牛分枝杆菌分子伴侣蛋白GroEL1及其编码基因与应用的制作方法
本发明属于生物和疫苗技术领域,具体涉及一种牛分枝杆菌免疫相关的分子伴侣蛋白groel1,该蛋白具有高黏附能力和高侵袭能力,可以提高机体的igg、iga抗体水平以及th2型细胞免疫水平,表明该蛋白能够引发高水平体液免疫和细胞免疫,具有一定的抗牛分枝杆菌感染能力,有作为牛结核病疫苗的潜能。
背景技术:
牛结核病是一种慢性消耗性人畜共患病,奶牛结核病被列为我国优先防治的动物病种之一。牛分枝杆菌(mycobacteriumbovis,m.bovis)是引起牛结核病的主要病原,,同时也能够从牛传染给人及其它动物,给社会带来巨大的经济损失和产业损失。
疫苗是动物疫病防控的最为重要且有效的手段之一。但由于奶牛为乳用动物,考虑到公共卫生因素,各国对牛结核病尤其是奶牛结核病的疫苗免疫格外谨慎。随着部分国家牛结核病防控形势的日趋严峻,越来越多的国家开始选择对牛进行疫苗免疫来预防结核病。卡介苗是在牛结核病免疫时优选的疫苗,但其免疫效果与预期还存在较大的差距。为有效防控牛结核病,疫苗研发越来越受到广泛关注。有效的疫苗靶标是研制高效疫苗的基础,寻找高免疫原性的蛋白是开发高效疫苗的重中之重。
技术实现要素:
本发明目的在于提供一种牛分枝杆菌免疫相关蛋白groel1,该蛋白是一类分子伴侣蛋白,具有提高机体免疫水平的能力。
为了实现本发明的目的,申请人所在的华中农业大学农业微生物学国家重点实验室反刍动物病原学分室从牛分枝杆菌卡介苗中扩增到了groel1基因并在耻垢分枝杆菌中进行了克隆,经westernblot分析验证,该蛋白成功表达,细胞模型验证了该重组菌具有非常高的黏附和侵袭能力。进一步将groel1连接至原核表达载体中并在大肠杆菌中进行表达纯化,经westernblot分析验证,证实了该蛋白具有良好的免疫原性。将该纯化的重组蛋白groel1与佐剂混合后进行小鼠免疫,发现蛋白能够显著提高小鼠igg、iga抗体水平及th2型细胞免疫反应水平,具有重要的抗牛分枝杆菌感染潜能,是研发新药和疫苗的重要潜在靶点。
本发明所提供的牛分枝杆菌免疫相关的分子伴侣蛋白,名称为groel1蛋白,来源于牛分枝杆菌(mycobacteriumbovis)卡介苗,是如下1)或2)的蛋白质:
1)由seqidno:6所示的氨基酸序列组成的蛋白质;
2)将seqidno:6的氨基酸序列经过一个或几个氨基酸残基的取代和/或缺失和/或添加且由1)衍生的蛋白质。
seqidno:6由539个氨基酸残基组成。
上述序列可人工合成,也可先合成其编码基因,再进行生物表达得到。上述2)中的groel1的编码基因可通过将seqidno:5所示的自5′末端第1-1620位碱基所示的dna序列中缺失一个或几个氨基酸残基的密码子,和/或进行一个或几个碱基对的错义突变得到。
上述蛋白的编码基因也属于本发明的保护范围。
groel1蛋白的编码基因具体可为如下1)-3)中任一项所述的基因:
1)其核苷酸序列是seqidno:5所示的序列;
2)在严格条件下可与seqidno:5限定的dna序列杂交且编码上述groel1蛋白的dna分子;
3)与1)限定的基因具有90%以上的同源性,且编码上述groel1蛋白的dna分子。
所述3)中的基因,与1)或2)限定的基因最好有95%以上的同源性。
seqidno:5所示的序列由1620个碱基对组成,为groel1基因的开放阅读框架(orf),编码氨基酸序列是seqidno:6所示的groel1蛋白。
含有上述groel1蛋白编码基因的重组载体、转基因细胞系和重组菌也属于本发明的保护范围。
本发明的具体技术方案如下所述:
申请人所用的牛分枝杆菌卡介苗巴斯德株bcg【美国模式培养物保藏所(atcc):35734】为美国俄勒冈州立大学luizbermudez教授馈赠。本发明的前期工作包括对保藏的牛分枝杆菌卡介苗巴斯德株基因组的提取。
本发明以牛分枝杆菌卡介苗巴斯德株bcg全基因组为模板克隆groel1基因,并连接至pmv261载体中,构建了重组质粒pmv261-groel1,将该重组质粒转化到耻垢分枝杆菌中,得到重组耻垢分枝杆菌,命名为ms-pmv-g1。westernblot结果证实了该蛋白成功表达并具有反应原性。黏附及侵袭能力的检测结果显示该重组耻垢分枝杆菌相较野生株具有高黏附及侵袭能力。进一步将克隆的groel1基因连接至pet-30a载体中,构建了重组质粒pet-30a-groel1,将该重组质粒转化大肠杆菌dh5α得到重组大肠杆菌菌株,命名为escherichiacolipet-30a-groel1,于2019年5月7日送交位于湖北省武汉市武汉大学的中国典型培养物保藏中心保藏,保藏编号为cctccm2019333。该菌株在iptg诱导下,表达groel1基因编码的重组蛋白。westernblot结果验证了该重组蛋白的高免疫原性和反应原性。为验证该蛋白在体内的免疫学作用,将纯化的重组蛋白groel1与佐剂混合后进行小鼠免疫,结果显示,groel1蛋白能够诱发高水平的igg、iga抗体及th2型细胞免疫,具有抵抗牛分枝杆菌,预防牛结核病的潜能。
本发明所提供的groel1蛋白还可用于制备牛结核病诊断试剂盒,例如牛分枝杆菌igg或iga抗体酶联免疫检测试剂盒。
本发明具有以下优点:
1、本发明的groel1重组蛋白由牛分枝杆菌groel1基因编码,被发明人证实具有高黏附能力和侵袭能力。
2、本发明的groel1重组蛋白已被发明人证实具有免疫原性。
3、本发明的groel1重组蛋白已被发明人在小鼠体内证实能够引起机体产生高水平的iga、igg抗体。
4、本发明的groel1重组蛋白已被发明人在小鼠体内证实能够引起机体产生高水平的th2型细胞免疫。
更详细的技术方案请参见《具体实施方式》。
序列表说明:
序列表seqidno:1是扩增groel1基因片段的引物groel1-f1的序列,用于构建pmv261-groel1。
序列表seqidno:2是扩增groel1基因片段的引物groel1-r1的序列,用于构建pmv261-groel1。
序列表seqidno:3是扩增groel1基因片段的引物groel1-f2的序列,用于构建pet-30a-groel1。
序列表seqidno:4是扩增groel1基因片段的引物groel1-r2的序列,用于构建pet-30a-groel1。
序列表seqidno:5是groel1基因的核苷酸的序列,其长度为1620bp。
序列表seqidno:6是groel1蛋白的氨基酸序列,共由539个氨基酸组成。
附图说明
图1:是pmv216的质粒图谱。
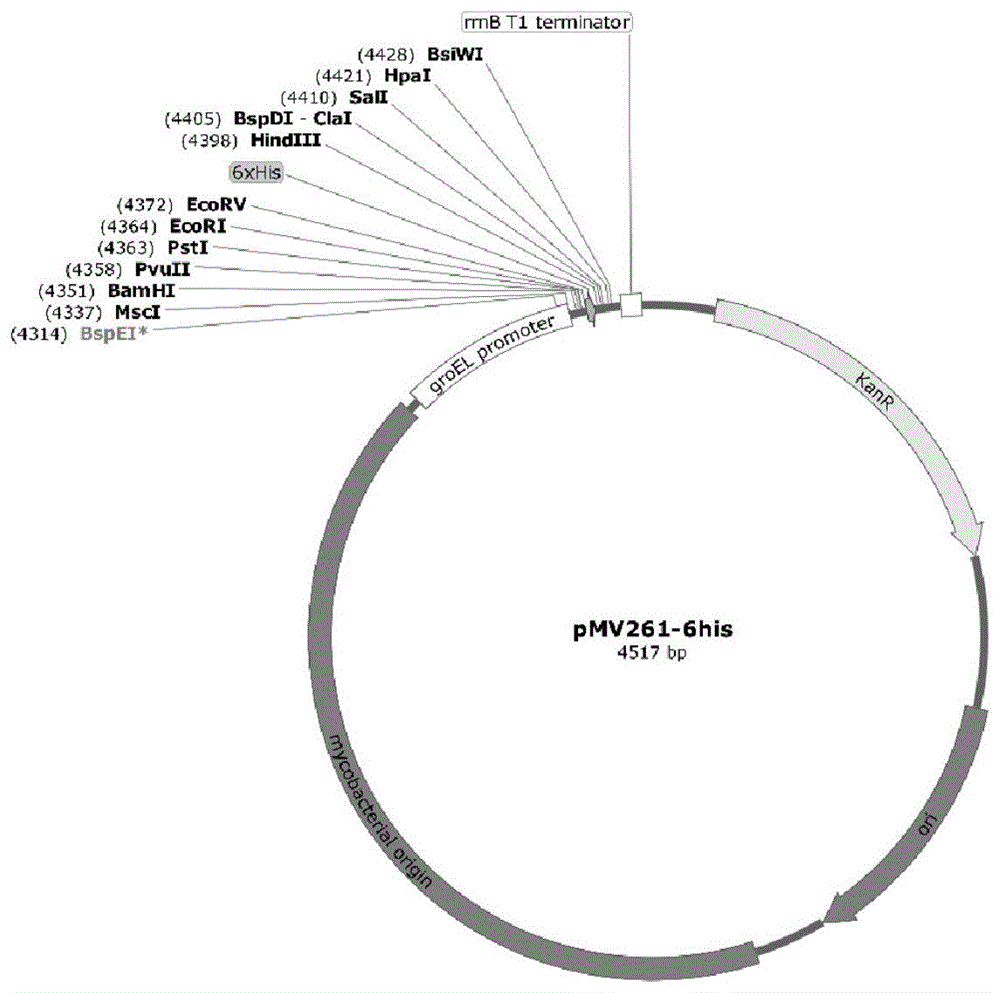
图2:是本发明构建的重组质粒pmv261-groel1的图谱。由pmv261质粒和groel1基因全长经过限制性内切酶消化后连接重组而成。
图3:westernblot验证ms-pmv-g1中groel1的表达。泳道m:高分子质量蛋白质标准;泳道1:ms-pmv-g1中groel1表达情况的检测;泳道2:耻垢分枝杆菌空菌对照。
图4:是ms-pmv-g1感染a549细胞后黏附及侵袭能力定量检测分析图。“***”表示p<0.001;a:黏附能力定量检测结果;b:侵袭能力定量检测结果。
图5:是pet-30a的质粒图谱。pet-30a为商业化质粒,购自novagen公司。
图6:是本发明构建的重组质粒pet-30a-groel1的图谱。是由pet-30a质粒和groel1基因全长经过限制性内切酶消化后连接重组而成。
图7:是纯化的牛分枝杆菌groel1蛋白sds-page图。泳道m:高分子质量蛋白质标准;泳道1:未诱导的含有pet-30a-groel1质粒的bl21重组菌;泳道2:iptg诱导后的含有pet-30a-groel1质粒的bl21重组菌;泳道3:iptg诱导后沉淀中表达情况;泳道4:iptg诱导后上清中表达情况;泳道5-6:洗脱液洗脱后蛋白检测;泳道7:纯化后的groel1蛋白。
图8:westernblot分析rgroel1蛋白的免疫原性。泳道m:高分子质量蛋白质标准;泳道1:含有pet-30a-groel1质粒的bl21重组菌。
图9:是各组小鼠免疫后的体重变化。图10:是各组小鼠免疫后的总igg和总iga抗体变化。a:总igg抗体检测结果;b:总iga抗体检测结果;*,p<0.05;**,p<0.01;***,p<0.001。
图11:是各组小鼠免疫后的特异性igg和特异性iga抗体变化。a:特异性igg抗体检测结果;b:特异性iga抗体检测结果;***,p<0.001。
图12:是各组小鼠免疫后的il-6及ifn-γ水平。a:il-6表达水平检测;b:ifn-γ表达水平检测;*,p<0.05;***,p<0.001。
具体实施方式
实施例1:重组耻垢分枝杆菌ms-pmv-g1的构建及验证
1.1重组耻垢分枝杆菌ms-pmv-g1的构建
以牛分枝杆菌卡介苗巴斯德株bcg(atcc:35734)全基因组为模板,利用如下设计的引物(序列表seqidno:1-2)克隆groel1基因,得到groel1基因的全序列,序列长度为1620bp。
扩增groel1基因的引物序列如下所示:
1、正向引物groel1-f1:gccaagacaattgcggatccatgagcaagctgatcgaatacgacg。下划线所示为bamhⅰ的酶切位点。对应序列表seqidno:1所示的序列。
2、反向引物groel1-r1:tggtggtggtggtggatatcgtgcgcgtgcccgtggt。下划线所示为ecorv的酶切位点。对应序列表seqidno:2所示的序列。
pcr反应体系如下:
模板dna2l,2×plantatmmastermix25l,引物各2l,超纯水19l。
pcr扩增条件如下:
98℃预变性5min,98℃变性30sec,62℃退火30sec,72℃延伸2min,进行30个循环反应;72℃彻底延伸10min。
回收pcr扩增产物,用bamhⅰ和ecorv酶切,同时将pmv261质粒(图1)用相同的酶进行双酶切。将酶切后的groel1基因和pmv261质粒用trelieftmsosoocloningkit(tsingke公司)进行连接,得到重组质粒pmv261-groel1(图2)。将该重组质粒pmv261-groel1转化耻垢分枝杆菌mc2155后,置于37℃摇床在110r/min下培养至对数期(od600=0.8~1.0)。采用菌液pcr鉴定正确后进行westernblot法测定表达。
1.2westernblot鉴定重组耻垢分枝杆菌ms-pmv-g1中groel1基因的表达
将1.1制备的重组耻垢分枝杆菌ms-pmv-g1经42℃热激诱导3-4h后收集菌液,4℃,l0000rpm/min离心5min后用pbs洗涤菌体3次,重悬菌体后超声破碎细胞进行sds-page电泳,将胶转移到pvdf膜上,膜洗涤后,将膜放入用tbst稀释的5%bsa中,室温封闭3h;充分洗涤后将pvdf膜放入2000倍稀释的his抗体中,4℃孵育过夜;充分洗涤后将pvdf膜放入10000倍稀释的辣根过氧化物酶标记的羊抗鼠igg中,室温1h;tbst洗涤三次后将混合好的ecl发光液均匀滴加在膜上,应用kodakimagestation化学发光检测仪检测信号。结果显示:ms-pmv-g1能与his抗体发生反应,可检测到特异性反应条带,分子量约为60kda(图3,泳道1),同时,耻垢分枝杆菌对照不能与his抗体发生反应,未检测到信号(图3,泳道2),因此ms-pmv-g1中groel1基因成功表达。
实施例2:重组耻垢分枝杆菌ms-pmv-g1黏附、侵袭能力的检测
2.1重组耻垢分枝杆菌ms-pmv-g1黏附能力的检测
将a549细胞(美国俄勒冈州立大学luizbermudez教授馈赠)在12孔细胞培养板中培养,待长成单层并达到2×105个/孔后,按照感染比10:1加入重组耻垢分枝杆菌ms-pmv-g1,设立空白耻垢分枝杆菌为对照,于4℃作用30min后用pbs充分洗涤,随后加入tritonx-100(bio-rad公司)裂解细胞,收集胞内菌涂布7h11固体(购自bd公司)平皿,于37℃、5%co2培养箱中培养5-7天。通过细菌计数发现重组菌ms-pmv-g1黏附能力显著高于野生菌株ms,p<0.001(图4a),为野生菌株的2.28倍。表明groel1具有提高细菌黏附力的功能。
2.2重组耻垢分枝杆菌ms-pmv-g1侵袭能力的检测
将a549细胞按照2×105个/孔铺至12孔细胞培养板,利用感染比为10:1将重组耻垢分枝杆菌ms-pmv-g1接种至a549细胞中,设置野生株耻垢分枝杆菌ms为对照。将细菌与a549细胞在37℃、5%co2培养箱中作用1h后加入tritonx-100(bio-rad公司)裂解细胞,利用菌落计数法定量测定菌落数。结果显示,重组耻垢分枝杆菌ms-pmv-g1侵袭能力显著高于野生株,p<0.001(图4b),为野生株的3.19倍。表明groel1具有提高细菌侵袭力的功能。
实施例3:groel1基因在大肠杆菌中的克隆、表达及纯化
3.1重组pet-30a-groel1质粒的克隆及表达
以牛分枝杆菌卡介苗巴斯德株(atcc:35734)全基因组为模板,利用如下设计的引物(序列表seqidno:3-4)克隆groel1基因,得到groel1基因的全序列,序列长度为1620bp。
扩增groel1基因的引物序列如下所示:
1、正向引物groel1-f2:cgggatccatgagcaagctgatcgaatacga。下划线所示为bamhi的酶切位点。对应序列表seqidno:3所示的序列。
2、反向引物groel1-r2:cggaattcgtgcgcgtgcccgtggt。下划线所示为ecori的酶切位点。对应序列表seqidno:4所示的序列。
pcr反应体系如下:模板dna2l,2×plantatmmastermix25l,引物各2l,超纯水19l。
pcr扩增条件如下:94℃预变性5min,94℃变性30sec,62℃退火30sec,72℃延伸45s,进行30个循环反应;72℃彻底延伸10min。
回收pcr扩增产物,用ecorⅰ和bamhi酶切,同时将pet-30a(+)质粒(图5)用相同的酶进行双酶切。将酶切后的groel1基因和pet-30a(+)质粒用t4dna连接酶进行连接,得到重组质粒pet-30a-groel1(图6)。将该重组质粒pet-30a-groel1转化大肠杆菌dh5α后,置于37℃摇床在180r/min下培养12h,提取质粒经过测序正确后(核苷酸序列对应序列表seqidno:5所示序列)转化大肠杆菌bl21菌,将该大肠杆菌重组菌在lb液体培养基中培养至od=0.6时取1ml菌液作为诱导前对照,同时加入异丙基硫代半乳糖苷(iptg)至终浓度为0.8mm,37℃摇床诱导表达4h。随后将样品12000r/min离心1min,弃掉上清,pbs重悬后12000r/min离心1min,弃上清,加入600μlpbs重悬,经过液压破碎仪进行菌体破碎,破碎压力1000kar破碎3次,至菌体澄清;然后12000r/min离心10min,分离上清和沉淀,沉淀用等体积600μlpbs重悬,分别取样进行加入适量的上样缓冲液,沸水煮10min,4000r/min离心2min。取上清及沉淀分别进行sds-page,观察目标蛋白的存在形式及表达情况。结果显示,groel1成功在上清中表达(图7,泳道4)。氨基酸序列对应序列表seqidno:6所示序列。
申请人将该大肠杆菌重组菌命名为escherichiacolipet-30a-groel1;于2019年5月7日送交位于湖北省武汉市武汉大学的中国典型培养物保藏中心(cctcc)保藏,保藏编号为cctccm2019333。
3.2groel1蛋白的纯化
将重组的大肠杆菌bl21按上述方法诱导表达裂解后,经过sds-page凝胶电泳后,确定groel1蛋白表达于上清中。利用镍柱对其进行纯化:
(1)将封闭柱子用的乙醇经柱子流出;
(2)向亲和层析柱中加入12ml灭菌去离子水洗涤;
(3)加入50ml的bindingbuffer平衡柱子;
(4)加入经过0.45μm孔径滤器过滤的蛋白上清液,收集前几滴滤出液体,编号为1。
(5)加入30ml的bindingbuffer平衡柱子,收集前几滴滤出液,编号为2。
(6)加入50ml的washingbuffer洗去杂蛋白,收集前几滴,编号为3。
(7)加入6ml的elutionbuffer洗脱目的蛋白,收集前几滴,编号为4。
(8)将编号为1-4的管中各加入50l上样缓冲液煮沸10min。
(9)配置sds-page聚丙烯酰胺凝胶,将处理好的样品加入孔中(20l/每孔),电泳(浓缩胶电泳条件为直流电压为80伏特,分离胶电泳条件为直流电压为120伏特),电泳完成后,取下凝胶用考马斯亮蓝染色过夜。然后脱色,确定得到纯化的目的蛋白(图7,泳道7)。
3.3groel1蛋白免疫原性的检测
采用westernblot方法进行rgroel1蛋白免疫原性鉴定,主要步骤为:将纯化的重组蛋白groel1(2.0μg/泳道)进行sds-page电泳,将胶上蛋白转移到pvdf膜上,膜洗涤后,将膜放入用tbst稀释的5%bsa中,室温封闭3h;充分洗涤后将pvdf膜放入5000倍稀释的牛阳性血清抗体中,4℃孵育过夜;充分洗涤后将pvdf膜放入5000倍稀释的辣根过氧化物酶标记的羊抗鼠igg中,室温1h;tbst洗涤三次后将混合好的ecl发光液均匀滴加在膜上,应用kodakimagestation化学发光检测仪检测信号。结果显示:groel1蛋白能与牛阳性血清发生反应,可检测到特异性反应条带,分子量约为60kda(图8),因此groel1蛋白具有良好的免疫原性。
实施例4:groel1蛋白引起小鼠免疫学反应的检测
4.1小鼠的免疫及体重监测
将groel1蛋白与佐剂(montanidetmims1313vgn)进行1:1混合,完全乳化后滴鼻免疫6-8周龄16-18g的雌性balb/c小鼠3只,每只25μg。同时设立单独免疫佐剂组和免疫pbs组为对照。第0天称重一次,随后每周称体重1次并做好记录,制成体重变化曲线表。结果显示,随着时间的延长,rgroel1蛋白免疫组、佐剂免疫组、pbs组小鼠的体重从18g缓慢增加至22g,各组之间体重变化无显著差异(p>0.05)(图9),小鼠精神状况均良好。
4.2免疫后小鼠总igg、iga抗体的检测
用欣博盛公司的小鼠iggelisa试剂盒及igaelisa试剂盒检测免疫后第0、7、14、21、28天的小鼠总igg、iga抗体。依照说明书进行检测,步骤简述如下:
将稀释后的血清加入回温后的反应板中,37℃孵育90min。充分洗涤后加入生物素化抗体工作液。37℃孵育60min。充分洗涤后加入酶结合物工作液,37℃避光孵育30min。充分洗涤后加入显色底物(tmb)避光37℃孵育15min。加入终止液后即刻测量od450值。
结果显示,自免疫后第14天起,groel1+佐剂免疫组小鼠的总igg抗体水平就显著高于单纯免疫佐剂及pbs组,p<0.05。第21和28天差异及其显著,p<0.001(图10a)。总iga抗体则从第7天起就持续显著增高,p<0.001(图10b)。总igg和总iga抗体水平呈现快速上升趋势。表明groel1蛋白能够在免疫早期就引起小鼠产生高水平的总igg和总iga抗体。
4.3免疫后小鼠特异性igg、iga抗体的检测
分别在免疫后第0、7、14、21、28天对小鼠进行断尾采血,收集血清后进行iga、igg抗体检测及细胞因子检测。具体步骤如下:
将groel1蛋白(终浓度为2μg/ml)作为抗原包被酶标板,4℃过夜后充分洗涤,加入封闭液后37℃封闭1h;充分洗涤后加入100μl100倍稀释的待检血清,37℃孵育2h;充分洗涤后加入辣根过氧化物酶标记的羊抗鼠igg或iga,37℃孵育2h,充分洗涤后加底物tmb显色10min;加入终止液2m硫酸,用酶联免疫检测仪读数,测定波长为450nm处的od值。结果显示,groel1+佐剂免疫组小鼠特异性igg(图11a)和特异性iga(图11b)抗体水平呈现持续性显著增高。自免疫后第7天起就与单纯免疫佐剂组和pbs组呈现极显著差异(p<0.001)。表明groel1蛋白能够在免疫早期就引起小鼠产生高水平的特异性igg和特异性iga抗体。
4.4免疫后小鼠细胞因子的检测
用欣博盛公司的小鼠ifn-γelisa试剂盒及il-6elisa试剂盒检测免疫后第0、7、14、21、28天的小鼠ifn-γ及il-6水平。依照说明书进行检测,步骤简述如下:
将稀释后的血清加入回温后的反应板中,37℃孵育90min。充分洗涤后加入生物素化抗体工作液。37℃孵育60min。充分洗涤后加入酶结合物工作液,37℃避光孵育30min。充分洗涤后加入显色底物(tmb)避光37℃孵育15min。加入终止液后即刻测量od450值。
结果显示:从第0天至第21天,各组小鼠ifn-γ水平均有一定程度的上升,第28天呈现下降趋势,但groel1+佐剂免疫组与佐剂对照组无显著差异(图12b),表明groel1蛋白并未引起小鼠产生高水平的ifn-γ。自免疫后第7天起,groel1+佐剂免疫组小鼠血清中il-6水平就显著高于佐剂对照组和pbs免疫组(p<0.001)(图12a),表明rgroel1蛋白能够在免疫早期就引起小鼠产生高水平的il-6,即th2型细胞免疫水平。
综上,groel1蛋白具有提高分枝杆菌黏附及侵袭能力的作用,指征该蛋白能够提高细菌的毒力;其原核表达蛋白能够引发小鼠产生高水平iga及igg抗体。虽然目前抗体对结核病的抵抗作用还存在一定的争论,但细胞因子il-6表达水平的显著上升验证了groel1蛋白能够引发高水平的th2型细胞免疫水平,表明该蛋白具有一定的抗牛分枝杆菌感染潜能,能够作为疫苗研发、诊断制剂的重要靶标。
序列表
<110>华中农业大学
<120>牛分枝杆菌分子伴侣蛋白groel1及其编码基因与应用
<160>6
<170>siposequencelisting1.0
<210>1
<211>45
<212>dna
<213>人工序列(artificialsequence)
<400>1
gccaagacaattgcggatccatgagcaagctgatcgaatacgacg45
<210>2
<211>37
<212>dna
<213>人工序列(artificialsequence)
<400>2
tggtggtggtggtggatatcgtgcgcgtgcccgtggt37
<210>3
<211>31
<212>dna
<213>人工序列(artificialsequence)
<400>3
cgggatccatgagcaagctgatcgaatacga31
<210>4
<211>25
<212>dna
<213>人工序列(artificialsequence)
<400>4
cggaattcgtgcgcgtgcccgtggt25
<210>5
<211>1620
<212>dna
<213>牛分枝杆菌(mycobacteriumbovis)
<400>5
atgagcaagctgatcgaatacgacgaaaccgcgcgtcgcgccatggaggtcggcatggac60
aagctggccgacaccgtgcgggtgacgctggggccgcgcggccggcatgtggtgctggcc120
aaggcgtttggcggacccacggttaccaacgacggcgtcacggtggcacgtgagatcgag180
ctggaagatccgtttgaagacttgggcgcccagctggtgaagtcggtggccaccaagacc240
aacgatgtggccggtgacggcaccaccaccgcaaccatcttggcgcaggcactgatcaag300
ggcggcctgaggctagtggccgccggcgtcaacccgatcgcgctcggcgtgggaatcggc360
aaggccgccgacgcggtatccgaggcgctgctggcatcggccacgccggtgtccggcaag420
accggcatcgcgcaggtggcgacggtgtcctcgcgcgacgagcagatcggtgacctggtt480
ggcgaagcgatgagcaaggtcggccacgacggcgtggtcagcgtcgaagaatcctcgacg540
ctgggcaccgagttggagttcaccgagggtatcggcttcgacaagggcttcttgtcggca600
tacttcgttaccgacttcgataaccagcaggcggtgctcgaggacgcgttgatcctgctg660
caccaagacaagatcagctcgcttcccgatctgttgccattgctggaaaaggttgcagga720
acgggtaagccactactgatcgtggctgaagacgtggagggcgaagcgttggcgacgctg780
gtcgtcaacgcgattcgcaagacgttgaaagcggtcgcggtcaaggggccgtacttcggt840
gaccgccgtaaggcgttccttgaggacctggcggtggtgacgggtggccaggtggtcaac900
cccgacgccggcatggtgctgcgcgaggtgggcttggaggtgctgggctcggcccgacgc960
gtggtggtcagcaaggacgacacggtcattgtcgacggcggcggcaccgcagaagcggtg1020
gccaaccgggcgaagcacttgcgtgccgagatcgacaagagcgattcggattgggatcgg1080
gaaaagcttggcgagcggctggccaaactggccggcggggttgctgtcatcaaggtgggt1140
gccgccaccgagaccgcactcaaggagcgcaaggaaagcgtcgaggatgcggtcgcggcc1200
gccaaggccgcggtcgaggagggcatcgtccctggtgggggagcctcgctcatccaccag1260
gcccgcaaggcgctgaccgaactgcgtgcgtcgctgaccggtgacgaggtcctcggtgtc1320
gacgtgttctccgaagcccttgccgcgccgttgttctggatcgccgccaacgctggcttg1380
gacggctcggtggtggtcaacaaggtcagcgagctacccgccgggcatgggctgaacgtg1440
aacaccctgagctatggtgacttggccgctgacggcgtcatcgacccggtcaaggtgact1500
aggtcggcggtgttgaacgcgtcatcggttgcccggatggtactcaccaccgagacggtc1560
gtggtcgacaagccggccaaggcagaagatcacgaccatcaccacgggcacgcgcactga1620
<210>6
<211>539
<212>rna
<213>牛分枝杆菌(mycobacteriumbovis)
<400>6
- 还没有人留言评论。精彩留言会获得点赞!
- 一种敲除c‑di‑AMP分解酶的重组耻垢分枝杆菌株及其应用的制造方法与工艺
- 检测结核分枝杆菌感染的抗原多肽池及其应用的制造方法与工艺
- 一种猕猴桃分枝定位器的制造方法与工艺
- 一种xanthone类化合物及其单晶的制备方法与作为抗海分枝杆菌药物的应用与制造工艺
- 一种糖肽类抗生素耐药基因检测试剂盒的制造方法与工艺
- 分枝杆菌快速分离培养方法与制造工艺
- 一种用于检测潜在结核的特异性标志物及其应用的制造方法与工艺
- 结核分枝杆菌抗原蛋白Rv2941及其T细胞表位肽的应用的制造方法与工艺
- 一种微流控芯片上基于荧光量子点磁性富集并分离结核分枝杆菌TB的方法及装置与制造工艺
- 二肽meso‑DAP‑D‑Ala和/或四肽L‑Ala‑D‑Glu‑L‑Lys‑D‑Ala的新用途的制造方法与工艺