一种γ-生育酚甲基转移酶突变体及其在提高α-生育酚及α-生育三烯酚产量的应用的制作方法
一种
γ
‑
生育酚甲基转移酶突变体及其在提高
α
‑
生育酚及
α
‑
生育三烯酚产量的应用
【技术领域】
1.本发明涉及基因工程技术领域,具体涉及一种γ
‑
生育酚甲基转移酶突变体,还涉及含有所述突变体的工程菌株,及其在提高α
‑
生育酚及α
‑
生育三烯酚产量的应用。
背景技术:
2.维生素e是生育酚和生育三烯酚的总称,它是一种两亲性分子,由莽草酸途径来源的极性芳香族头部和亲脂性类异戊二烯侧链组成。根据侧链的饱和度和芳香环上甲基数目和位置的不同,分为α、β、γ、δ
‑
生育酚和α、β、γ、δ
‑
生育三烯酚这8种化合物,其中α
‑
生育酚更易被机体吸收,因此,α
‑
生育酚被认为是最具活性的物质。生育酚是重要的抗氧化剂,具有清除自由基,终止脂质链式氧化反应的作用,且对心血管疾病、癌症等有积极作用,还可以提高免疫力、抗衰老和促进生育功能。
3.γ
‑
生育酚甲基转移酶(γ
‑
tmt)是生育酚合成途径最后阶段的关键酶,催化γ
‑
和δ
‑
生育酚(生育三烯酚)转化为α
‑
和β
‑
生育酚(生育三烯酚)。可以说γ
‑
tmt基因就控制着着α
‑
生育酚含量的高低。研究显示γ
‑
生育酚甲基转移酶(γ
‑
tmt)在进化的过程中高度保守,在拟南芥和集胞藻中几乎具有相同的活性。拟南芥γ
‑
生育酚甲基转移酶(γ
‑
tmt)可以改变种子中维生素e的组分,过量表达使种子中的γ
‑
生育酚大量转化为α
‑
生育酚。但是γ
‑
tmt的活性有限,不足以催化γ
‑
生育酚大量转化为α
‑
生育酚。于是相当多的研究人员对γ
‑
生育酚甲基转移酶进行了过量表达。shintani和dellapenna在拟南芥中过量表达γ
‑
生育酚甲基转移酶(γ
‑
tmt)使α
‑
生育酚的含量提高了80倍(shintani et al.,1998)。在水稻,紫苏,大豆中分别过量表达γ
‑
生育酚甲基转移酶(γ
‑
tmt),α
‑
生育酚的含量都得到了显著提高(cho et al.,2005,tavva et al.,2007,yusuf et al.,2007)。
4.随着目前维生素e强大功能的普及和对维生素e的不断深入研究,市场需求也在不断增加,促使天然维生素e的生产方法不断改进。目前的化学合成方法主要存在收率低、纯度不高、浪费较大、易污染环境、设备昂贵、操作复杂及质量差等问题。而通过对关键酶基因的克隆和调控基因高效表达等生物工程技术手段仍然存在产量低的问题。通过酶催化反应合成生产生育酚具有促进其生产和工业应用的潜力。
技术实现要素:
5.在生育酚和生育三烯酚的生产中,γ
‑
生育酚甲基转移酶是γ
‑
生育酚和γ
‑
生育三烯酚向α
‑
生育酚和α
‑
生育三烯酚转化的关键基因,在目前的研究中,α类型活性是最高的,但是α类型所占比例不高,存在α型生育酚和生育三烯酚转化率低导致的α
‑
生育酚和生育三烯酚产量低的问题。
6.本发明的思路是通过基因序列定点突变,对来自拟南芥的γ
‑
生育酚甲基转移酶进行分子改造。
7.基于此,本发明提供一种γ
‑
生育酚甲基转移酶突变体d81m,所述突变体是对来源
于拟南芥的γ
‑
生育酚甲基转移酶氨基酸序列进行突变,将第81位天冬氨酸突变为蛋氨酸得到的。
8.在本发明中,所述突变体d81m的氨基酸序列如seq id no.4所示。
9.进一步地,本发明还提供编码上述γ
‑
生育酚甲基转移酶突变体的基因,以及携带上述基因的载体,和表达上述基因或上述载体的基因工程菌。
10.特别地,对于所述基因工程菌,其宿主菌为大肠杆菌,以pet
‑
28a为载体。
11.进一步地,本发明还提供上述的γ
‑
生育酚甲基转移酶突变体d81m在制备α
‑
生育酚或α
‑
生育三烯酚中的应用。
12.特别是以γ
‑
生育酚或γ
‑
生育三烯酚为底物,制备α
‑
生育酚或α
‑
生育三烯酚。
13.根据一种优选的实施方式,底物浓度为0.6mmol/l,反应体系里含有1.5mmol/l的sam(s
‑
腺苷甲硫氨酸),上述工程菌株在温度25℃下反应时间为60
‑
90min,制备得到α
‑
生育酚或α
‑
生育三烯酚。
14.本发明通过实验证实突变体d81m显著提高了γ型生育酚和生育三烯酚向α型转化的效率,提高了α型生育酚和生育三烯酚的含量。经反应条件优化,d81m生产α
‑
生育酚的产量达到193.2mg/l,转化率为72.6%,α
‑
生育三烯酚的产量为210.8mg/l,转化率为79.2%,在野生型的基础上将转化率提高了约2.8倍,大大提高了α
‑
生育酚和α
‑
生育三烯酚的含量。
【附图说明】
15.图1为各反应条件及对应结果。
【具体实施方式】
16.以下实施例用于非限制性地解释本发明的技术方案。
17.在本发明中,如无特殊说明,用于说明浓度的“%”均为质量百分比,“:”均为质量比,“份”均为重量份。
18.本发明涉及以下培养基或缓冲液:
19.lb液体培养基(1l):胰蛋白胨10g,酵母提取物5g,nacl 10g,121℃高压灭菌20min。
20.lb固体培养基(i l):胰蛋白胨10g,酵母提取物5g,nacl 10g,121℃高压灭菌20min。
21.50mmol/l tris,ph8.5:称量6.055g tris置于1l烧杯中,加入约800ml的去离子水,充分搅拌溶解,加入浓盐酸调节所需要的ph值到8.2,将溶液定容至1l,高温高压灭菌后,室温保存。
22.实施例1γ
‑
生育酚甲基转移酶的三位结构分析及突变体质粒的构建
23.1.突变点的确定
24.来源于拟南芥的γ
‑
生育酚甲基转移酶(γ
‑
tmt)在ncbi数据库的编号为np_176677.1,核苷酸序列如seq id no.1所示,氨基酸序列如seq id no.2所示。对γ
‑
tmt进行三维建模,结合底物口袋构象、计算模拟、底物与酶分子相互作用力,理性设计单点突变,选择81位的天冬氨酸,将其突变为蛋氨酸。
25.2.质粒的构建
26.(1)构建原始质粒pet
‑
28a
‑
γ
‑
tmt
27.从ncbi上下载γ
‑
生育酚甲基转移酶基因(γ
‑
tmt)序列(geneid:842805),送生工合成,连接在载体pet
‑
28a的多克隆位点,测序验证,得到原始质粒pet
‑
28a
‑
γ
‑
tmt。
28.(2)构建突变质粒pet
‑
28a
‑
γ
‑
tmtd81m
29.设计定点突变的突变引物(如表2),以携带γ
‑
tmt基因的原始质粒pet
‑
28a
‑
γ
‑
tmt为模板进行单点突变,将γ
‑
生育酚甲基转移酶(γ
‑
tmt)的第81位的天冬氨酸替换为蛋氨酸,构建得到突变质粒pet
‑
28a
‑
γ
‑
tmtd81m。突变经pcr及模板消化反应两个步骤,模板消化的产物测序验证。突变体酶的核苷酸序列如seq id no.3所示,氨基酸序列如seq id no.4所示。
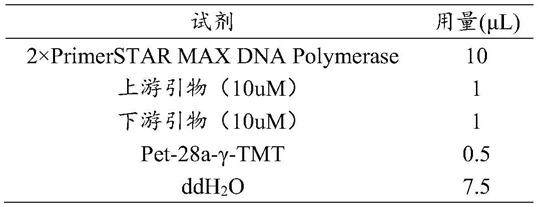
30.表1 pcr反应体系
[0031][0032]
pcr反应条件:95℃预变性3min,95℃变性30s,56℃退火30s,72℃延伸3min40s,32个循环,4℃保存。
[0033]
表2引物序列表
[0034][0035]
表3模板消化反应体系
[0036][0037]
反应条件:37℃反应90min。
[0038]
实施例2工程菌株的构建及突变体酶的表达纯化
[0039]
(1)工程菌株的构建
[0040]
将实施例1中得到的突变质粒pet
‑
28a
‑
γ
‑
tmtd81m和原始质粒pet
‑
28a
‑
γ
‑
tmt分别转化至大肠杆菌(e.coli)bl21(de3)感受态细胞中,涂布于含有100μg/ml卡那霉素的lb固体培养基中,于37℃培养12h后,分别得到长有重组工程菌e.coli bl21/pet
‑
28a
‑
γ
‑
tmtd81m和e.coli bl21/pet
‑
28a
‑
γ
‑
tmt的平板。
[0041]
(2)突变体酶的表达
[0042]
挑取步骤(1)中平板上的单菌落到4ml有50μg/ml卡那霉素的lb液体培养基,于37℃培养12h得到种子液,将种子液接入200ml含有50μg/ml卡那霉素的lb液体培养基,于37℃培养2~3h至od值为0.6~0.8,加终浓度为0.2mmol/l的iptg诱导蛋白的表达,于28℃培养6~8h,获得发酵液。将发酵液于4℃、4000rpm离心30min,收集菌体。
[0043]
(3)突变体酶的纯化。
[0044]
向步骤(2)中获得的菌体中加入用pbs缓冲液将菌体洗涤两次,并用pbs缓冲液将菌体重悬,移至金属容器,置于冰水浴中进行超声破碎。调节超声波探头于菌液液面以下,并不接触容器,设定超声破碎条件为:功率33%,工作3s,停歇3s,时间为15min。将全细胞破碎液转移至离心管,用提前预冷至4℃的离心机12000rpm,离心20min分离上清与沉淀。取离心后的上清用0.22um的滤膜进行过滤,用于ni
‑
nta纯化。
[0045]
使用镍离子亲和层析柱对粗酶液进行纯化:首先,使用平衡缓冲液(300mmol/lnacl,10mmol/l tris base,50mmol/l nah2po4,10mmol/l咪唑,ph7.0)平衡柱子;然后,将得到的粗酶液加入到柱子中;接着用含有低浓度咪唑的缓冲液(50mmol/l咪唑,10mmol/l tris base,300mmol/lnacl,50mmol/l nah2po4,ph7.0)冲洗杂蛋白;最后用含有高浓度咪唑的缓冲液(200mmol/l咪唑,10mmol/l tris base,300mmol/lnacl,50mmol/l nah2po4,ph7.0和300mmol/l咪唑,10mmol/l tris base,300mmol/lnacl,50mmol/l nah2po4,ph7.0)洗脱,得到纯化后的突变酶d81m和亲本酶γ
‑
tmt。
[0046]
(4)在最适温度和ph的条件下测定γ
‑
生育酚甲基转移酶的酶活
[0047]
在50mmol/l tris,ph8.5,25℃的反应条件下,以0.6mmol/l的γ
‑
生育酚为底物,添加1.5mmol/l sam(s
‑
腺苷甲硫氨酸)和60μl步骤(3)获得的突变体酶d81m和γ
‑
tmt,反应90min后将离心管于95℃水浴1min灭火酶终止反应。最终测得亲本酶活为30u/mg,突变体酶活为150u/mg,在亲本酶的基础上活性提高了5倍。
[0048]
实施例3:突变体酶d81m产α
‑
生育酚的条件优化
[0049]
产α
‑
生育酚条件优化:
[0050]
(1)优化加酶量
[0051]
以0.4mmol/l的γ
‑
生育酚为底物,向1.5ml的反应体系里(50mmol/l tris,ph8.5)分别加入5、10、20、30、40、60、80、100、120μl的突变体酶d81m和1.0mmol/l sam,于25℃反应120min。反应结束后将离心管于95℃水浴1min灭火酶终止反应。
[0052]
其中,α
‑
生育酚含量测定方法为:取5ml反应液用2ml丙酮分两次萃取,离心取上清,经0.22um滤膜滤后,进hplc分析。hplc检测条件为:流动相:0.01m kh2po4溶液(a)和甲醇(b);比例为90%a/10%b;流速:0.8ml/min;检测波长:290nm;色谱柱:ymc
‑
pack ods
‑
aq(4.6
×
250mm)。标准曲线制作:α
‑
生育酚标准曲线的制作:用万分之一天平称取10mgα
‑
生育酚,溶于10ml纯甲醇溶液中,浓度定为1g/l的母液,依次用纯甲醇溶液梯度稀释,稀释得到浓度分别为250mg/l,100mg/l,50mg/l,20mg/l,5mg/l的α
‑
生育酚溶液,并用相同的方法再稀释出两批同浓度的α
‑
生育酚溶液,对所有样品经0.22um滤膜过滤,再进行hplc分析,制作标准曲线。
[0053]
结果如图1a所示:当酶的添加量在60μl时,α
‑
生育酚的产量可以达到40mg/l以上。
[0054]
(2)优化底物浓度
[0055]
分别以0.1,0.2,0.3,0.4,0.5,0.6,0.7,0.8mmol/l的γ
‑
生育酚为底物,向1.5ml的反应体系里(50mmol/l tris,ph8.5)加入60μl的突变体酶d81m和1.0mmol/l sam,于25℃反应120min。反应结束后将离心管于95℃水浴1min灭火酶终止反应。
[0056]
结果如图1b所示:当γ
‑
生育酚的添加量在0.6mmol/l时α
‑
生育酚的产量可以达到90mg/l以上。
[0057]
(3)优化s
‑
腺苷甲硫氨酸sam浓度
[0058]
以0.6mmol/l的γ
‑
生育酚为底物,向1.5ml的反应体系里(50mmol/l tris,ph8.5)分别添加0.1,0.2,0.5,1.0,1.5,1.8,2.0mmol/l的sam,和60μl的突变体酶d81m,于25℃反应120min。反应结束后将离心管于95℃水浴1min灭火酶终止反应。
[0059]
结果如图1c所示,当sam的添加量在1.5mmol/l时α
‑
生育酚的产量可以达到160mg/l以上。
[0060]
(4)反应时间对α
‑
生育酚产量的影响
[0061]
以0.6mmol/l的γ
‑
生育酚为底物,向1.5ml的反应体系里(50mmol/l tris,ph8.5)加入1.5mmol/l sam和60μl的突变体酶d81m,于25℃反应10、20、40、60、90、120、150min。反应结束后将离心管于95℃水浴1min灭火酶终止反应。
[0062]
结果如图1d所示,当反应的时间在60
‑
90min时,α
‑
生育酚的产量可以达到180mg/l以上。
[0063]
可见,最优条件为50mmol/l tris、ph8.5,5mmol/l dtt,0.6mmol/l的底物,1.5mmol/l sam和60μl的酶,25℃反应90min。
[0064]
在最优反应条件下,以0.6mmol/lγ
‑
生育酚和0.6mmol/lγ
‑
生育三烯酚为底物,测定反应后突变体酶d81m的α
‑
生育酚和
‑
α生育三烯酚产量,以亲本酶γ
‑
tmt作为对照,结果如表4。
[0065]
其中,α
‑
生育三烯酚、γ
‑
生育三烯酚和γ
‑
生育酚含量的测定方法和hplc检测条件以及标准曲线的制作均同α
‑
生育酚。
[0066]
α
‑
生育酚转化率(%)=反应后α
‑
生育酚含量/反应前γ
‑
生育酚含量
×
100%
[0067]
α
‑
生育三烯酚转化率(%)=反应后α
‑
生育三烯酚含量/反应前γ
‑
生育三烯酚含量
×
100%
[0068]
表4反应前后底物与产物含量比较
[0069][0070]
由结果可知,在最优反应条件下,亲本酶γ
‑
tmt产生的α
‑
生育酚的最高产量为67.5mg/l,转化率为25.4%,α
‑
生育三烯酚的最高产量为75.3mg/l,转化率为28.3%;而突变体酶d81mα
‑
生育酚最高产量为193.2mg/l,转化率为72.6%,α
‑
生育三烯酚最高产量为210.8mg/l,转化率为79.2%,突变酶d81m的转化率比亲本酶提高约2.8倍。
[0071]
综上所述,突变体d81m显著提高了γ型生育酚和生育三烯酚向α型转化的效率,提高了α型生育酚和生育三烯酚的含量。本发明的d81m生产α
‑
生育酚的产量可达到193.2mg/l,转化率为72.6%,α
‑
生育三烯酚的产量为210.8mg/l,转化率为79.2%,显著提高了α
‑
生育酚和α
‑
生育三烯酚的含量。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1