qLIN6基因在调节或筛选稻米中油脂组分含量的应用的制作方法
本发明涉及水稻分子育种领域,更特别地,涉及qlin6基因在调节和筛选稻米中油脂组分含量的应用。
背景技术:
植物种子中脂质主要以甘油三酯的形式储存。大米中的油脂对大米的储存、食用品质和消费者的健康都非常重要,其含油率一般为2-4%。米饭的食用品质和适口性受到大米中的脂肪酸组成与含量的影响。目前,本领域中对大米的脂质合成通路知之甚少,liu等证实了osfad3将亚油酸(c18:2)转化成亚麻酸(c18:2)。zaplin等证实osfad2-1将油酸转化成亚油酸。一些研究者还初步鉴定了几个数量性状位点(qtl)。
但是,目前鉴定到的基因和qtl都太少,仍然无法知晓大米油脂合成的遗传和分子基础,因此需要找到影响大米中的油脂组分及含量的主效基因,用做筛选标志或遗传操作的靶点。
技术实现要素:
在本发明的发明人通过研究发现,鉴定了23个与油含量和油脂组成显著相关的位点,并且通过连锁分析的验证得到了11个有可能参与到了水稻中与稻米油脂代谢有关的通路的候选基因。在进一步的研究后,我们发现这些位点中,4个基因对油脂组成的自然变异贡献巨大,并且在亚群中有分化。其中之一就是qlin6基因,我们的研究显示,qlin6是负向调控c18:1含量、正向调控c18:2的主效基因。该基因的突变如果导致蛋白功能的降低或消失,使稻米中的c18:2含量降低,使c18:1含量升高。
基于以上研究,本发明提供了qlin6基因在调节稻米中油脂组分含量的应用,所述qlin6基因的序列如seqidno:1所示。
在一个优选实施方案中,所述油脂组分为c18:1和c18:2种的一种或多种组合。
本发明还提供了一种调节稻米中油脂组分含量的方法,包括导入qlin6基因、提高qlin6基因表达水平、降低qlin6基因表达水平或者突变qlin6基因使qlin6基因表达的蛋白功能提高、降低或消失的步骤,所述qlin6基因的序列如seqidno:1所示。
例如,如果目标稻种其他性状优良,但是稻米中c18:2含量太高或c18:1含量太低,并且该稻种含有正常功能的qlin6基因,可通过敲除或突变目标稻种中的qlin6基因,使qlin6基因不表达,或者表达的蛋白的磷脂酰胆碱:甘油二脂转磷酸胆碱酶2功能降低或没有磷脂酰胆碱:甘油二脂转磷酸胆碱酶2功能,即可降低稻米中的c18:2含量或提高c18:1含量。
反之,如果目标稻种其他性状优良,但是稻米中c18:2含量太低或c18:1含量太高,并且该稻种不含有正常功能的qlin6基因,可通过导入qlin6基因,或突变目标稻种中的不正常的qlin6基因使其回复突变成具有正常磷脂酰胆碱:甘油二脂转磷酸胆碱酶2功能的qlin6基因,即可提高稻米中的c18:2含量或降低c18:1含量。
本发明还提供了qlin6基因在水稻育种中作为稻米油脂组分含量的筛选标志的应用,所述qlin6基因的序列如seqidno:1所示。
本发明还提供了一种水稻杂交育种方法,包括使用qlin6基因或其突变体作为筛选标志来选择目标基因型的步骤。
在一个优选实施方案中,所述水稻杂交育种方法包括以下步骤:
s1:鉴定亲本中的qlin6基因及其突变状态;
s2:选择qlin6基因或其突变体作为筛选标志;
s3:将所述筛选标志与其他优势性状聚合到一起。
例如,已有稻种的其他性状优良,但稻米中c18:2含量太高或c18:1含量太低,并且含有正常功能的qlin6基因,要想通过杂交育种的方法降低稻米的c18:2含量或提高c18:1含量,可先选择qlin6基因发生了导致磷脂酰胆碱:甘油二脂转磷酸胆碱酶2功能降低或消失的突变的亲本与该稻种进行杂交,通过将qlin6基因突变体作为分子标志来使该突变基因与其他优势性状聚合到一起。
反之,已有稻种的其他性状优良,但稻米中c18:2含量太低或c18:1含量太高,并且不含正常功能的qlin6基因,要想通过杂交育种的方法提高稻米的c18:2含量或降低c18:1含量,可先选择具有正常磷脂酰胆碱:甘油二脂转磷酸胆碱酶2功能的qlin6基因亲本与该稻种进行杂交,通过将该正常qlin6基因作为分子标志来使该突变基因与其他优势性状聚合到一起。
附图说明
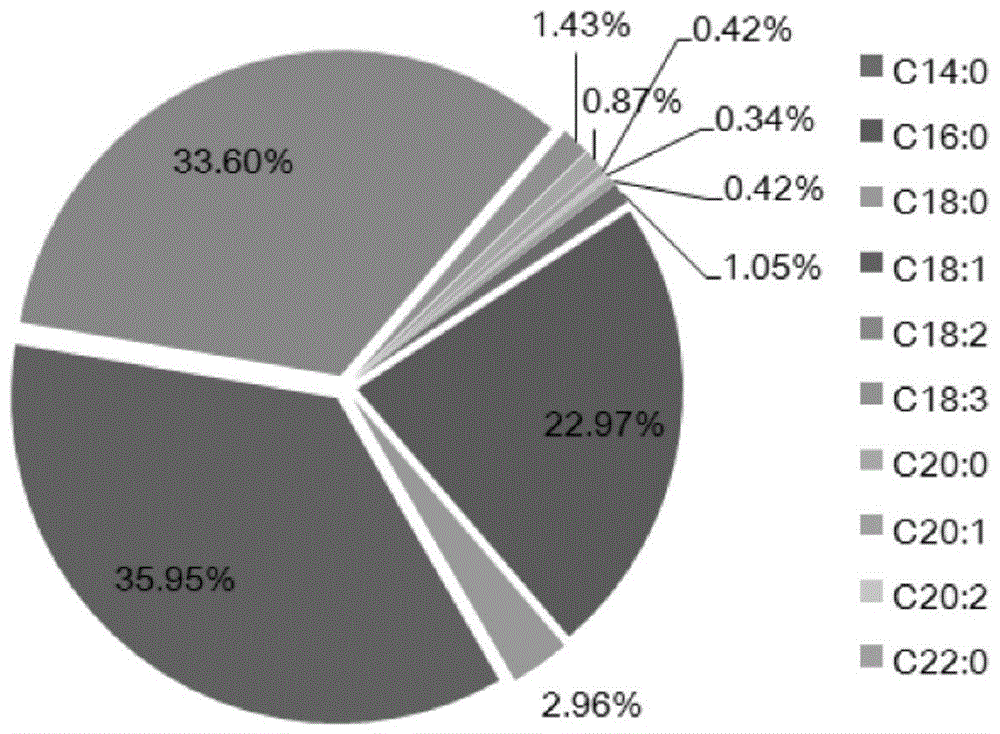
图1为通过gc-ms检测的533种稻米品种中的10种油脂组分的百分含量;
图2为533个稻米品种中澳洲稻(aus)、籼稻(ind)、粳稻(jap)的油脂相关性状的表型分布;
图3为籼稻亚群中c18:2含量关联分析的曼哈顿图(manhattanplot),虚线表示显著性阈值(-log10(p)=7.18)
图4为染色体6上的显著性峰周围的区域曼哈顿图和连锁不平衡(ld)热图,箭头表示候选基因的核苷酸变异位点,垂向虚线表示峰周围的候选区;
图5为qlin6的基因结构以及该基因的多态性;
图6为通过连锁分析鉴定qlin6作为w78/h6s的重组自交(ril)群体中c18:2含量的主效位点的qtl扫描结果的统计图;
图7为不同qlin6单倍型的c18:2含量统计图,其中a的测试对象为获取群体中的533个水稻品种,b的测试对象为由b805d/h6s衍生的ril群体;
图8为野生型和敲除突变家系中的10种脂肪酸含量的统计图,*p<0.05,**p<0.01;***p<0.001。
具体实施方式
以下结合附图对本发明的原理和特征进行描述,所举实例只用于解释本发明,并非用于限定本发明的范围。
1、样品来源
本发明使用的水稻材料包括533个栽培稻品种构成的自然群体和三个自己培育的重组自交群体。自然群体用于全基因组关联分析,三个重组自交群体用于连锁分析。
其中,533个栽培稻品种包括50个澳洲稻品种(aus)、305个籼稻品种(indica)和178个粳稻品种(japonica)。
三个重组近繁品种分别衍生自以下杂交系:珍汕97/明恢63(zs97/mh63)、88b-2/hd9802s和b805d/h6s。
所有四个群体均在武汉的华中农业大学的试验田中栽植生长,在五月中旬播种,六月中旬移栽到大田,收获后的稻谷晾干后室温储藏至少三个月,然后用于实验。
2、样品处理
将稻谷脱壳成糙米,然后碾粉,过80目筛,于80℃下干燥24h,然后进行脂质提取。0.2g米粉加入10ml玻璃管中,加入4.5ml硫酸:甲醇溶液(1:19)和100μl溶于氯仿的十七烷酸标准溶液,混匀。十七烷酸标准溶液的配置为1.07g十七烷酸标样溶于10ml色谱纯氯仿。将混合物在85℃水浴中孵育2h。冷却至室温后,使用正己烷提取脂肪酸甲酯,并通过气相色谱-质谱联用仪(gc-ms)分析。所有鉴定的脂肪酸含量的总和作为含油量。每个样品进行两次测试,将平均值用于后续分析。
3、gc-ms分析结果
gc-ms分析结果如图1所示,鉴定出的10种脂肪酸中,软脂酸(c16:0)、油酸(c18:1)和亚油酸(c18:2)占总脂肪酸含量的90%以上。我们在研究中发现了大量油脂相关性状存在广泛的变异,从c18:2含量的1.5倍差异到肉豆蔻酸(c14:0)含量的7.4倍差异。总脂肪酸含量介于22.99-40.96mg/g的范围。
其中,澳洲稻(aus)中的c16:0含量最高,其次是籼稻(ind)和粳稻(jap),c18:1和c18:2的含量则相反(图2)。
4、关联分析和连锁分析
全基因组关联研究(gwas):分别对籼稻、粳稻和所有样品实施gwas。在每个分析组中,仅选择次要位点频率(maf)>5%、缺失率<15%的snp进行关联分析。
连锁分析:对三个ril群体——zs97/mh63(96个株系)、88b-2/hd9802s(108个株系)和b805d/h6s(135个株系)——进行连锁分析。这些株系均通过二代测序,然后将50bp双末端读序比对到水稻参考基因组。其中比对质量≥40的序列和平均质量≥10的碱基用于鉴定snp。通过使用mpr算法来推断亲本株系的基因型,构建遗传bin图谱用于鉴定qtl。
根据以上工作,我们最终获得了520万个高质量snp,用于评估群体结构和亲缘关系,并在其中鉴定了23个显著的关联位点。在23个位点中,我们发现了qlin6是与c18:1和c18:2含量相关的主效qtl(图3)。
如图4所示,qlin6位于6号染色体的长臂上,我们基于点配对连锁不平衡(ld)相关系数(r2<0.6)分析了23.98-24.21mb(233kb)区域内的序列,最终确定该qtl对应的候选基因为loc_os06g40500(序列为seqidno:1),其在拟南芥中的同源基因编码磷脂酰胆碱:甘油二脂转磷酸胆碱酶2(pdct2)。
loc_os06g40500基因1号外显子上有两个snp位点导致氨基酸的改变,在整个gwas群中,第一snp位点对表型变异的贡献为27.5%,第二snp位点对表型变异的贡献为6.3%。我们推测,qlin6的1号外显子上第169位的g到a的突变(ex1-g/a)很可能导致了功能改变(图5),因此,以突变(ex1-g/a)作为研究对象。
qlin6的ex1-g/a位点的基因型在籼稻品种b805d与h6s之间存在差异。并且,使用b805d/h6s杂交系衍生的ril群体进行的qtl分析证实了qlin6单倍型对c18:1和c18:2含量的对比影响(图6,表1)。
表1油脂相关性状的表型分析
根据突变位点的g和a碱基,我们将qlin6划分为单倍型a(seqidno:1)和单倍型b(seqidno:2)。无论在获取群体中还是b805d/h6sril群体中,单倍型a的株系c18:2含量均高于具有单倍型b的株系(图7)。
5、转基因分析
为了进一步确定qlin6的功能,我们利用crispr-cas9系统敲除了zs97和nipponbare中的qlin6。在制备crispr/cas9敲除构建体上,我们仔细地检查了基因编码区,设计了带有c末端ngg的23bp靶点。确定靶点后,我们将20bp靶点插入中间载体per8-cas9-u6或per8-cas9-u3,并克隆至pcxun-cas9,得到敲除载体,命名为ko-lin6。将敲除构建体导入大肠杆菌straintrans5α感受态细胞中进行测序,验证正确后,导入根瘤农杆菌agrobacteriumtumefaciensstraineha105,然后分别转入zs97和nipponbare。获得转基因植株后,通过测序验证敲除系的基因型。
与野生型相比,zs97和nipponbare敲除系ko-qlin6c18:1含量显著升高,c18:2含量显著降低(图8)。但是nipponbare的敲除系表现出更明显的效果。可见,虽然qlin6的两种单倍型都有功能,但是单倍型b的功能比单倍型a要强。证明该位点可用于调节大米中的脂肪酸组成。
综合上面的实验可知,qlin6是负c18:1含量和正调控c18:2含量的主效基因。该基因的突变如果导致蛋白功能的降低或消失,使稻米中使c18:1升高和c18:2含量降低。
虽然以上的实例中仅展示了通过crispr-cas9系统得到敲除突变体和ex1-g/a突变体,但是,本领域技术人员在阅读本申请披露的内容后可知,只需要将该基因导入、敲除、敲降或突变成相关功能降低或无相关功能,即可达到调节稻米中c18:1和c18:2含量的目的。
以上所述仅为本发明的较佳实施例,并不用以限制本发明,凡在本发明的精神和原则之内,所作的任何修改、等同替换、改进等,均应包含在本发明的保护范围之内。
序列表
<110>华中农业大学
湖北双水双绿生物科技有限公司
<120>qlin6基因在调节或筛选稻米中油脂组分含量的应用
<160>2
<170>siposequencelisting1.0
<210>1
<211>924
<212>dna
<213>水稻(oryzasativa)
<400>1
atgccgccgccgccgccgcccagcctcacggccaacaccgcatcctccatgggcaacgcc60
gaggccgtcgtggtgctgcccgcgaacggcggcgcgcggcggcgcgccgacaaggtcgtc120
cacccggcgccgatgccggacagagcagctggtggcgcgatggagagggaaggcggcggc180
gtcggcggcggcggcgaggtgggtgggtggaggaggccggagtggtgctcggcggcgggg240
gtggcgggggtcctgcggcggcacccggcggcggcggcgttcgggtgcgggctgctgctg300
ttcatggccgtggagtacaccatccccatggtgccgcccgccgcgccgccggtcgacctc360
ggcttcgccgccaccgccgcgctccacgccgggatcgccgcccgcccatggctcaactcg420
ctcctcgccgcgctcaacacggtgttcgtggcgatgcaggcggcgtacatcctgtgggcg480
atcctcggcgagggccggccgcgcgccgccgtggcggcgatgatgatgttcacctgccgc540
ggcgcgctcggctgcgccacgcagctgccgctgccggccgagttcctgggctccggcatg600
gacttccccgtcggcaacgtctccttcttcctcttcttctccggccacgtcgccggcgcg660
gtgatcgccgccgaggacatgcgccgcgcggggcgccgcggcatggcgcgcctctacgac720
gcgctcaacctgctccagggcgtcaggctgctcgcctgcaggggccactacaccatcgac780
ctcgccgtcggcgtcggcgccggcctcctcttcgacatgctcgccggcaggtacctggac840
ggcaagaacaccgtcgacggcggcgccgccgtggcgccggggagccggtgctgcagctgc900
cacaaggctctcttgtcacagtag924
<210>2
<211>924
<212>dna
<213>水稻(oryzasativa)
<400>2
atgccgccgccgccgccgcccagcctcacggccaacaccgcatcctccatgggcaacgcc60
gaggccgtcgtggtgctgcccgcgaacggcggcgcgcggcggcgcgccgacaaggtcgtc120
cacccggcgccgatgccggacagagcagctggtggcgcgatggagaggaaaggcggcagc180
gtcggcggcggcggcgaggtgggtgggtggaggaggccggagtggtgctcggcggcgggg240
gtggcgggggtcctgcggcggcacccggcggcggcggcgttcgggtgcgggctgctgctg300
ttcatggccgtggagtacaccatccccatggtgccgcccgccgcgccgccggtcgacctc360
ggcttcgccgccaccgccgcgctccacgccgggatcgccgcccgcccatggctcaactcg420
ctcctcgccgcgctcaacacggtgttcgtggcgatgcaggcggcgtacatcctgtgggcg480
atcctcggcgagggccggccgcgcgccgccgtggcggcgatgatgatgttcacctgccgc540
ggcgcgctcggctgcgccacgcagctgccgctgccggccgagttcctgggctccggcatg600
gacttccccgtcggcaacgtctccttcttcctcttcttctccggccacgtcgccggcgcg660
gtgatcgccgccgaggacatgcgccgcgcggggcgccgcggcatggcgcgcctctacgac720
gcgctcaacctgctccagggcgtcaggctgctcgcctgcaggggccactacaccatcgac780
ctcgccgtcggcgtcggcgccggcctcctcttcgacatgctcgccggcaggtacctggac840
ggcaagaacaccgtcgacggcggcgccgccgtggcgccggggagccggtgctgcagctgc900
cacaaggctctcttgtcacagtag924
- 还没有人留言评论。精彩留言会获得点赞!