno.4)和5’catcctgtatatgaggcccat-3’(seqidno.5)。
第(1-3)组:
1.用安慰剂(冷冻保护剂重悬于pbs中)处理。
2.用长双歧杆菌ah010610^9总细胞处理。
3.用长双歧杆菌3562410^9总细胞处理。
每组小鼠的数量(第1-3组)=5
细胞因子和趋化因子的测量。
使用商业u-plexbiomarkergroup1mouse35-plex(mesoscalediscovery)平台,根据制造商的说明,测量小鼠血清和bal液中il-1β、il-2、il-4、il-5、il-6、il-9、il-10、il-12p70、il-12/il-23p40、il-13、il-15、il-16、il-17a、il-17a/f、il-17c、il-17e、il-17f、il-21、il-22、il-23、il-30、il-31、il-33、ip-10、mip3α、mip-2、mip-1β、mip-1α、mcp-1、kc/gro、tnf-α、vegf、epo、gm-csf、ifn-γ的浓度;使用elisa试剂盒(raybiotech,inc,)检测小鼠il-28(ifn-λ2/3)、小鼠g-csf、小鼠trail、小鼠areg;使用来自r&dsystems的quantikine试剂盒,按照制造商的说明书,测量制瘤素m和小鼠表面活性蛋白d(spd)。通过ifn-α铂elisa(thermofisherscientific)测量小鼠血清中和bal液中的ifn-α,使用verikinemouseinterferonbetahselisakits(pblassayscience)测量小鼠血清和bal液中干扰素β(ifnβ)的水平。
测量细胞浸润到bal中。
从bal液中分离细胞,并使用coultercounter(iginstrumenten-gesellschaftag,basel,switzerland)确定支气管肺泡灌洗(bal)液中的总细胞数。基于在用diff-quik溶液(dadebehring,siemenshealthcarediagnostics,deerfield,il)染色的细胞离心涂片器上的标准形态的和细胞化学的标准,进行差异细胞计数(200个细胞计数/样品)。
令人惊讶的是,施用长双歧杆菌ah0106比长双歧杆菌35624菌株更好地保护了小鼠。在两种菌株中病毒滴度降低了相似的水平(图10),但在长双歧杆菌ah0106处理的小鼠中死亡率降低更多(图11)。这种增强的存活与降低的干扰素α和干扰素β应答以及增强的表面活性蛋白d应答相关(图12)。此外,在ah0106处理的动物中,存在细胞因子和趋化因子的有效诱导(图13),其与第5天bal中巨噬细胞的增加有关(图14)。这种免疫应答有助于增强在长双歧杆菌ah0106处理的动物中观察到的病毒清除,并且愈合过程的开始使巨噬细胞流入肺中。
实施例7:细胞壁沉淀物生成
细菌收获/洗涤
方法:
1.通过离心(14000rpm,4℃,20min;转子ja-20(avantij-26xpbeckmancoulter)收获相当于250ml的原始细菌生物量(总细胞计数=1.5x1011)。细菌沉淀物用无菌pbs洗涤,弃去上清液并再次洗涤(重复两次)。
2.将活冻干细菌和非活冻干细菌(0.5g的3.0x1011粉末或总细胞计数=1.5x1011)重悬于50ml中,并通过离心(14000rpm,4℃,20min;转子ja-20(avantij-26xpbeckmancoulter)收获。用无菌pbs洗涤细菌沉淀物,弃去上清液并再次洗涤(重复两次)。
3.最后将沉淀物重悬于50ml无菌pbs中,将细菌溶液等分成两份25ml等分试样。
细胞破碎
该过程的目的是从整个细菌中产生细胞壁组分并消除细胞质组分和其他成分。这涉及选自以下的一种或多种步骤:
·用螯合剂处理,任选地与冻融过程结合,以在使用酶处理帮助裂解时阻止dna酶或蛋白酶活性。
·用糖苷水解酶和/或n-乙酰胞壁质酶(n-acetylmuramidase)进行酶处理。
·施加剪切力,如超声处理或施加高压,如使用弗氏压碎器。
·通过使用离心将细胞壁组分与细胞质组分分开。
·过滤细胞壁组分。
材料:
·edta(0.5mfluka)是用于去除金属离子(钙或镁)的螯合剂,以在使用酶进行细胞壁裂解时阻止dna酶或蛋白酶活性。
·溶菌酶(sigma10mg/ml)*不含内毒素,是一种糖苷水解酶,其催化n-乙酰胞壁酸和n-乙酰-d-氨基葡萄糖残基之间的1,4-β键水解。这种水解损害了细菌细胞壁的完整性,导致细菌裂解。
·变溶菌素(10kusigma稀释于1mlh2o)是n-乙酰胞壁质酶,它是一种溶胞壁酶,其切割细菌细胞壁的β-n-乙酰胞壁酰-(1→4)-n-乙酰葡糖胺键。这种切割损害了细菌细胞壁的完整性,导致细菌裂解。
·玻璃珠90-150μm粒径(vwr)与超声处理联用以帮助裂解。
方法:
1.向每个25ml等分试样中加入250μledta(0.5m原液fluka),因此终浓度为5mmedta。
2.通过将2个等分试样置于液氮中直至冷冻来冷冻等分试样,然后将等分试样在水中解冻;重复此过程两次。
3.将每个25ml细菌等分试样与250μl溶菌酶(10mg/ml)和250μl变溶菌素(2.5ku)在37℃孵育1小时,偶尔温和涡旋。从该步骤开始,通过超声波仪或弗氏压碎器使用剪切力来破碎细菌细胞。
超声处理
·在超声处理之前,将半茶匙的90-150μm粒径(vmr)的高压灭菌的玻璃珠加入到细菌等分试样中。
·一次超声处理25ml重悬的细菌材料,然后放在冰上,使用超声波仪(vibracellsonics)和50ml探针对其他等分试样进行超声处理。(设置调谐=50,频率=60)。该过程在冰上重复4次,持续10分钟。超声处理后,通过在4℃以1000rpm离心10分钟来除去玻璃珠、未破坏的细胞和细胞碎片。
用弗氏压碎器的高压
·可选地,使用弗氏压碎器(thermoelectroncorporationfa-078a)在高设定(20,000psi当量)下,通过1500psi的压力破碎细菌细胞。该过程在冰上重复三次。
·使用弗氏压碎器施加剪切力导致清除第三次运行后混浊的细菌细胞。在弗氏压碎器破碎后,通过在4℃以1000rpm离心10分钟除去未破坏的细胞和细胞碎片,并保留含有细胞壁材料和细胞质组分的上清液。
4.在施加剪切力(超声处理或高压)后,保留含有细胞壁材料和细胞质组分的上清液。将该上清液在4℃以14000rpm离心20分钟,以将细胞壁材料与细胞质材料分开。弃去上清液。
5.将每250ml的原始细菌生物量的含有细胞壁材料的沉淀物重悬于5mlpbs。将细胞壁材料沉淀物以1000rpm离心10分钟以除去沉淀物中的黑色残余物。将包含细胞壁材料的沉淀物储存在-80℃中。
离心机和转子
avantij-ecentrifugebeckmancoulter
转子:ja-20
超声仪
vibracellsonics,sonicsandmaterials
probe435-09
弗氏压碎器
弗氏压碎器fa-078a
cellfa-032(40kstandard)(standardfrenchpressure40,000psi,cell容量为35ml,压力高达40,000psi)
尺寸过滤后进行超滤
材料:
·聚偏二氟乙烯(pvdf)膜过滤器(0.45μm孔径,millipore,bedford,ma)
·100kdamwcouf装置(millipore).
1.将重悬的含有细胞壁材料的细菌沉淀物(5ml)解冻并通过聚偏二氟乙烯(pvdf)膜过滤器(0.45μm孔径,millipore,bedford,ma;在使用前用pbs彻底润洗)过滤。通过膜过滤器的材料不含完整细菌,并且是尺寸<0.45μm的细胞壁材料。通过该0.45μm过滤步骤产生4ml细胞壁材料。从最初的250ml细菌培养物种,留下4ml悬浮的细胞壁材料。
2.然后将<0.45μm的细胞壁材料进行超滤。
a.首先用15mlpbs润洗uf装置,在4℃以3500xg旋转。
b.取出<0.45μm的细菌沉淀物提取物,将约4ml载入至100kdamwcouf装置中。
c.当>100kda上层溶液体积变小时,用pbs渗滤。
d.通过100kda过滤器的材料将被保留并储存在-80℃中。
e.应将保留物重新悬浮于与加入超滤装置一样的体积的pbs中(4ml)。该材料被称为>100kda细菌沉淀物。在4ml保留物中,由此产生的细胞壁组分的最终干重是120mg,等于最终浓度为30mg细胞壁组分每1ml。
f.将该细胞壁组分浓缩,用于体外和体内试验以获得更高的溶解度。为了制备比30mg/ml更浓缩的材料,如300mg/ml(10x)或150mg/ml(5x)>100kda细菌沉淀物溶液,仅用比起始材料少10倍或5倍体积重悬细胞壁组分。
实施例8:mddcs中的来自ah0106的细胞壁组分(有益地阻断i型干扰素和由此产生的ip-10(一种促炎趋化因子)诱导
为了确定如实施例7产生的来自长双歧杆菌ah0106的细胞壁组分是否可能具有有益的抗病毒作用,将该组分在暴露于hrv16的人mddc上孵育,并监测i型干扰素和细胞ip-10应答。对于单核细胞衍生的巨噬细胞的产生:使用密度梯度离心从健康供体分离外周血单核细胞(pbmc)。利用macs系统(miltenyibiotec,130-050-201)使用cd14阳性分离来分离人外周血单核细胞。将细胞在含有白细胞介素41000u/ml(novartis)和粒细胞巨噬细胞集落刺激因子(peprotech,300-03)1000u/ml的crpmi培养基(lifetechnologies,21875-091)中培养6天,以将它们分化成mddc。将细胞在crpmi培养基(rpmi(lifetechnologies,21875-091)+10%胎牛血清(sigmacatalogf4135)和1%青霉素/链霉素(sigmacatalogp0781)中培养。
用来自长双歧杆菌ah0106的细胞组分(30mg/ml)预处理1小时后,用hrv16(moi)25:1)刺激37℃、5%co2的crpmi中的mddc24小时,或仅用hrv16单独刺激。通过bio-plex多重悬浮阵列(bio-radlaboratories)检测细胞因子分泌(ifn-α、ifn-β、ip-10)。
与用长双歧杆菌ah0106菌株预处理的hrv16刺激的mddc的上述结果一致,ip-10(图16(a))、ifn-α(图16(b))和ifn-β(图16(c))对hrv16的应答被来自长双歧杆菌ah0106菌株的细胞壁组分减弱。
实施例9:并非所有来自长双歧杆菌物种的细胞壁组分都具有相同的作用
还使用实施例4中描述的方法测试来自另一种双歧杆菌,短双歧杆菌(bifucc2003)的细胞壁组分,并且没有显示出相似的显著作用。该bifucc2003组分在病毒刺激后降低了ip-10的产生,但在24小时试验期间,没有达到长双歧杆菌ah0106细胞壁组分相同的程度(图17(a))。
此外,在发炎的粘膜内,不仅病毒本身诱导ip-10分泌;其他细胞因子也诱导ip-10产生。细胞因子如ifn-β是作为原发抗病毒宿主应答的一部分产生的。特别是ifn-β是ip-10的有效诱导剂。因此,我们检测了细胞壁组分对应答于ifn-β的ip-10分泌的影响(图17(b))。用细胞壁组分(30mg/ml)预处理1小时后用ifn-β(200ng/ml)刺激37℃、5%co2的crpmi中的mddc24小时,或仅用ifn-β单独刺激。长双歧杆菌ah0106细胞壁沉淀物组分抑制了针对这些刺激物的ip-10的分泌,而bifucc2003细胞壁沉淀物组分根本没有降低ip-10分泌。
实施例10:来自长双歧杆菌ah0106菌株的细胞壁组分在单核细胞衍生的树突状细胞中有额外的有益作用(阻断tnf-α),其有助于减少炎症,并且并非所有的双歧杆菌菌株都具有相同的作用
在发炎的粘膜内,不仅病毒本身诱导促炎性tnf-α分泌,而且其他(toll样受体)tlr配体也可以诱导其产生。因此,我们检测了对tlr配体poly:ic(tlr-3)和lps(tlr-4)应答的tnf-α的分泌。在暴露于lps(一种tlr-4激动剂)之前,用长双歧杆菌ah0106、长双歧杆菌35624和bifucc2003的细胞壁预处理mddc。监测ip-10和tnf的应答。用剂量为(30mg/ml)的来自双歧杆菌的细胞壁组分预处理1小时后,用tlr-4激动剂lps(50ng/ml)或tlr-3激动剂poly:ic(5μg/ml)刺激37℃、5%co2的crpmi中的mddc24小时,或仅用tlr激动剂单独刺激。通过quantikineelisa(r&dsystems)检测tnf-α的分泌。与来自整个长双歧杆菌ah0106菌株结果一致,来自长双歧杆菌ah106的细胞壁组分减弱了tnf-α对lps和poly:ic的应答,而相同剂量的来自长双歧杆菌35624和bifucc2003的细胞壁组分增加了tnf-α(图18(a)和18(b))。这是一个非常令人惊讶的结果,因为长双歧杆菌35624是一种熟知的抗炎菌株,而长双歧杆菌ah0106在该系统中具有优异的应答。
讨论
综上,如实施例2-10所示,我们已经显示存在增强的iii型干扰素应答,其是有益的,并且添加来自ah0106的细胞壁组分的引起了该理想应答。在作为之后宿主免疫系统应答的一部分的细胞(dc)中,细胞壁组分阻断了过度的可导致细胞损伤和继发感染的i型干扰素应答。这种靶向作用有益于流感病毒、普通感冒(鼻病毒)和rsv造成的感染,有益于儿童和成人以及肥胖个体中慢性呼吸道疾病如哮喘、copd和ards的病毒恶化。
级联应答
病毒感染的异常级联应答如图19(a)所示。
ah0106细胞壁组分介导的对病毒感染的级联应答如图19(b)所示。
主要的抗病毒应答由ifn控制。最明确定义的i型ifn是ifn-α和ifn-β。大多数细胞类型产生ifn-β,而造血细胞,特别是浆细胞样树突状细胞,是ifn-α的主要产生者(ivashkivanddonlin2014)。如前所述,i型干扰素应答,如ifn-α和ifn-β,已被证明与流感感染模型中发病率和死亡率的增加直接相关(davidsonetal,2014)。i型ifn可以刺激ip-10(也称为cxcl-10)的产生,该趋化因子与cxcr3结合,其主要功能是th1细胞的化学引诱物。λifn(ifnl、iii型ifn或il-28和il-29)构成了更新的一类干扰素,它们与i型干扰素共有同源性、表达模式和抗病毒功能(lazearetal.,2015b;wacketal.,2015)。它们诱导出现与i型ifn非常相似的下游信号传导,驱动isg的表达和抗病毒应答的诱导(durbinetal,2013;mendozaetal,2017)。然而,iii型干扰素在限制促炎应答或免疫病理学中起重要作用。
在流感病毒感染的肺中,ip-10升高(ichikawaetal,2013)。实际上,使用单克隆抗体阻断ip-10改善病毒诱导的肺损伤(wangetal,2013)。ip-10在ards患者的肺中升高,并且已经显示其是ards病理学中的重要因素(ichikawaetal,2013)。
sp-d通过与病毒的ha/na糖蛋白上富含甘露糖的聚糖结合,在对抗流感的固有宿主防御中具有重要作用(hartshornetal,1997;readingetal,1997;hartshornetal,2000)。sp-d在体外介导一系列抗病毒活性,包括中和病毒感染性和抑制病毒na的酶活性,并且sp-d缺陷型小鼠对高度糖基化的流感病毒感染更易感(hartshornetal,1997;readingetal,1997;tecleetal,2007;levineetal,2001;vigerustetal,2007;hawgoodetal,2004)。sp-d增强了吞噬rsv和肺清除rsv(levineetal,2004)。
继发细菌感染是病毒感染后的主要问题。充分认识到流感、鼻病毒和rsv的病毒-细菌共感染。呼吸道中的主要细菌感染包括肺炎链球菌、卡拉莫拉菌和流感嗜血杆菌,甚至金黄色葡萄球菌也已显示在病毒感染后引起严重感染(hewittetal,2016)。继发细菌感染最常发生在原发病毒感染后的5-10天,因此表明短暂的免疫抑制(原发应答)可能是细菌生长的缘由。所提出的针对流感和肺炎链球菌之间的协同作用的机制表明由原发流感病毒感染引起的抗病毒i型ifn(ifn-α/β)应答通过抑制抗菌免疫增强了宿主对继发细菌攻击的易感性(nakamuraetal,2011;shahangianetal,2009;lietal,2012)。相反,iii型干扰素,如ifn-λ可以限制病毒复制而不抑制ifn-α/β诱导后发生的继发细菌感染的清除。已经表明,与动物中的对照相比,在细菌攻击之前用ifnlr1fc蛋白直接减弱ifn-λ信号传导的影响显著增加了肺中的细菌负荷(richetal,2017)。此外,sp-d的缺失与上呼吸道和下呼吸道肺炎链球菌的定植和感染的增加,以及菌血症的较早发作和较长持续存在相关。显示sp-d在体外结合并凝集肺炎链球菌(一种继发细菌感染物质,其是哮喘和copd患者继发恶化的关键问题)(jounblatetal,2005)。
被病毒感染的失调或异常免疫应答急剧影响的不同疾病,总结如下;急性呼吸窘迫综合征(ards),哮喘包括儿童哮喘、copd、肥胖。
急性呼吸窘迫综合征(ards)影响全世界的许多人,且与非常高的死亡率(30-50%)相关。呼吸道病毒感染(如流感)与ards相关。
哮喘是气道的慢性炎性病症,通常与气道高应答性和可变气流相关。
哮喘是气道的慢性炎性病症,通常与气道高应答性和可变气流阻塞相关,其通常可自发地或在治疗期间可逆(who,2007)。在儿童、青少年和较低频率的成年人中,大约80至85%的哮喘恶化与病毒性上呼吸道病毒感染有关,而鼻病毒(rv)占这些病毒相关恶化的约60-70%。病毒感染与所有年龄段儿童的哮鸣(wheezing)疾病密切相关。rsv是细支气管炎或哮吼(croup)的主要致病物质,而鼻病毒(rv)在之后的哮鸣儿童中最常被检测到。由这些中任一引起的严重呼吸道疾病与随后的哮喘发展有关,并且对有rv感染的哮鸣的幼儿的风险最大(jarttiandgern,2017)。
肥胖与失调的免疫和炎性应答有关。肥胖对哮喘发生的影响似乎在女性和非过敏个体中更为突出,而体重指数(bmi)的增加对哮喘发病有剂量应答效应。越来越明显的是,肥胖与独特的哮喘表型相关,其特征在于对常规哮喘疗法具有不同应答的更严重的疾病。此外,在2009年甲型流感h1n1大流行期间,肥胖被鉴定为严重流感的危险因素,并且肥胖个体对呼吸道病毒的抗病毒防御受损(almondetal,2013)。有证据表明,不是病毒本身,而是针对rv的免疫应答的性质驱动了这种破坏性应答(steinkeetal,2016)。
copd是美国第三大死亡原因。事实上,copd是唯一发病率正在增加的主要死亡原因,预计到2030年将成为发达世界的第三大死亡原因(仅次于心脏病和中风)。它是由炎症诱导的气道损伤造成的,引起慢性支气管炎和/或肺气肿。更广谱的病毒可诱导copd患者的恶化,但又似乎不是病毒,而是针对病毒的免疫应答导致症状的恶化(zhouetal,2015)。
应当理解的是,本发明的菌株可以以常规制备物(如胶囊、微胶囊、片剂、颗粒、粉末、锭剂、丸剂、栓剂、悬浮剂和糖浆)以口服可摄入形式施用于动物(包括人)。合适的制剂可以通过通常采用的使用常规有机和无机添加剂的方法制备。医药组合物中活性成分的量可以是行使期望的治疗效果的水平。
制剂还可以包括细菌成分、药物实体或生物化合物。
此外,包含本发明菌株的疫苗可以使用任何合适的已知方法制备,并且可以包括药学上可接受的载体或佐剂。
人体免疫系统在广泛的人类疾病的病因学和病理学中起重要作用。高免疫应答和低免疫应答导致大多数疾病状态,或是大多数疾病状态的组成部分。一类生物实体,称为细胞因子,对控制免疫过程特别重要。这些微妙的细胞因子网络的扰动正在与许多疾病越来越相关。这些疾病包括但不限于炎性病症、免疫缺陷、炎性肠病、肠易激综合征、癌症(特别是胃肠和免疫系统的那些)、腹泻病、抗生素相关性腹泻、小儿腹泻、阑尾炎、自身免疫性病症、多发性硬化、阿尔兹海默病、类风湿性关节炎、乳糜泻、糖尿病、器官移植、细菌感染、病毒感染、真菌感染、牙周病、泌尿生殖系统疾病、性传播疾病、hiv感染、hiv复制、hiv相关性腹泻、手术相关创伤、外科手术诱导的转移性疾病、败血症、体重减轻、厌食、发烧控制、恶病质、伤口愈合、溃疡、肠屏障功能、过敏、哮喘、呼吸病症、循环病症、冠心病、贫血、凝血系统疾病、肾病、中枢神经系统病症、肝病、局部缺血、营养病症、骨质疏松症、内分泌病症、表皮病症、银屑病和寻常痤疮。对细胞因子产生的影响对于所检查的益生菌菌株是特异性的。因此,可以选择特异性的益生菌菌株以使特定疾病类型的特有的专有细胞因子失衡正常化。定制疾病特异性疗法可以使用an1206的单一菌株或其突变体或变体或选择这些菌株来完成。
肠道菌群对肠道免疫系统的发育和正常功能很重要。在没有肠道菌群的情况下,肠道免疫系统是未充分发育的,如在无菌动物模型所证明的,并且某些功能参数减少,如巨噬细胞吞噬能力和免疫球蛋白产生。肠道菌群在刺激非破坏性免疫应答中的重要性变得越来越明显。西方世界过敏的发生和严重程度的增加与医疗卫生(hygieneandsanitation)的增加有关,同时伴随着宿主遇到的传染性攻击的数量和范围的减少。这种免疫刺激的缺乏可能使宿主对非致病性但抗原性的物质反应,导致过敏或自身免疫。故意食用一系列非致病性免疫调节细菌将为宿主提供必要和适当的有教育意义的刺激,以正确开发和控制免疫功能。
炎症是用于描述在有持续物理损伤、感染或有持续免疫应答的部位的液体、血浆蛋白和白细胞的局部积聚的术语。炎性应答的控制在许多水平上发挥作用。控制因子包括细胞因子、激素(如氢化可的松)、前列腺素、或活性中间体和白三烯。细胞因子是低分子量生物活性蛋白,其参与免疫和炎性应答的产生和控制,同时还调节发育、组织修复和造血作用。它们提供了白细胞本身之间以及与其他细胞类型之间的交流手段。大多数细胞因子是多效的并且表达多种生物学上重叠的活性。细胞因子级联和网络控制炎性应答而不是特定细胞因子对特定细胞类型的作用。炎性应答的减弱导致适当的活化信号和其他导致炎性应答中止的炎性介质浓度更低。tnfα是一种关键的促炎细胞因子,因为他启动一系列细胞因子和生物效应的级联,导致炎性状态。因此,抑制tnfα的物质目前用于治疗炎性疾病,如英夫利昔单抗。
促炎细胞因子被认为在许多炎性疾病(包括炎性肠病(ibd))的致病机制中具有重要作用。目前用于治疗ibd的疗法旨在降低这些促炎细胞因子,包括il-8和tnfα的水平。此类疗法还可在治疗系统性炎性疾病如类风湿性关节炎中发挥重要作用。
本发明的菌株可以在一系列炎性疾病治疗中具有潜在的应用,特别是如果与其他抗炎疗法(如非固醇类抗炎药(nsaid)或英夫利昔单抗)组合使用。
在广谱的肿瘤类型中多功能细胞因子的产生表明癌症患者正在发生显著的炎性应答。目前尚不清楚这种应答对体内肿瘤细胞的生长和发育有何保护作用。然而,这些免疫应答可能对携带肿瘤的宿主产生不利影响。复杂的细胞因子相互作用涉及肿瘤和正常组织中细胞因子产生和细胞增殖的调节。人们早就认识到,体重减轻(恶病质)是癌症患者死亡的最常见原因,最初的营养不良表明预后不良。为了肿瘤的生长和扩散,它必须诱导新血管的形成并降解细胞外基质。炎性应答可能在上述机制中起重要作用,因此导致宿主的衰退和肿瘤的进展。由于长双歧杆菌的抗炎特性,这些细菌菌株它们可降低恶性细胞转化的速率。此外,肠道细菌可以从膳食化合物中产生具有遗传毒性、致癌的和肿瘤促进活性的物质,肠道细菌可以将促致癌剂活化为dna反应剂。通常,与肠道内的其他群体(如拟杆菌(bacteroide)、真杆菌(eubacteria)和梭菌(clostridia))相比,双歧杆菌物种具有低的异生物质代谢酶活性。因此,增加肠道中双歧杆菌细菌的数量可以有利地改变这些酶的水平。
大多数病原生物通过粘膜表面进入。这些部位的有效疫苗接种防止特定感染物质的入侵。迄今为止,口服疫苗接种策略已经集中在减毒活病原生物或纯化的胶囊化抗原的使用上。在体内,工程化以从感染物质中产生抗原的益生菌细菌可以提供有吸引力的替代品,因为这些细菌被认为对人类食用是安全的(grsa状态)。
小鼠研究表明,食用表达外源抗原的益生菌细菌可以引发保护性免疫应答。编码破伤风毒素c片段(ttfc)的基因在乳乳球菌(lactococcuslactis)中表达,并通过口服途径免疫小鼠。该系统能够诱导显著足够高的抗体滴度以保护小鼠免受致死毒素攻击。除抗原递呈外,活细菌载体还可以在体内产生生物活性化合物,如免疫刺激细胞因子。分泌生物活性人il-2或il-6和ttfc的乳乳球菌在鼻内免疫小鼠中诱导了10-15倍高的血清igg滴度。然而,对于这种特定的细菌菌株,通过与这些细胞因子共表达,没有增加总iga水平。其他细菌菌株,如格氏链球菌(streptococcusgordonii),也正在检测它们作为粘膜疫苗的可用性。定植于小鼠口腔和阴道腔的重组格氏链球菌诱导了对该细菌表达的抗原的黏膜和全身抗体应答。因此,使用益生菌细菌作为载体的口服免疫不仅保护宿主免受感染,而且可以取代病原体通常会引发的免疫刺激,因此有助于宿主的免疫教育。
益生元
益生菌生物的引入是通过摄取合适载体中的微生物来实现的。提供一种促进这些益生菌菌株在大肠中生长的培养基将是有利的。添加一种或多种寡糖、多糖或其他益生元增强了胃肠道中乳酸细菌的生长。益生元是指任何非活食物成分,其在结肠中由被认为具有积极价值的固有细菌(如双歧杆菌(bifidobacteria)、乳杆菌(lactobacilli))特异性发酵。益生元的类型可包括含有果糖、木糖、大豆、半乳糖、葡萄糖和甘露糖的那些。组合施用益生菌菌株和一种或多种益生元化合物可以增强所施用的益生菌在体内的生长,从而产生更显著的健康益处,并被称为合生素(synbiotic)。
其他活性成分
应当理解的是,益生菌菌株可以预防性施用或作为治疗方法单独或与如上所述的其他益生菌和/或益生元材料一起施用。此外,该细菌可以用作使用其他活性材料(例如用于治疗炎症或其他病症的那些,尤其是具有免疫参与的那些)的预防或治疗方案的一部分。这些组合可以在单一制剂中施用,或在相同或不同时间以单独制剂施用,以及用相同或不同的施用途径。
药物组合物
药物组合物是包含治疗有效量的药物活性物种或由治疗有效量的药物活性物质组成的组合物。它优选包括药学上可接受的载体、稀释剂或赋形剂(包括其组合)。用于治疗用途的可接受的载体或稀释剂在制药领域中是熟知的,并且描述于如remington'spharmaceuticalsciences中。药物载体、赋形剂或稀释剂的选择可以根据预期的施用途径和标准药学实践来选择。药物组合物可以包含载体、赋形剂或稀释剂,或除此之外-任何合适的粘合剂、润滑剂、悬浮剂、包衣剂、增溶剂、推进剂。
药学上可接受的载体的实例包括如水、盐溶液、醇、硅酮、蜡、凡士林、植物油、聚乙二醇、丙二醇、脂质体、糖、明胶、乳糖、直链淀粉、硬脂酸镁、滑石、表面活性剂、硅酸、粘稠石蜡、芳香油、脂肪酸单甘油酯和甘油二酯、石油脂肪酸酯、羟甲基纤维素、聚乙烯吡咯烷酮等。
在适当的时候,药物组合物可以通过以下任何一种或多种施用:吸入,以栓剂或子宫脱的形式,以洗剂、溶液、乳膏、软膏或粉剂(dustingpowder)的形式表面施用,通过使用皮肤贴剂,以含有赋形剂(如淀粉或乳糖)的片剂形式、或以单独或与赋形剂混合的胶囊或ovule、或以含有调味剂或着色剂的酏剂、溶液或悬浮液的形式口服施用,或可肠道外注射,如海绵体内、静脉内、肌肉内或皮下。对于肠道外施用,该组合物最好以无菌水溶液的形式使用,其可含有其他物质,如足够的盐或单糖,以使溶液与血等渗。对于口腔或舌下施用,该组合物可以以片剂或锭剂(其可以以常规方式配制)的形式施用。
可以使用鼻喷雾剂、鼻洗液、或鼻内直接应用完成鼻内施用。
对肺的施用可以是干粉形式,使用吸入装置吸入。在某些情况下,制剂是气溶胶形式。气溶胶可以是溶液、悬浮液、喷雾、雾、蒸气、液滴、颗粒或干粉,例如,使用连续喷雾器的包括hfa推进剂的方法剂量吸入器、具有非hfa推进剂的计量剂量吸入器、雾化器、加压罐。
可以将制剂设计成胶囊化、除去和/或灭活病毒。可选地或另外地,该制剂可以阻止病毒进一步感染呼吸道。
为了有助于在呼吸道中,如在鼻腔中,递送和维持,该制剂可具有1厘泊到2000厘泊的理想粘度,如5cps到500cps,或5cps到300cps。可以使用任何合适的粘度改良剂来获得理想粘度。这些物质可以是合适的天然的或合成的聚合物材料,如羟丙基甲基纤维素。
取决于不同的递送系统,可能存在不同的组合物/制剂要求。举例来说,本发明的药物组合物可以配制成使用微型泵或通过粘膜途径递送,如作为吸入或可摄入溶液的鼻喷雾剂或气溶胶,或肠胃外递送,其中组合物通过可注射形式配制,用于通过例如静脉内、肌肉内或皮下途径递送。可选地,可以将制剂设计成通过两种途径递送。
本发明不限于本文以上所述的实施方案,其细节可变化。
参考文献
bouhnik,y.,survieeteffetschezl’hommedesbactériesingéréesdansleslaitsfermentés.lait1993,73;241-247.
yangjw.,fanlc.,miaoxy.,maob.,limh.,luhw.,liangs.,xujf.corticosteroidsforthetreatmentofhumaninfectionwithinfluenzavirus:asystematicreviewandmeta-analysis.clinmicrobiolinfect.2015oct;21(10):956-63.
davidsons.,crottas.,mccabetm.,wacka.pathogenicpotentialofinterferonαβinacuteinfluenzainfection.natcommun.2014may21;5:3864.
nakamuras.,daviskm.,weiserjn.synergisticstimulationoftypeiinterferonsduringinfluenzaviruscoinfectionpromotesstreptococcuspneumoniaecolonizationinmice.jclininvest2011;121:3657–65.
shahangiana.,chowek.,tianx.etal.typeiifnsmediatedevelopmentofpostinfluenzabacterialpneumoniainmice.jclininvest2009;119:1910–20.
liw.,moltedob.,morantm.typeiinterferoninductionduringinfluenzavirusinfectionincreasessusceptibilitytosecondarystreptococcuspneumoniaeinfectionbynegativeregulationofgammadeltatcells.jvirol2012;86:12304–12.
hewittr.,farneh.,ritchiea.,lukee.,johnstonsl.,malliap.theroleofviralinfectionsinexacerbationsofchronicobstructivepulmonarydiseaseandasthma.theradvrespirdis.2016apr;10(2):158-74.doi:10.1177/1753465815618113.
galaniie.,triantafylliav.,eleminiadouee.,koltsidao.,stavropoulosa.,manioudakim.,thanosd.,doylese.,kotenkosv.,thanopoulouk.,andreakose.interferon-λmediatesnon-redundantfront-lineantiviralprotectionagainstinfluenzavirusinfectionwithoutcompromisinghostfitness.immunity.2017may16;46(5):875-890.
davidsons.,mccabetm.,crottas.,gadhh.,hesselem.,beinkes.,hartmannr.,wacka.ifn-λisapotentanti-influenzatherapeuticwithouttheinflammatorysideeffectsofifn-αtreatment.embomolmed.2016sep1;8(9):1099-112.
thiel,s.,andreidk.1989–structuresandfunctionsassociatedwiththegroupofmammalianlectinscontainingcollagen-likesequences.febslett.250:78.2.
sastry,k.,andezekowitzr.a.1993.collectins:patternrecognitionmoleculesinvolvedinfirstlinehostdefense.curr.opin.immunol.5:59.
shix.,zhouw.,huangh.,zhuh.,zhoup.,zhuh.,jud.inhibitionoftheinflammatorycytokinetumornecrosisfactor-alphawithetanerceptprovidesprotectionagainstlethalh1n1influenzainfectioninmice.crit.care,2013;17(6);r301.
bartlettnw1.,singanayagama.,johnstonsl.mousemodelsofrhinovirusinfectionandairwaysdisease.methodsmolbiol.2015;1221:181-8.doi:10.1007/978-1-4939-1571-2_14.ivashkivlb.,anddonlinlt.regulationoftypeiinterferonresponses.natrevimmunol.2014jan;14(1):36–49.
lazearhm.,nicetj.,diamondms.interferon-immunefunctionsatbarriersurfacesandbeyond.immunity,2015jul21;43(1):15-28.
wacka.,terczyńska-fylae.,hartmannr.guardingthefrontiers:thebiologyofthetypeiiiinterferons.natimmunol.2015aug;16(8):802-9.
durbinrk.,kotenkosv.,durbinje.interferoninductionandfunctionatthemuscolsurface.immunolrev.2013sep;255(1):25-39.
mendozajl.,schneiderwm.,hoffmannhh.,vercauterenk.,judekm.,xionga.,moragai.,hortontm.,glennjs.,dejongyp.,ricecm.,garciakc.theifn--ifn-r1-il-10rβcomplexrevealsstructuralfeaturesunderlyingtypeiiiifnfunctionalplasticity.immunity.2017mar21;46(3):379-392.
ichikawaa.,kubak.,moritam.,chidas.,tezukah.,harah.,sasakit.,ohtekit.,ranierivm.,dossantoscc.,kawaokay.,akiras.,lusterad.,lub.,penningerjm.,uhligs.,slutskyas.,imaiy.cxcl10-cxcr3enhancesthedevelopmentofneutrophil-mediatedfulminantlunginjuryofviralandnonviralorigin.amjrespircritcaremed.2013jan1;187(1):65-77.
weiwang.,penghuiyang.,etal.monoclonalantibodyagainstcxcl-10/ip-10amelioratesinfluenzaa(h1n1)virusinducedacutelunginjury.cellresearch(2013)23:577-580.
hartshornkl.,whitemr.,shepherdv.,reidk.,jenseniusjc.,crouchec.mechanismsofanti-influenzaactivityofsurfactantproteinsaandd:comparisonwithserumcollectins.amjphysiol1997;273:l1156–l1166.
readingpc.,moreyls.,crouchec.,andersem.collectin-mediatedantiviralhostdefenseofthelung:evidencefrominfluenzavirusinfectionofmice.jvirol1997;71:8204–8212.
hartshornkl.,whitemr.,voelkerdr.,coburnj.,zanerk.,crouchec.mechanismofbindingofsurfactantproteindtoinfluenzaaviruses:importanceofbindingtohaemagglutinintoantiviralactivity.biochemj2000;351(pt2):449–458.
teclet.,whitemr.,crouchec.,hartshornkl.inhibitionofinfluenzaviralneuraminidaseactivitybycollectins.archvirol2007;152:1731–1742.
levineam.,whitsettja.,hartshornkl.,crouchec.,korfhagentr.sp-denhancesclearanceofinfluenzaavirusfromthelungininvivomousemodels.jimmunol2001;167:5868–5873.
vigerustdj.,ulettkb.,boydkl.,madsenj.,hawgoods.,mccullersja.n-linkedglycosylationattenuatesh3n2influenzaviruses.jvirol2007;81:8593–8600.hawgoods.,brownc.,edmondsonj.,stumbaugha.,allenl.,goerkej.etal.pulmonarycollectinsmodulatestrain-specificinfluenzaavirusinfectionandhostresponses.jvirol2004;78:8565–8572.
levineam(1).,elliottj.,whitsettja.,srikiatkhachorna.,crouche.,desilvan.,korfhagent.surfactantprotein-denhancesphagocytosisandpulmonaryclearanceofrespiratorysyncytialvirus.amjrespircellmolbiol.2004aug;31(2):193-9.
richh.,robinsonke.,mchughkj.,clayme.,alcornjf.theroleofinterferonlambdaduringinfluenza,staphylococcusaureussuper-infection.jimmunolmay1,2017,198(1supplement)77.16.
jounblatr.,clarkh.,eggletonp.,hawgoods.,andrewpw.,kadioglua.theroleofsurfactantproteindinthecolonisationoftherespiratorytractandonsetofbacteraemiaduringpneumococcalpneumoniarespirres.2005oct28;6:126.
worldhealthorganisation2007.globalsurveillance,preventionandcontrolofchronicrespiratorydiseases,acomprehensiveapproach.(editorsjeanbousquetandnikolaikhaltaev).
jarttit.,gernje.roleofviralinfectionsinthedevelopmentandexacerbationofasthmainchildren.jallergyclinimmunol.2017oct;140(4):895-906.
almondmh1.,edwardsmr.,barclayws.,johnstonsl.obesityandsusceptibilitytosevereoutcomesfollowingrespiratoryviralinfection.thorax.2013jul;68(7):684-6.doi:10.1136/thoraxjnl-2012-203009.
steinkejw.,borishl.immuneresponsesinrhinovirus-inducedasthmaexacerbations.currallergyasthmarep.2016nov;16(11):78.
zhoux.,liq.,zhoux.exacerbationofchronicobstructivepulmonarydisease.cellbiochembiophys.2015nov;73(2):349-55.
序列表
<110>营养健康有限公司
<120>可以有益地调节针对呼吸道病毒感染的免疫应答的长双歧杆菌
<130>alim70/c/wo
<150>ep17158564.9
<151>2017-02-28
<150>ep17158567.2
<151>2017-02-28
<160>5
<170>patentinversion3.5
<210>1
<211>18
<212>dna
<213>bifidobacteriumlongum
<400>1
ctggtgccaaggcatcca18
<210>2
<211>19
<212>dna
<213>bifidobacteriumlongum
<400>2
gctggatcacctcctttct19
<210>3
<211>498
<212>dna
<213>bifidobacteriumlongum
<400>3
ttgctgggatcacctcctttttacggagaattcagtcggatgttcgtccgacggtgtgcg60
ccccgcgcgtcgcatggtgcgatggcggcggggttgctggtgtggaaaacgtcgttggct120
ttgccctgccggtcgtgcggtgggtgcggggtggtatggatgcgcttttgggctcccgga180
tcgccaccccaggctttttgcctggcgcgattcgatgcccgtcgtgcctgggggccggcc240
gtgtgccggcgcgatggcgtggcggtgcgtggtggcttgagaactggatagtggacgcga300
gcaaaacaagggtttttgaatctttgttttgctgttgatttcgaatcgaactctattgtt360
cgtttcgatcgttttgtgatcatttttagtgtgatgatttgtcgtcctgggaatttgcta420
gaggaatacttgcgggccatgcactttcgtggtgtgtgttgcttgcaagggcgtatggtg480
gaggccttggcaccagaa498
<210>4
<211>20
<212>dna
<213>bifidobacteriumlongum
<400>4
ggactgcagcgtagacgctt20
<210>5
<211>21
<212>dna
<213>bifidobacteriumlongum
<400>5
catcctgtatatgaggcccat21
pct/ro/134表
- 还没有人留言评论。精彩留言会获得点赞!
- 一种治疗感冒的中药组合物的制作方法
- 阿掏比克酸三唑基与1h?四氮唑基衍生物的组合物用于制备抗病毒药物的制作方法
- Schiglautone A衍生物的组合物用于制备抗病毒药物的制作方法
- Artalbic acid的O?(苯并咪唑基)乙基与O?(二羟乙胺基)乙基衍生物的组合物在抗病毒 ...的制作方法
- 一种白头翁与蛴螬有效部位配伍后抗rsv病毒的制备方法
- 表达信号肽替换的呼吸道合胞病毒f蛋白的重组新城疫耐热疫苗株及制备方法
- 一种蛴螬中可抗呼吸道合胞病毒的有效部位及制备方法
- Salviskinone A的苯并咪唑基和二(2-甲硫基乙基)胺基衍生物的组合物在抗病毒药物中 ...的制作方法
- 一种呼吸道合胞体病毒疫苗及其制备方法与应用
- Virosaine A的四氢吡咯基和吗啉基衍生物的组合物在抗病毒药物中的应用