调节细胞表达的生物活性的结合分子的制作方法
本发明涉及结合分子领域。具体地讲,其涉及用于治疗涉及变异细胞的疾病的治疗性结合分子领域。更具体地,其涉及结合两种或更多种膜结合蛋白的细胞外部分的结合分子,从而调节细胞表达的生物活性。
背景技术:
1、尽管在治疗疾病方面已经取得了许多进展并且关于导致癌症的分子事件的知识增加,但癌症仍然是世界上发病和死亡的主要原因。这是世界范围内第二位致死原因。根据世界卫生组织(world health organization),癌症在2015年造成880万人死亡。在全球范围内,近六分之一的死亡由癌症引起。例如,结肠直肠癌(crc)是全球第三大常见癌症。在2008年,123万人被诊断患有该疾病。其是欧洲第二大常见癌症,在2012年诊断出约447,000新病例(总数的13%)。结肠直肠癌是癌症死亡的第四大常见原因,据估计每年导致608,000(eu 148,000)人死亡。虽然crc方面的一些新疗法已经取得了进展,但许多在临床试验上失败了;转移性crc在很大程度上仍然不可治愈。
2、传统上,大多数癌症药物发现聚焦于阻断基本细胞功能和杀死分裂细胞的药物。然而,在晚期癌症的情况下,无论多么具有攻击性地施用,甚至是到患者因治疗遭受危及生命的副作用的程度,化学疗法也很难实现完全治愈。在大多数情况下,患者的肿瘤停止生长或暂时萎缩(称为缓解)仅仅是为了再次开始增殖,有时更快增殖(称为复发),并且变得越来越难以治疗。最近,癌症药物开发的焦点已经从广泛细胞毒性的化学疗法转向具有较低毒性的靶向细胞抑制疗法。晚期癌症治疗已在白血病及一些其他癌症中得到临床验证。然而,在大多数癌症中,靶向方法仍被证明不足以有效地完全消除大多数患者的癌症。黑素瘤是非常频繁发生的癌症的另一个示例。当没有及早地检测出时,癌症很可能在极难以治疗的阶段转移。免疫干预治疗已显示对至少某些黑素瘤已经转移的患者有效。非小细胞肺癌是一种很难在外科手术的早先阶段发现的癌症类型。另外,这些类型的癌症已通过免疫干预疗法成功地治疗。
3、对癌症的靶向已使用多种不同的方法实现,包括例如靶向癌症存活和/或生长依赖于其的信号蛋白的小分子;含有肿瘤特异性蛋白的疫苗;采用有效地杀伤肿瘤细胞的免疫细胞以及使细胞毒性分子靶向肿瘤;干扰信号转导和/或使宿主免疫系统(重新)定向至肿瘤细胞的抗体的细胞疗法。阻断ctla-4或pd-1轴的单克隆抗体已示出诱导黑素瘤、nsclc、肾细胞癌和尿路上皮癌患者亚群的持久临床反应。
4、本发明提供了用于(重新)定向免疫系统组分的新型装置和方法。本发明还涉及用于调节细胞所表达的生物活性的装置和方法。
技术实现思路
1、本发明提供了一种刺激细胞上tnf受体超家族的成员的活性的方法,包括提供第一细胞和第二细胞,其中第一细胞在细胞膜上具有所述成员并且所述第二细胞在细胞膜上具有第二膜蛋白,该方法包括使所述细胞与包含两个抗原结合位点的结合分子接触,其中第一抗原结合位点能够结合所述成员(第一膜蛋白)的细胞外部分,并且第二抗原结合位点能够结合所述第二膜蛋白的细胞外部分,从而刺激所述第一细胞上所述成员的活性。在一些实施方案中,所述方法是体外方法。在一些实施方案中,所述tnf受体超家族的成员是cd137或ox40,优选地cd137。在一些实施方案中,所述第二膜蛋白不为tnf受体超家族的成员。在一些实施方案中,所述第二膜蛋白是b7家族的成员。在一些实施方案中,所述第二膜蛋白是pd-l1。
2、在一个优选的实施方案中,该方法还包括提供包含能够结合所述tnf受体超家族的成员的细胞外部分的抗原结合位点以及能够结合所述第二膜蛋白的细胞外部分的抗原结合位点的另一结合分子(第二结合分子),其中所述第一结合分子和第二结合分子结合:
3、-所述第一膜蛋白上的不同表位;
4、-所述第二膜蛋白上的不同表位;或者
5、-所述第一膜蛋白上的不同表位以及所述第二膜蛋白上的不同表位;
6、该方法还包括将所述第一细胞和所述第二细胞与所述第一结合分子和所述第二结合分子一起温育,从而刺激或提高所述第一细胞上所述tnf受体超家族的成员的活化。
7、本发明还提供一种包含能够结合tnf受体超家族的成员(第一膜蛋白)的细胞外部分的抗原结合位点以及能够结合第二膜蛋白的细胞外部分的抗原结合位点的结合分子。tnf受体超家族成员优选地为cd137或ox40,优选地cd137。在一些实施方案中,所述第二膜蛋白不为tnf受体超家族的成员。所述第二膜蛋白优选地为b7家族的成员。在一些实施方案中,所述第二膜蛋白是pd-l1。
8、本发明还提供包含一种或多种结合分子的组合物或成套试剂盒,所述结合分子包含能够结合tnf受体超家族的成员(第一膜蛋白)的细胞外部分的抗原结合位点以及能够结合第二膜蛋白的细胞外部分的抗原结合位点。在一个优选的实施方案中,本发明提供了包含两种或更多种此类结合分子的组合物或成套试剂盒;其中至少两种所述结合分子能够结合:
9、-所述第一膜蛋白上的不同表位;
10、-所述第二膜蛋白上的不同表位;或者
11、-所述第一膜蛋白上的不同表位以及所述第二膜蛋白上的不同表位。优选的是至少两种结合分子结合所述第一膜蛋白上相同的表位,以及结合所述第二膜蛋白上不同的表位。
12、本发明还提供了刺激细胞上的cd137或ox40的活性的方法,该方法包括提供第一细胞和第二细胞,其中所述第一细胞在细胞膜上具有cd137或ox40并且所述第二细胞在细胞膜上具有第二膜蛋白,并且使所述第一细胞和所述第二细胞与如下结合分子(第一结合分子)相接触:包含能够结合至所述cd137或ox40(第一膜蛋白)的细胞外部分的抗原结合位点;以及能够结合至第二膜蛋白的细胞外部分的抗原结合位点;该方法还包括使所述第一细胞和所述第二细胞与所述第一结合分子一起温育,从而刺激所述第一细胞上所述cd137或ox40的活性。在一些实施方案中,所述第二膜蛋白不为tnf受体超家族的成员。在一些实施方案中,所述方法是体外方法。
13、在一个优选的实施方案中,该方法还包括提供另一结合分子(第二结合分子),所述另一结合分子包含能够结合所述第一膜蛋白的细胞外部分的抗原结合位点;以及能够结合所述第二膜蛋白的细胞外部分的抗原结合位点,其中所述第一结合分子和第二结合分子结合:
14、-所述第一膜蛋白上的不同表位;
15、-所述第二膜蛋白上的不同表位;或者
16、-所述第一膜蛋白上的不同表位;以及所述第二膜蛋白上的不同表位;
17、该方法还包括使所述第一细胞和所述第二细胞与所述第一结合分子和所述第二结合分子一起温育,从而刺激所述第一细胞上cd137或ox40的活性。
18、在一些实施方案中,根据本发明的结合分子包含能够结合tnf受体超家族的成员的细胞外部分的抗原结合位点以及能够结合b7家族的成员的抗原结合位点。在一些实施方案中,根据本发明的所述结合分子的抗原结合位点由能够结合tnf受体超家族的成员的细胞外部分的一个抗原结合位点以及能够结合b7家族的成员的一个抗原结合位点组成。在一些实施方案中,根据本发明的所述结合分子包含能够结合cd137的细胞外部分的抗原结合位点以及能够结合b7家族的成员的抗原结合位点。在一些实施方案中,根据本发明的所述结合分子的抗原结合位点由能够结合cd137的细胞外部分的一个抗原结合位点以及能够结合b7家族的成员的一个抗原结合位点组成。在一些实施方案中,根据本发明的所述结合分子包含能够结合cd137的抗原结合位点以及能够结合pd-l1的抗原结合位点。在一些实施方案中,根据本发明的所述结合分子的抗原结合位点由能够结合cd137的一个抗原结合位点以及能够结合pd-l1的一个抗原结合位点组成。在一些实施方案中,根据本发明的所述结合分子具有不超过两个抗原结合位点。
19、如本文所述的结合分子优选地为抗体。
20、本发明还提供了一种刺激细胞上tnf受体超家族的成员的活性的方法,包括提供第一细胞和第二细胞,其中所述第一细胞在细胞膜上具有所述成员(第一膜蛋白)并且所述第二细胞在细胞膜上具有第二膜蛋白,该方法包括使所述细胞与包含至少两个可变结构域的根据本发明的抗体接触,其中一个可变结构域包含能够结合所述第一膜蛋白的细胞外部分的第一抗原结合位点,并且另一个可变结构域包含能够结合所述第二膜蛋白的细胞外部分的第二抗原结合位点,从而刺激所述第一细胞上所述成员的活性。在一些实施方案中,所述方法是体外方法。
21、本发明还提供一种抗体或其功能部分、衍生物和/或类似物,其包含:
22、-能够结合tnf受体超家族的成员(第一膜蛋白)的细胞外部分的可变结构域;并且
23、-能够结合第二膜蛋白的细胞外部分的可变结构域。第一膜蛋白优选地为cd137或ox40,优选地cd137。在一些实施方案中,所述第二膜蛋白不为tnf受体超家族的成员。
24、结合分子优选地为双特异性抗体。本发明还提供了一种刺激细胞上tnf受体超家族的成员的活性的方法,包括提供第一细胞和第二细胞,其中所述第一细胞在细胞膜上具有所述成员(第一膜蛋白)并且所述第二细胞在细胞膜上具有第二膜蛋白,该方法包括使所述细胞与包含两个可变结构域的双特异性抗体接触,其中一个可变结构域包含能够结合所述第一膜蛋白的细胞外部分的第一抗原结合位点,并且另一个可变结构域包含能够结合所述第二膜蛋白的细胞外部分的第二抗原结合位点,从而刺激所述第一细胞上所述成员的活性。在一些实施方案中,所述方法是体外方法。还提供了一种双特异性抗体,该双特异性抗体包含具有能够结合tnf受体超家族成员(第一膜蛋白)的细胞外部分的抗原结合位点的可变结构域以及具有能够结合第二膜蛋白的细胞外部分的抗原结合位点的可变结构域。第一膜蛋白优选地为cd137或ox40,优选地cd137。第二膜蛋白优选地不为tnf受体超家族的成员。
25、在一些实施方案中,根据本发明的抗体包含含有能够结合tnf受体超家族的成员的细胞外部分的抗原结合位点以及能够结合b7家族的成员的一个抗原结合位点的可变结构域。在一些实施方案中,根据本发明的所述抗体的抗原结合位点由能够结合tnf受体超家族的成员的细胞外部分的一个抗原结合位点以及能够结合b7家族的成员的一个抗原结合位点组成。在一些实施方案中,根据本发明的所述抗体包含能够结合cd137的细胞外部分的抗原结合位点以及能够结合b7家族的成员的抗原结合位点。在一些实施方案中,根据本发明的所述抗体的抗原结合位点由能够结合cd137的细胞外部分的一个抗原结合位点以及能够结合b7家族的成员的一个抗原结合位点组成。在一些实施方案中,根据本发明的所述抗体包含能够结合cd137的抗原结合位点以及能够结合pd-l1的抗原结合位点。在一些实施方案中,根据本发明的所述抗体的抗原结合位点由能够结合cd137的一个抗原结合位点以及能够结合pd-l1的一个抗原结合位点组成。在一些实施方案中,根据本发明的所述抗体具有不超过两个抗原结合位点。
26、还提供了一种药物组合物,该药物组合物包含一种或多种结合分子,优选地本发明的抗体或其变体。
27、还提供了编码本发明的抗体或其变体的重链或重链可变区的一种核酸分子或核酸分子的集合体。
28、还提供了编码本发明的抗体的一种核酸分子或核酸分子的集合体。
29、本发明的抗体优选地包含含有如图3所示的mf的氨基酸序列的重链可变区。在一个优选的实施方案中,该抗体还包含含有如图1所示的轻链可变区的氨基酸序列的轻链可变区。在一个优选的实施方案中,轻链包含如图1a所示的氨基酸序列。在一个优选的实施方案中,重链包含igg1抗体,优选地人igg1抗体的恒定区。在一个优选的实施方案中,所述igg1恒定区的ch2区被工程化成使抗体的adcc和/或cdc活性下降。在一个优选的实施方案中,ch2区包含如图2e所示的序列。在一个优选的实施方案中,抗体的ch3区被工程化成有利于重链异源二聚化。在一个优选的实施方案中,一个重链包含如图2f所示的序列,且另一个重链包含如图2g所示的序列。
30、还提供了包含单独或一起编码本发明的抗体或其变体的一种或多种核酸分子的细胞。还提供了使用如所述的细胞产生本发明的抗体或其变体,优选地与此同时从细胞的培养物收获抗体或其变体的方法。
31、还提供了包含本发明的抗体或其变体的细胞体系。
32、还提供了一种用于治疗患有涉及异常细胞的疾病(诸如癌症)或患有慢性病毒或寄生物感染的个体的方法,该方法包括向对其有需要的个体施用结合分子,优选地本发明的抗体或其变体。
33、本发明还提供了一种结合分子,优选地本发明的抗体或其变体;优选地本发明的双特异性抗体或其变体用于治疗患有涉及异常细胞的疾病(诸如癌症)或患有慢性病毒或寄生物感染的个体的用途。
34、在一个优选的实施方案中,寄生物是细胞内寄生物。
35、还提供了一种在所述个体中刺激个体针对异常细胞的免疫应答的方法,该方法包括提供给(施用于)所述个体结合分子,优选地抗体或其变体,优选地本发明的双特异性抗体或其变体。异常细胞优选地为癌细胞、病毒感染细胞、寄生虫或寄生物感染细胞。在一个优选的实施方案中,细胞是癌细胞或赘生细胞。
36、发明详述
37、肿瘤坏死因子受体超家族(tnfrsf)是一类受体。它们的特征通常在于能够经由富含半胱氨酸的细胞外结构域结合肿瘤坏死因子(tnf)。除神经生长因子(ngf)之外,所有tnf均与原型tnfα同源。在其活性形式中,大多数tnf受体在质膜中形成三聚体复合物。因此,大多数tnf受体包含跨膜结构域(tmd)并且位于细胞膜上。然而,一些能够裂解为可溶形式(例如tnfr1),并且一些完全缺乏tmd(例如dcr3)。结合tnf受体超家族成员的本发明抗体结合至超家族的膜结合成员。仅以未与细胞膜结合的形式存在的成员不在本发明的范围之内。
38、tnf受体在结合受体的配体时参与信号转导到细胞内部。某些受体需要特异性衔接蛋白诸如tradd、traf、rip和fadd用于下游信号转导。在本发明的上下文中,tnf超家族的各成员是优选的。这些包括肿瘤坏死因子受体1;肿瘤坏死因子受体2;淋巴毒素β受体;ox40;cd40;fas受体;cd27;cd30;cd137;死亡受体3;死亡受体4;死亡受体5;死亡受体6;rank;troy;baff受体;b细胞成熟抗原(bcma)以及跨膜激活蛋白和钙离子信号调节亲环素配体相互作用蛋白(taci)。
39、肿瘤坏死因子受体1是肿瘤坏死因子α的主要受体之一。该受体具有多个别名,其中一些为肿瘤坏死因子受体超家族成员1a;tnfrsf1a;tnf-r1;tnf-ri;tnfr-i;tnfr1;tnfar;p60;p55;肿瘤坏死因子受体1a同种型β;肿瘤坏死因子结合蛋白1;肿瘤坏死因子受体1型;肿瘤坏死因子受体i型;肿瘤坏死因子-α受体;肿瘤坏死因子受体1;cd120a抗原;tnfr1-d2;tnf-r-i;tnf-r55;cd120a;tnfr55;tnfr60;tnf-r;p55-r;tbp1;fpf;以及ms5。肿瘤坏死因子受体1的外部标识为hgnc:11916;entrez基因:7132;ensembl:ensg00000067182;omim:191190和uniprotkb:p19438。
40、肿瘤坏死因子受体2是结合肿瘤坏死因子α(tnfα)的膜受体。该受体具有多个别名,其中一些为:肿瘤坏死因子受体超家族成员1b;tnfrsf1b;肿瘤坏死因子受体ii型;肿瘤坏死因子受体2;p80tnf-α受体;tnf-rii;tnf-r2;tnfr2;tnfbr;p75;肿瘤坏死因子结合蛋白2;肿瘤坏死因子β受体;p75tnf受体;cd120b抗原;依那西普;tnf-r-ii;tnf-r75;p75tnfr;tnfr-ii;cd120b;tnfr1b;tnfr80;tbpii。肿瘤坏死因子受体2的外部标识为hgnc:11917;entrez基因:7133;ensembl:ensg00000028137;omim:191191;以及uniprotkb:p20333。
41、淋巴毒素β受体表达于大多数细胞类型的表面,包括上皮细胞和骨髓谱系细胞,但通常不表达于正常t和b淋巴细胞上。蛋白质结合淋巴毒素膜形式(淋巴毒素α与淋巴毒素β的复合物)。编码的蛋白质及其配体在淋巴组织和转化细胞的发育和组织化中发挥作用。蛋白质的激活可例如引发细胞凋亡。蛋白质以大量别名已知,其中有:ltbr;肿瘤坏死因子受体2相关蛋白;肿瘤坏死因子受体iii型;肿瘤坏死因子c受体;d12s370;tnfrsf3;tnfcr;tnfr3;淋巴毒素β受体(tnfr超家族成员3);淋巴毒素b受体;lt-beta-r;tnf-r-iii;tnfr2-rp;tnf-riii;tnfr-iii;tnfr-rp;以及cd18。淋巴毒素β受体的外部标识为hgnc:6718;entrez基因:4055;ensembl:ensg00000111321;omim:600979和uniprotkb:p36941。
42、ox40不在静息幼稚t细胞上组成型表达,不同于cd28,ox40是在活化后24小时至72小时之后表达的次级共同刺激免疫检查点分子;其配体ox40l也不表达于静息抗原递呈细胞上,但在它们活化之后表达。ox40的表达取决于t细胞的活化。在不具有cd28的情况下,ox40的表达通常延迟并且以较低水平存在。蛋白质以大量别名已知,其中有:tnfrsf4;肿瘤坏死因子受体超家族成员4;tax转录活化的糖蛋白1受体;ox40l受体;act35抗原;cd134抗原;txgp1l;tax转录活化的糖蛋白1受体;淋巴细胞活化抗原act35(lymphoid activationantigene act35);ox40细胞表面抗原;atc35抗原;ox40抗原;act35;cd134;以及imd16。ox40的外部标识为hgnc:11918;entrez基因:7293;ensembl:ensg00000186827;omim:600315和uniprotkb:p43489。
43、cd40是存在于抗原递呈细胞上的共刺激蛋白质并且参与它们的活化。t辅助细胞上的cd154(cd40l)与cd40的结合活化了抗原递呈细胞并且诱导cd40的细胞内信号转导和多种下游效应。蛋白质以多种不同的别名已知,其中有:cd40分子;cd40分子tnf受体超家族成员5;cd40l受体;tnfrsf5;cdw40;bp50;肿瘤坏死因子受体超家族成员5;b细胞表面抗原cd40;b细胞表面抗原cd40;b细胞相关分子;cd40抗原;以及p50。cd40的外部标识为hgnc:11919;entrez基因:958;ensembl:ensg00000101017;omim:109535;以及uniprotkb:p25942。
44、fas受体是细胞表面上的致使程序性细胞死亡(细胞凋亡)的死亡受体。其构成一种较显著细胞凋亡途径的一部分。其以多个另选方案已知:诸如fas细胞表面死亡受体;肿瘤坏死因子受体超家族成员6;细胞凋亡介导的表面抗原fas;tnf受体超家族成员6;faslg受体;cd95抗原;tnfrsf6;apt1;fas1;apo-1细胞表面抗原;细胞凋亡抗原1;apo-1抗原;fasama;alps1a;apo-1;fastm;以及cd95。fas的外部标识为hgnc:11920;entrez基因:355;ensembl:ensg00000026103;omim:134637;以及uniprotkb:p25445。
45、据信cd27对于t细胞免疫的产生和长期维持很重要。其结合至配体cd70,并且在调节b细胞活化和免疫球蛋白合成中发挥作用。cd27转导致使nf-κb和mapk8/jnk活化的信号。cd27结合蛋白(siva)作为促凋亡蛋白能够结合该受体并且据信在该受体所诱导的细胞凋亡中发挥作用。该蛋白质的别名其中有:cd27分子;肿瘤坏死因子受体超家族成员7;t细胞激活抗原cd27;cd27l受体;tnfrsf7;t14;t细胞激活抗原s152;cd27抗原;s152。lpfs2;s152和tp55。cd27的外部标识为hgnc:11922;entrez基因:939;ensembl:ensg00000139193;omim:186711;以及uniprotkb:p26842。
46、cd30被活化的t和b细胞表达。据信traf2和traf5与该受体相互作用,并且介导致使nf-kb活化的信号转导。其为细胞凋亡的正调节因子,并且也示出限制自身反应性cd8效应t细胞的增殖潜能并且保护机体免于自身免疫。其以多个不同的名称已知,诸如tnfrsf8;肿瘤坏死因子受体超家族成员8;淋巴细胞活化抗原cd30;cd30l受体;ki-1抗原;d1s166e;cd30;细胞因子受体cd30;cd30抗原;以及ki-1。cd30的外部标识为hgnc:11923;entrez基因:943;ensembl:ensg00000120949;omim:153243;以及uniprotkb:p28908。
47、cd137可被活化的t细胞表达。其也存在于其他细胞上,诸如树突状细胞、自然杀伤细胞、粒细胞以及炎症部位的血管壁细胞。该蛋白质因其对t细胞活化的共刺激活性而为人所知。cd137以多个不同的名称已知,诸如tnfrsf9;tnf受体超家族成员9;肿瘤坏死因子受体超家族成员9;t细胞抗原4-1bb同系物;4-1bb配体受体;t细胞抗原ila;cd137抗原;cdw137;ila;白介素激活受体,鼠ly63的同系物;淋巴细胞活化诱导(ila);小鼠4-1bb的同系物;受体蛋白4-1bb;t细胞抗原ila;以及4-1bb。cd137的外部标识为hgnc:11924;entrez基因:3604;ensembl:ensg00000049249;omim:602250;以及uniprotkb:q07011。cd137是最常见地在活化cd8+ t细胞上有所上调的诱导型受体。cd137信号转导通过活化nf-κb来增强t细胞功能[arch等人,1998]。其他细胞免疫细胞类型,包括cd4+ t细胞、单核细胞、b细胞、树突状细胞(dc)亚群以及粒细胞和nk细胞能够以各种水平表达cd137[shao等人,2011]。在单核细胞中,cd137通过被脂多糖(lps)和il-1β活化而诱导。在b淋巴细胞中,cd137表达由针对细胞表面免疫球蛋白的抗体和ebv的转化而诱导。在dc中,除增强其产生炎性细胞因子(il-6和il-12)及其存活之外,cd137的连接还通过上调b7共同刺激分子(cd80和cd86)诱导其成熟[makkouk等人,2015]。中性粒上cd137连接的天然功能是增大细菌和寄生物感染的吞噬作用。此外,cd137的连接阻断体外中性粒和嗜酸性粒细胞中il-3/il-5/gm-csf受体所介导的抗细胞凋亡信号,从而防止粒细胞积聚[simon,2001;vinay等人,2011]。在非淋巴细胞诸如软骨细胞、内皮细胞和肿瘤细胞中,cd137表达由细胞因子刺激所驱动,诸如对于软骨细胞为il-1β,对于内皮细胞为炎性细胞因子tnfα/ifnγ/il-1β且对于肿瘤细胞为ifnγ。刺激cd137的配体(cd137l)表达于活化抗原递呈细胞上。cd137作为单体和二聚体存在于膜中[pollok等人,1993]。
48、死亡受体3由活化和经历过抗原的t淋巴细胞表达。受体也由foxp3阳性调节性t淋巴细胞表达。该受体的配体为tl1a(tnfsf15),其在toll样受体或fc受体活化之后,在抗原递呈细胞和一些内皮细胞中有所上调。已经报道了编码不同同种型的各种可变剪接的转录变体,它们大多数是潜在的分泌分子。据信该受体参与控制t细胞活化所诱导的淋巴细胞增殖。据信活化取决于t细胞受体的先前结合。配体的结合经由il-2受体提高t细胞对内源性il-2的敏感性并且增强t细胞增殖。体内活化对于遇到同源抗原的那些t细胞可能是特异性的。在静息时,并且对于不具有潜在自身免疫性的个体,经常遇到同源抗原的大多数t细胞是foxp3+调节性t细胞。在不存在任何其他外源信号情况下刺激的死亡受体3刺激了foxp3+调节性(cd4阳性)t细胞的显著和特异性增殖。死亡受体3的治疗性激动剂可用于刺激treg扩增,这可减少哮喘、实体器官的同种异体移植和眼角膜炎的实验模型中的炎症。另一方面,受体连同自身抗原或疫苗抗原的共同刺激可分别导致免疫病理学恶化或增强的疫苗刺激的免疫。受体刺激对于t细胞介导的免疫是特异性的,其可用于增强或减弱炎症,这取决于外来抗原相对于自体抗原可用性的时间背景和质量。人体内tnfrsf25的刺激可导致类似但更可控的效应,原因是共同刺激阻断靶向分子诸如ctla-4和pd-1。死亡受体3也以多个其他名称已知,诸如tnfrsf25;肿瘤坏死因子受体超家族成员25;肿瘤坏死因子受体超家族成员12(易位链相关膜蛋白);淋巴细胞相关的死亡受体;细胞凋亡介导受体tramp;细胞凋亡介导受体dr3;细胞凋亡诱导受体air;蛋白质wsl-1;tnfrsf12;apo-3;ddr3;lard;dr3;细胞凋亡诱导受体;死亡受体β;蛋白质wsl;wsl-lr;tramp;wsl-1;apo3;wsl1;tr3;以及wsl。死亡受体3的外部标识为hgnc:11910;entrez基因:8718;ensembl:ensg00000215788;omim:603366;以及uniprotkb:q93038。
49、死亡受体4是tnf受体超家族的细胞表面受体,其结合trail并且据信转导细胞死亡信号并诱导细胞凋亡。其以多个名称已知,诸如tnfrsf10a;肿瘤坏死因子受体超家族成员10a;tnf相关细胞凋亡诱导配体受体1;死亡受体4;trail受体1;trail-r1;trailr1;apo2;dr4;肿瘤坏死因子受体超家族成员10a变体2;细胞毒性trail受体;cd261抗原;trailr-1和cd261。死亡受体4的外部标识为hgnc:11904;entrez基因:8797;ensembl:ensg00000104689;omim:603611;以及uniprotkb:o00220
50、死亡受体5是结合trail并且介导细胞凋亡的tnf受体超家族的细胞表面受体。该受体可由肿瘤坏死因子相关细胞凋亡诱导配体(tnfsf10/trail/apo-2l)活化,并且转导细胞凋亡信号。该受体以多种不同的名称已知,其中有:tnfrsf10b;肿瘤坏死因子受体超家族成员10b;tnf相关细胞凋亡诱导配体受体2;死亡受体5;trail-r2;trailr2;killer;trick2;ztnfr9;dr5;p53调节的dna损伤诱导型细胞死亡受体(killer);肿瘤坏死因子类受体蛋白ztnfr9;trail/apo-2l的含死亡域受体;细胞凋亡诱导性蛋白trick2a/2b;细胞凋亡诱导性受体trail-r2;tnf受体超家族成员10b;细胞毒性trail受体-2;fas样蛋白质;trail受体2;cd262抗原;killer/dr5;trick2a;trick2b;trickb;以及cd262。死亡受体5的外部标识为:hgnc:11905;entrez基因:8795;ensembl:ensg00000120889;omim:603612;以及uniprotkb:o14763。
51、死亡受体6可在活化时诱导细胞凋亡。小鼠的敲除研究表明,受体在t辅助细胞活化中发挥作用,并且可参与炎症和免疫调节。还认为受体涉及致使阿尔茨海默病的脑中神经退化以及应激反应的信号转导和细胞存活。过表达诱导表达细胞发生细胞凋亡。app(淀粉样前体蛋白)是该受体的天然配体并且首先裂解为aβ和n-app。n-app是与dr6相互作用以触发阿尔茨海默病患者轴突退化的片段。死亡受体6也以多个其他名称已知,诸如tnfrsf21;肿瘤坏死因子受体超家族成员21;dr6;tnfr相关死亡受体6;cd358抗原;bm-018;以及cd358。死亡受体6的外部标识为hgnc:13469;entrez基因:27242;ensembl:ensg00000146072;omim:605732;uniprotkb:o75509。
52、rank是rank-配体(rankl)的受体以及调节破骨细胞分化和活化的rank/rankl/opg信号转导途径的一部分。其与骨再造和修复、免疫细胞功能、淋巴结发育、热调节和乳腺发育相关联。骨保护素(opg)是rank的诱饵受体,并且通过竞争rankl来调节rank信号转导途径的刺激。rank的胞质结构域将信号传送给下游靶标诸如nf-κb和jnk。rank表达于骨骼肌、胸腺、肝脏、结肠、小肠、肾上腺、破骨细胞、乳腺上皮细胞、前列腺、血管细胞和胰腺中。通常nf-κb的活化由rankl介导,但单独rank的过表达也可激活nf-κb途径。rank以多个不同的名称已知,诸如tnfrsf11a;肿瘤坏死因子受体超家族成员11a nfkb激活因子;杂合性丢失,18,染色体区1;破骨细胞分化因子受体;nf-kb的受体激活因子;骨派吉特氏病2;odfr;肿瘤坏死因子受体超家族成员11a,nfkb激活因子;肿瘤坏死因子受体超家族成员11a,nfkb激活因子;核因子kb的受体激活因子;cd265抗原;loh18cr1;trancer;cd265;optb7;osts;pdb2;feo和ofe。rank的外部标识为:hgnc:11908;entrez基因:8792;ensembl:ensg00000141655;omim:603499和uniprotkb:q9y6q6。
53、baff受体是识别baff的tnf受体超家族的膜蛋白。b细胞活化因子(baff)增强体外b细胞存活,并且是外周b细胞群的调节因子。baff受体是baff的受体,并且为包含单一细胞外富苯丙氨酸结构域的iii型跨膜蛋白。认为该受体是baff介导的成熟b细胞存活所需的主要受体。baff也被tnf受体b细胞成熟抗原(bcma)以及跨膜激活蛋白和钙离子信号调节亲环素配体相互作用蛋白(taci)结合。baff受体以多个不同的名称已知,诸如tnfrsf13c;肿瘤坏死因子受体超家族成员13c;b细胞活化因子受体;blys受体3;baff-r;baffr;b细胞活化因子受体;cd268抗原;氟奋乃静;bromix;cd268;cvid4;以及br3。baff受体的外部标识为hgnc:17755;entrez基因:115650;ensembl:ensg00000159958;omim:606269;以及uniprotkb:q96rj3。
54、b细胞成熟抗原(bcma)是识别b细胞活化因子(baff)的tnf受体超家族的细胞表面受体。该受体优先地表达于成熟b淋巴细胞中,并且据信对于b细胞发育和自体免疫应答很重要。已显示该受体特异性结合肿瘤坏死因子(配体)超家族成员13b(tnfsf13b/tall-1/baff),并且导致nf-kb和mapk8/jnk活化。b细胞成熟抗原也以多个别名已知,诸如tnfrsf17;肿瘤坏死因子受体超家族成员17;b细胞成熟蛋白;bcm;b细胞成熟因子;cd269抗原;tnfrsf13a;以及cd269。bcma的外部标识为hgnc:11913;entrez基因:608;ensembl:ensg00000048462;omim:109545;以及uniprotkb:q02223。
55、跨膜激活蛋白和钙离子信号调节亲环素配体相互作用蛋白(taci)。该基因编码的蛋白质是肿瘤坏死因子(tnf)受体超家族的淋巴细胞特异性成员。其与钙调节剂和亲环蛋白配体(caml)相互作用。该蛋白质也可结合baff和april(tnsf13或cd256)。taci诱导转录因子nfat、ap1和nf-k-b活化,并且通过与tnf配体相互作用在体液免疫中发挥作用。taci缺陷型小鼠具有增加的脾b细胞和血清ig,这暗示着意指taci在b细胞存活中的潜在负调节作用。然而,更简单的解释可能是缺乏taci使得更多的循环baff变得可用,其能够结合至br3并且增加b细胞数。taci相关的疾病包括常见变异型免疫缺陷症2和免疫球蛋白a缺陷症2。编码taci的基因位于17号染色体上的史密斯-马吉利氏综合征区。taci也以多个其他名称已知,诸如tnfrsf13b即肿瘤坏死因子受体超家族成员13b;跨膜激活剂和caml相互作用子;肿瘤坏死因子受体13b;cd267抗原;tnfrsf14b;cd267;cvid2;igad2;cvid;以及ryzn3。taci的外部标识为hgnc:18153;entrez基因:23495;ensembl:ensg00000240505;omim:604907;以及uniprotkb:o14836。
56、troy在胚胎发育期间表达。其已示出在细胞中过表达时激活jnk信号转导途径。受体的活化可诱导细胞凋亡。据信受体在胚胎发育中发挥作用。描述了编码不同同种型的可变剪接的转录变体。troy也以多个其他名称已知,诸如tnfrsf19;肿瘤坏死因子受体超家族成员19;毒性及jnk信号途径诱导子(toxicity and jnk inducer);trade;taj;以及taj-α。troy的外部标识为hgnc:11915;entrez基因:55504;ensembl:ensg00000127863;omim:606122;以及uniprotkb:q9ns68。
57、b7家族包括多种结构上相关的细胞表面蛋白,其结合至调节免疫应答的淋巴细胞上的受体。淋巴细胞的活化通过细胞表面抗原特异性t细胞受体或b细胞受体的结合来引发。b7配体同时递送的附加信号也决定了这些细胞的免疫应答。这些所谓的“共刺激”或“共抑制”信号通过淋巴细胞上cd28家族受体由b7家族成员递送。b7家族成员与共刺激受体的结合增强免疫应答,并且与共抑制受体的结合减弱免疫应答。目前,据信以下成员为该家族的成员:b7.1(cd80)、b7.2(cd86)、诱导型共刺激因子配体(icos-l)、程序性死亡-1配体(pd-l1)、程序性死亡-2配体(pd-l2)、b7-h3(cd276)、b7-h4、b7-h5、b7-h6和b7-h7。b7家族成员表达于淋巴和非淋巴组织中。在具有b7家族基因突变的小鼠中,在免疫缺陷和自身免疫疾病的发展中示出成员对调节免疫应答的效应。b7配体所递送的信号的操纵已显示潜在地治疗自体免疫性疾病、炎性疾病和癌症。
58、结合tnf受体超家族的成员的细胞外部分以及b7家族成员的细胞外部分的根据本发明的结合分子或抗体或其变体所提供的优点在于,所期望的免疫应答能够被特别好地促进,因为b7家族成员向淋巴细胞递送“共刺激”或“共抑制”信号,从而增强或减弱免疫应答。因此,通过靶向b7家族的成员,可以增强刺激信号和/或抵消抑制信号,从而诱导或增强例如针对异常细胞的期望免疫应答。
59、cd80是存在于活化b细胞和单核细胞上的蛋白质,提供t细胞活化和存活所需的共刺激信号。其为t细胞表面上两种不同蛋白质的配体:cd28和ctla-4。当结合至cd28时,其与共刺激相关联,而结合至ctla4则与免疫应答的减弱相关联。cd80与cd86协同作用来活化t细胞。据报道cd80也结合pd-l1。cd80以多个其他名称已知,诸如cd80分子;cd80抗原;cd28抗原配体1;b7-1抗原;b淋巴细胞活化抗原b7;ctla-4反受体b7.1;活化b7-1抗原;cd28lg1;cd28lg;lab7;bb1;b7;共刺激因子cd80;cd80抗原;以及b7-1。cd80的外部标识为hgnc:1700;entrez基因:941;ensembl:ensg00000121594;omim:112203;以及uniprotkb:p33681。
60、cd86是表达于抗原递呈细胞上的蛋白质。其可以为t细胞活化和存活提供共刺激信号。其为t细胞表面上两种不同蛋白质的配体:cd28和ctla-4。当结合至cd28时,其与共刺激相关联,而结合至ctla4则与免疫应答的减弱相关联。cd86与cd80协同作用来活化t细胞。其以多个不同的名称已知,诸如cd86分子;cd86抗原;cd28抗原配体2;b7-2抗原;ctla-4反受体b7.2;cd28lg2;fun-1;bu63;b70;b淋巴细胞活化抗原b7-2;b淋巴细胞抗原b7-2;活化b7-2抗原;cd86抗原;lab72;以及b7-2。cd86的外部标识为hgnc:1705;entrez基因:942;ensembl:ensg00000114013;omim:601020;以及uniprotkb:p42081。
61、可诱导t细胞共刺激分子配体(icosl或cd275)由apc以及多种非血液组织组成型表达。表达可以在正在发生的炎症情况下有所下调。目前已知icosl与人体内的icos、cd28和ctla-4相互作用。icosl/cd28相互作用似乎共同刺激人t细胞对同种异体抗原的初次应答和记忆/回忆应答。据信icosl/ctla-4导致共抑制信号。icosl也称为icoslg;b7相关蛋白1;b7同系物2;b7样蛋白gl50;b7同系物2;b7rp-1;b7-h2;b7rp1;b7h2;跨膜蛋白b7-h2icos配体;cd275抗原;kiaa0653;icos-l;licos;以及gl50。icosl的外部标识为hgnc:17087;entrez基因:23308;ensembl:ensg00000160223;omim:605717;以及uniprotkb:o75144。
62、pd-l1是在特定事件诸如妊娠、组织同种异体移植物、自身免疫疾病和其他疾病状态诸如肝炎期间抑制免疫应答中发挥作用的1型跨膜蛋白。pd-l1表达于各种类型的癌症中,尤其是nsclc(boland等人,2013;velcheti等人,2014)、黑素瘤、肾细胞癌、胃癌、肝细胞癌以及各种白血病和多发性骨髓瘤(bernstein等人,2014;thompson等人,2005)。pd-l1存在于癌细胞的细胞质和质膜中,但并非所有癌症或肿瘤内的所有细胞均表达pd-l1(dong等人,2002)。通过上调pd-l1表达,多个肿瘤微环境细胞有助于免疫抑制。该效应被称为“适应性免疫抗性”,因为肿瘤通过响应于活化t细胞所产生的ifn-γ而诱导pd-l1来保护自身(sharma等人,2017)。pd-l1也可通过癌基因调节,该机制被称为固有免疫抗性(akbay等人,2013)。在肿瘤微环境内,pd-l1也表达于骨髓细胞和活化t细胞上(tumeh等人,2014)。pd-l1的表达由多种促炎分子诱导产生,包括i和ii型ifn-γ、tnf-α、lps、gm-csf和vegf,以及细胞因子il-10和il-4,并且ifn-γ是最有效的诱导物(kondo等人,2010;sznol和chen,2013)。
63、pd-l1与pd-1或b7.1(cd80)的结合传送抑制信号,其使表达pd-1的t细胞的增殖下降。据信pd-1能够通过细胞凋亡来控制外来抗原特异性t细胞积聚。pd-l1由多种癌细胞表达,并且据信其表达至少部分地是造成针对癌细胞的免疫应答减弱的原因。pd-l1是b7家族蛋白质的成员并且以多种其他名称已知,诸如cd274分子;cd274抗原;b7同系物1;pdcd1配体1;pdcd1lg1;pdcd1l1;b7h1;pdl1;程序性细胞死亡1配体1;程序性死亡配体1;b7-h1;以及b7-h。cd274的外部标识为hgnc:17635;entrez基因:29126;ensembl:ensg00000120217;omim:605402;uniprotkb:q9nzq7。
64、pd-l2是pd-1的第二配体。pd-l2对pd-1的结合抑制了cd4+ t细胞的t细胞受体(tcr)介导的增殖和细胞因子产生。在低抗原浓度下,pd-l2/pd-1结合抑制了b7-cd28信号。在高抗原浓度下,pd-l2/pd-1结合减少细胞因子的产生。通过干扰素γ治疗,pd-l表达在抗原递呈细胞上有所上调。其表达于某些正常组织和多种肿瘤中。据信pd-l1和pd-l2具有重叠功能并且调节t细胞应答。该蛋白以多个其他名称已知,诸如程序性细胞死亡1配体2;b7树突状细胞分子;程序性死亡配体2;嗜乳脂蛋白b7-dc;pdcd1配体2;pd-1配体2;pdcd1l2;b7-dc;cd273;b7dc;pdl2;pd-1-配体2;cd273抗原;ba574f11.2;以及btdc。pd-l2的外部标识为hgnc:18731;entrez基因:80380;ensembl:ensg00000197646;omim:605723;以及uniprotkb:q9bq51。
65、b7-h3(cd276)表达在各种恶性肿瘤中提高,并且能够在正常和肿瘤衍生的循环内皮细胞之间有所区分(kraan等人british journal of cancer(2014)111,149-156)。受体的刺激指导人髓基质细胞分化为成骨细胞(xu等人2011;immunobiology 216(2011)1311-1317)。蛋白质包含4个人ig样结构域,而小鼠蛋白质似乎具有2个此类结构域。据信该蛋白质是髓系细胞触发受体(triggering receptor expressed on myeloid cells,trem)-样转录物2(tlt-2或tremml2)的首先鉴定出的配体。后一种蛋白质结合b7-h3(4ig-b7-h3)并且共刺激cd8 t细胞的活化(hofmeyer等人2009pnas105;10277-10278)。cd276被广泛表达。其充当t细胞共刺激物。cd276也以多个其他名称已知,诸如cd276分子;共同刺激分子;cd276抗原;b7同系物3;4ig-b7-h3;b7-h3;b7h3;以及b7rp-2。cd276的外部标识为hgnc:19137;entrez基因:80381;ensembl:ensg00000103855;omim:605715;以及uniprotkb:q5zpr3。
66、b7-h4(vtcn1)mrna似乎被广泛表达,但仅少数细胞有效地在膜上表达该蛋白质。b7-h4表达以及结合至活化t细胞经由细胞周期停滞、减少增殖、以及减少il-2产生来抑制t细胞效应子功能。b7-h4在多种人癌症的癌细胞和免疫抑制肿瘤相关巨噬细胞(tam)表面有所上调。通过b7-h4途径的信号转导导致tcr介导的cd4+和cd8+ t细胞增殖、细胞周期进程和il-2产生受到抑制。b7-h4也以多个其他名称已知,诸如含v-set结构域的t细胞活化抑制剂1;免疫共刺激蛋白质b7-h4;t细胞共同刺激分子b7x;b7超家族成员1;b7同系物4;b7h.5;b7h4;t细胞共同刺激分子b7x;b7家族成员h4;蛋白质b7s1;pro1291;vctn1;b7s1;b7x;以及h4 2。b7-h4的外部标识为hgnc:28873;entrez基因:79679;ensembl:ensg00000134258;omim:608162和uniprotkb:q7z7d3。
67、b7-h5(vista)是b7家族的55-65kda成员。其为表达于骨中、胚胎干细胞(esc)上、以及肿瘤细胞表面上的跨膜分子。在肿瘤细胞上,该蛋白质促进mt1-mmp的表达和活性并且充当mt1-mmp的底物。这提高了细胞运动的潜能。该蛋白以多个其他名称已知,诸如染色体10开放阅读框54;含v-set结构域免疫调节受体;t细胞活化的v-结构域ig抑制子;应激诱导分泌蛋白-1;sisp-1;sisp1;应激诱导分泌蛋白1;血小板受体gi24;血小板受体gi24;死亡结构域1α;dd1alpha;b7h5;以及gi24。蛋白质的外部标识为:hgnc:30085;entrez基因:64115;ensembl:ensg00000107738;omim:615608;以及uniprotkb:q9h7m9。
68、b7-h6属于b7家族(参见mim 605402)并且选择性地表达于肿瘤细胞上。b7-h6与nkp30的结合(ncr3;mim 611550)导致自然杀伤(nk)细胞的活化和细胞毒性(brandt等人,2009j exp med.2009年7月6日;206(7):1495-503)。自然杀伤(nk)细胞是参与肿瘤消除的先天性免疫系统的淋巴细胞。b7-h6是结合nkp30(即引发抗肿瘤nk细胞的细胞毒性和细胞因子分泌的人受体)的肿瘤细胞表面分子。b7-h6的其他名称为ncr3lg1;自然杀伤细胞细胞毒性受体3配体1;b7同系物6;b7h6;推定的含ig样结构域蛋白质dkfzp686o24166/dkfzp686i21167;以及dkfzp686o24166。b7-h6的外部标识为hgnc:42400;entrez基因:374383;ensembl:ensg00000188211;omim:613714;以及uniprotkb:q68d85。
69、b7-h7(hhla2)蛋白质在胎盘和肠道上皮的滋养细胞、肾、胆囊、以及乳腺(而非大多数其他器官)中检测到。hhla2蛋白质广泛表达于乳腺、肺、甲状腺、黑素瘤、胰腺、卵巢、肝脏、膀胱、结肠、前列腺、肾和食道的人癌症中。高hhla2表达与局部淋巴结转移和分级相关联(janakiram等人clin cancer res;21(10):2359-66;2015年5月15日)。tmigd2被鉴定为hhla2的受体之一。b7-h7以多个不同的名称已知,诸如herv-h ltr-关联2;人内源性逆转录病毒-h长末端重复序列关联蛋白2;b7h7和b7y。b7-h7的外部标识为hgnc:4905;entrez基因:11148;ensembl:ensg00000114455;omim:604371和uniprotkb:q9um44。
70、程序性细胞死亡1蛋白(pd-1)是属于cd28受体家族的细胞表面受体并且表达于t细胞和前b细胞上。目前已知pd-1结合两种配体-pd-l1和pd-l2。pd-1用作免疫检查点,通过抑制t细胞活化在下调免疫系统中发挥重要作用,这继而降低自体免疫并促进自身耐受性。据信pd-1的抑制作用通过双重机制来实现:促进淋巴结中抗原特异性t细胞的细胞凋亡(程序性细胞死亡),并同时减少调节性t细胞(抑制性t细胞)的细胞凋亡。pd-1也以多个不同的别名已知,诸如pdcd1;程序性细胞死亡1;系统性红斑狼疮易感性2;蛋白质pd-1;hpd-1;pd1;程序性细胞死亡1蛋白质;cd279抗原;cd279;hpd-l;hsle1;sleb2;以及pd-1。pd-1的外部标识为hgnc:8760;entrez基因:5133;ensembl:ensg00000188389;omim:600244;以及uniprotkb:q15116。阻断pd-1活性的新药物类型即pd-1抑制剂激活免疫系统攻击肿瘤,并因此被成功用于治疗某些类型的癌症。
71、clec12a也称为c型凝集素结构域家族12成员a;c型凝集素蛋白cll-1;micl;树突状细胞相关凝集素2;c型凝集素超家族;骨髓抑制型c型类凝集素受体;c型类凝集素分子-1;cll-1;dcal2;cll1;c型类凝集素分子1;dcal-2;杀伤细胞凝集素样受体亚家族l成员1(klrl1);cd371(bakker a.等人cancer res.2004,64,第8843 50页;genbanktm登录号:ay547296;zhang w.等人genbanktm登录号:af247788;a.s.marshall等人j biol chem2004,279,第14792-802页;genbanktm登录号:ay498550;y.han等人blood 2004,104,第2858 66页;h.floyd等人genbanktm登录号:ay426759;c.h.chen等人blood 2006,107,第1459 67页)。标识:hgnc:31713;entrez基因:160364;ensembl:ensg00000172322;omim:612088;uniprotkb:q5qgz9。clec12a是急性髓性白血病(aml)中表达于白血病母细胞和白血病干细胞上的抗原,包括cd34阴性或cd34低表达白血病干细胞(侧群细胞)(a.b.bakker等人cancer res 2004,64,第8443 50页;van rhenen等人2007 blood 110:2659;moshaver等人2008 stem cells 26:3059)。据信clec12a的表达原本受限于造血细胞谱系,尤其是外周血和骨髓内的髓系细胞,即粒细胞、单核细胞和树突状细胞前体。更重要地,clec12a不存在于造血干细胞上。该表达谱使得clec12a为aml中特别有利的靶标。全长形式的clec12a包含275个氨基酸残基,包括一段附加的细胞内10个氨基酸,其不存在于大多数其他同种型中,并且显示出严格地髓样表达谱(表面表达和mrna水平)。术语“clec12a或其功能型等同物”意指保留严格髓样表达谱(处于表面表达水平和mrn水平两者)的以上所提及的所有(诸如剪接和突变)变体及其同种型,如bakker等人cancer res 2004,64,第8443-50页和marshall 2004-j biol chem 279(15),第14792-14802页所述。本发明的clec12a结合抗体结合人clec12a。在本文提及clec12a时,除非另外特别说明,该提及是指人clec12a。
72、“erbb1”或“egfr”是四种受体酪氨酸激酶(rtk)家族的成员,称为her-或cerbb-1、-2、-3和-4。egfr具有由四个亚结构域构成的细胞外结构域(ecd),其中两个参与配体结合,并且其中一个参与同源二聚化和异源二聚化。该部分所用的参考标号是指标题为“说明书引用的参考文献”的列表中参考文献的编号。egfr整合来自各种配体的细胞外信号以产生各种不同的细胞内应答。由egfr激活的主要信号转导途径由ras-分裂素活化蛋白激酶(mapk)有丝分裂信号级联构成。该途径的激活通过将grb2募集到酪氨酸磷酰化的egfr来引发。这导致ras通过grb2结合的ras-鸟嘌呤核苷酸交换因子(sos)激活。此外,pi3-激酶-akt信号转导途径也通过egfr激活,但该激活在存在her3共表达的情况下更强。egfr涉及多种人类上皮恶性肿瘤,值得注意的是乳腺癌、膀胱癌、非小细胞肺癌、肺癌、结肠癌、卵巢癌、头颈癌和脑癌。已经发现激活基因中的突变以及受体及其配体的过表达,产生自分泌激活循环。该rtk因此广泛用作癌症治疗的靶标。已经开发了靶向rtk的小分子抑制剂和针对细胞外配体结合结构域的单克隆抗体(mab)并且迄今为止已经示出多项临床成功,虽然大部分针对一组选择的患者。人egfr蛋白和编码其的基因的数据库登录号是(基因库nm_005228.3)。该登录号主要用于提供鉴定作为靶标的egfr蛋白质的其他方法,由抗体结合的egfr蛋白质的实际序列可有所不同,例如由于编码基因中的突变,例如在一些癌症中发生的那些等。在本文提及egfr时,除非另行指出,否则该提及是指人egfr。结合egfr的抗原结合位点,结合egfr及其多种变体,诸如在一些egfr阳性肿瘤上表达的那些。
73、如本文所用,“erbb-2”或“her2”是指在人类中由erbb-2基因编码的蛋白质。基因或蛋白质的别名包括:cd340;her-2;her-2/neu;mln19;neu;ngl;tkr1。erbb-2基因经常被称为her2(得自人表皮生长因子受体2)。在本文提及erbb-2时,所述提及是指人erbb-2。包含结合erbb-2的抗原结合位点的抗体结合人erbb-2。由于人和其他哺乳动物直系同源物之间的序列和三级结构相似性,erbb-2抗原结合位点也可结合此类直系同源物,但不一定如此。人erbb-2蛋白质和编码它的基因的数据库登录号为(np_001005862.1、np_004439.2、nc_000017.10、nt_010783.15、nc_018928.2)。所述登录号主要用于提供鉴定作为靶标的erbb-2的其他方法,结合抗体的erbb-2蛋白质的实际序列可有所不同,例如由于编码基因中的突变,例如在一些癌症中发生的那些等。erbb-2抗原结合位点结合erbb-2及其多种变体,诸如由一些erbb-2阳性肿瘤细胞表达的变体。
74、如本文所用,“erbb-3”或“her3”是指在人类中由erbb-3基因编码的蛋白质。基因或蛋白质的替代名称为:her3;lccs2;mda-bf-1;c-erbb-3;c-erbb-3;erbb-3-s;p180-erbb-3;p45-serbb-3;以及p85-serbb-3。在本文提及erbb-3时,所述提及是指人erbb-3。包含结合erbb-3的抗原结合位点的抗体结合人erbb-3。由于人和其他哺乳动物直系同源物之间的序列和三级结构相似性,erbb-3抗原结合位点也可结合此类直系同源物,但不一定如此。人erbb-3蛋白质和编码它的基因的数据库登录号为(np_001005915.1、np_001973.2、nc_000012.11、nc_018923.2、nt_029419.12)。所述登录号主要用于提供鉴定作为靶标的erbb-3的其它方法,由抗体结合的erbb-3蛋白质的实际序列可有所不同,例如由于编码基因中的突变,例如在一些癌症中发生的那些等。erbb-3抗原结合位点结合erbb-3及其多种变体,诸如由一些erbb-2阳性肿瘤细胞表达的变体。
75、lgr4为含富含亮氨酸重复序列的g蛋白偶联受体4,基因或蛋白质的别名为:gpr48;g-蛋白偶联受体48;bnmd17;含富含亮氨酸重复序列的g蛋白偶联受体4;含富含亮氨酸重复序列的g蛋白偶联受体4;g-蛋白偶联受体48;
76、结合lgr4的本发明蛋白质或抗体结合人lgr4。由于人和其它哺乳动物直系同源物之间的序列和三级结构相似性,本发明的lgr4结合蛋白质或抗体也结合此类直系同源物,但不一定如此。人lgr4蛋白质和编码其的基因的数据库登录号为(nc_000011.10;nc_018922.2;nt_009237.19;np_060960.2)。所述登录号主要用于提供鉴定作为靶标的lgr4的其它方法,被结合的lgr4蛋白质的实际序列可有所不同,例如由于编码基因中的突变,例如在一些癌症中发生的那些等。lgr4抗原结合位点结合lgr4及其多种变体,诸如由一些lgr4阳性肿瘤细胞表达的变体。
77、lgr5为含富含亮氨酸重复序列的g蛋白偶联受体5,基因或蛋白质的另选的名字为含富含亮氨酸重复序列的g蛋白偶联受体5;含富含亮氨酸重复序列的g蛋白偶联受体5;g-蛋白偶联受体hg38;g-蛋白偶联受体49;g-蛋白偶联受体67;gpr67;gpr49;孤儿g蛋白偶联受体hg38;g-蛋白偶联受体49;gpr49;hg38和fex。
78、结合lgr5的本发明蛋白质或抗体结合人lgr5。由于人和其他哺乳动物直系同源物之间的序列和三级结构相似性,本发明的lgr5结合蛋白质或抗体也结合此类直系同源物,但不一定如此。人lgr5蛋白质和编码它的基因的数据库登录号为(nc_000012.12;nt_029419.13;nc_018923.2;np_001264155.1;np_001264156.1;np_003658.1)。所述登录号主要用于提供鉴定作为靶标的lgr5的其它方法,被结合的lgr5蛋白质的实际序列可有所不同,例如由于编码基因中的突变,例如在一些癌症中发生的那些等。lgr5抗原结合位点结合lgr5及其多种变体,诸如由一些lgr5阳性肿瘤细胞表达的变体。
79、znrf3为锌环状指样蛋白3。基因或蛋白质的另选的名字为锌环状指样蛋白3;锌/环状指样蛋白3;环状指样蛋白203;kiaa1133;rnf203;新型c3hc4型锌指样蛋白(环指样蛋白);e3泛素-蛋白质连接酶znrf3;cta-292e10.6;ec 6.3.2.;和bk747e2.3 3。
80、结合znrf3的本发明蛋白质或抗体结合人znrf3。由于人和其他哺乳动物直系同源物之间的序列和三级结构相似性,本发明的znrf3结合蛋白质或抗体也结合此类直系同源物,但不一定如此。人znrf3蛋白质和编码其的基因的数据库登录号为(nc_000022.11;nt_011520.13;nc_018933.2;np_001193927.1;np_115549.2)。所述登录号主要用于提供鉴定作为靶标的znrf3的其它方法,被结合的znrf3蛋白质的实际序列可有所不同,例如由于编码基因中的突变,例如在一些癌症中发生的那些等。znrf3抗原结合位点结合znrf3及其多种变体,诸如由一些znrf3阳性肿瘤细胞表达的变体。
81、rnf43为环状指样蛋白43。基因或蛋白质的另选的名字为环状指样蛋白43;rnf124;e3泛素-蛋白质连接酶rnf43;环状指样蛋白43;ec6.3.2.;urcc。
82、结合rnf43的本发明蛋白质或抗体结合人rnf43。由于人和其他哺乳动物直系同源物之间的序列和三级结构相似性,本发明的rnf43结合蛋白质或抗体也结合此类直系同源物,但不一定如此。人rnf43蛋白质和编码它的基因的数据库登录号为(nc_000017.11;nt_010783.16;nc_018928.2;np_001292473.1;np_001292474.1;np_060233.3)。所述登录号主要用于提供鉴定作为靶标的rnf43的其它方法,被结合的rnf43蛋白质的实际序列可有所不同,例如由于编码基因中的突变,例如在一些癌症中发生的那些等。nrf43抗原结合位点结合rnf43及其多种变体,诸如由一些rnf43阳性肿瘤细胞表达的那些变体。
83、对序列标识符进行标引以识别哪个蛋白质作为目标。只要抗体的相应可变结构域所要识别的表位未受到影响,结合分子诸如本发明的抗体还识别其至少一些变体,诸如其等位变体、剪接变体和突变变体。一些别名可以或不可也被用来指其他蛋白质。名称只是为了参考的目的给出。结合分子诸如本发明的抗体结合至如表达于细胞上的蛋白质。只要结合分子所结合的表位可用,其还能够结合至蛋白质的变体。因此,只要表位可用,剪接变体或突变变体(如果有的话)也将被结合。结合分子结合至所指定蛋白质的事实意味着其能够作为特性结合至蛋白质,且并非暗示结合分子实际结合至靶标(尽管其可以)。这也不意味着抗体不结合至其他蛋白质。结合分子诸如本发明抗体的此类交叉反应性目前是未知的;然而,不明确地排除的是,可存在此类交叉反应性。
84、本发明公开了结合tnf受体超家族成员(第一膜蛋白)的细胞外部分以及第二膜蛋白的细胞外部分的结合分子。所述第二膜蛋白优选地不为tnf受体超家族的成员。此类结合分子还被进一步称为“本发明的结合分子”。结合分子优选地为结合蛋白。在一个优选的实施方案中,结合分子为抗体或其变体,优选地双特异性抗体或其变体。还提供了包含如本文所述的两种或更多种结合分子的组合物和成套试剂盒。
85、本发明的结合分子优选地为抗体(或其变体,如本技术的其他地方所述)、抗体模拟物、多肽、核酸配体或它们的组合。这些蛋白质或核酸配体通常结合到一个靶标上。本发明的结合分子结合两个或更多个靶标。结合分子优选地结合两个靶标。抗体或双特异性抗体的变体维持该方面。结合蛋白或核酸配体具有靶标所要结合的结合位点(抗原结合位点)。结合蛋白或核酸配体优选地包含两个或更多个结构域,该两个或更多个结构域优选地具有用于靶标(抗原)的结合位点,优选地每个结构域有一个结合位点。此类结构域优选地为抗体可变结构域或其变体。抗体可变结构域已成为许多研究的主题。制备出保留正常可变结构域的结合特异性的类似可变结构域或其部分的多种变体。此类变体的非限制性示例在本文别处有所描述。
86、应当理解这些抗体、抗体模拟物、多肽和核酸配体的任何组合可通过本领域已知的方法连接在一起。例如,在一些实施方案中,本发明的结合分子为缀合物或融合蛋白。就抗体而言,制备多特异性抗体的技术已经进展到还包括双特异性抗体,其具有与正常单特异性抗体相同的总体结构,但其中抗体的两个臂各自结合不同的靶标。
87、抗体模拟物为多肽,与抗体相同,其可特异性结合抗原,但在结构上与抗体不相关。抗体模拟物通常为人工肽或蛋白质,摩尔质量为约3kda至20kda。优于抗体的共同优点是更好的溶解度、组织渗透、针对热和酶的稳定性、以及相对低生产成本。抗体模拟物的非限制性示例为亲和分子(通常基于蛋白质a的z结构域);亲和素(通常基于γ-b结晶或泛素);affimer(通常基于半胱氨酸蛋白酶抑制剂);affitin(通常基于来自嗜酸热硫化叶菌(sulfolobus acidocaldarius)的sac7d);α体(通常基于三重螺旋卷曲螺旋);anticalin(通常基于脂质运载蛋白);avimer(通常基于各种膜受体的a结构域)darpin(通常基于锚蛋白重复基序);fynomer(通常基于fyn 7的sh3结构域);kunitz结构域肽(通常基于各种蛋白酶抑制剂的kunitz结构域);和单体(通常基于纤粘蛋白的iii型结构域)。
88、单体是合成的结合蛋白,其使用纤连蛋白iii型结构域(fn3)作为分子支架来构造。单体是用于形成靶向结合蛋白质的抗体的简单且稳健的替代形式。术语“单体”由koide集团于1998年创造出来,其发表了使用人纤粘蛋白的第十fn3结构域证明单体概念的第一篇论文。
89、单体和其它抗体模拟物通常由组合库形成,其中使用分子展示和定向进化技术(例如噬菌体展示、mrna展示和酵母表面展示)使支架的部分多样化。大量抗体模拟物具有对其相应靶标的高亲和力和高特异性。
90、核酸配体是与特定靶分子结合的寡核苷酸或肽分子。核酸配体通常通过从大随机序列库中选择它们而形成,但天然的核酸配体也存在于核糖开关中。核酸配体可以大分子形式用于基础研究和临床目的。
91、如本文所用,术语“缀合物”是指已经任选地通过连接区共价接合的两个或更多个分子。例如,在一些实施方案中,缀合物是通过连接区连接至第二蛋白质或非蛋白质部分的第一蛋白质或非蛋白质部分。例如,在本发明结合分子的一些实施方案中,其包括或由两种或更多种已经共价接合的抗体组成。缀合物不限于第一部分和第二部分,但在一些实施方案中,也可具有通过其它连接区接合的第三、第四或更多部分。如本专利申请的其它地方所述,蛋白质部分的示例包括但不限于:多肽、拟肽或抗体(或如本专利申请的其它地方所述的抗体部分、衍生物或类似物)。非蛋白质部分的示例包括但不限于核酸配体。可使用多种类型的连接子,并且根据缀合物中的分子类型和连接子的期望特性(长度、柔性、对蛋白酶活性的抗性和其它类似特性),连接子将被选择为合适的。此类连接子可包括核苷酸、多肽、或适宜的合成材料。例如,连接子可以为柔性肽连接子。在某些实施方案中,连接子可以为可裂解连接子,其使得缀合物的部分彼此分离。在其它实施方案中,肽连接子可以为螺旋连接子。用于连接蛋白质和其他分子的各种示例和试剂盒在本领域中为人们所知。如本文所用,术语“融合蛋白”是指包括两个或更多个多肽的蛋白质,或者通过重组在dna水平上连接并且作为单个多肽一起表达的蛋白质。融合蛋白还可包括肽连接区,其也由dna编码并且与融合蛋白一起表达。作为融合蛋白的一部分的肽连接子可被设计成具有特定特性,诸如柔性、亲水性、蛋白酶抗性、可裂解性等。所有这些性质均可设计在dna序列中,并且设计连接子的方法在本领域中为人们所熟知。例如,抗体可通过本领域中熟知的方法连接在一起,并且如本文所述,形成双特异性抗体或多靶向抗体。另外,双特异性抗体可通过本领域已知的各种方法构造,例如通过使用诸如的技术(参见例如wo 2013/157954)。双特异性单克隆抗体(bsmab、bsab)通常包括两种不同单克隆抗体的结合结构域,并且因此结合两个不同的表位。分子,而且另外其他全长igg双特异性抗体具有两种不同的抗原结合特异性,所述结合特异性由scfv的fab的全长igg分子的两个不同可变区编码。可通过用编码两个不同的共同轻链(clc)抗体的基因构建体共转染个体细胞来产生,如本文其他地方详述的。ch3工程化确保有效的异源二聚反应和基本上纯的特异性抗体的形成。
92、本发明的结合分子优选地为抗体或其变体。本发明的结合分子优选地为双特异性抗体或其变体。
93、抗体通常通过所谓的抗原结合位点结合其靶标。未修饰的抗原结合位点通常由抗体的可变结构域形成并存在于抗体的可变结构域中。可变结构域包含所述抗原结合位点。结合抗原的可变结构域为包含结合抗原的抗原结合位点的可变结构域。
94、在一个实施方案中,抗体可变结构域包含重链可变区(vh)和轻链可变区(vl)。抗原结合位点可以存在于组合的vh/vl可变结构域中,或仅存在于vh区域中或仅存在于vl区域中。当抗原结合位点存在于可变结构域的两个区域中的一个中时,对应的可变区可有助于结合可变区的折叠和/或稳定性,但不显著有利于抗原本身的结合。
95、如本文所用,抗原结合是指抗体与其抗原的典型结合能力。抗体与抗原的结合可以各种方式评估。一种方式是将抗体与抗原(优选地表达抗原的细胞)一起温育,去除未结合的抗体(优选通过洗涤步骤),并使用结合至经结合抗体的标记抗体来检测经结合的抗体。
96、通过抗体结合抗原通常通过抗体的互补决定区(cdr)来介导,并且抗原和可变结构域两者的特异性三维结构允许这两种结构精确地结合在一起(类似于锁和钥匙的相互作用),与蛋白质的随机、非特异性粘附相反。因为抗体通常识别被称为抗原表位的抗原部分,并且因为此类表位也可存在于其它化合物中,所以如果此类其它化合物含有相同的表位,则根据本发明的抗体也可识别其它蛋白质。因此,术语“结合”不排除抗体与另一种或多种包含相同表位的蛋白质的结合。此类一种或多种其它蛋白质优选不是人蛋白质。
97、本发明的蛋白质诸如抗体通常不与除了出生后,优选地成人细胞膜上的特异性靶蛋白之外的其它蛋白质结合。
98、如本文所用,术语“抗体”是指蛋白质分子,优选地属于蛋白质的免疫球蛋白类别,其包含结合抗原上的表位的一个或多个可变结构域,其中此类结构域衍生自抗体的可变结构域或共享与抗体的可变结构域的序列同源性。用于治疗应用的抗体优选尽可能地接近待治疗的受试者的天然抗体(例如,对于人受试者而言,人抗体)。可根据特异性和亲和性来表达抗体结合。特异性决定哪个抗原或其表位被结合结构域特异性结合。亲和性是对特定抗原或表位的结合强度的量度。优选地,根据本发明的抗体的单独臂的亲和力在纳摩尔范围内。抗体诸如本发明的双特异性抗体通常包含天然抗体的恒定结构域(fc部分),其可如本文别处所述的那样工程化成例如使adcc和/或cdc活性下降。本发明的抗体通常是双特异性全长抗体,优选地人igg亚类。
99、术语“可变结构域”、“vh/vl对”、“vh/vl”在本文互换使用。可变结构域由重链的可变区和轻链的可变区构成。重链的可变区通常由重排vdj区形成。轻链的可变区通常由重排vj区形成。vdj/vj区目前也可使用例如功能性抗体可用的大量序列信息人工产生。
100、在一些实施方案中,根据本发明的结合分子或抗体或变体包含能够结合tnf受体超家族的成员的细胞外部分的抗原结合位点以及能够结合b7家族的成员的一个抗原结合位点。在一些实施方案中,根据本发明的结合分子或抗体或变体包含能够结合cd137的细胞外部分的抗原结合位点以及能够结合b7家族的成员的抗原结合位点。在一些实施方案中,根据本发明的结合分子或抗体或变体包含能够结合cd137的抗原结合位点以及能够结合pd-l1的抗原结合位点。
101、在一些实施方案中,根据本发明的结合分子或抗体或变体具有不超过两个抗原结合位点。这意味着此类结合分子或抗体或变体的抗原结合部分由两个抗原结合位点组成,而不存在附加的抗原结合位点。两个抗原结合位点各自优选地包含免疫球蛋白vh/vl对。优选地,本发明的结合分子或抗体或变体的抗原结合部由能够结合tnf受体超家族的成员的细胞外部分的一个免疫球蛋白可变结构域以及能够结合第二膜蛋白的一个免疫球蛋白可变结构域组成。特定优选的实施方案是具有igg形式的免疫球蛋白,所提供的优点是根据本发明的二价结合分子/抗体/变体的半衰期通常相比于多价化合物较长。此外,相比于多价化合物,根据本发明的二价结合分子的免疫原性通常较低。根据这些实施方案的分子/抗体/变体优选地保留天然igg的结构,并因此维持与天然igg的该种结构相关的所有益处。
102、如本文所用,术语“多价”涵盖三种或更多种特异性,其例如存在于三价结合分子和四价结合分子中。
103、一些实施方案提供根据本发明的结合分子或抗体或变体,其中所述结合分子或抗体或变体的抗原结合位点由能够结合tnf受体超家族的成员的细胞外部分的一个抗原结合位点以及能够结合b7家族的成员的一个抗原结合位点组成。在一些实施方案中,根据本发明的所述结合分子或抗体或变体的抗原结合位点由能够结合cd137的细胞外部分的一个抗原结合位点以及能够结合b7家族的成员的一个抗原结合位点组成。在一些实施方案中,根据本发明的所述结合分子或抗体或变体的抗原结合位点由能够结合cd137的一个抗原结合位点以及能够结合pd-l1的一个抗原结合位点组成。
104、如本文所用,术语“抗原结合位点”意指特异性结合抗原表位的结合分子或抗体的位点。此类抗原结合位点优选地源于抗体的可变结构域(特别地其cdr区)或与之共享序列同源性。在一些优选的实施方案中,所述抗原结合位点是免疫球蛋白vh/vl对所形成的免疫球蛋白可变结构域。在其他实施方案中,所述抗原结合位点来源于抗体模拟物,例如,前文所述的亲和分子、亲和素、affimer、affitin、α体、抗运载蛋白(anticalin)、avimer、darpin、fynomer、kunitz结构域肽或单体。
105、本发明的抗体优选为“全长”抗体。根据本发明的术语“全长”被定义为包括基本完全抗体,而不具有尺寸大于20个氨基酸残基的一个或多个人工加入的部分,例如附加抗原结合位点或附加活性位点或附加配体或附加配体结合部分。然而,全长抗体并非必须具有完整抗体的所有功能。为避免疑义,全长抗体包含两条重链和两条轻链。每条链包含恒定(c)区域和可变(v)区域,其可分解为命名为ch1、ch2、ch3和vh的重链结构域和命名为cl、vl的轻链结构域。重链的结构域优选地以天然抗体的顺序存在(vh-ch1-ch2-ch3;意指vh结构域与ch1结构域相邻,继之以ch2结构域,随后接着是ch3结构域)。轻链的结构域也优选地以天然抗体的顺序存在(vl-cl;意指vl结构域与cl结构域相邻)。抗体经由包含在fab片段部分中的可变结构域结合到抗原。抗体可通过恒定结构域,主要通过fc部分与免疫系统的分子和细胞相互作用。在一些实施方案中,本发明的抗体为igg,优选地全长igg。全长igg抗体是优选的,因为它们通常具有有利的半衰期并且由于免疫原性的原因期望保持接近全自体同源的(人)分子。在一些实施方案中,本发明的抗体为全长igg1、全长igg2、全长igg3或全长igg4抗体。
106、根据本发明的全长抗体包括其中可存在突变的抗体,所述突变提供所需特性或仅是原始链中的突变的替代物。此类突变应当不是任何区域的大部分的缺失。然而,其中一个或多个氨基酸残基插入、缺失、置换或它们的组合,但基本上不改变所得抗体的抗原结合特性的抗体涵盖在术语“全长抗体”内。例如,igg抗体在恒定区中可具有1-20个氨基酸残基插入、置换、缺失或它们的组合。
107、本发明的抗体或其功能部分、衍生物和/或类似物优选地为双特异性抗体或其功能部分、衍生物和/或类似物。在一个优选的实施方案中,双特异性igg抗体的效应子功能下降。在一个优选的实施方案中,本发明的抗体为双特异性全长抗体。本发明的抗体优选地为双特异性全长igg抗体,优选地在ch2/下铰链区中突变以降低效应子功能。ch2/下铰链区内突变以降低效应子功能的igg1基于其在人体内较长的循环半衰期是有利的。为了防止人类的任何免疫原性,优选根据本发明的双特异性抗体是人抗体。
108、术语“双特异性”(bs)意指抗体的一部分(如上所定义)结合抗原上的一个表位,而第二部分结合相同抗原或不同抗原上的不同表位。不同的表位通常存在于不同的抗原上。然而,不同的表位也可以存在于相同的抗原上。根据本发明,所述第一抗原和第二抗原实际上是两种不同的蛋白质。优选的双特异性抗体是包含两种不同单克隆抗体的部分并因此可以结合至两种不同的表位,优选两种不同抗原的抗体。取决于由双特异性抗体识别的两种抗原的表达水平,(亚)细胞定位和化学计量,抗体的两个fab臂可以或不可以同时结合其表位。双特异性抗体的一个臂通常包含一个抗体的可变结构域,并且另一个臂包含另一个抗体的可变结构域(即,双特异性抗体的一个臂由与一个轻链配对的一个重链形成,然而其它臂由与轻链配对的不同重链形成)。本发明的双特异性抗体的重链可变区通常彼此不同,然而轻链可变区优选在本发明的双特异性抗体中相同。其中不同重链可变区与相同或共同轻链可变区缔合的双特异性抗体也被称为具有共同轻链可变区(clcv)的双特异性抗体。优选的是轻链恒定区也是如此。此类双特异性抗体被称为具有共同轻链(clc)。因此,本发明还提供根据本发明的双特异性抗体,其中两个臂均包含共同的轻链。
109、如本文所述的双特异性抗体优选地包含共同轻链可变结构域,优选地共同轻链。根据本发明的术语“共同轻链”是指可以一致的或具有一些氨基酸序列差异然而全长抗体的结合特异性不受影响的轻链。例如,在如本文所用的共同轻链的定义范围内可能的是,例如通过引入和测试保守性氨基酸改变,制备或找到不一致但仍然在功能上等同的轻链,当与重链配对时区域中氨基酸改变不有助于或仅部分有助于结合特异性等。在添加或不添加术语“重排”的情况下,术语“共同轻链”、“共同lc”、“clc”、“单个轻链”均可在本文中互换使用。在添加或不添加术语“重排”的情况下,术语“共同轻链可变区”、“共同vl”、“共同lcv”、“clcv”、“单个vl”均可在本文中互换使用。本发明的优选方面是双特异性抗体具有能够与至少两个,并优选地多个不同结合特异性的重链(可变区)组合的共同轻链(可变区),以形成功能性抗原结合结构域的抗体(wo2004/009618,wo2009/157771)。共同轻链(可变区)优选地为人轻链(可变区)。共同轻链(可变区)优选地具有种系序列。优选的种系序列是经常用于人类库中的轻链可变区,并且具有良好的热力学稳定性、产量和溶解度。优选的种系轻链为o12。共同轻链优选地为重排种系人k轻链igvκ1-39*01/igjκ1*01(图1a)。共同轻链可变区优选地为重排种系人k轻链igvκ1-39*01/igjκ1*01的可变区。共同轻链优选地包含具有0-5个氨基酸插入、缺失、取代、添加或它们的组合的如图1b或1d所示的轻链可变区。共同轻链优选地还包括轻链恒定区,优选地k轻链恒定区。编码共同轻链的核酸可被密码子优化成用于用来表达共同轻链蛋白质的细胞体系。编码核酸可偏离种系核酸序列。
110、在一个优选的实施方案中,轻链包含含有如图1a所示的o12/igvκ1-39*01基因片段的氨基酸序列的轻链区,该氨基酸序列具有0-10个,优选地0-5个氨基酸插入、缺失、取代、添加或它们的组合。短语“o12轻链”将在整个说明书中用作“包含含有如图1a所示的o12/igvκ1-39*01基因片段的氨基酸序列的轻链可变区的轻链,该氨基酸序列具有0-10个,优选地0-5个氨基酸插入、缺失、取代、添加或它们的组合”的简称。igvκ1-39是免疫球蛋白可变k 1-39基因的简称。该基因也称为免疫球蛋白k可变1-39;igkv139;igkv1-39;o12a或o12。基因的外部标识为hgnc:5740;entrez基因:28930;ensembl:ensg00000242371。igvκ1-39的优选氨基酸序列在图1e中给出。这列出了v区的序列。v区可与五个j区之一组合。图1b和1d描述了与j区组合的igvκ1-39的两个优选的序列。所连接的序列示为igkv1-39/jk1和igkv1-39/jk5;别名为igvκ1-39*01/igjκ1*01或igvκ1-39*01/igjκ5*01(根据imgt.org处的imgt数据库全球网站命名)。
111、优选的是包含轻链可变区的o12/igvκ1-39*01为种系序列。还优选的是包含轻链可变区的igjκ1*01或/igjκ5*01为种系序列。在一个优选的实施方案中,igkv1-39/jk1或igkv1-39/jk5轻链可变区为种系序列。
112、在一个优选的实施方案中,轻链可变区包含种系o12/igvκ1-39*01。在一个优选的实施方案中,轻链可变区包含k轻链igvκ1-39*01/igjκ1*01或igvκ1-39*01/igjκ5*01。在一个优选的实施方案中包含igvκ1-39*01/igjκ1*01。轻链可变区优选地包含种系k轻链igvκ1-39*01/igjκ1*01或种系k轻链igvκ1-39*01/igjκ5*01,优选地种系igvκ1-39*01/igjκ1*01。
113、产生具有o12轻链的抗体的成熟b-细胞通常产生相对于种系序列(即生物体的非淋巴细胞中的正常序列)发生一个或多个突变的轻链。造成这些突变的过程通常称为体细胞(超变)突变。所得的轻链称为亲和力成熟的轻链。此类轻链衍生自o12种系序列时为o12-衍生的轻链。在本说明书中,短语“o12轻链”将包括o12-衍生的轻链。经体细胞超变引入的突变当然也可以在实验室人工引入。在实验室也可引入其他突变而不影响轻链在性质上但并不一定在数量上的特性。如果轻链包含具有0-10个、优选地0-5个氨基酸插入、缺失、取代、添加或它们的组合的如图1a、图1b;图1d或图1e所示的序列,轻链至少为o12轻链。在一个优选的实施方案中,o12轻链是包含具有0-9个、0-8个、0-7个、0-6个、0-5个、0-4个氨基酸插入、缺失、取代、添加或它们的组合的如图1a;图1b;图1d或图1e所示的序列的轻链。在一个优选的实施方案中,o12轻链是包含具有0-5个,优选地0-4个,更优选地0-3个氨基酸插入、缺失、取代、添加或它们的组合的如图1a、图1b;图1d或图1e所示的序列的轻链。在一个优选的实施方案中,o12轻链是包含具有0-2个,更优选地0-1个,最优选地0个氨基酸插入、缺失、取代、添加或它们的组合的如图1a、图1b;图1d或图1e所示的序列的轻链。在一个优选的实施方案中,o12轻链是包含具有所提及氨基酸插入、缺失、取代、添加或它们的组合的如图1a或图1b所示的序列的轻链。在一个优选的实施方案中,轻链包含图1a的序列。在一个优选的实施方案中,轻链可变区包含图1b的序列。
114、共同轻链(可变区)可为λ轻链,这因此也提供于本发明的上下文中,然而,k轻链是优选的。本发明的共同轻链的恒定部分可为k或λ轻链的恒定区。其优选地为k轻链的恒定区,优选地其中所述共同轻链为种系轻链,优选地包含igvkl-39基因片段的重排种系人k轻链,最优选地重排种系人k轻链igvkl-39*01/igjkl*01(图1)。术语重排的种系人κ轻链igvκ1-39*01/igjκ1*01、igkv1-39/igkj1、huvκ1-39轻链或简称huvκ1-39、或简称1-39在整个申请中可互换使用。显然,本领域技术人员将认识到“共同”还是指氨基酸序列不一致的轻链的功能性等同物。存在所述轻链的许多变体,其中存在不影响功能性结合区形成的突变(缺失、取代、添加)。
115、igvκ1-39是免疫球蛋白可变k 1-39基因的简称。该基因也称为免疫球蛋白k可变1-39;igkv139;igkv1-39;o12a或o12。基因的外部标识为hgnc:5740;entrez基因:28930;ensembl:ensg00000242371。igvκ1-39的优选氨基酸序列在图1中给出。这列出了v区的序列。v区可与五个j区之一组合。图1描述了与j区组合的igvκ1-39的两个优选的序列。所连接的序列示为igkv1-39/jk1和igkv1-39/jk5;别名为igvκ1-39*01/igjκ1*01或igvκ1-39*01/igjκ5*01(命名根据imgt数据库万维网,imgt.org)。
116、共同轻链可变区优选地连接至k轻链恒定区。在一个优选的实施方案中,轻链包含k轻链igvκ1-39*01/igjκ1*01或igvκ1-39*01/igjκ5*01。在一个优选的实施方案中包含igvκ1-39*01/igjκ1*01。
117、产生共同轻链的细胞能够产生例如重排种系人k轻链igvκ1-39*01/igjκ1*01和包含融合于λ恒定区的所提及轻链的可变区的轻链。
118、如本文所述的双特异性抗体或其变体优选地具有结合tnf受体超家族的成员的细胞外部分的一个重链可变区/轻链可变区(vh/vl)组合,以及结合第二膜蛋白的细胞外部分的第二vh/vl组合,其中所述第二膜蛋白不为tnf受体超家族的成员。
119、如本文所述的双特异性抗体或其变体优选地具有结合tnf受体超家族的成员的细胞外部分的一个重链可变区/轻链可变区(vh/vl)组合,以及结合b7家族的成员的细胞外部分的第二vh/vl组合。如本文所述,这样提供的优点在于,所期望的免疫应答能够被特别好地促进,因为b7家族成员向淋巴细胞递送“共刺激”或“共抑制”信号,从而增强或减弱免疫应答。因此,通过靶向b7家族的成员,可以增强刺激信号和/或抵消抑制信号,从而诱导或增强例如针对异常细胞的期望免疫应答。
120、在一个优选的实施方案中,所述第一vh/vl组合中的vl类似于所述第二vh/vl组合中的vl。在一个更优选的实施方案中,第一vh/vl组合和第二vh/vl组合中的vl是相同的。在一个优选的实施方案中,双特异性抗体是具有结合tnf受体超家族的成员的细胞外部分的一个重/轻(h/l)链组合以及结合b7家族成员的细胞外部分的一个h/l链组合的全长抗体。在一个优选的实施方案中,所述第一h/l链组合中的轻链类似于所述第二h/l链组合中的轻链。在一个更优选的实施方案中,第一h/l链组合和第二h/l链组合中的轻链是相同的。
121、已经公开了几种方法以利于产生双特异性抗体,或者相反单特异性抗体。在本发明中,优选的是相比于产生相应的单特异性抗体,细胞有利于产生双特异性抗体。这通常通过修饰重链的恒定区来实现,使得其相比于同源二聚化而有利于异源二聚化(即与其他重链/轻链组合的重链二聚体化)。在一个优选的实施方案中,本发明的双特异性抗体包含具有相容性异源二聚化结构域的两个不同免疫球蛋白重链。各种相容性异源二聚化结构域已在本领域中有所描述。相容性异源二聚化结构域优选地为相容性免疫球蛋白重链ch3异源二聚化结构域。当使用野生型ch3结构域时,两种不同重链(a和b)和共同轻链的共表达将产生三种不同的抗体物质aa、ab和bb。aa和bb是两种单特异性二价抗体的命名,ab是双特异性抗体的命名。为增加期望的双特异性产物(ab)的百分比,可以使用ch3工程化,或换句话讲,可使用具有相容的异源二聚化结构域的重链,如下文所定义的。现有技术描述了其中可实现重链的此类异源二聚化的各种方式。一种方式是产生“杵臼结构”双特异性抗体。参见美国专利申请20030078385(arathoon等人)。
122、如本文所用,术语“相容性异源二聚化结构域”是指被工程化使得工程化结构域a'将优先与工程化结构域b'形成异源二聚体,反之亦然,a'-a'与b’-b’之间的同源二聚化减少的蛋白质结构域。
123、在us13/866,747(现公开为us 9,248,181),us14/081,848(现公开为us 9,358,286)和pct/nl2013/050294(公布为wo2013/157954)中;(以引用方式并入本文),公开了使用相容性异源二聚化结构域产生双特异性抗体的方法和装置。这些装置和方法还可有利地用于本发明。具体地,本发明的双特异性抗体优选地包含突变,以基本上仅产生双特异性全长igg分子。优选的突变是第一ch3结构域(“kk-变体”重链)中的氨基酸取代l351k和t366k(eu编号)以及第二结构域(“de-变体”重链)中的氨基酸取代l351d和l368e,反之亦然。先前已在我们的us 9,248,181和us9,358,286专利以及wo2013/157954pct专利申请中示出,de-变体和kk-变体优先配对形成异源二聚体(所谓的“dekk”双特异性分子)。几乎不进行de-变体重链(dede同源二聚体)的同源二聚化,这是因为相同重链之间的ch3-ch3界面中带电残基之间的相斥。
124、可通过编码一个轻链和两个不同重链的一种或多种质粒的(瞬时)转染来生成双特异性抗体,所述重链被ch3工程化成确保有效异源二聚化并形成双特异性抗体。这些链在单细胞中的产生导致相比于形成单特异性抗体而言有利于形成双特异性抗体。基本上仅产生双特异性全长igg1分子的优选突变是在351位和366位处的氨基酸取代,例如,在第一ch3结构域(“kk-变体”重链)中的l351k和t366k(根据eu编号法进行编号)和在351位和368位处的氨基酸取代,例如第二ch3结构域(“de-变体”重链)中的l351d和l368e,反之亦然。
125、在一个实施方案中,包含结合cd137的可变结构域的重链/轻链组合包含重链的de变体。在该实施方案中,包含能够结合除cd137之外的抗原的可变结构域的重链/轻链组合包含重链的kk变体。将认识到,本发明的实施方案也可包括结合cd137的可变结构域,并且包括重链的kk变体,以及本领域的技术人员已知用于促进与能够结合除cd137之外的抗原的可变结构域异源二聚体化的其他变型。
126、fc区介导抗体的效应子功能,诸如补体依赖性细胞毒性(cdc)、抗体依赖性细胞毒性(adcc)以及抗体依赖性细胞吞噬作用(adcp)。根椐治疗性抗体或fc融合蛋白应用,可能期望减小或增大效应子功能。减小的效应子功能在本发明中是优选的。当免疫应答如本发明的一些实施方案中的那样被活化、增强或刺激时,可能期望减小的效应子功能。效应子功能减小的抗体能够被用于靶向免疫细胞的细胞表面分子等等。
127、发现igg与fcγrs或c1q结合需要位于铰链区和ch2结构域内的残基。ch2结构域(图2d)的两个区与fcγrs和c1q结合相关。第233-236位处的igg2残基以及第327、330和331位处的igg4残基置换成人igg1显示极大地降低了adcc和cdc(armour等人,1999.eur jimmunol.29(8):2613-24;shields等人,2001。j biol chem.276(9):6591-604)。此外,idusogie等人表明,不同位置处的丙氨酸取代(包括k322)使补体活化显著下降(idusogie等人,2000.j immunol.164(8):4178-84。
128、由于它们减小的效应子功能,igg4抗体代表用于受体阻断而不清除细胞的igg亚型。igg4分子可在动态过程中交换半分子,称为fab臂交换。该现象可在治疗性抗体与内源性igg4之间发生。s228p突变是确保fab臂交换能力下降的突变的示例(labrijn等人,2009.nat biotechnol.27(8):767-71)。
129、效应子功能减小的抗体优选地为包含经修饰ch2/下铰链区的igg抗体,例如从而降低fc受体相互作用或降低c1q结合。在一些实施方案中,本发明的抗体是具有突变ch2和/或下铰链结构域的igg抗体,使得双特异性igg抗体与fcγ受体的相互作用降低。包含突变ch2区的抗体优选地为igg1抗体。此类突变igg1ch2和/或下铰链结构域优选地包含第235和/或236位(eu编号)的氨基取代,优选地l235g和/或g236r取代(图2e)。
130、如本文所述的抗体或双特异性抗体的变体包含抗体或双特异性抗体的功能部分、衍生物和/或类似物。变体保留(双特异性)抗体的结合特异性。功能部分、衍生物和/或类似物保留(双特异性)抗体的结合特异性。结合特异性由结合如本文所述的第一膜蛋白和第二膜蛋白的细胞外部分的能力来定义。
131、抗体的功能部分,或优选地如本文所述的双特异性抗体的功能部分是含有结合tnf受体超家族的成员的细胞外部分的可变结构域以及结合所述第二膜蛋白的细胞外部分的可变结构域的部分。合适的部分例如为f(ab’)2片段,如用胃蛋白酶消化双特异性抗体所产生。包含所述可变结构域的其他部分包括在本发明中。
132、抗体的功能性衍生物,或优选地如本文所述的双特异性抗体的功能性衍生物是包含经由接头连接的结合tnf受体超家族的成员的细胞外部分的可变结构域以及结合所述第二膜蛋白的细胞外部分的可变结构域的蛋白质。可变结构域可为这样的可变结构域、或fab片段、或可变结构域样分子,诸如包含经由接头连接在一起的vh和vl的单链fv片段。可变结构域样分子的其他示例为所谓的单结构域抗体片段。单结构域抗体片段(sdab)是具有单个单体可变抗体区的抗体片段。如同整个抗体,其能够选择性结合特异性抗原。在分子量仅12kda-15kda的情况下,单结构域抗体片段比由两条重蛋白链和两条轻链构成的常见抗体(150kda-160kda)小得多,并且甚至小于fab片段(~50kda,一条轻链和半条重链)和单链可变片段(~25kda,两个可变区,一个来自轻链且另一个来自重链)。单结构域抗体本身并不比正常抗体(通常为90kda-100kda)小得多。单结构域抗体片段大部分由存在于骆驼科中的重链抗体工程化;这些被称为vhh片段某些鱼类也具有仅重链的抗体(ignar,“免疫球蛋白新抗原受体”),由此可获得被称为vnar片段的单结构域抗体片段。另一种方法是使来自人类或小鼠的常见免疫球蛋白g(igg)的二聚体可变结构域分裂成单体。尽管目前大部分对单结构域抗体的研究均基于重链可变结构域,但是来源于轻链的纳米抗体也示出与靶表位特异性结合。可变结构域样分子的其他非限制性示例是vhh、人结构域抗体(dab)和unibodies。优选的功能部分是包含含有重链可变区和轻链可变区的可变结构域的部分。此类可变结构域的非限制性示例是f(ab)-片段和单链fv片段。可变结构域(-样)键的双特异性形式例如为结合至两个不同scfv的人血清白蛋白(hsa);包含经由二聚化基序或自身结合的二级结构诸如螺旋束或卷曲螺旋结合在一起的两个不同scfv的双特异性小抗体,以引起scfv片段的二聚作用(morrison(2007)nat.biotechnol 25:1233-34)。合适的hsa接头和使scfv偶联于接头的方法的示例描述于wo2009/126920。
133、抗体或其功能部分、衍生物和/或类似物,或优选地本发明的双特异性抗体或其功能部分、衍生物和/或类似物优选地用于人。为此,本发明的抗体或其功能部分、衍生物和/或类似物优选是人或人源化抗体。人类对多肽的耐受性受许多不同方面的支配。免疫力,无论是t细胞介导的、b细胞介导的还是其它介导的均是涵盖于人类对多肽的耐受性中的变量之一。本发明的双特异性抗体的恒定区优选地包含:优选地含有如图2所示序列的人重链恒定区;以及优选地含有如图1c所示序列的人轻链恒定区。恒定区可包含与天然存在的人抗体的恒定区的一个或多个,优选不超过10个,优选不超过5个氨基酸差异。优选恒定部分完全来源于天然存在的人抗体。本文产生的各种抗体来源于用相应靶免疫的普通轻链小鼠,如wo2009/157771中所述。本文产生的各种抗体来源于人抗体可变结构域库。因此,这些可变结构域是人可变结构域。独特的cdr区域可来源于人,是合成的或来源于另一种生物体。当其除了cdr区域之外,具有与天然存在的人抗体可变区的氨基酸序列一致的氨基酸序列时,该可变区至少为人可变区。在此类实施方案中,结合tnf受体超家族的成员或b7家族的膜缔合成员的抗体的可变区的vh,或本发明抗体中的轻链可包含与天然存在的人抗体的可变区的一个或多个,优选不超过10个,优选不超过5个氨基酸差异,不计数cdr区的氨基酸序列的可能差异。在体细胞超突变的情况下,此类突变也天然存在。
134、至少就重链可变区而言,抗体可来源于各种动物物种。其是人源化诸如鼠重链可变区的常见操作。存在可实现这种人源化的各种方式,其中存在嫁接到人重链可变区中的cdr,其具有匹配小鼠重链可变区的3d结构的3d结构;将小鼠重链可变区去免疫,优选地通过从小鼠重链可变区去除已知或可疑的t细胞或b细胞表位进行。该去除通常是通过将表位中的一个或多个氨基酸取代另一(通常保守性)氨基酸,使得表位的序列被修饰,使得其不再是t-细胞或b-细胞表位。
135、去免疫的鼠重链可变区比初始鼠重链可变区在人类中具有更少的免疫原性。优选地,本发明的可变区或结构域进一步人源化,例如镶饰。通过使用镶饰技术,容易被免疫系统遇到的外部残基选择性地用人残基取代,以提供包含弱免疫原性或基本上非免疫原性的镶面表面的杂交分子。如用于本发明的动物优选哺乳动物,更优选灵长类,最优选地人。
136、根据本发明的抗体或双特异性抗体或其功能部分、衍生物和/或类似物优选地包含人抗体的恒定区。根据其重链恒定结构域的差异,将抗体分组成五类或同种型:igg、iga、igm、igd和ige。这些类或同种型包含所述重链中的至少一个,其用相应的希腊字母命名。在一个优选的实施方案中,本发明提供了根据本发明的抗体,其中所述恒定区选自igg恒定区,即选自igg1、igg2、igg3和igg4。优选地所述恒定区为igg4或igg1恒定区(图2),更优选地突变igg1恒定区。igg1恒定区中的一些变化在自然界中发生和/或在不改变所得抗体的免疫学特性的情况下获得。通常在恒定区中获得介于约1-10个氨基酸插入、缺失、取代或它们的组合。恒定区可如本文所指出的那样突变成允许有效的异源二聚化,或者减小效应子功能或出于其他原因包括半衰期、稳定性等等。
137、合理的方法已朝向使人类环境下的非人类残基含量最小化演变。各种方法可用于成功地将抗体的抗原结合特性嫁接到另一种抗体上。抗体的结合特性可主要位于cdr3区的适当序列中,通常由可变结构域中的cdr1区和cdr2区的序列与可变结构域的适当结构的组合作为整体支持。目前各种方法适用于将cdr区嫁接到另一抗体的适宜可变结构域上。这些方法中的一些在j.c.almagro1和j.fransson(2008)frontiers in bioscience 13,1619-1633中综述,其以引用方式包括在本文中。
138、包含如图3所示的可变重链序列的可变结构域的轻链可变区优选为o12或基于o12的种系轻链,优选为重排的种系人κ轻链igvκ1-39*01/igjκ1*01或其片段或功能性衍生物(根据imgt.org处的imgt数据库全球网站命名)。使用术语重排的种系人κ轻链igvκ1-39*01/igjκ1*01、igkv1-39/igkj1、huvκ1-39轻链或简称huvκ1-39。轻链可具有1个、2个、3个、4个或5个氨基酸插入、缺失、取代或它们的组合。提及的1个、2个、3个、4个或5个氨基酸取代优选为保守氨基酸取代,所述插入、缺失、取代或它们的组合优选不在vl链的cdr3区中,优选地不在vl链的cdr1、cdr2或cdr3区中或vl链的fr4区中。共同轻链的优选的序列示于图1中。
139、各种方法可用于产生双特异性抗体。一种方法涉及两个不同重链和两个不同轻链在细胞中的表达,并且收集由细胞产生的抗体。以这种方式产生的抗体将通常包含具有重链和轻链的不同组合的抗体的集合,其中一些是期望的双特异性抗体。双特异性抗体随后可以从该集合中纯化。由细胞产生的双特异性抗体与其它抗体的比率可以各种方式增加。在本发明的优选实施方案中,通过在细胞中不表达两种不同的轻链而是两种基本上相同的轻链来增加该比率。两个基本上相同的轻链可为具有基本相同的轻链可变区以及不同的轻链恒定区,或优选地两个基本相同的轻链恒定区的轻链。这种概念在本领域中也被称为“共同轻链”方法。当基本上相同的轻链与两个不同的重链一起工作从而允许形成具有不同抗原结合位点和伴随的不同结合特性的可变结构域时,由细胞产生的双特异性抗体与其他抗体的比率相比于两种基本上不同轻链的表达显著改善。与两个相同重链的配对相比,由细胞产生的双特异性抗体的比率可通过刺激两个不同重链的彼此配对进一步改善。现有技术描述了其中可实现重链的此类异源二聚化的各种方式。一个优选方法在美国临时申请61/635,935中有所描述,其由美国常规申请13/866,747和pct专利申请pct/nl2013/050294(wo2013/157954a1)进行了追踪,所述专利文献以引用方式并入本文。公开了用于产生双特异性抗体(由单一细胞)的方法和装置,由此提供与形成单特异性抗体相比,有利于形成双特异性抗体的装置。这些方法还可有利地用于本发明。因此,本发明提供了一种用于制备根据本发明的双特异性抗体(由单一细胞)的方法,其中所述双特异性抗体包含能够形成界面的两个ch3结构域,所述方法包括在所述细胞中提供:a)编码包含重链的第一ch3结构域的第一核酸分子,b)编码包含重链的第二ch3结构域的第二核酸分子,其中所述核酸分子设置有用于优先将包含重链的所述第一ch3结构域和第二ch3结构域配对的装置,所述方法还包括以下步骤:培养所述宿主细胞并且允许所述两种核酸分子表达并且从培养物中收获所述双特异性抗体。所述第一核酸分子和第二核酸分子可以为相同的核酸分子、载体或基因递送载体的一部分,并可以整合在宿主细胞的基因组的相同位点处。另选地,所述第一核酸分子和第二核酸分子独立地向所述细胞提供。宿主细胞包含至少一个轻链,并优选地共同轻链。
140、一个优选的实施方案提供了用于由单一细胞产生根据本发明的双特异性抗体的方法,其中所述双特异性抗体包含能够形成界面的两个ch3结构域,所述方法包括提供:
141、-细胞,其具有:a)编码重链的第一核酸分子,所述重链包含能够结合tnf受体超家族的膜缔合成员的细胞外部分并包含第一ch3结构域的抗原结合位点,b)编码重链的第二核酸分子,所述重链包含能够结合膜缔合第二蛋白质的细胞外部分并且包含第二ch3结构域的抗原结合位点,其中所述核酸分子具有用于优先将所述第一ch3结构域和第二ch3结构域配对的装置,
142、所述方法还包括以下步骤:培养所述细胞并允许表达由所述两种核酸分子编码的蛋白质并从培养物中收获所述双特异性igg抗体。在一个特别优选的实施方案中,所述细胞还具有编码共用轻链的第三核酸分子。所述第一核酸分子、第二核酸分子和第三核酸分子可以为相同的核酸分子、载体或基因递送载体的一部分,并可以整合在宿主细胞的基因组的相同位点处。另选地,所述第一核酸分子、第二核酸分子和第三核酸分子独立地向所述细胞提供。优选的共同轻链基于o12,优选地其为重排种系人k轻链igvκ1 39*01/igjκ1*01,如上所述。优先配对所述第一ch3结构域和所述第二ch3结构域的装置优选为重链编码区的ch3结构域中的相应突变。用于基本上仅产生双特异性抗体的优选的突变是第一ch3结构域中的氨基酸取代l351k和t366k(根据eu编号进行编号)以及第二ch3结构域中的氨基酸取代l351d和l368e,或反之亦然(图2)。因此,本发明还提供了根据本发明用于产生双特异性抗体的方法,其中所述第一ch3结构域包含氨基酸取代l351k和t366k(根据eu编号进行编号),并且其中所述第二ch3结构域包含氨基酸取代l351d和l368e,所述方法还包括以下步骤:培养所述细胞并允许表达由所述核酸分子编码的蛋白质并从培养物中收获所述双特异性抗体。本发明还提供了根据本发明用于产生双特异性抗体的方法,其中所述第一ch3结构域包含氨基酸取代l351d和l368e(根据eu编号进行编号),并且其中所述第二ch3结构域包含氨基酸取代l351k和t366k,所述方法还包括以下步骤:培养所述细胞并允许表达所述核酸分子并从培养物中收获所述双特异性抗体。可由这些方法产生的抗体也是本发明的一部分。ch3异源二聚化结构域优选为igg1异源二聚化结构域。包含ch3异源二聚化结构域的重链恒定区优选为igg1恒定区。
143、tnf受体超家族的成员(第一膜蛋白)优选地为cd137;ox40;cd40或cd30。在一个优选的实施方案中,第一膜蛋白为cd137或ox40,优选地cd137。本发明的结合分子优选地包含用于所述第一膜蛋白的一个(抗原)结合位点。在一些实施方案中,本发明的结合分子对于所述第一膜蛋白为单价。结合分子优选地包含用于所述第二膜蛋白的一个(抗原)结合位点。在一些实施方案中,本发明的结合分子对于所述第二膜蛋白为单价。在一些实施方案中,本发明的结合分子对于所述第一膜蛋白为单价且对于所述第二膜蛋白为单价。在一些实施方案中,本发明的结合分子对于tnf受体超家族成员为单价且对于b7家族成员为单价。在一些实施方案中,本发明的结合分子对于cd137为单价且对于b7家族成员为单价。在一些实施方案中,本发明的结合分子对于cd137为单价且对于pd-l1为单价。二价单克隆抗cd137抗体在本领域中已知用于活化cd137,而现有技术的单价cd137结合分子却通常不活化。
144、如本文所述的第一膜蛋白是tnf受体超家族成员(即细胞膜蛋白)。如果蛋白质具有存在于其所处的细胞的细胞膜中的跨膜区,该蛋白质被称为细胞上的膜蛋白。这通常为如本文所述的第一细胞。蛋白质还可具有跨膜区。在此类情况下,存在于细胞膜中的所有跨膜区存在于同一细胞的细胞膜中。第一膜蛋白为存在于细胞膜上时具有细胞外部分的细胞膜蛋白。细胞膜是使细胞内侧与细胞外侧隔开的细胞的膜。第一膜蛋白通常处于如本文所述的第一细胞的细胞膜上。在本发明的结合分子或本发明的方法或用途的上下文中,第一膜蛋白通常处于如本文所述的第一细胞的细胞膜上。第一膜蛋白可存在于所述第二细胞上,但优选的是第一膜蛋白的表达在所述第二细胞上忽略不计。通常,所述第一膜蛋白在所述第二细胞上的水平为相比于第一细胞上第一膜蛋白的表达的最多10%。第二细胞优选地不显著表达所述第一膜蛋白。如果采用对所述第一膜蛋白特异的抗体,在facs测定中借助于免疫荧光不能检出(高于背景)第一膜蛋白,则表达至少不显著。
145、第二膜蛋白同样为细胞膜蛋白。其具有存在于其所处的细胞的细胞膜中的跨膜区。这通常为如本文所述的第二细胞。第二蛋白质还可具有跨膜区。在此类情况下,存在于细胞膜中的所有跨膜区存在于同一细胞的细胞膜中。第二膜蛋白为存在于细胞膜上时具有细胞外部分的细胞膜蛋白。在本发明的结合分子或本发明的方法或用途的上下文中,第二膜蛋白通常处于如本文所述的第二细胞的细胞膜上。第二膜蛋白可存在于所述第一细胞上,但优选的是第二膜蛋白的表达在所述第一细胞上忽略不计。通常,所述第二膜蛋白在所述第一细胞上的水平为相比于第二细胞上的第二膜蛋白的表达的最多10%。第一细胞优选地不显著表达所述第二膜蛋白。如果采用对所述第二膜蛋白特异的抗体,在facs测定中借助于免疫荧光不能检出(高于背景)第二膜蛋白,则表达至少不显著。
146、根据一些实施方案,根据本发明的结合分子或(双特异性)抗体或变体具有能够结合tnf受体超家族的成员的细胞外部分的一个抗原结合位点,以及能够结合不为tnf受体超家族的成员的第二膜蛋白的第二抗原结合位点。其提供的优点在于,表达tnf受体超家族的几个不同成员的(免疫)细胞诸如t细胞的顺式活化被至少部分地避免,从而减小了归因于非特异性t细胞活化的潜在不利副作用和毒性。本发明的这些实施方案与涉及结合至tnf超家族受体的结合剂,尤其是结合至tnf超家族的至少两个不同的受体的结合剂的现有技术方法形成对比。此类现有技术方法可导致t细胞顺式活化,这意味着不存在第二靶标,并且可能涉及过度t细胞应答的风险,例如导致细胞因子风暴。因此,相比于具有能够结合tnf受体超家族的成员的细胞外部分的第一抗原结合位点以及能够结合不为tnf受体超家族的成员的膜蛋白的第二抗原结合位点的根据本发明的结合分子,此类现有技术方法潜在的不利副作用增大。
147、还提供了包含能够结合cd137或ox40的细胞外部分的抗原结合位点以及能够结合第二膜蛋白的细胞外部分的抗原结合位点的抗体或其功能部分、衍生物和/或类似物,其中所述第二膜蛋白不为tnf受体超家族的成员。还提供了一种刺激细胞上tnf受体超家族的成员的活性的方法,包括提供第一细胞和第二细胞,其中所述第一细胞在细胞膜上具有所述tnf受体超家族成员并且所述第二细胞在细胞膜上具有第二膜蛋白,所述方法包括使所述细胞与包含两个可变结构域的抗体或其功能部分、衍生物和/或类似物接触,其中一个可变结构域包含能够结合所述tnf受体超家族的成员的细胞外部分的第一抗原结合位点,并且另一个可变结构域包含能够结合所述第二膜蛋白的细胞外部分的第二抗原结合位点,从而刺激所述第一细胞上所述成员的活性;其中所述第二膜蛋白不为tnf受体超家族的成员。在一些实施方案中,所述方法是体外方法。
148、在一些实施方案中,所述抗体或其功能部分、衍生物和/或类似物包含能够结合所述tnf受体超家族成员的一个抗原结合位点以及能够结合不为tnf受体超家族成员的所述第二膜蛋白的一个抗原结合位点。在一些实施方案中,本发明的所述抗体或功能部分、衍生物和/或类似物的抗原结合部分由能够结合所述tnf受体超家族的成员的细胞外部分的一个免疫球蛋白可变结构域以及能够结合不为tnf受体超家族成员的所述第二膜蛋白的一个免疫球蛋白可变结构域组成。所述双特异性抗体优选地为全长抗体。在一些实施方案中,所述双特异性抗体为全长igg,即全长igg1、igg2、igg3或igg4,优选地全长igg1或全长igg4。
149、还提供了包含能够结合cd137或ox40的细胞外部分的抗原结合位点以及能够结合第二膜蛋白的细胞外部分的抗原结合位点的抗体或其功能部分、衍生物和/或类似物,其中所述第二膜蛋白为b7家族的成员,优选pd-l1。还提供了一种刺激细胞上tnf受体超家族的成员的活性的方法,包括提供第一细胞和第二细胞,其中所述第一细胞在细胞膜上具有所述tnf受体超家族的成员并且所述第二细胞在细胞膜上具有第二膜蛋白,所述方法包括使所述细胞与包含两个可变结构域的抗体或其功能部分、衍生物和/或类似物接触,其中一个可变结构域包含能够结合所述tnf受体超家族的成员的细胞外部分的第一抗原结合位点,并且另一个可变结构域包含能够结合所述第二膜蛋白的细胞外部分的第二抗原结合位点,从而刺激所述第一细胞上所述成员的活性;其中所述第二膜蛋白为b7家族的成员,优选地pd-l1。在一些实施方案中,所述方法是体外方法。
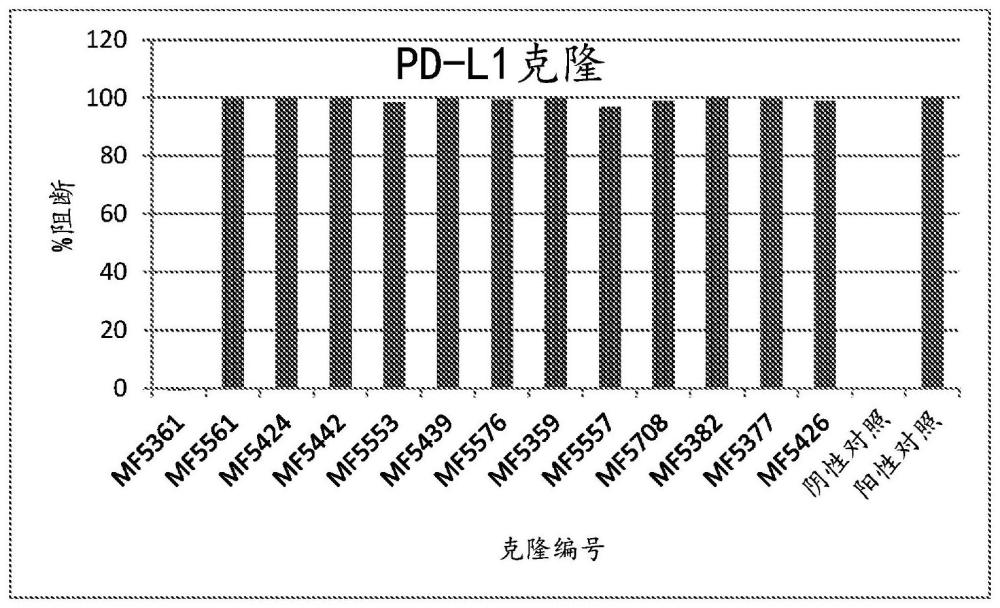
150、在一些实施方案中,所述抗体或其功能部分、衍生物和/或类似物包含能够结合所述tnf受体超家族成员的一个抗原结合位点以及能够结合所述第二膜蛋白即b7家族的成员、优选地pd-l1的一个抗原结合位点。在一些优选的实施方案中,本发明的所述抗体或功能部分、衍生物和/或类似物的抗原结合部分由能够结合所述tnf受体超家族的成员的细胞外部分的一个免疫球蛋白可变结构域以及能够结合所述第二膜蛋白即b7家族成员,优选地pd-l1的一个免疫球蛋白可变结构域组成。所述双特异性抗体优选地为全长抗体。在一些实施方案中,所述双特异性抗体为全长igg,即igg1、igg2、igg3或igg4,优选地全长igg1或全长igg4。
151、“阻断”所述第一膜蛋白与其结合配偶体结合的可变结构域干扰第一膜蛋白与所述结合配偶体的结合。此类可变结构域能够结合第一膜蛋白。此类阻断可变结构域能够结合所述第一膜蛋白上的表位并且与第一膜蛋白的结合配偶体竞争性结合表位。此类阻断可变结构域和第一膜蛋白的结合配偶体也可结合至所述第一膜蛋白上的不同表位。在此类情况下,阻断活性可例如归因于结合配偶体的减弱的结合,和/或结合配偶体在已经结合至所述第一膜蛋白时被置换,和/或阻断可变结构域可通过空间位阻防止结合配偶体结合至第一膜蛋白。所有这些及其他机制可至少部分地防止所述结合配偶体结合至所述第一膜蛋白。
152、在一个实施方案中,包含结合tnf受体超家族成员的抗原结合位点的结构域阻断成员与成员配体的结合。tnf受体超家族成员-配体相互作用已经在本领域得到广泛地研究。一般来讲,通常已知tnf受体超家族成员具有至少一种配体。已知的受体配体对的示例为tnf受体肿瘤坏死因子受体1和2以及配体tnf-α;受体ox40和配体ox40l;受体cd40和配体cd154;fas受体和配体fasl;cd30受体和配体cd153;以及受体cd137和配体cd137l。在一些实施方案中,包含结合tnf受体超家族成员的抗原结合位点的可变结构域不阻断成员与成员配体的结合。
153、在一些实施方案中,结构域包含结合tnf受体超家族成员的抗原结合位点,并且阻断其tnf受体超家族靶膜蛋白与该靶膜蛋白的结合配偶体相结合。所述可变结构域的特征还可使得当提供在包含两个所述可变结构域的单特异性二价抗体中时,其并不刺激细胞上tnf-受体超家族成员的活性而无交联。在一些实施方案中,包含结合tnf受体超家族成员的抗原结合位点的结构域包含阻断cd137与cd137l结合的可变结构域,所述可变结构域的特征还在于这样的事实:当提供于包含两个所述可变结构域的单特异性二价抗体中时,其并不刺激细胞上cd137的活性。
154、术语“结合配偶体;结合对;受体-配体对等是指能够彼此结合并且作为结合的结果产生活性的蛋白质。配偶体或对中的至少一者是细胞的细胞膜上的膜蛋白。活性通常作用于在细胞上具有该膜蛋白的细胞。
155、当相比于不存在可变结构域情况下的结合时,阻断如本文所述的膜蛋白的特异性结合对的结合的可变结构域通常使对的结合下降。这优选地在体外测定中测量。通常,这通过使可变结构域与其能够结合的膜蛋白一起温育,并随后使混合物与对的其他成员一起温育来进行。随后将对的结合与不存在可变结构域情况下对的结合进行比较。可变结构域可完全防止第一膜蛋白与其结合配偶体的结合。其也可部分地防止结合对的结合。当相比于不存在可变结构域情况下的结合,阻断膜蛋白的特异性结合对的结合的可变结构域优选地使对的结合下降至少50%,优选地至少60%,优选地至少70%,优选地至少80%,并且更优选地至少90%。可变结构域对于结合的阻断在本文定义为用包含两个相同的所述可变结构域的二价单克隆抗体所获得的阻断。可变结构域当然也在存在于包含所述可变结构域和结合第二膜蛋白的可变结构域的抗体中时阻断结合。
156、能够结合cd137的细胞外部分并且至少部分地阻断cd137配体与cd137结合的特异性可变结构域是包含以下项的vh的氨基酸序列的可变结构域:mf6783;mf6861;mf6795;mf6808;mf6798;mf6754;mf6763;mf6744;mf6785;mf6825;mf6737;mf6749;mf6788;或mf6797。
157、能够结合pd-l1的细胞外部分并且阻断pd1与pd-l1结合的特异性可变结构域是包含以下项的vh的氨基酸序列的可变结构域:mf5554;mf5576;mf5578;mf9375;mf9376;mf7702;mf5359;mf5377;mf5382;mf5424;mf5426;mf5439;mf5442;mf5553;mf5557;mf5561;mf5576;mf5594;或mf5708。氨基酸序列示于图3中。
158、结合tnf受体超家族成员的可变结构域优选地为提供于包含两个所述结构域的单特异性二价抗体中时不刺激细胞上tnf受体超家族成员的活性的可变结构域。所述可变结构域在二价单特异性抗体上下文中包含两个相同的所述可变结构域,并且不刺激包含tnf受体超家族成员的细胞的活性。
159、对细胞上tnf受体超家族的成员活性的刺激通常通过测量细胞的生物活性进行测量。活性的类型取决于要分析tnf受体超家族的哪个成员。就ox40和cd137而言,例如,可测量ox40和/或cd137阳性t细胞的活化状态。ox40和cd137是所谓的共同刺激蛋白质,其刺激活化t细胞的活化。用于测量t细胞的活化的合适方法提供于实施例部分中。一种方法是测量活化t细胞或包含所述t细胞的组合物所产生的il-2、ifnγ和/或tnfα。其他tnf受体具有不同的生物活性。例如cd30表达于活化的t-细胞和b-细胞上,并且是细胞凋亡的正调节因子。cd30刺激的测量可通过测量响应于本发明的结合分子的活化b-细胞或t-细胞的细胞凋亡。cd40是抗原递呈细胞诸如巨噬细胞上所见的共刺激蛋白质,并且刺激进一步刺激了抗原递呈细胞的活化。当在如本文所讨论的结合分子存在下,优选地在根据本发明的抗体或功能部分、衍生物和/或类似物存在下所测量的活性高于在不存在根据本发明的结合分子,优选地抗体或功能部分、衍生物和/或类似物情况下其他方面相同的条件下所测量的活性时,tnf受体超家族的成员的活性被刺激。对活性的刺激包括诱导活性以及增强已存在的活性。
160、cd137或ox40的刺激活性优选地通过测量含有cd137或ox40的细胞的生物活性进行测量。生物活性优选地为表达cd137或ox40的细胞的活化状态。cd137和ox40是表达于免疫细胞,包括活化t细胞上的共同刺激分子。优选地通过测定免疫细胞(例如活化t细胞)的活化水平来测量cd137或ox40的活性刺激。在个体中,优选地通过测量个体的免疫细胞和/或t细胞的活化来测量cd137或ox40的活性刺激。另选地,其也可通过如下方式测定:在适用情况下,测量个体中的肿瘤应答;个体的病毒载量;或个体的寄生物载量。
161、本发明还提供了一种结合和/或活化t细胞的方法,该方法包括:提供包含t细胞以及所述t细胞待结合或活化的细胞(第二细胞)的体系;以及提供具有包含能够结合tnf受体超家族的成员的可变结构域以及能够结合第二蛋白质的细胞外部分的可变结构域的至少一种抗体,优选地至少一种双特异性抗体的所述体系;以及在容许t细胞结合和/或活化的条件下温育所述体系。在一些实施方案中,所述方法是体外方法。所述tnf受体超家族成员优选地为cd137或ox40,最优选地cd137,并且所述第二膜蛋白优选地不为tnf受体超家族的成员。所述第二膜蛋白优选地为b7家族的成员,最优选地pd-l1。所述t细胞待结合或活化的细胞优选地为免疫细胞(例如抗原递呈细胞或巨噬细胞)、赘生细胞、病毒感染细胞、或细胞内寄生物感染细胞。结合和/或活化t细胞将t细胞引导向特定靶标。活化t细胞是活化所述t细胞的t细胞受体。结合t细胞通常为活化t细胞。结合也可将已活化t细胞引导向抗体特异性靶标。容许所述t细胞变得结合和/或活化的条件通常为培养条件,但也可在非人动物中温育。该条件使得t细胞在不存在抗体时不结合。如果测量t细胞的集合,这些中的一些可以已经结合或活化,前提条件是该集合包含未结合或活化的足够t细胞。
162、本发明的抗体可使两种细胞靠近在一起,这允许由除本发明抗体所结合的tnf受体超家族的成员之外的蛋白质和第二膜蛋白所介导的细胞之间发生相互作用。一种此类相互作用是一种细胞上的t细胞受体与另一细胞上mhc的相互作用。
163、所述第一细胞和所述第二细胞优选地为不同的细胞。不同的细胞可同时表达所述第一膜蛋白和第二膜蛋白。然而通常,第一膜蛋白仅表达于所述第一细胞上,且所述第二膜蛋白仅表达于所述第二细胞上。如果第一细胞不表达所述第二膜蛋白;所述第二细胞不表达所述第一膜蛋白;或更优选地它们的组合,生物活性通常受到更多刺激。当tnf受体超家族成员为ox40;cd137;cd30或cd40时,优选的是所述第一细胞是免疫细胞,优选地t细胞。在这些情况下,优选的是第二细胞为异常细胞、肿瘤细胞或免疫细胞(例如,巨噬细胞或抗原递呈细胞)。异常细胞是通常不存在于健康个体中的细胞。此类细胞的非限制性优选示例为癌细胞、病毒感染细胞、寄生物感染细胞或被诱导表达第二膜蛋白的细胞。合适的第二细胞也为免疫细胞。此类细胞的优选示例为树突状细胞、巨噬细胞、骨髓谱系的其他细胞或b细胞。在一些情况下,细胞可由于相邻细胞诸如免疫细胞、成纤维细胞或癌细胞所释放的抑制因子而表达第二膜蛋白。在一些实施方案中,在主要组织相容性复合体(mhc)上下文中,所述第二细胞是呈递肿瘤抗原或病原性抗原(如例如病毒抗原或寄生物抗原)的抗原递呈细胞。所述mhc复合体优选地为人白细胞抗原(hla)复合体。在该上下文中,本发明的抗体可同时增强体外抗原幼稚t细胞的扩增和分化。诱导和增强针对肿瘤抗原的新t细胞应答通常将导致更有效的肿瘤免疫和癌细胞清除。
164、cd137可被活化的t细胞表达。其也存在于其他细胞上,诸如树突状细胞、自然杀伤细胞、粒细胞以及炎症部位的血管壁细胞。该蛋白质因其对t细胞活化的共刺激活性而为人所知。配体cd137l表达于单核细胞、巨噬细胞及其他细胞上。cd137与cd137配体的结合在包含受体和配体的细胞中发挥作用。配体或受体的活化能够以各种方式实现。常见的方式是涂布到组织培养平板上或经由抗体交联(综述见schwartz,2004)。先天性及适应性免疫细胞上cd137的表达加上其加强抗肿瘤反应的能力使其建立为治疗靶标来增强肿瘤免疫。各种靶向cd137的免疫治疗已达到临床开发阶段(综述见makkouk等人2016)。受体或配体的活化似乎需要细胞膜中的同聚体缔合来发挥其效应。具有用于cd137的两个结合位点的抗体能够活化受体或配体。从而此类抗体对于cd137为二价。还产生仅具有一个cd137的结合位点的分子。现有技术的此类单价结合分子不可活化受体(mcnamara 2008)。相比之下,根据本发明的结合分子或抗体或变体能够活化cd137,即使在其对于cd137为单价的情况下也是如此。
165、ox40是t细胞上表达的次级共同刺激免疫检查点分子。ox40表达不为组成型,其通常在活化后24小时至72小时表达;其配体ox40l也不表达于静息抗原递呈细胞上,但在它们活化之后表达。
166、在本发明中,据发现如果结合分子还具有用于另一细胞(第二细胞)上的第二膜蛋白的结合位点,包括当第二膜蛋白不为tnf受体超家族成员的成员时,仅具有一个用于tnf受体超家族成员(第一膜蛋白)的结合位点的结合分子可被活化。第二膜蛋白可为tnf受体超家族受体的配体。然而,通常其不为tnf受体超家族成员(第一膜蛋白)的配体。第二膜蛋白可为tnf受体超家族成员的配体,不为第一膜蛋白的配体。
167、细胞上tnf受体超家族的成员的活性的刺激通常需要将两种或更多种受体复合物结合在一起。细胞表面膜上排列的同型三聚配体通常通过结合相邻细胞上的多个tnf受体超家族成员复合物来实现这一点。认为tnf受体超家族复合物的簇聚有利于刺激细胞的活性。这例如在仅具有cd137受体的细胞质部分的人工受体中也是显而易见的。各种人工受体的细胞质部分的紧密靠近也刺激包含人工受体的细胞。tnf受体超家族成员特异的二价单特异性抗体还能够模拟配体效应并刺激活性。认为抗体的臂使受体结合在一起或簇聚。此类活性也称为受体的交联。活性有时可通过提供抗-抗体抗体而被进一步刺激,这进一步提供了受体的交联。仅具有一个tnf受体超家族成员的结合位点的现有技术结合分子通常不能刺激活性。此类分子无法簇聚或交联tnf受体超家族成员。然而,根据本发明的结合分子或抗体或变体能够刺激tnf受体超家族成员的活性,包括当其对于所述tnf受体超家族成员为单价的情况。不受任何理论的束缚,据信根据本发明的双特异性抗体也可使tnf受体超家族受体复合物簇聚,从而促进tnf受体超家族成员活化。当根据本发明的抗体所结合的第二膜蛋白是b7家族的成员诸如pd-l1时,这尤其适用。
168、在本发明的一个方面,第二膜蛋白作为包括所述第二膜蛋白的两个或更多个实例的多聚体蛋白的一部分存在于细胞膜上。此类多聚体蛋白质可为本发明的结合分子的抗原结合位点提供两个或更多个表位。在这种情况下,其他细胞上结合的tnf受体超家族成员将变得簇聚并刺激含有tnf受体的细胞的活性。本发明的结合分子可通过结合不同细胞上的另一蛋白质而迫使tnf受体超家族成员靠近。该特征也称为反交联,这与结合相同细胞上tnf受体超家族成员的两个或更多个实例的结合分子的顺交联相反。在一些实施方案中,第二膜蛋白为同源二聚体或同源三聚体。同源二聚体是由两个相同的多肽单元构成的蛋白质。同源三聚体是由三个相同的多肽单元构成的蛋白质。不受任何理论的束缚,据信用于第二膜蛋白的抗原结合位点的表位共同结合多个结合分子。这些从而充当迫使两个或更多个tnf受体超家族成员紧密靠近的锚。紧密地靠近足以刺激细胞上tnf受体超家族成员的活性。
169、在另一个实施方案中,所述第二膜蛋白为存在于细胞膜上的一个或多个离散区内的蛋白质。细胞膜不提供细胞膜的所有组分的均一分布。目前已知细胞膜具有相比于膜的其他部分而言细胞膜的一种或多种组分更常见地存在的区(综述见vereb,g.,等人2003proc.natl.acad.sci.100.14:8053-8058)。所述区优选地为细胞膜上的蛋白质簇、结构域、微结构域或隔室,优选地免疫突触。不受任何理论的束缚,据信非随机分布有利于tnf受体超家族成员紧密靠近。
170、在其他实施方案中,细胞上tnf受体超家族成员的活性的刺激通过提供结合tnf受体超家族的相同成员(第一膜蛋白)和相同第二膜蛋白的两种或更多种结合分子来实现。涉及两种或更多种结合分子的实施方案也称为实施方案。如例如图15和图16所示,实施方案可导致t细胞活化。用于制备此类产物的一般方法公开于wo 2013/157953和wo2004/009618中并且以引用方式并入本文。术语“oligoclonics”是用表示的注册商标。在实施方案中,至少两种结合分子结合所述第一膜蛋白上的不同表位;所述第二膜蛋白上的不同表位;或所述第一膜蛋白上的不同表位以及所述第二膜蛋白上的不同表位。实施方案允许两种或更多种结合分子结合至第一膜蛋白和/或第二膜蛋白的相同分子,从而刺激细胞上tnf受体超家族成员的活性。优选的是至少两种结合分子结合所述第二膜蛋白上不同的表位。或结合所述第一膜蛋白上的不同表位以及所述第二膜蛋白上的不同表位。在一个特别优选的实施方案中,至少两种结合分子结合所述第一膜蛋白上的相同表位以及结合所述第二膜蛋白上的不同的表位。在一些实施方案中,两种结合分子阻断tnf受体超家族成员-配体相互作用。在其他实施方案中,两种结合分子不阻断tnf受体超家族成员-配体相互作用。
171、第一膜蛋白和第二膜蛋白上的不同表位是优选的,使得结合一个表位的结合分子和结合不同的表位的结合分子的同时结合成为可能。在一个优选的实施方案中,不同的表位为非竞争性表位。在一个优选的实施方案中,所述第一结合分子和所述第二结合分子能够结合cd137或ox40的相同结构域。在一个优选的实施方案中,所述第一结合分子和所述第二结合分子结合cd137或ox40上相同的表位。
172、第二膜蛋白优选地为多聚体细胞因子受体、b7家族的成员、cd28家族的成员;atp结合盒转运蛋白(abc转运蛋白)的成员;水通道蛋白;丝氨酸/苏氨酸激酶受体家族的成员;受体酪氨酸激酶家族的成员。在一个优选的实施方案中,第二膜蛋白是b7家族的成员。在一个优选的实施方案中,b7家族成员是cd80;cd86;pd-l1;pd-l2;icosl;b7-h3;b7-h4;b7-h5;b7-h6;或b7-h7。优选的是第二膜蛋白是b7家族的共抑制蛋白。在该优选的实施方案中,优选的是结合第二膜蛋白的可变结构域阻断b7家族成员与cd28家族的其结合配偶体结合。这样,第二膜蛋白所提供的对第一细胞潜在共抑制信号下降。在一个特别优选的实施方案,第二膜蛋白是pd-l1或pd-l2,优选地pd-l1。在另一个优选的实施方案中,第二膜蛋白是egf受体家族(erbb);胰岛素受体家族;igf受体家族;fgf受体家族;vegf受体家族;hgf受体家族;或axl受体家族的成员。第二膜蛋白优选地为egf受体家族(erbb)的成员,优选地egfr;erbb-2或erbb-3,优选地erbb-2。优选的是结合egf受体家族的egfr、erbb-3或erbb-4成员的可变结构域阻断生长因子与所述成员结合。在该实施方案中,所述第二细胞上的egf受体家族成员的活性下降。
173、在本发明的实施方案中,第二膜蛋白是结合对的成员。例如,egf受体(egfr)与egf形成结合对。合适的结合对的其他非限制性示例为her3和调蛋白;lgr5-rspondin;lgr4-rspondin;或b7家族成员配体以及cd28家族的其受体。在本发明的一个优选的实施方案中,如本文所述的结合分子阻断第二膜蛋白与结合对的互补成员结合。此类结合分子通常刺激细胞上tnf受体超家族的成员的活性并且阻断第二膜蛋白的活性。此类结合分子特别好地适于第二细胞为肿瘤细胞的情况,或适于治疗患有癌症的个体。在一个优选的实施方案中,第二膜蛋白是b7家族的成员,优选地pd-l1或pd-l2,优选地pd-l1,并且所述至少一种结合分子优选地阻断所述b7家族成员与cd28家族的其正常受体结合。在一个优选的实施方案中,第二膜蛋白是pd-l1,并且所述至少一种结合分子优选地阻断pd-l1与pd-1结合。
174、本发明提供了增强表达cd137的细胞的生物效应的方法,该方法包括提供具有第一细胞和第二细胞的体系,其中所述第一细胞在细胞膜上包含cd137,并且所述第二细胞在细胞膜上包含蛋白质,所述蛋白质的细胞外部分上具有相同表位的两个或更多个实例(即,存在相同表位的两个或更多个拷贝);以及提供具有包含cd137的细胞外部分的结合位点以及所述表位的结合位点的结合分子的所述体系;该方法还包括在允许所述生物活性增强的条件下温育所述体系。在一些实施方案中,所述方法是体外方法。在一些实施方案中,所述表达cd137的细胞是免疫细胞,优选地t细胞,并且所述第二细胞是肿瘤细胞。在一些实施方案中,所述表达cd137的细胞是免疫细胞,优选地t细胞,并且所述第二细胞是另一种免疫细胞。在一些实施方案中,所述表达cd137的细胞是免疫细胞,优选地t细胞,并且所述第二细胞是骨髓谱系细胞。在一些实施方案中,所述表达cd137的细胞是免疫细胞,优选地t细胞,并且所述第二细胞是原递呈细胞。所述抗原递呈细胞在mhc环境中,优选地hla环境中优选地呈递肿瘤抗原或病原性抗原。
175、如果成员被细胞表达,细胞通常在膜上具有tnf受体超家族成员。表达能够以各种方式进行测量。通常使用定量rna特异性pcr。也通常使用采用免疫荧光的免疫组织化学或facs分析。
176、其中提供第一细胞和第二细胞的合适体系是细胞培养物。另一种合适的体系是包含第一细胞和第二细胞的非人动物。其他合适的体系是体外体系,其中细胞以活性形式维持,但其中不一定有利于细胞的生长。可使第一细胞和第二细胞在例如不一定有利于生长但允许测量生物活性的测定条件下一起温育。
177、在容许细胞表达所述第一膜蛋白与所述第二膜蛋白的结合所介导的所述生物活性的条件下温育所述体系意指,使该体系维持在第一细胞和第二细胞能够由于结合配偶体而表现出生物活性的条件之下。体内或体外温育不必涉及超出经过足以允许生物活性变得显而易见的时间。
178、当相比于不存在可变结构域情况下的结合时,未阻断如本文所述的膜蛋白的特异性结合对的结合的可变结构域通常不使对的结合下降。这优选地在体外测定中测量。通常,这通过使可变结构域与其能够结合的膜蛋白一起温育,并随后使混合物与对的其他成员一起温育来进行。随后将对的结合与不存在可变结构域情况下对的结合进行比较。当相比于不存在可变结构域情况下的结合,如果其使对的结合下降不超过50%,优选地不超过40%,优选地不超过30%,优选地不超过20%,并且更优选地不超过10%,则认为可变结构域不阻断膜蛋白的特异性结合对的结合。可变结构域的结合以及阻断或不阻断与结合对的另一成员的结合在本文定义为使用包含所述两个相同的所述可变结构域的二价单克隆抗体所获得的阻断。阻断或不阻断定义为使用包含所述两个相同的所述可变结构域的二价单特异性抗体获得。
179、能够结合cd137的细胞外结构域并且不阻断cd137与cd137l结合的特异性可变结构域是包含以下项的vh的氨基酸序列的可变结构域:mf6860;mf6848;mf6805;mf6832;mf6870;mf6862;mf6875;或mf6873。
180、能够结合pd-l1的细胞外结构域并且不阻断pd1与pd-l1结合的特异性可变结构域是包含mf5361的vh的氨基酸序列的可变结构域。
181、可变结构域在性质上但并不一定在数量上的功能方面,诸如结合至抗原、阻断受体配体相互作用的能力、可变结构域的生物活性等能够以各种方式进行测定。合适的形式为fab片段或抗体。合适的抗体形式是包含两个可变结构域的单特异性二价抗体。另一种合适的形式例如是包含待测试可变结构域和另一可变结构域的双特异性抗体。另一可变结构域优选地为关于待实施的测定具有中性特异性的可变结构域。合适的中性可变结构域是能够结合破伤风类毒素的可变结构域。
182、本发明的抗体或其功能部分、衍生物和/或类似物优选地包含阻断其tnf受体超家族靶膜蛋白与该靶膜蛋白的结合配偶体结合的可变结构域。在一些实施方案中,本发明的抗体或其功能部分、衍生物和/或类似物包含这样的可变结构域:阻断其tnf受体超家族靶膜蛋白与该靶膜蛋白的结合配偶体结合,并且当提供于包含两个所述可变结构域的单特异性二价抗体中时,不刺激细胞上tnf受体超家族成员的活性。在一些实施方案中,本发明的抗体或其功能部分、衍生物和/或类似物包含这样的可变结构域:阻断cd137与cd137l结合,并且当提供于包含两个所述可变结构域的单特异性二价抗体中时,不刺激细胞上cd137的活性。
183、本发明还提供了用于治疗患有癌症的个体的方法,该方法包括向对其有需要的个体施用本发明的结合分子,优选本发明的抗体或功能部分、衍生物和/或类似物或本发明的双特异性抗体。个体优选为患有癌症的个体。在一些实施方案中,癌症是包括表达所述第二膜蛋白的癌细胞的癌症。在一些实施方案中,癌症是包括表达b7家族的成员的癌细胞的癌症。在一些实施方案中,所述个体的免疫细胞和/或骨髓谱系细胞表达所述第二膜蛋白,优选地b7家族的成员。在一些实施方案中,所述个体的抗原递呈细胞(apc)表达所述第二膜蛋白,优选地b7家族的成员。根据这些实施方案,癌细胞可以或不可表达所述第二膜蛋白。当apc表达所述第二膜蛋白时,所述癌症的抗原由个体的此类apc呈递,并且免疫细胞(优选地t细胞)的反式激活可由能够结合个体的免疫细胞以及免疫细胞、apc或肿瘤细胞的本发明抗体或功能部分、衍生物和/或类似物所诱导。在一些实施方案中,使用结合cd137和b7家族成员,优选地pd-l1的本发明的抗体或功能部分、衍生物和/或类似物。此类本发明的抗体或功能部分、衍生物和/或类似物可通过结合表达cd137的免疫细胞(优选地t细胞)以及表达所述b7家族的成员的肿瘤细胞和/或免疫细胞和/或骨髓谱系细胞和/或apc来反式激活免疫细胞。
184、癌症优选为腺癌。优选的癌症为结肠直肠癌;胰腺癌;肺癌;乳腺癌;肝癌;前列腺癌;卵巢癌;宫颈癌;子宫内膜癌;头颈癌;黑素瘤;睾丸癌;尿路上皮癌;肾癌;胃癌;或者良性肿瘤癌。在一个优选的实施方案中,癌症为结肠直肠癌;胰腺癌;肺癌;乳腺癌;肝癌;前列腺癌;卵巢癌;宫颈癌;子宫内膜癌;头颈癌;或黑素瘤。在一个特别优选的实施方案中,癌症为结肠直肠癌;胰腺癌;肺癌;乳腺癌;或肝癌。在一个特别优选的实施方案中,癌症为胃肠癌。在一个优选的实施方案中,癌症为结肠直肠癌。在该实施方案中,结合分子,优选地抗体或其功能部分、衍生物和/或类似物优选地为含有能够结合cd137或ox40的可变结构域以及能够结合pd-l1的可变结构域的抗体。结合cd137或ox40的可变结构域优选地阻断cd137与cd137配体结合,或者在ox40情况下阻断ox40与ox40配体结合。结合pd-l1的可变结构域优选地阻断pd-1与pd-l1结合。
185、还提供了一种体外体系,该体外体系包含抗体或其功能部分、衍生物和/或类似物,或本发明的双特异性抗体或其功能部分、衍生物和/或类似物,以及所述第一细胞、所述第二细胞。第一细胞和所述第二细胞优选地分别在细胞膜上表达所述第一膜蛋白和所述第二膜蛋白。体系优选地为适于所述第一细胞的维持和/或生长的细胞体系。细胞体系优选地适于所述第二细胞的维持和/或生长。诸如体系例如适于升高和/或增加针对异常细胞的免疫细胞。此类免疫细胞可随后施用给对其有需要的个体,例如癌症患者。免疫细胞优选地包括t细胞或nk细胞,优选地细胞毒性t细胞。免疫细胞优选地对于对其有需要的个体是自体同源的。
186、还提供了一种在所述个体中刺激个体针对异常细胞的免疫应答的方法,该方法包括提供给所述个体本发明的抗体或其功能部分、衍生物和/或类似物。异常细胞优选地为癌细胞、病毒感染细胞、寄生虫或寄生物感染细胞。在一个优选的实施方案中,细胞是癌细胞或赘生细胞。在该实施方案中,抗体或其功能部分、衍生物和/或类似物优选地为含有能够结合cd137或ox40的细胞外部分的可变结构域以及能够结合pd-l1的可变结构域的抗体。结合cd137或ox40的可变结构域优选地阻断cd137与cd137配体结合,或者在ox40情况下阻断ox40与ox配体结合。结合pd-l1的可变结构域优选地阻断pd-1与pd-l1结合。
187、瘤是组织的异常生长,并且当其还形成肿块时通常称为肿瘤。本发明的瘤通常形成肿块。赘生细胞是来自已形成肿块的瘤的细胞。世界卫生组织(who)将瘤分类为四个主要类型:良性肿瘤、原位肿瘤、恶性肿瘤、以及不确定或未知行为的瘤。恶性肿瘤也简单地称为癌症。
188、刺激免疫应答涵盖诱导免疫应答以及增强已存在的免疫应答。个体内的免疫应答可通过测量(适用情况下)以下项进行测量:个体的肿瘤载量;个体的病毒载量;个体的寄生物载量。
189、所述病毒感染细胞优选地为感染有免疫缺陷病毒、疱疹病毒(优选地单纯疱疹病毒)、水痘病毒、细胞巨化病毒或埃-巴二氏病毒、乳头状瘤病毒、肝炎病毒(优选地甲型肝炎病毒、乙型肝炎病毒或丙型肝炎病毒)、麻疹病毒或腺病毒的细胞。病毒优选地为已知能够在个体内持续存在的病毒。持续感染的特征是病毒未被清除但留在受感染个体的特定细胞内的那些。持续感染可包括隐性感染期和生产性感染期而不迅速杀伤宿主细胞或甚至产生宿主细胞的过度损伤。持续病毒-宿主相互作用可为潜伏性、慢性和/或长程感染。
190、寄生物感染细胞是感染有细胞内寄生物的细胞。此类寄生物是能够在宿主细胞内生长和繁殖的寄生微生物。一些细胞内寄生物也可在细胞外存活。此类寄生物是所谓的兼性细胞内寄生物。非限制性示例是单核细胞增多性李斯特菌(listeria monocytogenes)、军团菌属(legionella)、分枝杆菌(mycobacterium)和新型隐球菌(cryptococcusneoformans)的特定物种。优选的细胞内寄生物是不能在宿主细胞外生长的寄生物,优选的示例为衣原体(chlamydia),以及分枝杆菌的近缘种、特定种,诸如麻风分枝杆菌(mycobacterium leprae),特定原生动物,包括:顶复虫类(apicomplexans)(疟原虫属(plasmodium spp.),刚地弓形虫(toxoplasma gondii)以及小球隐孢子虫(cryptosporidium parvum)和锥虫(trypanosomatids))。
191、本发明还提供编码根据本发明的抗体重链可变区的核酸分子。核酸分子(通常体外、分离或重组核酸分子)优选编码如图3所示的任一重链可变区,或具有3个、2个、3个、4个或5个氨基酸插入、缺失、取代或它们的组合的如图1所示的重链可变区。在一个优选的实施方案中,核酸分子包含如图3所示的序列。核酸分子优选地使用被最优化以在待使用的产抗体细胞中表达的密码子。优选地,编码如图3所示的重链可变区或具有1个、2个、3个、4个或5个氨基酸插入、缺失、取代或它们的组合的如图3所示的重链可变区的核酸经密码子优化以在人细胞,优选地per.c6tm;或中国仓鼠,优选地cho中表达。本发明还提供了编码所提及重链可变区连同图2的重链恒定区的核酸分子。
192、如本发明所用的核酸分子典型地但非排他性地为核糖核酸(rna)或脱氧核糖核酸(dna)。对于本领域技术人员而言,另选的核酸是可用的。根据本发明的核酸分子例如包含在细胞中。当所述核酸分子在所述细胞中表达时,所述细胞可产生根据本发明的抗体。因此,在一个实施方案中,本发明提供包含根据本发明的抗体和/或根据本发明的核酸分子的细胞。当所述细胞产生重链和轻链时,产生抗体。提供了能够产生本发明的抗体的细胞。细胞优选地包含编码包含抗体重链可变区的抗体重链的核酸分子,该抗体重链与共同轻链组合时能够结合所述第一膜蛋白。所述细胞优选地还包含编码包含抗体重链可变区的抗体重链的核酸分子,该抗体重链与共同轻链组合时能够结合所述第二膜蛋白。所述细胞优选地还包含编码共同轻链的核酸分子。所述细胞优选为动物细胞,更优选地哺乳动物细胞,更优选地灵长类细胞,最优选地人类细胞。出于本发明的目的,合适的细胞是能够包含并优选生产根据本发明的抗体和/或根据本发明的核酸的任何细胞。
193、本发明还提供包含根据本发明的抗体的细胞。还提供了包含单独或一起编码本发明抗体的一种或多种核酸分子的细胞。一种或多种核酸分子是可表达核酸分子,这意指它们包含rna转录和翻译蛋白质编码结构域所需的顺式信号。优选地,所述细胞(通常为体外、分离或重组细胞)产生所述抗体。在一个优选的实施方案中,所述细胞是杂交瘤细胞、中国仓鼠卵巢(cho)细胞、ns0细胞或per-c6tm细胞。在一个特别优选的实施方案中,所述细胞为cho细胞。还提供了包含根据本发明的细胞的细胞培养物。各种机构和公司已经开发了用于大规模生产抗体的细胞系,例如用于临床应用。此类细胞系的非限制性示例为cho细胞、ns0细胞或per.c6tm细胞。这些细胞也用于其它目的,诸如蛋白质的生产。被开发用于工业规模生产蛋白质和抗体的细胞系在本文中还被称为工业细胞系。因此,在一个优选的实施方案中,本发明提供了被开发用于大规模生产抗体的细胞系以用于生产本发明的抗体的用途。本发明还提供了一种细胞,其用于产生包含编码如图3、图1和图2所示的vh、vl和/或重链的核酸分子的抗体。优选地,所述核酸分子包含如图1和图2所示的序列。
194、本发明还提供了一种生产抗体的方法,所述方法包括培养本发明的细胞并从所述培养物中收获所述抗体。优选地,所述细胞在无血清培养基中培养。优选地,所述细胞适于悬浮生长。本发明还提供了可通过用于生产根据本发明的抗体的方法获得的抗体。所述抗体优选由培养物的培养基中纯化。优选所述抗体经亲和纯化。
195、本发明的细胞例如为杂交瘤细胞系、cho细胞、293f细胞、ns0细胞或本领域已知适用于出于临床目的而产生抗体,尤其是产生用于施用给人类的抗体的任何其他细胞类型。在一个特别优选的实施方案中,所述细胞是人细胞,优选地由腺病毒e1区转化的细胞或其功能等同物。此类细胞系的优选示例为per.c6tm细胞系或其等同物。在一个特别优选的实施方案中,所述细胞是cho细胞或其变体,优选地利用表达抗体的谷氨酰胺合成酶(gs)载体体系的变体。
196、本发明还提供包含根据本发明的一种或多种抗体或其变体的药物组合物。药物组合物优选地包含药学上可接受的赋形剂或载体。
197、本发明的抗体或其变体还可包含标记,优选用于体内成像的标记。此类标记对于治疗应用而言通常不是必要的。在例如诊断装置中,标记可能是有帮助的。例如,体内的可视化靶细胞中。各种标签是适合的,并且许多在本领域为人们所熟知。在一个优选的实施方案中,标记是用于检测的放射性标记。在另一个优选的实施方案中,标记是红外标记。优选地,红外标记适用于体内成像。对于本领域技术人员而言,各种红外标记是可用的。优选的红外标记例如为:irdye 800;irdye 680rd;irdye 680lt;irdye 750;irdye 700dx;irdye800rs irdye 650;irdye 700亚磷酰胺;irdye 800亚磷酰胺(li-cor usa;4647superiorstreet;lincoln,nebraska)。
198、待施用于患者的根据本发明的抗体量通常在治疗窗口内,这是指为获得治疗效果使用足够的量,然而该量不超过阈值从而导致不可接受的程度的副作用。为获得期望的治疗效果所需的抗体量越低,则治疗窗口通常将越大。因此,优选在低剂量下发挥足够治疗效果的根据本发明的抗体。剂量可以在纳武单抗的给药方案的范围内。剂量也可以更低。
199、在肿瘤抑制环境中,周围细胞上pd-l1的表达预计达到将致使t细胞上的cd137活化的密度阈值,如实施例8所述(图28a和图28b)。这样特异性抗体将能够活化肿瘤内的t细胞,并且不-或者在显著较小程度上-作用于表达低pd-l1细胞表面水平的细胞。在cd137×pd-l1双特异性抗体包含阻断pd-l1的fab臂的情况下,抗体此外将克服pd-1/pd-l1阻断。通过以“反式”作用,cd137×pd-l1抗体将释放pd-1/pd-l1阻断,同时通过活化cd137来活化t细胞。因此,cd137×pd-l1抗体可增强局部t细胞应答,导致大量细胞因子释放(实施例9),其继而可活化肿瘤微环境中的其他免疫细胞,并且至少部分地克服肿瘤的局部免疫抑制。本发明人已经示出,相比于基于具有同种类型特异性的现有技术基准抗体的抗体,如例如基于优卢单抗(urelumab)(抗cd137)或基于阿特朱单抗(抗pd-l1)的抗体,根据本发明的双特异性抗体通常具有较好的t细胞活化特性。在示例中,相比于两种此类基于基准的抗体的混合物,根据本发明的双特异性抗体实现了较强的t细胞活化活性。这例如示于当前示例的t细胞反式激活测定和seb刺激测定中。还已证明,根据本发明的双特异性抗体能够逆转肿瘤相关m2巨噬细胞所诱导的免疫抑制,并且能够(重新)刺激体外从患者肿瘤分离的肿瘤特异性t细胞。根据本发明的双特异性抗体能够(重新)刺激肿瘤特异性cd4+效应记忆t细胞、肿瘤特异性cd8+效应记忆t细胞以及肿瘤特异性cd8+终末分化t细胞,而基于阿特朱单抗的基准抗pd-l1抗体通常仅(重新)刺激cd4+ t细胞。从而,相比于基准抗体,根据本发明的双特异性抗体具有(重新)刺激抗原特异性t细胞的更可变子集(包括cd8+t细胞)的潜能。
200、除了通过活化经历过抗原的cd8 t细胞而针对肿瘤再生出现有细胞毒性t细胞应答之外,cd137×pd-l1双特异性抗体还可提高新发cd8+ t细胞抗肿瘤应答。通过濒死肿瘤细胞或抗原递呈细胞所吞噬的肿瘤细胞而脱落到环境中的肿瘤(新)抗原被转运至引流淋巴结或三级淋巴结构(其为存在于肿瘤组织中的异位淋巴形成物)。在局部肿瘤环境中,肿瘤抗原被呈递给将在抗原识别时扩增和分化的幼稚cd8+ t细胞。
201、如示例中所示,相比于基于优卢单抗或基于阿特朱单抗的基准抗体,根据本发明的抗体可在cd8+ t细胞引发后使t细胞扩增增强到较高的程度。在示例中,已经表明相比于基于优卢单抗和基于阿特朱单抗的基准抗体的混合物,根据本发明的抗体可更大地同时诱导抗原特异性cd8+ t细胞的扩增和分化,这将有利于产生肿瘤特异性记忆和终末分化杀伤t细胞的较大群体。
202、相比于具有可变结构域的二价单特异性抗体的组合,抗体或其变体并且尤其是根据本发明的双特异性抗体或其变体可具有较低的副作用。阻断抑制和/或共同刺激分子的抗体组合有益于不对现有免疫疗法作出反应的患者。然而,免疫调节受体(imod)的双重阻断已示出提高免疫相关的细胞毒性。抗体或其变体并且尤其是根据本发明的双特异性抗体或其变体适于解决imod的双重阻断,因为它们可发挥不能由单克隆抗体组合再生的功能活性,并且可更具选择性地靶向特定细胞群,这使患者的安全责任下降。不受任何理论的束缚,据信相比于(组合的)单特异性抗体,本发明的双特异性抗体或变体的不利副作用的几率减小至少部分地因为本发明的双特异性抗体或变体通常表现出反式t细胞活化,而其具有较低的顺式t细胞活化活性。在本发明上下文中,使用具有低顺式t细胞活化活性的抗体是优选的,因为这减弱了潜在的非特异性t细胞应答。相比于具有可变结构域的二价单特异性抗体的组合,根据本发明的抗体或其双特异性抗体或功能部分、衍生物和/或类似物具有较小的免疫相关的细胞毒性。
203、鉴于上述内容,根据本发明的双特异性抗体、或其功能部分、衍生物和/或类似物对于治疗性应用是优选的。
204、通过将抗体克隆到互补表达载体中产生双特异性抗体形式的抗体,所述互补表达载体包含在ch3区中的突变,所述突变驱动重链的异源二聚化。许多双特异性抗体以较小规模生产并在结合和功能测定中对癌细胞系进行测试。本发明的抗体,具体地讲本发明的双特异性抗体可将低毒性分布与高功效组合。本发明的抗体可用于各种类型和系列的免疫靶向治疗。但与两个臂结合相同抗原的抗体相比,本发明的抗体可具有增加的治疗窗口。
205、本发明还提供根据本发明的双特异性抗体或其功能部分、衍生物和/或类似物用于制备治疗或预防异常细胞、肿瘤和/或癌细胞转移形成的药物的用途。所述转移瘤起源于其的肿瘤优选地为对所述第二细胞膜蛋白呈阳性,优选地对b7家族的成员呈阳性的肿瘤。
206、本发明的抗体可在悬浮293f细胞中瞬时转染之后以>50mg/l的含量制备。可纯化双特异性抗体至大于98%的纯度,收率>70%。分析表征研究示出与二价单特异性igg1相当的双特异性igg1抗体特征。
207、本发明还提供了双特异性抗体或其功能部分、衍生物和/或类似物,其可结合tnf受体超家族的膜结合成员的细胞外部分和膜结合的第二膜蛋白,优选地b7家族的成员的细胞外部分。在一些实施方案中,所述双特异性抗体或其功能部分、衍生物或类似物包含能够结合所述tnf受体超家族的成员的一个抗原结合位点以及能够结合所述第二蛋白质,优选地所述b7家族的成员的一个抗原结合位点。在一些优选的实施方案中,本发明的所述双特异性抗体或功能部分、衍生物或类似物的抗原结合部分由能够结合所述tnf受体超家族的成员的细胞外部分的一个免疫球蛋白可变结构域以及能够结合所述b7家族的成员的一个免疫球蛋白可变结构域组成。所述双特异性抗体或其功能部分、衍生物或类似物优选地对于所述tnf受体超家族的成员为单价且对于所述b7家族的成员为单价。所述双特异性抗体优选地为全长抗体。在一些实施方案中,所述双特异性抗体为全长igg,即全长igg1、igg2、igg3或igg4,优选地全长igg1或全长igg4。
208、本发明还提供能够结合cd137的细胞外部分以及pd-l1的细胞外部分的双特异性抗体或其功能部分、衍生物和/或类似物。所述双特异性抗体或其功能部分、衍生物或类似物优选地包含两个抗原结合位点。所述双特异性抗体或其功能部分、衍生物或类似物优选地包含能够结合cd137的一个抗原结合位点以及能够结合pd-l1的一个抗原结合位点。在一些优选的实施方案中,本发明的所述双特异性抗体或功能部分、衍生物或类似物的抗原结合部分由能够结合cd137的细胞外部分的一个免疫球蛋白可变结构域以及能够结合pd-l1的一个免疫球蛋白可变结构域组成。所述双特异性抗体或其功能部分、衍生物或类似物优选地对于cd137为单价且对于pd-l1为单价。在一些实施方案中,能够结合cd137的抗原结合位点能够阻断cd137与cd137l结合。在一些实施方案中,能够结合cd137的抗原结合位点不能阻断cd137与cd137l结合。在一些实施方案中,能够结合pd-l1的抗原结合位点能够阻断pd-l1与pd-1结合。在一些实施方案中,能够结合pd-l1的抗原结合位点不能阻断pd-l1与pd-1结合。所述双特异性抗体优选地为全长抗体。在一些实施方案中,所述双特异性抗体为全长igg,即全长igg1、igg2、igg3或igg4,优选地全长igg1或全长igg4。
209、本发明还提供了用于治疗患有癌症的个体的方法,所述方法包括向对其有需要的个体施用本发明的结合分子或本发明的双特异性抗体或功能部分、衍生物或类似物。
210、本发明还提供了本发明的结合分子或本发明的双特异性抗体或功能部分、衍生物或类似物在治疗患有癌症的个体中的用途。
211、还提供了一种细胞体系,该细胞体系包含本发明的抗体或双特异性抗体或其功能部分、衍生物和/或类似物,以及表达tnf受体超家族的膜结合成员的第一细胞以及表达膜结合的第二膜蛋白,优选地b7家族的成员的第二细胞。
212、本发明提供一种刺激细胞上tnf受体超家族的成员的活性的方法,包括提供第一细胞和第二细胞,其中所述第一细胞在细胞膜上具有所述成员并且所述第二细胞在细胞膜上具有第二膜蛋白,所述方法包括使所述细胞与包含两个可变结构域的双特异性抗体或其变体接触,其中一个可变结构域包含能够结合所述tnf受体超家族的成员的细胞外部分的第一抗原结合位点,并且其中另一个可变结构域包含能够结合所述第二膜蛋白的细胞外部分的第二抗原结合位点,从而刺激所述第一细胞上所述成员的活性。在一些实施方案中,所述双特异性抗体包含能够结合所述tnf受体超家族的成员的一个抗原结合位点。在一些实施方案中,所述方法是体外方法。在一个优选的实施方案中,所述tnf受体超家族的成员是cd137或ox40。所述第二膜蛋白优选地不为tnf受体超家族的成员。所述双特异性抗体优选地对于所述tnf受体超家族的成员为单价且对于所述第二膜蛋白为单价,优选地对于b7家族的成员为单价。所述双特异性抗体优选地为全长抗体。在一些实施方案中,所述双特异性抗体为全长igg,即全长igg1、igg2、igg3或igg4,优选地全长igg1或全长igg4。
213、所述第一细胞优选地不在细胞膜上显著地表达所述第二膜蛋白。所述第二膜蛋白优选地为存在于细胞膜上的一个或多个区中的蛋白质。所述区优选地为细胞膜上的簇、结构域、微结构域或隔室,优选地免疫突触。所述第二膜蛋白优选地作为包括所述第二膜蛋白的两个或更多个实例的多聚体蛋白的一部分存在于细胞膜上。在一些实施方案中,所述第二膜蛋白作为同源二聚体或同源三聚体的一部分存在于细胞膜上。在一个优选的实施方案中,所述第二膜蛋白为多聚体细胞因子受体、b7家族的成员、cd28家族的成员;atp结合盒转运蛋白(abc转运蛋白)成员;水通道蛋白;丝氨酸/苏氨酸激酶受体家族的成员;受体酪氨酸激酶家族的成员。第二膜蛋白优选地为b7家族的成员,优选地pd-l1或pd-l2,优选地pd-l1。在一个优选的实施方案中,第二膜蛋白是egf受体家族(erbb);igf受体家族;fgf受体家族;vegf受体家族;hgf受体家族;或axl受体家族的成员。第二膜蛋白优选地为egf受体家族(erbb)的成员,优选地egfr;erbb-2或erbb-3,优选地erbb-2。优选地,结合tnf受体超家族的成员的可变结构域阻断了配体与成员的结合。结合所述tnf受体超家族的成员的细胞外部分的可变结构域优选地被定义为如下可变结构域:当处于包括两个结合所述tnf受体超家族的成员的所述可变结构域的二价单特异性抗体形式时,不刺激细胞上所述tnf受体超家族成员的活性。方法优选地还包括提供包含能够结合所述tnf受体超家族的成员的细胞外部分的抗原结合位点以及能够结合所述第二膜蛋白的细胞外部分的抗原结合位点的另一种双特异性抗体,其中所述第一双特异性抗体和所述第二双特异性抗体结合:
214、-所述第一膜蛋白上的不同表位;
215、-所述第二膜蛋白上的不同表位;或者
216、-所述第一膜蛋白上的不同表位以及所述第二膜蛋白上的不同表位;
217、所述方法还包括将所述第一细胞和所述第二细胞与所述第一双特异性抗体和所述第二双特异性抗体一起温育,从而刺激所述第一细胞上所述tnf受体超家族的成员的活性。在一些实施方案中,所述方法是体外方法。在一个优选的实施方案中,tnf受体超家族成员是cd137或ox40。在一些实施方案中,所述第一双特异性抗体和所述第二双特异性抗体各自包含能够结合所述tnf受体超家族的成员的一个抗原结合位点。所述第二膜蛋白优选地不为tnf受体超家族的成员。所述第一双特异性抗体和/或所述第二双特异性抗体优选地对于所述tnf受体超家族的成员为单价且对于所述第二膜蛋白为单价,优选地对于所述b7家族的成员为单价。所述第一双特异性抗体和/或所述第二双特异性抗体优选地为全长抗体。在一些实施方案中,所述第一双特异性抗体和/或所述第二双特异性抗体为全长igg,即全长igg1、igg2、igg3或igg4,优选地全长igg1或全长igg4。
218、能够结合所述第二膜蛋白的第一双特异性抗体和第二双特异性抗体的抗原结合位点优选地结合所述第二膜蛋白的细胞外部分上的不同表位。所述第二膜蛋白的细胞外部分上的不同表位优选地为非竞争性表位。
219、还提供了一种双特异性抗体,其包含能够结合cd137或ox40的细胞外部分的抗原结合位点以及能够结合第二膜蛋白的细胞外部分的抗原结合位点。在一些实施方案中,所述双特异性抗体包含能够结合所述cd137或ox40的一个抗原结合位点。所述第二膜蛋白优选地不为tnf受体超家族的成员。第二膜蛋白优选地不被t细胞显著程度地表达。第二膜蛋白优选地表达于免疫细胞、骨髓谱系细胞、抗原递呈细胞、肿瘤细胞、病毒感染细胞或寄生物感染细胞上。优选地,所述第二膜蛋白为存在于细胞膜上的一个或多个区中的蛋白质。所述区优选地为细胞膜上的簇、结构域、微结构域或隔室,优选地免疫突触。在一些实施方案中,所述第二膜蛋白是作为包括两个或更多个所述第二膜蛋白的多聚体蛋白的一部分存在于细胞膜上的蛋白质。在一些实施方案中,所述第二膜蛋白作为同源二聚体或同源三聚体的一部分存在于细胞膜上。优选地,所述第二膜蛋白为多聚体细胞因子受体、b7家族的成员、cd28家族的成员;atp结合盒转运蛋白(abc转运蛋白)成员;水通道蛋白;丝氨酸/苏氨酸激酶受体家族的成员;受体酪氨酸激酶家族的成员。第二膜蛋白优选地为b7家族的成员,优选地pd-l1或pd-l2,优选地pd-l1。在一些实施方案中,第二膜蛋白是egf受体家族(erbb);胰岛素受体家族;igf受体家族;fgf受体家族;vegf受体家族;hgf受体家族;或axl受体家族的成员。在一些实施方案中,第二膜蛋白是egf受体家族(erbb)的成员,优选地egfr;erbb-2或erbb-3,优选地erbb-2。结合所述cd137或ox40的可变结构域优选地阻断配体与成员结合。结合所述cd137或ox40的细胞外部分的可变结构域优选地被定义为如下可变结构域:当处于包括两个结合所述cd137或ox40的所述可变结构域的二价单特异性抗体形式时,不刺激细胞上cd137或ox40的活性。所述双特异性抗体优选地对于cd137或ox40为单价且对于所述第二膜蛋白为单价。所述双特异性抗体优选地为全长抗体。在一些实施方案中,所述双特异性抗体为全长igg,即全长igg1、igg2、igg3或igg4,优选地全长igg1或全长igg4。
220、本发明还提供了包含根据本发明的一种或多种双特异性抗体的组合物。还提供了一种包含本发明的两种或更多种双特异性抗体的组合物或成套试剂盒,其中第一双特异性抗体和第二双特异性抗体的能够结合cd137或ox40的抗原结合位点结合所述cd137或ox40上的不同表位。还提供了一种刺激细胞上cd137或ox40的活性的方法,包括提供第一细胞和第二细胞,其中所述第一细胞在细胞膜上具有所述cd137或ox40(第一膜蛋白)并且所述第二细胞在细胞膜上具有第二膜蛋白,所述方法包括使所述细胞与包含两个可变结构域的根据本发明的双特异性抗体(第一双特异性抗体)接触,其中一个可变结构域包含能够结合所述第一膜蛋白的细胞外部分的第一抗原结合位点,并且另一个可变结构域包含能够结合所述第二膜蛋白的细胞外部分的第二抗原结合位点,从而刺激所述第一细胞上所述第一膜蛋白的活性。在一些实施方案中,所述双特异性抗体包含能够结合所述第一膜蛋白的一个抗原结合位点。在一些实施方案中,所述方法是体外方法。该方法优选地还包括提供根据本发明的另一双特异性抗体(第二双特异性抗体),该另一双特异性抗体包含具有能够结合所述第一膜蛋白的细胞外部分的抗原结合位点的可变结构域;以及具有能够结合所述第二膜蛋白的细胞外部分的抗原结合位点的可变结构域,其中所述第一双特异性抗体和所述第二双特异性抗体结合:
221、-所述第一膜蛋白上的不同表位;
222、-所述第二膜蛋白上的不同表位;或者
223、-所述第一膜蛋白上的不同表位;以及所述第二膜蛋白上的不同表位;
224、该方法还包括将所述第一细胞和所述第二细胞与所述第一双特异性抗体和所述第二双特异性抗体一起温育,从而刺激所述第一细胞上cd137或ox40的活性。第二膜蛋白优选地为b7家族的成员,更优选地pd-l1。
225、如下文所示的mf序列所定义的抗体优选地为具有两个不同的可变结构域的双特异性抗体,其中这些可变结构域之一包含所示序列。
226、包含能够结合cd137的细胞外部分的可变结构域的抗体或其功能部分、衍生物和/或类似物优选地包含具有cdr3区的重链可变区,该cdr3区含有mf6754;mf6763;mf6785;或mf6797的可变重链区的cdr3区的氨基酸序列(图3)。
227、包含能够结合cd137的细胞外部分的可变结构域的抗体或其功能部分、衍生物和/或类似物优选地包含具有cdr1、cdr2和cdr3区的重链可变区,该cdr1、cdr2和cdr3区含有mf6754;mf6763;mf6785;或mf6797所示的vh之一的可变重链区的cdr1、cdr2和cdr3的氨基酸序列(图3)。cdr1、cdr2和cdr3序列优选地选自相同的vh区。
228、包含能够结合cd137的细胞外部分的可变结构域的抗体或其功能部分、衍生物和/或类似物优选地包含mf6754;mf6763;mf6785;或mf6797的可变重链区的氨基酸序列,其关于所示mf的vh的氨基酸序列具有最多15个,优选地0个、1个、2个、3个、4个、5个、6个、7个、8个、9个或10个,并且优选地具有0个、1个、2个、3个、4个或5个氨基酸插入、缺失、取代或它们的组合。氨基酸插入、缺失、取代或它们的组合(如果有的话)优选地不处于cdr区的氨基酸序列中。
229、包含能够结合pd-l1的细胞外部分的可变结构域的抗体或其功能部分、衍生物和/或类似物优选地包含具有cdr3区的重链可变区,该cdr3区含有mf5554;mf5576;mf5578;mf9375;mf9376;mf7702;mf5424;mf5561;mf5439;mf5553;mf5594;mf5426;mf5442或mf5361的可变重链区的cdr3区的氨基酸序列(图3)。
230、包含能够结合pd-l1的细胞外部分的可变结构域的抗体或其功能部分、衍生物和/或类似物优选地包含具有cdr1、cdr2和cdr3区的重链可变区,该cdr1、cdr2和cdr3区含有mf5554;mf5576;mf5578;mf9375;mf9376;mf7702;mf5424;mf5561;mf5439;mf5553;mf5594;mf5426;mf5442或mf5361所示的vh之一的可变重链区的cdr1、cdr2和cdr3的氨基酸序列(图3)。cdr1、cdr2和cdr3序列优选地选自相同的vh区。
231、包含能够结合pd-l1的细胞外部分的可变结构域的抗体或其功能部分、衍生物和/或类似物优选地包含mf5554;mf5576;mf5578;mf9375;mf9376;mf7702;mf5424;mf5561;mf5439;mf5553;mf5594;mf5426;mf5442或mf5361的可变重链区的氨基酸序列,其关于所示mf的vh的氨基酸序列具有最多15个,优选地0个、1个、2个、3个、4个、5个、6个、7个、8个、9个或10个,并且优选地具有0个、1个、2个、3个、4个或5个氨基酸插入、缺失、取代或它们的组合。氨基酸插入、缺失、取代或它们的组合(如果有的话)优选地不处于cdr区的氨基酸序列中。
232、抗体或其功能部分、衍生物和/或类似物优选地包含能够结合cd137的细胞外部分、阻断cd137与cd137配体结合的可变结构域,以及能够结合pd-l1的细胞外部分、阻断pd-1与pd-l1结合的可变结构域。该抗体或其功能部分、衍生物和/或类似物中结合pd-l1的细胞外部分的可变结构域优选地包含具有以下项的vh之一的cdr3的氨基酸序列或cdr1、cdr2和cdr3的氨基酸序列的vh区:mf5554;mf5576;mf5578;mf9375;mf9376;mf7702;mf5424;mf5561;mf5439;mf5553;mf5594;mf5426;mf5442或mf5361(图3)。在一个优选的实施方案中,结合pd-l1的细胞外部分的可变结构域包含具有以下项的vh的氨基酸序列的vh区:mf5554;mf5576;mf5578;mf9375;mf9376;mf7702;mf5424;mf5561;mf5439;mf5553;mf5594;mf5426;mf5442或mf5361,其关于所示mf的vh的氨基酸序列具有最多15个,优选地0个、1个、2个、3个、4个、5个、6个、7个、8个、9个或10个,并且优选地具有0个、1个、2个、3个、4个或5个氨基酸插入、缺失、取代或它们的组合。
233、该抗体或其功能部分、衍生物和/或类似物中结合cd137的细胞外部分的可变结构域优选地包含具有以下项的vh之一的cdr3的氨基酸序列或cdr1、cdr2和cdr3的氨基酸序列的vh区:mf6754;mf6763;mf6785;或mf6797(图3)。在一个优选的实施方案中,结合cd137的细胞外部分的可变结构域包含具有以下项的vh的氨基酸序列的vh区:mf6754;mf6763;mf6785;或mf6797,其关于所示mf的vh的氨基酸序列具有最多15个,优选地0个、1个、2个、3个、4个、5个、6个、7个、8个、9个或10个,并且优选地具有0个、1个、2个、3个、4个或5个氨基酸插入、缺失、取代或它们的组合。氨基酸插入、缺失、取代或它们的组合(如果有的话)优选地不处于cdr区的氨基酸序列中。该抗体或功能部分、衍生物和/或类似物中特别优选的组合是包含以下项所示序列或其变体的可变结构域的组合:mf6797和mf7702;mf6763和mf7702;mf6785和mf7702;mf6797和mf5553;mf6763和mf5553;mf6785和mf5553;mf6754和mf5424;mf6763和mf5561;mf6785和mf5439;mf6797和mf5553;mf6744和mf5594;mf6744和mf5361;mf6783和mf5361;或mf6783和mf5594。
234、如本文所述的抗体或其功能部分、衍生物和/或类似物优选地包含:
235、-结合cd137的可变结构域,包含具有mf6754的vh的cdr3的氨基酸序列或cdr1、cdr2和cdr3的氨基酸序列的vh区;以及
236、-结合pd-l1的可变结构域,包含具有以下项的vh的cdr3的氨基酸序列或cdr1、cdr2和cdr3的氨基酸序列的vh区:mf5554mf5576;mf5578;mf9375;mf9376;mf7702;mf5594;mf5424;mf5426;mf5553;mf5442;mf5561;mf5439或mf5361(图3)。
237、抗体或其功能部分、衍生物和/或类似物优选地包含:
238、-结合cd137的可变结构域,包含具有mf6754的vh的氨基酸序列的vh区,其关于mf6754的vh的氨基酸序列具有最多15个,优选地0个、1个、2个、3个、4个、5个、6个、7个、8个、9个或10个,并且优选地具有0个、1个、2个、3个、4个或5个氨基酸插入、缺失、取代或它们的组合;以及
239、-结合pd-l1的可变结构域,包含具有以下项的vh的氨基酸序列的vh区:mf5554;mf5576;mf5578;mf9375;mf9376;mf7702;mf5594;mf5424;mf5426;mf5553;mf5442;mf5561;mf5439或mf5361(图3),其关于所示mf的vh的氨基酸序列具有最多15个,优选地0个、1个、2个、3个、4个、5个、6个、7个、8个、9个或10个,并且优选地具有0个、1个、2个、3个、4个或5个氨基酸插入、缺失、取代或它们的组合。
240、如本文所述的抗体或其功能部分、衍生物和/或类似物优选地包含:
241、-结合cd137的可变结构域,包含具有mf6763的vh的cdr3的氨基酸序列或cdr1、cdr2和cdr3的氨基酸序列的vh区;以及
242、-结合pd-l1的可变结构域,包含具有以下项的vh的cdr3的氨基酸序列或cdr1、cdr2和cdr3的氨基酸序列的vh区:mf5554mf5576;mf5578;mf9375;mf9376;mf7702;mf5594;mf5424;mf5426;mf5553;mf5442;mf5561;mf5439或mf5361(图3)。
243、抗体或其功能部分、衍生物和/或类似物优选地包含:
244、-结合cd137的可变结构域,包含具有mf6763的vh的氨基酸序列的vh区,其关于mf6763的vh的氨基酸序列具有最多15个,优选地0个、1个、2个、3个、4个、5个、6个、7个、8个、9个或10个,并且优选地具有0个、1个、2个、3个、4个或5个氨基酸插入、缺失、取代或它们的组合;以及
245、-结合pd-l1的可变结构域,包含具有以下项的vh的氨基酸序列的vh区:mf5554;mf5576;mf5578;mf9375;mf9376;mf7702;mf5594;mf5424;mf5426;mf5553;mf5442;mf5561;mf5439或mf5361(图3),其关于所示mf的vh的氨基酸序列具有最多15个,优选地0个、1个、2个、3个、4个、5个、6个、7个、8个、9个或10个,并且优选地具有0个、1个、2个、3个、4个或5个氨基酸插入、缺失、取代或它们的组合。
246、如本文所述的抗体或其功能部分、衍生物和/或类似物优选地包含:
247、-结合cd137的可变结构域,包含具有mf6785的vh的cdr3的氨基酸序列或cdr1、cdr2和cdr3的氨基酸序列的vh区;以及
248、-结合pd-l1的可变结构域,包含具有以下项的vh的cdr3的氨基酸序列或cdr1、cdr2和cdr3的氨基酸序列的vh区:mf5554mf5576;mf5578;mf9375;mf9376;mf7702;mf5594;mf5424;mf5426;mf5553;mf5442;mf5561;mf5439或mf5361(图3)。
249、抗体或其功能部分、衍生物和/或类似物优选地包含:
250、-结合cd137的可变结构域,包含具有mf6785的vh的氨基酸序列的vh区,其关于mf6785的vh的氨基酸序列具有最多15个,优选地0个、1个、2个、3个、4个、5个、6个、7个、8个、9个或10个,并且优选地具有0个、1个、2个、3个、4个或5个氨基酸插入、缺失、取代或它们的组合;以及
251、-结合pd-l1的可变结构域,包含具有以下项的vh的氨基酸序列的vh区:mf5554;mf5576;mf5578;mf9375;mf9376;mf7702;mf5594;mf5424;mf5426;mf5553;mf5442;mf5561;mf5439或mf5361(图3),其关于所示mf的vh的氨基酸序列具有最多15个,优选地0个、1个、2个、3个、4个、5个、6个、7个、8个、9个或10个,并且优选地具有0个、1个、2个、3个、4个或5个氨基酸插入、缺失、取代或它们的组合。
252、如本文所述的抗体或其功能部分、衍生物和/或类似物优选地包含:
253、-结合cd137的可变结构域,包含具有mf6797的vh的cdr3的氨基酸序列或cdr1、cdr2和cdr3的氨基酸序列的vh区;以及
254、-结合pd-l1的可变结构域,包含具有以下项的vh的cdr3的氨基酸序列或cdr1、cdr2和cdr3的氨基酸序列的vh区:mf5554mf5576;mf5578;mf9375;mf9376;mf7702;mf5594;mf5424;mf5426;mf5553;mf5442;mf5561;mf5439或mf5361(图3)。
255、抗体或其功能部分、衍生物和/或类似物优选地包含:
256、-结合cd137的可变结构域,包含具有mf6797的vh的氨基酸序列的vh区,其关于mf6797的vh的氨基酸序列具有最多15个,优选地0个、1个、2个、3个、4个、5个、6个、7个、8个、9个或10个,并且优选地具有0个、1个、2个、3个、4个或5个氨基酸插入、缺失、取代或它们的组合;以及
257、-结合pd-l1的可变结构域,包含具有以下项的vh的氨基酸序列的vh区:mf5554;mf5576;mf5578;mf9375;mf9376;mf7702;mf5594;mf5424;mf5426;mf5553;mf5442;mf5561;mf5439或mf5361(图3),其关于所示mf的vh的氨基酸序列具有最多15个,优选地0个、1个、2个、3个、4个、5个、6个、7个、8个、9个或10个,并且优选地具有0个、1个、2个、3个、4个或5个氨基酸插入、缺失、取代或它们的组合。
258、如示例中所示,含有与不同的结合cd137的可变结构域(包括mf6754、mf6763、mf6785和mf6797)组合的基于mf5553的结合pd-l1的可变结构域的抗体提供特别好的t细胞活化结果。
259、因此,还提供了双特异性抗体或其功能部分、衍生物和/或类似物,其包含:
260、-结合cd137的可变结构域,包含具有mf6754的vh的cdr3区的氨基酸序列的vh区;以及
261、-结合pd-l1的可变结构域,包含具有mf5553的vh的cdr3区的氨基酸序列的vh区。
262、还提供了双特异性抗体或其功能部分、衍生物和/或类似物,其包含:
263、-结合cd137的可变结构域,包含具有mf6754的vh的cdr1、cdr2和cdr3区的氨基酸序列的vh区;以及
264、-结合pd-l1的可变结构域,包含具有mf5553的vh的cdr1、cdr2和cdr3区的氨基酸序列的vh区。
265、还提供了双特异性抗体或其功能部分、衍生物和/或类似物,其包含:
266、-结合cd137的可变结构域,包含具有mf6754的vh的氨基酸序列的vh区,其关于mf6754的vh的氨基酸序列具有最多15个,优选地0个、1个、2个、3个、4个、5个、6个、7个、8个、9个或10个,并且优选地具有0个、1个、2个、3个、4个或5个氨基酸插入、缺失、取代或它们的组合;以及
267、-结合pd-l1的可变结构域,包含具有mf5553的vh的氨基酸序列的vh区,其关于mf5553的vh的氨基酸序列具有最多15个,优选地0个、1个、2个、3个、4个、5个、6个、7个、8个、9个或10个,并且优选地具有0个、1个、2个、3个、4个或5个氨基酸插入、缺失、取代或它们的组合。
268、还提供了双特异性抗体或其功能部分、衍生物和/或类似物,其包含:
269、-结合cd137的可变结构域,包含具有mf6763的vh的cdr3区的氨基酸序列的vh区;以及
270、-结合pd-l1的可变结构域,包含具有mf5553的vh的cdr3区的氨基酸序列的vh区。
271、还提供了双特异性抗体或其功能部分、衍生物和/或类似物,其包含:
272、-结合cd137的可变结构域,包含具有mf6763的vh的cdr1、cdr2和cdr3区的氨基酸序列的vh区;以及
273、-结合pd-l1的可变结构域,包含具有mf5553的vh的cdr1、cdr2和cdr3区的氨基酸序列的vh区。
274、还提供了双特异性抗体或其功能部分、衍生物和/或类似物,其包含:
275、-结合cd137的可变结构域,包含具有mf6763的vh的氨基酸序列的vh区,其关于mf6763的vh的氨基酸序列具有最多15个,优选地0个、1个、2个、3个、4个、5个、6个、7个、8个、9个或10个,并且优选地具有0个、1个、2个、3个、4个或5个氨基酸插入、缺失、取代或它们的组合;以及
276、-结合pd-l1的可变结构域,包含具有mf5553的vh的氨基酸序列的vh区,其关于mf5553的vh的氨基酸序列具有最多15个,优选地0个、1个、2个、3个、4个、5个、6个、7个、8个、9个或10个,并且优选地具有0个、1个、2个、3个、4个或5个氨基酸插入、缺失、取代或它们的组合。
277、还提供了双特异性抗体或其功能部分、衍生物和/或类似物,其包含:
278、-结合cd137的可变结构域,包含具有mf6785的vh的cdr3区的氨基酸序列的vh区;以及
279、-结合pd-l1的可变结构域,包含具有mf5553的vh的cdr3区的氨基酸序列的vh区。
280、还提供了双特异性抗体或其功能部分、衍生物和/或类似物,其包含:
281、-结合cd137的可变结构域,包含具有mf6785的vh的cdr1、cdr2和cdr3区的氨基酸序列的vh区;以及
282、-结合pd-l1的可变结构域,包含具有mf5553的vh的cdr1、cdr2和cdr3区的氨基酸序列的vh区。
283、还提供了双特异性抗体或其功能部分、衍生物和/或类似物,其包含:
284、-结合cd137的可变结构域,包含具有mf6785的vh的氨基酸序列的vh区,其关于mf6785的vh的氨基酸序列具有最多15个,优选地0个、1个、2个、3个、4个、5个、6个、7个、8个、9个或10个,并且优选地具有0个、1个、2个、3个、4个或5个氨基酸插入、缺失、取代或它们的组合;以及
285、-结合pd-l1的可变结构域,包含具有mf5553的vh的氨基酸序列的vh区,其关于mf5553的vh的氨基酸序列具有最多15个,优选地0个、1个、2个、3个、4个、5个、6个、7个、8个、9个或10个,并且优选地具有0个、1个、2个、3个、4个或5个氨基酸插入、缺失、取代或它们的组合。
286、还提供了双特异性抗体或其功能部分、衍生物和/或类似物,其包含:
287、-结合cd137的可变结构域,包含具有mf6797的vh的cdr3区的氨基酸序列的vh区;以及
288、-结合pd-l1的可变结构域,包含具有mf5553的vh的cdr3区的氨基酸序列的vh区。
289、还提供了双特异性抗体或其功能部分、衍生物和/或类似物,其包含:
290、-结合cd137的可变结构域,包含具有mf6797的vh的cdr1、cdr2和cdr3区的氨基酸序列的vh区;以及
291、-结合pd-l1的可变结构域,包含具有mf5553的vh的cdr1、cdr2和cdr3区的氨基酸序列的vh区。
292、还提供了双特异性抗体或其功能部分、衍生物和/或类似物,其包含:
293、-结合cd137的可变结构域,包含具有mf6797的vh的氨基酸序列的vh区,其关于mf6797的vh的氨基酸序列具有最多15个,优选地0个、1个、2个、3个、4个、5个、6个、7个、8个、9个或10个,并且优选地具有0个、1个、2个、3个、4个或5个氨基酸插入、缺失、取代或它们的组合;以及
294、-结合pd-l1的可变结构域,包含具有mf5553的vh的氨基酸序列的vh区,其关于mf5553的vh的氨基酸序列具有最多15个,优选地0个、1个、2个、3个、4个、5个、6个、7个、8个、9个或10个,并且优选地具有0个、1个、2个、3个、4个或5个氨基酸插入、缺失、取代或它们的组合。
295、此外在示例中示出,含有与不同的结合cd137的可变结构域(包括mf6763、mf6785和mf6797)组合的基于mf7702的结合pd-l1的可变结构域的抗体提供特别好的t细胞活化结果。
296、因此,还提供了双特异性抗体或其功能部分、衍生物和/或类似物,其包含:
297、-结合cd137的可变结构域,包含具有mf6797的vh的cdr3区的氨基酸序列的vh区;以及
298、-结合pd-l1的可变结构域,包含具有mf7702的vh的cdr3区的氨基酸序列的vh区。
299、还提供了双特异性抗体或其功能部分、衍生物和/或类似物,其包含:
300、-结合cd137的可变结构域,包含具有mf6797的vh的cdr1、cdr2和cdr3区的氨基酸序列的vh区;以及
301、-结合pd-l1的可变结构域,包含具有mf7702的vh的cdr1、cdr2和cdr3区的氨基酸序列的vh区。
302、还提供了双特异性抗体或其功能部分、衍生物和/或类似物,其包含:
303、-结合cd137的可变结构域,包含具有mf6797的vh的氨基酸序列的vh区,其关于mf6797的vh的氨基酸序列具有最多15个,优选地0个、1个、2个、3个、4个、5个、6个、7个、8个、9个或10个,并且优选地具有0个、1个、2个、3个、4个或5个氨基酸插入、缺失、取代或它们的组合;以及
304、-结合pd-l1的可变结构域,包含具有mf7702的vh的氨基酸序列的vh区,其关于mf7702的vh的氨基酸序列具有最多15个,优选地0个、1个、2个、3个、4个、5个、6个、7个、8个、9个或10个,并且优选地具有0个、1个、2个、3个、4个或5个氨基酸插入、缺失、取代或它们的组合。
305、还提供了双特异性抗体或其功能部分、衍生物和/或类似物,其包含:
306、-结合cd137的可变结构域,包含具有mf6763的vh的cdr3区的氨基酸序列的vh区;以及
307、-结合pd-l1的可变结构域,包含具有mf7702的vh的cdr3区的氨基酸序列的vh区。
308、还提供了双特异性抗体或其功能部分、衍生物和/或类似物,其包含:
309、-结合cd137的可变结构域,包含具有mf6763的vh的cdr1、cdr2和cdr3区的氨基酸序列的vh区;以及
310、-结合pd-l1的可变结构域,包含具有mf7702的vh的cdr1、cdr2和cdr3区的氨基酸序列的vh区。
311、还提供了双特异性抗体或其功能部分、衍生物和/或类似物,其包含:
312、-结合cd137的可变结构域,包含具有mf6763的vh的氨基酸序列的vh区,其关于mf6763的vh的氨基酸序列具有最多15个,优选地0个、1个、2个、3个、4个、5个、6个、7个、8个、9个或10个,并且优选地具有0个、1个、2个、3个、4个或5个氨基酸插入、缺失、取代或它们的组合;以及
313、-结合pd-l1的可变结构域,包含具有mf7702的vh的氨基酸序列的vh区,其关于mf7702的vh的氨基酸序列具有最多15个,优选地0个、1个、2个、3个、4个、5个、6个、7个、8个、9个或10个,并且优选地具有0个、1个、2个、3个、4个或5个氨基酸插入、缺失、取代或它们的组合。
314、还提供了双特异性抗体或其功能部分、衍生物和/或类似物,其包含:
315、-结合cd137的可变结构域,包含具有mf6785的vh的cdr3区的氨基酸序列的vh区;以及
316、-结合pd-l1的可变结构域,包含具有mf7702的vh的cdr3区的氨基酸序列的vh区。
317、还提供了双特异性抗体或其功能部分、衍生物和/或类似物,包含:
318、-结合cd137的可变结构域,包含具有mf6785的vh的cdr1、cdr2和cdr3区的氨基酸序列的vh区;以及
319、-结合pd-l1的可变结构域,包含具有mf7702的vh的cdr1、cdr2和cdr3区的氨基酸序列的vh区。
320、还提供了双特异性抗体或其功能部分、衍生物和/或类似物,其包含:
321、-结合cd137的可变结构域,包含具有mf6785的vh的氨基酸序列的vh区,其关于mf6785的vh的氨基酸序列具有最多15个,优选地0个、1个、2个、3个、4个、5个、6个、7个、8个、9个或10个,并且优选地具有0个、1个、2个、3个、4个或5个氨基酸插入、缺失、取代或它们的组合;以及
322、-结合pd-l1的可变结构域,包含具有mf7702的vh的氨基酸序列的vh区,其关于mf7702的vh的氨基酸序列具有最多15个,优选地0个、1个、2个、3个、4个、5个、6个、7个、8个、9个或10个,并且优选地具有0个、1个、2个、3个、4个或5个氨基酸插入、缺失、取代或它们的组合。
323、此外在示例中示出,含有基于mf6744的结合cd137的可变结构域以及基于mf5594的结合pd-l1的可变结构域的双特异性抗体提供特别好的t细胞活化;参见例如图14-图16。重要地,相比于基于抗体优卢单抗(urelumab)的抗体,此类抗体具有较强的t细胞活化潜能。
324、因此,还提供了双特异性抗体或其功能部分、衍生物和/或类似物,其包含:
325、-结合cd137的可变结构域,包含具有mf6744的vh的cdr3区的氨基酸序列的vh区;以及
326、-结合pd-l1的可变结构域,包含具有mf5594的vh的cdr3区的氨基酸序列的vh区。
327、还提供了双特异性抗体或其功能部分、衍生物和/或类似物,其包含:
328、-结合cd137的可变结构域,包含具有mf6744的vh的cdr1、cdr2和cdr3区的氨基酸序列的vh区;以及
329、-结合pd-l1的可变结构域,包含具有mf5594的vh的cdr1、cdr2和cdr3区的氨基酸序列的vh区。
330、还提供了双特异性抗体或其功能部分、衍生物和/或类似物,其包含:
331、-结合cd137的可变结构域,包含具有mf6744的vh的氨基酸序列的vh区,其关于mf6744的vh的氨基酸序列具有最多15个,优选地0个、1个、2个、3个、4个、5个、6个、7个、8个、9个或10个,并且优选地具有0个、1个、2个、3个、4个或5个氨基酸插入、缺失、取代或它们的组合;以及
332、-结合pd-l1的可变结构域,包含具有mf5594的vh的氨基酸序列的vh区,其关于mf5594的vh的氨基酸序列具有最多15个,优选地0个、1个、2个、3个、4个、5个、6个、7个、8个、9个或10个,并且优选地具有0个、1个、2个、3个、4个或5个氨基酸插入、缺失、取代或它们的组合。
333、此外在示例中示出,含有基于mf6744的结合cd137的可变结构域以及基于mf5361的结合pd-l1的可变结构域的双特异性抗体提供特别好的t细胞活化;参见例如图14-图16。重要地,相比于基于抗体优卢单抗的抗体,此类抗体具有较强的t细胞活化潜能。
334、因此,还提供了双特异性抗体或其功能部分、衍生物和/或类似物,其包含:
335、-结合cd137的可变结构域,包含具有mf6744的vh的cdr3区的氨基酸序列的vh区;以及
336、-结合pd-l1的可变结构域,包含具有mf5361的vh的cdr3区的氨基酸序列的vh区。
337、还提供了双特异性抗体或其功能部分、衍生物和/或类似物,其包含:
338、-结合cd137的可变结构域,包含具有mf6744的vh的cdr1、cdr2和cdr3区的氨基酸序列的vh区;以及
339、-结合pd-l1的可变结构域,包含具有mf5361的vh的cdr1、cdr2和cdr3区的氨基酸序列的vh区。
340、还提供了双特异性抗体或其功能部分、衍生物和/或类似物,其包含:
341、-结合cd137的可变结构域,包含具有mf6744的vh的氨基酸序列的vh区,其关于mf6744的vh的氨基酸序列具有最多15个,优选地0个、1个、2个、3个、4个、5个、6个、7个、8个、9个或10个,并且优选地具有0个、1个、2个、3个、4个或5个氨基酸插入、缺失、取代或它们的组合;以及
342、-结合pd-l1的可变结构域,包含具有mf5361的vh的氨基酸序列的vh区,其关于mf5361的vh的氨基酸序列具有最多15个,优选地0个、1个、2个、3个、4个、5个、6个、7个、8个、9个或10个,并且优选地具有0个、1个、2个、3个、4个或5个氨基酸插入、缺失、取代或它们的组合。
343、此外在示例中示出,含有基于mf6783的结合cd137的可变结构域以及基于mf5361的结合pd-l1的可变结构域的双特异性抗体提供特别好的t细胞活化;参见例如图14-图15。重要地,相比于基于抗体优卢单抗的抗体,此类抗体具有较强的t细胞活化潜能。
344、因此,还提供了双特异性抗体或其功能部分、衍生物和/或类似物,其包含:
345、-结合cd137的可变结构域,包含具有mf6783的vh的cdr3区的氨基酸序列的vh区;以及
346、-结合pd-l1的可变结构域,包含具有mf5361的vh的cdr3区的氨基酸序列的vh区。
347、还提供了双特异性抗体或其功能部分、衍生物和/或类似物,其包含:
348、-结合cd137的可变结构域,包含具有mf6783的vh的cdr1、cdr2和cdr3区的氨基酸序列的vh区;以及
349、-结合pd-l1的可变结构域,包含具有mf5361的vh的cdr1、cdr2和cdr3区的氨基酸序列的vh区。
350、还提供了双特异性抗体或其功能部分、衍生物和/或类似物,其包含:
351、-结合cd137的可变结构域,包含具有mf6783的vh的氨基酸序列的vh区,其关于mf6783的vh的氨基酸序列具有最多15个,优选地0个、1个、2个、3个、4个、5个、6个、7个、8个、9个或10个,并且优选地具有0个、1个、2个、3个、4个或5个氨基酸插入、缺失、取代或它们的组合;以及
352、-结合pd-l1的可变结构域,包含具有mf5361的vh的氨基酸序列的vh区,其关于mf5361的vh的氨基酸序列具有最多15个,优选地0个、1个、2个、3个、4个、5个、6个、7个、8个、9个或10个,并且优选地具有0个、1个、2个、3个、4个或5个氨基酸插入、缺失、取代或它们的组合。
353、此外在示例中示出,含有基于mf6783的结合cd137的可变结构域以及基于mf5594的结合pd-l1的可变结构域的双特异性抗体提供特别好的t细胞活化;参见例如图14-图15。
354、因此,还提供了双特异性抗体或其功能部分、衍生物和/或类似物,其包含:
355、-结合cd137的可变结构域,包含具有mf6783的vh的cdr3区的氨基酸序列的vh区;以及
356、-结合pd-l1的可变结构域,包含具有mf5594的vh的cdr3区的氨基酸序列的vh区。
357、还提供了双特异性抗体或其功能部分、衍生物和/或类似物,其包含:
358、-结合cd137的可变结构域,包含具有mf6783的vh的cdr1、cdr2和cdr3区的氨基酸序列的vh区;以及
359、-结合pd-l1的可变结构域,包含具有mf5594的vh的cdr1、cdr2和cdr3区的氨基酸序列的vh区。
360、还提供了双特异性抗体或其功能部分、衍生物和/或类似物,其包含:
361、-结合cd137的可变结构域,包含具有mf6783的vh的氨基酸序列的vh区,其关于mf6783的vh的氨基酸序列具有最多15个,优选地0个、1个、2个、3个、4个、5个、6个、7个、8个、9个或10个,并且优选地具有0个、1个、2个、3个、4个或5个氨基酸插入、缺失、取代或它们的组合;以及
362、-结合pd-l1的可变结构域,包含具有mf5594的vh的氨基酸序列的vh区,其关于mf5594的vh的氨基酸序列具有最多15个,优选地0个、1个、2个、3个、4个、5个、6个、7个、8个、9个或10个,并且优选地具有0个、1个、2个、3个、4个或5个氨基酸插入、缺失、取代或它们的组合。
363、此外,在示例中示出,含有基于mf6744的结合cd137的可变结构域以及基于mf5361的结合pd-l1的可变结构域的双特异性抗体,连同含有基于mf6744的结合cd137的可变结构域以及基于mf5594的结合pd-l1的可变结构域的双特异性抗体的组合(用作双重双特异性,例如实施方案)提供优异的t细胞活化(参见图14-图16)并且优于基于优卢单抗的抗体。
364、因此,还提供了混合物或成套试剂盒,其包含:
365、-第一双特异性抗体或其功能部分、衍生物和/或类似物,包含含有具有mf6744的vh的cdr3区的氨基酸序列的vh区的结合cd137的可变结构域,以及含有具有mf5594的vh的cdr3区的氨基酸序列的vh区的结合pd-l1的可变结构域;以及
366、-第二双特异性抗体或其功能部分、衍生物和/或类似物,包含含有具有mf6744的vh的cdr3区的氨基酸序列的vh区的结合cd137的可变结构域,以及含有具有mf5361的vh的cdr3区的氨基酸序列的vh区的结合pd-l1的可变结构域。
367、还提供了混合物或成套试剂盒,其包含:
368、-第一双特异性抗体或其功能部分、衍生物和/或类似物,包含含有具有mf6744的vh的cdr1、cdr2和cdr3区的氨基酸序列的vh区的结合cd137的可变结构域,以及含有具有mf5594的vh的cdr1、cdr2和cdr3区的氨基酸序列的vh区的结合pd-l1的可变结构域;以及
369、-第二双特异性抗体或其功能部分、衍生物和/或类似物,包含含有具有mf6744的vh的cdr1、cdr2和cdr3区的氨基酸序列的vh区的结合cd137的可变结构域,以及含有具有mf5361的vh的cdr1、cdr2和cdr3区的氨基酸序列的vh区的结合pd-l1的可变结构域。
370、还提供了混合物或成套试剂盒,其包含:
371、-第一双特异性抗体或其功能部分、衍生物和/或类似物,包含含有具有mf6744的vh的氨基酸序列的vh区的结合cd137的可变结构域,其关于mf6744的vh的氨基酸序列具有最多15个,优选地0个、1个、2个、3个、4个、5个、6个、7个、8个、9个或10个,并且优选地具有0个、1个、2个、3个、4个或5个氨基酸插入、缺失、取代或它们的组合,以及含有具有mf5594的vh的氨基酸序列的vh区的结合pd-l1的可变结构域,其关于mf5594的vh的氨基酸序列具有最多15个,优选地0个、1个、2个、3个、4个、5个、6个、7个、8个、9个或10个,并且优选地具有0个、1个、2个、3个、4个或5个氨基酸插入、缺失、取代或它们的组合;以及
372、-第二双特异性抗体或其功能部分、衍生物和/或类似物,包含含有具有mf6744的vh的氨基酸序列的vh区的结合cd137的可变结构域,其关于mf6744的vh的氨基酸序列具有最多15个,优选地0个、1个、2个、3个、4个、5个、6个、7个、8个、9个或10个,并且优选地具有0个、1个、2个、3个、4个或5个氨基酸插入、缺失、取代或它们的组合,以及含有具有mf5361的vh的氨基酸序列的vh区的结合pd-l1的可变结构域,其关于mf5361的vh的氨基酸序列具有最多15个,优选地0个、1个、2个、3个、4个、5个、6个、7个、8个、9个或10个,并且优选地具有0个、1个、2个、3个、4个或5个氨基酸插入、缺失、取代或它们的组合。
373、此外,在示例中示出,相比于基于优卢单抗的抗体,含有基于mf6744的结合cd137的可变结构域以及基于mf5361的结合pd-l1的可变结构域的双特异性抗体,连同具有基于mf6783的结合cd137的可变结构域以及基于mf5594的结合pd-l1的可变结构域的双特异性抗体的组合(用作双重双特异性,例如实施方案)提供优异的t细胞活化。
374、因此,还提供了混合物或成套试剂盒,其包含:
375、-第一双特异性抗体或其功能部分、衍生物和/或类似物,包含含有具有mf6783的vh的cdr3区的氨基酸序列的vh区的结合cd137的可变结构域,以及含有具有mf5594的vh的cdr3区的氨基酸序列的vh区的结合pd-l1的可变结构域;以及
376、-第二双特异性抗体或其功能部分、衍生物和/或类似物,包含含有具有mf6744的vh的cdr3区的氨基酸序列的vh区的结合cd137的可变结构域,以及含有具有mf5361的vh的cdr3区的氨基酸序列的vh区的结合pd-l1的可变结构域。
377、还提供了混合物或成套试剂盒,其包含:
378、-第一双特异性抗体或其功能部分、衍生物和/或类似物,包含含有具有mf6783的vh的cdr1、cdr2和cdr3区的氨基酸序列的vh区的结合cd137的可变结构域,以及含有具有mf5594的vh的cdr1、cdr2和cdr3区的氨基酸序列的vh区的结合pd-l1的可变结构域;以及
379、-第二双特异性抗体或其功能部分、衍生物和/或类似物,包含含有具有mf6744的vh的cdr1、cdr2和cdr3区的氨基酸序列的vh区的结合cd137的可变结构域,以及含有具有mf5361的vh的cdr1、cdr2和cdr3区的氨基酸序列的vh区的结合pd-l1的可变结构域。
380、还提供了混合物或成套试剂盒,其包含:
381、-第一双特异性抗体或其功能部分、衍生物和/或类似物,包含含有具有mf6783的vh的氨基酸序列的vh区的结合cd137的可变结构域,其关于mf6783的vh的氨基酸序列具有最多15个,优选地0个、1个、2个、3个、4个、5个、6个、7个、8个、9个或10个,并且优选地具有0个、1个、2个、3个、4个或5个氨基酸插入、缺失、取代或它们的组合,以及含有具有mf5594的vh的氨基酸序列的vh区的结合pd-l1的可变结构域,其关于mf5594的vh的氨基酸序列具有最多15个,优选地0个、1个、2个、3个、4个、5个、6个、7个、8个、9个或10个,并且优选地具有0个、1个、2个、3个、4个或5个氨基酸插入、缺失、取代或它们的组合;以及
382、-第二双特异性抗体或其功能部分、衍生物和/或类似物,包含含有具有mf6744的vh的氨基酸序列的vh区的结合cd137的可变结构域,其关于mf6744的vh的氨基酸序列具有最多15个,优选地0个、1个、2个、3个、4个、5个、6个、7个、8个、9个或10个,并且优选地具有0个、1个、2个、3个、4个或5个氨基酸插入、缺失、取代或它们的组合,以及含有具有mf5361的vh的氨基酸序列的vh区的结合pd-l1的可变结构域,其关于mf5361的vh的氨基酸序列具有最多15个,优选地0个、1个、2个、3个、4个、5个、6个、7个、8个、9个或10个,并且优选地具有0个、1个、2个、3个、4个或5个氨基酸插入、缺失、取代或它们的组合。
383、还在示例中示出,具有良好的t细胞活化特性的mf6797的cd137特异性vh的结合与包含如图42所示的cd137氨基酸序列的arg66、gly70和phe72的氨基酸的存在相关联。
384、本发明因此提供了能够结合至cd137的分离、合成或组合的抗体,或其功能部分、衍生物和/或类似物,其中所述抗体或功能部分、衍生物或类似物与cd137的结合与包含如图42所示的cd137氨基酸序列的arg66、gly70和phe72的氨基酸的存在相关联。所述抗体或功能部分、衍生物或类似物与cd137的结合还优选地与包含如图42所示的cd137氨基酸序列的val71的氨基酸相关联。
385、术语“arg66”是指如图42所示的cd137序列的第66位精氨酸残基。术语“gly70”是指如图42所示的cd137序列的第70位甘氨酸残基。术语“val71”是指如图42所示的cd137序列的第71位缬氨酸残基。术语“phe72”是指如图42所示的cd137序列的第72位苯丙氨酸残基。
386、抗体或功能部分、衍生物或类似物与cd137的结合与所述氨基酸残基的存在相关联,如果当这些残基中的任一者被丙氨酸取代时,抗体或功能部分、衍生物或类似物与所得cd137蛋白质的结合下降。
387、一些实施方案提供了能够结合至cd137的分离、合成或组合的抗体,或其功能部分、衍生物和/或类似物,其中所述抗体或功能部分、衍生物或类似物特异性地结合如图42所示的cd137氨基酸序列的氨基酸arg66、gly70和phe72。所述抗体或功能部分、衍生物或类似物还特异性地结合如图42所示的cd137氨基酸序列的氨基酸val71。
388、一些优选的实施方案提供了能够结合至cd137且结合至pd-l1并且具有基于mf6797的结合cd137的可变结构域以及基于mf7702的结合pd-l1的可变结构域的双特异性抗体、或其功能部分、衍生物和/或类似物。具有特别良好的t细胞活化特性的此类双特异性抗体与cd137和pd-l1的结合与包含以上所提及cd137氨基酸残基的氨基酸相关联。
389、现在已鉴定出以上所提及的cd137氨基酸残基,已经有可能生成或选择出特异性地结合这些氨基酸残基的抗体或其变体。生成和/或选择特异性结合特定氨基酸残基的结合分子可使用本领域公知的方法进行,如例如通过用包含含有目标氨基酸残基的特定结构域的抗原片段来免疫能够产生抗体的转基因非人动物。另选地,通过针对结合至所鉴定的氨基酸残基的噬菌体筛选抗体噬菌体展示库。
390、还提供了与抗体pb17311竞争性结合cd137和/或pd-l1的抗体或其变体。例如采用使包含cd137和/或pd-l1的细胞与pb17311以及与候选抗体或其变体一起温育竞争测定法,鉴定出竞争性抗体或其变体。相比于在不具有候选抗体或其变体情况下使包含cd137和/或pd-l1的细胞与pb17311一起温育的对照,能够减少所结合pb17311的量的候选抗体或其变体是竞争性抗体或变体。
391、一些实施方案提供了与抗体pb17311竞争性结合cd137和/或pd-l1的分离、合成或重组的抗体,或其功能部分、衍生物和/或类似物。
392、一些实施方案提供了分离、合成或重组的抗体,或其功能部分、衍生物和/或类似物,其与抗体pb17311竞争性结合如图42所示的cd137氨基酸序列的氨基酸arg66、gly70和phe72,更优选地竞争性结合如图42所示的cd137氨基酸序列的氨基酸arg66、gly70、val71和phe72。
393、一些实施方案提供了与抗体pb17309竞争性结合cd137和/或pd-l1的分离、合成或重组的抗体,或其功能部分、衍生物和/或类似物。
394、一些实施方案提供了与抗体pb17310竞争性结合cd137和/或pd-l1的分离、合成或重组的抗体,或其功能部分、衍生物和/或类似物。
395、例如使用竞争测定法分离出与pb17309或pb17310竞争性结合cd137和/或pd-l1的抗体或其变体,其中将不存在候选抗体或其变体时pb17309或pb17310与包含cd137和/或pd-l1的细胞的结合与存在所述候选抗体或其变体时pb17309或pb17310与包含cd137和/或pd-l1的细胞的结合进行比较。相比于在不具有所述候选抗体或其变体情况下使包含cd137和/或pd-l1的细胞与pb17309或pb17310一起温育的对照,能够减少所结合pb17309或pb17310的量的候选抗体或其变体被鉴定为竞争性抗体或变体。
396、如本文所述的ox40×pd-l1双特异性抗体或其功能部分、衍生物和/或类似物优选地包含:
397、-结合ox40的可变结构域,包含具有以下项的vh的cdr3的氨基酸序列或cdr1、cdr2和cdr3的氨基酸序列的vh区:mf6629mf6630;mf6637;mf6643;mf6645;mf6648;mf6655;mf6658;mf6660;mf6675;mf6686;mf6690;mf6692;mf6700;mf6706;mf6714;mf6721;mf6722;mf6724;mf6728;mf6729;mf6826;mf6940;mf6942;mf6943;mf6944;mf6947;mf6949;mf7331;mf7332;mf7334;mf7341;mf7345;mf7350;mf7351;mf7352;mf7353;mf7356;mf7358;mf7365;mf7366;mf7371;mf7372;mf7374;mf7378;mf7382;mf7383;mf7394;mf7395;或mf7397;以及
398、-结合pd-l1的可变结构域。结合pd-l1的可变结构域优选地包含具有如图3所示的pd-l1特异性vh的vh的cdr3的氨基酸序列或cdr1、cdr2和cdr3的氨基酸序列的vh区。在一个优选的实施方案中,pd-l1特异性vh如mf5594;mf5424;mf5426;mf5553;mf5442;mf5561;mf5426;或mf5439所示(图3)。
399、抗体或其功能部分、衍生物和/或类似物优选地包含:
400、-结合ox40的可变结构域,包含具有以下项的vh的氨基酸序列的vh区:mf6629;mf6630;mf6637;mf6643;mf6645;mf6648;mf6655;mf6658;mf6660;mf6675;mf6686;mf6690;mf6692;mf6700;mf6706;mf6714;mf6721;mf6722;mf6724;mf6728;mf6729;mf6826;mf6940;mf6942;mf6943;mf6944;mf6947;mf6949;mf7331;mf7332;mf7334;mf7341;mf7345;mf7350;mf7351;mf7352;mf7353;mf7356;mf7358;mf7365;mf7366;mf7371;mf7372;mf7374;mf7378;mf7382;mf7383;mf7394;mf7395;或mf7397,其关于所示vh的氨基酸序列具有最多15个,优选地0个、1个、2个、3个、4个、5个、6个、7个、8个、9个或10个,并且优选地具有0个、1个、2个、3个、4个或5个氨基酸插入、缺失、取代或它们的组合;以及
401、-结合pd-l1的可变结构域。结合pd-l1的可变结构域优选地包含具有以下项的vh的氨基酸序列的vh区:mf5594;mf5424;mf5426;mf5553;mf5442;mf5561;mf5426;或mf5439(图3),其关于所示mf(图3)的vh的氨基酸序列具有最多15个,优选地0个、1个、2个、3个、4个、5个、6个、7个、8个、9个或10个,并且优选地具有0个、1个、2个、3个、4个或5个氨基酸插入、缺失、取代或它们的组合。
402、在所提及的h、vh、l和vl区中提及的最多15个,优选地1个、2个、3个、4个、5个、6个、7个、8个、9个或10个,并且优选地1个、2个、3个、4个或5个氨基酸取代优选为保守氨基酸取代,所述插入、缺失、取代或它们的组合优选不在h、vh、l或vl链的cdr3区中,优选地不在vh或vl链的cdr1、cdr2或cdr3区中,并且优选地不在fr4区中。
403、出于清楚和简要描述的目的,本文将特征结构描述为相同或独立实施方案的一部分,然而,应该理解,本发明的范围可包括具有所述特征的全部或一些的组合的实施方案。
404、本发明在以下实施例中进一步说明。这些实施例不限制本发明的范围,而是仅用于澄清本发明。
- 还没有人留言评论。精彩留言会获得点赞!