蛋白质构象变化的生物正交拉曼原位检测方法与流程
本发明涉及蛋白质化学和构象变化的检测,尤其涉及一种利用生物正交拉曼报告基团特异性标记、检测蛋白质,利用生物正交拉曼技术体外检测蛋白构象变化,以及利用生物正交拉曼技术在活细胞原位检测蛋白构象变化的方法。
背景技术:
蛋白质分子的空间构象是其功能、活性的基础。蛋白质对环境和刺激的响应是通过蛋白质或者蛋白复合体其构象的变化进而实现其功能和活性的调节。其中某些重要氨基酸的位置、取向、运动在蛋白质的变构过程中发生显著的变化。因而实现快速、灵敏和高效的检测蛋白质构象变化(特别是在生理条件下原位检测蛋白质的构象变化)对于研究蛋白质结构与功能联系至关重要。
目前,研究蛋白结构的方法有很多种,包括x-射线晶体衍射法、核磁共振法、红外光谱法、荧光光谱法、拉曼光谱法、圆二色谱法、质谱法等。这些方法各有利弊和适用的范围。比如,x射线晶体衍射已经发展了几十年,样品不需要标记,分子量没有限制,数据采集和分析相对成熟,它能够提供原子分辨率的蛋白质结构;但是x射线晶体衍射法要求对分析蛋白质的晶体(制备蛋白质单晶是很复杂和困难的),且要求样品量大、费时、灵敏度差,适合稳态、易于获得晶体的蛋白,对于溶液中的、瞬时的构象变化难以适用。红外光谱法灵敏度低(在组分分析中要求含量一般应大于0.1%),受水的信号干扰严重,后来发展的近红外技术也未能解决该问题,使得红外光谱法无法用于接近生理环境的体系。核磁共振法、质谱法、荧 光光谱法和拉曼光谱法目前能够检测溶液中的蛋白质构象变化。但核磁共振法仅适合于分子量小于40kda的样品,灵敏度低,分析浓度通常需要达到mm级,且数据处理复杂,分析成本高。质谱法可根据电荷价态分布来研究蛋白质构象,灵敏度高、能够实时监测和定量分析,但常用的esi-ms以及改进了的冷喷雾电离csi-ms等,均是进行气态化离子的分析检测,所给出的信息不能十分准确地表征蛋白质在溶液中的真实构象。荧光法中应用较多的是荧光共振能量转移法(fret),需要引入较大体积的供体和受体,对蛋白结构和功能会产生一定影响,并且荧光法也有较强的运用现在(比如fret的效果在10nm范围内是比较灵敏的,超出该限制条件就很难适用了)。相反,拉曼光谱法简单易行,不受水的干扰,无需对被测样品作标记及特殊制备,能够提供物质的分子组成和结构等样品内在的信息,获得丰富的指纹谱峰,已经在细胞、组织甚至活体中得到了应用。但是传统基于拉曼光谱的检测方法,其灵敏度较低,且天然生物分子的谱峰重叠、干扰严重,严重限制了拉曼法的进一步应用。
技术实现要素:
本发明的目的是针对现有技术中存在的难题,提供一种利用生物正交拉曼报告基团特异性标记、检测蛋白质的方法。
为了提高拉曼光谱法的灵敏度,近年来研究者发展了各种增强技术,如表面增强拉曼(sers)。sers是au、ag等金属表面对于拉曼信号的一种增强现象,一般可以实现106-1012的增强效果,可达到与荧光光谱相近的灵敏度。另一方面,为了解决生物分子的拉曼谱峰重叠的问题,申请人的发明人研究了正交拉曼标签标记的策略,即在生物分子上引入拉曼信号正交基团(如炔基、叠氮、c-d等),它们的拉曼信号在细胞拉曼信号的沉默区(1800-2800cm-1)。
为达到上述目的,本发明的使用生物正交拉曼报告基团特异性标记、检测蛋白质的方法,包括如下步骤:
a)制备携带生物正交拉曼报告基团的非天然氨基酸;
b)利用基因扩展技术,将步骤a)制备的非天然氨基酸插入目标蛋白质,从而实现对目标蛋白质进行生物正交拉曼报告基团标记;
c)利用生物正交拉曼技术对步骤b)得到的蛋白质进行检测。
优选地,上述生物正交拉曼报告基团为炔基、叠氮、氰基或碳氘键。
优选地,上述方法还包括利用金纳米离子的表面增强效应。
优选地,利用金纳米离子的表面增强效应中的金纳米颗粒aunps通过采用柠檬酸钠还原法制备。
优选地,柠檬酸钠还原法中所用的haucl4的终浓度为0.25mm,柠檬酸钠的终浓度为0.01%(w/w)。
优选地,上述使用生物正交拉曼报告基团特异性标记、检测蛋白质的方法还可包括增加蛋白与aunps的亲和力。
进一步优选地,上述增加蛋白与aunps的亲和力包括使用含半胱氨酸的标签、含甲硫氨酸的标签或蛋白化学修饰上含巯基的小分子。
本发明的另一目的是提供一种使用生物正交拉曼技术体外检测蛋白构象变化的方法,包括如下步骤:
a)在不同溶液条件下,利用上述使用生物正交拉曼报告基团特异性标记、检测蛋白质的方法来检测蛋白质构象变化;
b)在基于步骤a)检测蛋白质构象变化的基础上,研究蛋白质构象变化和环境改变关系。
本发明的又一目的是提供一种使用生物正交拉曼技术在活细胞原位检测蛋白构象变化的方法,包括如下步骤:
a)利用上述使用生物正交拉曼报告基团特异性标记、检测蛋白质的方法检测活细胞表面的蛋白质构象变化;
b)在基于步骤a)检测蛋白质构象变化的基础上,在活细胞层面上,原位研究蛋白质构象变化和其蛋白质活性受刺激调节的关系。
优选地,上述刺激包括生长因子刺激、配体刺激或环境改变刺激。
本发明中使用生物正交拉曼报告基团特异性标记、检测蛋白质的方 法,利用基因扩展技术和携带生物正交拉曼报告基团(包含但不仅限于炔基、叠氮、氰基、碳氘键等)的非天然氨基酸技术实现对目标蛋白质进行生物正交拉曼报告基团标记的方法。
利用金纳米离子的表面增强效应,实现在溶液中检测特定蛋白质上生物正交拉曼报基团的方法。该方法具备高效、灵敏和目标蛋白质特异性。
利用生物正交拉曼标记结合金纳米离子表面增强效应,实现在活细胞层面上检测特定蛋白质的方法。该方法生物相容性好,操作简单,且兼具高效、灵敏和目标蛋白质特异性。
本发明还提供了使用生物正交拉曼策略体外检测蛋白构象变化的方法;使用炔基等生物正交拉曼报告基团,结合金纳米离子表面增强效应,在不同溶液条件下,检测蛋白质构象变化的方法;在基于生物正交拉曼检测蛋白质构象变化的基础上,研究蛋白质构象变化和环境改变关系的新方法。
使用含半胱氨酸的标签增加蛋白与aunps的亲和力,当然包括其他增加亲和力的方法用于sers增强的生物正交raman策略检测蛋白局部构象变化,包含但不仅限于含甲硫氨酸的标签或蛋白化学修饰上含巯基的小分子。
本发明还提供了使用生物正交拉曼策略在活细胞原位检测蛋白构象变化的方法;使用炔基等生物正交拉曼报告基团,结合金纳米离子表面增强效应,检测活细胞表面蛋白质构象变化的方法;在基于生物正交拉曼检测蛋白质构象变化的基础上,在活细胞层面上,原位研究蛋白质构象变化和其蛋白质活性受刺激(生长因子、配体和环境改变等)调节的新方法。
本发明提供的利用生物正交拉曼报告基团特异性标记、检测蛋白质的方法,利用生物正交拉曼技术体外检测蛋白构象变化的方法,以及利用生物正交拉曼技术在活细胞原位检测蛋白构象变化的方法,通过基因密码子扩增技术和sers增强的方法,实现快速、灵敏和高效地检测带有生物正交基团的蛋白质在不同的微环境或活性条件下其构象的变化,特别是实现活细胞上蛋白质构象变化的原位检测,进而解析蛋白质构象与功能之间的联系。该技术基于在蛋白质某特定位点引入生物正交拉曼报告基团的方法 和拉曼基团的分子振动信号对微环境改变非常敏感并随环境的变化会发生偏移的理论基础,实现对溶液中蛋白构象变化的检测以及活细胞上蛋白质构象变化的原位和灵敏的检测。
本发明的上述方法适用于蛋白质溶液中和活细胞生理条件下表面蛋白质构象变化的生物正交拉曼检测。本发明的上述方法具有简单易行,体系接近生理条件、灵敏度高、正交拉曼信号干扰小、数据处理简单,对蛋白分子量没有限制,适合多种蛋白体系的特点。
附图说明
图1是基于生物正交拉曼报告基团(炔基)的蛋白质构象变化检测;
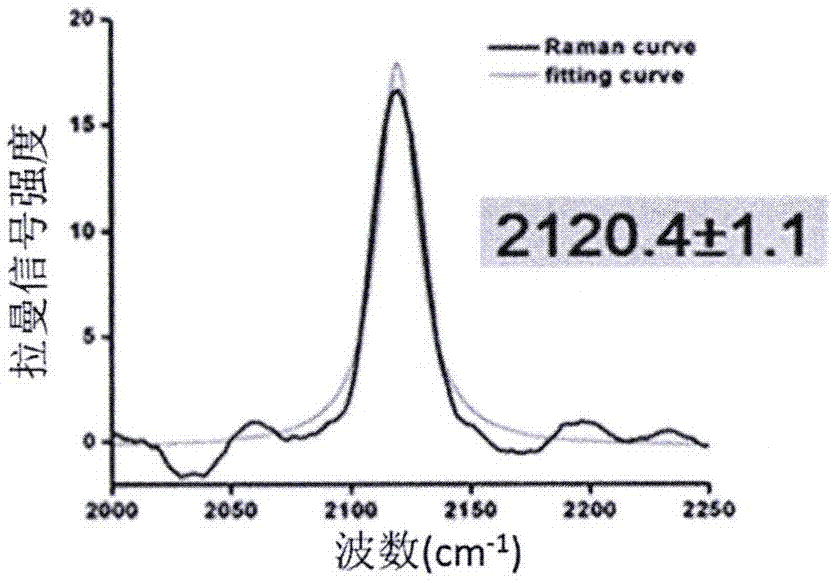
图2是hdea蛋白35位炔基在ph7的条件下的raman位移;
图3是hdea蛋白35位炔基在ph2的条件下的raman位移;
图4是hdea蛋白58位炔基在ph7的条件下的raman位移;
图5是hdea蛋白58位炔基在ph2的条件下的raman位移;
图6(a)是细胞上未做突变的egfr(wt-egfr)的raman光谱;
图6(b)是细胞上356位突变并插入penk但未经egf刺激的egfr的raman光谱;
图6(c)是细胞上356位突变并插入penk经过egf刺激的egfr的raman光谱。
具体实施方式
为了使本发明的目的、技术方案及优点更加清楚明白,下面结合附图及实施例,对本发明进行进一步详细说明。应当理解,此处所描述的具体实施例仅用以解释本发明,并不用于限定本发明。
本发明利用基因扩增技术在蛋白上特定位点插入含有正交基团的非天然氨基酸,利用金纳米离子能对拉曼信号进行等离子增强,实现对蛋白质高效、灵敏和快速的检测。因为基团的拉曼信号会受基团附近微观环境(比如,溶液种类、基团种类、电性等)的影响,并发生拉曼峰位的偏移, 实现对拉曼正交报告基团其微观环境变化的检测。而拉曼正交报告基团微观环境的变化,是和蛋白质的构象变化相关联的,这种变化反映了蛋白质构象的转变。将蛋白质与金纳米粒子靠近后(小于10nm)时,检测不同条件下,生物正交拉曼报告基团(此处为炔基)拉曼信号,得到该目标蛋白质构象变化的信息。具体见示意图1。
生物正交拉曼标签标记的策略,即在生物分子上引入拉曼信号正交基团(如炔基、叠氮、c-d等),它们的拉曼信号在细胞拉曼信号的沉默区(1800-2800cm-1)。这一策略解决了生物分子的拉曼谱峰重叠的问题,大大简化了拉曼数据的分析过程。基因密码子扩增技术是利用真核生物中终止密码子tag不编码氨基酸且使用概率低的性质,引入经过筛选的与tag和非天然氨基酸正交的氨酰转移酶-trna对,使细胞将非天然氨基酸插入蛋白的tag突变位点。利用基因扩增技术将含有拉曼正交基团的非天然氨基酸插入蛋白中,进行生物正交拉曼检测。
实施例1溶液中含炔基hdea蛋白在不同ph下的构象变化的检测
hdea是细菌膜间质中的分子伴侣蛋白。在ph7的生理环境下,hdea以同源二聚形式存在;而在ph2的生理环境中hdea解聚暴露其活性位点与其底物蛋白结合。在ph2的生理环境中hdea处于disorder(混乱)的状态,不能使用x-射线晶体衍射法研究其结构和构象变化。penk是含有炔基的赖氨酸衍生物(如下图所示),可以用正交的penk-mbpylrs/trna将penk插入蛋白的tag位点。
aunps的合成:采用柠檬酸钠还原法制备金纳米颗粒(aunps)溶胶,其中haucl4的终浓度为0.25mm,柠檬酸钠的终浓度为0.01%(w/w)。在煮沸的超纯水中,剧烈搅拌下分别加入haucl4和柠檬酸钠,30min后停止加热。
含炔基hdea的表达纯化:在细菌hdea蛋白n-端信号肽后引入含多个半胱氨酸的标签(gccpgccggsgs),在c-端引入his6标签。其35位或58位氨基酸对应密码子序列突变为tag,引入含炔基的非天然氨基酸penk。将改造的hdea基因序列插入pbad载体中。使用e.coli进行蛋白表达,在非天然氨基酸的存在下,使用阿拉伯糖诱导表达。然后使用ni柱纯化表达的蛋白。获得含有炔基的hdea蛋白。
aunps和hdea孵育:0.1mg/ml含炔基的hdea加入aunps溶液中,使用半胱氨酸的标签与aunps进行结合。室温孵育0.5h后,加入10x缓冲溶液使缓冲溶液终浓度为1xpbs(ph7)或柠檬酸钠缓冲溶液(10mm,含150mmnacl,ph2)。然后使用horiba拉曼显微镜检测溶液的raman光谱。
拉曼检测结果如下:
1、蛋白hdea(35位炔基),ph2到7
如图2所示,在ph7的条件下,hdea处于同源二聚的非活性状态,此时35位炔基的raman峰为单一的峰2120.4cm-1。如图3所示,而在ph2的条件下,hdea解聚为单体,逐渐形成活化构象,此时35位炔基的raman峰出现拖尾的肩峰,可以分峰为2126.1cm-1和2146.7cm-1。都相对于2120.4cm-1向高波数位移,分别对应于“开始解聚”和“近活化”的构象。
2、蛋白hdea(58位炔基),ph2到7
如图4和5所示,hdea58位炔基的构象变化和hdea35位差不多。但是二者“开始解聚”和“近活化”构象的比例不同。因为hdea在解聚逐渐形成活化构象的过程中不同氨基酸位点的变化不完全同步,所以不同氨基酸位点反映的“开始解聚”和“近活化”构象的比例不同。本发明提供一种使用生物正交拉曼报告基团特异性标记、检测蛋白质的方法,能够研究不同氨基酸位点在蛋白构象变化过程中的构象变化有无、快慢和比例。
实施例2细胞表面含炔基egfr蛋白在egf刺激前后构象变化的检测
egfr是表皮生长因子受体蛋白,位于细胞膜表面。egfr信号通路对细胞的生长、增殖和分化等生理过程发挥重要的作用,与多种癌症密切相关。egfr与表皮生长因子egf结合后会发生二聚,egf释放后主要以单体形式存在。
aunps的合成:采用柠檬酸钠还原法制备。
含炔基egfr在细胞上的表达:将egfr蛋白的210、250、356或440位的密码子分别突变为tag。在非天然氨基酸penk的存在下,将含有改造的egfr和penk-mbpylrs/trnapylcua基因序列的质粒瞬转入hek293t细胞中。40h后,用不含血清的培养基对细胞进行8h的饥饿处理,之后用100ng/ml的egf溶液刺激实验组细胞,15min后用4%多聚甲醛溶液固定细胞。金溶胶中加入1/10体积的10×pbs,快速加入细胞体系中孵育1h。使用nanophton拉曼显微镜检测细胞的拉曼光谱。
拉曼结果如下:
如图6(a)-6(c)所示,没有用egf刺激(egf-)的细胞上egfr主要以单体形式存在,经egf刺激(egf+)的细胞egfr主要以二聚体存在,在两种构象中同一位点的炔基周围的化学环境(如电场、亲疏水性)不同,其拉曼信号的位置会发生偏移。以356位点为例,未做突变的wildtype-egfr对照组没有炔基的拉曼信号,356位突变并插入penk的实验组均可测到炔基的拉曼信号,其中,egf-组细胞的炔基拉曼信号在2122cm-1,egf+组细胞的炔基拉曼信号在2126cm-1,即egf刺激前后egfr构象变化导致炔基的拉曼信号发生了位移。4个不同位点突变的统计结果如下表中所示,其中250、356和440位点,egf刺激后拉曼信号高波数位移,而210位点低波数位移,反映了egfr不同位点的化学环境在构象转化中经历了不同的变化趋势。
表1显示了细胞上egfr的不同位点炔基在egf刺激前后的拉曼位移,具体如下:
表1
溶液和细胞的结果表明,插入蛋白的炔基在不同条件下的拉曼信号的变化能够反映蛋白构象变化。利用生物正交拉曼技术能够对蛋白构象变化进行原位检测。
虽然本发明已以较佳实施例披露如上,然其并非用以限定本发明,任何所属技术领域的技术人员,在不脱离本发明的精神和范围内,当可作些许的更动与改进,因此本发明的保护范围当视权利要求所界定者为准。
- 还没有人留言评论。精彩留言会获得点赞!