一种在水稻花粉中特异表达的启动子PCHF45及其应用的制作方法
本发明涉及基因工程领域,具体地说,涉及水稻花粉特异性启动子pchf45及其应用。
背景技术:
转录调控是植物基因表达调控的主要形式之一,由顺式作用元件和反式作用因子协调完成。启动子是植物基因转录调控中最重要顺式元件之一,一般位于基因5′端上游区,是rna聚合酶及一些反式作用因子的识别与结合位点。启动子主要包含核心启动子区与转录调控区两个功能区。核心启动子区是启动转录的最短启动子片段,一般为40nt,是一段被rna聚合酶家族i、ii和iii识别和结合的dna序列。此区域包含一些重要的功能元件,能够准确的定位转录起点及方向,是基因表达调控的基础。转录调控区域位于核心启动子的上游(或下游),可与特异的转录因子相结合从而对转录的时空和强弱起调控作用,如增强子和沉默子等。深入研究启动子的表达模式既有利于理解基因的表达调控机制和生物学功能,又有助于控制外源基因的表达。
启动子按其表达方式可分为组成型启动子、诱导型启动子和时空特异型启动子等三类。组成型启动子能在所有或大多数组织中启动基因转录,使基因表达具有时空持续性和表达量恒定性。烟草花叶病毒的35s启动子,水稻的actin启动子以及玉米的ubiquitin启动子都属于组成型启动子。组成型启动子广泛应用于植物的基因工程研究中,用于目标基因,如抗虫和抗除草剂基因的超表达。诱导型启动子能在某些物理或化学信号的刺激下启动或大幅提高基因表达。它们具增强子、沉默子或类似功能的序列结构,并有明显的专一性。诱导型启动子根据诱导信号的不同可以分为光诱导启动子、热诱导启动子、低温诱导启动子、干旱诱导启动子、创伤诱导启动子、激素诱导启动子等。时空特异型启动子只在特定的生长阶段或部位中启动基因表达。组织特异性启动子是时空特异型启动子的一种,只在特定细胞、组织或器官中启动表达。在植物的遗传转化中运用组织特异表达的启动子控制目标基因的表达能更有效的避免使用组成型启动子带来的潜在副作用,如能减少组成型表达增加的代谢负担,降低转基因食品安全风险及对与环境的不良影响,以及重复使用相同启动子引发的基因沉默等。目前已开发的水稻组织特异启动子的种类多种多样,在根、茎、叶、种子和果实等几乎各种组织中都发现了有组织特异性表达的启动子。
雄性不育是杂种优势利用的基础,是杂交水稻产业的核心技术,研究水稻育性调控的机理具有重要的理论和实践意义。花药是水稻产生成熟雄配子体(即花粉或小孢子)的器官。在花药发育的早期阶段,存在一种原生造孢细胞,该细胞经过一系列分裂和分化,产生绒毡层和花粉母细胞。绒毡层为雄配子体的发育提供营养,包括各种酶类和非酶类蛋白质,并参与花粉外壁的合成。花粉母细胞则进行减数分裂,产生四分体。四分体中的单细胞经绒毡层分泌胼胝酶的作用分离产生单倍体小孢子体。单倍体小孢子体进一步发育,到晚期时形成中央大液泡。中央大液泡将细胞核排挤到细胞边缘(称为单核靠边期),形成细胞极性。这种细胞极性促进小孢子体进行一次不均等有丝分裂,产生一个大的营养细胞和一个小的生殖细胞。小的生殖细胞被完全包含在大的营养细胞内,形成细胞中细胞的特异现象。生殖细胞接着进行第二次有丝分裂,产生两个精细胞,形成成熟的花粉粒。因此,筛选和确定水稻花粉特异启动子将为水稻的基因工程,特别是在育性调控和转基因漂移防控等方面提供新的选择。
技术实现要素:
本发明的目的是提供一种植物花粉特异性启动子及其应用。
本发明提供的一种植物花粉特异性启动子pchf45,其为具有:
1)seqidno:1所示的核苷酸序列,
或2)在seqidno:1所示的核苷酸序列中经取代、缺失或添加一个或几个核苷酸且具有同等花粉特异性启动功能的由1)衍生的核苷酸序列;
或3)与seqidno:1所示的核苷酸序列互补的序列。
其中,2)所述的由1)衍生的核苷酸序列,与1)所述的核苷酸序列为与1)相比具有70%以上同源性、80%以上同源性、85%以上同源性、90%以上同源性、95%以上同源性、98%以上同源性或99%以上同源性,且具有同等花粉特异性启动子功能的核苷酸序列。
本领域技术人员根据相同目的能够容易地鉴定并利用与植物花粉特异性启动子pchf45核苷酸序列互补的dna分子,因此,具有启动子活性并在严格条件下能与本发明启动子序列或其片段杂交的dna序列包括在本发明中。其中,所述核苷酸序列互补,是指在严格条件下能与pchf45杂交。
严格条件是指探针将与其靶序列杂交至可探测程度超过与其它序列杂交(如至少2倍于背景)的条件。严格条件具有序列依赖性,且因实验其它条件的不同而不同。通过控制杂交和/或洗涤条件的严格性,可以鉴定与探针100%互补的靶序列(同源探测)。可选择地,可以调节严格条件以允许一些序列错配,使得探测到较低程度的相似性(异源探测)。通常,探针长度不超过1000个核苷酸,优选地短于500个核苷酸。
典型地,严格条件是在ph值7.0-8.3下盐浓度低于大约1.5mna离子,典型地大约0.01-1.0mna离子浓度(或其它盐类),温度对短探针(如10-50个核苷酸)至少大约30℃,对长探针(如超过50个核苷酸)至少大约60℃。通过添加去稳定剂如甲酰胺也可获得严格条件。低严格条件,例如,包括在30-35%甲酰胺、1mnacl、l%sds(十二烷基磺酸钠)的缓冲溶液中37℃杂交,在1×至2×ssc(20×ssc=3.0mnacl/0.3m柠檬酸三钠)中50-55℃洗涤。中度严格条件,例如,包括在40-45%甲酰胺、1.0mnacl、l%sds的缓冲溶液中37℃杂交,在0.5×至1×ssc中55-60℃洗涤。高度严格条件,例如,包括在50%甲酰胺、1mnacl、l%sds的缓冲溶液中37℃杂交,在0.1×ssc中60-65℃洗涤。非必要地,洗涤缓冲液可含有大约0.1%-1%的sds。杂交时间一般少于大约24小时,通常大约4-12小时。
特别典型地是杂交后洗涤的函数,关键因素是最终洗涤溶液的离子强度和温度。对于dna-dna杂交体,tm可以从meinkoth和wahl(analbiochem,1984,138:267-284)的方程估算:tm=81.5℃+16.6(logm)+0.41(%gc)-0.61(%form)-500/l;其中m是单价阳离子的摩尔浓度,%gc是鸟嘌呤核苷酸和胞嘧啶核苷酸在dna中的百分比,%form是甲酰胺在杂交溶液中的百分比,l是杂交体在碱基对中的长度。tm是50%互补靶序列与完全配对探针杂交的温度(在规定的离子强度和ph下)。每1%的错配需tm降低大约l℃;因此,tm杂交和/或洗涤条件可被调节以与所需同一性的序列杂交。例如,如果探寻的序列具有≥90%的同一性,tm可以降低10℃。一般地,选择的严格条件是低于特定序列的热解链温度(tm)大约5℃,且其在规定的离子强度和ph下互补。但是,高度严格条件可以应用低于热解链温度(tm)1、2、3或4℃的杂交和/或洗涤;中度严格条件可以应用低于热解链温度(tm)6、7、8、9或10℃的杂交和/或洗涤;低度严格条件可以应用低于热解链温度(tm)11、12、13、14、15或20℃的杂交和/或洗涤。本领域普通技术人员会理解杂交和/或洗涤溶液的条件随严格度的变化而变化,并应用此方程计算杂交和洗涤组合物和所需的tm。如果所需的错配程度使tm低于45℃(水溶液)或32℃(甲酰胺溶液),优选增加ssc浓度以能够使用较高的温度。核酸杂交的指南见于tijssen(1993)生物化学和分子生物学实验室技术一用核酸探针杂交,第i部分,第2章(elsevier,newyork);和ausubel等人编辑(1995)分子生物学现代方法第2章(greenepublishingandwiley-interscience,newyork)。见sambrook等人(1989)分子克隆:实验室手册(第二版,coldspringharborlaboratorypress,plainview,newyork)。
所述严格条件优选为在6×ssc(柠檬酸钠)、0.5%sds(十二烷基磺酸钠)的溶液中,在65℃下杂交,然后用2×ssc、0.1%sds和1×ssc、0.1%sds各洗膜1次。
本发明提供含有本发明所述植物花粉特异性启动子pchf45的基因表达盒、表达载体和含有所述表达载体的宿主细胞。
所述基因表达盒为在花粉特异性启动子pchf45的下游连接有结构基因、调节基因、结构基因的反义基因、调节基因的反义基因或能够干扰内源基因表达的小rna基因的表达盒。
本发明进一步提供含有上述花粉特异性启动子pchf45的基因表达盒的表达载体。
本发明提供了上述水稻花粉特异性启动子pchf45或含有其的表达盒、表达载体或宿主细胞在驱动外源基因在植物花粉中特异性表达的应用。
本发明提供了水稻花粉特异性启动子pchf45或含有其的表达盒、表达载体或宿主细胞在制备转基因植物中的应用。
所述转基因植物为外源基因在花粉中特异表达的转基因植物,优选为授粉、受精能力增强或削弱的转基因植物,更优选为雄性不育转基因植物。
所述植物包括而不限于水稻、玉米、高梁、大麦、燕麦、小麦、粟、甘蔗、大豆、芸苔属物种、棉花、红花、烟草、苜蓿和向日葵。
本发明还提供了扩增所述花粉特异性启动子pchf45的引物对,所述引物对的核苷酸序列为seqidno:2-3或seqidno:6-7。
本发明提供一种分离花粉特异性启动子pchf45的方法,其为使用seqidno:2-3或seqidno:6-7引物对pcr扩增花粉特异性启动子pchf45的核苷酸序列。
本发明还提供驱动外源基因在花粉中特异表达的方法,包括如下步骤:
将本发明的水稻花粉特异性启动子pchf45和目的外源基因克隆到载体中得到含有pchf45和目的外源基因的表达盒的重组表达载体,并将其引入植物基因组,得到外源基因在花粉中特异表达的转基因植物。
本发明提供的花粉特异启动子pchf45具有如下优点:
1)pchf45为水稻内源dna序列,转基因安全风险极低。
2)pchf45能驱动外源基因在花粉中特异性表达,表达水平精确。
3)本发明提供了一种驱动外源基因在花粉中特异性表达的新方法。
附图说明
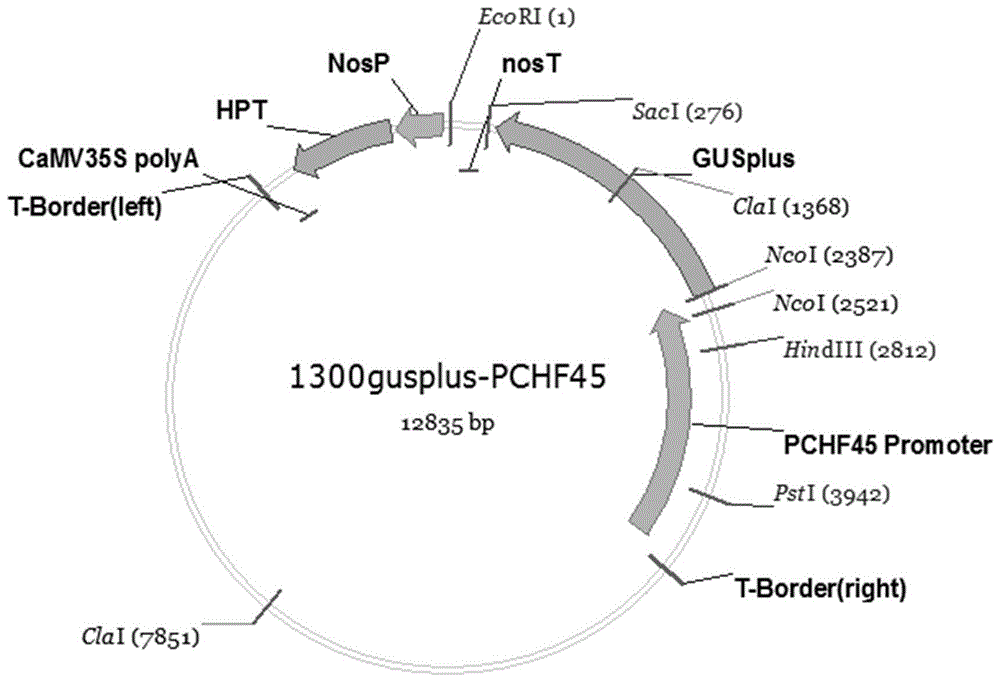
图1是实施例2中花粉特异性启动子pchf45的重组表达载体1300gus-pchf45载体图谱。
图2是实施例3中花粉特异性启动子pchf45的重组表达载体dx2182-pchf45图谱。
图3是非转基因中花11水稻花药与花粉gus染色照片,其中没有花粉被染成蓝色。
图4是实施例6中1300gus-pchf45转基因水稻花药与花粉gus染色照片。
图5是实施例6中1300gus-pchf45转基因水稻雌蕊gus染色照片。
图6是实施例6中1300gus-pchf45转基因水稻颖壳gus染色照片。
图7是实施例6中1300gus-pchf45转基因水稻根gus染色照片。
图8是实施例6中1300gus-pchf45转基因水稻茎gus染色照片。
图9是实施例6中1300gus-pchf45转基因水稻叶gus染色照片。
图10是非转基因中花11水稻花粉在绿色荧光显微镜下照片,其中没有花粉发出荧光。
图11是实施例7中actin启动子转基因水稻花粉在绿色荧光显微镜下照片,其中大多数花粉能发出荧光。
图12是实施例7中dx2182-pchf45转基因水稻花粉在绿色荧光显微镜下的照片,其中有一半花粉能发出荧光。
图13是实施例7中dx2182-pchf45转基因水稻花药在绿色荧光显微镜下的照片。
图14是实施例7中dx2182-pchf45转基因水稻颖壳在绿色荧光显微镜下的照片。
图15是实施例7中dx2182-pchf45转基因水稻叶片在绿色荧光显微镜下的照片。
图16是实施例7中dx2182-pchf45转基因水稻雌蕊在绿色荧光显微镜下的照片。
具体实施方式
以下实施例用于说明本发明,但不用来限制本发明的范围。在不背离本发明精神和实质的情况下,对本发明方法、步骤或条件所作的修改或替换,均属于本发明的范围。
若未特别指明,实施例中所用的技术手段为本领域技术人员所熟知的常规手段。
实施例1水稻花粉特异性启动子pchf45的获取
1.水稻基因组dna的提取
使用植物dna分离试剂盒(成都福际生物技术有限公司)提取水稻基因组dna。基因组来源于水稻品种日本晴的新鲜叶子。提取的基因组dna分装后于-20℃保存备用。
2.pchf45的pcr引物设计与扩增
引物设计使用gibsonassembly方法,扩增产物插入到1300gusplus载体(将gusplus元件插入pc1300多克隆位点获得)的ncoi和hindiii酶切位点和插入到dx2182载体的xmai、sali酶切位点。扩增pchf45的引物序列如seqidno:2-3和seqidno:6-7所示。引物序列seqidno:2-3上下游引物5’端均有15个核苷酸序列(cgacggccagtgcca)与载体相应连接位置重叠,以便gibsonassembly连接。引物序列seqidno:6-7上游引物5’端含有xmai酶切位点序列cccggg,下游引物5’端含有sali酶切位点序列gtcgac。
pcr反应体系(100μl):水稻基因组dnadna模板:3μl(50ng),kod聚合酶(购自东洋坊公司):2μl,10×buffer:10μl,10μm正向引物:3μl,10μm反向引物:3μl,10μmdntp:10μl,mgso4:4μl,1/10dmso:20μl,ddh2o:45μl。
pcr程序:预变性95℃,4min。变性94℃,30s;退火50℃,30s;延伸68℃,2min;35个循环。延伸68℃,10min。
扩增产物包含2051bp的花粉特异性启动子pchf45(序列如seqidno.1)。
实施例2构建启动子pchf45的重组表达载体p1300gusplus-pchf45
将实施例1中获得的pcr产物在1%琼脂糖凝胶中进行电泳,回收2051bp左右大小的条带。用ncoi及hindiii双酶切载体p1300gusplus,回收线性酶切载体。
用lighteningcloningkit(金福塞(北京)生物科技有限公司)对pcr回收产物和线性化p1300gusplus空载体进行连接,10μl体系如下:2.5μl回收产物(50ng/μl),0.5μl酶切载体(100ng/μl),2.5μlligationmix。连接程序:50℃,60min。
取上述连接产物5μl电击转化大肠杆菌感受态细胞。使用引物seqidno:4和seqidno:5进行菌落pcr,挑选阳性克隆测序验证。测序正确的载体命名为p1300gusplus-pchf45,图谱见图1。p1300gusplus载体含有gus基因。表达gus基因的组织经染色后呈现蓝色,可用于指示启动子的表达部位和强度。
实施例3构建含pchf45启动子序列重组表达载体dx2182-pchf45
1.构建含pchf45启动子序列的重组克隆载体pgem-pchf45
参照实施例1的方法,以序列如seqidno:6和seqidno:7所示的引物对替换实施例1中序列seqidno:2和seqidno:3所示的引物对进行扩增,得到的扩增产物进行加a尾(往未纯化产物(50μl)中加入3μldntp,0.5μltaq酶,继续72℃反应20min)。加a尾产物纯化回收后与pgem-t载体进行连接,操作步骤按promega公司产品pgem-t载体说明书进行,得到重组克隆载体pgem-pchf45。转化大肠杆菌感受态细胞,进一步挑选阳性克隆,测序。将测序正确的克隆提取质粒。
2.构建含pchf45启动子序列的重组表达载体dx2182-pchf45
以限制性内切酶xmai、sali双酶切实施例3所得的重组克隆载体pgem-pchf45和表达载体dx2182,将切下的含pchf45启动子的2051bp左右大小片段插入到线性酶切载体dx2182的xmai、sali酶切位点。
用t4连接酶对pcr回收产物和线性化dx2182空载体进行连接,10μl体系如下:4μl回收产物(50ng/μl),4μl酶切载体(100ng/μl),1μlt4连接酶。连接程序:22℃,2小时。
取上述连接产物5μl电击转化大肠杆菌感受态细胞。使用引物seqidno:8和seqidno:9进行菌落pcr,挑选阳性克隆测序验证。测序正确的载体命名为dx2182-pchf45,图谱见图2。dx2182载体含有egfp基因(一种增强型gfp基因)。表达egfp基因的组织在绿色荧光显微镜下发出荧光,可用于指示启动子的表达部位和强度。
实施例4p1300gusplus-pchf45转基因水稻的获得
取-70℃保存的农杆菌eha105于含50μg/ml利福平平板划线,28℃培养。挑取单菌落接种于50mlyeb液体培养基中,220rpm28℃振荡培养12-16hr。取2ml菌液转接于100ml(含有抗生素)yeb液体培养基中,28℃220rpm振荡培养至od600=0.5。冰上预冷10分钟,5000rpm离心10min(冷冻离心机预冷到4℃)。无菌去离子水洗2次(每次10ml),10%甘油洗1次溶于3ml10%甘油中。分别往两管100μl感受态细胞中加1μl实施例2和实施例3中得到的p1300gusplus-pchf45质粒,2.5kv电击转化。在含有卡那霉素和利福平的yeb培养板上28℃培养,挑选阳性克隆,用p1300gusplus载体特异性引物seqidno:4-5进行pcr验证。
验证正确的克隆,通过农杆菌介导的遗传转化法侵染水稻中花11(hieiyohtas,komarit,kumashirot(1994)efficienttransformationofrice(oryzasatival.)mediatedbyagrobacteriumandsequenceanalysisoftheboundariesofthet-dna.theplantjournal6:271-282)。经共培养、筛选、分化、生根等环节获取t0代转基因幼苗。提取转化植株的叶片总dna,使用引物seqidno:10和seqidno:11进行pcr阳性检测,选取经pcr验证的阳性植株栽培,自交结实获得t1代。取t0或t1代植株进行后续分析。
实施例5dx2182-pchf45转基因水稻的获得
取-70℃保存的农杆菌eha105于含50μg/ml利福平平板划线,28℃培养。挑取单菌落接种于50mlyeb液体培养基中,220rpm28℃振荡培养12-16hr。取2ml菌液转接于100ml(含有抗生素)yeb液体培养基中,28℃220rpm振荡培养至od600=0.5。冰上预冷10分钟,5000rpm离心10min(冷冻离心机预冷到4℃)。无菌去离子水洗2次(每次10ml),10%甘油洗1次溶于3ml10%甘油中。往100μl感受态细胞中加1μl实施例3中得到的dx2182-pchf45质粒,2.5kv电击转化。在含有卡那霉素和利福平的yeb培养板上28℃培养,挑选阳性克隆,用dx2182载体特异性引物seqidno:8-9进行pcr验证。
验证正确的克隆,通过农杆菌介导的遗传转化法侵染水稻中花11(hieiyohtas,komarit,kumashirot(1994)efficienttransformationofrice(oryzasatival.)mediatedbyagrobacteriumandsequenceanalysisoftheboundariesofthet-dna.theplantjournal6:271-282)。经共培养、筛选、分化、生根等环节获取t0代转基因幼苗。提取转化植株的叶片总dna,使用引物seqidno:10和seqidno:11进行pcr阳性检测,选取经pcr验证的阳性植株栽培,自交结实获得t1代。取t0或t1代植株进行后续分析。
实施例6转基因水稻gus染色分析
配制gus染色液x-gluc反应液(50mm磷酸纳缓冲液,ph值7.0,0.5mm铁氰化钾,0.5mm亚铁氰化钾,0.5mg/mlx-gluc,体积百分含量20%甲醇,0.1%tritonx-100),随机选取5个以上在实施例4中获得的1300gus-pchf45转基因阳性株系采集花药、雌蕊、颖壳、根、叶、茎等组织样品,浸泡在x-gluc反应液中37℃2h或过夜,然后用体积百分含量75%乙醇脱去组织的叶绿体颜色后观察照相。结果显示,野生型中花11的花粉不能染色(图3),而转基因水稻的花粉有一半被染成蓝色(图4),其它雌蕊(图5)、颖壳(图6)、根(图7)、茎(图8)、叶(图9)等组织均不能被染色,表明pchf45启动子可驱动gus基因在水稻花粉中特异性表达,而在水稻花药、颖壳、根、茎、叶片、雌蕊等组织均不能驱动外源基因的表达。
实施例7转基因水稻花粉egfp绿色荧光分析
随机选取5个以上在实施例5中获得的dx2182转基因阳性株系采集成熟期花药及花粉制片,置于绿色荧光显微镜下分析结果。结果显示,野生型中花11的花粉没有发荧光(图10),而阳性对照actin启动子(也构建在dx2182载体egfp报告基因的上游)的转基因花粉能发荧光(图11)。参照阳性对照结果,pchf45启动子的转基因花粉能发出明显荧光(图12),其他花药(图13)、颖壳(图14)、叶片(图15)、雌蕊(图16)等组织均不能发荧光,表明本发明的pchf45启动子可驱动egfp基因在水稻花粉中特异性表达,而在水稻花药、颖壳、叶片、雌蕊等组织均不能驱动外源基因的表达。
虽然,上文中已经用一般性说明及具体实施方案对本发明作了详尽的描述,但在本发明基础上,可以对之作一些修改或改进,这对本领域技术人员而言是显而易见的。因此,在不偏离本发明精神的基础上所做的这些修改或改进,均属于本发明要求保护的范围。
序列表
<110>海南波莲水稻基因科技有限公司
<120>一种在水稻花粉中特异表达的启动子pchf45及其应用
<130>khp191111511.1
<160>11
<170>siposequencelisting1.0
<210>1
<211>2051
<212>dna
<213>人工序列(artificialsequence)
<400>1
cctagctctatgtacttcccataagcctattattgattgttcacagtttcctcggtgatg60
aatttgcaaatggattagattgtgtgatacttaactaataacctgacaaatacagcatgc120
acctatgcaaaaataacaccaattcaaaaccttcttcctctccaaaaaaaaattgtaaaa180
aaaatcactcataaaaatattaaggccaaatttggcatatcataattcattcgatcattc240
atgagctaatgacacctcaataaaggtgaaaacagcaatacggaggtgccttgaaaactg300
cttcgatttacctccaaattaagccagttacagactctgaagtgttgtggatttgcacta360
gaacaagtttggcaataagaaacaccaattctgaagaacatggtgatttagacatggtat420
tgtataatcaaaatgggacttccattactaattggtgttttggcccaaaccaccaacgaa480
atgtaacttacactgcagctgtatgctcgaaaaatttaggccattcaggtaaaatttagg540
agatttcaagccacatacatggtaactggattctgattgatgatgaacatgtttgagcag600
caccactaaaactttcggtttaggtcaaaatttcgatgattttggtggtcactgaaaatt660
cacaaatttcagtttttttgaacctttttgaacaaaattatttcaaattttgactaaatt720
tgaataaatttgatcaaattcacaaataatacttgggaaaatctgaaaagtttgggcaag780
gtagttattactaaaatttccgaaattttggaaatttcgaactgaaattttaatccctac840
ctataagattgtgacatcgcacagattcaggatggaaaaacataacaacaacccttgaga900
tcaaacttggtattttctttgtttcatattataagacgttttggattttgaatagattca960
tgcatggatctacgtattatgattcatatatgtgtccaaattgatatggatattagtgaa1020
tctaggtacgggataaagaggctaacttcttataatatggaacggagagactaccttgta1080
agcattattatcttaataaaggcagggtggtcgagaaggcttgtttttctattcctagtt1140
tctatcactcctacatcgtaagttgtcaaagacgatagatttgagaacatgtgttgtgtt1200
gtgtttgagagagcacgaggaggcgtcatctaaatttatacagtgttagttgtactgttt1260
tctataggatttgaaaccggtggatggaaaatcataatgctctacctagctctatgtgct1320
gtccatctgccttttattaattagcatttcaaagttttctcaatgatgcatttgcaaaat1380
ggattagattatgtgaaatttaactaacttggcaaatacagcattcatccaagcattcat1440
gcttaaacaacaatacaataccaattcacaaccttcttcctcctctccaaaattggaaaa1500
taaattcactaaaaaaaaactactaagcccaaaattaccatctcatgactcattccttca1560
atcatcatgagttaatgacacctcaaacaaacaaaaaaaaattctaaaaaacttcctcaa1620
gctttatgagtgaaaaactttctaatacaactacagtataattatgccgtagttccattg1680
catgtaactaaacaaaaagcgcatcttgacggccagaaaatcgaaagtttctccaacaaa1740
aatccatcgccagtttttttttagcaaaaaccgaaatatcaataaacctaaacagaaaac1800
tgcatttctctcaaccatgtccaaaacgtgggtcccatgcatcttctccaactcgattcc1860
cgccaaaaatagcgcgccaaaccaccggaacacgcgcgccgccgcacggccatggccacc1920
gccgctgccctatatctcccctctctcaccaccctccatataggcaggcaggcaccgtcg1980
ccggtggtggcccacctcgtgcctcctcccccaccccccggccctctcctcccccacctc2040
cagccatggag2051
<210>2
<211>38
<212>dna
<213>人工序列(artificialsequence)
<400>2
cgacggccagtgccacctagctctatgtacttcccata38
<210>3
<211>42
<212>dna
<213>人工序列(artificialsequence)
<400>3
gaaatttaccctcagatctaccatctccatggctggaggtgg42
<210>4
<211>27
<212>dna
<213>人工序列(artificialsequence)
<400>4
gatcagtttaaagaaagatcaaagctc27
<210>5
<211>25
<212>dna
<213>人工序列(artificialsequence)
<400>5
ctgcaaggcgattaagttgggtaac25
<210>6
<211>33
<212>dna
<213>人工序列(artificialsequence)
<400>6
cccgggcctagctctatgtacttcccataagcc33
<210>7
<211>26
<212>dna
<213>人工序列(artificialsequence)
<400>7
gtcgacggtgcctgcctgcctatatg26
<210>8
<211>21
<212>dna
<213>人工序列(artificialsequence)
<400>8
aactgcctggcacagcaattg21
<210>9
<211>25
<212>dna
<213>人工序列(artificialsequence)
<400>9
gacgttgtggctgttgtagttgtac25
<210>10
<211>21
<212>dna
<213>人工序列(artificialsequence)
<400>10
cttagccagacgagcgggttc21
<210>11
<211>19
<212>dna
<213>人工序列(artificialsequence)
<400>11
gcttctgcgggcgatttgt19
- 还没有人留言评论。精彩留言会获得点赞!