靶向EpCAM的激动性CD28抗原结合分子的制作方法
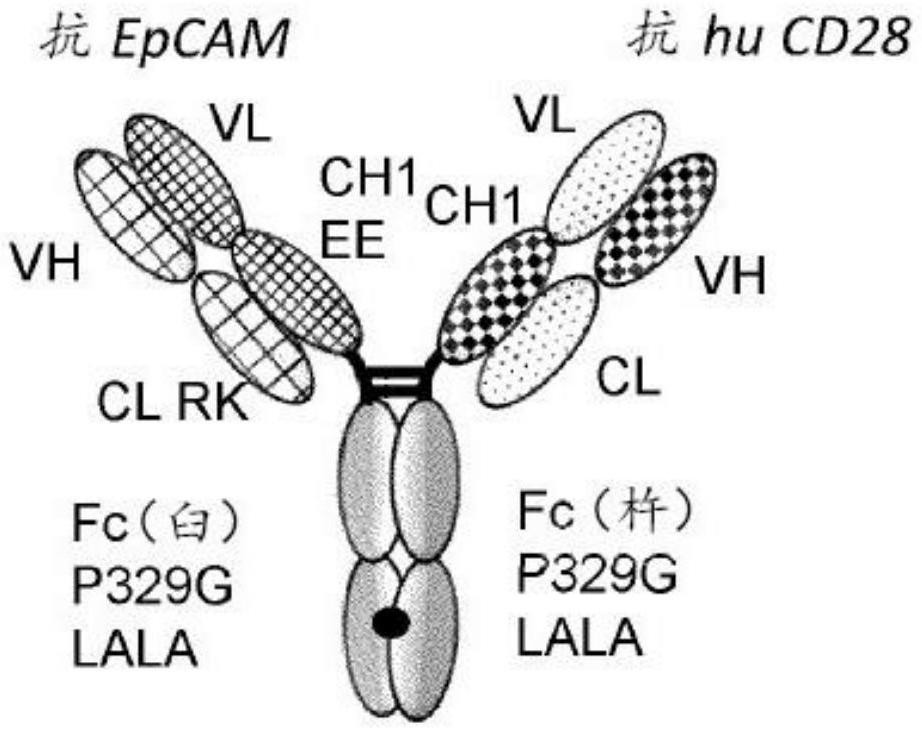
本发明涉及特征在于与cd28单价结合的包含新的人源化epcam抗体的双特异性激动性cd28抗原结合分子、其生产方法、含有这些分子的药物组合物,以及它们在疾病、特别是癌症的治疗中作为免疫调节剂和/或共刺激物的用途。
背景技术:
1、癌症免疫疗法正在成为越来越有效的疗法选择,其可以在黑色素瘤、非小细胞肺癌和肾细胞癌等癌症类型中产生显著和持久的反应。这主要是由几种免疫检查点阻断剂的成功推动的,包括抗pd-1(例如keytruda、merck;opdivo、bms)、抗ctla-4(例如yervoy、bms)和抗pd-l1(例如tecentriq、罗氏)。这些药剂可能作为许多癌症类型的标准治疗,或作为组合疗法的支柱,然而,只有一小部分患者(<25%)从这些疗法中受益。此外,各种癌症(前列腺癌、结直肠癌、胰腺癌、肉瘤、非三阴性乳腺癌等)对这些免疫调节剂存在原发耐药性。许多报告表明,缺乏预先存在的抗肿瘤t细胞会导致一些患者缺乏反应或反应不佳。总之,尽管现有免疫疗法具有令人印象深刻的抗癌作用,但对于解决较大癌症患者群体和开发旨在诱导和增强新型肿瘤特异性t细胞反应的疗法来说,显然存在医学需求。
2、cd28是共刺激分子亚家族的创始成员,其特征在于成对的v-set免疫球蛋白超家族(igsf)结构域连接到单个跨膜结构域和包含关键信号传导基序的细胞质结构域(carreno和collins,2002)。该亚家族的其他成员包括icos、ctla-4、pd1、pd1h、tigit和btla(chen和flies,2013)。cd28表达仅限于t细胞,并且普遍存在于所有初始型亚群和大多数经历抗原刺激的亚群中,包括那些表达pd-1或ctla-4的亚群。cd28和ctla-4高度同源并竞争结合相同的b7分子cd80和cd86,这两种分子在树突状细胞、b细胞、巨噬细胞和肿瘤细胞上表达(linsley等人,1990)。ctla-4对b7配体家族的更高亲和力使ctla-4在配体结合方面胜过cd28并抑制效应t细胞应答(engelhardt等人,2006)。相比之下,pd-1被证明通过使cd28的细胞质结构域部分去磷酸化来抑制cd28信号转导(hui等人,2017)。通过专职性抗原呈递细胞表面上的cd80或cd86连接cd28对于初始t细胞的功能性从头启动、随后的克隆扩增、细胞因子产生、靶细胞裂解和长期记忆的形成是严格必需的。cd28配体的结合还促进诱导型共刺激受体如ox-40、icos和4-1bb的表达(综述于acuto和michel,2003)。在连接cd28(二硫键连接的同型二聚体)后,膜近端ymnm基序和远端pyap基序已被证明与几种激酶和衔接蛋白复合(boomer和green,2010)。这些基序对于诱导il2转录来说很重要,该转录由nfat、ap-1和nfκb家族转录因子的cd28依赖性活化介导(fraser等人,1991)(june等人,1987)(thompson等人,1989)。然而,在cd28的细胞质结构域内发现了另外的表征不佳的磷酸化和泛素化位点。正如(esensten等人,2016)所综述,该cd28启动的通路在促进常规t细胞的增殖和效应子功能方面具有关键作用。cd28连接还促进调节性t细胞的抗炎功能。cd28通过部分增强来自t细胞受体的信号来共刺激t细胞,但也被证明介导独特的信号传导事件(acuto和michel,2003;boomer和green,2010;june等人,1987)。由cd28特异性触发的信号控制t细胞功能的许多重要方面,包括下游蛋白的磷酸化和其他翻译后修饰(例如,pi3k介导的磷酸化)、转录变化(例如bcl-xl表达)、表观遗传变化(例如il-2启动子)、细胞骨架重塑(例如微管组织中心的取向)和糖酵解速率的变化(例如糖酵解通量)。cd28缺陷小鼠对传染性病原体、同种异体移植物抗原、移植物抗宿主病、接触性过敏反应和哮喘的反应降低(acuto和michel,2003)。缺乏cd28介导的共刺激导致t细胞体外和体内增殖减少,严重抑制生发中心形成和免疫球蛋白同种型转换,减少辅助性t(th)细胞分化和th2-型细胞因子表达。cd4依赖性细胞毒性cd8+t细胞应答也受到影响。重要的是,cd28缺陷型初始t细胞的增殖反应降低,尤其是在较低抗原浓度下。越来越多的文献主体支持使cd28结合在t细胞上具有抗肿瘤潜力的观点。最近的证据表明,pd-l1/pd-1和ctla-4检查点抑制剂的抗癌作用取决于cd28(kamphorst等人,2017;tai等人,2007)。调查ctla-4和pd-1阻断治疗效果的临床研究在晚期黑色素瘤和其他癌症患者中显示出非常有希望的结果。此外,输注表达人工嵌合t细胞受体的基因工程t细胞已显示出在b细胞癌症和其它癌症中的高应答率和耐久性,该人工嵌合t细胞受体包含与细胞内tcr信号结构域(cd3z)和细胞内共刺激结构域(cd28和/或4-1bb结构域)融合的细胞外抗原识别结构域。
3、cd28激动性抗体可分为两类:(i)cd28超激动抗体和(ii)cd28常规激动性抗体。通常,为了活化初始t细胞,需要t细胞抗原受体(tcr,信号1)的参与和cd28的共刺激信号(信号2)。cd28超级激动剂(cd28sa)是cd28特异性单克隆抗体,能够在没有明显t细胞受体参与的情况下自主活化t细胞(hünig,2012)。在啮齿动物中,cd28sa活化常规和调节性t细胞。cd28sa抗体在多种自身免疫、炎症和移植模型中具有治疗效果。然而,人类cd28sa抗体tgn1412的i期研究在2006年引发了一场危及生命的细胞因子风暴。后续研究表明,该毒性是由于人类t细胞和临床前动物模型的t细胞的cd28反应性差异导致的剂量错误引起的。目前正在针对ra患者和转移性或不可切除的晚期实体恶性肿瘤患者的开放标签、多中心剂量递增研究中对tgn1412进行重新评估。cd28常规激动性抗体,例如克隆9.3,模拟cd28天然配体,并且仅在存在t细胞受体信号(信号1)的情况下能够增强t细胞活化。已发表的见解表明,抗体的结合表位对激动性抗体是超级激动剂还是常规激动剂具有重大影响(beyersdorf等人,2005)。超激动tgn1412与cd28的横向基序结合,而常规激动性分子9.3与配体结合表位紧密结合。由于不同的结合表位,超激动抗体和常规激动性抗体在t细胞表面形成cd28分子线性复合物的能力不同。准确地说,tgn1412能够有效地形成cd28的线性阵列,这可能导致聚合的信号成分足以超过t细胞活化的阈值。另一方面,常规激动剂9.3会导致复合物的结构不是线性的。先前已经发表了基于9.3克隆的常规激动结合剂的转化尝试(otz等人,2009),该尝试使用针对黑素瘤相关蛋白聚糖和cd28的重组双特异性单链抗体。尽管基于双特异性单链抗体形成多聚体构建体的内在趋势,使用了常规cd28激动结合剂9.3,但据报道,所报告的双特异性单链抗体仍发挥“超激动”活性。
4、上皮细胞粘附分子(epcam)–也称为肿瘤相关钙信号转导子1(tacstd1)、17-1a和cd326–是一种约40kda的i型跨膜糖蛋白,在上皮癌中高表达,并且在正常单上皮中表达水平较低。例如,在schnell等人,biochimica et biophysica acta–biomembranes(2013),1828(8):1989-2001;trzpis等人,am j pathol.(2007)171(2):386–395以及baeuerle和gires,br.j.cancer,(2007)96:417–423中回顾了epcam的结构和功能。
5、epcam在基底侧膜处表达,并且在不依赖于钙的嗜同性细胞粘附中发挥作用。成熟的epcam分子(在尽显处理以去除23个氨基酸信号肽之后)包含n末端、242氨基酸细胞外结构域(该氨基酸细胞外结构域包含表皮生长因子样重复区、人甲状腺球蛋白(ty)重复区和半胱氨酸缺乏区)、单程23个氨基酸跨膜结构域和c末端、26氨基酸胞质结构域(该氨基酸细胞质结构域包含用于α-辅肌动蛋白和npxy内化基序的两个结合位点)。epcam经常在上皮来源的癌症中过度表达并且是由癌症干细胞表达,并且因此是一种对治疗和诊断具有重要意义的分子。由于其在癌症及其转移瘤中频繁和高表达,epcam用作预后标志物、治疗靶标以及循环和扩散性肿瘤细胞(ctc/dtc)的锚定分子,这些细胞被认为是转移性癌细胞的主要来源。细胞外结构域epcam可以被裂解以产生可溶性细胞外结构域分子epex和细胞内分子epicd。epicd已经被证明与其他蛋白质缔合形成核复合物,该核复合物上调促进细胞增生的基因的表达。epcam还可能参与上皮细胞向间质细胞的转变(emt),并且可能促成大转移瘤的形成。
6、已经进行了临床试验,以用于抗epcam抗体治疗各种癌症的用途。epcam特异性抗体(依决洛单抗(edrecolomab);17-1a)于1995年首次在德国获得市场批准用于治疗结直肠癌,但从未获得fda批准。此外,epcam用于富集、识别和表征从晚期癌症患者的原发性肿瘤扩散到血液和骨髓中的转移细胞。尽管存在挑战,epcam仍然是临床上用于分离具有预后价值和转移潜力的循环肿瘤细胞(ctc)的首选表面抗原。
7、已经发现,当有限量的抗cd3双特异性抗体,即t细胞双特异性抗体(tcb),诸如cea-tcb,与激动性抗cd28分子组合时,可以实现更好的t细胞活化。鉴于cd28在各种肿瘤适应症中t细胞上以基线表达(lavin等人,2017;tirosh等人,2016;zheng等人,2017)并且cd28信号传导的活化增强了t细胞受体信号,tcb分子与靶向epcam的cd28分子的组合预期协同作用以诱导强而持久的抗肿瘤应答。wo 2020/127618 a1描述了靶向肿瘤的激动性cd28抗原结合分子。其中描述了各种肿瘤靶点。
8、然而,已经发现分子的活性严格取决于肿瘤靶向抗体的特性。因此,我们在本文中描述了新颖靶向epcam的激动性cd28分子,该分子显示出与tcb的强大协同作用,并且在存在tcb信号的情况下需要cd28结合单价以实现严格的肿瘤靶标依赖性。
技术实现思路
1、本发明描述了新的靶向epcam的双特异性激动性cd28抗原结合分子,其实现肿瘤依赖性t细胞活化和肿瘤细胞杀伤,而无需形成多聚体。本发明的双特异性cd28抗原结合分子的特征在于与cd28单价结合,并且在于它们包含如本文所定义的能够与上皮细胞粘附分子(epcam)特异性结合的特异性抗原结合结构域。此外,它们具有由能够稳定缔合的第一亚基和第二亚基组成的fc结构域,该第一亚基和第二亚基包含一个或多个氨基酸取代,该一个或多个氨基酸取代降低该抗原结合分子与fc受体的结合亲和力和/或效应子功能。从而消除了fc受体介导的交联,并且通过结合能够与epcam特异性结合的第二抗原结合结构域的交联来实现肿瘤特异性活化。
2、因此,本发明提供了双特异性激动性cd28抗原结合分子,其特征在于与cd28单价结合,该双特异性激动性cd28抗原结合分子包含
3、(a)能够与cd28特异性结合的第一抗原结合结构域,
4、(b)能够与如下抗原结合结构域特异性结合的第二抗原结合结构域:该抗原结合结构域能够与上皮细胞粘附分子(epcam)特异性结合,以及
5、(c)由能够稳定缔合的第一亚基和第二亚基构成的fc结构域,其包含一个或多个氨基酸取代,该一个或多个氨基酸取代降低抗原结合分子与fc受体的结合亲和力和/或效应子功能,
6、其中能够与epcam特异性结合的第二抗原结合结构域包含
7、(i)重链可变区(vhepcam)和轻链可变区(vlepcam),该重链可变区包含以下重链互补决定区:seq id no:309的cdr-h1、seq id no:310的cdr-h2和seq id no:311的cdr-h3,该轻链可变区包含以下轻链互补决定区:seq id no:312或seq id no:313的cdr-l1、seqid no:314的cdr-l2和seq id no:315的cdr-l3;或者
8、(ii)重链可变区(vhepcam)和轻链可变区(vlepcam),该重链可变区包含以下重链互补决定区:seq id no:2的cdr-h1、seq id no:3的cdr-h2和seq id no:4的cdr-h3,该轻链可变区包含以下轻链互补决定区:seq id no:5的cdr-l1、seq id no:6的cdr-l2和seqid no:7的cdr-l3;或者
9、(iii)重链可变区(vhepcam)和轻链可变区(vlepcam),该重链可变区包含以下重链互补决定区:seq id no:10的cdr-h1、seq id no:11的cdr-h2和seq id no:12的cdr-h3,该轻链可变区包含以下轻链互补决定区:seq id no:13的cdr-l1、seq id no:14的cdr-l2和seq id no:15的cdr-l3。
10、在一个方面,提供了如下定义的双特异性激动性cd28抗原结合分子,其中该fc结构域是igg,特别是igg1 fc结构域或igg4 fc结构域。在一个特定方面,由能够稳定缔合的第一亚基和第二亚基构成的fc结构域是igg1 fc结构域。在一个方面,fc结构域包含氨基酸取代l234a和l235a(根据kabat eu索引编号)。在一个方面,fc结构域属于人igg1亚类,并且包含氨基酸突变l234a、l235a和p329g(根据kabat eu索引编号)。
11、在一个方面,提供了如上文所定义的双特异性激动性cd28抗原结合分子,其中能够与cd28特异性结合的第一抗原结合结构域包含
12、(i)重链可变区(vhcd28)和轻链可变区(vlcd28),该重链可变区包含以下重链互补决定区:seq id no:26的cdr-h1、seq id no:27的cdr-h2和seq id no:28的cdr-h3,该轻链可变区包含以下轻链互补决定区:seq id no:29的cdr-l1、seq id no:30的cdr-l2和seqid no:31的cdr-l3;或者
13、(ii)重链可变区(vhcd28)和轻链可变区(vlcd28),该重链可变区包含seq id no:18的cdr-h1、seq id no:19的cdr-h2和seq id no:20的cdr-h3,该轻链可变区包含seq idno:21的cdr-l1、seq id no:22的cdr-l2和seq id no:23的cdr-l3。
14、在一个方面,双特异性激动性cd28抗原结合分子的能够与cd28特异性结合的抗原结合结构域包含重链可变区(vhcd28)和轻链可变区(vlcd28),该重链可变区包含seq idno:26的cdr-h1、seq id no:27的cdr-h2和seq id no:28的cdr-h3,该轻链可变区包含seqid no:29的cdr-l1、seq id no:30的cdr-l2和seq id no:31的cdr-l3。
15、在另一个方面,双特异性激动性cd28抗原结合分子的能够与cd28特异性结合的抗原结合结构域包含重链可变区(vhcd28)和轻链可变区(vlcd28),该重链可变区包含seq idno:18的cdr-h1、seq id no:19的cdr-h2和seq id no:20的cdr-h3,该轻链可变区包含seqid no:21的cdr-l1、seq id no:22的cdr-l2和seq id no:23的cdr-l3。
16、而且,提供了如上文所定义的双特异性激动性cd28抗原结合分子,其中能够与cd28特异性结合的第一抗原结合结构域包含重链可变区(vhcd28)和轻链可变区(vlcd28),该重链可变区包含与seq id no:24的氨基酸序列至少约95%、96%、97%、98%、99%或100%相同的氨基酸序列,该轻链可变区包含与seq id no:25的氨基酸序列至少约95%、96%、97%、98%、99%或100%相同的氨基酸序列。
17、在进一步方面,提供了双特异性激动性cd28抗原结合分子,其中能够与cd28特异性结合的第一抗原结合结构域包含重链可变区(vhcd28)和轻链可变区(vlcd28),该重链可变区包含选自由seq id no:32、seq id no:33、seq id no:34、seq id no:35、seq id no:36、seq id no:37、seq id no:38、seq id no:39、seq id no:40和seq id no:41组成的组的氨基酸序列,该轻链可变区包含选自由seq id no:25、seq id no:42、seq id no:43、seqid no:44、seq id no:45、seq id no:46、seq id no:47、seq id no:48、seq id no:49、seqid no:50和seq id no:51组成的组的氨基酸序列。
18、在另一个方面,提供了双特异性激动性cd28抗原结合分子,其中能够与cd28特异性结合的第一抗原结合结构域包含
19、(a)重链可变区(vhcd28)和轻链可变区(vlcd28),该重链可变区包含seq id no:37的氨基酸序列,该轻链可变区包含seq id no:44的氨基酸序列,或者
20、(b)重链可变区(vhcd28)和轻链可变区(vlcd28),该重链可变区包含seq id no:37的氨基酸序列,该轻链可变区包含seq id no:25的氨基酸序列,或者
21、(c)重链可变区(vhcd28)和轻链可变区(vlcd28),该重链可变区包含seq id no:41的氨基酸序列,该轻链可变区包含seq id no:51的氨基酸序列,或者
22、(d)重链可变区(vhcd28)和轻链可变区(vlcd28),该重链可变区包含seq id no:36的氨基酸序列,该轻链可变区包含seq id no:43的氨基酸序列,或者
23、(e)重链可变区(vhcd28)和轻链可变区(vlcd28),该重链可变区包含seq id no:36的氨基酸序列,该轻链可变区包含seq id no:44的氨基酸序列,或者
24、(f)重链可变区(vhcd28)和轻链可变区(vlcd28),该重链可变区包含seq id no:36的氨基酸序列,该轻链可变区包含seq id no:49的氨基酸序列,或者
25、(g)重链可变区(vhcd28)和轻链可变区(vlcd28),该重链可变区包含seq id no:36的氨基酸序列,该轻链可变区包含seq id no:25的氨基酸序列,或者
26、(h)重链可变区(vhcd28)和轻链可变区(vlcd28),该重链可变区包含seq id no:33的氨基酸序列,该轻链可变区包含seq id no:25的氨基酸序列,或者
27、(i)重链可变区(vhcd28)和轻链可变区(vlcd28),该重链可变区包含seq id no:32的氨基酸序列,该轻链可变区包含seq id no:43的氨基酸序列,或者
28、(j)重链可变区(vhcd28)和轻链可变区(vlcd28),该重链可变区包含seq id no:32的氨基酸序列,该轻链可变区包含seq id no:49的氨基酸序列,或者
29、(k)重链可变区(vhcd28)和轻链可变区(vlcd28),该重链可变区包含seq id no:32的氨基酸序列,该轻链可变区包含seq id no:25的氨基酸序列。
30、在一个特定的方面,提供了双特异性激动性cd28抗原结合分子,其中能够与cd28特异性结合的第一抗原结合结构域包含重链可变区(vhcd28)和轻链可变区(vlcd28),该重链可变区包含以下重链互补决定区:seq id no:52的cdr-h1、seq id no:53的cdr-h2和seqid no:54的cdr-h3,该轻链可变区包含以下轻链互补决定区:seq id no:55的cdr-l1、seqid no:56的cdr-l2和seq id no:57的cdr-l3。在一个方面,能够与cd28特异性结合的第一抗原结合结构域包含重链可变区(vhcd28)的cdr和轻链可变区(vlcd28)的cdr,该重链可变区包含seq id no:37的氨基酸序列,该轻链可变区包含seq id no:44的氨基酸序列。在一个特定方面,能够与cd28特异性结合的第一抗原结合结构域包括重链可变区(vhcd28)和轻链可变区(vlcd28),该重链可变区包含seq id no:37的氨基酸序列,该轻链可变区包含seqid no:44的氨基酸序列。
31、在另一个特定方面,提供了双特异性激动性cd28抗原结合分子,其中能够与cd28特异性结合的第一抗原结合结构域包含重链可变区(vhcd28)和轻链可变区(vlcd28),该重链可变区包含以下重链互补决定区:seq id no:58的cdr-h1、seq id no:59的cdr-h2和seqid no:60的cdr-h3,该轻链可变区包含以下轻链互补决定区:seq id no:61的cdr-l1、seqid no:62的cdr-l2和seq id no:63的cdr-l3。在一个方面,能够与cd28特异性结合的第一抗原结合结构域包含重链可变区(vhcd28)的cdr和轻链可变区(vlcd28)的cdr,该重链可变区包含seq id no:36的氨基酸序列,该轻链可变区包含seq id no:43的氨基酸序列。在一个方面,能够与cd28特异性结合的第一抗原结合结构域包含重链可变区(vhcd28)和轻链可变区(vlcd28),该重链可变区包含seq id no:36的氨基酸序列,该轻链可变区包含seq idno:43的氨基酸序列。在进一步的特定方面,提供了双特异性激动性cd28抗原结合分子,其中能够与cd28特异性结合的第一抗原结合结构域包含重链可变区(vhcd28)和轻链可变区(vlcd28),该重链可变区包含以下重链互补决定区:seq id no:64的cdr-h1、seq id no:65的cdr-h2和seq id no:66的cdr-h3,该轻链可变区包含以下轻链互补决定区:seq id no:67的cdr-l1、seq id no:68的cdr-l2和seq id no:69的cdr-l3。在一个方面,能够与cd28特异性结合的第一抗原结合结构域包含重链可变区(vhcd28)的cdr和轻链可变区(vlcd28)的cdr,该重链可变区包含seq id no:32的氨基酸序列,该轻链可变区包含seq id no:25的氨基酸序列。在一个方面,能够与cd28特异性结合的第一抗原结合结构域包含重链可变区(vhcd28)和轻链可变区(vlcd28),该重链可变区包含seq id no:32的氨基酸序列,该轻链可变区包含seq id no:25的氨基酸序列。
32、在一个方面,提供了双特异性激动性cd28抗原结合分子,其中能够与epcam特异性结合的第二抗原结合结构域包含重链可变区(vhepcam)和轻链可变区(vlepcam),该重链可变区包含以下重链互补决定区:seq id no:309的cdr-h1、seq id no:310的cdr-h2和seqid no:311的cdr-h3,该轻链可变区包含以下轻链互补决定区:seq id no:312或seq idno:313的cdr-l1、seq id no:314的cdr-l2和seq id no:315的cdr-l3。在一个方面,能够与epcam特异性结合的第二抗原结合结构域包含
33、(i)含有seq id no:258的氨基酸序列的该重链可变区(vhepcam)的cdr和含有seqid no:264的氨基酸序列的该轻链可变区(vlepcam)的cdr,或者
34、(ii)含有seq id no:258的氨基酸序列的该重链可变区(vhepcam)的cdr和含有seq id no:266的氨基酸序列的该轻链可变区(vlepcam)的cdr,或者
35、(iii)含有seq id no:258的氨基酸序列的该重链可变区(vhepcam)的cdr和含有seq id no:267的氨基酸序列的该轻链可变区(vlepcam)的cdr,或者
36、(iv)含有seq id no:258的氨基酸序列的该重链可变区(vhepcam)的cdr和含有seq id no:269的氨基酸序列的该轻链可变区(vlepcam)的cdr,或者
37、(v)含有seq id no:259的氨基酸序列的该重链可变区(vhepcam)的cdr和含有seqid no:266的氨基酸序列的该轻链可变区(vlepcam)的cdr。
38、在一个方面,能够与epcam特异性结合的第二抗原结合结构域包含
39、(i)重链可变区(vhepcam)和轻链可变区(vlepcam),该重链可变区包含seq id no:258的氨基酸序列,该轻链可变区包含seq id no:264的氨基酸序列,或者
40、(ii)重链可变区(vhepcam)和轻链可变区(vlepcam),该重链可变区包含seq idno:258的氨基酸序列,该轻链可变区包含seq id no:266的氨基酸序列,或者
41、(iii)重链可变区(vhepcam)和轻链可变区(vlepcam),该重链可变区包含seq idno:258的氨基酸序列,该轻链可变区包含seq id no:267的氨基酸序列,或者
42、(iv)重链可变区(vhepcam)和轻链可变区(vlepcam),该重链可变区包含seq idno:258的氨基酸序列,该轻链可变区包含seq id no:269的氨基酸序列,或者
43、(v)重链可变区(vhepcam)和轻链可变区(vlepcam),该重链可变区包含seq id no:259的氨基酸序列,该轻链可变区包含seq id no:266的氨基酸序列。
44、在一个方面,提供了双特异性激动性cd28抗原结合分子,其中能够与epcam特异性结合的第二抗原结合结构域包含重链可变区(vhepcam)和轻链可变区(vlepcam),该重链可变区包含以下重链互补决定区:seq id no:2的cdr-h1、seq id no:3的cdr-h2和seq idno:4的cdr-h3,该轻链可变区包含以下轻链互补决定区:seq id no:5的cdr-l1、seq idno:6的cdr-l2和seq id no:7的cdr-l3。在一个方面,能够与epcam特异性结合的抗原结合结构域包括包含seq id no:8的氨基酸序列的重链可变区(vhepcam)和包含seq id no:9的氨基酸序列的轻链可变区(vlepcam)。在一个特定方面,能够与epcam特异性结合的第二抗原结合结构域包括包含seq id no:8的氨基酸序列的重链可变区(vhepcam)和包含seq idno:9的氨基酸序列的轻链可变区(vlepcam)。
45、在另一个方面,提供了如本文该的双特异性激动性cd28抗原结合分子,其中能够与epcam特异性结合的抗原结合结构域包含重链可变区(vhepcam)和轻链可变区(vlepcam),该重链可变区包含以下重链互补决定区:seq id no:10的cdr-h1、seq id no:11的cdr-h2和seq id no:12的cdr-h3,该轻链可变区包含以下轻链互补决定区:seq id no:13的cdr-l1、seq id no:14的cdr-l2和seq id no:15的cdr-l3。在一个方面,能够与epcam特异性结合的抗原结合结构域包括包含seq id no:16的氨基酸序列的重链可变区(vhepcam)和包含seq id no:17的氨基酸序列的轻链可变区(vlepcam)。在一个特定方面,能够与epcam特异性结合的抗原结合结构域包括包含seq id no:16的氨基酸序列的重链可变区(vhepcam)和包含seq id no:17的氨基酸序列的轻链可变区(vlepcam)。
46、在进一步的方面,提供了如上文定义的双特异性激动性cd28抗原结合分子,其中能够与cd28特异性结合的第一抗原结合结构域和/或能够与epcam特异性结合的第二抗原结合结构域是fab片段或crossfab片段。在一个方面,提供了如本文所述的双特异性激动性cd28抗原结合分子,其包含(a)能够与cd28特异性结合的fab片段,(b)能够与epcam特异性结合的crossfab片段和(c)由能够稳定缔合的第一亚基和第二亚基构成的fc结构域,其包含一个或多个氨基酸取代,该取代降低抗原结合分子对fc受体的结合亲和力和/或效应子功能。在另一个方面,提供了如本文所述的双特异性激动性cd28抗原结合分子,其包含(a)能够与cd28特异性结合的crossfab片段,(b)能够与epcam特异性结合的fab片段和(c)由能够稳定缔合的第一亚基和第二亚基构成的fc结构域,其包含一个或多个氨基酸取代,该一个或多个氨基酸取代降低抗原结合分子对fc受体的结合亲和力和/或效应子功能。
47、在一个方面,能够与cd28特异性结合的第一抗原结合结构域为fab片段,其中fab轻链和fab重链的可变结构域vl和vh彼此替换或恒定结构域cl和ch1彼此替换,特别是可变结构域vl和vh彼此替换。在一个方面,能够与epcam特异性结合的第二抗原结合结构域是常规fab片段。在一个方面,能够与epcam特异性结合的第二抗原结合结构域是fab分子,其中在恒定结构域cl中,第123位(根据kabat eu索引编号)处的氨基酸被选自赖氨酸(k)、精氨酸(r)或组氨酸(h)的氨基酸取代,并且第124位(根据kabat eu索引编号)处的氨基酸独立地被赖氨酸(k)、精氨酸(r)或组氨酸(h)取代,并且其中在恒定结构域ch1中,第147位(根据kabat eu索引编号)处的氨基酸独立地被谷氨酸(e)或天冬氨酸(d)取代,并且第213位(根据kabat eu索引编号)处的氨基酸独立地被谷氨酸(e)或天冬氨酸(d)取代(根据kabat eu索引编号)。
48、在一个特定的方面,提供了双特异性激动性cd28抗原结合分子,其包含
49、(i)含有seq id no:92的氨基酸序列的第一轻链、含有seq id no:91的氨基酸序列的第一重链、含有seq id no:104的氨基酸序列的第二重链和含有seq id no:105的氨基酸序列的第二轻链,或者
50、(ii)含有seq id no:92的氨基酸序列的第一轻链、含有seq id no:91的氨基酸序列的第一重链、含有seq id no:100的氨基酸序列的第二重链和含有seq id no:101的氨基酸序列的第二轻链。
51、在进一步的方面,提供了如本文所述的双特异性激动性cd28抗原结合分子,其中能够与epcam特异性结合的第二抗原结合结构域为fab分子,其中fab轻链和fab重链的可变结构域vl和vh彼此替换或恒定结构域cl和ch1彼此替换,特别是可变结构域vl和vh彼此替换。在一个方面,能够与cd28特异性结合的第一抗原结合结构域是常规fab分子。在一个方面,能够与cd28特异性结合的第一抗原结合结构域是fab分子,其中在恒定结构域cl中,第123位(根据kabat eu索引编号)氨基酸被选自赖氨酸(k)、精氨酸(r)或组氨酸(h)的氨基酸取代,并且第124位(根据kabat eu索引编号)氨基酸独立地被赖氨酸(k)、精氨酸(r)或组氨酸(h)取代,其中在恒定结构域ch1中,第147位(根据kabat eu索引编号)氨基酸独立地被谷氨酸(e)或天冬氨酸(d)取代,并且第213位(根据kabat eu索引编号)氨基酸独立地被谷氨酸(e)或天冬氨酸(d)取代(根据kabat eu索引编号)。
52、在一个特定的方面,提供了双特异性激动性cd28抗原结合分子,其包含
53、(i)含有seq id no:83的氨基酸序列的第一轻链、含有seq id no:74的氨基酸序列的第一重链、含有seq id no:271的氨基酸序列的第二重链和含有seq id no:272的氨基酸序列的第二轻链,或者
54、(ii)含有seq id no:83的氨基酸序列的第一轻链、含有seq id no:74的氨基酸序列的第一重链、含有seq id no:273的氨基酸序列的第二重链和含有seq id no:272的氨基酸序列的第二轻链,或者
55、(iii)含有seq id no:83的氨基酸序列的第一轻链、含有seq id no:74的氨基酸序列的第一重链、含有seq id no:274的氨基酸序列的第二重链和含有seq id no:272的氨基酸序列的第二轻链,或者
56、(iv)含有seq id no:83的氨基酸序列的第一轻链、含有seq id no:74的氨基酸序列的第一重链、含有seq id no:275的氨基酸序列的第二重链和含有seq id no:272的氨基酸序列的第二轻链,或者
57、(v)含有seq id no:83的氨基酸序列的第一轻链、含有seq id no:74的氨基酸序列的第一重链、含有seq id no:273的氨基酸序列的第二重链和含有seq id no:276的氨基酸序列的第二轻链。
58、在另一个方面,提供了如本文所述的双特异性激动性cd28抗原结合分子,其中第一抗原结合结构域和第二抗原结合结构域各自是fab分子,并且fc结构域由能够稳定缔合的第一亚基和第二亚基构成;并且其中(i)第一抗原结合结构域在fab重链的c末端与fc结构域的第一亚基的n末端融合,并且第二抗原结合结构域在fab重链的c末端与fc结构域的第二亚基的n末端融合,或(ii)第二抗原结合结构域在fab重链的c末端与fc结构域的第一亚基的n末端融合,并且第一抗原结合结构域在fab重链的c末端与该fc结构域的第二亚基的n末端融合。在一个方面,fc结构域包含促进fc结构域的第一亚基和第二亚基缔合的修饰。在一个方面,fc结构域的第一亚基包含氨基酸取代s354c和t366w(eu编号),fc结构域的第二亚基包含氨基酸取代y349c、t366s和y407v(根据kabat eu索引编号)。
59、根据本发明的另一个方面,提供了一种或多种经分离的多核苷酸,其编码本发明的双特异性激动性cd28抗原结合分子。本发明进一步提供了一种或多种载体(特别是表达载体)和宿主细胞,该载体包含本发明的经分离的多核苷酸,该宿主细胞包含本发明的经分离的多核苷酸或表达载体。在一些方面,宿主细胞是真核细胞,特别是哺乳动物细胞。在另一个方面,提供了制备如本文所述的双特异性激动性cd28抗原结合分子的方法,该方法包含在适合于双特异性激动性cd28抗原结合分子的表达的条件下培养宿主细胞。任选地,该方法还包含回收双特异性激动性cd28抗原结合分子。本发明还涵盖通过本发明的方法制备的双特异性激动性cd28抗原结合分子。
60、本发明进一步提供了药物组合物,该药物组合物包含本发明的双特异性激动性cd28抗原结合分子和至少一种药用赋形剂。在一个方面,药物组合物用于疾病,特别是癌症的治疗。
61、本发明还包括使用本发明的双特异性激动性cd28抗原结合分子或药物组合物的方法。在一个方面,本发明提供了用作药物的根据本发明的双特异性激动性cd28抗原结合分子或药物组合物。在一个方面,提供了用于(a)增强细胞活化或(b)增强t细胞效应子功能的如本文所述的双特异性激动性cd28抗原结合分子。在一个方面,提供了用于疾病的治疗的根据本发明的双特异性激动性cd28抗原结合分子或药物组合物。在具体方面,疾病为癌症。在另一个方面,提供了根据本发明的用于癌症的治疗的双特异性激动性cd28抗原结合分子或药物组合物,其中该激动性cd28抗原结合分子用于与化疗剂、放射疗法和/或用于癌症免疫疗法的其他药剂组合施用。在进一步的方面,提供了用于癌症的治疗的双特异性激动性cd28抗原结合分子或药物组合物,其中双特异性激动性cd28抗原结合分子用于与t细胞活化性抗cd3双特异性抗体组合施用。在又一个方面,提供用于在癌症的治疗中使用的双特异性激动性cd28抗原结合分子或药物组合物,其中双特异性激动性cd28抗原结合分子用于与抗pd-l1抗体或抗pd-1抗体组合施用。
62、还提供了根据本发明的双特异性激动性cd28抗原结合分子或药物组合物在制备用于治疗疾病的药物中的用途;以及治疗个体疾病的方法,该方法包含向该个体施用治疗有效量的根据本发明的双特异性激动性cd28抗原结合分子或以药用形式包含根据本发明的双特异性激动性cd28抗原结合分子的组合物。在具体方面,疾病为癌症。在一个方面,提供了一种方法(a)增强个体中的细胞活化或(b)增强t细胞效应子功能,包括向所述个体以药用形式施用根据本发明的双特异性激动性cd28抗原结合分子或包含该双特异性激动性cd28抗原结合分子的组合物。在另一个方面,提供了根据本发明的双特异性激动性cd28抗原结合分子在制备用于治疗疾病的药物中的用途,其中该治疗包含与化疗剂、放射疗法和/或用于癌症免疫疗法的其他药剂共同施用。在进一步方面,提供了治疗个体中的疾病的方法,该方法包含向该个体施用治疗有效量的根据本发明的双特异性激动性cd28抗原结合分子或以药用形式包含根据本发明的双特异性激动性cd28抗原结合分子的组合物,其中该方法包含与化疗剂、放射疗法和/或用于癌症免疫疗法的其他药剂共同施用。在进一步方面,提供了治疗个体中的疾病的方法,包含向该个体施用治疗有效量的根据本发明的双特异性激动性cd28抗原结合分子或以药用形式包含根据本发明的双特异性激动性cd28抗原结合分子的组合物,其中该方法包含与t细胞活化抗cd3双特异性抗体共同施用。在另一个方面,提供了治疗个体中的疾病的方法,该方法包含向该个体施用治疗有效量的根据本发明的双特异性激动性cd28抗原结合分子或以药用形式包含根据本发明的双特异性激动性cd28抗原结合分子的组合物,其中该方法包含与抗pd-l1抗体或抗pd-1抗体共同施用。还提供了抑制个体中肿瘤细胞生长的方法,该方法包含向个体施用有效量的根据本发明的双特异性激动性cd28抗原结合分子,或以药用形式包含根据本发明的双特异性激动性cd28抗原结合分子的组合物,以抑制肿瘤细胞的生长。在上述方面中的任何方面,个体优选为哺乳动物,特别是人。
- 还没有人留言评论。精彩留言会获得点赞!