一种高效重组且无筛选标记的痘苗病毒载体及其建立方法与流程
本发明涉及基因重组技术领域,尤其涉及一种高效重组且无筛选标记的痘苗病毒载体及其建立方法。
背景技术:
痘病毒(poxvirus)是一类有包膜的基因组较大的线型双链dna病毒,包括痘苗病毒(vacciniavirus)、天花病毒(variolavirus)、牛痘病毒(cowpoxvirus)和猴痘病毒(monkeypoxvirus)等。其具有非必需基因多,对外源基因的容量大(<25kbp)及胞质复制等特点,因而痘苗病毒和其它类型痘病毒能够以载体的形式可用于抵抗人类免疫缺陷病毒(humanimmunodeficiencyvirus,hiv)、流感病毒(influenzavirus)等的疫苗构建;更重要的是,通过对痘苗病毒特定基因进行敲除,能够使其具有肿瘤选择性,可以作为溶瘤病毒的载体成为肿瘤免疫治疗的新手段。
获得重组痘苗病毒载体的主要方法包括传统同源重组、直接体外连接法、细菌人工染色体等。但是这些方法存在重组效率低,病毒纯化耗时长等弊端,严重阻碍了痘苗病毒载体在临床上的应用,亟需高效重组痘苗病毒载体的技术方法。crispr(clusteredregularlyinterspacedshortpalindromicrepeat)-cas9是一种grna引导cas9蛋白对靶点切割的基因编辑技术,已成功编辑腺病毒(adenovirus)、单纯疱疹病毒(herpessimplexvirus)和痘病毒(poxvirus)lister株、westernreserve株基因组。而痘苗病毒天坛株(vacciniavirustiantanstrain,vtt)是中国自主研发、世界卫生组织文献记载中成功用于预防天花的疫苗毒株,在消灭天花的过程中发挥了重要的作用,已作为中国防止天花再次出现的应急储备疫苗毒种。因此,本申请拟在痘苗病毒天坛株的基础上通过crispr-cas9建立高效重组痘苗病毒载体系统。在重组痘苗病毒载体的建立过程中,为便于筛选,常引入外源筛选标记基因。利用筛选标记基因,可以从大量阴性克隆中将阳性重组克隆筛选出来,但随着重组病毒的传代,筛选标记基因不再有用,但却长期存在持续表达,产生安全性问题。为了满足重组痘苗病毒实际应用的需要,需要删除重组病毒中的筛选标记。因此,本申请在建立高效重组痘苗病毒载体系统之后,利用cre-loxp技术删除重组病毒中的筛选标记,建立高效重组且无筛选标记的痘苗病毒载体系统。
技术实现要素:
本发明针对上述技术问题,经过大量试验细节条件摸索,利用crispr-cas9技术和cre-loxp系统建立一种高效重组且无筛选标记的痘苗病毒载体。
为了实现上述目的,本发明提供如下技术方案:
一种高效重组且无筛选标记的痘苗病毒载体的建立方法,包括以下步骤:
s1、采用crispr-cas9技术敲除痘苗病毒tk基因,然后转染带有egfp的重组质粒pj2r-egfp-loxp,获得缺失tk带有egfp的重组病毒vacv-δtk-egfp-loxp;
s2、采用cre-loxp系统将带有egfp的重组病毒vacv-δ
tk-egfp-loxp中的筛选标记egfp删除,最终获得无egfp的重组病毒载体。
本申请首先对痘苗病毒基因组进行扫描分析,利用网站(http://www.e-crisp.o-rg/e-crisp/)预测靶向痘苗病毒tk区的3条grna,对tk区进行编辑,建立高效重组技术方法。为此,本研究通过瞬时转染grna-cas9质粒后,感染vtt并转入重组质粒pj2r-egfp-loxp,经过噬斑实验纯化、测序鉴定证明获得了带有egfp-loxp替换了tk区的重组痘苗病毒,而且此利用crispr-cas9获得重组痘苗病毒载体系统的重组效率比传统的同源重组方法大大提高,建立起初步的高效重组痘苗病毒载体系统。
cre-loxp是一种利用cre酶识别特异dna序列(loxp位点),使两个loxp位点间发生基因重组的系统。cre重组酶能够介导三种条件下两个loxp位点间的重组。一是在两个loxp位点位于一条dna链上并且方向相同的条件下,cre重组酶可以切除loxp位点间的序列;二是在两个loxp位点位于一条dna链上,但在方向相反的条件下,cre重组酶能够导致loxp位点间的序列倒位;三是在两个loxp位点分别位于两条不同的dna链或染色体上,cre酶能介导两条dna链的交换或染色体易位。本申请利用的是cre酶的删除功能,构建的重组质粒在荧光标记egfp两侧带有相同方向的loxp,在上述获得带有egfp-loxp的重组病毒的基础上,通过表达cre酶删除重组病毒中的筛选标记,获得删除tk且无筛选标记的重组痘苗病毒载体。
作为优选的,上述的高效重组且无筛选标记的痘苗病毒载体的建立方法,步骤s1通过转染grna-cas9质粒获得重组病毒或者通过grna-cas9细胞系获得重组病毒。
作为优选的,上述的高效重组且无筛选标记的痘苗病毒载体的建立方法,步骤s1病毒转染用细胞为293t细胞或hela细胞。
更具体地,步骤s1包括以下步骤:
(1)grna-cas9质粒的构建与鉴定
设计靶向tk区的grna序列;通过分子克隆方法将lenticrispr-v2中cas9的核定位信号nls缺失,然后将靶向tk区的grna序列导入lenti-delnls,获得带有胞质定位cas9并靶向tk的grna质粒;在293t细胞中转染lenti-delnls、lenti-delnls-grna,48h后裂解细胞,通过westernblot检测cas9的表达;
(2)重组质粒pj2r-egfp-loxp的构建与鉴定
在293t细胞中感染vtt,然后转染pj2r-egfp-loxp重组质粒;48h后,免疫荧光观察egfp的表达,构建的重组质粒pj2r-egfp-loxp主要包括p7.5、p11、h5启动子、egfp、loxp位点和tk区同源臂;
(3)重组病毒vacv-δtk-egfp-loxp的筛选
在293t细胞密度为60%~70%时,转染grna-cas9质粒;24h后,用dmem稀释野生型病毒,感染细胞;2h后,转染pj2r-egfp-loxp;4h后,换成2%fbs的培养基;48h后,收获重组病毒悬液,在vero细胞中利用病毒噬斑形成实验进行多轮筛选,最终获得重组病毒vacv-δ
tk-egfp-loxp;
(4)重组病毒vacv-δtk-egfp-loxp的鉴定
提取步骤(3)重组病毒dna,然后利用pcr扩增验证,再根据pcr产物测序,确证重组病毒在vtt的tk区准确插入egfp基因。
作为优选的,上述的高效重组且无筛选标记的痘苗病毒载体的建立方法,步骤s2的具体方法为:
(1)pqcxip-cre质粒的构建与鉴定
通过分子克隆方法将cre构建至pqcxip载体中;在293t细胞中转染质粒pqcxip-cre,48h后裂解细胞,通过westernblot检测;
(2)重组病毒vacv-δtk的筛选
在293t细胞密度为60%~70%时,转染pqcxip-cre质粒;24h后,用dmem稀释重组病毒vacv-δtk-egfp-loxp,感染细胞;2h后,换成2%fbs的培养基;48h后,收获重组病毒悬液,在vero细胞中利用病毒噬斑形成实验进行多轮筛选,最终获得无筛选标记的重组病毒vacv-δtk;
(3)重组病毒vacv-δtk的鉴定
提取步骤(6)重组病毒vacv-δtk的基因组,然后利用pcr扩增验证,再根据pcr产物测序,确证vacv-δtk在vacv-δtk-egfp-loxp的tk区准确删除egfp基因。
本发明还提供了采用上述建立方法制备得到的高效重组且无筛选标记的痘苗病毒载体。优选地,痘苗病毒采用中国天坛株痘苗病毒。天坛株痘苗病毒是我国自主研发用于预防天花的痘苗病毒。同时也是世界卫生组织允许预防天花的疫苗毒株。现在天坛株痘苗病毒已经作为防止天花再次出现的应急储备疫苗毒种。痘苗病毒作为载体可以携带外源蛋白,可以用于抗病毒疫苗载体构建与肿瘤的免疫治疗中。
与现有技术相比,本发明的有益效果为:
本发明提供的高效重组痘苗病毒载体的建立方法,通过分子克隆技术、westernblot、免疫荧光、病毒空斑形成实验,发现在293t细胞密度
60%~70%,转染grna-cas9质粒。24h后,用dmem稀释野生型病毒,感染细胞;2h后,转染pj2r-egfp-loxp,获得重组病毒vacv-δ
tk-egfp-loxp,重组效率能够达到10%以上。与传统方法比,能够提高10~20倍。在vacv-δtk-egfp-loxp基础上,利用cre-loxp系统将重组病毒载体中的筛选标记egfp删除,最终获得无egfp的重组病毒载体。本发明利用crispr-cas9技术建立的高效重组痘苗载体,提高重组效率,为以痘苗病毒为载体的疫苗构建和肿瘤免疫治疗节省时间成本。同时通过cre-loxp系统将重组病毒载体中的筛选标记egfp删除,使其提高实际应用的可能性。综上所述,本研究通过crispr-cas9与cre-loxp技术建立起高效重组且无筛选标记的痘苗病毒天坛株载体系统,为其在疫苗载体构建、肿瘤免疫治疗等方面提供参考价值。
附图说明
为了更清楚地说明本申请实施例或现有技术中的技术方案,下面将对实施例中所需要使用的附图作简单地介绍,显而易见地,下面描述中的附图仅仅是本发明中记载的一些实施例,对于本领域普通技术人员来讲,还可以根据这些附图获得其他的附图。
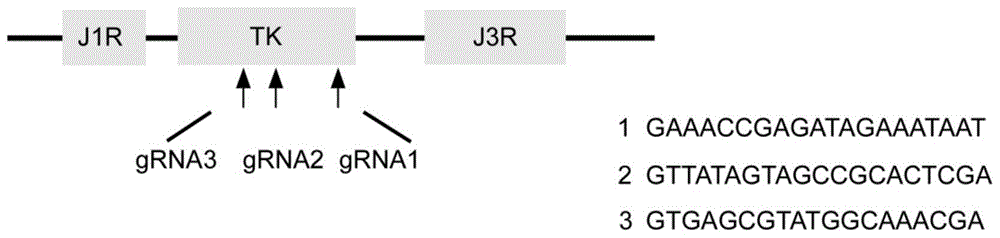
图1为grna设计靶向的位置及序列。
图2为grna载体质粒lenti-delnls及靶向tk区的3个grna质粒lenti-delnls-grna1、lenti-delnls-grna2、lenti-delnls-grna3中的cas9蛋白的表达(n为293t细胞)。
图3为lenti-delnls、lenti-delnls-grna1、lenti-delnls-grna2、lenti-delnls-grna3质粒的cas9定位在细胞质。
图4为重组质粒pj2r-egfp-loxp及同源重组示意图。
图5为在293t细胞中感染野生型痘苗病毒后,重组质粒pj2r-egfp-loxp的荧光表达情况。
图6用荧光显微镜观察重组病毒第一轮筛选纯化的结果。
图7为重组病毒第一轮筛选的重组效率。
图8为经过几轮纯化,获得的重组病毒中绿色荧光蛋白的表达情况。
图9为重组病毒vacv-δtk-egfp-loxp鉴定所需pcr引物示意图。
图10为对重组病毒vacv-δtk-egfp-loxp的pcr鉴定。wt为野生型病毒,a、b、c、d为重组病毒。
图11为在293t细胞中转染pqcxip-cre质粒,cre蛋白表达情况(control为293t细胞)。
图12为转染质粒pqcxip-cre,获得的重组病毒vacv-δtk的第一轮筛选结果。
图13为统计无色噬斑与所有噬斑的比例,计算得到删除效率。
图14为对重组病毒vacv-δtk进行pcr鉴定。wt为野生型病毒,vgfp为重组病毒vacv-δtk-egfp-loxp,a、b、c、d为重组病毒vacv-δtk。
具体实施方式
为了使本领域的技术人员更好地理解本发明的技术方案,下面将结合附图对本发明作进一步的详细介绍。
本发明实施例中使用的痘苗病毒是中国自主研发的天坛株,由该毒株由中国疾病预防控制中心病毒病所应急技术中心提供。天坛株痘苗病毒是曾在中国广泛使用了50年,用于种痘预防天花的疫苗株。由于该毒株已在人体使用超过数亿人次,其免疫效果及副作用均比较清楚。该毒株的全基因组序列资料已存入国际基因库中,编号为genebank,no.af095689。
该病毒在非洲猴肾细胞(vero)中扩增,细胞冻融裂解后,离心,将上清分装后-80℃保存。vero细胞及感染用293t细胞用含10%胎牛血清、100u/ml青霉素、100u/ml链霉素的dmem培养液,在37℃,5%co2培养箱中培养,每2-3天传代一次。
实施例1通过crispr-cas9系统获得重组病毒
1、方法
1.1、grna的设计
针对天坛株痘苗病毒tk区,通过网站(http://crispr.mit.edu/,http://www.e-crisp.org/e-crisp/)设计grna序列,包括grna1、grna2和grna3(如图1所示)。
grna1的序列为:gaaaccgagatagaaataat;
grna2的序列为:gttatagtagccgcactcga;
grna3的序列为:gtgagcgtatggcaaacga。
1.2、grna-cas9质粒中cas9蛋白的表达及定位的检测
1.2.1、蛋白的提取
在293t(5×105)细胞中转染lenti-delnls、lenti-delnls-grna1、lenti-delnls-grna2、lenti-delnls-grna3,48h后收取细胞。3000rpm离心3min,pbs洗一次,加入100μl细胞裂解液ripa,-20℃保存备用。
1.2.2、westernblot检测cas9的表达
(l)首先制备12%的分离胶(4ml30%丙烯酰胺溶液,2.5mltris/hciph8.8,100μl10%sds,100μl10%过硫酸氨,5μltemed),灌胶,液面顶端加入水,室温下聚合60min。
(2)倒干净上层水,再制备5%的积层胶(0.5ml30%丙烯酰胺溶液,0.5mltris/hciph6.8,40μl10%sds,50μl10%过硫酸氨,5μltemed),在积层胶上插入梳齿,室温下聚合50min。待胶完全凝聚后,小心拔下梳齿。
(3)收获的裂解液12000rpm,4℃离心10min。取60μl,加入4×上样缓冲液20μl,98℃煮10min。
(4)制备的电泳胶装进电泳槽,在内槽加满新鲜配制的电泳缓冲液(没过制胶板),将上步处理好的样品取20μl缓缓加入积层胶梳齿孔中,再在电泳槽外槽内加入适量电泳缓冲液,进行电泳:首先是120v,电泳20分钟,观察样品被很好的压缩成一条线,蛋白预染marker开始分离,说明样品进入分离胶,可调整电流为160v,继续电泳40分钟,观察蛋白预染marker很好的分开,结束电泳。
(5)切掉多余胶块,按照从负极到正极分别为滤纸、胶、硝酸纤维素膜(nc膜)、滤纸的顺序叠放在转膜夹板中,放入转膜装置,加满新配的转膜液,冰上或4℃,插上电极进行转膜,80v120min,将胶上的蛋白转移到nc膜上。
(6)转好的膜用5%脱脂奶粉封闭,水平摇床上25rpm,室温孵育lh。
(7)将膜浸在含适当稀释抗体(flag抗体1:1000稀释)的5%脱脂奶粉中,水平摇床上25rpm,4℃过夜。
(8)50rpm水平摇床上,用tbst洗膜4次,5min/次。
(9)弃掉洗液,加入1:10000稀释的alexa
(10)弃掉二抗,25rpm水平摇床上用tbst洗膜4次,5min/次。
(11)取出膜,利用li-corodyssey仪器扫膜。
1.2.3、通过免疫荧光实验检测grna-cas9质粒中cas9蛋白的定位
在hela细胞中转染lenti-delnls、lenti-delnls-grna1、lenti-delnls-grna2、lenti-delnls-grna3,24h后pbs洗2遍,用4%的多聚甲醛固定15min,再用含0.1%triton的多聚甲醛室温打孔10min。pbs洗4次,再用封闭液室温封闭2h。上一抗flag,室温2h或者4℃过夜。0.1%triton的pbs洗4次,上二抗alexa
1.3、重组病毒vacv-δtk-egfp-loxp的获得
1.3.1、重组过程
293t细胞(5×105)接种在六孔板无抗生素dmem完全培养基中,培养过夜,细胞生长平铺60%~70%时,转染grna-cas9质粒。24h后,用dmem稀释野生型病毒,感染细胞;2h后,转染pj2r-egfp-loxp;4h后,换成2%fbs的培养基;48h后,吸弃1.2ml上清,吹取细胞,冻融三次。
1.3.2、通过病毒噬斑形成实验进行纯化过程
vero(1×106)接种在六孔板无抗生素dmem完全培养基中,培养过夜,细胞生长平铺80%~90%时,将收取的病毒悬液10倍梯度稀释重新感染vero细胞。2h后,pbs洗一遍,换成终浓度为1.2%的甲基纤维素培养基。三天后,免疫荧光显微镜下统计重组病毒的重组效率。挑取绿色荧光噬斑重复上述方法进行多轮筛选,得到重组病毒。
1.3.3、重组病毒vacv-δtk-egfp-loxp的鉴定
获得的重组病毒和野生型病毒样品按
引物设计根据ncbi数据库中提供的vtt12病毒株的核苷酸全序列,依据引物设计的基本原则,运用软件primerpremier5.0设计编码病毒蛋白基因序列的特异性引物。
pcr操作步骤
(1)pcr扩增的操作步骤如下:
(2)按照下面表格剂量的添加到0.2ml无核酸的薄壁pcr管中。
(3)轻轻的混匀,通过短暂离心确保全部反应成分在扩增管的管底。
(4)将反应体系置入已经按照上述方法设置的pcr仪器中反应。收集pcr反应产物,-20℃保存。
pcr产物电泳
利用1%琼脂糖凝胶电泳检测病毒pcr产物:
(1)在已加有30ml1×tae电泳缓冲液的三角锥瓶中加入准确称量的琼脂糖粉0.3g。
(2)锥瓶放入微波炉中用中火加热至琼脂糖完全溶解后,室温下冷却至60~70℃左右,加入溴化乙锭(eb)2μl,轻轻摇晃使其充分混匀。
(3)将胶模放入胶槽中固定好,并在距底板0.5-1.0mm的位置放置梳子,以便加入琼脂糖后可以形成完好的加样孔。
(4)将温热的琼脂糖溶液倒入胶模中,凝胶的厚度在3-5mm之间。
(5)在凝胶完全凝固后(于室温放置10-25min或置于4℃冰箱5min),小心移去梳子,将凝胶放入电泳槽中。
(6)加入恰好没过胶面约1mm深的足量电泳缓冲液。
(7)取10μldna样品与2μl的6×loadingbuffer混合后,用微量移液器缓慢将混合物加入加样孔内。
(8)盖上电泳槽并通电,使dna向阳极移动。几分钟后,溴酚蓝从加样孔中迁移到凝胶中。继续电泳直至溴酚蓝在凝胶中迁移出适当的距离。
(9)切断电流,从电泳槽上拔下电线,打开槽盖。取出凝胶。
(10)紫外灯下观察样品的移动情况并拍照。
2、结果
2.1grna-cas9质粒的构建与鉴定
本实验室通过分子克隆方法将lenticrispr-v2中cas9的核定位信号nls缺失,然后将三条靶向tk区的grna序列(如图1所示)导入lenti-delnls,获得带有胞质定位cas9并靶向tk的grna质粒。在293t(5×105)细胞中转染lenti-delnls、lenti-delnls-grna1、lenti-delnls-grna2、lenti-delnls-grna32μg,48h后裂解细胞。通过westernblot检测,结果显示在293t细胞中flag标签的cas9能够成功表达(如图2所示)。在hela(1×105)细胞中转染0.5μglenti-delnls、lenti-delnls-grna1、lenti-delnls-grna2、lenti-delnls-grna3,通过激光共聚焦显微镜观察,显示在转染了lenti-delnls、lenti-delnls-grna1、lenti-delnls-grna2、lenti-delnls-grna3的细胞中,缺失nls的cas9在细胞质中表达(如图3所示)。
2.2重组质粒pj2r-egfp-loxp的构建及鉴定
构建的重组质粒pj2r-egfp-loxp主要包括p7.5、p11、h5启动子、绿色荧光蛋白(enhancegreenfluorescentprotein,egfp)、loxp位点、tk区同源臂(leftarm,rightarm)。重组质粒及同源重组的示意图如图4所示。重组质粒经测序验证后,在293t细胞中感染vtt,然后转染pj2r-egfp-loxp质粒。48h后,免疫荧光观察egfp的表达,证明pj2r-egfp-loxp构建成功,且绿色荧光蛋白成功表达(如图5所示)。
2.3重组病毒vacv-δtk-egfp-loxp的筛选
293t细胞中转染grna-cas9质粒,24h后感染天坛株痘苗病毒vtt,2h后,转染重组质粒pj2r-egfp-loxp,4h后,换成2%fbs的培养基。48h后,收获重组病毒悬液。在vero细胞中利用病毒噬斑形成实验进行第一轮筛选。筛选结果如图6所示,能观察到有绿色荧光蛋白表达的重组病毒噬斑。计算得到的重组效率如图7所示,利用crispr-cas9技术获得重组病毒的效率能达到10%以上,与传统的同源重组构建重组病毒的效率只有0.1%-1%相比,重组的效率大大提高。为得到重组病毒,用相同方法进行多轮筛选,最终获得重组病毒,荧光显微镜下观察到所有噬斑均有绿色荧光蛋白表达(如图8所示)。
2.4重组病毒vacv-δtk-egfp-loxp的鉴定
将重组病毒按
实施例2通过cre-loxp技术删除重组病毒中的筛选标记egfp
1.方法
1.1pqcxip-cre质粒的构建与鉴定
通过分子克隆的方法将cre构建至pqcxip载体中,测序证明序列正确。
1.2、重组病毒vacv-δtk的获得
1.2.1、通过转染pqcxip-cre质粒删除vacv-δtk-egfp-loxp病毒中的egfp
293t细胞(5×105)接种在六孔板无抗生素dmem完全培养基中,培养过夜,细胞生长平铺60%~70%时,转染对照及pqcxip-cre质粒,4~6h后换成10%fbs的培养基。24h后,用dmem稀释重组病毒
vacv-δtk-egfp-loxp,感染细胞;2h后,换成2%fbs的培养基;48h后,吸弃1.2ml上清,吹取细胞,冻融三次。
1.2.2、通过病毒噬斑形成实验进行纯化过程
vero(1×106)接种在六孔板无抗生素dmem完全培养基中,培养过夜,细胞生长平铺80%~90%时,将1.2.1中收取的病毒悬液10倍梯度稀释重新感染vero细胞。2h后,pbs洗一遍,换成终浓度为1.2%的甲基纤维素培养基。三天后,免疫荧光显微镜下统计重组病毒中egfp的删除效率。挑取无色噬斑重复上述方法进行多轮筛选,得到重组病毒vacv-δtk。
1.3、重组病毒vacv-δtk的鉴定
重组病毒基因组提取及鉴定同实验案例一。
2、结果
2.1、pqcxip-cre质粒的构建与鉴定
在293t(5×105)细胞中转染对照及pqcxip-cre,48h后收取细胞。3000rpm离心3min,pbs洗一次,加入100μl细胞裂解液ripa,-20℃保存备用。然后通过westernblot检测蛋白表达(如图11所示)。
2.2、重组病毒vacv-δtk的纯化
在293t细胞中转染质粒pqcxip-cre,4~6h后换成10%fbs的培养基。24h后,用dmem稀释重组病毒vacv-δtk-egfp-loxp,感染细胞;2h后,换成2%fbs的培养基;48h后,收获病毒悬液。在vero细胞中利用病毒噬斑形成实验进行第一轮筛选,转染质粒pqcxip-cre获得的重组病毒出现没有绿色荧光的噬斑(结果如图12所示)。荧光显微镜下能观察到有无色噬斑与所有噬斑的比例,计算得到的删除效率约为50%,如图13所示。为得到重组病毒vacv-δtk,挑选没有绿色荧光的噬斑,用相同方法进行多轮筛选,最终获得没有绿色荧光的重组病毒。
2.3重组病毒vacv-δtk的鉴定
利用实验案例一方法将病毒扩大培养,提取病毒dna,进行pcr验证(如图14所示)。并进行测序验证,证明获得删除tk且不含egfp的重组病毒。
以上只通过说明的方式描述了本发明的某些示范性实施例,毋庸置疑,对于本领域的普通技术人员,在不偏离本发明的精神和范围的情况下,可以用各种不同的方式对所描述的实施例进行修正。因此,上述附图和描述在本质上是说明性的,不应理解为对本发明权利要求保护范围的限制。
- 还没有人留言评论。精彩留言会获得点赞!