一种控制稻米品质的热激蛋白家族基因HSP110-3及其应用
一种控制稻米品质的热激蛋白家族基因hsp110-3及其应用
技术领域
1.本发明属于基因工程技术领域,具体涉及一种控制稻米品质的热激蛋白家族基因hsp110-3及其应用。
背景技术:
2.水稻是世界上最重要的粮食作物之一,全世界种植水稻的国家和地区有 100多个。随着人们生活水平的不断改善,对稻米品质的要求也越来越高,不断改善稻米的品质也一直是水稻育种家们追求的育种目标。稻米品质主要包含研磨品质、外观品质、营养品质和蒸煮食味品质等四个主要方面,稻米垩白是评价稻米外观品质的重要指标,影响稻米外观品质的因素有很多,而垩白的形成主要是由遗传基因决定,且在品种间差异比较大。因此,对控制稻米垩白的基因进行发掘,并解析其影响稻米垩白的形成机理,并应用于水稻等禾谷类作物优质品种的培育,具有重要的理论意义和生产应用潜力。
3.在过去20年,随着dna标记和基因组测序技术的快速发展,影响稻米品质的qtl逐渐被发现并被克隆研究。而影响垩白形成的主要原因有遗传因素和环境因素。遗传因素主要有淀粉合成相关基因如:wx、agpase、ssi、 ssii、siii等,这些基因突变会直接导致稻米淀粉合成受阻,进而直接导致籽粒垩白形成;蛋白合成及转运相关基因如:osaap6、osglua2、osrab5a、 osvpei等,这些基因的突变会导致蛋白合成以及运输过程受阻,间接影响蛋白体的形成以及导致垩白的产生;脂肪酸合成及转运基因如:osltpl36、 lox3、osfad2-1等基因突变会导致脂类物质合成及转运受阻导致垩白形成。此外,受高温环境影响因子基因如osbzip58、flo2、osmsd7、cphsp70-2 会直接或间接影响一些营养物质合成相关基因的表达进而导致籽粒垩白形成。仅利用某一个控制垩白性状的qtl或基因较大幅度降低水稻的垩白性状几乎是不可能的,而多个有利基因的聚合才是降低垩白的有效途径,利用性状关联开展水稻的品质改良也是降低稻米垩白简洁有效的方法。目前被发现的控制垩白性状的基因还不足够,需要克隆更多的控制垩白性状的基因或发掘更多的低垩白等位基因,并开发出实用的与垩白性状紧密连锁的分子标记或基因内的功能标记,以用于垩白性状改良的分子育种。因此研究参与并调控垩白形成的基因,对于提高稻米品质具有重要的研究意义。
技术实现要素:
4.针对现有技术中的上述不足,本发明提供一种控制稻米品质的热激蛋白家族基因hsp110-3及其应用,该基因参与并调控垩白形成,有利于提高稻米品质。
5.为实现上述目的,本发明解决其技术问题所采用的技术方案是:
6.一种控制稻米品质的热激蛋白家族基因hsp110-3,该基因的核苷酸序列如seq id no.1所示。
7.一种热激蛋白家族基因hsp110-3编码的蛋白质,该蛋白质的氨基酸序列如seq id no.2所示。
8.进一步地,热激蛋白家族基因hsp110-3在调控稻米品质的应用。
9.进一步地,热激蛋白家族基因hsp110-3在水稻等禾谷类作物品种培育中的应用。
10.一种包含热激蛋白家族基因hsp110-3的表达载体。
11.一种包含热激蛋白家族基因hsp110-3的工程菌。
12.一种调控稻米品质的生物制剂,包括热激蛋白家族基因hsp110-3。
13.本发明的有益效果为:
14.本发明克隆了一种控制稻米品质的热激蛋白基因hsp110-3,通过 crispr/cas9基因编辑技术对野生型稻米中的基因hsp110-3进行敲除后,转基因株系籽粒表现出垩白粒率显著升高,表明该基因参与并调控垩白的形成,具有调控稻米品质的功能,能够用于水稻等禾谷类作物的优质品种培育,也为其他物种中同源基因的克隆和研究提供了参考。
附图说明
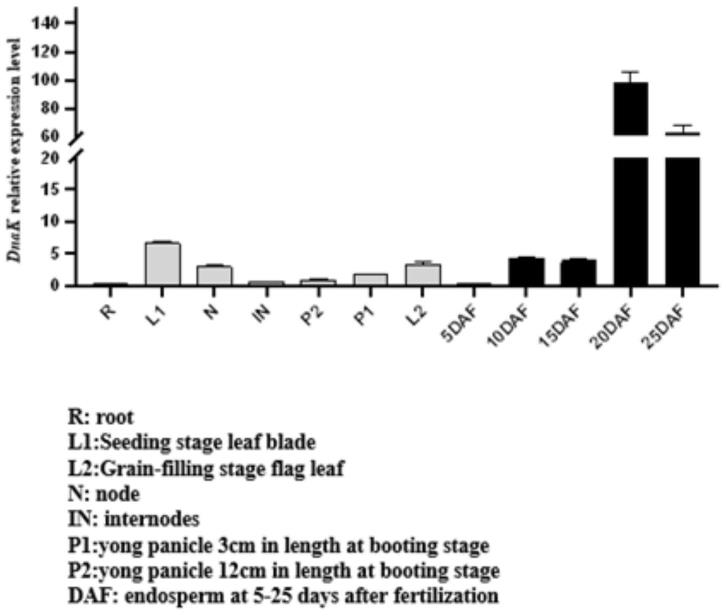
15.图1为hsp110-3基因在水稻全生育期时空表达谱。
16.图2为crispr/cas9系统载体图谱。
17.图3为hsp110-3基因crispr/cas9靶位点信息。
18.图4为报告基因gus载体图谱。
19.图5为hsp110-3的gus转基因植株不同组织部位染色结果。
20.图6为hsp110-3转基因株系表型及考种数据;其中,a为转基因株系与野生型株系的植株株叶形态比较图;b为转基因株系与野生型株系穗部比较图;c为转基因株系与野生型株系的籽粒粒长和粒宽比较图;d为转基因株系与野生型株系的糙米垩白面积比较图;e为转基因株系与野生型株系的精米外观比较图;f为转基因株系与野生型株系的穗长比较;g为转基因株系与野生型株系的分蘖数比较;h为基因株系与野生型株系植株株高比较;i 为转基因株系与野生型株系粒长比较;j为转基因株系与野生型株系粒宽比较;k为转基因株系与野生型株系垩白粒率比较;l为转基因株系与野生型株系籽粒精米垩白面积比较;m为转基因株系与野生型株系精米垩白度比较; n为转基因株系与野生型株系精米白度比较;o为转基因株系与野生型株系精米透明度比较;“**”表示在0.01水平下差异显著。
具体实施方式
21.下面对本发明的具体实施方式进行描述,以便于本技术领域的技术人员理解本发明,但应该清楚,本发明不限于具体实施方式的范围,对本技术领域的普通技术人员来讲,只要各种变化在所附的权利要求限定和确定的本发明的精神和范围内,这些变化是显而易见的,一切利用本发明构思的发明创造均在保护之列。
22.实施例
23.1、qpcr引物设计
24.hsp110-3蛋白由578个氨基酸组成,对应基因包含了1737个核苷酸;使用软件primer5分别在hsp110-3基因的exon区设计了一对定量引物, lhy53f(5
’‑
gctggcaatgtcctcagggtat-3’), lhy53r(5
’‑
caggtcaagcgtactgccataca-3’)。
25.2、hsp110-3基因在水稻全生育期时空表达谱分析
26.提取水稻根、苗期叶片、灌浆期旗叶、茎、节间、幼穗3cm处、幼穗 12cm处和开花后
5d、10d、15d、20d和25d的颖果中rna,反转录后利用 qpcr进行表达量分析,如图1所示。
27.结果表明:hsp110-3基因主要在颖果表达,并在颖果发育中后期高表达。
28.3、hsp110-3基因的crispr/cas9敲除载体构建及遗传转化
29.(1)向导rna靶位点序列的设计与选择
30.根据hsp100-3的基因组序列,设计选择并合成1个hsp110-3的向导 rna靶序列。
31.(2)靶位点crispr/cas9-grna载体构建
32.将步骤(1)中合成的向导rna靶序列的寡核苷酸链95℃变性,然后通过与pyl-u6a-grna中间载体混合,加入bsa1内切酶和t4连接酶及 buffer,pcr仪上37℃5min,20℃5min,5个循环进行酶切连接,之后再经过两轮pcr获得2个分别含有这个靶位点的grna表达盒,然后通过golden gate cloning的方法将grna表达盒依次装载到crispr/cas9载体上,得到单个靶位点crispr/cas9-grna载体,crispr/cas9系统载体图谱,如图2所示;hsp110-3基因crispr/cas9靶位点信息,如图3所示。
33.(3)遗传转化
34.将中花11号种子经次氯酸钠溶液灭菌后置于诱导培养基上,约2周后将诱导出的愈伤挑下进行继代培养,1周后挑取生长旺盛的愈伤用于农杆菌浸染。
35.将带有敲除靶位点crispr/cas9-grna载体转入农杆菌菌株eh105中,挑单克隆摇菌,浸染愈伤,在黑暗条件下25℃共培养3天,然后转移到含有 g418抗生素的筛选培养基上筛选10天左右。将筛选出的愈伤转移至分化培养基上进行培养,待长成正常幼苗后移出试管进行移栽。最后取植株叶片提 dna,通过pcr进行阳性植株鉴定。
36.4、阳性植株检测
37.取步骤(3)所得植株叶片提取dna,首先用潮霉素检测引物筛选获得阳性植株,接着分别用跨靶位点的扩增引物,ak773 (5
’‑
agcattcctcaccggatgtt-3’)和ak774 (5
’‑
agtcgcagatgtcgtttcct-3’)进行扩增,并将阳性单株中序列扩增后送擎科测序公司测序。
38.阳性植株测序获得序列经过比对分析显示hsp110-3基因共获得2种蛋白翻译提前终止和1种氨基酸缺失的转基因植株。
39.5、gus载体构建及转化
40.以野生型zh11的dna为模板,通过pcr扩增atg起始密码子上游的 2kb(hsp110-3)基因组片段,并将pcr产物胶回收用单片段重组酶连接到以线性化的dx2181载体上。并送百格基因转粳稻zh11材料。gus载体图谱,如图4所示。
41.6、gus材料染色
42.取阳性gus转基因植株的各个组织器官进行gus染色分析。
43.(1)按1:50的比例将x-gluc solution(50x)和gus buffer充分混匀,配成x-gluc染色液。
44.(2)将各个组织放入5ml离心管中,加入适量染色液,使组织完全浸没于染液中。
45.(3)37℃避光孵育1-24h,直至组织上显现蓝色。
46.(4)用70%乙醇反复脱去组织中的叶绿素,直至叶绿素被彻底清除。
47.(5)样品保存于70%乙醇中,并用体式显微镜进行观察和拍照。
48.如图5所示。
49.7、表型分析
50.在t0和t1代转基因植株种植中,对不同株系农艺性状观察发现,与野生型zh11相比,转基因植株除垩白外,其它农艺性状无显著差异。获得稳定遗传的t2代转基因植株后,田间种植三次重复,每个重复种植2行,首先测序检测确认靶位点突变。随后考察农艺性状时,每个重复去掉边行和杂株,选择5-10株稳定株系,考察单株籽粒垩白粒率,并进行统计分析,如图6所示。
51.结果显示,与野生型zh11相比,转基因植株籽粒垩白粒率均显著增加。由此说明,hsp110-3基因是水稻籽粒垩白调控基因,hsp110-3基因可以调控稻米品质。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1