FHY3/FAR1蛋白在调控植物生长和防御平衡中的应用的制作方法
本发明涉及fhy3/far1蛋白的新应用,尤其涉及拟南芥fhy3/far1蛋白在植物面临避荫反应时调控生长和防御平衡中的应用,属于fhy3/far1蛋白的新应用领域。
背景技术:
在模式植物拟南芥中,far-redelongatedhypocotyls3(fhy3)和far-redimpairedresponse1(far1)是光敏色素a信号转导途径的两个关键正向调控因子,其功能缺失导致植物对远红光的反应大大下降(wanganddeng,2002)。fhy3和far1编码两个同源的从古老的mutator-like转座酶衍变而来的新型转录因子,其蛋白氮末端c2h2型锌指结构域具有dna结合活性,能特异地结合下游靶基因启动子的结合位点(fhy3/far1bindingsite,fbs,5′-cacgcgc-3′)上,调节下游基因表达,比如激活fhy1和fhl的表达,后者能与phya相互作用,并介导phya从细胞质到细胞核的转运过程和随后的光应答反应(linetal.,2007)。最近的研究表明,fhy3和far1蛋白可以直接与dna直接结合从而调节众多下游基因的表达,从而调控植物的生长发育(ouyangetal.,2011;wangandwang,2015)。
迄今为止,尚未发现fhy3/far1蛋白应用于调控植物生长和防御平衡等方面。
技术实现要素:
本发明的主要目的是提供fhy3/far1蛋白的一种新的用途;
本发明的上述目的是通过以下技术方案来实现的:
本发明提供fhy3/far1蛋白在植物面临避荫反应时调控植物生长以及防御平衡中的应用。
本发明首先提供fhy3/far1蛋白在抑制植物生长或加强植物防御反应上的应用。
本发明通过试验发现在面临避荫反应时,fhy3/far1蛋白通过直接激活避荫反应的抑制子par1和par2基因的转录来抑制植物生长加速。fhy3/far1通过与jaz蛋白和myc2蛋白相互作用促进茉莉酸介导的防御反应。
因此,本发明提供了一种抑制植物生长或加强植物防御反应的方法,包括:将fhy3/far1蛋白编码基因在植物中进行过表达得到转基因植物;所得到的转基因植物在面临避荫反应时过快的生长速度被抑制,同时对于病原菌的抵抗能力增强;譬如:将fhy3/far1蛋白编码基因可操作的与表达调控元件相连接,得到在植物中表达该编码基因的重组植物表达载体;将所述重组植物表达载体转化目标植物,使fhy3/far1蛋白编码基因在植物中进行过表达。
本发明通过进一步的试验发现,突变fhy3/far1蛋白的植株,一方面直接调控的靶基因par1和par2表达水平下降,导致生长速度加快,避荫反应增强,下胚轴伸长,避荫反应下游基因表达水平升高;另一方面,由于突变导致与myc2蛋白的相互作用丧失,对坏死菌(botrytiscinerea)的抵抗能力下降,下游防御相关的基因表达水平降低,使防御能力减弱;因此,在植物中突变fhy3/far1蛋白的编码基因,可以增强植物的避荫反应,抑制植物的防御反应。
因此,本发明另一方面提供了一种增强植物的避荫反应或抑制植物的防御反应的方法,包括:将植物中的fhy3/far1蛋白的编码基因进行突变,得到fhy3/far1功能缺陷的突变体,该突变体在面临避荫反应时生长速度会加块,对于茉莉酸介导的防御反应减弱。
本领域人员可以采用各种常规的方法对植物中的fhy3/far1蛋白进行突变使fhy3/far1蛋白的功能缺失或出现缺陷。
本发明进一步公开了含有所述fhy3/far1蛋白编码基因的重组表达载体以及含有所述重组表达载体的重组宿主细胞。将所述fhy3/far1蛋白编码基因可操作的与表达调控元件相连接得到重组植物表达载体;该重组植物表达载体可以由5′端非编码区,fhy3/far1蛋白编码基因和3′非编码区组成;其中,所述的5′端非编码区可以包括启动子序列、增强子序列或/和翻译增强序列;所述的启动子可以是组成性启动子、诱导型启动子、组织或器官特异性启动子;所述的3′非编码区可以包含终止子序列、mrna切割序列等。合适的终止子序列可取自根癌农杆菌的ti-质粒,例如章鱼碱合成酶和胭脂碱合成酶终止区。
所述重组植物表达载体还可含有用于选择转化细胞的选择性标记基因,用于选择经转化的细胞或组织。所述标记基因包括:编码抗生素抗性的基因以及赋予除草化合物抗性的基因等。此外,所述的标记基因还包括表型标记,例如β-半乳糖苷酶和荧光蛋白等。
所述的目标植物包括但不限于:单子叶植物或双子叶植物。更优选的,所述的目标植物包括农作物、蔬菜或观赏植物、果树等,例如,可以是玉米、水稻、高粱、小麦、大豆、马铃薯、大麦、番茄、菜豆、花生或甘蔗等。
转化方案以及将所述多核苷酸或多肽引入植物的方案可视用于转化的植物(单子叶植物或双子叶植物)或植物细胞的类型而变化。将所述多核苷酸引入植物细胞的合适方法包括:显微注射、电穿孔、农杆菌介导的转化、直接基因转移以及高速弹道轰击等。在特定的实施方案中,可利用多种瞬时转化法将fhy3/far1蛋白编码基因提供给植物。利用常规方法可使已转化的细胞再生稳定转化植株(mccormicketal.plantcellreports.1986.5:81-84)。
本发明中所述的拟南芥fhy3/far1蛋白的氨基酸序列为seqidno.1所示,其编码基因的核苷酸序列为seqidno.2所示。另外,本领域技术人员还可以将seqidno.2所示的核苷酸进行优化以增强在拟南芥中的表达效率。
本发明整体技术方案的详述
本发明首先构建了拟南芥fhy3far1突变体并观察了突变体在避荫反应的表型,试验结果发现,突变体比野生型的下胚轴更长,这说明突变体的避荫反应表型增强,生长加快。同时观察了fhy3far1突变体的下胚轴的细胞变化,观察到突变体的细胞长度明显变长,进一步检测突变体中的基因表达变化发现,一些避荫反应诱导的基因在fhy3和fhy3far1突变体中明显升高。
本发明进一步探讨了fhy3和far1影响避荫反应导致的生长变化的机制试验:将正常生长条件下的fhy3和fhy3far1突变体转到低r/fr条件下(30-50μem-2s-1,r/frratio=8),分别0h、1h、2h取材,采用定量pcr技术手段检测par1和par2基因的表达水平。试验结果显示par1和par2基因在突变体中的表达模式显著降低。进一步采用chip-pcr(染色质免疫共沉淀)和emsa(凝胶阻滞)实验验证了fhy3直接结合par1基因的启动子,调控其转录。同时,par1基因的过量表达可以回复fhy3far1突变体的避荫反应增强的表型,即fhy3far1par1-gfp的下胚轴长度变短。试验结果表明,fhy3和far1通过调控par1和par2基因表达从而影响避荫反应导致的生长变化。
本发明采用酵母双杂交实验和萤光素酶互补实验发现,fhy3和myc2蛋白能够相互作用。
本发明将生长三周的fhy3far1的突变体拟南芥叶片进行botrytiscinerea接菌,分别置于正常光照和lowr/fr条件下培养3天,结果发现,fhy3far1突变体的botrytiscinerea菌斑面积显著增大,表明fhy3和far1在正常光照和lowr/fr条件下都可以抑制botrytiscinerea菌的侵染。进一步检测了ja介导的防御基因的表达水平变化,结果发现lox2、pdf1.2、tat1、vsp2基因的表达水平在fhy3far1突变体中都显著的降低。
本发明所涉及到的术语定义
除非另外定义,否则本文所用的所有技术及科学术语都具有与本发明所属领域的普通技术人员通常所了解相同的含义。
术语“多核苷酸”或“核苷酸”意指单股或双股形式的脱氧核糖核苷酸、脱氧核糖核苷、核糖核苷或核糖核苷酸及其聚合物。除非特定限制,否则所述术语涵盖含有天然核苷酸的已知类似物的核酸,所述类似物具有类似于参考核酸的结合特性并以类似于天然产生的核苷酸的方式进行代谢。除非另外特定限制,否则所述术语也意指寡核苷酸类似物,其包括pna(肽核酸)、在反义技术中所用的dna类似物(硫代磷酸酯、磷酰胺酸酯等)。除非另外指定,否则特定核酸序列也隐含地涵盖其保守修饰的变异体(包括(但不限于)简并密码子取代)和互补序列以及明确指定的序列。特定而言,可通过产生其中一个或一个以上所选(或所有)密码子的第3位经混合碱基和/或脱氧肌苷残基取代的序列来实现简并密码子取代(batzer等人,nucleicacidres.19:5081(1991);ohtsuka等人,j.biol.chem.260:2605-2608(1985);和cassol等人,(1992);rossolini等人,molcell.probes8:91-98(1994))。
术语“多肽”、“肽”和“蛋白”在本文中互换使用以意指氨基酸残基的聚合物。即,针对多肽的描述同样适用于描述肽和描述蛋白,且反之亦然。所述术语适用于天然产生氨基酸聚合物以及其中一个或一个以上氨基酸残基为非天然编码氨基酸的氨基酸聚合物。如本文中所使用,所述术语涵盖任何长度的氨基酸链,其包括全长蛋白(即抗原),其中氨基酸残基经由共价肽键连接。
术语“重组宿主细胞株”或“宿主细胞”意指包含本发明多核苷酸的细胞,而不管使用何种方法进行插入以产生重组宿主细胞,例如直接摄取、转导、f配对或所属领域中已知的其它方法。外源性多核苷酸可保持为例如质粒的非整合载体或者可整合入宿主基因组中。宿主细胞可为原核细胞或真核细胞,宿主细胞还可为单子叶或双子叶植物细胞。
术语“可操作的连接”指两个或更多个元件之间功能性的连接,可操作的连接的元件可为邻接或非邻接的。
术语“重组植物表达载体”意指一种或多种用于实现植物转化的dna载体;本领域中这些载体常被称为二元载体。二元载体连同具有辅助质粒的载体是大多常用于土壤杆菌介导转化的。二元载体通常包括:t-dna转移所需要的顺式作用序列、经工程化处理以便能够在植物细胞中表达的选择标记物,待转录的异源性dna序列等。
术语“转化”指将异源性dna序列引入到宿主细胞或有机体的方法。
术语“表达”指内源性基因或转基因在植物细胞中的转录和/或翻译。
附图说明
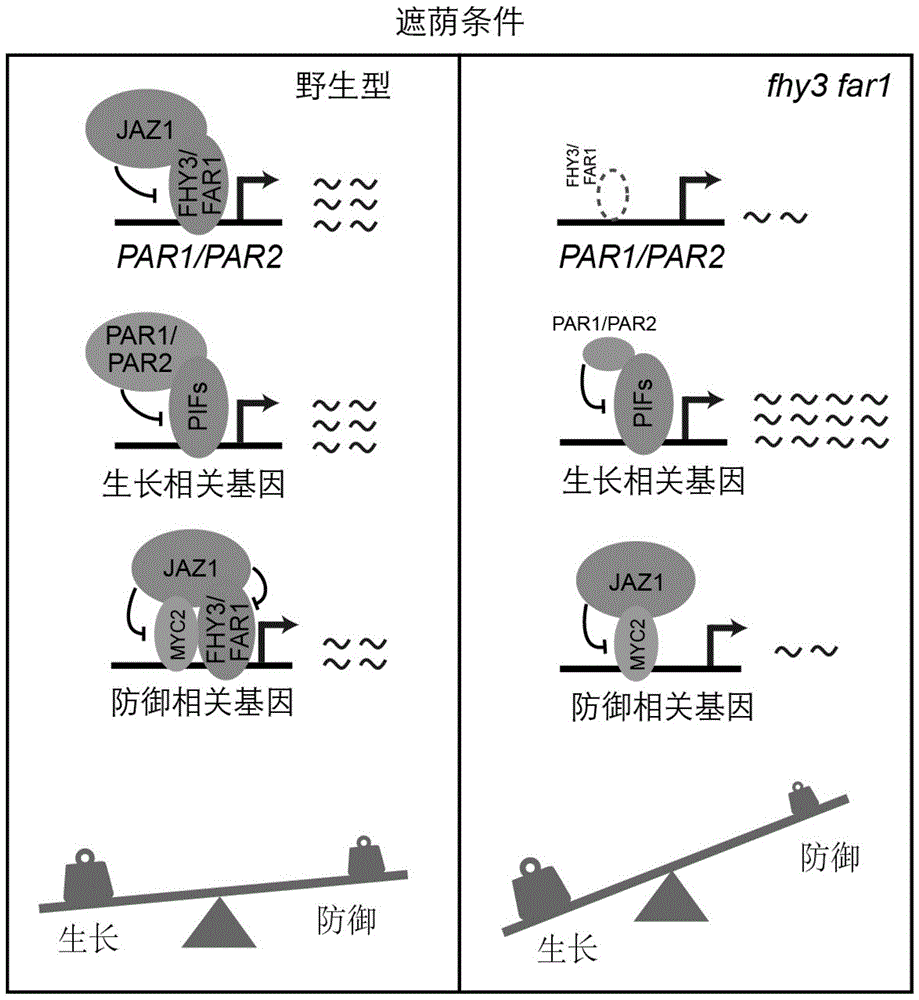
图1fhy3/far1调制植物避荫反应条件下的生长防御平衡模式图;植物面临遮荫条件时,jaz1蛋白蛋白含量升高。一方面jaz1抑制fhy3/far1蛋白的转录活性,使par1/2维持在稳定的水平,继而抑制pifs蛋白的活性和生长相关基因的表达水平。另一方面,jaz1抑制fhy3/far1和myc2蛋白对防御基因的转录激活能力,从而使防御能力减弱,最终生长和防御维持一定的平衡。在fhy3far1缺失功能的突变体中,由于fhy3/far1的缺失,par1/2转录水平显著降低,使pifs下游的生长相关基因表达水平显著上升。此外,缺失的fhy3/far1也导致防御基因显著下降,最终使避荫反应条件下的生长和防御平衡被打破,即植物加速生长,防御能力明显下降。
图2为fhy3和far1影响植物避荫反应导致的生长加快。从图a和图b中可以看出,缺失功能的fhy3和fhy3far1突变体的下胚轴伸长变长。从图c和图d中可以看出,这种下胚轴生长变化的情况主要原因是细胞伸长引起的;从图e中可以看出,避荫反应下游基因表达在突变体中升高,这表明fhy3和far1在正常情况下抑制这些基因的表达。
图3为fhy3和far1通过直接调控par1和par2影响避荫反应。其中图a为fhy3和far1影响par1和par2的表达水平;图b、图c和图d为fhy3蛋白直接结合par1和par2的启动子;图e和图f为fhy3和far1调控par1表达从而影响避荫反应的遗传证据。
图4为fhy3与myc2蛋白相互作用,共同调控myc2靶基因的转录。图a和图b显示fhy3可以与myc2蛋白相互作用。图c和图d显示fhy3和myc2共同调控lox2基因的转录。图d显示lox2基因表达水平在fhy3far1myc2突变体中表达水平显著降低。
图5为fhy3和far1参与ja介导的防御反应。其中图a和图b为缺失功能的fhy3far1突变体对坏死菌(botrytiscinerea)的抗性明显减弱。图c为ja相关的防御反应基因表达水平在fhy3far1突变体中明显下降。
具体实施方式
以下结合具体实施例来进一步描述本发明,本发明的优点和特点将会随着描述而更为清楚。但这些实施例仅是范例性的,并不对本发明的范围构成任何限制。本领域技术人员应该理解的是,在不偏离本发明的精神和范围下可以对本发明的细节和形式进行修改或替换,但这些修改和替换均落入本发明的保护范围内。
试验例1拟南芥fhy3far1突变体的构建及避荫反应表型观察试验
fhy3突变体(salk_002711)和far1突变体(salk_031652)购买于美国abrc突变体库(https://abrc.osu.edu/),fhy3far1双突变体由fhy3和far1单突变体杂交获得。
将fhy3突变体(salk_002711)和fhy3far1突变体(salk_002711,salk_031652)长在持续白光条件下3天,随后置于低r/fr条件下(30-50μem-2s-1,r/frratio=8)继续生长3天,观察幼苗生长情况。
图2a和2b显示突变体比野生型(col)的下胚轴更长,这说明突变体的避荫反应表型增强,生长加快。同时观察了fhy3far1突变体的下胚轴的细胞变化,采用pi(propidiumiodide)染色后,可以看到细胞形态的变化,观察到突变体的细胞长度明显变长,进一步检测突变体中的基因表达变化发现,一些避荫反应诱导的基因在fhy3和fhy3far1突变体中明显升高。
试验例2fhy3和far1影响避荫反应导致生长变化的机制试验
将正常生长条件下的fhy3和fhy3far1突变体转到低r/fr条件下(30-50μem-2s-1,r/frratio=8),分别0h、1h、2h取材,采用定量pcr技术手段检测par1和par2基因的表达水平。
图3a显示par1和par2基因在突变体中的表达模式显著降低。进一步,采用chip-pcr(染色质免疫共沉淀)和emsa(凝胶阻滞)实验验证了fhy3直接结合par1基因的启动子,调控其转录(图3b-3d)。同时,par1基因的过量表达(35s::par1-gfp)可以回复fhy3far1突变体的避荫反应增强的表型,即fhy3far1par1-gfp的下胚轴长度变短(图3e)。上述试验结果表明,fhy3和far1通过调控par1和par2基因表达从而影响避荫反应导致的生长变化。
试验例3fhy3与myc2蛋白相互作用试验
采用酵母双杂交实验,将质粒pb42ad-myc2和lexa-fhy3共同转进egy48酵母株系中,在sd-uth营养缺陷性平板上生长,3天后,将生长状况良好的酵母移到sd-uth+x-gal平板上显色。
试验结果如图4a显示,fhy3和myc2蛋白可以相互作用。
采用萤光素酶互补实验(lci)在烟草叶片中检测fhy3和myc2蛋白的互作:分别构建cluc-fhy3和myc2-nluc载体,并转化农杆菌,然后共同转化烟草叶片。
试验结果如图4b显示,fhy3和myc2蛋白可以相互作用。图4c和4d显示,增加fhy3蛋白可以促进myc2对靶基因lox2的转录激活。进一步采用定量pcr技术检测了fhy3far1myc2突变体中的lox2基因表达水平。试验结果如图4e显示,lox2水平在三突变体中显著的降低。
试验例4fhy3far1突变体对病菌的抵抗能力试验
对生长三周的拟南芥叶片进行botrytiscinerea接菌,分别置于正常光照和lowr/fr条件下培养3天,结果如图5a和5b所示,fhy3far1突变体的botrytiscinerea菌斑面积显著增大,表明fhy3和far1在正常光照和lowr/fr条件下都可以抑制botrytiscinerea菌的侵染。进一步,本试验检测了ja介导的防御基因的表达水平变化。如图5c-5e所示,lox2、pdf1.2、tat1、vsp2基因的表达水平在fhy3far1突变体中都显著的降低。
sequencelisting
<110>中国农业科学院生物技术研究所、华南农业大学
<120>fhy3/far1蛋白在调控植物生长和防御平衡中的应用
<130>bj-2002-190320a
<160>2
<170>patentinversion3.5
<210>1
<211>839
<212>prt
<213>arabidopsisthaliana
<400>1
metaspileaspleuargleuhisserglyaspleucyslysglyasp
151015
aspgluaspargglyleuaspasnvalleuhisasnglugluaspmet
202530
aspileglylysilegluaspvalservalgluvalasnthraspasp
354045
servalglymetglyvalprothrglygluleuvalglutyrthrglu
505560
glymetasnleugluproleuasnglymetgluphegluserhisgly
65707580
glualatyrserphetyrglnglutyrserargalametglypheasn
859095
thralaileglnasnserargargserlysthrthrargglupheile
100105110
aspalalysphealacysserargtyrglythrlysargglutyrasp
115120125
lysserpheasnargproargalaargglnserlysglnaspproglu
130135140
asnmetalaglyargargthrcysalalysthraspcyslysalaser
145150155160
methisvallysargargproaspglylystrpvalilehisserphe
165170175
valarggluhisasnhisgluleuleuproalaglnalavalserglu
180185190
glnthrarglysiletyralaalametalalysglnphealaglutyr
195200205
lysthrvalileserleulysseraspserlysserserpheglulys
210215220
glyargthrleuservalgluthrglyaspphelysileleuleuasp
225230235240
pheleuserargmetglnserleuasnserasnphephetyralaval
245250255
aspleuglyaspaspglnargvallysasnvalphetrpvalaspala
260265270
lysserarghisasntyrglyserphecysaspvalvalserleuasp
275280285
thrthrtyrvalargasnlystyrlysmetproleualailepheval
290295300
glyvalasnglnhistyrglntyrmetvalleuglycysalaleuile
305310315320
seraspgluseralaalathrtyrsertrpleumetgluthrtrpleu
325330335
argalaileglyglyglnalaprolysvalleuilethrgluleuasp
340345350
valvalmetasnserilevalprogluilepheproasnthrarghis
355360365
cysleupheleutrphisvalleumetlysvalsergluasnleugly
370375380
glnvalvallysglnhisaspasnphemetprolyspheglulyscys
385390395400
iletyrlysserglylysaspgluaspphealaarglystrptyrlys
405410415
asnleualaargpheglyleulysaspaspglntrpmetileserleu
420425430
tyrgluasparglyslystrpalaprothrtyrmetthraspvalleu
435440445
leualaglymetserthrserglnargalaaspserileasnalaphe
450455460
pheasplystyrmethislyslysthrservalglngluphevallys
465470475480
valtyraspthrvalleuglnaspargcysglugluglualalysala
485490495
aspserglumettrpasnlysglnproalametlysserproserpro
500505510
pheglulysservalsergluvaltyrthrproalavalphelyslys
515520525
pheglnilegluvalleuglyalailealacysserproarggluglu
530535540
asnargaspalathrcysserthrpheargvalglnaspphegluasn
545550555560
asnglnaspphemetvalthrtrpasnglnthrlysalagluvalser
565570575
cysilecysargleupheglutyrlysglytyrleucysarghisthr
580585590
leuasnvalleuglncyscyshisleuserserileproserglntyr
595600605
ileleulysargtrpthrlysaspalalysserarghisphesergly
610615620
gluproglnglnleuglnthrargleuleuargtyrasnaspleucys
625630635640
gluargalaleulysleuasngluglualaserleuserglngluser
645650655
tyrasnilealapheleualailegluglyalaileglyasncysala
660665670
glyileasnthrserglyargserleuproaspvalvalthrserpro
675680685
thrglnglyleuileservalglugluaspasnhisserargserala
690695700
glylysthrserlyslyslysasnprothrlyslysarglysvalasn
705710715720
progluglnaspvalmetprovalalaalaprogluserleuglngln
725730735
metasplysleuserproargthrvalglyileglusertyrtyrgly
740745750
thrglnglnservalglnglymetvalglnleuasnleumetglypro
755760765
thrargaspasnphetyrglyasnglnglnthrmetglnglyleuarg
770775780
glnleuasnserilealaprosertyraspsertyrtyrglyprogln
785790795800
glnglyilehisglyglnglyvalaspphepheargproalaasnphe
805810815
sertyraspileargaspaspproasnvalargthrthrglnleuhis
820825830
gluaspalaserarghisser
835
<210>2
<211>2520
<212>dna
<213>arabidopsisthaliana
<400>2
atggatatagatcttcgactacattcaggtgacctttgcaaaggagatgatgaggataga60
ggacttgataatgtgttacacaatgaagaagatatggatattggcaaaattgaggatgtt120
tctgttgaggttaatactgatgacagtgtaggcatgggtgttccaactggggagttggtt180
gagtacactgagggtatgaatcttgaacctcttaatggtatggagtttgagtcacacggt240
gaagcgtattcgttttaccaggagtattctcgagctatggggtttaacactgcgatacag300
aacagtcgtagatcgaaaactacaagagagtttattgatgcgaagtttgcttgctctagg360
tatggtactaaaagagaatatgacaaatcttttaatcggccacgagctagacaaagtaag420
caagatccagagaatatggctggtaggagaacatgtgcgaaaaccgattgcaaagctagt480
atgcatgttaagagaaggccggatgggaaatgggtaattcacagctttgtgagggagcat540
aatcatgagcttttacctgcacaagctgtgagtgaacagaccagaaagatatatgctgca600
atggctaagcagtttgctgagtacaaaactgttataagtctaaaaagcgattcgaaaagt660
tcttttgaaaaaggtcggactttatcagtggaaacaggggatttcaaaatcctgcttgac720
ttcttgtcacggatgcaatccttgaattccaacttcttttatgcagtagatctgggcgat780
gatcagcgagtgaagaatgtattttgggtagatgcaaagagcaggcataactatggcagt840
ttttgtgatgttgtttctctagacaccacctacgtaagaaacaagtacaagatgccactt900
gctatttttgttggagtgaaccagcactaccagtatatggtccttgggtgcgccttaata960
tccgatgagagtgcagcaacatattcttggttaatggagacctggctgagagcaatcggt1020
ggacaagctccaaaggttttgatcactgagctagacgttgtaatgaactccattgttccc1080
gaaatcttccccaatacacggcattgcctttttctttggcatgtattgatgaaggtttct1140
gagaatcttggtcaagttgttaagcaacatgataattttatgcctaagtttgagaagtgc1200
atctataaatcagggaaagatgaagactttgcgagaaaatggtataagaatcttgctaga1260
ttcggcctcaaagatgatcaatggatgatttctttatatgaagaccgcaagaaatgggca1320
ccaacctacatgactgatgttcttttggctgggatgtctacatctcaacgagctgatagt1380
attaatgctttcttcgacaagtatatgcataagaagactagtgtacaagaatttgtaaag1440
gtatatgatacagttttgcaagatagatgtgaagaagaagccaaagctgattctgaaatg1500
tggaacaagcagccagcaatgaagtctccatcgccatttgagaagagtgtttctgaagtc1560
tatactcctgcagttttcaaaaagttccagattgaagttttgggtgcgattgcgtgtagt1620
cctagggaggagaaccgagatgccacatgttctacttttagagtgcaggattttgaaaac1680
aaccaagacttcatggtaacatggaaccaaacaaaggctgaagtctcttgcatctgtcgg1740
ttatttgagtataaagggtatctctgcagacacactcttaatgttcttcagtgttgtcat1800
ctttcttccatcccctcgcagtacatattgaagcggtggacaaaagatgcaaaaagccgg1860
catttttcaggagaacctcagcagttgcagactaggctgctgcggtataacgatttgtgc1920
gaacgagcattaaagctgaatgaagaggcatccctctctcaagagagttacaatattgca1980
ttccttgctattgaaggagctataggaaactgtgctggtattaatacttccggcagaagt2040
cttccagatgttgtaacgtcaccaactcaaggtctgatttctgtggaggaggacaaccat2100
agcagaagtgcaggcaaaaccagcaagaaaaagaatccaactaagaaaagaaaagtgaac2160
ccagagcaggatgttatgccagttgcagcgccagaaagcttgcaacagatggataagctg2220
agcccaaggacggttggcattgaaagctattatggaacacagcagagcgtacaaggaatg2280
gtccagctgaacttaatggggccaactcgcgataatttctatgggaatcaacaaacaatg2340
caaggattgcgacagttgaactcgatagcaccaagctacgacagttactacggtcctcag2400
caaggcatccatggacagggagtagatttcttcaggcctgccaatttttcttacgacatc2460
cgggatgatcctaatgtgaggactacgcagttgcatgaggacgcgtctagacactcgtaa2520
- 还没有人留言评论。精彩留言会获得点赞!