基于CRISPR/Cas9系统在Beta-TC-6细胞株中剔除Sidt2基因方法与流程
本发明涉及医疗基因工程领域,尤其涉及基于crispr/cas9系统在beta-tc-6细胞株中剔除sidt2基因方法。
背景技术:
crispr/cas9基因编辑系统成为继锌指蛋白(zfps)、转录激活子样效应因子核酸酶(tales)之后的第三代基因编辑技术。与传统的基因编辑工具相比具有诸多的优点,首先crispr/cas9基因编辑系统适应性和操作性强,来自产脓链球菌和嗜热链球菌的cas9由于pam识别序列仅为2个碱基(gg),几乎可以在所有的基因中找到大量靶点。cas9蛋白在目前测试过的几乎所有生物和细胞中均有活性,包括细菌、酵母、植物、鱼、以及哺乳动物细胞;其次,crispr/cas9基因编辑系统简单易学,目前已有多种成熟的crispr/cas9基因编辑系统质粒载体,常规的分子生物学实验室即可开展构建,只需要将长约25bp左右的sgrna双链连接到线性化的载体中,构建难度相对于常见的表达载体和报告基因载体小,构建完成的载体只需要通过菌落pcr或者测序即可鉴定,不必像构建表达载体担心长片段的碱基突变问题。但是crispr/cas9质粒需要表达cas9蛋白相对于其他的载体偏大,一般常常在10kbp以上,转染细胞难度较大,尤其对于难转染细胞更是雪上加霜。虽然可以通过对成功转染的细胞表达的抗生素筛选或者荧光标记可进行筛选,但是转染效率低对于后续的筛选和单克隆培养造成很大的困难,并且cas9蛋白具有一定的细胞毒性,后续的抗生素筛选和流式分选也会对转染成功的细胞也会有影响。因此提高crispr/cas9质粒的转染效率是急需解决的问题。
目前现有质粒转染主流的方法包括脂质体转染、阳离子非脂质体转染、磷酸钙转染等方法。这些方法有各自的优点,同时也存在着缺点。比如脂质体转染效率高,但是对于一些原代细胞或者难转染的分泌性的细胞转染效率较低。电穿孔也称为电转染,是通过高强度的电场作用,使细胞膜上形成可逆的瞬时通道,质粒在电场的作用下通过通道进入细胞内。电穿孔转染相比于其他转染方法具有操作简单、对于原代细胞和难转染细胞效率高、重复性好等优点。但是电穿孔转染对于不同的细胞系转染参数差异较大,因此需要针对不同的细胞系进行电转染参数的优化,目前比较成熟的运用电穿孔的细胞都是常见细胞系或者原代细胞,对胰岛细胞系使用电穿孔较少,也没对其进行电转染参数进行优化。另外质粒的纯度以及浓度、细胞的状态等都会对电转的效率产生影响。电转参数优化时一般在保证转染质粒纯度浓度以及细胞状态的前提下,通过固定穿孔时间,摸索不同穿孔电压梯度,以摸索细胞系最优穿孔电压。
根据上述,本发明针对小鼠胰岛素瘤胰岛β细胞(beta-tc-6)难转染以及crispr/cas9质粒分子较大的特点,采取电穿孔进行cas9质粒的递送,并且设置不同的穿孔电压梯度进行电转参数的优化,从而实现了在小鼠胰岛素瘤胰岛β细胞(beta-tc-6)中高效的递送crispr/cas9质粒;同时,本发明通过将crispr/cas9基因编辑系统和电穿孔相结合,克服了crispr/cas9质粒难转导的特点,为常规的分子生物学实验室开展crispr/cas9基因基因编辑系统提供了切实可行的办法。
技术实现要素:
本发明所要解决的技术问题在于:提供基于crispr/cas9系统在beta-tc-6细胞株中剔除sidt2基因方法,来克服crispr/cas9质粒难转导的特点,从而实现了在小鼠胰岛素瘤胰岛β细胞(beta-tc-6)中高效的递送crispr/cas9质粒,为常规的分子生物学实验室开展crispr/cas9基因基因编辑系统提供了切实可行的办法。
为解决上述技术问题,本发明的技术方案是:基于crispr/cas9系统在beta-tc-6细胞株中剔除sidt2基因方法,具体实验步骤如下:
(1)、sgrnaoligodna序列合成;利用sgrna在线设计工具针对小鼠sidt2基因设计长度为20bp左右的oligodna,sgrnassdna序列f链5’端需要添加cacc从而可以和bbsi酶切后的载体互补,需要注意的是f链的第一个碱基必须是g,如果选取的guide序列的第一个碱基不是g,可自行添加一个g以增强u6启动子的活性,r链的5’端添加aaac。另外,需在位点上下游各设计一条引物,用于后续检测阳性克隆,引物能扩增约300bp的dna片段,上游引物距突变位点约100bp,下游引物距突变位点约200bp。将设计后的序列进行合成,纯化级别为page;
(2)、oligodna退火:合成的sgrna稀释为终浓度为100um,配制退火反应体系:稀释的sgrna各1ul,2ulannealingbufferfordnaoligos(5x),用超纯水补足到10ul。pcr仪中95℃10分钟,随后取出室温冷却两个小时。稀释10倍后储存在-20℃备用;
(3)、px459重组质粒的构建:px459质粒为含有u6启动子的sgrna骨架的表达载体。(a)px459载体的线性化:使用bbsi限制性内切酶线性化px459载体,使用胶回收试剂盒回收线性化px459载体。(b)sgrna与线性化px459连接:按照t4连接酶说明书配置反应体系,pcr中25℃反应3小时。(c)连接产物转化感受态细胞dh5α,37℃培养过夜。次日挑取菌落摇菌抽提质粒;
(5)、px459重组质粒的验证:质粒送生物公司测序,测序引物为u6启动子通用引物,根据测序结果鉴定sgrna是否正确连接到载体中。测序成功的质粒命名为px459-sidt2,使用去内毒素质粒大量提取试剂盒抽提质粒,要求质粒浓度大于1ug/ul并且纯度达到电转染要求;
(6)、beta-tc-6细胞的培养:细胞培养条件为含15%胎牛血清的dmem高糖培养基,细胞汇合度达到70%左右进行传代;
(7)、px459-sidt2质粒的电转:电转前细胞融合度达到80%左右,胰酶消化细胞后加入opti-mem使其成为单细胞悬液并且调整细胞数量为1x106个/90ul,吸取90ul细胞悬液加入10ul质粒轻轻混匀后加入电转杯中。设置导入电转参数:脉冲电压为20v,脉冲时间为20ms,脉冲间隔为50ms,脉冲次数为5次,电压衰减为40%,电穿孔模式为正方向。穿孔电转参数为:固定脉冲时间5ms,脉冲间隔50ms,脉冲次数2次,电压衰减10%,电穿孔模式为正方向,设置脉冲电压梯度为125v、150v、175v、200v。完成电转染的细胞在六孔板中培养过夜;
(8)、阳性细胞的筛选:电转染48小时加入嘌呤霉素至终浓度为3ug/ml进行筛选(提前需要进行嘌呤霉素筛选梯度的摸索,本实验室小鼠胰岛素瘤胰岛β细胞beta-tc-6的筛选浓度为3ug/ml),72小时后换为正常培养条件,扩增冻存;
(9)、t7酶和westernblot检测:野生型细胞以及阳性细胞一部分使用试剂盒提取dna,一部分使用细胞裂解液提取全细胞蛋白。提取的dna通过pcr对sgrna的靶向区域进行扩增,扩增引物的退火温度54℃。(1)t7酶切分析:按照说明书进行操作,酶切后的产物通过2%琼脂糖凝胶分析。(2)westernblot检测:使用微量分光光度计测定蛋白浓度,取等量的蛋白加入蛋白上样缓冲液,煮沸后进行sds-page电泳,电泳完成后湿转到pvdf上,用5%的脱脂奶粉封闭一个小时,一抗孵育过夜,第二天进行二抗孵育,加入曝光底物后使用超灵敏多功能成像仪进行曝光,以β-actin作为内参;
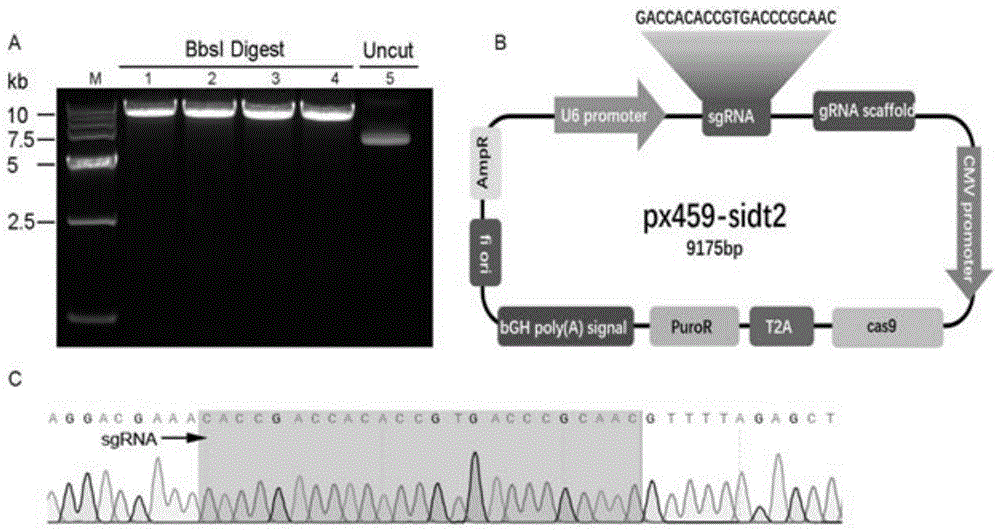
(10)、px459-sidt2克隆构建:px459空载体经bbsi酶切线性化后,从琼脂糖凝胶中回收酶切后的线性化片段。将退火的sgrna双链和回收的线性化px459载体相连,构建px459-sidt2重组质粒。构建的px459-sidt2经测序表明插入序列的位置、方向均正确,重组质粒px459-sidt2构建成功;
(11)、t7e1酶及westernblot检测小鼠sidt2基因的敲除效果:t7e1酶结果显示,电转染px459-sidt2的混合克隆细胞有一条明显的切开的条带,即混合克隆细胞中出现碱基错配,表明px459-sidt2在beta-tc-6细胞中实现靶向敲除。westernblot检测敲除效果,结果显示同野生型细胞比较,混合克隆细胞sidt2蛋白表达显著降低(t=6.668,p=0.0026)。进一步实验需要进行单克隆培养,从而得到敲除效率为100%的克隆细胞株。
进一步,所述的步骤(1)-步骤(11)中使用的材料为质粒载体pspcas9(bb)-2a-puro(px459)(addgeneplasmid#62988);感受态细胞dh5α;beta-tc-6小鼠胰岛素瘤胰岛β细胞。
进一步,所述的步骤(1)-步骤(11)中使用的主要试剂及仪器为bbsi限制性内切酶、t4dnaligase、t7核酸内切酶ⅰ;opti-mem减血清培养基;胎牛血清(标准级);annealingbufferfordnaoligos(5x)、β-actin抗体;sidt2兔多克隆抗体;pcr仪;蛋白印记系统;nepa21高效基因转染系统;电转杯。
进一步,所述的第(7)步骤之后电转染后抗生素筛选,结果显示脉冲电压为150v时转染效率最高并且细胞状态没有明显改变,因此确定beta-tc-6细胞电转染px459-sidt2最优电转穿孔电转参数为:脉冲电压150v,脉冲时间5ms,脉冲间隔50ms,脉冲次数2次,电压衰减10%,电穿孔模式为正方向。
该基于crispr/cas9系统在beta-tc-6细胞株中剔除sidt2基因方法,针对小鼠胰岛素瘤胰岛β细胞(beta-tc-6)难转染以及crispr/cas9质粒分子较大的特点,通过采取电穿孔进行cas9质粒的递送,并设置不同的穿孔电压梯度进行电转参数的优化,从而实现了在小鼠胰岛素瘤胰岛β细胞(beta-tc-6)中高效的递送crispr/cas9质粒;同时,通过将crispr/cas9基因编辑系统和电穿孔相结合,克服了crispr/cas9质粒难转导的特点,为常规的分子生物学实验室开展crispr/cas9基因基因编辑系统提供了切实可行的办法,具有极高的推广应用价值。
附图说明
图1是成功构建px489-sidt2克隆;
图2是设置脉冲电压梯度加压筛选72小时后的阳性细胞;
图3是t7e1酶及westemblot检测小鼠sidt2基因的敲除效果;
图4是小鼠sidt2sgrna及验证引物序列。
如下具体实施方式将结合上述附图进一步说明。
具体实施方式
在下文中,阐述了多种特定细节,以便提供对构成所描述实施例基础的概念的透彻理解。然而,对本领域的技术人员来说,很显然所描述的实施例可以在没有这些特定细节中的一些或者全部的情况下来实践。在其他情况下,没有具体描述众所周知的处理步骤。
实施例1
如图1、图2、图3和图4所示,基于crispr/cas9系统在beta-tc-6细胞株中剔除sidt2基因方法,具体实验步骤如下:
sgrnaoligodna序列合成;利用sgrna在线设计工具针对小鼠sidt2基因设计长度为20bp左右的oligodna,sgrnassdna序列f链5’端需要添加cacc从而可以和bbsi酶切后的载体互补,需要注意的是f链的第一个碱基必须是g,如果选取的guide序列的第一个碱基不是g,可自行添加一个g以增强u6启动子的活性,r链的5’端添加aaac。另外,需在位点上下游各设计一条引物,用于后续检测阳性克隆,引物能扩增约300bp的dna片段,上游引物距突变位点约100bp,下游引物距突变位点约200bp。将设计后的序列进行合成,纯化级别为page。oligodna退火:合成的sgrna稀释为终浓度为100um,配制退火反应体系:稀释的sgrna各1ul,2ulannealingbufferfordnaoligos(5x),用超纯水补足到10ul。pcr仪中95℃10分钟,随后取出室温冷却两个小时。稀释10倍后储存在-20℃备用。px459重组质粒的构建:px459质粒为含有u6启动子的sgrna骨架的表达载体。(a)px459载体的线性化:使用bbsi限制性内切酶线性化px459载体,使用胶回收试剂盒回收线性化px459载体。(b)sgrna与线性化px459连接:按照t4连接酶说明书配置反应体系,pcr中25℃反应3小时。(c)连接产物转化感受态细胞dh5α,37℃培养过夜。次日挑取菌落摇菌抽提质粒。px459重组质粒的验证:质粒送生物公司测序,测序引物为u6启动子通用引物,根据测序结果鉴定sgrna是否正确连接到载体中。测序成功的质粒命名为px459-sidt2,使用去内毒素质粒大量提取试剂盒抽提质粒,要求质粒浓度大于1ug/ul并且纯度达到电转染要求。beta-tc-6细胞的培养:细胞培养条件为含15%胎牛血清的dmem高糖培养基,细胞汇合度达到70%左右进行传代。px459-sidt2质粒的电转:电转前细胞融合度达到80%左右,胰酶消化细胞后加入opti-mem使其成为单细胞悬液并且调整细胞数量为1x106个/90ul,吸取90ul细胞悬液加入10ul质粒轻轻混匀后加入电转杯中。设置导入电转参数:脉冲电压为20v,脉冲时间为20ms,脉冲间隔为50ms,脉冲次数为5次,电压衰减为40%,电穿孔模式为正方向。穿孔电转参数为:固定脉冲时间5ms,脉冲间隔50ms,脉冲次数2次,电压衰减10%,电穿孔模式为正方向,设置脉冲电压梯度为125v、150v、175v、200v。完成电转染的细胞在六孔板中培养过夜。阳性细胞的筛选:电转染48小时加入嘌呤霉素至终浓度为3ug/ml进行筛选(提前需要进行嘌呤霉素筛选梯度的摸索,本实验室小鼠胰岛素瘤胰岛β细胞beta-tc-6的筛选浓度为3ug/ml),72小时后换为正常培养条件,扩增冻存。t7酶和westernblot检测:野生型细胞以及阳性细胞一部分使用试剂盒提取dna,一部分使用细胞裂解液提取全细胞蛋白。提取的dna通过pcr对sgrna的靶向区域进行扩增,扩增引物的退火温度54℃。(1)t7酶切分析:按照说明书进行操作,酶切后的产物通过2%琼脂糖凝胶分析。(2)westernblot检测:使用微量分光光度计测定蛋白浓度,取等量的蛋白加入蛋白上样缓冲液,煮沸后进行sds-page电泳,电泳完成后湿转到pvdf上,用5%的脱脂奶粉封闭一个小时,一抗孵育过夜,第二天进行二抗孵育,加入曝光底物后使用超灵敏多功能成像仪进行曝光,以β-actin作为内参。px459-sidt2克隆构建:px459空载体经bbsi酶切线性化后(图1a),从琼脂糖凝胶中回收酶切后的线性化片段。将退火的sgrna双链和回收的线性化px459载体相连,构建px459-sidt2重组质粒。构建的px459-sidt2经测序表明插入序列的位置、方向均正确(图1c),重组质粒px459-sidt2构建成功(图1b)。t7e1酶及westernblot检测小鼠sidt2基因的敲除效果:t7e1酶结果显示,电转染px459-sidt2的混合克隆细胞有一条明显的切开的条带(图3a),即混合克隆细胞中出现碱基错配,表明px459-sidt2在beta-tc-6细胞中实现靶向敲除。westernblot检测敲除效果,结果显示同野生型细胞比较,混合克隆细胞sidt2蛋白表达显著降低(t=6.668,p=0.0026)(图3b、c)。进一步实验需要进行单克隆培养,从而得到敲除效率为100%的克隆细胞株,所述的步骤(1)-步骤(11)中使用的材料为质粒载体pspcas9(bb)-2a-puro(px459)(addgeneplasmid#62988);感受态细胞dh5α;beta-tc-6小鼠胰岛素瘤胰岛β细胞,所述的步骤(1)-步骤(11)中使用的主要试剂及仪器为bbsi限制性内切酶、t4dnaligase、t7核酸内切酶ⅰ;opti-mem减血清培养基;胎牛血清(标准级);annealingbufferfordnaoligos(5x)、β-actin抗体;sidt2兔多克隆抗体;pcr仪;蛋白印记系统;nepa21高效基因转染系统;电转杯,所述的第(7)步骤之后电转染后抗生素筛选,结果显示脉冲电压为150v时转染效率最高并且细胞状态没有明显改变,因此确定beta-tc-6细胞电转染px459-sidt2最优电转穿孔电转参数为:脉冲电压150v,脉冲时间5ms,脉冲间隔50ms,脉冲次数2次,电压衰减10%,电穿孔模式为正方向(图2)。
该基于crispr/cas9系统在beta-tc-6细胞株中剔除sidt2基因方法,针对小鼠胰岛素瘤胰岛β细胞(beta-tc-6)难转染以及crispr/cas9质粒分子较大的特点,通过采取电穿孔进行cas9质粒的递送,并设置不同的穿孔电压梯度进行电转参数的优化,从而实现了在小鼠胰岛素瘤胰岛β细胞(beta-tc-6)中高效的递送crispr/cas9质粒;同时,通过将crispr/cas9基因编辑系统和电穿孔相结合,克服了crispr/cas9质粒难转导的特点,为常规的分子生物学实验室开展crispr/cas9基因基因编辑系统提供了切实可行的办法,具有极高的推广应用价值。
本发明不局限于上述具体的实施方式,本领域的普通技术人员从上述构思出发,不经过创造性的劳动,所做出的种种变换,均落在本发明的保护范围之内。
- 还没有人留言评论。精彩留言会获得点赞!