中和抗甲型流感抗体及其用途的制作方法
中和抗甲型流感抗体及其用途
1.本技术是申请日为2014年10月1日的中国专利申请201480053969.3“中和抗甲型流感抗体及其用途”的分案申请。
技术领域
2.本发明涉及具有针对甲型流感病毒的广泛中和活性的抗体并且涉及这类抗体的用途。
背景技术:
3.流感病毒引起年度流感流行和偶然的大流行,这对全世界公共卫生构成显著威胁。季节性流感感染与每年200,000
‑
500,000例死亡相关联,特别是在幼儿、免疫功能低下患者和老年人中。死亡率典型地在具有大流行性流感爆发的季节期间进一步增加。对于开发用于预防和治疗流感感染(特别是缺乏服务的群体中)的有效抗病毒治疗剂仍然存在显著未满足的医学需求。
4.存在三种类型的流感病毒,甲型、乙型和丙型。甲型流感病毒可以感染各种各样的鸟类和哺乳动物,包括人、猪、鸡、以及雪貂。甲型流感病毒可以基于编码表面糖蛋白血凝素(ha)和神经氨酸酶(na)的两种基因的抗原区中的等位基因变异而分类为亚型。ha是受体结合和膜融合糖蛋白,其介导病毒附着和进入靶细胞;ha是保护性体液免疫应答的主要靶标。ha蛋白是三聚体结构的并且包含单一多肽前体ha0的三个相同拷贝,该多肽前体在蛋白水解成熟时裂解成含有球状头部(ha1)和柄区(ha2)的ph依赖性、亚稳定的中间体。膜远侧“球状头部”构成ha1结构的大部分并且包含用于病毒进入的唾液酸结合口袋和主要抗原结构域。从ha2和一些ha1残基组装的膜近侧“柄”结构包含融合机器,其在晚期内体的低ph环境中经历构象变化以便触发膜融合和渗透至细胞中。甲型流感亚型之间的序列同源性程度在ha1中(亚型之间34%
‑
59%同源性)比在ha2区中(51%
‑
80%同源性)低。由流感病毒感染引发的中和抗体通常靶向可变ha1球状头部以便防止病毒受体结合且通常是毒株特异性的。罕见地,已经鉴别了靶向ha的球状头部的广泛交叉反应性单克隆抗体(克劳斯j.c.(krause j.c.)等人,2011病毒学杂志(j.virol.)85;惠特尔j.(whittle j.)等人,2011美国国家科学院院刊(pnas)108;爱可尔特dc(ekiert dc)等人,2012自然(nature)489;李ps(lee ps)等人,2012美国国家科学院院刊(pnas)109)。相比之下,柄区的结构是相对保守的并且最近已鉴别了结合ha柄以便防止用于病毒进入的ph引发的融合步骤的少数广泛中和抗体(爱可尔特dc等人,2009科学(science)324;瑞j.(sui j.)等人,自然结构与分子生物学(nat struct mol biol)16;拉马特j(wrammert j)等人,2011实验医学杂志(j exp med)208;爱可尔特dc等人,2011科学333;科尔蒂d(corti d)等人,2010临床研究杂志(j clin invest)120;思罗斯比m.(throsby m.),2008公共科学图书馆
·
综合(plos one)3)。这些柄反应性中和抗体中的大多数是对甲型流感1组病毒特异性的或对2组病毒特异性的。最近,分离了与1组病毒和2组病毒两者交叉反应的柄结合抗体(科尔蒂d等人,2011科学333;李gm(li gm)等人,2012美国国家科学院院刊109和西里尔d(cyrille d)等人,2012科学337;纳卡穆
拉g(nakamura g)等人,2013,细胞宿主与微生物(cell host&microbe)14)。
5.迄今为止,不存在广泛地中和或抑制所有甲型流感病毒感染或减轻由甲型流感病毒引起的疾病的市售抗体。因此,对于保护免受甲型流感病毒的多种1组亚型和2组亚型的新型抗体仍然存在需要。
技术实现要素:
6.本发明提供一种针对甲型流感病毒的抗体或其结合片段,该抗体或其结合片段能够结合甲型流感病毒血凝素并且中和甲型流感病毒的至少一种1组亚型和至少一种2组亚型。
7.优选地,本发明的抗体或结合片段能够结合甲型流感病毒血凝素并且中和至少1、2、3、4、5、6、7、8、9或10种甲型流感病毒1组亚型和至少1、2、3、4、5或6种甲型流感病毒2组亚型。进一步优选地,本发明的抗体或结合片段能够结合甲型流感病毒血凝素并且中和至少5种甲型流感病毒1组亚型和至少1或2种甲型流感病毒2组亚型。
8.甲型流感病毒的血凝素亚型分为两个主要系统发育分组,这些分组被鉴别为1组,该组包括亚型h1、h2、h5、h6、h8、h9、h11、h12、h13、h16、以及h17;和2组,该组包括亚型h3、h4、h7、h10、h14、以及h15。在一个实施例中,根据本发明的抗体或结合片段能够结合和/或中和选自h1、h2、h5、h6、h8、h9、h11、h12、h13、h16和h17及其变体的一种或多种甲型流感病毒1组亚型;以及选自h3、h4、h7、h10、h14和h15及其变体的一种或多种甲型流感病毒2组亚型。在另一个实施例中,根据本发明的抗体或结合片段能够结合和/或中和甲型流感病毒1组亚型h1、h2、h5、h6、h8、h9、h11、h12、h13、h16和h17;以及甲型流感病毒2组亚型h3、h4、h7、h10、h14和h15。在另一个实施例中,该抗体或结合片段能够结合和/或中和1组亚型h1、h2、h5、h6和h9以及2组亚型h3和h7。在另一个实施例中,该抗体或结合片段能够结合和/或中和1组亚型h1、h2、h5和h6以及2组亚型h3和h7。
9.本发明是基于从作为起始材料的自单个供体收集的igg记忆b细胞分离天然存在的人单克隆抗体(mab)。优化用于产生具有改进的特征的抗体变体,如在此所描述。这些优化的抗体变体不是天然存在的;它们是使用重组技术产生的。本发明的抗体或其片段结合ha的柄区并且中和甲型流感病毒的多于一种亚型的感染,这些亚型分别选自1组亚型和2组亚型。为抗甲型流感ha柄结合抗体的本发明的抗体展示相较于来自公开文献(抗体fi6v4,wo2013/011347a1中所描述)且在实例5的表6中所示的抗体更宽的气息覆盖范围或针对甲型流感病毒的更好中和活性。此外,本发明的抗体可在阻断ha成熟方面比一种或多种其他mab更有效,如在实例6的图1中所示。
10.在一些实施例中,该抗体或其结合片段包含一组六个cdr,其中该组六个cdr选自下组,该组由以下各项组成:
11.(a)seq id no.:3的hcdr1、seq id no.:4的hcdr2、seq id no.:5的hcdr3、seq id no.:8的lcdr1、seq id no.:9的lcdr2、以及seq id no.:10的lcdr3;
12.(b)seq id no.:13的hcdr1、seq id no.:14的hcdr2、seq id no.:15的hcdr3、seq id no.:18的lcdr1、seq id no.:19的lcdr2、seq id no.:20的lcdr3;
13.(c)seq id no.:23的hcdr1、seq id no.:24的hcdr2、seq id no.:25的hcdr3、seq id no.:28的lcdr1、seq id no.:29的lcdr2、以及seq id no.:30的lcdr3;
14.(d)seq id no.:33的hcdr1、seq id no.:34的hcdr2、seq id no.:35的hcdr3、seq id no.:38的lcdr1、seq id no.:39的lcdr2、以及seq id no.:40的lcdr3;
15.(e)seq id no.:43的hcdr1、seq id no.:44的hcdr2、seq id no.:45的hcdr3、seq id no.:48的lcdr1、seq id no.:49的lcdr2、以及seq id no.:50的lcdr3;
16.(f)seq id no.:53的hcdr1、seq id no.:54的hcdr2、seq id no.:55的hcdr3、seq id no.:58的lcdr1、seq id no.:59的lcdr2、以及seq id no.:60的lcdr3;
17.(g)seq id no.:63的hcdr1、seq id no.:64的hcdr2、seq id no.:65的hcdr3、seq id no.:68的lcdr1、seq id no.:69的lcdr2、以及seq id no.:70的lcdr3;
18.(h)seq id no.:73的hcdr1、seq id no.:74的hcdr2、seq id no.:75的hcdr3、seq id no.:78的lcdr1、seq id no.:79的lcdr2、以及seq id no.:80的lcdr3;
19.(i)seq id no.:83的hcdr1、seq id no.:84的hcdr2、seq id no.:85的hcdr3、seq id no.:88的lcdr1、seq id no.:89的lcdr2、seq id no.:90的lcdr3;
20.(j)seq id no.:93的hcdr1、seq id no.:94的hcdr2、seq id no.:95的hcdr3、seq id no.:98的lcdr1、seq id no.:99的lcdr2、以及seq id no.:100的lcdr3;
21.(k)seq id no.:103的hcdr1、seq id no.:104的hcdr2、seq id no.:105的hcdr3、seq id no.:108的lcdr1、seq id no.:109的lcdr2、以及
22.seq id no.:110的lcdr3;
23.(l)seq id no.:113的hcdr1、seq id no.:114的hcdr2、seq id no.:115的hcdr3、seq id no.:118的lcdr1、seq id no.:119的lcdr2、以及
24.seq id no.:110的lcdr3;
25.(m)seq id no.:123的hcdr1、seq id no.:124的hcdr2、seq id no.:125的hcdr3、seq id no.:128的lcdr1、seq id no.:129的lcdr2、以及seq id no.:130的lcdr3;
26.(n)seq id no.:133的hcdr1、seq id no.:134的hcdr2、seq id no.:135的hcdr3、seq id no.:138的lcdr1、seq id no.:139的lcdr2、以及seq id no.:140的lcdr3;以及
27.(o)seq id no.:143的hcdr1、seq id no.:144的hcdr2、seq id no.:145的hcdr3、seq id no.:148的lcdr1、seq id no.:149的lcdr2、以及seq id no.:150的lcdr3;
28.(p)根据(a)至(o)中任一项的一组六个cdr包含一个或多个氨基酸取代、缺失或插入;
29.(q)根据(a)至(p)中任一项的一组六个cdr包含1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、或24或25个氨基酸取代;
30.(r)根据(a)至(q)中任一项的一组六个cdr hcdr1、hcdr2、hcdr3、lcdr1、lcdr2、lcdr3包含:
31.(i)hcdr1,其具有与seq id no:3一致或相对于seq id no:3包含3个或更少个氨基酸残基取代的氨基酸序列;
32.(ii)hcdr2,其具有与seq id no:4一致或相对于seq id no:4包含5个或更少个氨基酸残基取代的氨基酸序列;
33.(iii)hcdr3,其具有与seq id no:5一致或相对于seq id no:5包含6个或更少个氨基酸残基取代的氨基酸序列;
34.(iv)lcdr1,其具有与seq id no:6一致或相对于seq id no:6包含5个或更少个氨
基酸残基取代和/或一个缺失的氨基酸序列;
35.(v)lcdr2,其具有与seq id no:7一致或相对于seq id no:7包含5个或更少个氨基酸残基取代的氨基酸序列;以及
36.(vi)lcdr3,其具有与seq id no:8一致或相对于seq id no:8包含1个或更少个氨基酸残基取代的氨基酸序列;
37.(s)根据(a)至(r)中任一项的一组六个cdr hcdr1、hcdr2、hcdr3、lcdr1、lcdr2、lcdr3包含:
38.(i)hcdr1,其中:
39.kabat残基31为s,
40.kabat残基32为n或y,
41.kabat残基33为n、s或r,
42.kabat残基34为a,
43.kabat残基35为v或t,
44.kabat残基35a为w
45.kabat残基35b为n;
46.(ii)hcdr2,其中:
47.kabat残基50为r,
48.kabat残基51为t,
49.kabat残基52为y,
50.kabat残基52a为y,
51.kabat残基53为r,
52.kabat残基54为s,
53.kabat残基55为k或g,
54.kabat残基56为w,
55.kabat残基57为y,
56.kabat残基58为n或y,
57.kabat残基59为d,
58.kabat残基60为y,
59.kabat残基61为a,
60.kabat残基62为e、v或d,
61.kabat残基63为s或f,
62.kabat残基64为v或l,
63.kabat残基65为k;
64.(iii)hcdr3,其中:
65.kabat残基95为s或g,
66.kabat残基96为g,
67.kabat残基97为h,
68.kabat残基98为i,
69.kabat残基99为t,
70.kabat残基100为v或e,
71.kabat残基100a为f,
72.kabat残基100b为g,
73.kabat残基100c为v或l,
74.kabat残基100d为n,
75.kabat残基100e为v或i,
76.kabat残基100f为d,
77.kabat残基100g为a,
78.kabat残基100f为f或y,
79.kabat残基101为d,
80.kabat残基102为m、i或v;
81.(iv)lcdr1,其中:
82.kabat残基24为r,
83.kabat残基25为t、a或不存在,
84.kabat残基26为s或a,
85.kabat残基27为q,
86.kabat残基28为s或r,
87.kabat残基29为l,
88.kabat残基30为s、n或r
89.kabat残基31为s,
90.kabat残基32为y,
91.kabat残基33为l、t或d,
92.kabat残基34为h;
93.(v)lcdr2,其中:
94.kabat残基50为a,
95.kabat残基51为a、t或s,
96.kabat残基52为s或t,
97.kabat残基53为s或t,
98.kabat残基54为l或r,
99.kabat残基55为q、l或g,
100.kabat残基56为s;以及,
101.(vi)lcdr3,其中:
102.kabat残基89为q,
103.kabat残基90为q或l,
104.kabat残基91为s,
105.kabat残基92为r,并且
106.kabat残基93为t。
107.本发明提供抗体及其结合片段,该抗体及其结合片段包含一组六个cdr:hcdr1、hcdr2、hcdr3、lcdr1、lcdr2、lcdr3,其中该组六个cdr在表11和13中示出。
108.本发明的变体抗体序列可以与本技术中列举的这些序列共有75%或更高(例如,80%、85%、90%、95%、97%、98%、99%或更高)氨基酸序列一致性。在一些实施例中,序列一致性是关于参考序列(即,在本技术中列举的序列)的全长计算的。在一些另外的实施例中,如在此所提及的一致性百分比是如使用blast 2.1.3版、使用由ncbi(国家生物技术信息中心(national center for biotechnology information);http://www.ncbi.nlm.nih.gov/)指定的默认参数[blosum 62矩阵;空位开放罚分=l 1并且空位扩展罚分=l]来确定的。
[0109]
变体抗体也包括在本发明的范围内。因此,在本技术中列举的这些序列的变体也包括在本发明的范围内。具有改进的亲和力和/或效力的抗体序列的变体可以使用本领域中已知的方法来获得并且包括在本发明的范围内。例如,氨基酸取代可以用于获得具有进一步改进的亲和力的抗体。可替代地,核苷酸序列的密码子优化可以用于提高用以产生抗体的表达系统中的翻译效率。此外,包含通过将定向进化方法应用于本发明的任何核酸序列来针对抗体特异性或中和活性优化的序列的多核苷酸也在本发明的范围内。
[0110]
本发明提供一种根据本发明的抗体或其结合片段,该抗体或其结合片段包含与选自下组的vh具有至少75%一致性的vh和/或与选自下组的vl具有至少75%一致性的vl,该组由以下各项组成:
[0111]
(a)seq id no.:2的vh和seq id no.:7的vl,
[0112]
(b)seq id no.:12的vh和seq id no.:17的vl,
[0113]
(c)seq id no.:22的vh和seq id no.:27的vl,
[0114]
(d)seq id no.:32的vh和seq id no.:37的vl,
[0115]
(e)seq id no.:42的vh和seq id no.:47的vl,
[0116]
(f)seq id no.:52的vh和seq id no.:57的vl,
[0117]
(g)seq id no.:62的vh和seq id no.:67的vl,
[0118]
(h)seq id no.:72的vh和seq id no.:77的vl,
[0119]
(i)seq id no.:82的vh和seq id no.:87的vl,
[0120]
(j)seq id no.:92的vh和seq id no.:97的vl,
[0121]
(k)seq id no.:102的vh和seq id no.:107的vl,
[0122]
(l)seq id no.:112的vh和seq id no.:117的vl,
[0123]
(m)seq id no.:122的vh和seq id no.:127的vl,
[0124]
(n)seq id no.:132的vh和seq id no.:137的vl,
[0125]
(o)seq id no.:144的vh和seq id no.:147的vl,以及
[0126]
(p)seq id no:152的vh和seq id no:157的vl。
[0127]
根据本发明的抗体或其结合片段可以包含选自下组的vh和vl,该组由以下各项组成:
[0128]
(a)seq id no.:2的vh和seq id no.:7的vl,
[0129]
(b)seq id no.:12的vh和seq id no.:17的vl,
[0130]
(c)seq id no.:22的vh和seq id no.:27的vl,
[0131]
(d)seq id no.:32的vh和seq id no.:37的vl,
[0132]
(e)seq id no.:42的vh和seq id no.:47的vl,
[0133]
(f)seq id no.:52的vh和seq id no.:57的vl,
[0134]
(g)seq id no.:62的vh和seq id no.:67的vl,
[0135]
(h)seq id no.:72的vh和seq id no.:77的vl,
[0136]
(i)seq id no.:82的vh和seq id no.:87的vl,
[0137]
(j)seq id no.:92的vh和seq id no.:97的vl,
[0138]
(k)seq id no.:102的vh和seq id no.:107的vl,
[0139]
(l)seq id no.:112的vh和seq id no.:117的vl,
[0140]
(m)seq id no.:122的vh和seq id no.:127的vl,
[0141]
(n)seq id no.:132的vh和seq id no.:137的vl,
[0142]
(o)seq id no.:144的vh和seq id no.:147的vl,以及
[0143]
(p)seq id no:152的vh和seq id no:157的vl。
[0144]
根据本发明的抗体或其结合片段可以选自下组,该组由以下各项组成:免疫球蛋白分子、单克隆抗体、嵌合抗体、cdr移植抗体、人源化抗体、fab、fab'、f(ab')2、fv、二硫键连接的fv、scfv、单结构域抗体、双抗体、多特异性抗体、双重特异性抗体、以及双特异性抗体。
[0145]
根据本发明的抗体或其结合片段可包含vh,其包含人种系框架,优选vh6
‑
1;和/或vl,其包含人种系框架,优选vk1
‑
39。优选地,根据本发明的抗体或其结合片段包含vh,其包含人种系框架vh6
‑
1;以及vl,其包含人种系框架vk1
‑
39。vh6框架很少用于抗体中。
[0146]
根据本发明的抗体或其结合片段可包含fc区,优选地该抗体是igg1、igg2或igg4或其结合片段。
[0147]
在一个实施例中,本发明的抗体包含人igg恒定结构域,该恒定结构域具有相对于野生型人igg恒定结构域的一个或多个氨基酸取代。本发明的抗体可以包含具有m252y、s254t和t256e(“yte”)氨基酸取代的人igg恒定结构域,其中氨基酸残基是根据卡巴特(kabat)中的eu索引进行编号的。
[0148]
本发明还提供一种针对甲型流感病毒的抗体或其结合片段,该抗体或其结合片段能够结合甲型流感病毒血凝素并且中和甲型流感病毒的至少一种1组亚型和至少一种2组亚型,其特征在于该抗体或其结合片段与以上所述的本发明的抗体竞争结合甲型流感病毒血凝素。因此,本发明包括一种与本发明的抗体结合相同表位的抗体或其片段,或一种与本发明的抗体竞争结合的抗体。
[0149]
本发明进一步提供一种分离的核酸,该核酸编码根据本发明的抗体或其片段。优选地,该核酸是cdna。本发明还包括编码本发明的抗体的轻链和重链以及cdr中的部分或全部的核酸序列。因此,在此提供编码本发明的示例性抗体的轻链和重链以及cdr中的部分或全部的核酸序列。提供编码本发明的示例性抗体的cdr、重链可变区和轻链可变区的核酸序列的seq id编号。由于遗传密码的冗余性,将存在这些序列的变体,这些变体编码相同的氨基酸序列。
[0150]
本发明再进一步提供一种包含根据本发明的分离的核酸的载体;优选地该载体是表达载体。
[0151]
此外,本发明提供一种包含根据本发明的分离的核酸或载体的宿主细胞。适合的宿主细胞包括哺乳动物细胞系,如来源于hek或cho细胞的那些。
[0152]
此外,本发明提供一种用于制造本发明的抗体或片段的方法,该方法包括在适合用于表达该抗体或其片段的条件下培养本发明的宿主细胞。
[0153]
这类方法可以进一步包括从该宿主细胞培养物分离该抗体或其片段并且任选地将该分离的抗体或片段配制成组合物。
[0154]
本发明再进一步提供一种包含根据本发明的抗体或其片段和药学上可接受的载体的组合物。
[0155]
本发明还提供一种组合物,该组合物包含根据本发明的抗体或其片段、组氨酸和nacl(在从约5.5至约6.5范围内的ph下,优选在约ph 6.0下);又更优选地包含根据本发明的抗体或其片段、约20至约30mm组氨酸和约0.1至约0.2m nacl(在从约5.5至约6.5范围内的ph下,优选在约ph 6.0下);最优选地包含25mm his和0.15m nacl(在从约5.5至约6.5范围内的ph下,例如在约ph 6.0下)。
[0156]
此外,本发明提供:
[0157]
‑
一种根据本发明的抗体或其片段,该抗体或其片段用于在预防或治疗受试者的甲型流感感染中使用;
[0158]
‑
根据本发明的抗体或其片段在制造用于预防或治疗受试者的甲型流感感染的药剂中的用途;
[0159]
‑
一种用于预防或治疗受试者的甲型流感感染的方法,该方法包括给予根据本发明的抗体或其片段;
[0160]
‑
根据本发明的抗体或其片段防止用于甲型流感病毒进入细胞的ph触发的融合步骤的用途;或
[0161]
‑
根据本发明的抗体或其片段用于抑制甲型流感病毒ha成熟的用途。
[0162]
本发明的示例性抗体包括但不限于:抗体3、抗体5、抗体6、抗体8、抗体10、抗体11、抗体12、抗体13、抗体14、以及抗体15。
[0163]
本发明还提供根据本发明的抗体或其结合片段在体外诊断受试者的甲型流感感染中的用途。
[0164]
详细说明
[0165]
引言
[0166]
本发明提供结合甲型流感病毒血凝素(ha)柄且中和如在此所述的甲型流感病毒感染1组亚型和2组亚型的抗体(包括人形式)以及其片段、衍生物/轭合物和组合物;这类抗甲型流感病毒ha柄抗体在此被称为本发明的抗体。
[0167]
如在此所用,术语“中和”是指抗体或其结合片段结合传染因子(如甲型流感病毒)且降低该传染因子的生物活性(例如,毒力)的能力。中和的最低要求是该抗体或其结合片段结合该传染因子的能力。在一个实施例中,本发明的抗体或其结合片段免疫特异性地结合甲型流感病毒的至少一个特定表位或抗原决定簇。在一个更具体的实施例中,本发明的抗体或其结合片段免疫特异性地结合甲型流感病毒ha柄蛋白的至少一个特定表位或抗原决定簇。
[0168]
抗体能够在病毒的生命周期过程中的不同点中和传染因子(如甲型流感病毒)的活性。例如,抗体可以通过干扰病毒与一种或多种细胞表面受体的相互作用来干扰该病毒附着至靶细胞。可替代地,抗体可以例如通过干扰经由受体介导的内吞作用进行的病毒内
化来干扰该病毒与其受体的一种或多种附着后相互作用。
[0169]
在一个实施例中,该抗体或其结合片段通过干扰融合过程,例如通过干扰病毒与内体膜的融合来中和甲型流感的活性。在另一个实施例中,该抗体或其结合片段干扰蛋白酶介导的ha0裂解,从而干扰病毒成熟和ha2病毒融合肽的形成。例如,在一个实施例中,该抗体或其结合片段干扰蛋白酶介导的ha0裂解,这是甲型流感病毒的活化所需的。
[0170]
如在此所用,术语“抗体(antibody)”和“抗体(antibodies)”(也被称为免疫球蛋白)涵盖单克隆抗体(包括全长单克隆抗体)、人抗体、人源化抗体、骆驼抗体、嵌合抗体、单链fv(scfv)、单链抗体、单结构域抗体、结构域抗体、fab片段、f(ab')2片段、表现出所希望的生物活性的抗体片段(例如,抗原结合部分)、二硫键连接的fv(dsfv)、以及抗独特型(抗id)抗体(包括(例如)针对本发明的抗体的抗id抗体)、胞内抗体、以及任何以上的表位结合片段。具体地说,抗体包括免疫球蛋白分子和免疫球蛋白分子的免疫活性片段,即,含有至少一个抗原结合位点的分子。免疫球蛋白分子可以具有任何同种型(例如,igg、ige、igm、igd、iga和igy)、亚同种型(例如igg1、igg2、igg3、igg4、iga1和iga2)或同种异型(例如,gm,如g1m(f、z、a或x)、g2m(n)、g3m(g、b、或c),am,em,以及km(1、2或3))。
[0171]
人抗体通常是约150,000道尔顿的异四聚体糖蛋白,由两个相同的轻(l)链和两个相同的重(h)链构成。每条轻链通过一个共价二硫键与一条重链相连接,而二硫键的数目在不同免疫球蛋白同种型的重链之间变化。每条重链和轻链还具有规律性间隔的链内二硫桥。每条重链在一端具有一个可变结构域(vh),之后跟随多个恒定结构域(ch)。每条轻链在一端具有一个可变结构域(vl)并且在其另一端具有一个恒定结构域(cl);轻链的恒定结构域与重链的第一恒定结构域对准,并且轻链的可变结构域与重链的可变结构域对准。基于轻链恒定区的氨基酸序列,轻链被分类为λ链或κ链。κ轻链的可变结构域在此还可以表示为vk。
[0172]
本发明的抗体包括全长或完整抗体、抗体片段(包括抗原结合片段)、天然序列抗体或氨基酸变体,人、人源化、翻译后修饰、嵌合或融合抗体,免疫轭合物、以及其功能性片段。抗体可以在fc区中被修饰以便提供所希望的效应子功能或血清半衰期。如在以下部分中更详细地论述,在具有适当的fc区的情况下,在细胞表面上结合的裸抗体可以诱导细胞毒性:例如经由抗体依赖性细胞毒性(adcc)或通过在补体依赖性细胞毒性(cdc)中募集补体,或通过募集表达一个或多个效应子配体的非特异性细胞毒性细胞,这些效应子配体识别甲型流感病毒ha柄上的结合的抗体并且随后在抗体依赖性细胞介导的吞噬作用(adcp)中引起该细胞的吞噬作用;或一些其他机制。可替代地,在希望消除或减少效应子功能以便于最小化副作用或治疗并发症的情况下,可以使用某些其他fc区。本发明的抗体的fc区可以被修饰来提高对fcrn的结合亲和力并且因此增加血清半衰期。可替代地,fc区可以轭合至peg或白蛋白以便增加血清半衰期,或可以是产生所希望的作用的一些其他轭合作用。
[0173]
本发明的抗甲型流感病毒ha柄抗体适用于诊断、预防、治疗和/或减轻哺乳动物中的甲型流感病毒感染的一种或多种症状。
[0174]
本发明提供一种包含本发明的抗甲型流感病毒ha柄抗体和载体的组合物。出于预防或治疗甲型流感病毒感染的目的,组合物可以被给予至需要这种治疗的患者。本发明还提供包含本发明的抗甲型流感病毒ha柄抗体和载体的配制品。在一个实施例中,该配制品是包含药学上可接受的载体的治疗性配制品。
[0175]
在某些实施例中,本发明提供适用于预防或治疗哺乳动物中的甲型流感感染的方法,这些方法包括向该哺乳动物给予治疗有效量的抗体。可以如医师所指导进行短期(急性地)、慢性地、或间歇性地给予抗体治疗性组合物。
[0176]
在某些实施例中,本发明还提供包含至少一种抗甲型流感病毒ha柄抗体的制品,如无菌剂型和试剂盒。可以提供含有用于例如在elisa或蛋白质印迹中体外检测和定量甲型流感病毒的抗体的试剂盒。适用于检测的这种抗体可以提供有标记,如荧光或放射性标记。
[0177]
术语
[0178]
在详细描述本发明之前,应当了解的是,本发明并不限于特定的组合物或方法步骤,因为这些组合物或方法步骤可以改变。必须注意的是,除非上下文中另外清楚地指出,如在本说明书及所附权利要求中所使用,单数形式“一种/一个(a/an)”和“该”包括复数指示物。
[0179]
除非另外定义,在此所用的所有技术和科学术语具有与本发明所涉及领域的普通技术人员通常所理解的相同的意义。例如,生物医学与分子生物学简明词典(the concise dictionary of biomedicine and molecular biology),juo,pei
‑
show,第二版,2002,crc出版社;细胞与分子生物学词典(the dictionary of cell and molecular biology),第三版,1999,学术出版社(academic press);以及生物化学与分子生物学牛津词典(the oxford dictionary of biochemistry and molecular biology),修订版,2000,牛津大学出版社(oxford university press)为技术人员提供在本发明中使用的许多术语的常用词典。
[0180]
氨基酸可以在此通过它们的通常己知的三字母符号或通过由iupac
‑
iub生物化学命名委员会(iupac
‑
iub biochemical nomenclature commission)推荐的单字母符号来提及。同样地,核苷酸可以通过它们的普遍公认的单字母代码而被提及。
[0181]
除非另有说明,否则抗体的可变结构域、互补决定区(cdr)以及框架区(fr)中的氨基酸编号遵循kabat定义,该定义如列出于卡巴特等人,具有免疫学重要性的蛋白质序列(sequences of proteins of immunological interest),第5版,美国国立卫生研究院,公共卫生事业部,马里兰州贝塞斯达市(1991)。使用这个编号系统,实际的线性氨基酸序列可以含有对应于可变结构域的fr或cdr的缩短或插入该fr或cdr中的较少的或另外的氨基酸。例如,重链可变结构域可以包含在h2的残基52之后的单个氨基酸插入(根据kabat的残基52a)以及在重链fr残基82之后的插入残基(例如,根据kabat的残基82a、82b、以及82c等)。可以通过抗体序列与“标准”kabat编号序列在同源区的比对而针对给定的抗体确定残基的kabat编号。框架残基的最大比对常常需要在编号系统中插入有待用于fv区的“间隔”残基。另外,由于种间或等位基因差异,在任何给定的kabat位点编号处的某些单独残基的身份可以从抗体链到抗体链变化。
[0182]
抗甲型流感病毒ha柄抗体
[0183]
在某些实施例中,这些抗体是分离的和/或纯化的和/或无热原的抗体。如在此使用的术语“纯化”是指已经从其天然环境的组分中鉴别和分离和/或回收的其他分子,例如多肽、核酸分子。因此,在一个实施例中,本发明的抗体是纯化的抗体,其中它们已经与其天然环境的一种或多种组分分离。如在此使用的术语“分离的抗体”是指基本上不含具有不同
抗原特异性的其他抗体分子的抗体(例如,特异性地结合甲型流感病毒ha柄的分离的抗体基本上不含特异性地结合不同于甲型流感病毒ha柄的抗原的抗体)。因此,在一个实施例中,本发明的抗体是分离的抗体,其中它们已经与具有不同特异性的抗体分离。典型地,分离的抗体是单克隆抗体。此外,本发明的分离抗体可以是基本上不含一种或多种其他细胞材料和/或化学品的并且在此被称为分离和纯化抗体。在本发明的一个实施例中,“分离的”单克隆抗体的组合涉及具有不同特异性且组合在明确定义的组合物中的抗体。在下文更详细地描产生和纯化/分离抗体的方法。
[0184]
本发明的分离抗体包含在此披露的由任何适合的多核苷酸编码的抗体氨基酸序列,或任何分离的或配制的抗体。
[0185]
本发明的抗体免疫特异性地结合对甲型流感病毒ha柄蛋白特异的至少一个特定表位。如在此使用的术语“表位”是指能够结合抗体的蛋白质决定簇。表位通常包括如氨基酸或糖侧链的分子的化学活性表面基团,并且通常具有特定的三维结构特征以及特定的电荷特征。构象和非构象表位的区别在于:在变性溶剂存在下,与前者的结合丧失但与后者的结合不丧失。
[0186]
在一个实施例中,该抗体或其结合片段结合在至少h1、h2、h3、h4、h5、h6、h7、h8、h9、h10、h11、h12、h13、h14、h15、h16或h17或所有甲型流感ha亚型之中为保守的表位。在另一个实施例中,该抗体或其结合片段结合在选自h1、h2、h5、h6、h8、h9、h11、h12、h13和h16的一种或多种、或至少1、2、3、4、5、6、7、8、9或10种甲型流感病毒1组亚型以及选自h3、h4、h7、h10、h14和h15的一种或多种、或至少1、2、3、4、5或6种2组亚型之中为保守的表位。
[0187]
在一个实施例中,该抗体或其结合片段以约0.01ug/ml与约5ug/ml之间、或约0.01ug/ml与约0.5ug/ml之间、或约0.01ug/ml与约0.1ug/ml之间、或少于约5ug/ml、1ug/ml、0.5ug/ml、0.1ug/ml或0.05ug/ml的ec
50
结合至少17种h1、h2、h3、h4、h5、h6、h7、h8、h9、h10、h11、h12、h13、h14、h15、h16或h17或所有甲型流感亚型。在另一个实施例中,该抗体或其结合片段以约0.01ug/ml与约5ug/ml之间、或约0.01ug/ml与约0.5ug/ml之间、或约0.01ug/ml与约0.1ug/ml之间、或少于约5ug/ml、1ug/ml、0.5ug/ml、0.1ug/ml或0.05ug/ml的ec
50
结合选自h1、h2、h5、h6、h8、h9、h11、h12、h13和h16的一种或多种、或至少1、2、3、4、5、6、7、8、9或10种甲型流感病毒1组亚型以及选自h3、h4、h7、h10、h14和h15的一种或多种、或至少1、2、3、4、5或6种2组亚型。
[0188]
在一个实施例中,该抗体或其结合片段识别为线性表位抑或连续表位的表位。在另一个实施例中,该抗体或其结合片段识别非线性或构象表位。在一个实施例中,该表位位于ha2的高度保守的柄区中。在一个更具体的实施例中,该抗体结合片段结合ha2的高度保守的柄区中的构象表位。在一个实施例中,该表位包含选自ha2的柄区中的位置18、19、42、45(位置是根据如描述于韦斯(weiss)等人,分子生物学杂志(j.mol.biol.)(1990)212,737
‑
761(1990)中的h3编号系统编号的)的一个或多个氨基酸作为接触残基。在一个更具体的实施例中,该表位包含选自ha2的柄区中的18、19、42和45的一个或多个氨基酸作为接触残基。在另一个实施例中,该表位包含ha2的柄区中的氨基酸18、19、42和45作为接触残基。在又另一个实施例中,该表位包含ha2的柄区中的氨基酸18、19和42作为接触残基。
[0189]
由本发明的抗体或其结合片段识别的一个或多个表位可以具有多种用途。例如,呈纯化或合成形式的表位可以用于产生免疫应答(即,作为疫苗,或用于产生用于其他用途
的抗体)或用于针对与该表位发生免疫反应的抗体筛选血清。在一个实施例中,由本发明的抗体或其结合片段识别的表位或具有这种表位的抗原可以用作用于产生免疫应答的疫苗。在另一个实施例中,本发明的抗体和结合片段可以用于例如通过测定疫苗中的抗原是否包含呈正确构象的正确免疫原性表位来监测疫苗的质量。
[0190]
可变区
[0191]
如在此所用,术语“亲本抗体”是指由用于制备在此定义的变体或衍生物的氨基酸序列编码的抗体。亲本多肽可以包含天然抗体序列(即,天然存在的,包括天然存在的等位基因变体)或天然存在的序列的具有事先存在的氨基酸序列修饰(如其他插入、缺失和/或取代)的抗体序列。亲本抗体可以是人源化抗体或人抗体。在具体实施例中,本发明的抗体是亲本抗体的变体。如在此所用,术语“变体”是指借助在亲本抗体序列中添加、缺失和/或取代一个或多个氨基酸残基而在氨基酸序列上不同于“亲本”抗体氨基酸序列的抗体。
[0192]
抗体的抗原结合部分包含抗体的保留了特异性地结合抗原的能力的一个或多个片段。已经显示抗体的抗原结合功能可以由全长抗体的片段来执行。涵盖在术语抗体的“抗原结合部分”内的结合片段的实例包括:(i)fab片段,其是由vl结构域、vh结构域、cl结构域、以及ch1结构域组成的单价片段;(ii)f(ab')2片段,其是在铰链区包含由二硫桥键连接的两个fab片段的二价片段;(iii)fd片段,其由vh结构域和ch1结构域组成;(iv)fv片段,其由抗体的单臂的vl结构域和vh结构域组成;(v)dab片段(华德(ward)等人,(1989)自然(nature)341:544
‑
546),其由vh结构域组成;以及(vi)分离的互补决定区(cdr)。此外,尽管fv片段的两个结构域vl和vh是由单独的基因编码的,但是它们可以使用重组方法通过合成连接子来接合,该合成连接子使它们能够被制备为单一蛋白链,其中vl区和vh区配对以便形成单价分子(被称为单链fv(scfv);参见例如,博尔德(bird)等人(1988)科学(science)242:423
‑
426;和休斯顿(huston)等人(1988)美国科学院院刊(proc.natl.acad.sci.usa)85:5879
‑
5883)。这类单链抗体还旨在涵盖在术语抗体的“抗原结合部分”内。这些抗体片段是使用本领域的技术人员已知的常规技术获得的,并且以与完整抗体相同的方式针对效用来筛选这些片段。可以通过重组dna技术或通过完整免疫球蛋白的酶裂解或化学裂解来产生抗原结合部分。
[0193]
本发明的抗体包含至少一个抗原结合结构域,该抗原结合结构域包含在此所述的vh和vl结构域。
[0194]
在某些实施例中,纯化抗体包含与表1中披露的vh和/或vl序列中的至少一个具有给定一致性百分比的vh和/或vl。如在此所用,术语“序列一致性百分比(%)”(还包括“同源性”)被定义为在比对序列并且(必要时)引入空位以便获得最大的序列一致性百分比并且不将任何保守取代视为序列一致性的部分之后,在候选序列中的氨基酸残基或核苷酸与在参考序列(如亲本抗体序列)中的氨基酸残基或核苷酸一致的百分比。除了手动之外,用于比较的最佳序列比对可以通过以下各项来产生:史密斯(smith)和沃特曼(waterman)的局部同源性算法,1981,应用数学进展(ads app.math.)2,482;尼德曼(needleman)和翁施(wunsch)的局部同源性算法,1970,分子生物学杂志48,443;皮尔森(pearson)和利普曼(lipman)的相似性搜索方法,1988,美国科学院院刊85,2444;或使用这些算法的计算机程序(威斯康星遗传学软件包(wisconsin genetics software package),遗传学计算机组,575大学道,麦迪逊,威斯康星州中的gap、bestfit、fasta、blast p、blast n和tfasta)。
[0195]
本发明的抗体可以包含与在此所述的vh氨基酸序列具有至少65%、70%、75%、80%、85%、90%、95%或具有100%一致性的vh氨基酸序列。这些抗体可以具有与在此所述的vh氨基酸序列的氨基酸序列具有至少90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或具有100%一致性的vh氨基酸序列。
[0196]
本发明的抗体可以包含与在此所述的vl氨基酸序列具有至少65%、70%、75%、80%、85%、90%、95%或具有100%一致性的vl氨基酸序列。这些抗体可以具有与在此所述的vl氨基酸序列具有至少90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或具有100%一致性的vl氨基酸序列。
[0197]
在本发明的范围内的抗体能够中和甲型流感病毒的一种或多种1组亚型以及一种或多种2组亚型,如在此所描述。
[0198]
互补决定区(cdr)
[0199]
虽然可变结构域(vh和vl)包含抗原结合区;但是可变性不是均匀地分布于抗体的这些可变结构域内的。它被集中在轻链(vl或vk)和重链(vh)可变结构域两者中的称为互补决定区(cdr)的区段中。这些可变结构域的更高保守性的部分被称为框架区(fr)。天然重链和轻链的可变结构域各自包含由三个cdr连接的四个fr,这四个fr大体上采用β
‑
片层构型,这三个cdr形成连接β
‑
片层结构的环并且在一些情况下形成β
‑
片层结构的一部分。每条链中的cdr由fr紧密靠近地保持在一起,与来自另一条链的cdr促成了抗体的抗原结合位点的形成(参见卡巴特等人,上文)。重链的三个cdr被指定为cdr
‑
h1、cdr
‑
h2和cdr
‑
h3,并且轻链的三个cdr被指定为cdr
‑
l1、cdr
‑
l2和cdr
‑
l3。在此使用kabat编号系统。因而,cdr
‑
h1在大致氨基酸31处(即,在第一个半胱氨酸残基之后大致9个残基处)开始,包括大致5
‑
7个氨基酸,并且在下一个酪氨酸残基处结束。cdr
‑
h2在cdr
‑
h1末端之后的第十五个残基处开始,包括大致16
‑
19个氨基酸,并且在下一个精氨酸或赖氨酸残基处结束。cdr
‑
h3在cdr
‑
h2末端之后的大致第三十三个氨基酸残基处开始;包括3
‑
25个氨基酸;并且在序列w
‑
g
‑
x
‑
g处结束,其中x是任何氨基酸。cdr
‑
l1在大致残基24(即,在半胱氨酸残基之后)处开始;包括大致10
‑
17个残基;并且在下一个酪氨酸残基处结束。cdr
‑
l2在cdr
‑
l1末端之后大致第十六个残基处开始,并且包括大致7个残基。cdr
‑
l3在cdr
‑
l2末端之后的大致第三十三个残基处开始;包括大致7
‑
11个残基并且在序列f
‑
g
‑
x
‑
g处结束,其中x是任何氨基酸。注意cdr在抗体间变化极大(并且通过定义将不展现与kabat共有序列的同源性)。
[0200]
本发明涵盖中和抗甲型流感ha柄抗体,这些抗体包含与在此所述的氨基酸序列基本相同的序列中的氨基酸。与在此所述的这些序列基本相同的氨基酸序列包括以下序列,这些序列包含例如抗体11、抗体12、抗体13、抗体14或抗体15的氨基酸序列中或seq id no:102、112、122、132或142中所示的氨基酸序列中的保守氨基酸取代以及氨基酸缺失和/或插入。保守氨基酸取代是指第一氨基酸被第二氨基酸置换,该第二氨基酸具有与该第一氨基酸的化学和/或物理性质相似的化学和/或物理性质(例如,电荷、结构、极性、疏水性/亲水性)。保守取代包括以下组内的一个氨基酸被另一个氨基酸置换:赖氨酸(k)、精氨酸(r)和组氨酸(h);天冬氨酸(d)和谷氨酸(e);天冬酰胺(n)、谷氨酰胺(q)、丝氨酸(s)、苏氨酸(t)、酪氨酸(y)、k、r、h、d、以及e;丙氨酸(a)、缬氨酸(v)、亮氨酸(l)、异亮氨酸(i)、脯氨酸(p)、苯丙氨酸(f)、色氨酸(w)、蛋氨酸(m)、半胱氨酸(c)、以及甘氨酸(g);f、w和y;c、s和t。
[0201]
框架区
[0202]
重链和轻链的可变结构域各自包含四个框架区(fr1、fr2、fr3、fr4),这些框架区是可变结构域的更高保守性的部分。重链的四个fr被指定为fr
‑
h1、fr
‑
h2、fr
‑
h3、以及fr
‑
h4,并且轻链的四个fr被指定为fr
‑
l1、fr
‑
l2、fr
‑
l3、以及fr
‑
l4。在此使用kabat编号系统,参见表1,卡巴特等人,同上。因而,fr
‑
h1开始于位置1并且在大致氨基酸30处结束,fr
‑
h2大致是从氨基酸36至49,fr
‑
h3大致是从氨基酸66至94,并且fr
‑
h4大致是氨基酸103至113。fr
‑
l1开始于氨基酸1并且在大致氨基酸23处结束,fr
‑
l2大致是从氨基酸35至49,fr
‑
l3大致是从氨基酸57至88,并且fr
‑
l4大致是从氨基酸98至107。在某些实施例中,框架区可以包含根据kabat编号系统的取代,例如在fr
‑
l1中106a处的插入。除了天然存在的取代之外,fr残基的一种或多种改变(例如,取代)也可以被引入本发明的抗体中,只要它保留中和能力。在某些实施例中,这些改变产生针对甲型流感病毒ha柄的抗体的结合亲和力的改进或优化。用于修饰的框架区残基的实例包括直接非共价结合抗原的那些(阿密特(amit)等人,科学(science),233:747
‑
753(1986));与cdr相互作用/实现cdr的构象的那些(乔西亚(chothia)等人,分子生物学杂志,196:901
‑
917(1987));和/或参与vl
‑
vh界面的那些(美国专利号5,225,539)。
[0203]
在另一个实施例中,出于“种系化(germlining)”的目的,fe可以包含一个或多个氨基酸变化。例如,将所选择的抗体重链和轻链的氨基酸序列与种系重链和轻链氨基酸序列进行比较,并且其中所选择的vl链和/或vh链的某些框架残基不同于种系构型(例如,由于用来制备噬菌体文库的免疫球蛋白基因的体细胞突变),可能希望的是使所选择抗体的改变的框架残基“回复突变”为种系构型(即,改变所选择的抗体的框架氨基酸序列,这样使得它们与种系框架氨基酸序列相同)。框架残基的这种“回复突变”(或“种系化”)可以通过用于引入特定突变的标准分子生物学方法(例如,定点诱变;pcr介导的诱变,等等)来完成。
[0204]
编码本发明的抗体的核苷酸序列
[0205]
除了以上所述的氨基酸序列之外,本发明进一步提供对应于这些氨基酸序列并且编码本发明的人抗体的核苷酸序列。在一个实施例中,本发明提供多核苷酸,这些多核苷酸包含编码在此所述的抗体或其片段的核苷酸序列。这些包括但不限于编码以上提及的氨基酸序列的核苷酸序列。因此,本发明还提供编码包含在此所述的抗体的cdr和fr的vh和vl框架区的多核苷酸序列以及用于它们在细胞(例如哺乳动物细胞)中有效表达的表达载体。使用多核苷酸制备抗体的方法在下文更详细地描述。
[0206]
本发明还涵盖在严格或较低严格性杂交条件(例如,如在此所定义)下与编码在此所述的本发明的抗体的多核苷酸杂交的多核苷酸。如在此使用的术语“严格性”是指杂交实验的实验条件(例如,温度和盐浓度),指示在探针与过滤器结合的核酸之间的同源性的程度;严格性越高,探针与过滤器结合的核酸之间的同源性百分比越高。
[0207]
严各杂交条件包括但不限于:在6x氯化钠/柠檬酸钠(ssc)中、在大约45℃下杂交至过滤器结合的dna,随后在大约50℃
‑
65℃下在0.2x ssc/0.1%sds中洗涤一次或多次;高度严格条件如在6x ssc中、在大约45℃下杂交至过滤器结合的dna,随后在大约65℃下在0.1x ssc/0.2%sds中洗涤一次或多次;或者本领域的技术人员已知的任何其他严格杂交条件(参见例如,奥苏贝尔(ausubel)f.m.等人编辑,1989当代分子生物学实验指南(current protocols in molecular biology),第1卷,格林出版协会公司(green publishing associates,inc.)和约翰威利父子出版公司(john wiley and sons,inc.),
纽约,第6.3.1至6.3.6页和第2.10.3页)。
[0208]
基本上一致的序列可以是多态性序列,即种群中的可替代序列或等位基因。等位基因差异可以小至一个碱基对。基本上一致的序列还可以包含诱变序列,包括包含沉默突变的序列。突变可以包含一个或多个残基改变、一个或多个残基的缺失、或一个或多个另外残基的插入。
[0209]
可以通过本领域中已知的任何方法获得多核苷酸并且测定多核苷酸的核苷酸序列。例如,如果抗体的核苷酸序列是已知的,那么编码该抗体的多核苷酸可以从化学合成的寡核苷酸来组装(例如,如库特梅尔(kutmeier)等人,生物技术(biotechniques)17:242(1994)中所描述),简单地说,这涉及合成含有编码该抗体的序列的部分的重叠寡核苷酸,退火并且连接那些寡核苷酸,并且然后通过pcr来扩增所连接的寡核苷酸。
[0210]
编码抗体的多核苷酸还可以从来自适合来源的核酸产生。如果含有编码具体抗体的核酸的克隆是不可获得的,但抗体分子的序列是已知的,则编码免疫球蛋白的核酸可以化学合成,或通过pcr扩增使用与序列的3'和5'端可杂交合成引物,或通过使用对具体基因序列具有特异性的寡核苷酸探针以便鉴别例如来自cdna文库的编码抗体的cdna克隆来进行克隆,而从适合的来源(例如,抗体cdna文库、或从表达抗体的任何组织或细胞产生的cdna文库、或从这些组织或细胞分离的核酸(优选polya+rna),这些细胞如被选择为表达抗体的杂交瘤细胞)获得。接着可以使用本领域中熟知的任何方法将通过pcr产生的扩增核酸克隆至可复制的克隆载体中。
[0211]
一旦确定抗体的核苷酸序列和相应氨基酸序列,就可以使用本领域中熟知用于操纵核苷酸序列的方法,例如重组dna技术、定点诱变、pcr等(参见,例如以下文献中所描述的技术:萨布鲁克(sambrook)等人,1990,分子克隆:实验手册(molecular cloning,a laboratory manual),第2版,冷泉港实验室(cold spring harbor laboratory),冷泉港(cold spring harbor),纽约;和奥苏贝尔等人编辑,1998,当代分子生物学实验指南,约翰&威利父子公司,纽约)来对抗体的核苷酸序列进行操纵以产生具有不同氨基酸序列的抗体,例如以产生氨基酸取代、缺失和/或插入。
[0212]
结合特征
[0213]
如上所述,本发明的抗甲型流感病毒ha柄抗体专有地抑或相对于其他多肽优先地免疫特异性地结合甲型流感病毒ha柄蛋白、肽、亚基、片段、部分或其任何组合的至少一个特定表位或抗原决定簇。如在此使用的术语“表位”或“抗原决定簇”是指能够结合抗体的蛋白质决定簇,其中在此的术语“结合”优选地涉及特异性结合。这些蛋白质决定簇或表位通常由分子(如氨基酸或糖侧链)的化学活性表面基团组成,并且通常具有特定的三维结构特征以及特定的电荷特征。构象和非构象表位的区别在于:在变性溶剂存在下,与前者的结合丧失但与后者的结合不丧失。如在此使用的术语“非连续表位”是指在从蛋白质的一级序列中的至少两个分开的区域形成的蛋白质抗原上的构象表位。
[0214]
抗原与抗体之间的相互作用与其他非共价蛋白质
‑
蛋白质相互作用相同。一般来说,抗原与抗体之间存在四种类型的结合相互作用:(i)氢键,(ii)分散力,(iii)路易斯酸与路易斯碱之间的静电力以及(iv)疏水相互作用。疏水相互作用是抗体
‑
抗原相互作用的主要驱动力,并且是基于经由非极性基团的水的排斥而不是分子的吸引(丹佛(tanford),1978)。然而,某些物理力也促成抗原
‑
抗体结合,例如表位形状与不同抗体结合位点的配合
或互补。此外,其他材料和抗原可以与抗体发生交叉反应,由此竞争可获得的游离抗体。
[0215]
抗原与抗体之间的结合的亲和力常数和特异性的测量是确定使用本发明的抗体的预防、治疗、诊断或研究方法的功效的关键要素。“结合亲和力”通常是指分子(例如,抗体)的单一结合位点与其结合配偶体(例如,抗原)之间的非共价相互作用的总和的强度。除非另外指明,否则如在此所用,“结合亲和力”是指反映结合对的成员(例如,抗体与抗原)之间的1:1相互作用的固有结合亲和力。分子x对其配偶体y的亲和力通常可以由平衡解离常数(kd)表示,计算为比率k
off
/k
on
。参见,例如陈,y.(chen,y.)等人,(1999)分子生物学杂志293:865
‑
881。可以通过本领域中已知的常见方法来测量亲和力,包括在此描述和例举的那些方法。用于动力学表征的可商购的系统的一个实例包括仪器家族。低亲和力抗体通常缓慢地结合抗原并且倾向于容易解离,而高亲和力抗体通常更快地结合抗原并且倾向于更长地保持结合。测量结合亲和力的多种方法是本领域中已知的,其中的任何方法均可以用于本发明的目的。
[0216]
结合亲和力的确定可以使用在实例部分中进一步描述的特定技术以及本领域中熟知的方法来测量。这样一种方法包括通过用感兴趣的抗体的fab型式以及它的抗原来进行的放射性标记抗原结合测定(ria)而测量解离常数“kd”,该放射性标记抗原结合测定如由以下测定所描述,该测定通过在未标记抗原的滴定系列存在下平衡fab与(
125
i)
‑
标记抗原的最小浓度、然后用抗fab抗体包被的板捕获结合的抗体而测量fab对抗原的溶液结合亲和力(陈等人,(1999)分子生物学杂志293:865
‑
881)。为了建立该测定的条件,将微量滴定板(dynex)用在50mm碳酸钠(h 9.6)中的5μg/ml捕获性抗fab抗体(cappel labs)包被过夜,并且随后在室温(大致23℃)下用pbs中的2%(w/v)牛血清白蛋白封闭二至五个小时。在非吸附板(nunc#269620)中,将100pm或26pm[
125
i]
‑
抗原与感兴趣的fab的连续稀释液进行混合(例如,与普雷斯塔(presta)等人,(1997)癌症研究(cancer res.)57:4593
‑
4599中的抗vegf抗体fab
‑
12的评定一致)。然后将感兴趣的fab孵育过夜;然而,孵育可以连续持续较长的时期(例如,65小时)以便确保达到平衡。此后,将混合物转移至捕获板,以用于在室温下孵育(例如,持续一小时)。然后将溶液去除并且将板用在pbs中的0.1%吐温20(tween
‑
20)洗涤八次。当这些板已经干燥时,添加150μl/孔的闪烁剂(microscint
‑
20;帕卡德(packard)),并且将这些板在topcountγ计数器(帕卡德)上进行计数持续十分钟。选择给出小于或等于20%的最大结合的各fab的浓度以供在竞争性结合测定中使用。
[0217]
在另一种情况下,可以通过使用表面等离子体共振测定来测量kd值,该测定使用biacore
tm
‑
2000或biacore
tm
‑
3000(biacore公司,皮斯卡塔韦(piscataway),新泽西州)在25℃下用在约10个反应单位(ru)的固定抗原cm5芯片进行。简言之,根据供应商的说明书,将羧甲基化的葡聚糖生物传感器芯片(cm5,biacore公司)用n
‑
乙基
‑
n'
‑
(3
‑
二甲基氨基丙基)
‑
碳二亚胺盐酸盐(edc)以及n
‑
羟基琥珀酰亚胺(nhs)活化。将抗原用110mm乙酸钠(ph 4.8)稀释为5ug/ml(约0.2um),之后以5ul/分钟的流速注入以便获得偶联蛋白的大致10个反应单位(ru)。在注入抗原后,注入1m乙醇胺以便封闭未反应的基团。为了动力学测量,在25℃下将fab的两倍连续稀释液(0.78nm至500nm)以大致25ul/分钟的流速注入具有0.05%吐温20的pbs(pbst)中。通过同时拟合缔合和解离传感图,使用简单的一对一朗缪尔结合模型(biacore评估软件3.2版)来计算缔合速率(k
on
)和解离速率(k
off
)。
[0218]
如果通过以上的表面等离子体共振测定缔合速率超过106m
‑1s
‑1,则可以通过使用
荧光淬灭技术来测定缔合速率,该技术在25℃下在增加浓度的抗原存在下测量pbs(ph 7.2)中的20nm抗
‑
抗原抗体(fab形式)的荧光发射强度(激发=295nm;发射=340nm,16nm带通)的增加或降低,如在分光光度计中所测量,该分光光度计如配备有停止
‑
流动的分光光度计(aviv仪器公司)或具有搅拌红色小池的8000
‑
系列slm
‑
aminco分光光度计(thermospectronic)。根据本发明的“缔合速率(on
‑
rate或rate of association或association rate)”或“k
on”也可以使用如上所述的biacore
tm
‑
2000或biacore
tm
‑
3000(biacore公司,皮斯卡塔韦,新泽西州)用以上所述的相同的表面等离子体共振技术来测定。
[0219]
适用于确定本发明的抗体或其改变/突变的衍生物(以下讨论)的结合特征的方法和试剂是本领域中已知的和/或是可商购的(美国专利号6,849,425;6,632,926;6,294,391;6,143,574)。此外,被设计用于这类动力学分析的设备和软件是可商购的(例如a100和仪器;biacore international ab公司,乌普萨拉(uppsala),瑞典)。
[0220]
在一个实施例中,本发明的抗体(包括其结合片段或变体)还可以在它们对甲型流感病毒多肽的结合亲和力方面进行描述或说明。典型地,具有高亲和力的抗体具有小于10
‑7m的kd。在一个实施例中,抗体或其结合片段结合甲型流感多肽或其片段或变体的解离常数或kd小于或等于5
×
10
‑7m、10
‑7m、5
×
10
‑8m、10
‑8m、5
×
10
‑9m、10
‑9m、5
×
10
‑
10
m、10
‑
10
m、5
×
10
‑
11
m、10
‑
11
m、5
×
10
‑
12
m、10
‑
12
m、5
×
10
‑
13
m、10
‑
13
m、5
×
10
‑
14
m、10
‑
14
m、5
×
10
‑
15
m或10
‑
15
m。甲型流感多肽可以包括ha多肽。在一个更具体的实施例中,抗体或其结合片段结合甲型流感多肽或其片段或变体的解离常数或kd小于或等于5
×
10
‑
10
m、10
‑
10
m、5
×
10
‑
11
m、10
‑
11
m、5
×
10
‑
12
m或10
‑
12
m。本发明涵盖结合甲型流感多肽的解离常数或kd在任何单独列举的值之间的范围内的抗体。
[0221]
在另一个实施例中,本发明的抗体或其结合片段结合甲型流感多肽或其片段或变体的解离速率(k
off
)小于或等于5
×
10
‑2sec
‑1、10
‑2sec
‑1、5
×
10
‑3sec
‑1、或10
‑3sec
‑1、5
×
10
‑4sec
‑1、10
‑4sec
‑1、5
×
10
‑5sec
‑1、或10
‑5sec
‑1、5
×
10
‑6sec
‑1、10
‑6sec
‑1、5
×
10
‑7sec
‑1或10
‑7sec
‑1。在一个更具体的实施例中,本发明的抗体或其结合片段结合甲型流感多肽或其片段或变体的解离速率(k
off
)小于或等于5
×
10
‑4sec
‑1、10
‑4sec
‑1、5
×
10
‑5sec
‑1、或10
‑5sec
‑1、5
×
10
‑6sec
‑1、10
‑6sec
‑1、5
×
10
‑7sec
‑1、或10
‑7sec
‑1。本发明还涵盖结合甲型流感多肽的解离速率(k
off
)在任何单独列举的值之间的范围内的抗体。
[0222]
在另一个实施例中,本发明的抗体或其结合片段结合甲型流感多肽或其片段或变体的缔合速率(k
on
)大于或等于103m
‑1sec
‑1、5
×
103m
‑1sec
‑1、104m
‑1sec
‑1、5
×
104m
‑1sec
‑1、105m
‑1sec
‑1、5
×
105m
‑1sec
‑1、106m
‑1sec
‑
1、5
×
106m
‑1sec
‑1、107m
‑1sec
‑
1、或5
×
107m
‑1sec
‑1。在一个更具体的实施例中,本发明的抗体或其结合片段结合甲型流感多肽或其片段或变体的缔合速率(k
on
)大于或等于105m
‑1sec
‑1、5
×
105m
‑1sec
‑1、106m
‑1sec
‑
1、5
×
106m
‑1sec
‑1、107m
‑1sec
‑1或5
×
107m
‑1sec
‑1。本发明涵盖结合甲型流感多肽的缔合速率(k
on
)在任何单独列举的值之间的范围内的抗体。
[0223]
在一个实施例中,可以将结合测定作为直接结合测定抑或作为竞争结合测定而进行。可以使用标准elisa或标准流式细胞术测定来检测结合。在直接结合测定中,测试了候选抗体与其同源抗原的结合。在另一方面,竞争结合测定评定了候选抗体与已知的抗体或
结合甲型流感病毒ha柄的其他化合物竞争的能力。一般来说,允许抗体与能够被检测的甲型流感病毒ha柄结合的任何方法被涵盖在用于检测和测量抗体的结合特征的本发明的范围内。本领域的技术人员将认识到这些熟知的方法并且出于这一原因未在此详细地提供。还利用这些方法来筛选提供所希望的特征的那些的一组抗体。
[0224]
本发明的抗体免疫特异性地结合甲型流感病毒ha柄并且能够中和甲型流感病毒感染。中和测定可以如在此实例部分中所描述或使用本领域中已知的其他方法来进行。术语“50%抑制浓度”(缩写为“ic
50”)表示用于甲型流感病毒的50%中和所需的抑制剂(例如,本发明的抗体)的浓度。本领域的普通技术人员将了解,更低的ic
50
值对应于更有效的抑制剂。
[0225]
在一个实施例中,根据本发明的抗体或其结合片段在微量中和测定中具有表示为50%抑制浓度(ic
50 ug/ml)的在从约0.01ug/ml至约50ug/ml范围内、或在从约0.01ug/ml至约5ug/ml的抗体范围内、或在从约0.01ug/ml至约0.1ug/ml的用于中和甲型流感病毒的抗体范围内的中和效力。在在此所述的微量中和测定中使用的抗体的最高浓度是50ug/ml。本发明的抗体的高效力意味着较低浓度的抗体可以用于获得甲型流感病毒的50%中和。
[0226]
在某些实施例中,本发明的抗体可以诱导细胞死亡。“诱导细胞死亡”的抗体是引起活细胞变得无活力的抗体。体外细胞死亡可以在不存在补体和免疫效应细胞的情况下来测定以便区分由抗体依赖性细胞介导的细胞毒性(adcc)或补体依赖性细胞毒性(cdc)诱导的细胞死亡。因此,用于细胞死亡的测定可以使用热灭活的血清(即,在不存在补体的情况下)且在不存在免疫效应细胞的情况下进行。为了测定抗体是否能够诱导细胞死亡,可以相对于未处理的细胞评定膜完整性的损失,如通过碘化丙啶(pi)、台盼蓝(参见摩尔(moore)等人,细胞技术学(cytotechnology)17:1
‑
11(1995))、7aad的摄取或本领域中熟知的其他方法所评价。
[0227]
在一个具体实施例中,本发明的抗体可以经由细胞凋亡诱导细胞死亡。“诱导细胞凋亡”的抗体是诱导程序性细胞死亡的抗体,如通过膜联蛋白v的结合、dna断裂、细胞收缩、内质网的扩张、细胞破碎和/或膜囊泡(称为凋亡小体)的形成所测定。不同方法可供用于评价与细胞凋亡相关的细胞事件。例如,可以通过膜联蛋白结合测量磷酯酰丝氨酸(ps)易位;可以通过dna梯化(laddering)评价dna断裂;并且可以通过亚二倍体细胞的任何增加来评价核/染色质凝聚连同dna断裂。优选地,诱导细胞凋亡的抗体是在膜联蛋白结合测定中相对于未处理的细胞产生约2至50倍、优选地约5至50倍、并且最优选地约10至50倍膜联蛋白结合诱导的抗体。
[0228]
在另一个具体实施例中,本发明的抗体可以经由抗体依赖性细胞毒性(adcc)和/或补体依赖性细胞介导的细胞毒性(cdc)和/或抗体依赖性细胞介导的吞噬作用(adcp)来诱导细胞死亡。人igg1亚类抗体的adcc活性和cdc活性的表达通常涉及抗体的fc区结合至存在于效应细胞(如杀伤细胞、自然杀伤细胞或活化的巨噬细胞)表面上的针对抗体的受体(下文称为“fcγr”)。不同的补体组分可以被结合。关于结合,已经表明抗体的铰链区和c区的第二结构域(下文称为“cγ2结构域”)中的若干氨基酸残基是重要的(欧洲免疫学杂志(eur.j.immunol.),23,1098(1993);免疫学(immunology),86,319(1995);化学免疫学(chemical immunology),65,88(1997)),并且cγ2结构域中的糖链(化学免疫学,65,88(1997))也是重要的。
[0229]
为了评定感兴趣的抗体的adcc活性,可以使用体外adcc测定,如美国专利号5,500,362中描述的测定。该测定还可以使用可商购的试剂盒,例如cytotox 96(普洛麦格公司(promega))来进行。用于这类测定的有用的效应细胞包括但不限于,外周血单核细胞(pbmc)、自然杀伤(nk)细胞、以及nk细胞系。表达转基因fc受体(cd16)和相关信号传导多肽(例如fc
ε
ri
‑
γ)的nk细胞系也可以充当效应细胞(wo 2006/023148)。例如,可以测定任何特定抗体通过补体活化和/或adcc介导细胞溶解的能力。在体外生长且标记感兴趣的细胞;将抗体与免疫细胞组合添加至细胞培养物,这些免疫细胞可以通过抗原抗体复合物活化;即,,效应细胞参与adcc应答。还可以针对补体活化对抗体进行测试。在任一情况下,通过标记从溶解的细胞的释放来检测细胞溶解。细胞溶解的程度还可以通过检测细胞质蛋白(例如ldh)至上清液中的释放来测定。事实上,可以使用患者自己的血清作为补体和/或免疫细胞的来源来筛选抗体。能够在体外测试中介导人adcc的抗体随后可以治疗性地用于该特定患者。感兴趣的分子的adcc活性也可以例如在如克莱因斯(clynes)等人,美国国家科学院院刊(美国)95:652
‑
656(1998)中披露的动物模型中进行体内评定。此外,用于调节(即,增加或降低)抗体的adcc和任选地cdc活性的水平的技术是本领域中熟知的(例如,美国专利号5,624,821;6,194,551;7,317,091)。本发明的抗体可能能够诱导adcc和/或cdc,或者可能已经进行修饰以便具有诱导adcc和/或cdc的能力。用于确定adcc功能的测定可以使用人效应细胞来实施以便评定人adcc功能。这类测定还可以包括旨在筛选通过坏死和/或凋亡机制诱导、介导、增强、阻断细胞死亡的抗体的那些测定。包括利用活性染料的测定的这类方法、检测和分析半胱天冬酶的方法、以及测量dna断裂的测定可以用于评定在体外用感兴趣的抗体培养的细胞的凋亡活性。
[0230]
抗体的产生
[0231]
以下描述用于产生适用于本发明的抗体的示例性技术。
[0232]
单克隆抗体
[0233]
可以使用本领域中已知的各种各样的技术来制备单克隆抗体,这些技术包括使用杂交瘤(科勒(kohler)等人,自然,256:495(1975);哈洛(harlow)等人,抗体:实验室手册(antibodies:a laboratory manual),(冷泉港实验室出版社,第2版1988);哈默林(hammerling)等人,在:单克隆抗体和t细胞杂交瘤(monoclonal antibodies and t
‑
cell hybridomas)563
‑
681(爱思唯尔(elsevier)出版公司,纽约,1981)中)、重组、以及噬菌体展示技术、或其组合。如在此所用的术语“单克隆抗体”是指从基本上同种或分离的抗体的群体中获得的抗体,例如单独的抗体,这些抗体包含除了可能天然存在的突变(可能以少量存在)之外的相同的群体。单克隆抗体是高特异性的,针对单一抗原位点的。此外,与包含针对不同决定簇(表位)的不同抗体的多克隆抗体制剂相比,每种单克隆抗体是针对该抗原上的相同决定簇的。除了它们的特异性之外,单克隆抗体的有利之处在于它们可以在不被其他抗体污染的情况下合成。修饰词“单克隆”不应被理解为需要通过任何特定的方法而产生抗体。以下是用于产生单克隆抗体的代表性方法的描述(不是旨在为限制性的),并且这些方法可以用来产生例如单克隆哺乳动物抗体、嵌合抗体、人源化抗体、人抗体、结构域抗体、双抗体(diabody)、疫苗体(vaccibody)、线性和多特异性抗体。
[0234]
杂交瘤技术
[0235]
使用杂交瘤技术产生和筛选特异性抗体的方法在本领域中是常规的并且是众多
周知的。在杂交瘤方法中,小鼠或其他适当的宿主动物(如仓鼠)如上所述被免疫以便引出淋巴细胞,这些淋巴细胞产生或能够产生将特异性地结合用于免疫的抗原的抗体。可替代地,淋巴细胞可以被体外免疫。在免疫之后,分离淋巴细胞,并且然后使用适合的融合剂或融合配偶体(如聚乙二醇)与骨髓瘤细胞系融合从而形成杂交瘤细胞(戈丁(goding),单克隆抗体:原理与实践(monoclonal antibodies:principles and practice),第59
‑
103页(学术出版社(academic press),1986))。在某些实施例中,所选择的骨髓瘤细胞是有效融合、支持由所选择的抗体产生细胞的稳定高水平抗体产生、并且对针对未融合的亲代细胞而选择的选择性培养基敏感的那些骨髓瘤细胞。在一方面,骨髓瘤细胞系是鼠类骨髓瘤系,如来源于从美国加利福尼亚州圣地亚哥市的索尔克研究所细胞分配中心(salk institute cell distribution center,san diego,california,usa)可获得的mopc
‑
21和mpc
‑
11小鼠肿瘤的、以及从美国马里兰州罗克维尔市的美国典型培养物保藏中心(american type culture collection,rockville,maryland,usa)可获得的sp
‑
2和衍生物(例如x63
‑
ag8
‑
653细胞)的那些。也已经描述了人骨髓瘤和小鼠
‑
人杂骨髓瘤细胞系用于产生人单克隆抗体(科兹博(kozbor),免疫学杂志(j.immunol.),133:3001(1984);和布罗德尔(brodeur)等人,单克隆抗体生产技术和应用(monoclonal antibody production techniques and applications),第51
‑
63页(马塞尔德克尔公司(marcel dekker,inc.),纽约,1987))。
[0236]
一旦产生具有所希望的特异性、亲和力、和/或活性的抗体的杂交瘤细胞被鉴别出,就可以通过有限稀释程序而将这些克隆进行亚克隆并且通过标准方法使其生长(戈丁,同上)。用于这一目的的适合的培养基包括例如d
‑
mem或rpmi
‑
1640培养基。另外,杂交瘤细胞可以在动物中体内生长为腹水瘤,例如通过将细胞腹膜内(i.p.)注射至小鼠中。
[0237]
将由亚克隆分泌的单克隆抗体通过常规抗体纯化程序(例如像,亲和色谱(例如使用蛋白a或蛋白g
‑
琼脂糖凝胶)或离子交换色谱、亲和标签、羟基磷灰石色谱、凝胶电泳、渗析等)而从培养基、腹水液、或血清适当地分离。示例性纯化方法在下文进行更详细地描述。
[0238]
重组dna技术
[0239]
使用重组dna技术产生和筛选特异性抗体的方法在本领域中是常规的并且是众所周知的(例如美国专利号4,816,567)。可以容易地使用常规程序(例如,通过使用能够特异性地结合至编码鼠抗体的重链和轻链的基因的寡核苷酸探针)来将编码单克隆抗体的dna分离和/或测序。一旦分离,可以将dna置于表达载体中,然后将其转染至宿主细胞中,如大肠杆菌细胞、猴cos细胞、中国仓鼠卵巢(cho)细胞、或不以另外的方式产生抗体蛋白的骨髓瘤细胞,从而在重组宿主细胞中获得单克隆抗体的合成。关于在细菌中重组表达编码抗体的dna的文献综述包括斯盖拉(skerra)等人,免疫学当前观点(curr.opinion in immunol.),5:256
‑
262(1993)和普吕克通(pluckthun),免疫学评论(immunol.revs.),130:151
‑
188(1992)。如下对于通过噬菌体展示和抗体人源化所产生的抗体所描述,可以从除了杂交瘤之外的产生本发明的抗体的一种或多种来源获得用于重组抗体的dna或基因材料。
[0240]
抗体或其变体的重组表达通常需要构建含有编码抗体的多核苷酸的表达载体。因此,本发明提供可复制的载体,这些载体包含可操作地连接至启动子的,编码抗体分子、抗体的重链或轻链、抗体的重链或轻链可变结构域或其一部分、或者重链或轻链cdr的核苷酸序列。这类载体可以包含编码抗体分子的恒定区的核苷酸序列(参见例如,美国专利号5,981,216;5,591,639;5,658,759和5,122,464),并且可以将抗体的可变结构域克隆至这种
载体中以用于表达整个重链、整个轻链或整个重链和轻链两者。
[0241]
一旦通过常规技术将表达载体转移至宿主细胞中,然后就通过常规技术培养转染的细胞以便产生抗体。因此,本发明包括可操作地连接至异源启动子的含有编码本发明的抗体或其片段、或其重链或轻链、或其部分、或本发明的单链抗体的多核苷酸的宿主细胞。在用于表达双链抗体的某些实施例中,可以将编码重链和轻链的载体在宿主细胞中共表达,以用于表达整个免疫球蛋白分子,如下所详述。
[0242]
作为用于表达重组抗体的宿主可获得的哺乳动物细胞系是在本领域中已知的,并且包括许多可从美国典型培养物保藏中心(atcc)获得的永生化细胞系,包括但不限于中国仓鼠卵巢(cho)细胞、海拉细胞、幼仓鼠肾(bhk)细胞、猴肾细胞(cos)、人肝细胞癌细胞(例如,hep g2)、人上皮肾293细胞、以及多种其他细胞系。不同的宿主细胞具有针对蛋白质和基因产物的翻译后加工和修饰的特征性和特异性机制。可以选择适当的细胞系或宿主系统以便确保所表达的抗体或其部分的正确修饰和加工。为此,可以使用具有用于初级转录物的适当加工、基因产物的糖基化和磷酸化的细胞机器(cellular machinery)的真核宿主细胞。这类哺乳动物宿主细胞包括但不限于cho、very、bhk、hela、cos、mdck、293、3t3、w138、bt483、hs578t、htb2、bt2o和t47d、ns0(不内源性地产生任何功能性免疫球蛋白链的鼠骨髓瘤细胞系)、sp20、crl7o3o以及hss78bst细胞。通过使人淋巴细胞永生化而产生的人细胞系可以用来重组产生单克隆抗体。人细胞系per.c6.(库赛尔(crucell),荷兰)可以用来重组产生单克隆抗体。
[0243]
可以用作表达重组抗体的宿主的另外的细胞系包括但不限于昆虫细胞(例如sf21/sf9,粉纹夜蛾(trichoplusia ni)bti
‑
tn5b1
‑
4)或酵母细胞(例如啤酒酵母(s.cerevisiae)、毕赤酵母属(pichia)、us7326681;等)、植物细胞(us20080066200);以及鸡细胞(wo2008142124)。
[0244]
在某些实施例中,本发明的抗体被表达于具有抗体的稳定表达的细胞系中。稳定表达可以用于重组蛋白的长期、高产量生产。例如,可以产生稳定表达抗体分子的细胞系。可以用适当的包含表达控制元件(例如,启动子、增强子、转录终止子、聚腺苷酸化位点等)和选择性标志基因的工程化的载体转化宿主细胞。在引入外源dna后,可以允许细胞在富集培养基中生长1
‑
2天,并且然后转换至选择性培养基。在重组质粒中的选择性标记赋予对选择的抗性,并且允许将质粒稳定整合进其染色体中的细胞生长并且形成病灶,进而可以将其克隆并且扩充成细胞系。用于以高产量产生稳定细胞系的方法是在本领域中熟知的,并且试剂通常是可商购的。
[0245]
在某些实施例中,本发明的抗体被表达于具有抗体的瞬时表达的细胞系中。瞬时转染是这样一种方法:在该方法中引入细胞的核酸不会整合至该细胞的基因组或染色体dna中。事实上该核酸被维持作为细胞中的染色体外元件,例如,作为附加体。附加体的核酸的转录过程不受影响,并且产生由附加体的核酸编码的蛋白质。
[0246]
稳定抑或瞬时转染的细胞系被维持在本领域中熟知的产生单克隆抗体的表达和产生的细胞培养基和条件中。在某些实施例中,哺乳动物细胞培养基是基于可商购的培养基配制品,包括例如dmem或汉姆氏(ham's)f12。在其他实施例中,将细胞培养基进行改良以便支持细胞生长和生物蛋白表达两者的增长。如在此所用,术语“细胞培养基”、“培养基”和“培养基配制品”是指在多细胞生物体或组织以外的人工体外环境中用于细胞的维持、生
长、增殖、或扩增的营养液。可以将细胞培养基优化以用于特定的细胞培养用途,包括例如,被配制来促进细胞生长的细胞培养物生长培养基,或者被配制来促进重组蛋白产生的细胞培养物生产培养基。术语营养素、成分(ingredient)、以及组分(component)在此可互换地使用,是指组成细胞培养基的组分(constituent)。
[0247]
在一个实施例中,使用补料分批法维持细胞系。如在此所用,“补料分批法”是指在首先用基础培养基孵育之后,用另外的营养素供应给补料分批细胞培养物的方法。例如,补料分批法可以包括根据所确定的补料时间表在给定时期内添加补充培养基。因此,“补料分批细胞培养”是指这样一种细胞培养:其中最初将细胞(典型地为哺乳动物细胞)和培养基供应至培养容器,并且在培养期间,连续地或以不连续递增方式将另外的培养营养素馈送至培养物,有或没有在培养终止之前的定期的细胞和/或产物的收获。
[0248]
所使用的细胞培养基以及其中含有的营养素是本领域的技术人员已知的。在一个实施例中,细胞培养基包含基础培养基以及产生改良的基础培养基的至少一种水解产物,例如基于大豆的水解产物、基于酵母的水解产物、或这两种类型的水解产物的组合。在另一个实施例中,另外的营养素可以仅包含基础培养基如浓缩的基础培养基,或者可以仅包含水解产物或浓缩的水解产物。适合的基础培养基包括但不限于达尔伯克改良的伊格尔培养基(dmem)、dme/f12、最低必需培养基(mem)、伊格尔基础培养基(bme)、rpmi 1640、f
‑
10、f
‑
12、α
‑
最低必需培养基(α
‑
mem)、格拉斯哥最低必需培养基(g
‑
mem)、pf cho(参见,例如,cho无蛋白质培养基(西格玛)或用于无蛋白质的cho细胞的ex
‑
cell
tm 325pf cho无血清培养基(safc生物科学公司))、以及伊斯科夫改良的达尔伯克培养基(iscove's modified dulbecco's medium)。可以用于本发明的基础培养基的其他实例包括bme基础培养基(gibco
‑
英杰(invitrogen);还参见伊格尔,h(eagle,h)(1965)实验生物学与医学会会报(proc.soc.exp.biol.med.)89,36);达尔伯克改良的伊格尔培养基(dmem,粉末)(gibco
‑
英杰(#31600);还参见达尔伯克(dulbecco)和弗里曼(freeman)(1959)病毒学(virology)8,396;史密斯(smith)等人(1960)病毒学12,185.体外组织培养标准委员会(tissue culture standards committee,in vitro)6:2,93);cmrl 1066培养基(gibco
‑
英杰(#11530);还参见帕克r.c.(parker r.c.)等人(1957)特别出版物(special publications),纽约科学院,5,303)。
[0249]
基础培养基可以是无血清的,意指该培养基不含血清(例如,胎牛血清(fbs)、马血清、山羊血清、或本领域的技术人员已知的任何其他动物源的血清),或没有动物蛋白的培养基或化学上限定的培养基。
[0250]
可以改良基础培养基以便去除在标准基础培养基中发现的某些非营养组分,如不同的无机和有机缓冲液、一种或多种表面活性剂、以及氯化钠。此类组分从基础细胞培养基的去除允许剩余营养组分的浓度增加,并且可以改进总体的细胞生长和蛋白质表达。另外,可以根据细胞培养条件的要求,将省去的组分添加回至含有改良的基础细胞培养基的细胞培养基中。在某些实施例中,细胞培养基包含改良的基础细胞培养基以及至少一种以下营养素:铁源、重组生长因子;缓冲液;表面活性剂;渗透性调节剂;能量源;以及非动物水解产物。另外,改良的基础细胞培养基可以任选地含有氨基酸、维生素、或氨基酸和维生素两者的组合。在另一个实施例中,改良的基础培养基进一步包含谷氨酰胺,例如l
‑
谷氨酰胺和/或氨甲蝶呤。
[0251]
可以通过使用本领域中已知的补料分批、分批、灌注的生物反应器工艺或连续进料生物反应器方法而大量地进行抗体产生。大规模生物反应器具有至少1000升的容量,优选约1,000至100,000升的容量。这些生物反应器可以使用叶轮搅拌器来分布氧和营养素。小规模生物反应器通常是指在不大于大致100升的体积容量中的细胞培养,并且范围可以是从约1升至约100升。可替代地,一次性生物反应器(sub)可以用于大规模抑或小规模培养。
[0252]
温度、ph、搅拌、曝气以及接种密度将取决于所使用的宿主细胞和有待表达的重组蛋白而不同。例如,重组蛋白细胞培养可以被维持在30℃与45℃之间的温度下。可以在培养过程期间监测培养基的ph,这样使得ph停留在最佳水平,对于某些宿主细胞来说,该最佳水平可以是在6.0至8.0的ph范围内。叶轮驱动的混合可以用于此类培养方法以用于搅拌。叶轮的转速可以大致是50至200cm/秒叶尖速度,但可以取决于所培养的宿主细胞的类型而使用本领域中已知的其他气升或其他混合/曝气系统。提供充足的曝气以便在培养物中维持大致20%至80%空气饱和度的溶解氧浓度,这再次取决于所培养的所选择的宿主细胞。可替代地,生物反应器可以直接将空气或氧气喷射至培养基之中。存在其他供氧方法,包括采用中空纤维膜曝气器的无泡曝气系统。
[0253]
噬菌体展示技术
[0254]
可以从使用麦卡弗蒂(mccafferty)等人,自然,348:552
‑
554(1990)、克拉克森(clackson)等人,自然,352:624
‑
628(1991)以及马克斯(marks)等人,分子生物学杂志(j.mol.biol.),222:581
‑
597(1991)中描述的技术产生的抗体噬菌体文库分离单克隆抗体或抗体片段。在这类方法中,抗体可以通过重组组合抗体文库(优选scfv噬菌体展示文库)的筛选而分离,这些文库是使用从源自人淋巴细胞的mrna制备的人vl和vh cdna制备的。用于制备和筛选这类文库的方法学是本领域中已知的。除了用于产生噬菌体展示文库的可商购的试剂盒(例如,法玛西亚重组噬菌体抗体系统(pharmacia recombinant phage antibody system),目录号27
‑
9400
‑
01;和stratagene surfzap
tm
噬菌体展示试剂盒,目录号240612)之外,特别适合用于在产生和筛选抗体展示文库中使用的方法和试剂的实例可以在例如美国专利号6,248,516;us 6,545,142;6,291,158;6,291,159;6,291,160;6,291,161;6,680,192;5,969,108;6,172,197;6,806,079;5,885,793;6,521,404;6,544,731;6,555,313;6,593,081;6,582,915;7,195,866中找到。因此,这些技术是用于产生和分离单克隆抗体的传统单克隆抗体杂交瘤技术的可行替代。
[0255]
在噬菌体展示方法中,将功能抗体结构域展示在噬菌体颗粒的表面上,这些颗粒携带对其进行编码的多核苷酸序列。在一个具体实施例中,这种噬菌体可以用于展示从谱系或组合抗体文库(例如,人或鼠类)表达的抗原结合结构域。表达结合所感兴趣的抗原的抗原结合结构域的噬菌体可以用抗原来选择或鉴别,例如,使用标记的抗原或结合或捕获于固体表面或珠粒上的抗原。这些方法中使用的噬菌体典型地是丝状噬菌体,该噬菌体包括从具有fab、fv的噬菌体表达的fd和m13结合结合域或重组融合至噬菌体基因iii或基因viii蛋白的二硫化物稳定的fv抗体结构域。
[0256]
如上文参考文献中所述,在噬菌体选择之后,来自噬菌体的抗体编码区可以被分离并且用于产生完整抗体,包括人抗体、人源化抗体或任何其他所希望的抗原结合片段,并且表达在任何所希望的宿主之中,包括哺乳动物细胞、昆虫细胞、植物细胞、酵母、以及细
菌,例如如下文详细描述的。例如,还可以使用本领域中已知的方法采用重组产生fab、fab’和f(ab')2片段的技术,这些方法如以下文献中所披露的那些:pct公布wo 92/22324;马利纳克斯(mullinax)等人,生物技术(biotechniques)12(6):864
‑
869(1992);以及贝特尔(better)等人,科学240:1041
‑
1043(1988)。
[0257]
可以用于产生单链fv和抗体的技术的实例包括在美国专利号4,946,778和5,258,498中描述的那些。因此,以上所述的和本领域中熟知的那些技术可以用于产生重组抗体,其中从噬菌体展示文库分离结合结构域(例如scfv)。
[0258]
抗体纯化和分离
[0259]
一旦抗体分子已经通过重组或杂交瘤表达而产生,可以通过本领域中已知用于纯化免疫球蛋白分子的任何方法将其纯化,例如,通过色谱法(例如,离子交换色谱,特别是通过对于特定抗原蛋白a或蛋白g的亲和力的亲和色谱,以及尺寸柱色谱(sizing column chromatography))、离心、差别溶解度,或者通过用于纯化蛋白质的任何其他标准技术。此外,本发明的抗体或其片段可以融合至异源多肽序列(在此称为“标签”),以便促进纯化。
[0260]
在使用重组技术时,抗体可以细胞内、在周质间隙中产生,或直接分泌至培养基之中。如果抗体是在细胞内产生,那么作为第一步,(例如)通过离心或超滤去除宿主细胞抑或溶解片段的颗粒碎片。卡特(carter)等人,生物/技术(bio/technology),10:163
‑
167(1992)描述了一种用于分离分泌到大肠杆菌的周质空间中的抗体的程序。在抗体分泌到培养基中的情况下,通常首先使用可商购的蛋白质浓缩滤器(例如amicon或millipore pellicon超滤单元)对来自这类表达系统的上清液进行浓缩。蛋白酶抑制剂如pmsf可以被包括在任何前述步骤中以便抑制蛋白水解,并且抗生素可以被包括来防止外来污染物的生长。
[0261]
从细胞制备的抗体组合物可以单独使用例如羟基磷灰石色谱、疏水相互作用色谱、离子交换色谱、凝胶电泳、渗析和/或亲和色谱或与其他纯化步骤组合来纯化。蛋白a作为亲和配体的适合性取决于抗体中存在的任何免疫球蛋白fc结构域的种类和同种型并且将由本领域的技术人员理解。附着有亲和配体的基质最常为琼脂糖,但其他基质也是可用的。在机械上稳定的基质如可控多孔玻璃或聚(苯乙烯二乙烯基)苯允许比用琼脂糖可以实现的更快的流速和更短的处理时间。在抗体包含ch3结构域的情况下,bakerbond abx树脂(马林克罗特贝克有限公司(j.t.baker),菲利普斯堡(phillipsburg),新泽西)可用于纯化。取决于待回收的抗体也可用其他蛋白质纯化技术,如离子交换柱上分级分离、乙醇沉淀、反相hplc、二氧化硅上色谱、肝素上色谱、阴离子或阳离子交换树脂(如聚天冬氨酸柱)上sepharose色谱、色谱聚焦、sds
‑
page、以及硫酸铵沉淀。
[0262]
在任何一个或多个初步纯化步骤之后,包含感兴趣的抗体和污染物的混合物可以经受低ph疏水性相互作用色谱,该色谱使用ph介于约2.5
‑
4.5之间的洗脱缓冲液并且在低盐浓度(例如,从约0至0.25m盐)下进行。
[0263]
因此,在某些实施例中提供基本上纯化/分离的本发明的抗体。在一个实施例中,这些分离/纯化的重组表达的抗体可以被给予至患者以便介导预防或治疗作用。预防剂是被设计且用于预防疾病、病症或感染发生的药物或治疗。治疗剂具体地涉及特定疾病、病症或感染的治疗。治疗剂量是用于治疗特定疾病、病症或感染所需的量。在另一个实施例中,这些分离/纯化的抗体可以用于诊断甲型流感病毒感染。
[0264]
人抗体
[0265]
人抗体可以使用本领域中熟知的方法来产生。人抗体避免一些与具有鼠类或大鼠可变区和/或恒定区的抗体相关的问题。这种鼠类或大鼠源性蛋白质的存在可以导致抗体的快速清除或可以导致患者产生针对该抗体的免疫应答。
[0266]
可以通过体外方法获得人抗体。适合的实例包括但不限于噬菌体展示(医学免疫公司(medimmune(以前为cat)、莫弗西斯生物公司(morphosys)、dyax、biosite/梅达瑞克斯、xoma、symphogen、亚力兄公司(alexion,(以前为proliferon)、affimed)、核糖体展示(医学免疫公司(以前为cat))、酵母展示等等。可以使用噬菌体展示技术(参见例如美国专利号5,969,108)从来自未免疫供体的免疫球蛋白可变(v)结构域基因谱系体外产生人抗体和抗体片段。根据这项技术,抗体v结构域基因是以框内方式克隆至丝状噬菌体(如m13或fd)的主要或次要外壳蛋白基因中,并且在噬菌体颗粒的表面上展示为功能性抗体片段。因为丝状颗粒含有噬菌体基因组的单链dna拷贝,所以基于抗体的功能特性的选择还导致编码表现那些特性的抗体的基因的选择。因而,噬菌体模拟b细胞的一些特性。噬菌体展示可以例如综述于约翰逊凯文s.(johnson,kevin s.)和奇斯韦尔戴维德j.(chiswell,david j.),结构生物学当前观点(current opinion in structural biology)3:564
‑
571(1993)中的多种形式进行。若干v基因区段来源可以用于噬菌体展示。克拉克森等人,自然,352:624
‑
628(1991)从源自免疫小鼠的脾脏的v基因的小随机组合文库分离了抗噁唑酮抗体的不同阵列。可以构建来自未免疫人供体的v基因谱系,并且可以按照由马克斯(marks)等人,分子生物学杂志222:581
‑
597(1991)或格里菲斯(griffith)等人,欧洲分子生物学学会杂志(embo j.)12:725
‑
734(1993)描述的技术而基本上分离针对抗原(包括自身抗原)的不同阵列的抗体。还参见美国专利号5,565,332和5,573,905。
[0267]
如上所讨论,还可以由体外活化的b细胞产生人抗体(参见美国专利号5,567,610和5,229,275)。
[0268]
在免疫应答的成熟过程中,免疫球蛋白基因经历不同的修饰,包括在v、d和j基因区段之间的重组、同种型转换、以及可变区中的超突变。重组和体细胞超突变是产生抗体多样性和亲和力成熟的基础,但是它们还可以产生序列文库,这些序列文库可以使作为治疗剂的这种免疫球蛋白的商业生产有困难或者增加抗体的免疫原性风险。通常,虽然在框架区中的突变可以增加免疫原性的风险,cdr区中的突变可能促成改进的亲和力和功能。这种风险可以通过将框架突变回复为种系而降低,同时确保抗体的活性未受不利影响。多样化过程还可以产生一些结构文库,或这些结构文库可以存在于促成重链和轻链可变域的种系序列中。不论来源,可能希望的是将潜在的结构文库去除,这些结构文库可以导致产物的不稳定性、聚集、异质性、或者增加的免疫原性。不希望的文库的实例包括不成对的半胱氨酸(可以导致二硫键混乱或可变的巯基加合物形成)、n
‑
连接糖基化位点(导致结构和活性的异质性)、以及脱酰胺作用(例如ng、ns)、异构化(dg)、氧化(暴露的甲硫氨酸)、以及水解(dp)位点。
[0269]
因此,为了降低免疫原性的风险并且改进药学特性,可能希望的是将框架序列回复为种系、将cdr回复为种系、和/或去除结构倾向(structural liability)。
[0270]
因此,在一个实施例中,在特定的抗体在氨基酸水平上不同于其对应的种系序列的情况下,可以将抗体序列突变回种系序列。通过使用标准分子生物学技术,这种纠正突变
可以在一个、二个、三个或更多个位置、或任何突变位置的组合处发生。
[0271]
抗体片段
[0272]
在某些实施例中,本发明抗体是抗体片段或包含这些片段的抗体。抗体片段包括全长抗体的一部分,通常是其抗原结合区或可变区。抗体片段的实例包括fab、fab'、f(ab')2、fd以及fv片段。双抗体;线性抗体(美国专利号5,641,870)和单链抗体分子。
[0273]
在传统上,这些片段是使用本领域中熟知的技术、经由完整抗体的蛋白水解消化而衍生的。然而,现在可以通过重组宿主细胞直接产生这些片段。所有fab、fv和scfv抗体片段都可以表达在大肠杆菌中并且从其分泌,因而允许大量这些片段的容易生产。在一个实施例中,可以从以上讨论的抗体噬菌体文库分离抗体片段。可替代地,fab'
‑
sh片段还可以直接从大肠杆菌中回收并且化学连接以便形成f(ab')2片段(卡特(carter)等人,生物/技术,10:163
‑
167(1992))。根据另一个途径,可以直接从重组宿主细胞培养物分离f(ab')2片段。用于产生抗体片段的其他技术对于熟练的从业人员将是清楚的。在其他实施例中,选择的抗体是单链fv片段(scfv)。在某些实施例中,抗体不是fab片段。fv和scfv是没有恒定区的具有完整组合位点的仅有种类;因此,它们适合用于在体内使用过程中减少非特异性结合。可以将scfv融合蛋白构建为在scfv的氨基末端抑或羧基末端产生效应蛋白融合。
[0274]
在某些实施例中,本发明的抗体是结构域抗体,例如对应于人抗体的重链可变区(vh)或轻链可变区(vl)的、包含抗体的小功能性结合单位的抗体。结构域抗体的实例包括但不限于,多曼提斯(domantis)的那些结构域抗体(参见例如,wo04/058821;wo04/081026;wo04/003019;wo03/002609;美国专利号6,291,158;6,582,915;6,696,245;以及6,593,081)。
[0275]
在本发明的某些实施例中,本发明的抗体是线性抗体。线性抗体包括一对串联的fd区段(vh
‑
ch1
‑
vh
‑
ch1),这些区段形成一对抗原结合区。,参见,萨帕塔(zapata)等人,蛋白质工程化(protein eng.),8(10):1057
‑
1062(1995)。
[0276]
其他氨基酸序列修饰
[0277]
除了以上描述的人、人源化和/或嵌合抗体外,本发明还涵盖本发明的抗体的进一步修饰以及其变体和其片段,这些修饰包括可变轻链(vl)结构域和/或可变重链(vh)结构域和/或fc区中的一个或多个氨基酸残基和/或多肽取代、添加和/或缺失以及翻译后修饰。包括在这些修饰中的是其中抗体已经共价连接至一个部分的抗体结合物。适用于连接至抗体的部分包括但不限于蛋白质、肽、药物、标记物、以及细胞毒素。可以作出针对抗体的这些改变以便改变或微调这些抗体的特征(生物化学的、结合和/或功能性的),适用于治疗和/或诊断甲型流感病毒感染。用于形成轭合物、作出氨基酸和/或多肽改变以及翻译后修饰的方法是本领域中熟知的,其中一些在下文详述。
[0278]
针对抗体的氨基酸变化必然产生与以上鉴别的抗体序列或亲本抗体序列小于100%一致的序列。在某些实施例中,在这种情况下,抗体可以与如在此所述的抗体的重链或轻链可变结构域的氨基酸序列具有约25%至约95%序列一致性。因此,在一个实施例中,修饰的抗体可以具有这样一种氨基酸序列:该氨基酸序列与如在此所述的抗体的重链或轻链可变结构域的氨基酸序列具有至少25%、35%、45%、55%、65%、75%、80%、85%、90%、95%、96%、97%、98%或99%氨基酸序列一致性或相似性。在另一个实施例中,改变的抗体可以具有这样一种氨基酸序列:该氨基酸序列与如在此所述的抗体的重链或轻链cdr1、
cdr2或cdr3的氨基酸序列具有至少25%、35%、45%、55%、65%、75%、80%、85%、90%、95%、96%、97%、98%或99%氨基酸序列一致性或相似性。在另一个实施例中,改变的抗体可以具有这样一种氨基酸序列:该氨基酸序列与如在此所述的抗体的重链或轻链fr1、fr2、fr3或fr4的氨基酸序列具有至少25%、35%、45%、55%、65%、75%、80%、85%、90%、95%、96%、97%、98%或99%氨基酸序列一致性或相似性。
[0279]
在某些实施例中,通过引入抗体的一个或多个可变区中的一种或多种氨基酸改变(例如,置换、缺失和/或添加)而产生改变的抗体。在另一个实施例中,氨基酸改变被引入框架区。框架区残基的一种或多种改变可以导致对于抗原的抗体的结合亲和力的改进。当这些变化是针对人源化抗体(不同于cdr区,框架区可以来自不同的物种)做出时,尤其如此。用于修饰的框架区残基的实例包括直接非共价结合抗原的那些(阿密特(amit)等人,科学,233:747
‑
753(1986));与cdr相互作用/实现cdr的构象的那些(乔西亚(chothia)等人,分子生物学杂志,196:901
‑
917(1987));和/或参与vl
‑
vh界面的那些(美国专利号5,225,539和6,548,640)。在一个实施例中,可以改变从约一个至约五个框架残基。有时,甚至在没有任何高变区残基已被改变时,这可能足以产生适合于在临床前试验中使用的抗体突变体。然而,通常改变的抗体将包含另外的一种或多种高变区改变。
[0280]
用于产生改变的抗体的一种有用程序被称为“丙氨酸扫描诱变”(坎宁安(cunningham)和威尔斯(wells),科学,244:1081
‑
1085(1989))。在这种方法中,一个或多个高变区残基被一个或多个丙氨酸或聚丙氨酸残基置换,从而改变氨基酸与靶抗原的相互作用。然后,对于取代显示功能敏感度的那些高变区残基通过在(或针对)取代位点引入另外的或其他突变而被改善(refined)。t因此,尽管用于引入氨基酸序列变异的位点被预先确定,本身突变的本性不需要预先确定。如在此所述,针对以此方式产生的ala突变体的生物活性来筛选它们。
[0281]
在某些实施例中,取代变体涉及取代亲本抗体(例如,人源化或人抗体)的一个或多个高变区残基。通常,所得的被选择用于进一步开发的一种或多种变体相对于它们从其产生的亲本抗体而言将具有改进的生物特性。用于产生这类取代变体的方便的方式涉及使用噬菌体的亲和力成熟(霍金斯(hawkins)等人,分子生物学杂志,254:889
‑
896(1992)和洛曼(lowman)等人,生物化学(biochemistry),30(45):10832
‑
10837(1991))。简言之,若干高变区位点(6
‑
7个位点)被突变以在每个位点产生所有可能的氨基酸取代。因而产生的抗体突变体被展示为来自丝状噬菌体颗粒的、作为与包装在每个颗粒中的m13的基因iii产物融合的单价形式。然后,如在此披露,针对噬菌体展示突变体的生物活性(例如,结合亲和力)来筛选它们。
[0282]
在抗体序列中的突变可以包括取代、缺失(包括内部缺失)、添加(包括产生融合蛋白的添加)、或在氨基酸序列之内和/或邻近处的氨基酸残基的保守性取代,但是那导致“沉默”改变,导致这种改变产生功能上等效的抗体。可以在涉及残基的极性、电荷、溶解度、疏水性、亲水性、和/或两亲性的相似性的基础上做出保守性氨基酸取代。例如,非极性(疏水性)氨基酸包括丙氨酸、亮氨酸、异亮氨酸、缬氨酸、脯氨酸、苯丙氨酸、色氨酸和蛋氨酸;极性中性氨基酸包括甘氨酸、丝氨酸、苏氨酸、半胱氨酸、酪氨酸、天冬酰胺和谷氨酰胺;带正电荷的(碱性)氨基酸包括精氨酸、赖氨酸和组氨酸;并且带负电荷的(酸性)氨基酸包括天冬氨酸和谷氨酸。另外,甘氨酸和脯氨酸是能影响链取向的残基。非保守性取代将使得必需
将这些类别之一的成员与另一类别中的成员交换。此外,如果希望的话,非典型氨基酸或化学氨基酸类似物可以作为取代或添加而被引入抗体序列中。非典型氨基酸总体上包括但不限于普通氨基酸的d
‑
异构体、α
‑
氨基异丁酸、4
‑
氨基丁酸、abu、2
‑
氨基丁酸、γ
‑
abu、ε
‑
ahx、6
‑
氨基己酸、aib、2
‑
氨基异丁酸、3
‑
氨基丙酸、鸟氨酸、正亮氨酸、正缬氨酸、羟脯氨酸、肌氨酸、瓜氨酸、磺基丙氨酸、叔丁基甘氨酸、叔丁基丙氨酸、苯基甘氨酸、环己基丙氨酸、β
‑
丙氨酸、氟代氨基酸、设计者(designer)氨基酸(例如β
‑
甲基氨基酸、cα
‑
甲基氨基酸、nα
‑
甲基氨基酸、以及氨基酸类似物。
[0283]
在另一个实施例中,不参与维持抗体的适当构象的任何半胱氨酸残基还可以被(通常用丝氨酸)取代以便改进分子的氧化稳定性并且预防异常交联。相反,可以将半胱氨酸键添加至抗体以改进它的稳定性(特别是在抗体是如fv片段的抗体片段的情况下)。
[0284]
变体fc区
[0285]
已知fc区的变体(例如,氨基酸取代和/或添加和/或缺失)增强或减小抗体的效应子功能(参见,例如,美国专利号5,624,821;5,885,573;6,538,124;7,317,091;5,648,260;6,538,124;wo 03/074679;wo 04/029207;wo 04/099249;wo 99/58572;美国公开号2006/0134105;2004/0132101;2006/0008883)并且可以改变该抗体的药代动力学特性(例如,半衰期)(参见,美国专利6,277,375和7,083,784)。因此,在某些实施例中,本发明的抗体包含改变的fc区(在此也被称为“变体fc区”),其中已经在该fc区中做出一种或多种改变以便改变这些抗体的功能特性和/或药代动力学特性。此类改变可以导致clq结合和补体依赖的细胞毒性(cdc)或者对于igg的fcγr结合以及抗体依赖的细胞的细胞毒性(adcc)、或者抗体依赖的细胞介导的吞噬作用(adcp)的减少或增加。本发明涵盖在此所述的具有变体fc区的抗体,其中已经做出变化以便微调效应子功能,从而增强或减弱、提供所希望的效应子功能。因此,本发明的抗体包含变体fc区(即,已经如下文所讨论进行改变的fc区)。本发明的包含变体fc区的抗体在此还被称为“fc变体抗体”。如在此所用,天然的是指未修饰的亲本序列,并且包含天然fc区的抗体在此被称为“天然fc抗体”。fc变体抗体可以通过本领域已熟知的多种方法产生。非限制性实例包括分离抗体编码区(例如,从杂交瘤)以及在分离的抗体编码区的fc区中形成一个或多个所希望的取代。可替代地,可以将抗体的抗原结合部分(例如,可变区)亚克隆至编码变体fc区的载体中。在一个实施例中,如与天然fc区相比,变体fc区表出相似的诱导效应子功能的水平。在另一个实施例中,如与天然fc相比的,变体fc区展现出较高的效应子功能的诱导作用。在下文详述了变体fc区的一些具体实施例。用于测量效应子功能的方法是本领域中熟知的。
[0286]
通过fc区的变化来改变抗体的效应子功能,这些变化包括但不限于氨基酸取代、氨基酸添加、氨基酸缺失以及对于fc氨基酸的翻译后修饰(例如,糖基化)的变化。以下所述的方法可以用于微调本发明抗体的效应子功能、fc区对于fcr的结合特性(例如,亲和力和特异性)的比率,从而产生具有所希望特性的治疗性抗体。
[0287]
应理解,如在此使用的fc区包括含有抗体的除第一恒定区免疫球蛋白结构域之外的恒定区的多肽。因此,fc是指iga、igd、以及igg的最后两个恒定区免疫球蛋白结构域、以及ige和igm的最后三个恒定区免疫球蛋白结构域、以及在这些结构域的n末端的柔性铰链。对于iga和igm,fc可以包括j链。对于igg,fc包含免疫球蛋白结构域cγ2和cγ3(cγ2和cγ3)以及cγ1(cγ1)与cγ2(cγ2)之间的铰链。尽管fc区的界限可以改变,人igg重链fc区通
常被定义为在其羧基末端包括残基c226或p230,其中编号是根据如卡巴特(kabat)中列出的eu索引进行的。fc可以是指孤立地这个区,或在抗体、抗体片段或fc融合蛋白的环境中的这个区。已在多个不同fc位置处观察到多态性,包括但不限于如由eu索引编号的位置270、272、312、315、356、以及358,并且因此所呈现的序列与现有技术序列之间可能存在略微的差异。
[0288]
在一个实施例中,与天然fc抗体相比,fc变体抗体对于一种或多种fc受体表现出改变的结合亲和力,这些fc受体包括但不限于fcrn、fcγri(cd64)(包括同种型fcγria、fcγrib和fcγric);fcγrii(cd32,包括同种型fcγriia、fcγriib和fcγriic);以及fcγriii(cd16,包括同种型fcγriiia和fcγriiib)。
[0289]
在一个实施例中,相对于天然fc抗体,fc变体抗体具有增强的对一种或多种fc配体的结合。在另一个实施例中,fc变体抗体表现出增加的或降低的对fc配体的亲和力,该亲和力比天然fc抗体所表现的对于fc配体的亲和力高或低至少2倍、或至少3倍、或至少5倍、或至少7倍、或至少10倍、或至少20倍、或至少30倍、或至少40倍、或至少50倍、或至少60倍、或至少70倍、或至少80倍、或至少90倍、或至少100倍、或至少200倍、或在2倍与10倍之间、或在5倍与50倍之间、或在25倍与100倍之间、或在75倍与200倍之间、或在100倍与200倍之间。在另一个实施例中,fc变体抗体表现的对fc配体的亲和力比天然fc抗体所表现的对fc配体的亲和力高或低至少90%、至少80%、至少70%、至少60%、至少50%、至少40%、至少30%、至少20%、至少10%、或至少5%。在某些实施例中,fc变体抗体具有增加的对fc配体的亲和力。在其他实施例中,fc变体抗体具有降低的对fc配体的亲和力。
[0290]
在一个具体实施例中,fc变体抗体对fc受体fcγriiia具有增强的结合。在另一个具体实施例中,fc变体抗体对fc受体fcγriib具有增强的结合。在另一个具体实施例中,fc变体抗体对fc受体fcγriiia和fcγriib两者具有增强的结合。在某些实施例中,与天然fc抗体相比,对fcγriiia具有增强的结合的fc变体抗体在结合fcγriib受体方面不具有伴随的增加。在一个具体实施例中,fc变体抗体对fc受体fcγriiia具有降低的结合。在另一个具体实施例中,fc变体抗体对fc受体fcγriib具有降低的结合。在仍然另一个具体实施例中,对fcγriiia和/或fcγriib表现出改变的亲和力的fc变体抗体对fc受体fcrn具有增强的结合。在又另一个具体实施例中,相对于天然fc抗体,对fcγriiia和/或fcγriib表现出改变的亲和力的fc变体抗体对c1q具有改变的结合。
[0291]
在一个实施例中,fc变体抗体表现的对fcγriiia受体的亲和力比天然fc抗体所表现的对fcγriiia受体的亲和力高或低至少2倍、或至少3倍、或至少5倍、或至少7倍、或至少10倍、或至少20倍、或至少30倍、或至少40倍、或至少50倍、或至少60倍、或至少70倍、或至少80倍、或至少90倍、或至少100倍、或至少200倍、或在2倍与10倍之间、或在5倍与50倍之间、或在25倍与100倍之间、或在75倍与200倍之间、或在100倍与200倍之间。在另一个实施例中,fc变体抗体表现的对fcγriiia的亲和力比天然fc抗体所表现的对fcγriiia的亲和力高或低至少90%、至少80%、至少70%、至少60%、至少50%、至少40%、至少30%、至少20%、至少10%、或至少5%。
[0292]
在一个实施例中,fc变体抗体表现的对fcγriib受体的亲和力比天然fc抗体所表现的对fcγriib受体的亲和力高或低至少2倍、或至少3倍、或至少5倍、或至少7倍、或至少10倍、或至少20倍、或至少30倍、或至少40倍、或至少50倍、或至少60倍、或至少70倍、或至少
80倍、或至少90倍、或至少100倍、或至少200倍、或在2倍与10倍之间、或在5倍与50倍之间、或在25倍与100倍之间、或在75倍与200倍之间、或在100倍与200倍之间。在另一个实施例中,fc变体抗体表现的对fcγriib的亲和力比天然fc抗体所表现的对fcγriib的亲和力高或低至少90%、至少80%、至少70%、至少60%、至少50%、至少40%、至少30%、至少20%、至少10%、或至少5%。
[0293]
在一个实施例中,相对于天然fc抗体,fc变体抗体表现出增加的或降低的对c1q的亲和力。在另一个实施例中,fc变体抗体表现的对c1q受体的亲和力比天然fc抗体所表现的对c1q受体的亲和力高或低至少2倍、或至少3倍、或至少5倍、或至少7倍、或至少10倍、或至少20倍、或至少30倍、或至少40倍、或至少50倍、或至少60倍、或至少70倍、或至少80倍、或至少90倍、或至少100倍、或至少200倍、或在2倍与10倍之间、或在5倍与50倍之间、或在25倍与100倍之间、或在75倍与200倍之间、或在100倍与200倍之间。在另一个实施例中,fc变体抗体表现的对c1q的亲和力比天然fc抗体所表现的对c1q的亲和力高或低至少90%、至少80%、至少70%、至少60%、至少50%、至少40%、至少30%、至少20%、至少10%、或至少5%。在仍然另一个具体实施例中,对ciq表现出改变的亲和力的fc变体抗体对fc受体fcrn具有增强的结合。在又另一个具体实施例中,相对于天然fc抗体,表现出改变的对c1q的亲和力的fc变体抗体具有改变的对fcγriiia和/或fcγriib的结合。
[0294]
在本领域熟知的是抗体能够通过在本领域中共同称为抗体效应子功能的多种方法引导攻击和破坏。被称为“抗体依赖性细胞介导的细胞毒性”或“adcc”的这些方法之一指的是细胞毒性的一种形式,其中结合至存在于某些细胞毒性细胞(例如,天然杀伤(nk)细胞、嗜中性粒细胞、以及巨噬细胞)上的fc受体(fcr)上的分泌的ig使得这些细胞毒性效应细胞能够特异性地结合至带有抗原的细胞并且随后用细胞毒素杀伤该细胞。针对细胞表面的特异性高亲和力igg抗体“武装”细胞毒性细胞并且是这种杀伤所需要的。细胞的溶解是细胞外的,要求直接的细胞与细胞接触,并且不涉及补体。
[0295]
术语效应子功能所涵盖的另一种方法是补体依赖性细胞毒性(下文称为“cdc”),该方法指的是通过补体系统进行的细胞破坏的生物化学事件。补体系统是在正常血浆中发现的一种蛋白质复合系统,该系统与抗体组合以消灭病原菌以及其他外源细胞。
[0296]
术语效应子功能所涵盖的仍然另一种方法是抗体依赖性细胞介导的吞噬作用(adcp),该方法指的是细胞介导的反应,其中表达一种或多种效应配体的非特异性细胞毒性细胞识别细胞上的结合抗体并且随后引起该细胞的吞噬作用。
[0297]
期望的是,通过用于确定一种或多种fcγr介导的效应细胞功能的体外功能测定来表征fc变体抗体。在某些实施例中,fc变体抗体具有类似的结合特性,并且效应细胞如在基于体外的测定中那样在体内模型(如在此描述和披露的那些)中起作用。然而,本发明不排除在基于体外的测定中未表现出所希望的表型但在体内表现出所希望的表型的fc变体抗体。
[0298]
在某些实施例中,包含fc变体的抗体具有相对于包含天然fc区的抗体增强的细胞毒性或吞噬活性(例如,adcc、cdc和adcp)。在一个具体实施例中,fc变体抗体具有的细胞毒性或吞噬活性比天然fc抗体的细胞毒性或吞噬活性大至少2倍、或至少3倍、或至少5倍、或至少10倍、或至少50倍、或至少100倍、或至少200倍、或2倍与10倍之间、或5倍与50倍之间、或25倍与100倍之间、或75倍与200倍之间、或100与200倍之间。可替代地,fc变体抗体具有
hakko kirin co.,ltd.))表现出相较于天然fc抗体增加的cdc活性,该技术增强抗体的主要作用机制之一,cdc。使用被称为同种型嵌合现象的方法,其中igg3(抗体的同种型)的多个部分被引入igg1(治疗性抗体的标准同种型)的相应区中,技术使cdc活性显著增强超过igg1抑或igg3的cdc活性,同时保留igg1的所希望的特征,如adcc、pk曲线和蛋白质a结合。此外,该技术可以与技术一起使用,从而产生具有增强的adcc和cdc活性的更优异的治疗性mab
[0305]
本发明的fc变体抗体可以具有相对于天然fc抗体增强的adcc活性和增强的血清半衰期。
[0306]
本发明的fc变体抗体可以具有相对于天然fc抗体增强的cdc活性和增强的血清半衰期。
[0307]
本发明的fc变体抗体可以具有相对于天然fc抗体增强的adcc活性、增强的cdc活性以及增强的血清半衰期。
[0308]
可以通过增加fc区对fcrn的结合亲和力而增加包含fc区的蛋白质的血清半衰期。如在此使用的术语“抗体半衰期”表示一种抗体的药物代谢动力学特性,它是抗体分子在它们的给予之后的平均存活时间的量度。抗体半衰期可以表示为从患者(或其他哺乳动物)的身体或其特定区室(例如,如在血清中测量的,即循环半衰期),或在其他组织中消除50%已知量的免疫球蛋白所需要的时间。从一种免疫球蛋白或免疫球蛋白类别到另一种免疫球蛋白或免疫球蛋白类别,半衰期可以不同。通常,抗体半衰期的增加导致循环中所给予抗体的平均停留时间(mrt)的增加。
[0309]
半衰期的增加允许供给患者的药物量的减少以及给药频率的减少。为了增加抗体的血清半衰期,可以例如将补救受体结合表位并入到抗体(尤其是抗体片段)中,如在美国专利号5,739,277中所描述的。如在此所用,术语“补救受体结合表位”是指引起igg分子的体内血清半衰期的增加的igg分子(例如,igg1、igg2、igg3、或igg4)的fc区的表位。
[0310]
可替代地,具有延长的半衰期的本发明的抗体可以通过对鉴别为参与fc与fcrn受体之间的相互作用的氨基酸残基进行修饰而产生(参见例如美国专利号6,821,505和7,083,784;以及wo 09/058492)。另外,可利用本领域中广泛使用的技术通过与peg或白蛋白轭合来使本发明抗体的半衰期增加。在一些实施例中,本发明的包含fc变体区的抗体与包含天然fc区的抗体相比具有增加约5%、约10%、约15%、约20%、约25%、约30%、约35%、约40%、约45%、约50%、约60%、约65%、约70%、约80%、约85%、约90%、约95%、约100%、约125%、约150%或更多的半衰期。在一些实施例中,包含fc变体区的抗体与包含天然fc区的抗体相比具有增加大约2倍、大约3倍、大约4倍、大约5倍、大约10倍、大约20倍、大约50倍或更多、或在2倍与10倍之间、或在5倍与25倍之间、或在15倍与50倍之间的半衰期。
[0311]
在一个实施例中,本发明提供fc变体,其中fc区包含在选自下组的一个或多个位置处的修饰(例如,氨基酸取代、氨基酸插入、氨基酸缺失),该组由以下各项组成:如通过如在kabat中列出的eu索引来编号的221、225、228、234、235、236、237、238、239、240、241、243、244、245、247、250、251、252、254、255、256、257、262、263、264、265、266、267、268、269、279、280、284、292、296、297、298、299、305、308、313、316、318、320、322、325、326、327、328、329、330、331、332、333、334、339、341、343、370、373、378、392、416、419、421、428、433、434、435、436、440、以及443。任选地,fc区可以包含在本领域技术人员已知的另外的和/或替代位置
处的修饰(参见例如美国专利5,624,821、6,277,375、6,737,056、7,083,784、7,317,091、7,217,797、7,276,585、7,355,008、2002/0147311、2004/0002587、2005/0215768、2007/0135620、2007/0224188、2008/0089892、wo 94/29351、以及wo 99/58572)。另外,有用的氨基酸位置以及特定取代在us 6,737,056的表2以及表6
‑
10、us 2006/024298的图41中呈现的表、us 2006/235208的图5、12以及15中呈现的表、us 2006/0173170的图8、9以及10中呈现的表、以及wo 09/058492的图8
‑
10、13以及14中呈现的表中举例说明。
[0312]
在一个具体实施例中,本发明提供fc变体,其中fc区包含至少一个选自下组的取代,该组由以下各项组成:如通过如在kabat中列出的eu索引来编号的221k、221y、225e、225k、225w、228p、234d、234e、234n、234q、234t、234h、234y、234i、234v、234f、235a、235d、235r、235w、235p、235s、235n、235q、235t、235h、235y、235i、235v、235e、235f、236e、237l、237m、237p、239d、239e、239n、239q、239f、239t、239h、239y、240i、240a、240t、240m、241w、241l、241y、241e、241r、243w、243l、243y、243r、243q、244h、245a、247l、247v、247g、250e、250q、251f、252l、252y、254s、254t、255l、256e、256f、256m、257c、257m、257n、262i、262a、262t、262e、263i、263a、263t、263m、264l、264i、264w、264t、264r、264f、264m、264y、264e、265a、265g、265n、265q、265y、265f、265v、265i、265l、265h、265t、266i、266a、266t、266m、267q、267l、268e、269h、269y、269f、269r、270e、280a、284m、292p、292l、296e、296q、296d、296n、296s、296t、296l、296i、296h、296g、297s、297d、297e、298a、298h、298i、298t、298f、299i、299l、299a、299s、299v、299h、299f、299e、305i、308f、313f、316d、318a、318s、320a、320s、322a、322s、325q、325l、325i、325d、325e、325a、325t、325v、325h、326a、326d、326e、326g、326m、326v、327g、327w、327n、327l、328s、328m、328d、328e、328n、328q、328f、328i、328v、328t、328h、328a、329f、329h、329q、330k、330g、330t、330c、330l、330y、330v、330i、330f、330r、330h、331g、331a、331l、331m、331f、331w、331k、331q、331e、331s、331v、331i、331c、331y、331h、331r、331n、331d、331t、332d、332s、332w、332f、332e、332n、332q、332t、332h、332y、332a、333a、333d、333g、333q、333s、333v、334a、334e、334h、334l、334m、334q、334v、334y、339t、370e、370n、378d、392t、396l、416g、419h、421k、428l、428f、433k、433l、434a、424f、434w、434y、436h、440y以及443w。任选地,fc区可以包含本领域的技术人员已知的另外的和/或替代的氨基酸取代,这些氨基酸取代包括但不限于在us 6,737,056的表2以及表6
‑
10、us 2006/024298的图41中呈现的表、us 2006/235208的图5、图12以及图15中呈现的表、us 2006/0173170的图8、图9以及图10中呈现的表、以及wo 09/058492的图8、图9以及图10中呈现的表中举例说明的那些。
[0313]
在一个具体实施例中,本发明提供fc变体抗体,其中fc区包含在选自下组的一个或多个位置处的至少一个修饰(例如,氨基酸取代、氨基酸插入、氨基酸缺失),该组由以下各项组成:如通过如在kabat中列出的eu索引来编号的228、234、235以及331。在一个实施例中,该修饰是选自下组的至少一个取代,该组由以下各项组成:如通过如在kabat中列出的eu索引来编号的228p、234f、235e、235f、235y、以及331s。
[0314]
在另一个具体实施例中,本发明提供fc变体抗体,其中fc区是igg4 fc区并且包含在选自下组的一个或多个位置处的至少一个修饰,该组由以下各项组成:如通过如在kabat中列出的eu索引来编号的228和235。在仍然另一个具体实施例中,fc区是igg4 fc区并且非天然存在的氨基酸是选自下组,该组由以下各项组成:如通过如在kabat中列出的eu索引来
编号的228p、235e以及235y。
[0315]
在另一个具体实施例中,本发明提供fc变体,其中fc区包含在选自下组的一个或多个位置处的至少一个非天然存在的氨基酸,该组由以下各项组成:如通过如在kabat中列出的eu索引来编号的239、330以及332。在一各实施例中,该修饰是选自下组的至少一个取代,该组由以下各项组成:如通过如在kabat中列出的eu索引来编号的239d、330l、330y以及332e。
[0316]
在一个具体实施例中,本发明提供fc变体抗体,其中fc区包含在选自下组的一个或多个位置处的至少一个非天然存在的氨基酸,该组由以下各项组成:如通过如在kabat中列出的eu索引来编号的252、254以及256。在一个实施例中,该修饰是选自下组的至少一个取代,该组由以下各项组成:如通过如在kabat中列出的eu索引来编号的252y、254t以及256e。在本发明的特别优选的抗体中,该修饰是如通过如在kabat中列出的eu索引来编号的三个取代252y、254t和256e(称为“yte”),参见u.s.7,083,784。
[0317]
在某些实施例中,由igg抗体引发的效应子功能很大程度上取决于连接至蛋白质的fc区的碳水化合物部分(克劳蒂亚费拉拉(claudia ferrara)等人,2006,生物技术与生物工程(biotechnology and bioengineering)93:851
‑
861)。因此,可以修饰fc区的糖基化以便提高或降低效应子功能(参见例如乌马纳(umana)等人,1999,自然生物技术(nat.biotechnol)17:176
‑
180;戴维斯(davies)等人,2001,生物技术和生物工程74:288
‑
294;希尔兹(shields)等人,2002,生物化学杂志277:26733
‑
26740;新川(shinkawa)等人,2003,生物化学杂志278:3466
‑
3473;美国专利号6,602,684;6,946,292;7,064,191;7,214,775;7,393,683;7,425,446;7,504,256;美国公开号2003/0157108;2003/0003097;2009/0010921;potillegent
tm
技术(biowa有限公司(biowa,inc.)新泽西州普林斯顿(princeton,n.j.));glycomab
tm
糖基化工程技术(格黎卡特生物技术股份公司(glycart biotechnology ag),瑞士苏黎世(zurich,switzerland)))。因此,在一个实施例中,本发明的抗体的fc区包含氨基酸残基的改变的糖基化。在另一个实施例中,氨基酸残基的改变的糖基化导致效应功能降低。在另一个实施例中,氨基酸残基的改变的糖基化导致效应功能增强。在一个具体实施例中,fc区具有减少的岩藻糖基化。在另一个实施例中,fc区未被岩藻糖基化(参见例如美国专利申请公开号2005/0226867)。在一方面,如在宿主细胞(例如,cho细胞,浮萍(lemna minor))中产生的、具有增加的效应子功能(尤其是adcc)的这些抗体被工程化来产生adcc比通过亲本细胞产生的抗体高出100倍以上的高度去岩藻糖基化的抗体(莫里(mori)等人,2004,生物技术与生物工程88:901
–
908;考克斯(cox)等人,2006,自然生物技术24:1591
‑
7)。
[0318]
向igg分子上的低聚糖添加唾液酸可以增强它们的抗炎症活性并且改变它们的细胞毒性(肯内高(keneko)等人,科学,2006,313:670
‑
673;斯卡隆(scallon)等人,分子免疫学(mol.immuno.)2007年3月;44(7):1524
‑
34)。以上参考的研究论证了具有增加的唾液酸化的igg分子具有抗炎症特性,而具有降低的唾液酸化的igg分子具有提高的免疫刺激特性(例如,提高adcc活性)。因此,对于具体治疗性应用,抗体可以被修饰为具有适当的唾液酸化分布(美国公开号2009/0004179和国际公开号wo 2007/005786)。
[0319]
在一个实施例中,与天然fc区相比,本发明的抗体的fc区包含改变的唾液酸化分布。在一个实施例中,与天然fc区相比,本发明的抗体的fc区包含增加的唾液酸化分布。在
另一个实施例中,与天然fc区相比,本发明的抗体的fc区包含降低的唾液酸化分布。
[0320]
在一个实施例中,本发明的fc变体可以与其他已知的fc变体组合,这些其他已知的fc变体如在贵特(ghetie)等人,1997,自然生物技术,15:637
‑
40;邓肯(duncan)等人,1988,自然,332:563
‑
564;伦德(lund)等人,1991,免疫学杂志(j.immunol),147:2657
‑
2662;伦德等人,1992,分子免疫学,29:53
‑
59;阿莱格里(alegre)等人,1994,移植(transplantation),57:1537
‑
1543;哈钦斯(hutchins)等人,1995,美国美国科学院院刊92:11980
‑
11984;杰弗里斯(jefferis)等人,1995,免疫学快报(immunol lett.),44:111
‑
117;伦德等人,1995,美国实验生物学学会联合会杂志(faseb j)9:115
‑
119;杰弗里斯等人,1996,免疫学快报,54:101
‑
104;伦德等人,1996,免疫学杂志,157:4963
‑
4969;阿穆尔(armour)等人,1999,欧洲免疫学杂志(eur j immunol),29:2613
‑
2624;idusogie等人,2000,免疫学杂志,164:4178
‑
4184;雷迪(reddy)等人,2000,免疫学杂志,164:1925
‑
1933;许(xu)等人,2000,细胞免疫学(cell immunol),200:16
‑
26;idusogie等人,2001,免疫学杂志,166:2571
‑
2575;希尔兹等人,2001,生物化学杂志,276:6591
‑
6604;杰弗里斯等人,2002,免疫学快报,82:57
‑
65;普雷斯塔(presta)等人,2002,生物化学学会汇刊(biochem soc trans)30:487
‑
490);美国专利号5,624,821、5,885,573、5,677,425、6,165,745、6,277,375、5,869,046、6,121,022、5,624,821、5,648,260、6,528,624、6,194,551、6,737,056、7,122,637、7,183,387、7,332,581、7,335,742、7,371,826、6,821,505、6,180,377、7,317,091、7,355,008;2004/0002587;以及wo 99/58572中披露的那些。fc结构域的其他修饰和/或取代和/或添加和/或缺失将易于对本领域技术人员显而易见。
[0321]
糖基化
[0322]
除了糖基化的改变抗体的效应子功能的能力之外,在可变区中的修改的糖基化还可以改变抗体对于抗原的亲和力。在一个实施例中,在本发明抗体的可变区中的糖基化模式被修改。例如,可以制作无糖基化的抗体(即,抗体缺乏糖基化)。可以改变糖基化作用例如以增加该抗体对抗原的亲和性。此类糖类修饰可以通过例如改变在抗体序列之内的一个或多个糖基化位点来完成。例如,可以进行一个或多个氨基酸取代换,导致一个或多个可变区框架糖基化位点的消除,由此消除在那个位点的糖基化。这种无糖基化可以增加抗体对于抗原的亲和力。这种方法在美国专利号5,714,350和6,350,861中更详细地描述。还可以进行一个或多个氨基酸取代,氨基酸取代导致存在于fc区中的糖基化位点(例如,igg的天冬酰胺297)的消除。此外,无糖基化的抗体可以在缺乏必要的糖基化机器(machinery)的细菌细胞中产生。
[0323]
抗体轭合物
[0324]
在某些实施例中,使用本领域中熟知的方法将本发明的抗体轭合或共价连接至一种物质。在一个实施例中,连接的物质是治疗剂、可检测的标记(在此还被称为报道分子)或固体支撑物。用于连接至抗体的适合的物质包括但不限于氨基酸、肽、蛋白质、多糖、核苷、核苷酸、寡核苷酸、核酸、半抗原、药物、激素、脂质、脂质组装体、合成聚合物、聚合微粒、生物细胞、病毒、荧光团、生色团、染料、毒素、半抗原、酶、抗体、抗体片段、放射性同位素、固体基质、半固体基质、及其组合。将另一物质轭合或共价连接至抗体的方法是本领域中熟知的。
[0325]
在某些实施例中,本发明的抗体被轭合至固体支撑物。抗体可轭合于固体支撑物
作为筛选和/或纯化和/或制造方法的一部分。可替代地,本发明的抗体可以轭合至固体支撑物作为诊断方法或组合物的一部分。适用于在本发明中使用的固体支撑物典型地在液相中基本上不可溶。有大量支撑物可用并且为本领域中的普通技术人员所已知。因此,固体支撑物包括固体和半固体基质,如气凝胶和水凝胶、树脂、珠粒、生物芯片(包括薄膜涂布生物芯片)、微流体芯片、硅芯片、多孔板(也称为微量滴定板或微板)、膜、导体和非导体金属、玻璃(包括显微镜载玻片)和磁性支撑物。固体支撑物的更具体实例包括硅胶、聚合物膜、粒子、衍生化塑胶膜、玻璃珠、棉花、塑胶珠粒、氧化铝凝胶、多糖(如琼脂糖凝胶)、聚(丙烯酸酯)、聚苯乙烯、聚(丙烯酰胺)、多元醇、琼脂糖、琼脂、纤维素、葡聚糖、淀粉、聚蔗糖(ficoll)、肝素、糖原、支链淀粉、甘露聚糖、菊糖、硝酸纤维素、重氮纤维素、聚氯乙烯、聚丙烯、聚乙烯(包括聚(乙二醇))、尼龙、乳胶珠粒、磁性珠粒、顺磁性珠粒、超顺磁性珠粒、淀粉等。
[0326]
在一些实施例中,固体支撑物可包括反应性官能团,包括但不限于羟基、羧基、氨基、硫醇基、醛、卤素、硝基、氰基、酰胺基、脲、碳酸酯、氨基甲酸酯、异氰酸酯、砜、磺酸酯、磺酰胺、亚砜等,用于连接本发明的抗体。
[0327]
适合的固相支撑物可根据期望的最终用途和对于各种合成方案的适合性进行选择。例如,在需要形成酰胺键以便将本发明抗体连接至固体支撑物的情况下,可采用一般适用于肽合成的树脂,如聚苯乙烯(例如,获自巴亨公司(bachem inc.),半岛实验室(peninsula laboratories)的pam树脂等)、polyhipe
tm
树脂(获自阿米诺技术公司(aminotech),加拿大)、聚酰胺树脂(获自半岛实验室)、接枝有聚乙二醇的聚苯乙烯树脂(tentagel
tm
,拉普聚合物公司(rapp polymere),图宾根(tubingen),德国)、聚二甲基
‑
丙烯酰胺树脂(可获自米利根公司(milligen)/biosearch公司,加利福利亚州)或pega珠粒(获自聚合物实验室(polymer laboratories))。
[0328]
在某些实施例中,本发明的抗体出于诊断和其他测定的目的轭合至标记,其中可以检测到抗体和/或其相关的配体。轭合至抗体且在在此所述的本发明的方法和组合物中使用的标记是任何化学部分(有机或无机的),该部分在大于280nm的波长下表现出最大吸收并且在共价连接至抗体时保留其光谱特性。标记物包括但不限于发色团、荧光团、荧光蛋白、磷光性染料、叠层染料(tandem dye)、颗粒、半抗原、酶以及放射性同位素。
[0329]
在某些实施例中,抗体轭合至荧光团。如此,用于标记本发明的抗体的荧光团包括但不限于:芘(包括在美国专利5,132,432中披露的相应衍生物化合物中的任一种)、蒽、萘、吖啶、芪、吲哚或苯并吲哚、噁唑或苯并噁唑、噻唑或苯并噻唑、4
‑
氨基
‑7‑
硝基苯并
‑2‑
氧杂
‑
1,3
‑
二唑(nbd)、花青(包括美国专利号6,977,305和6,974,873中的任何相应化合物)、羰花青(包括美国序列号09/557,275;美国专利号4,981,977;5,268,486;5,569,587;5,569,766;5,486,616;5,627,027;5,808,044;5,877,310;6,002,003;6,004,536;6,008,373;6,043,025;6,127,134;6,130,094;6,133,445;以及公开wo 02/26891、wo 97/40104、wo 99/51702、wo 01/21624;ep 1 065 250 a1中的任何相应化合物)、喹诺酮(carbostyryl)、卟啉、水杨酸酯、邻氨基苯甲酸酯、薁、苝、吡啶、喹啉、硼聚氮杂引达省(borapolyazaindacene)(包括美国专利号4,774,339;5,187,288;5,248,782;5,274,113;以及5,433,896中披露的任何相应化合物)、呫吨(包括美国专利号6,162,931;6,130,101;6,229,055;6,339,392;5,451,343;5,227,487;5,442,045;5,798,276;5,846,737;4,945,
171;美国序列号09/129,015和09/922,333中披露的任何相应化合物)、噁嗪(包括美国专利号4,714,763中披露的任何相应化合物)或苯并噁嗪、卡巴嗪(包括美国专利号4,810,636中披露的任何相应化合物)、次联苯甲酮(phenalenone)、香豆素(包括美国专利号5,696,157;5,459,276;5,501,980以及5,830,912中披露的任何相应化合物)、苯并呋喃(包括美国专利号4,603,209和4,849,362中披露的任何相应化合物)、以及苯次联苯甲酮(benzphenalenone)(包括美国专利号4,812,409中披露的任何相应化合物)及其衍生物。如在此所用,噁嗪包括试卤灵(包括5,242,805中披露的任何相应化合物)、氨基噁嗪酮、二氨基噁嗪、以及其苯并取代的类似物。
[0330]
在一个具体实施例中,轭合至在此描述的抗体的荧光团包括呫吨(对甲氨基酚、若丹明、荧光素、以及其衍生物)、香豆素、花青、芘、噁嗪、以及硼聚氮杂引达省。在其他实施例中,这类荧光团是磺化的呫吨、氟化的呫吨、磺化的香豆素、氟化的香豆素、以及磺化的花青。还包括以下列商品名出售且通常已知为以下各项的染料:alexadylight、染料、oregonpacific blue
tm
、fam、fitc、以及rox
tm
。
[0331]
连接至抗体上的荧光团的选择将确定被轭合的抗体的吸收和荧光发射特性。可以用于抗体和抗体结合的配体的荧光团标记的物理特性包括但不限于:光谱特征(吸收、发射和斯托克斯位移)、荧光强度、寿命、极化以及光致漂白率、或其组合。所有这些物理特性可以用于将一个荧光团与另一个荧光团区分开,并且从而允许多元分析。在某些实施例中,荧光团在大于480nm的波长下具有最大吸收。在其他实施例中,荧光团在处于或接近488nm至514nm(特别适合用于由氩离子激光激发源的输出激发)或在接近546nm(特别适合用于由水银弧光灯激发)下吸收。在另一实施例中,荧光团可以针对组织或完整生物体应用在nir(近红外区域)中发射。荧光标记的其他所希望的特性可以包括细胞渗透性和低毒性,例如,如果将在细胞或生物体(例如,活体动物)中执行抗体的标记。
[0332]
在某些实施例中,酶是标记并且轭合至在此所述的抗体。因为可以获得可检测的信号的放大,从而得到提高的测定灵敏度,所以酶是合乎需要的标记物。酶自身不产生可检测的响应,但是当它被适当的底物接触时,起到分解底物的作用,这样使得被转化的底物产生荧光、比色或发光信号。因为标记试剂上的一种酶可以导致多个底物被转化成可检测的信号,所以酶将可检测的信号放大。选择酶底物以产生优选的可测量的产物,例如,比色、荧光或化学发光。这类底物在本领域中广泛使用且是本领域的技术人员熟知的。
[0333]
在一个实施例中,比色或荧光底物和酶组合使用氧化还原酶如辣根过氧化物酶和底物如3,3
’‑
二氨基联苯胺(dab)和3
‑
氨基
‑9‑
乙基咔唑(aec),这产生有区别的颜色(分别棕色和红色)。产生可检测的产物的其他比色氧化还原酶底物包括但不限于:2,2
‑
连氮基
‑
双(3
‑
乙基苯并噻唑啉
‑6‑
磺酸)(abts)、邻苯二胺(opd)、3,3’,5,5
’‑
l四甲基联苯胺(tmb)、邻联茴香胺、5
‑
氨基水杨酸、4
‑
氯
‑1‑
萘酚。荧光底物包括但不限于,高香草酸或4
‑
羟基
‑3‑
甲氧基苯乙酸;还原的吩噁嗪和还原的苯并噻嗪,包括红试剂及其变体(美国专利号4,384,042);和还原的二氢呫吨,包括二氢荧光素(美国专利号6,162,931);以及二氢若丹明,包括二氢若丹明123。为酪酰胺的过氧化物酶底物(美国专利号5,196,306;5,583,001和5,731,158)表示一类独特的过氧化物酶底物,在于它们可以在酶的作用之前固有地可检测,但在如酪酰胺信号放大(tsa)描述的过程中通过过氧化物酶的作用“固定在适当位置”。
这些底物广泛地用于标记样品(这些样品是细胞、组织或阵列)中的抗原以用于其随后通过显微术、流式细胞术、光学扫描和荧光测定法进行的检测。
[0334]
在另一个实施例中,比色(并且在一些情况下荧光)底物和酶组合使用磷酸酶(如酸性磷酸酶、碱性磷酸酶或这种磷酸酶的重组型式)与比色底物(如5
‑
溴
‑6‑
氯
‑3‑
吲哚基磷酸酯(bcip)、6
‑
氯
‑3‑
吲哚基磷酸酯、5
‑
溴
‑6‑
氯
‑3‑
吲哚基磷酸酯、对硝基苯基磷酸酯或邻硝基苯基磷酸酯)或与荧光底物(如4
‑
甲基伞形酮基磷酸酯、6,8
‑
二氟
‑7‑
羟基
‑4‑
甲基香豆素基磷酸酯(difmup,美国专利号5,830,912)、荧光素二磷酸酯、3
‑
邻甲基荧光素磷酸酯、试卤灵磷酸酯、9h
‑
(1,3
‑
二氯
‑
9,9
‑
二甲基吖啶
‑2‑
酮
‑7‑
基)磷酸酯(ddao磷酸酯)或elf 97、elf 39或相关磷酸酯(美国专利号5,316,906和5,443,986))的组合。
[0335]
糖苷酶,具体地说β
‑
半乳糖苷酶、β
‑
葡糖醛酸糖苷酶和β
‑
葡糖苷酶是另外适合的酶。适当的比色底物包括但不限于,5
‑
溴
‑4‑
氯
‑3‑
吲哚基β
‑
d
‑
吡喃半乳糖苷(x
‑
gal)和类似的吲哚基半乳糖苷、葡糖苷和葡糖苷酸、邻硝基苯基β
‑
d
‑
吡喃半乳糖苷(onpg)、以及对硝基苯基β
‑
d
‑
吡喃半乳糖苷。在一个实施例中,荧光底物包括试卤灵β
‑
d
‑
吡喃半乳糖苷、荧光素二半乳糖苷(fdg)、荧光素二葡糖苷酸以及它们的结构变体(美国专利号5,208,148;5,242,805;5,362,628;5,576,424以及5,773,236)、4
‑
甲基伞形酮基lβ
‑
d
‑
吡喃半乳糖苷、羧基伞形酮基β
‑
d
‑
吡喃半乳糖苷、以及氟化香豆素β
‑
d
‑
吡喃半乳糖苷(美国专利号5,830,912)。
[0336]
另外的酶包括但不限于,水解酶如胆碱酯酶和肽酶、氧化酶如葡萄糖氧化酶和细胞色素氧化酶、以及还原酶,对于这些酶适合的底物是已知的。
[0337]
酶以及其产生化学发光的适当的底物优选用于一些测定。这些包括但不限于天然和重组形式的荧光素酶和水母发光蛋白。磷酸酶、糖苷酶和氧化酶的产生化学发光的底物,如含有稳定的二氧杂环丁烷、鲁米诺、异氨基苯二酰肼以及吖啶酯的那些也是有用的。
[0338]
在另一个实施例中,也利用如生物素的半抗原作为标记。生物素是有用的,这是因为它可以在酶系统中起作用以进一步放大可检测的信号并且它可以充当一个有待在亲和色谱中使用用于分离目的的标签。出于检测目的,使用对于生物素具有亲和力的酶轭合物,如抗生物素蛋白
‑
hrp。随后添加一种过氧化物酶底物以产生可检测的信号。
[0339]
半抗原还包括激素、天然存在和合成的药物、污染物、过敏原、影响分子、生长因子、趋化因子、细胞因子、淋巴因子、氨基酸、肽、化学中间体、核苷酸及类似物。
[0340]
在某些实施例中,荧光蛋白可以轭合至抗体作为标记。荧光蛋白的实例包括绿色荧光蛋白(gfp)和藻胆蛋白以及其衍生物。荧光蛋白,特别是藻胆蛋白,对于产生叠层染料标记的标记试剂尤其有用。出于获得较大的斯托克斯位移的目的,这些叠层染料包含荧光蛋白和荧光团,其中发射光谱从荧光蛋白的吸收光谱的波长发生更远的位移。对于检测样品中的较低量的抗原来说这是特别有利的,其中发射的荧光被最大限度地优化,换言之很少至没有发射光被荧光蛋白重吸收。为了实现这一点,荧光蛋白和荧光团充当能量转移对,其中荧光蛋白在荧光团所吸收的波长下发射,并且荧光团随后在远离仅存在荧光蛋白的情况下所能够获得的波长的波长下发射。特别有用的组合是美国专利号4,520,110;4,859,582;5,055,556中披露的藻胆蛋白和美国专利号5,798,276中披露的磺基若丹明荧光团,或美国专利号6,977,305和6,974,873中披露的磺化花青荧光团;或美国专利号6,130,101中披露的磺化呫吨衍生物,以及美国专利号4,542,104中披露的那些组合。可替代地,荧光团充当能量供体并且荧光蛋白是能量受体。
[0341]
在某些实施例中,标记是放射性同位素。适合的放射性材料的实例包括但不限于:碘(
121
i、
123
i、
125
i、
131
i)、碳(
14
c)、硫(
35
s)、氚(3h)、铟(
111
in、
112
in、
113m
in、
115m
in)、锝(
99
tc、
99m
tc)、铊(
201
ti)、镓(
68
ga、
67
ga)、钯(
103
pd)、钼(
99
mo)、氙(
135
xe)、氟(
18
f)、
153
sm,
177
lu、
159
gd、
149
pm、
140
la、
175
yb、
166
ho、
90
y、
47
sc、
186
re、
188
re、
142
pr、
105
rh、以及
97
ru。
[0342]
医学治疗和用途
[0343]
本发明的抗体及其结合片段以及其变体可以用于治疗甲型流感病毒感染、用于预防甲型流感病毒感染和/或用于甲型流感病毒感染的检测、诊断和/或预后。
[0344]
诊断方法可以包括使抗体或抗体片段与样品相接触。这类样品可以是从例如鼻腔通道、鼻窦腔、唾液腺、肺、肝、胰腺、肾、耳、眼、胎盘、消化道、心脏、卵巢、垂体、肾上腺、甲状腺、脑或皮肤取得的组织样品。检测、诊断和/或预后方法还可以包括检测抗原/抗体复合物。
[0345]
在一个实施例中,本发明提供一种通过向受试者给予有效量的根据本发明的抗体或其结合片段或包含该抗体或其结合片段的药用组合物来治疗该受试者的方法。在一个实施例中,该抗体或其结合片段是基本上纯化的(即,基本上不含限制其作用或产生不希望的副作用的物质)。在一个实施例中,本发明的抗体或其结合片段在暴露后给予,或在受试者已经暴露于甲型流感病毒或感染甲型流感病毒之后给予。在另一个实施例中,本发明的抗体或其结合片段在暴露前给予,或给予至尚未暴露于甲型流感病毒或尚未感染甲型流感病毒的受试者。在一个实施例中,本发明的抗体或其结合片段被给予至针对一种或多种甲型流感亚型血清反应阴性的受试者。在另一个实施例中,本发明的抗体或其结合片段被给予至针对一种或多种甲型流感亚型血清反应阳性的受试者。在一个实施例中,本发明的抗体或其结合片段被给予至在感染或症状发作的1、2、3、4、5天内的受试者。在另一个实施例中,本发明的抗体或其结合片段可以被给予至在感染或症状发作之后1、2、3、4、5、6或7天之后的受试者和在感染或症状发作之后2、3、4、5、6、7、8、9或10天之内的受试者。
[0346]
在一个实施例中,该方法减轻受试者的甲型流感病毒感染。在另一个实施例中,该方法预防受试者的甲型流感病毒感染、降低受试者的甲型流感病毒感染的风险或延迟受试者的甲型流感病毒感染。在一个实施例中,受试者是哺乳动物。在一个更具体的实施例中,受试者是人。在一个实施例中,受试者包括但不限于,特别处于甲型流感病毒感染的风险或易患甲型流感病毒感染的受试者,例如,免疫功能低下的受试者。
[0347]
治疗可以是单剂量方案或多剂量方案,并且本发明的抗体或其结合片段可以用于被动免疫。
[0348]
在一个实施例中,本发明的抗体或其结合片段与一种或多种抗病毒药物组合给予至受试者。在一个实施例中,本发明的抗体或其结合片段与一种或多种小分子抗病毒药物组合给予至受试者。小分子抗病毒药物包括神经氨酸酶抑制剂,如奥司他韦扎那米韦以及金刚烷,如金刚烷胺和金刚乙胺。
[0349]
在另一个实施例中,本发明提供一种用作用于预防或治疗甲型流感病毒感染的药剂的组合物。在另一个实施例中,本发明提供本发明的抗体或其结合片段和/或包含本发明的抗体或其结合片段所结合的表位的蛋白质在制造用于治疗受试者和/或诊断受试者的药剂中的用途。
[0350]
如在本发明中描述的抗体及其片段还可以在用于诊断甲型流感病毒感染的试剂
盒中使用。此外,能够结合本发明的抗体的表位可以用于试剂盒中,该试剂盒用于通过检测保护性抗甲型流感病毒抗体的存在来监测疫苗接种程序的功效。如在本发明中描述的抗体、抗体片段或其变体和衍生物还可以在用于监测具有所希望的免疫原性的疫苗制造的试剂盒中使用。
[0351]
本发明还提供一种制备药用组合物的方法,该方法包括将单克隆抗体与一种或多种药学上可接受的载体掺混的步骤,其中该抗体是在此所述的根据本发明的单克隆抗体。
[0352]
各种递送系统是已知的并且可以用于给予本发明的抗体或其结合片段,包括但不限于,封装在脂质体中、微颗粒、微胶囊、能够表达该抗体或抗体片段的重组细胞、受体介导的内吞作用、构建作为逆转录病毒或其他载体的一部分的核酸、通过电穿孔递送技术递送裸核苷酸(如描述于穆舒马尼(muthumani)等人,公共科学图书馆
·
综合(plos one.)2013年12月31日;8(12):e84234.doi:10.1371/journal.pone.0084234.ecollection 2013)等。引入方法包括但不限于,皮内、肌肉内、腹膜内、静脉内、皮下、鼻内、硬膜外、以及口服途径。可以通过任何方便的途径给予这些组合物,例如通过输注或单次快速静脉注射(bolus injection)、通过经由上皮或皮肤粘膜内层(例如口腔粘膜、直肠和肠道粘膜等)吸收,并且可以将其与其他生物活性剂(包括但不限于小分子抗病毒组合物)一起给予。给予可以是全身性的或局部的。也可以例如通过使用吸入器或雾化器以及具有雾化剂的制剂而采用肺部给药。在又另一个实施例中,可以在控制释放系统中递送该组合物。
[0353]
本发明还提供药用组合物。这类组合物包含治疗有效量的本发明的抗体或其结合片段以及药学上可接受的载体。如在此所用的术语“药学上可接受的”意指由联邦政府或州政府的监管机构批准的或在美国药典或其他普遍认可的药典中列出的,用于在动物体内并且更具体地在人体内使用。术语“载体”是指与治疗剂一起给予的稀释剂、佐剂、赋形剂或媒介物。此类药物载体可以为无菌液体,如水和油,包括石油、动物、植物或合成来源的油,如花生油、豆油、矿物油、芝麻油等。当静脉内给予药用组合物时,水是优选的载体。还可以将盐水溶液、水性右旋糖和甘油溶液用作液体载体,特别是用于注射溶液。适合的药用赋形剂包括淀粉、葡萄糖、乳糖、蔗糖、明胶、麦芽、米、面粉、白垩、硅胶、硬脂酸钠、单硬脂酸甘油酯、滑石、氯化钠、脱脂奶粉、甘油、丙烯、二醇、水、乙醇以及类似物。如果希望的话,该组合物还可包含少量湿润剂或乳化剂或ph缓冲剂。这些组合物可以采取以下形式:溶液、悬浮液、乳液、片剂、丸剂、胶囊、粉剂、持续释放配制品以及类似物。该组合物可以与传统的结合剂和载体如甘油三酯一起配制为栓剂。口服配制品可以包含标准载体,如药物级甘露醇、乳糖、淀粉、硬脂酸镁、糖精钠、纤维素、碳酸镁等。在一个实施例中,该药物组合物含有治疗有效量的抗体或其结合片段(优选呈纯化形式)连同适合量的载体以便提供用于正确给予至患者的形式。该制剂应该符合施用方式的要求。
[0354]
典型地,对于抗体治疗剂,给予至患者的剂量在约0.1mg/kg至100mg/kg患者体重之间。
附图说明
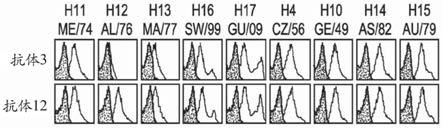
[0355]
图1a示出抗体3和抗体12与亚型h11、h12、h13、h16、h17、h4、h10、h14、以及h15的表面表达的ha蛋白的结合。直方图描绘细胞的数目对比与ha转染的细胞(白色)或模拟转染的细胞(灰色)结合的抗体的荧光强度。图1b示出在抗体3、抗体12或mpe8v3(不相关的病毒融
合蛋白抗体)(科尔蒂d(corti d)等人,2013,自然501)存在下鸡红细胞与a/波多黎各/8/34的低ph诱导的融合的抑制百分比。图1c和图1d示出在用胰蛋白酶消化5、10或20分钟之后未裂解的(ha0)、重组h1 ha的免疫印迹。消化反应包括在1c中单独ha(输入)抑或用fi6v4(披露于wo2013/011347a1中)、抗体3、fe17.23(球状头部特异性mab)(科尔蒂d等人,2010,临床研究杂志(j clin invest)120)或不相关的对照抗体(对照igg)预先处理的ha,并且在1d中单独ha(输入)或用抗体3、抗体12、抗体14或不相关的对照抗体(对照igg)预先处理的ha。
[0356]
图2示出在增加量的抗体3、抗体11、抗体12以及抗体14存在下a/hk/8/68感染的mdck细胞的nk细胞介导的杀伤的百分比。
[0357]
图3示出在增加量的抗体3、抗体11、抗体12以及抗体14或不相关的同种型对照(对照igg)存在下吞噬表达a/hk/8/68ha的mdck靶细胞的巨噬细胞的百分比。
[0358]
图4示出在增加量的抗体3、抗体11、抗体12以及抗体14存在下a/pr/8/34感染的mdck细胞的补体依赖性杀伤的百分比。
[0359]
图5示出当在用致死剂量的h1n1流感病毒感染之前4小时将不同浓度的抗体3或不相关的对照抗体(对照igg)给予至小鼠时,研究的每组中存活动物的百分比。
[0360]
图6示出其中在用致死剂量的h3流感病毒感染之前4小时将不同浓度的抗体3或不相关的对照抗体(对照igg)给予至小鼠的研究的每组中存活动物的百分比。
[0361]
图7示出当将小鼠用致死剂量的h1n1流感病毒感染并且在不同时间点(感染后第1天和第2天)用30mg/kg的抗体3或不相关的对照抗体(对照igg)处理时,每组中存活动物的百分比。
[0362]
图8示出其中将小鼠用致死剂量的h3流感病毒感染并且在不同时间点(感染后第3天、第4天和第5天)用30mg/kg的抗体3或不相关的对照抗体(对照igg)处理的研究的每组中存活动物的百分比。
[0363]
图9示出其中将小鼠用致死剂量的h1n1流感病毒感染并且在感染后第1天用2mg/kg的抗体3、抗体11、抗体12、抗体14或不相关的对照抗体(对照igg)处理的研究的每组中存活动物的百分比。
[0364]
图10示出其中将小鼠用致死剂量的h3流感病毒感染并且在感染后第2天用3mg/kg的抗体3、抗体11、抗体12、抗体14或不相关的对照抗体(对照igg)处理的研究的每组中存活动物的百分比。
[0365]
图11示出其中将小鼠用致死剂量的h1n1流感病毒感染,并且25mg/kg bid奥司他韦持续5天、10mg/kg的抗体12或10mg/kg的不相关的对照抗体(对照igg)的处理在不同时间点(在感染之前4小时、感染后第1天或感染后2天)开始的研究的每组中存活动物的百分比。
[0366]
图12示出其中将小鼠用致死剂量的h3流感病毒感染,并且25mg/kg口服奥司他韦每日两次(bid)持续5天、或单一剂量10mg/kg的抗体12或10mg/kg的不相关的对照抗体(对照igg)的处理在不同时间点(感染后第1、2、3、或4天)开始的研究的每组中存活动物的百分比。
[0367]
图13示出将小鼠用致死剂量的h3流感病毒感染且在感染后2天用2.5mg/kg或0.3mg/kg单一剂量的抗体12、25mg/kg的奥司他韦bid持续5天、或2.5mg/kg或0.3mg/kg的抗体12与25mg/kg的奥司他韦bid持续5天的组合处理的研究中的每组中存活动物的百分比。
[0368]
图14示出在用致死剂量的h5n1流感病毒感染且在感染后第1、2或3天用25mg/kg单
id no.:98的lcdr1、seq id no.:99的lcdr2、以及seq id no.:100的lcdr3;
[0386]
(k)seq id no.:103的hcdr1、seq id no.:104的hcdr2、seq id no.:105的hcdr3、seq id no.:108的lcdr1、seq id no.:109的lcdr2、以及seq id no.:110的lcdr3;
[0387]
(l)seq id no.:113的hcdr1、seq id no.:114的hcdr2、seq id no.:115的hcdr3、seq id no.:118的lcdr1、seq id no.:119的lcdr2、以及seq id no.:110的lcdr3;
[0388]
(m)seq id no.:123的hcdr1、seq id no.:124的hcdr2、seq id no.:125的hcdr3、seq id no.:128的lcdr1、seq id no.:129的lcdr2、以及seq id no.:130的lcdr3;
[0389]
(n)seq id no.:133的hcdr1、seq id no.:134的hcdr2、seq id no.:135的hcdr3、seq id no.:138的lcdr1、seq id no.:139的lcdr2、以及seq id no.:140的lcdr3;以及
[0390]
(o)seq id no.:143的hcdr1、seq id no.:144的hcdr2、seq id no.:145的hcdr3、seq id no.:148的lcdr1、seq id no.:149的lcdr2、以及seq id no.:150的lcdr3
[0391]
(p)根据(a)至(o)中任一项的一组六个cdr包含一个或多个氨基酸取代、缺失或插入;
[0392]
(q)根据(a)至(p)中任一项的一组六个cdr包含1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、或24或25个氨基酸取代;
[0393]
(r)根据(a)至(q)中任一项的一组六个cdr hcdr1、hcdr2、hcdr3、lcdr1、lcdr2、lcdr3包含:
[0394]
(i)hcdr1,其具有与seq id no:3一致或相对于seq id no:3包含3个或更少个氨基酸残基取代的氨基酸序列;
[0395]
(ii)hcdr2,其具有与seq id no:4一致或相对于seq id no:4包含5个或更少个氨基酸残基取代的氨基酸序列;
[0396]
(iii)hcdr3,其具有与seq id no:5一致或相对于seq id no:5包含6个或更少个氨基酸残基取代的氨基酸序列;
[0397]
(iv)lcdr1,其具有与seq id no:6一致或相对于seq id no:6包含5个或更少个氨基酸残基取代和/或一个缺失的氨基酸序列;
[0398]
(v)lcdr2,其具有与seq id no:7一致或相对于seq id no:7包含5个或更少个氨基酸残基取代的氨基酸序列;以及
[0399]
(vi)lcdr3,其具有与seq id no:8一致或相对于seq id no:8包含1个或更少个氨基酸残基取代的氨基酸序列;
[0400]
(s)根据(a)至(r)中任一项的一组六个cdr hcdr1、hcdr2、hcdr3、lcdr1、lcdr2、lcdr3包含:
[0401]
(i)hcdr1,其中:
[0402]
kabat残基31为s,
[0403]
kabat残基32为n或y,
[0404]
kabat残基33为n、s或r,
[0405]
kabat残基34为a,
[0406]
kabat残基35为v或t,
[0407]
kabat残基35a为w
[0408]
kabat残基35b为n;
[0409]
(ii)hcdr2,其中:
[0410]
kabat残基50为r,
[0411]
kabat残基51为t,
[0412]
kabat残基52为y,
[0413]
kabat残基52a为y,
[0414]
kabat残基53为r,
[0415]
kabat残基54为s,
[0416]
kabat残基55为k或g,
[0417]
kabat残基56为w,
[0418]
kabat残基57为y,
[0419]
kabat残基58为n或y,
[0420]
kabat残基59为d,
[0421]
kabat残基60为y,
[0422]
kabat残基61为a,
[0423]
kabat残基62为e、v或d,
[0424]
kabat残基63为s或f,
[0425]
kabat残基64为v或l,
[0426]
kabat残基65为k;
[0427]
(iii)hcdr3,其中:
[0428]
kabat残基95为s或g,
[0429]
kabat残基96为g,
[0430]
kabat残基97为h,
[0431]
kabat残基98为i,
[0432]
kabat残基99为t,
[0433]
kabat残基100为v或e,
[0434]
kabat残基100a为f,
[0435]
kabat残基100b为g,
[0436]
kabat残基100c为v或l,
[0437]
kabat残基100d为n,
[0438]
kabat残基100e为v或i,
[0439]
kabat残基100f为d,
[0440]
kabat残基100g为a,
[0441]
kabat残基100f为f或y,
[0442]
kabat残基101为d,
[0443]
kabat残基102为m、i或v;
[0444]
(iv)lcdr1,其中:
[0445]
kabat残基24为r,
[0446]
kabat残基25为t、a或不存在,
[0447]
kabat残基26为s或a,
[0448]
kabat残基27为q,
[0449]
kabat残基28为s或r,
[0450]
kabat残基29为l,
[0451]
kabat残基30为s、n或r
[0452]
kabat残基31为s,
[0453]
kabat残基32为y,
[0454]
kabat残基33为l、t或d,
[0455]
kabat残基34为h;
[0456]
(v)lcdr2,其中:
[0457]
kabat残基50为a,
[0458]
kabat残基51为a、t或s,
[0459]
kabat残基52为s或t,
[0460]
kabat残基53为s或t,
[0461]
kabat残基54为l或r,
[0462]
kabat残基55为q、l或g,
[0463]
kabat残基56为s;以及,
[0464]
(vi)lcdr3,其中:
[0465]
kabat残基89为q,
[0466]
kabat残基90为q或l;
[0467]
kabat残基91为s,
[0468]
kabat残基92为r,并且
[0469]
kabat残基93为t。
[0470]
6.根据以上实施方案中任一项所述的抗体或其结合片段,包含与选自下组的vh具有至少75%一致性的vh和/或与选自下组的vl具有至少75%一致性的vl,该组由以下各项组成:
[0471]
(a)seq id no.:2的vh和seq id no.:7的vl,
[0472]
(b)seq id no.:12的vh和seq id no.:17的vl,
[0473]
(c)seq id no.:22的vh和seq id no.:27的vl,
[0474]
(d)seq id no.:32的vh和seq id no.:37的vl,
[0475]
(e)seq id no.:42的vh和seq id no.:47的vl,
[0476]
(f)seq id no.:52的vh和seq id no.:57的vl,
[0477]
(g)seq id no.:62的vh和seq id no.:67的vl,
[0478]
(h)seq id no.:72的vh和seq id no.:77的vl,
[0479]
(i)seq id no.:82的vh和seq id no.:87的vl,
[0480]
(j)seq id no.:92的vh和seq id no.:97的vl,
[0481]
(k)seq id no.:102的vh和seq id no.:107的vl,
[0482]
(l)seq id no.:112的vh和seq id no.:117的vl,
[0483]
(m)seq id no.:122的vh和seq id no.:127的vl,
[0484]
(n)seq id no.:132的vh和seq id no.:137的vl,
[0485]
(o)seq id no.:144的vh和seq id no.:147的vl以及
[0486]
(p)seq id no:152的vh和seq id no:157的vl。
[0487]
7.根据以上实施方案中任一项所述的抗体或其结合片段,包含选自下组的vh和vl,该组由以下各项组成:
[0488]
(a)seq id no.:2的vh和seq id no.:7的vl,
[0489]
(b)seq id no.:12的vh和seq id no.:17的vl,
[0490]
(c)seq id no.:22的vh和seq id no.:27的vl,
[0491]
(d)seq id no.:32的vh和seq id no.:37的vl,
[0492]
(e)seq id no.:42的vh和seq id no.:47的vl,
[0493]
(f)seq id no.:52的vh和seq id no.:57的vl,
[0494]
(g)seq id no.:62的vh和seq id no.:67的vl,
[0495]
(h)seq id no.:72的vh和seq id no.:77的vl,
[0496]
(i)seq id no.:82的vh和seq id no.:87的vl,
[0497]
(j)seq id no.:92的vh和seq id no.:97的vl,
[0498]
(k)seq id no.:102的vh和seq id no.:107的vl,
[0499]
(l)seq id no.:112的vh和seq id no.:117的vl,
[0500]
(m)seq id no.:122的vh和seq id no.:127的vl,
[0501]
(n)seq id no.:132的vh和seq id no.:137的vl,
[0502]
(o)seq id no.:144的vh和seq id no.:147的vl以及
[0503]
(p)seq id no:152的vh和seq id no:157的vl。
[0504]
8.根据以上实施方案中任一项所述的抗体或其结合片段,其中该抗体或结合片段选自下组,该组由以下各项组成:免疫球蛋白分子、单克隆抗体、嵌合抗体、cdr移植抗体、人源化抗体、fab、fab'、f(ab')2、fv、二硫键连接的fv、scfv、单结构域抗体、双抗体、多特异性抗体、双重特异性抗体、以及双特异性抗体。
[0505]
9.根据以上实施方案中任一项所述的抗体或其结合片段,其中该vh包含人种系框架vh6
‑
1,该vl包含人种系框架vk1
‑
39,以及其组合。
[0506]
10.根据以上实施方案中任一项所述的抗体或其结合片段,包含fc区。
[0507]
11.根据以上实施方案中任一项所述的抗体或其结合片段,其中该抗体是igg1、igg2或igg4或其片段。
[0508]
12.一种针对甲型流感病毒的抗体或其结合片段,该抗体或其结合片段能够结合甲型流感病毒血凝素并且中和甲型流感病毒的至少一种1组亚型和至少一种2组亚型,其中该抗体或其结合片段结合在选自h1、h2、h5、h6、h8、h9、h11、h12、h13和h16的一种或多种甲型流感病毒1组亚型以及选自h3、h4、h7、h10、h14和h15的一种或多种2组亚型之中保守的表位。
[0509]
13.一种针对甲型流感病毒的抗体或其结合片段,该抗体或其结合片段能够结合甲型流感病毒血凝素并且中和甲型流感病毒的至少一种1组亚型和至少一种2组亚型,其中该抗体或其结合片段结合位于ha2的保守柄区中的表位。
[0510]
14.根据实施方案12所述的抗体或其结合片段,其中该表位包含选自ha2的根据h3编号系统的位置18、19、42和45的一个或多个氨基酸。
[0511]
15.一种针对甲型流感病毒的抗体或其结合片段,该抗体或其结合片段能够结合甲型流感病毒血凝素并且中和甲型流感病毒的至少一种1组亚型和至少一种2组亚型,该抗体或其结合片段与根据以上实施方案中任一项所述的抗体结合相同表位或竞争结合甲型流感病毒血凝素。
[0512]
16.根据实施方案14所述的抗体或其结合片段,其中该抗体或结合片段与具有seq id no:112中所示的氨基酸序列的抗体结合相同表位或竞争结合甲型流感病毒血凝素。
[0513]
17.一种分离的核酸,该分离的核酸编码根据实施方案1至16中任一项所述的抗体或其结合片段。
[0514]
18.一种载体,该载体包含根据实施方案17所述的分离的核酸。
[0515]
19.一种宿主细胞,该宿主细胞包含根据实施方案17所述的核酸或根据实施方案18所述的载体。
[0516]
20.一种用于制造根据实施方案1至16中任一项所述的抗体或其结合片段的方法,该方法包括在适合用于表达该抗体或其片段的条件下培养根据实施方案19所述的宿主细胞。
[0517]
21.根据实施方案20所述的方法,进一步包括从该宿主细胞培养物分离该抗体或其结合片段。
[0518]
22.一种组合物,该组合物包含根据实施方案1至16中任一项所述的抗体或其结合片段以及药学上可接受的载体。
[0519]
23.一种组合物,该组合物包含根据实施方案1至16中任一项所述的抗体或其结合片段、在ph 6.0的25mm his和0.15m nacl。
[0520]
24.根据实施方案1至16中任一项所述的抗体或其结合片段,该抗体或其结合片段用于在预防或治疗受试者的甲型流感感染中使用。
[0521]
25.根据实施方案1至16中任一项所述的抗体或其结合片段的用途,该抗体或其结合片段用于制造用以预防或治疗受试者的甲型流感感染的药剂。
[0522]
26.一种用于预防或治疗受试者的甲型流感感染的方法,该方法包括向该受试者给予有效量的根据实施方案1至16中任一项所述的抗体或其结合片段。
[0523]
27.一种用于预防或治疗受试者的甲型流感感染的方法,该方法包括给予有效量的根据实施方案1至16中任一项所述的抗体或其结合片段与小分子抗病毒组合物的组合。
[0524]
28.根据实施方案27所述的方法,其中该小分子抗病毒组合物是神经氨酸酶抑制剂或金刚烷。
[0525]
29.根据实施方案27所述的方法,其中该小分子抗病毒组合物选自奥塞米韦、扎那米韦、金刚烷胺、金刚乙胺、以及其组合。
[0526]
30.根据实施方案1至16中任一项所述的抗体或其片段的用途,该抗体或其片段用于体外诊断受试者的甲型流感感染。
[0527]
实例
[0528]
实例1.从记忆b细胞分离的人单克隆抗体的构建和优化
[0529]
从针对异种亚型抗体的高效价选择的供体的冷冻保存的外周血单核细胞(pbmc)分选cd22+igg+b细胞且使用埃
‑
巴二氏病毒(ebv)和cpg寡脱氧核苷酸2006以及饲养细胞以3个细胞/孔永生化。在14天之后收获含有抗体的培养上清液并且通过elisa结合测定进行
筛选以便测定分别针对h5(a/越南/1203/04)和h7(a/nld/03)血凝素(ha)的结合活性。四种b细胞克隆(抗体1、抗体4、抗体7、以及抗体9)被发现特异性地结合两种ha并且因此被收集。对这些克隆的vh基因和vl基因进行测序并且据发现根据使用kabat数据库对对vh和vl v、d、以及j片段进行的同源性分析克隆地相关。值得注意的是,抗体4的vh据发现在编码缬氨酸(在抗体5中编码的)或谷氨酸(在抗体6中编码的)的hcdr3中具有简并核苷酸位点。将将四种抗体的vh基因和vl基因克隆至igg1表达载体中(进行较小序列修饰以便有助于克隆和或密码子优化产生五种抗体;抗体3、抗体5、抗体6、抗体8、以及抗体10;用于以下实例中)并且通过瞬时转染源自hek或cho细胞的哺乳动物细胞系来产生重组抗体。在培养7
‑
10天之后收集来自转染的细胞的上清液,并且通过蛋白质a色谱法亲和力纯化igg,且渗析至pbs中。将抗体3进一步优化以产生变体,其中将位于框架区中的非种系编码的体细胞突变改变为种系编码的氨基酸,并且使cdr区经受简约诱变。如上所述表达含有不同突变的完整igg构建体并且通过elisa筛选粗上清液以便选择对h3和h1 ha蛋白具有增加的结合活性的克隆。使用0.15μg/ml的兔抗人igg的包被浓度进行elisa以便捕获且标准化来自上清液的igg,并且然后添加0.5μg/ml的生物素化ha亚型h1(a/加利福利亚/7/04(h1n1))或亚型h3(a/珀斯/16/09(h3n2))且孵育一小时。通过添加链霉抗生物素蛋白
‑
hrp(1:5000)来检测结合,并且在450nm下读取显影吸光度。将赋予更好结合的有益单一突变组合且克隆至组合文库中,这些突变如上所述表达且通过elisa进行筛选。该文库方法导致产生5种另外的抗体3变体,对这些变体进行进一步表征(抗体11
‑
15)。
[0530]
实例2.抗ha中和抗体(nab)结合不同亚型的ha
[0531]
为了测试抗ha抗体的表位是否在不同亚型的ha之中为保守的,进行ha交叉反应性elisa结合测定。在4℃下将384孔maxisorb elisa板(nunc)用pbs中的0.5ug/ml重组ha(rha)、亚型h1(a/加利福利亚/7/09(h1n1))、亚型h2(a/猪/mo/06(h2n3))、亚型h3(a/珀斯/16/09(h3n2))、亚型h5(a/越南/1203/04(h5n1))、亚型h6(a/水鸭/hk/w312/97(h6n1))、亚型h7(a/netherlands/219/03(h7n7))、以及亚型h9(a/鸡/hk/g9/97(h9n2))包被过夜。将板用含有0.1%v/v吐温
‑
20的pbs洗涤以便去除未包被的蛋白质并且随后添加含有1%(w/v)酪蛋白(赛默科技公司(thermo scientific))的封闭溶液且在室温下孵育1小时。将封闭溶液废弃且添加封闭溶液中的3倍连续稀释的抗ha抗体(酪蛋白
‑
pbs(赛默科技公司)且在室温下孵育1小时。将板洗涤三次并且使用过氧化物酶轭合的小鼠抗人igg抗体(杰克逊(jackson))检测结合的抗体。通过在添加supersignal pico底物(赛默科技公司)之后测量化学发光信号抑或通过在与四甲基联苯胺(tmb)单组分底物(kpl)孵育、接着添加2n硫酸以便终止反应之后测量在450nm下的颜色变化来计算抗体的结合活性。
[0532]
表1
[0533][0534]
表1示出所测试的所有抗ha igg结合至亚型h1、h2、h3、h5、h6、h9、以及h7的重组ha。在所测试的最高抗体浓度(6ug/ml)下,亚型h9的重组ha由抗体3和抗体10识别,但不由抗体5、抗体6和抗体8识别。这指示这些抗ha igg中的大多数的表位在不同亚型的ha分子之中是保守的。
[0535]
表2
[0536][0537]
表2示出所测试的所有抗ha igg变体结合至1组亚型h1、h2、h5、h6、以及h9的重组ha,具有类似的ec
50
值。所有变体结合至2组ha蛋白(h3和h7),然而,相较于抗体12
‑
15,抗体11和抗体3显示降低的活性与增加的ec
50
值。
[0538]
为了将这些结合结果扩展至包括更多不同的ha亚型,使用与ha转染的细胞的基于流式细胞术的结合进行了另外的结合研究。在该测定中,将hek细胞用亚型h4 a/鸭/捷克斯洛伐克/56(h4n6))、亚型h10(a/鸡/德国/n49(h10n7))、亚型h11(a/鸭/孟斐斯/546/74(h11n9))、亚型h12(a/鸭/艾伯塔/60/76(h12n5))、亚型h13(a/鸥/马里兰/704/77(h13n6))、亚型h14(a/野鸭/阿斯特拉罕/263/82(h14n5))、亚型h15(a/海鸥/西澳大利亚/2576/79(h15n9))、亚型h16(a/黑头鸥/瑞典/2/99(h16n3))、以及亚型h17(a/黄肩蝠/危地马拉/164/2009(h17n10))的表达全长野生型ha的质粒瞬时转染。在转染之后四十八小时,将细胞用胰蛋白酶分散,并且与5ug/ml的抗体3或抗体12在冰上孵育1小时。在该小时孵育之后,然后将结合至细胞表面表达的ha蛋白的抗体用山羊抗人igg daylight 649(杰克逊免疫研究公司(jackson immunoresearch))染色,并且将其通过流式细胞术进行检测。图1a示出当抗体结合至表达来自这些亚型中的每种的ha的细胞(白色)对模拟转染的细胞(灰色)时荧光强度的移动。抗体3结合至所测试的所有ha,除了h12以外,而抗体12结合至来自
两组(1组h11、h12、h13、h16、和h17以及2组h4、h10、h14和h15)的所测试的所有ha。
[0539]
实例3通过使用octet进行的与抗体3和抗体5igg1结合的ha的动力学表征
[0540]
在384倾斜孔板中使用fortebio octet qk 384动力学分析仪(门洛帕克(menlo park),加利福利亚州(ca))进行亲和力测量。将所有试剂在octet动力学缓冲液(fortebio公司)中稀释。将不同亚型:亚型h1(a/加利福利亚/7/04(h1n1))和亚型h3(a/珀斯/16/09(h3n2))的his标记的ha以10μg/ml固定到抗his传感器上。然后在来自100nm的2倍稀释液加零mab对照中监测抗ha mab缔合/解离。
[0541]
将缔合和解离原始数据针对零mab对照中的任何漂移进行校正,并且然后输出至graphpadprism(圣地亚哥(san diego),加利福利亚州)以用于进行亲和力曲线拟合。使用全局缔合/解离拟合来对数据进行拟合,其中所施加的限制为>5
×
10
‑6sec
‑1。如表3中所示,两种抗体均在pm水平下对结合h1具有非常高的亲和力,其中在检测限下具有慢解离速率。在亚
‑
nm水平下在h3三聚体情况下观察到两种抗体的相似k
on
、k
off
、以及kd。
[0542]
表3
[0543][0544]
实例4针对不同病毒亚型的抗ha igg1的体外交叉反应中和活性
[0545]
微量中和测定(mna)是从先前描述的加速的病毒抑制测定修改的,使用神经氨酸酶活性(na)作为读出(hassantoufighi,a.等人,2010,疫苗(vaccine)28:790)。简言之,对在补充有抗生素、谷氨酰胺(完全mem培养基)以及10%(v/v)胎牛血清的mem培养基(英杰公司)中培养的mdck细胞进行mna。将60tcid
50
(50%组织培养物感染剂量)的病毒添加至在384孔板中一式两份孔中的含有0.75ug/ml胰蛋白酶(沃辛顿(worthington))的完全mem培养基中的抗体的三倍稀释液,在室温下30分钟孵育之后,将2
×
104个细胞/孔添加至该板。在33℃ 5%co2孵育器中孵育大约40小时之后,通过将荧光标记的底物甲基伞形酮基
‑
n
‑
乙酰基神经氨酸(mu
‑
nana)(西格玛公司)添加至每个孔并且在37℃下孵育1小时来测量na活性。使用以下设置通过在荧光计envison(珀金埃尔默公司(perkinelmer))中读取荧光来定量由na活性表示的病毒复制:激发355nm,发射460nm;每孔10次发光。中和滴度(50%抑制浓度[ic
50
])被表示为相较于细胞对照孔使荧光信号降低50%的最终抗体浓度。表4和5显示抗ha抗体中和以下测试的不同亚型的甲型流感病毒:h1
‑
pr34(a/波多黎各/8/34(h1n1));h1
‑
pr34
‑
or(含有赋予奥司他韦耐药性的na 274y(n2编号)突变的a/波多黎各/8/34(h1n1));h1
‑
fm47(a/蒙默斯堡(fort monmouth)/1/47(h1n1));h1
‑
nj76(a/新泽西州/8/76(h1n1));h1
‑
kaw86(a/川崎/9/86(h1n1));h1
‑
tx91(caa/德克萨斯州/36/91(h1n1));h1
‑
bj95(ca a/北京/262/95(h1n1));h1
‑
ncal99(ca a/新喀里多尼亚(new caledonia/20/99)(h1n1));h1
‑
sd07(ca a/南达科他州(south dakota/6/07)(h1n1));h1
‑
ca09(ca a/加利福利亚州/7/09(h1n1));h1
‑
ca09
‑
or(含有赋予奥司他韦耐药性的na 274y(n2编号)突变的ca a/加利福利亚州/7/09(h1n1));h5
‑
vn04(ca a/越南/1203/04(h5n1));h5
‑
hk03(ca a/香港/213/
03(h5n1));h9
‑
hk97(ca a/鸡/香港/g9/97(h9n2);h2
‑
jp57(ca a/日本/57(h2n2));h2
‑
mo06(ca a/猪/密苏里州/06(h2n3));h6
‑
hk97(ca a/水鸭/香港/w312/97(h6n1));h6
‑
ab85(ca a/野鸭/艾伯塔省/89/85(h6n2));h3
‑
hk68(a/香港/8/68(h3n2));h3
‑
vic75(a/维多利亚/3/75(h3n2));h3
‑
la87(a/洛杉矶/7/09(h3n2));h3
‑
sd93(a/山东/9/93(h3n2));h3
‑
wh95(ca a/武汉/359/95(h3n2));h3
‑
syd97(ca a/悉尼/5/97(h3n2));h3
‑
wh95
‑
or(含有赋予奥司他韦耐药性的na 274y(n2编号)突变的ca a/武汉/359/95(h3n2));h3
‑
pa99(ca a/巴拿马/2007/99(h3n2));h3
‑
wy03(a/怀俄明州(wyoming)/03/03(h3n2));h3
‑
wi05(a/威斯康星州(wisconsin)/67/05(h3n2));h3
‑
珀斯09(ca a/珀斯/16/09(h3n2));h3
‑
vc11(a/维多利亚/361/11(h3n2));h7
‑
nld03(ca a/荷兰/219/03(h7n7));h7
‑
bc04(ca a/不列颠哥伦比亚/cn
‑
6/04(h7n3
‑
lp);h7
‑
anu13(ca a/安徽/1/13(h7n9)。
[0546]
表4
[0547][0548]
表4示出抗ha抗体中和所测试的所有1组甲型流感病毒。所有抗ha抗体(除了抗体8外)展示针对所测试的所有h3甲型流感病毒的中和活性,并且所有抗ha抗体(除了抗体6外)表现出针对h7
‑
nld03(ca a/荷兰/219/03(h7n7));h7
‑
bc04(ca a/不列颠哥伦比亚/cn
‑
6/04(h7n3
‑
lp)的中和活性。
[0549]
表5
[0550][0551][0552]
表5示出抗体变体(抗体11
‑
15)在中和所测试的所有1组和2组甲型流感病毒方面比亲本抗体3更有效,具有降低的ic
50
值。此外,抗体还中和3种病毒,这些病毒具有工程化至na蛋白中的赋予奥司他韦耐药性(or)的突变。
[0553]
实例5.抗ha igg针对猪源性h3n2病毒的中和活性。
[0554]
如实例4中描述在微量中和测定中测量抗ha抗体3和变体(抗体11
‑
15)针对新突发性猪源性h3n2病毒(a/明尼苏达州/11/2010和a/印第安纳州/10/2011)的中和活性。抗体fi6v4(在wo2013/011347a1中所描述)用作对照抗体。如在来自两个独立实验的表6中所示,抗体3和抗体变体(抗体11
‑
15)在中和猪源性a/印第安纳州/10/2011h3n2病毒方面比fi6v4更有效。抗体3和抗体变体有效力地中和猪源性a/明尼苏达州/11/2010h3n2病毒,而fi6v4在所测试的抗体的最高浓度(50ug/ml)下未能中和。
[0555]
表6
[0556][0557][0558]
实例6.抗ha中和抗体抑制流感融合和蛋白酶介导的ha0裂解。
[0559]
为了测试抗体介导的融合抑制,通过先前描述的修改的方案进行低ph诱导的红细胞融合测定(王t.t.(wang t.t.)等人,2010科学公共图书馆
·
病原学(plos pathog).6)。简言之,将a/波多黎各/8/34病毒(10
×
106tcid50)与人红细胞(2%最终红细胞浓度)在冰上孵育10分钟。将抗体3、抗体12和不相关的抗体mpe8v3与病毒在室温下孵育30分钟。然后在37℃下将红细胞添加至病毒
‑
抗体混合物持续30分钟且最终在37℃下添加乙酸钠缓冲液(0.5m ph 5.0)持续另外45分钟。将样品在400xg下离心6分钟并且在室温下孵育另外45分钟,且随后在400xg下再次离心6分钟以便沉淀红细胞。然后将上清液转移至elisa板以便通过测量在540nm下的吸光度来测定所释放的nadph的量(图1b)。结果显示抗体3和抗体12有效力地抑制病毒融合,而mpe8v3(针对副粘液病毒的融合蛋白的人单克隆抗体)(科尔蒂等人,2013自然501)不能抑制低ph诱导的融合。
[0560]
为了测试抗体介导的ha突变的阻断,将a/新喀里多尼亚/20/99(h1n1)的重组ha与抗体3、fi6v4、fe17.23或同种型对照抗体以15:1(mab:ha)的摩尔比孵育40分钟。然后将该抗体
‑
ha混合物暴露于2.5ug/ml的tpck处理的胰蛋白酶并且在37℃下孵育5、10和20分钟。将样品在聚丙烯酰胺凝胶上分离且然后转移至硝酸纤维素膜以用于使用识别甲型流感毒株的ha2和ha0的生物素化的人mab(fo32)(humab)进行蛋白质印迹分析(图1c)。结果显示抗体3在阻断蛋白酶介导的ha0裂解方面比fi6v4更有效。相比之下,fe17.23(识别ha球状头部的人单克隆抗体)和对照抗体不能抑制蛋白酶介导的ha0裂解。在一个单独实验中,使用以上所述的相同条件,相较于抗体3对抗体12和抗体14的蛋白酶裂解抑制进行了比较(图1d)。结果显示抗体12、抗体13具有与抗体3类似的阻断蛋白酶裂解的能力。
[0561]
实例7.抗ha抗体表现出fc效应子功能
[0562]
抗体具有通过fc效应子功能如抗体依赖性细胞毒性(adcc)、抗体依赖性细胞吞噬作用(adcp)和补体依赖性杀伤(cdc)清除病毒感染的细胞的潜力。为了证实抗ha抗体表现出adcc活性;对这些抗体在人自然杀伤(nk)细胞存在下杀死病毒感染的细胞的能力进行了测试。对用a/香港/8/68以20的moi感染的mdck细胞进行adcc测定。将感染的细胞与抗体的稀释系列一起孵育,且然后与从人pbmc(美天旎公司(miltenyi))阴性选择的纯化的nk细胞以6:1的效应子与靶比率进行孵育。将感染的细胞、抗体和nk细胞混合物孵育4小时,并且通过ldh释放(罗氏公司(roche))测量细胞杀死。图2示出所有四种抗ha柄抗体表现出感染的mdck细胞的剂量依赖性杀死。
[0563]
为了测量抗ha抗体介导吞噬作用的能力,使用了用源自a/香港/8/68的ha稳定转染的mdck细胞作为靶细胞。从pbmc分离人单核细胞,并且在m
‑
csf存在下培养7天以便分化成巨噬细胞。将人巨噬细胞和表达ha的靶细胞分别用紫色和绿色(celltrace紫或csfe,英杰公司)荧光标记。将标记的效应子和靶细胞在抗体的稀释系列存在下以6:1比率孵育2小时,且然后通过流式细胞术进行分析。将吞噬作用百分比测量为针对绿色靶细胞也为阳性的紫色染色的巨噬细胞(双阳性)的百分比。图3示出所有抗ha抗体显示类似水平的adcp,如所预期非特异性对照抗体不显示吞噬作用。
[0564]
为了测量抗ha抗体与补体合作来介导感染的细胞的杀死的能力,进行了cdc测定。在该测定中,将mdck细胞用a/波多黎各/8/34以2的moi感染,与抗体的稀释系列和源自兔的补体(cedarlane公司)以1:18的效应子与靶比率孵育。通过ldh释放(罗氏公司)测量细胞杀死。图4示出所有抗ha抗体显示在补体存在下介导细胞杀死的能力。
[0565]
实例8.抗ha抗体的预防和治疗作用
[0566]
在六至八周龄balb/c(哈兰实验室(harlan laboratories))小鼠模型中评价了人中和抗体(nab)针对流感病毒感染的保护功效。将小鼠在致死病毒激发之前抑或之后用不同剂量的nab进行处理。
[0567]
预防活性(图5和图6)将每组8只的小鼠作为单次腹膜内注射(ip)给予0.1、0.3、1、3和10mg/kg的抗体3或给予100μl体积中的10mg/kg的人同种型不相关的对照igg。在给药之后四小时,将小鼠用7倍50%小鼠致死剂量(7mld
50
)的a/加利福利亚州/7/09(h1n1)(h1
‑
ca09)或50μl体积中的7:1a/pr/8:a/hk/8/68ha(h3n1)(h3
‑
hk68)重排病毒鼻内接种。将小鼠在病毒激发当天或前一天称重并且每日监测体重损失和存活率持续14天(具有体重损失≥25%的小鼠被安乐死)。抗体3以剂量依赖性方式赋予保护。ip注射1mg/kg或更多的抗体3在用h1
‑
ca09(图5)和h3
‑
hk68(图6)激发的动物中提供完全保护。较低的抗体剂量(0.3mg/kg)也是高度保护性的,具有90%保护。如所预期,接受10mg/kg的同种型对照mab的小鼠没有一个在致死感染激发之后存活。
[0568]
治疗活性(图7和8)将小鼠用3mld
50
的h1
‑
ca09接种并且在感染后第24和48小时(h.p.i.)注射抗体3(图7)或在感染后第72、96和120小时注射5mld
50
的h3
‑
hk68(图8)。在感染后第24和48小时用30mg/kg的抗体3ip处理保护75%
‑
100%的用h1
‑
ca09激发的小鼠,并且在感染后第72和96小时用30mg/kg的抗体3ip处理保护87.5%
‑
100%的用h3
‑
hk68激发的小鼠。在h1和h3模型中在感染后第0或24小时用相同剂量的不相关的同种型对照抗体处理未能保护小鼠免于致死激发,其中存活率分别为0%或12.5%。
[0569]
抗体3变体的治疗活性(图9和10)将小鼠用3mld
50
的h1
‑
ca09接种并且在感染后第24小时注射抗体(图9)或用7mld
50 h3
‑
hk68接种且在感染后第48小时注射抗体(图10)。用2mg/kg的抗体3和变体mab(抗体11、抗体12和抗体14)ip处理保护87.5%
‑
100%的用h1
‑
ca09激发的小鼠,并且3mg/kg剂量的不同nab保护50%
‑
87.5%的用h3
‑
hk68激发的小鼠。如所预期,在h1和h3模型中在感染后第24或48小时用相同剂量的不相关的同种型对照抗体处理未能保护小鼠,其中存活率分别为0%或12.5%。
[0570]
实例9.抗ha抗体和小分子抑制剂奥司他韦的治疗作用
[0571]
为了直接比较抗ha nab与小分子神经氨酸酶(na)抑制剂奥司他韦的保护功效和组合疗法的作用,使用了在实例8中描述的流感鼠感染模型。
[0572]
抗ha nab与奥司他韦(图11和12)的治疗性比较将小鼠用3mld
50
的h1
‑
ca09接种且在感染之前4小时、感染后第1天抑或感染后2天用10mg/kg的抗体12处理或25mg/kg bid持续5天的奥司他韦引发(图11)。在感染之前和感染后1天用抗体12处理保护了100%的用h1
‑
ca09激发的小鼠,而所有用奥司他韦处理的动物死于感染。所有在感染之前4小时用相同剂量的不相关的同种型对照处理的动物以0%的存活率死亡。此外,将小鼠用7mld
50
的h3
‑
hk68接种,然后在感染后第1、2、3抑或4天用10mg/kg的抗体12处理或25mg/kg bid持续5天的奥司他韦引发(图12)。在感染后第1、2或3天用抗体12处理的动物显示100%的存活率,而在这些相同时间点用奥司他韦处理显示仅60%
‑
20%的存活率。如所预期,在感染后第1天用相同剂量的不相关的同种型对照抗体处理的小鼠以10%的存活率死于感染。
[0573]
抗ha nab与奥司他韦的治疗组合(图13)为了评定抗ha mab与奥司他韦的组合的相加作用,将小鼠用7mld
50
的h3
‑
hk68接种,并且在感染后第3天用次最优浓度的抗体12(2.5或0.3mg/kg)、25mg/kg bid持续5天的奥司他韦或抗体12(2.5或0.3mg/kg)与25mg/kg bid持续5天的奥司他韦的组合处理(图13)。用单独抗体12抑或奥司他韦处理保护仅10%
‑
20%的动物,而用2.5mg/kg的抗体12与奥司他韦的组合处理保护80%的动物,并且用0.3mg/kg的抗体12与奥司他韦的组合处理保护50%的动物。
[0574]
实例10.抗ha抗体和小分子抑制剂针对雪貂中的h5n1流感感染的治疗作用
[0575]
在五至六月龄流感血清阴性雪貂(triple f农场)中评价了抗ha nab和奥司他韦针对高致病性流感病毒感染的保护功效。将所有雪貂用1.0ml中的1ld
90
的a/vn/1203/04(h5n1)高致病性禽流感病毒鼻内激发(大约0.5ml/鼻孔),并且然后在感染后第1、2或3天用单一剂量的25mg/kg抗体12处理抑或25mg/kg bid持续5天的奥司他韦引发。计算每组的存活百分比(n=7)(图14)。在感染后第1、2或3天用抗体12处理的雪貂以及在感染后第1天用奥司他韦处理的那些雪貂受到保护,具有100%存活率。然而,当奥司他韦在感染后第2和3天引发时,雪貂仅分别具有71%存活(平均死亡天数12)和29%存活(平均死亡天数9)。如所预期,在感染后第1天用25mg/kg的不相关的同种型对照抗体处理的动物未能存活,具有0%存活率。
[0576]
实例11.通过选择单克隆抗体抗性突变体(marm)进行的表位鉴别。
[0577]
使用两种不同的方法从三种h3n2病毒分离抗体抗性突变体。将a/爱知/2/68(爱知/68)h3n2与高浓度的抗体12(125
×
ic
50
)孵育1小时,之后将病毒与抗体的混合物在10
×
96孔板中以30,000tcid50/孔吸附至mdck细胞并且在抗体12(10
×
ic
50
)存在下培养。分离在感染后表现出对感染的细胞的细胞病变作用(cpe)达3天的3种推定抗体12hk2/68marm。通过rt
‑
pcr扩增ha基因并且随后测序。序列分析揭示相较于亲本序列的2个非同义取代(表7)。这两个核苷酸改变分别编码在ha2的高度保守的柄区中的氨基酸位置18和19处的从异亮氨酸(i)至精氨酸(r)和从天冬氨酸(d)至酪氨酸(y)的单一氨基酸取代。可替代地,将流感h3n2病毒a/威斯康星州/67/2005(wi05)和ca a/巴拿马/2007/1999(pa99)的连续传代在增加浓度的抗体12(从2
‑5×
ic
50
至100
×
ic
50
)存在下繁殖。将潜在逃逸突变体通过有线稀释进行亚克隆柄区使其同源ha基因经受序列分析。鉴别了ha2中在位置19的从d至y的单一氨基酸改变和在位置42的从谷氨酰胺(q)至r的单一氨基酸改变。此外,在ha1与d19y的组合中在位置156的从组氨酸(h)至q的氨基酸取代;或在ha2中在位置19的从d至精氨酸(n)的氨基酸取代与残基45处的从i至n的氨基酸改变的组合;或在ha1与q42r的组合中在位置196的
从丙氨酸(a)至苏氨酸(t)的氨基酸取代的情况下观察到双突变(表7)。类似地,当使pa99在浓度达100
×
ic
50
的抗体12存在下连续传代时,在ha2残基42(q42r)和45(i45t)处选择单一氨基酸取代(表7)。表7中所示的代表性marm变体用于微量中和测定中以便进一步评价这些marm被抗体12中和的表型易感性。结果显示体外选择的含有突变d19y、h156q/d19y,、d19n/i45n、q42r或a196t/q42r的wi05 marm;含有q42r或i45t的pa99marm、以及具有突变d19y或i18r的爱知/68marm对抗体中和不太易感,其中当分别与其亲本野生型毒株相比较时,计算的ic
50
值的增加在对于pa99抗性克隆>8倍至对于wi05抗性变体>180倍的范围内(表8)。为了评定这些氨基酸取代对于对被抗体12中和的易感性的作用,产生了编码单独突变的重组a/香港/1
‑
5/68(rhk68)h3变体且使用微量中和测定进行了评价。如在表9中所示,h3 rhk68_i18r和rhk68_d19y变体在所测试的最高浓度(约200μg/ml)下表现出对抗体12的抗性并且相较于野生型rhk68病毒赋予对抗体12中和的易感性的>130倍降低。rhk68中的单一氨基酸改变q42r导致对被抗体12中和的易感性的适度约8倍降低。然而,在微量中和测定中在所选择的marm的ha蛋白中鉴别的氨基酸取代(k156q、a196t、i45n或i45t)不会改变编码这类取代的重组hk68病毒对抗体12的易感性。这些结果表明抗体12识别ha2的高度保守的柄区中的构象表位并且位置18、19、42或45的氨基酸是关键接触残基。
[0578]
表7.在抗体12抗性突变体的h3 ha中鉴别的氨基酸取代
[0579][0580]
表8.h3抗性变体对抗体12中和(neut)的易感性
[0581][0582]
表9.rhk68 h3变体对抗体12中和(neut)的易感性
[0583][0584][0585]
差异
[0586]
[0587][0588]
差异
[0589]
[0590][0591]
甲型流感参考文献:
[0592]
科尔蒂d.等人2010.异种亚型中和抗体由用季节性流感疫苗免疫的个体产生(heterosubtypic neutralizing antibodies are produced by individuals immunized with a seasonal influenza vaccine.)临床研究杂志120:1663
‑
1673。
[0593]
科尔蒂d.等人2011.一种选自浆细胞的结合1组和2组甲型流感血凝素的中和抗体
(a neutralizing antibody selected from plasma cells that binds to group 1and group 2influenza a hemagglutinins.)科学333:850
‑
856。
[0594]
科尔蒂d.等人2013.通过人单克隆抗体交叉中和四种副粘液病毒(cross
‑
neutralization of four paramyxoviruses by a human monoclonal antibody.)自然501(7467):439
‑
43。
[0595]
爱可尔特d.c.等人2009.高度保守的流感病毒表位的抗体识别(antibody recognition ofa highly conserved influenza virus epitope.)自然324:246
‑
251。
[0596]
爱可尔特d.c.等人2011.2组甲型流感病毒上的高度保守的中和表位(a highly conserved neutralizing epitope on group 2influenza a viruses.)自然333:843
‑
850。
[0597]
爱可尔特d.c.等人2012.由单一抗体环介导的甲型流感病毒的交叉中和(cross
‑
neutralization ofinfluenza a viruses mediated by a single antibody loop.)自然489:526
‑
532。
[0598]
克劳斯j.c.等人2011.识别流感h1n1病毒血凝素的球状头部上的保守的新颖表位的广泛中和人单克隆抗体(a broadly neutralizing human monoclonal antibody that recognizes a conserved,novel epitope on the globular head of the influenza h1n1 virus hemagglutinin.)病毒学杂志85:10905
‑
10908。
[0599]
李p.s.等人2012.通过亲合力增强的流感病毒血凝素受体结合位点的异种亚型抗体识别(heterosubtypic antibody recognition of the influenza virus hemagglutinin receptor binding site enhanced by avidity.)美国国家科学院院刊109:17040
‑
17045
[0600]
李g.m.等人2012.大流行性h1n1疫苗诱导人中的有利于广泛交叉反应性记忆b细胞的回忆应答(pandemic h1n1 influenza vaccine induces a recall response in humans that favors broadly cross
‑
reactive memory b cells.)美国国家科学院院刊109:9047
‑
9052。
[0601]
纳卡穆拉g.等人2013.体内人浆母细胞富集技术允许快速鉴别治疗性甲型流感抗体(an in vivo human
‑
plasmablast enrichment technique allows rapid identification oftherapeutic influenza a antibodies.)细胞宿主与微生物14:93
‑
103瑞j.等人2009.(禽类和人甲型流感病毒的广谱中和的结构和功能基础(structure and functional bases for broad
‑
spectrum neutralization of avian and human influenza a viruses.)自然结构与分子生物学16:265
‑
273。
[0602]
思罗斯比m.等人2008.从人igm+记忆b细胞回收的针对h5n1和h1n1交叉保护性的异种亚型中和单克隆抗体(heterosubtypic neutralizing monoclonal antibodies cross
‑
protective against h5n1 and h1n1 recovered from human igm+memory b cells.)公共科学图书馆
·
综合3:e3492
[0603]
王t.t.等人,2010.在用不同血凝素顺序免疫之后针对h3流感病毒的广泛保护性单克隆抗体(broadly protective monoclonal antibodies against h3 influenza viruses following sequential immunization with different hemagglutinins.)科学公共图书馆
·
病原学.6(2):e1000796。
[0604]
惠特尔j.r.r.等人2011.识别流感病毒血凝素的受体结合口袋的广泛中和人抗体(broadly neutralizing human antibody that recognizes the receptor
‑
binding pocket of influenza virus hemagglutinin.)美国国家科学院院刊108:14216
‑
14221。
[0605]
拉马特j.等人2011.广泛交叉反应性抗体主导针对2009大流行性h1n1流感病毒感染的人b细胞应答(broadly cross
‑
reactive antibodies dominate the human b cell response against 2009pandemic h1n1 influenza virus infection.)实验医学杂志208:181
‑
193。
[0606]
序列表信息
[0607]
抗体1(原始cdna)
[0608]
[0609]
抗体2(抗体1的表达形式)
[0610][0611]
抗体3(密码子优化的抗体2)
[0612][0613]
抗体4(原始cdna)hcdr3中的简并核苷酸,t或a
[0614][0615]
抗体5(抗体4hcdr3 v的表达形式)
[0616][0617]
抗体6(抗体4hcdr3 e的表达形式)
[0618][0619]
抗体7(原始cdna)
[0620][0621]
抗体8(抗体7的表达形式)
[0622][0623]
抗体9(原始cdna)
[0624][0625]
抗体10(抗体9的表达形式)
[0626][0627]
抗体11
[0628][0629]
抗体12
[0630][0631]
抗体13
[0632][0633]
抗体14
[0634][0635]
抗体15
[0636][0637]
抗体3
‑
gl
[0638]
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1