糖基转移酶及利用该糖基转移酶的制备甜菊醇糖苷的方法与流程
本发明涉及一种糖基转移酶以及利用该糖基转移酶的用于生产甜菊醇糖苷的组合物以及制备方法,更具体地涉及一种向甜菊醇糖苷转移葡萄糖的莱鲍迪苷的糖基转移酶、表达所述酶的重组菌株以及利用它们的生产作为甜菊醇糖苷的莱鲍迪苷的方法。
背景技术:
1、已知甜味剂是食品、饮料或糖果行业中最常使用的成分。甜味剂可以在生产过程中加入到最终食品产物中,或者可以适当稀释单独使用,可以用作在餐桌甜味剂或烘焙中替代白糖的家庭用替代物。甜味剂包括例如蔗糖、高果糖玉米糖浆、糖蜜、枫糖浆和蜂蜜等天然甜味剂和例如阿斯巴甜、糖精和三氯蔗糖(sucralose)等人造甜味剂。
2、甜叶菊(stevia)提取物是一种可从多年生灌木甜叶菊(stevia rebaudiana)提取的天然甜味剂。纯化至各种水平的甜叶菊提取物在食品和混合物中用作高敏感性调味料,或作为餐桌甜味剂单独出售。
3、虽然甜叶菊植物的提取物含有莱鲍迪苷和其他有助于甜味的甜菊醇糖苷,但现有的市售产品主要是莱鲍迪苷a,还有少量的莱鲍迪苷c、莱鲍迪苷d和莱鲍迪苷f等其它糖苷。从植物中提取的甜叶菊提取物可能含有导致异味(off-flavors)的化合物等污染物。根据所选的食品系统或用途,这些异味通常会成为问题。
4、此外,甜叶菊提取物的组合物可能因植物生长的土壤和气候而有很大差异。根据原料植物、气候条件和提取工艺,在商业制备过程中,据报道,莱鲍迪苷a的量占总甜菊醇糖苷含量的20-97%。其他甜菊醇糖苷以不同的量存在于甜叶菊提取物中。
5、从甜叶菊植物生产的甜菊提取物含有多种甜菊醇糖苷和导致异味的化合物,并且由于回收和纯化劳动密集且效率低下,仍然需要可以以高收率积累诸如reb d和reb m的甜菊醇糖苷的重组生产系统。为此,仍然需要开发一种高效酶。此外,仍然需要改善reb d的生产,以便在用于商业用途的重组宿主中生产甜菊醇糖苷reb m。
技术实现思路
1、要解决的技术问题
2、本发明的一个实例涉及一种将葡萄糖转移至甜菊醇糖苷的糖基转移酶、编码所述酶蛋白的核酸分子,包含所述核酸分子的重组载体和转化微生物。
3、本发明的一个实例涉及一种用于生产甜菊醇糖苷的组合物,所述组合物包含将葡萄糖转移至所述甜菊醇糖苷的糖基转移酶蛋白、表达所述酶蛋白的重组微生物、所述微生物的细胞、所述微生物的细胞裂解物、所述微生物的培养物和它们的提取物中的一种以上。
4、本发明的一个实例涉及一种甜菊醇糖苷的生产方法,其包括使选自将葡萄糖转移至所述甜菊醇糖苷的糖基转移酶蛋白、表达所述酶蛋白的重组微生物、所述微生物的细胞、所述微生物的细胞裂解物、所述微生物的培养物和它们的提取物中的一种以上与所述酶的底物进行反应的步骤。
5、技术方案
6、以下,对本发明进行更具体的说明。
7、本发明的一个实例涉及一种源自微生物且将葡萄糖转移至甜菊醇糖苷的糖基转移酶,更具体地涉及一种udp-葡糖基转移酶(即尿苷二磷酸葡糖基转移酶,简称为ugt)。
8、所述udp-葡糖基转移酶涉及一种包含与seq id no:1的氨基酸序列具有92%以上、95%以上、97%以上、99%以上、99.3%以上或99.5%以上的序列同一性的氨基酸序列的酶蛋白。例如,只要是具有这种氨基酸序列的同一性且显示出与由所述seq id no:1的氨基酸序列组成的蛋白对应的功效的氨基酸序列,则即使其是包括部分序列被删除、修饰、取代或添加的氨基酸序列,也显而易见地包含在本发明的范围内。所述酶蛋白可以被seq idno:2的核苷酸序列或与seq id no:2的核苷酸序列的序列同一性为40%以上、50%以上、60%以上、70%以上、80%以上、90%以上、95%以上、97%以上或99%以上的核苷酸序列编码。
9、在上文中,术语“同源性”或“同一性”是指与给定氨基酸序列或碱基序列的一致程度,并且可以表示为百分比。在本说明书中,与给定氨基酸序列或碱基序列具有相同或相似活性的同源序列用“%同源性”或“%同一性”表示。
10、根据本发明的另一个实例,包含与seq id no:1的氨基酸序列的序列同一性为92%以上、95%以上、97%以上或99%以上的氨基酸序列的酶蛋白包括具有与seq id no:1相同的氨基酸序列的野生型酶,和部分序列被删除、修饰、取代或添加的变异型酶蛋白,只要它们表现出与由所述seq id no:1的氨基酸序列组成的蛋白相对应的功效即可。
11、根据本发明的udp-葡糖基转移酶具有在葡萄糖供体的存在下将选自甜菊苷和莱鲍迪苷a(reb a)中的一种以上的底物转化为甜菊醇糖苷的活性,例如具有转化为选自莱鲍迪苷d(reb d)和莱鲍迪苷e(reb e)中的一种以上的活性,更具体地可以将reb a转化为rebd,并且可以将甜菊苷转化为reb e(参考以下反应式1),并且对于reb a和甜菊苷的单独底物或混合底物均具有转化活性。
12、reb a向reb d的转化是葡萄糖(glucose)通过甜菊醇环(steviol ring)上的β1-2糖苷键(beta 1-2glycosidic bond)的结合而发生的转移,主要发生在甜菊醇环的19位。甜菊醇糖苷可以包括1-3个通过β键结合的葡萄糖分子。
13、[反应式1]
14、
15、根据本发明udp-葡糖基转移酶具有在2-24小时,更具体为3–18小时的酶促反应中将reb a的40重量%以上转化为reb d的活性,具体地,具有将reb a的40重量%以上、50重量%以上、60重量%以上、65重量%以上或70重量%以上转化为reb d的活性。此外,根据本发明的酶蛋白在对reb a单独底物的2-24小时酶促反应,更具体为3-18小时的酶促反应中,以100%的eugt11的reb d转化率为基准,可以具有85%以上,90%以上或93%以上,例如85-110%、90-110%、93-110%、85-100%、90-100%、93-100%、85-99%、90-99%或93-99%的reb d转化率的活性。所述酶的转化活性可以通过reb a单独底物在30℃、ph7.2、150rpm条件下进行的酶反应来测量,对于通过hplc法分析的峰面积(peak area)值,利用以下数学式1计算转化率(%)。
16、[数学式1]
17、reb d转化率(%)=reb d的hplc峰面积/总hplc峰面积
18、此外,udp-葡糖基转移酶具有在包含甜菊苷和reb a的混合底物中在2-24小时,更具体为5-20小时的酶促反应中,将包含reb a和甜菊苷的混合底物的75重量%以上、80重量%、85重量%、90重量%或95重量%以上转化为reb d和reb e的活性。所述酶的转化活性可以通过在30℃、ph7.2和150rpm下对含有reb a和甜菊苷的混合底物进行的酶反应来测量,对于通过hplc法分析的峰面积值,利用以下数学式2计算转化率(%)。
19、[数学式2]
20、reb d/e转化率(%)=reb d/e的hplc峰面积/总hplc峰面积
21、此外,根据本发明的酶蛋白在包含reb a和甜菊苷的混合底物的2-24小时,更具体为5-20小时的酶促反应中,以100%的eugt11的reb d/e总转化率为基准,可以具有105%以上、110%以上、120%以上、130%以上、140%以上或150%以上,例如105-200%、110-200%、120-200%、130-200%、140-200%、150-200%、105-190%、110-190%、120-190%、130-190%、140-190%或150-190%的reb d/e转化率的活性。
22、根据本发明的酶蛋白在hplc分析图中不产生具有对应于14.5-15.5分钟的洗脱时间的峰的化合物,或者产生非常少量的副产物。因此,根据本发明的酶蛋白在包含reb a和甜菊苷的混合底物中具有比eugt11更高的reb d/e转化率,而且具有随酶反应时间的推移而增加的特性。
23、根据本发明的udp-葡糖基转移酶涉及一种通过寻找源自植物的酶而源自小麦(triticum aestivum)的酶。通过基因检索发现的taugt是作为现有的推定蛋白(putativeprotein)存在的,是功能尚未明确的蛋白。确认了该蛋白的功能,确认了ugt酶的活性,确认了将甜菊苷和reb a转化为reb e和reb d的酶的活性。
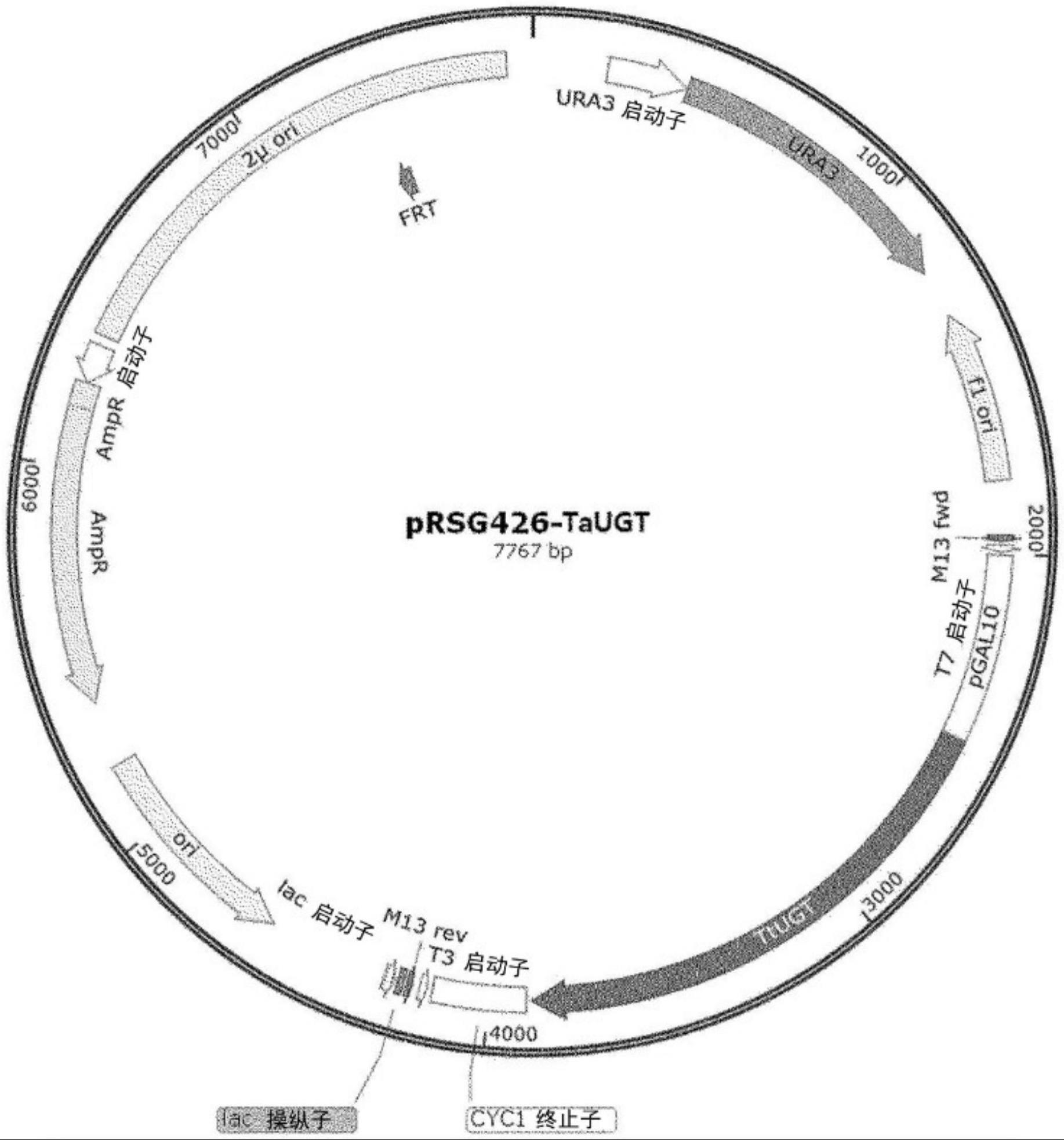
24、甜度高的reb d是由甜叶菊植物中少量存在的糖reb a产生的,参与该过程的酶是ugt91d2。具有与其相似的活性的酶是稻米中存在的eugt11酶。在甜叶菊的ugt91d2的情况下,具有将甜菊醇糖苷reb a转化为reb d的活性,但由于转化活性低,因此难以生产高含量的reb m。
25、此外,以往已知的ugt酶有源自甜叶菊(steviareb audiana)的ugt、源自水稻(oryza sativa)的ugt(以下称为eugt11)、源自大麦(hordeum vulgare)的ugt(以下称为hvugt)。与以往已知的eugt11相比,根据本发明的udp-葡糖基转移酶具有不产生反应副产物的优点。
26、与hvugt和eugt11的氨基酸序列同一性分析结果是,根据本发明的酶与hvugt的氨基酸序列同一性为91%,与eugt11的氨基酸序列同一性为66%,多核苷酸序列同一性分析的结果是,与hvugt的多核苷酸序列同一性为36%,与eugt11的多核苷酸序列同一性为37%。
27、根据本发明udp-葡糖基转移酶可以在反应温度为10-50℃且ph为5.0至9.0的含水体系中进行。优选地,反应可以在温度为25℃至40℃且ph为6.0至8.0的含水体系(aqueoussystem)中进行,更优选在温度为30℃且ph为7.2的含水体系中进行。根据优选的实施方案,反应可以在ph为7.2的磷酸缓冲液中进行。
28、本发明的一个实例涉及一种编码将葡萄糖转移至甜菊醇糖苷的糖基转移酶蛋白的核酸分子、包含所述核酸分子的重组载体和转化微生物。
29、根据本发明,重组微生物或细胞可以为微生物细胞,优选为大肠杆菌(e.coli)、酵母属(saccharomyces)菌株,例如包括酿酒酵母(saccharomyces cerevisiae)或毕赤酵母(pichia)等,但不限于此。
30、本说明书中的术语“转化”是指将含有编码靶蛋白的核酸的载体引入宿主细胞中,使得由所述核酸编码的蛋白可以在宿主细胞中表达。只要转化的核酸能够在宿主细胞中表达,则可以插入宿主细胞的染色体中或位于染色体外,或者可以无关地全部包括这些情况。此外,所述核酸包括编码靶蛋白的dna和rna。所述核酸只要可以引入宿主细胞并表达,则可以以任何形式引入。例如,可以将核酸以表达盒(expression cassette)的形式引入宿主细胞,所述表达盒是包含自我表达所需的所有要求的遗传构建体。所述表达盒可以包括可操作地连接至所述核酸的启动子(promoter)、转录终止信号、核糖体结合位点和翻译终止信号。所述表达盒可以是可自我复制的表达载体的形式。此外,所述核酸可以以其自身的形式引入宿主细胞并可操作地连接至在宿主细胞中表达所必需的序列,但不限于此。
31、此外,在上文中,术语“可操作地连接”是指基因序列功能性连接至启动子序列,所述启动子序列启动和介导编码本发明的靶蛋白的核酸的转录。
32、根据本发明的一个实例的包含编码将葡萄糖转移至甜菊醇糖苷的糖基转移酶蛋白的核酸分子的重组载体的一个实例示于图1的切割图中。例如,它可以包括编码根据本发明的酶的核酸分子和可操作地连接于所述核酸分子的作为转录控制序列的gal10启动子和cyc1终止子。转录启动子可以包括gal10、gal1、gal2、tef1、gpd1和tdh3启动子等,转录终止子可以包括cyc1、tef1和pgk1等。
33、转化本发明的载体的方法包括将核酸引入细胞的任何方法,根据宿主细胞,可以选择本领域中公知的适合的标准技术并进行。例如,有电穿孔(electroporation)、磷酸钙(capo4)沉淀、氯化钙(cacl2)沉淀、显微注射法(microinjection)、聚乙二醇(peg)法、deae-葡聚糖法、阳离子脂质体法和醋酸锂-dmso法等,但不限于此。
34、本发明的一个实例涉及一种用于生产甜菊醇糖苷的组合物,其包含选自将葡萄糖转移至所述甜菊醇糖苷的糖基转移酶蛋白、表达所述酶蛋白的重组微生物、所述微生物的细胞、所述微生物的细胞裂解物、所述微生物的培养物和它们的提取物中的一种以上。
35、所述用于生产甜菊醇糖苷的组合物可以进一步包含含有选自甜菊苷和reb a中的至少一种的底物,例如可以是甜菊苷或reb a单独底物或它们的混合底物。所述混合底物可以是甜菊苷或reb a的混合物或者包含甜菊苷或reb a的甜叶菊提取物。
36、所述组合物可以进一步包含将reb d或reb e转化为reb m的酶,转化为所述reb m用的酶是第二udp-糖基转移酶,例如包含源自甜叶菊的ugt76g1等。
37、所述组合物可以进一步包含葡萄糖供体,例如包含udp-葡萄糖或可以生成udp-葡萄糖的糖类。根据本发明的将葡萄糖转移至甜菊醇糖苷的糖基转移酶蛋白将葡萄糖从葡萄糖供体转移至甜菊醇糖苷。
38、所述培养物包含由生产udp-葡糖基转移酶的微生物生产的酶,可以包含菌株或是不包含菌株的无细胞(cell-free)形式。所述裂解物是指将产生udp-葡糖基转移酶的微生物的细胞破碎得到的裂解物或将所述裂解物离心分离获得的上清液,并且包括由生产所述udp-葡糖基转移酶的微生物生产的酶。
39、所述菌株的培养物包括由生产所述udp-葡糖基转移酶的微生物生产的酶,并且可以包含微生物细胞,或是不包含所述微生物细胞的无细胞形式。在本说明书中,除非另有说明,使用的生产udp-葡糖基转移酶的微生物是指包含选自所述菌株的细胞、所述菌株的培养物、所述细胞的裂解物、所述裂解物的上清液和它们的提取物中的一种以上。
40、当利用所述甜菊醇糖苷生产用组合物生产甜菊醇糖苷时,利用udp-葡糖基转移酶或生产酶的微生物的反应温度和反应ph条件如所述酶的反应温度和反应ph条件中所述。
41、本发明的一个实例涉及一种甜菊醇糖苷的生产方法,其包括使选自将葡萄糖转移至所述甜菊醇糖苷的糖基转移酶蛋白、表达所述酶蛋白的重组微生物、所述微生物的细胞、所述微生物的细胞裂解物、所述微生物的培养物和它们的提取物中的一种以上与所述酶的底物反应的步骤。
42、本发明的一个实例涉及一种制备选自reb d和reb e中的至少一种甜菊醇糖苷的方法,其包括在葡萄糖供体的存在下使选自根据本发明的udp-葡糖基转移酶蛋白、表达所述酶蛋白的重组微生物、所述微生物的细胞、所述微生物的细胞裂解物、所述微生物的培养物和它们的提取物中的一种以上与选自甜菊苷和reb a中的至少一种的底物发生反应的步骤。
43、本发明的附加的一个实例涉及一种制备reb m的方法,其包括以下步骤:在葡萄糖供体的存在下,使选自根据本发明的udp-葡糖基转移酶蛋白、表达所述酶蛋白的重组微生物、所述微生物的细胞、所述微生物的细胞裂解物、所述微生物的培养物和它们的提取物中的一种以上与选自甜菊苷和reb a中的至少一种的底物反应,从而制备选自reb d和reb e中的至少一种的甜菊醇糖苷;以及在葡萄糖供体的存在下,使所述制备的甜菊醇糖苷与选自第二udp-葡糖基转移酶蛋白、表达所述酶蛋白的重组微生物、所述微生物的细胞、所述微生物的细胞裂解物、所述微生物的培养物和它们的提取物中的至少一种发生反应。
44、所述reb m的制备步骤中使用的第二udp-葡糖基转移酶蛋白可以是ugt76g1等。
45、本发明方法可以显著缩短生产周期,并提高生产能力,成本低,并且可以提供纯度更高的产品,因此可以经济地用于食品、饮料产业。
46、根据本发明的udp-葡糖基转移酶(glucosyltransferase)蛋白的底物是来自中化(sinochem)(中国)公司的甜叶菊提取物,在该提取物中包含50%的reb a和90%的甜菊醇糖苷,从而用于进行酶促转化。
47、利用根据本发明的udp-葡糖基转移酶蛋白和表达所述酶蛋白的重组微生物制备的甜菊醇糖苷是高甜度甜味剂,添加于各种食品和饮料等中并使用或者单独作为餐桌甜味剂销售。
48、有益效果
49、与现有的已知的ugt相比,根据本发明的udp-葡糖基转移酶蛋白具有作用于甜菊醇糖苷的各种底物的底物特异性,并且具有不产生反应副产物的优点,并且可以在葡萄糖供体的存在下,通过使选自所述酶蛋白、表达所述酶蛋白的重组微生物、所述微生物的细胞、所述微生物的细胞裂解物、所述微生物的培养物和它们的提取物中的一种以上与选自甜菊苷和reb a中的至少一种的底物发生反应来制备甜菊醇糖苷,并且甜菊醇糖苷可以作为高甜度甜味剂添加到各种食品、饮料等中来使用。
- 还没有人留言评论。精彩留言会获得点赞!