一种基于固体核磁共振技术检测RNA结构的方法与流程
本发明涉及生物大分子结构检测领域,尤其涉及一种基于固体核磁共振技术检测rna结构的方法。
背景技术:
rna是生物体内必需的生物大分子,广泛存在于病毒、原核生物、真核生物中。rna在传递遗传信息、调控基因表达、抵御和治疗疾病等方面发挥着重要的作用(crickf,nature,1970,227,561-3;mehdizadehaghdame,etal.,gene,2016,0378111916305613;wangj,etal.,oncotargets&therapy,2016,9,681;fangy,fullwoodmj.,genomicsproteomics&bioinformatics,2016,14(1):42-54.)。rna功能的多样性与其rna复杂的空间结构密切相关(mortimersa,etal.,naturereviewsgenetics,2014,15,469-479.)。rna结构研究的实验技术主要包括x-ray晶体衍射技术、液体核磁共振技术和低温冷冻电镜技术。近年来rna结构发展迅速,但与蛋白质相比,rna结构研究的进展仍然落后于蛋白质结构的研究,已有的三维结构数据信息非常有限。据统计(http://www.rcsb.org,截止至2018年4月28日),已知的rna结构总量仅为已知蛋白质结构总量的1%。因此发展新型的rna结构解析技术尤为重要。
固态核磁共振技术是rna研究的新技术,可以在结晶或沉淀的rna中获取rna的结构信息(riedelk,etal.,journalofbiomolecularnmr,2005,31,49-57;marchankaa,simonb,carlomagnot.angewandtechemieinternationaledition,2013,52,9996-10001.)、分析rna动态性质(olsengl,etal.,journaloftheamericanchemicalsociety,2010,132,303-308.)等;对于大分子量rna(leppert,j.nucleicacidsresearch,2004,32,1177-1183.)以及rna-蛋白相互作用等领域(carlomagno,teresa.journalofmagneticresonance,2014,241,126-136.),也有独特的优势。因此,固体核磁研究rna的实验方法是传统rna结构解析技术的有效补充。
在rna固体核磁实验设计中,通常在魔角旋转(magicanglespinning)的条件下结合交叉极化方法(crosspolarization,cp)实现磁共振信号在不同磁性原子核之间的转移(pines,a.thejournalofchemicalphysics,1973,59,569.);既可以实现低灵敏核的信号增强,也可以实现异核低灵敏原子核之间的磁共振极化量的转移,实现化学位移相关谱的建立。利用交叉极化脉冲序列,设计复杂的信号传递途径,将同核或异核的原子核串联起来,可以获取原子核之间的长程距离相关,得到重要的结构信息,因此多用于多维固体核磁共振实验的设计中。以n-c(或c-n)传递为例,常用的固体核磁共振实验方法,2dspecific15n-13ccp方法通常用于构建rna二维15n-13c相关谱(langea,lucas,baldusm.journaloftheamericanchemicalsociety,2002,124(33):9704-9705.),可以实现芳香环内部n核和c核之间信号的传递,如尿嘧啶中n1-c2、n1-c6、n3-c2和n3-c4之间的传递,或构建13c化学位移选择性过滤的二维1h-15n相关谱等。
在cp传递中,由于传递的效率与传递原子核之间的距离成反比,并且与传递原子核磁旋比相关,因此13c-15n之间的传递效率往往低于1h-x(x=13c或15n)的传递效率。此外,在魔角旋转条件下,cp传递必须满足hartmann-hahn条件(hartmanns,hahne.physrev,1962,128(5):2042-2053.)。在高转速(如40khzmas)条件下,低灵敏核之间cp的匹配条件更为苛刻。
技术实现要素:
为了获得核糖核苷酸碱基中氨基、亚氨基上的1h原子和碱基上13c之间的相关信息,需要收集经过15n过滤的2d1h-13c相关谱。传统的hcnh脉冲序列,通过1h-13ccp实现13c的极化,并纪录13c的化学位移,然后将13c信号传递到相邻的15n原子,再由15n原子将信号传给直接相连的1h原子,进行1h化学位移记录。然而该序列传递效率较低,原因有两点:(1)cp传递的两核之间需要满足hartmann-hahn条件,即两核(i和s)的射频频率(rf)需要满足公式:ωi±ωs=nωr,ωr代表魔角旋转的频率,n为整数,ωi、ωs代表两核的射频频率。在高速魔角旋转条件下,13c与15n之间传递的频率条件较难匹配,并且化学位移各向异性使得13c-15n传递受到激发相位的影响,实验中优化匹配条件更难。(2)cp传递的效率与原子核磁旋比相关,1h的磁旋比是13c的4倍,是15n的10倍,因此1h-x(x=13c、15n)之间的传递效率高,而13c-15n之间的cp传递效率低。
针对上述现有技术中存在的问题与不足,本发明提供了一种基于固体核磁共振技术检测rna结构的方法,通过设定一种新型hchnh脉冲序列,不仅可以获得与hcnh相同的2d1h-13c相关谱,而且大幅提高了磁化转移的效率,提高了二维谱图信号的灵敏度。hchnh序列在hcnh序列的基础上,改进传递效率低的13c-15ncp传递,变为连续的两个含有氢原子核的cp传递,即13c-1h和1h-15n传递。利用1h的辅助,先将13c信号传递给1h,再通过1h将信号传给15n,实现了高效的磁共振极化率的传递,将信号灵敏度提高了60%。
本发明通过如下技术方案实现:
一种基于固体核磁共振技术检测rna结构的方法,包括以下步骤:
(1)待测rna样品的制备,所述rna样品中至少一种碱基由15n和13c标记;
(2)使用固体核磁共振波谱仪对rna样品进行检测;
(3)检测图谱的收集与分析;其中,
步骤(2)中在使用固体核磁共振波谱仪对rna样品进行检测之前,首先进行脉冲序列hchnh的设定(如图2所示):首先将1h信号全部激发,1h-13ccp实现低灵敏度13c核的极化,以及化学位移记录,设定接触时间为1ms-10ms,;在13c化学位移演化过程中,采用tppm脉冲对1h去耦;之后13c-1hcp将13c极化量转移给1h原子,再由1h-15ncp传递给15n核,设定接触时间为100μs-1000μs,最后将15n信号传回与15n核直接相连的1h原子核,记录1h化学位移。
进一步地,步骤(1)中所述rna样品的制备包括通过体外转录法、固相合成方法或生物提取法得到的13c、15n同位素标记的rna样品。所述rna样品纯度在90%以上,固体样品中rna质量大于1mg。
进一步地,步骤(3)中所述检测图谱包括一维1h-15ncp谱图,一维1h-13ccp谱图,一维1h-31pcp谱图,二维1h-15nhnh谱图,二维1h-13chch谱图,二维13c-15nhnc谱图,二维1h-15nhcnh谱图,二维1h-15nhcn(par)nh谱图,二维15n-13ctedor谱图,二维13c-13cdarr谱图。
相较于现有技术,本发明的有益效果如下:
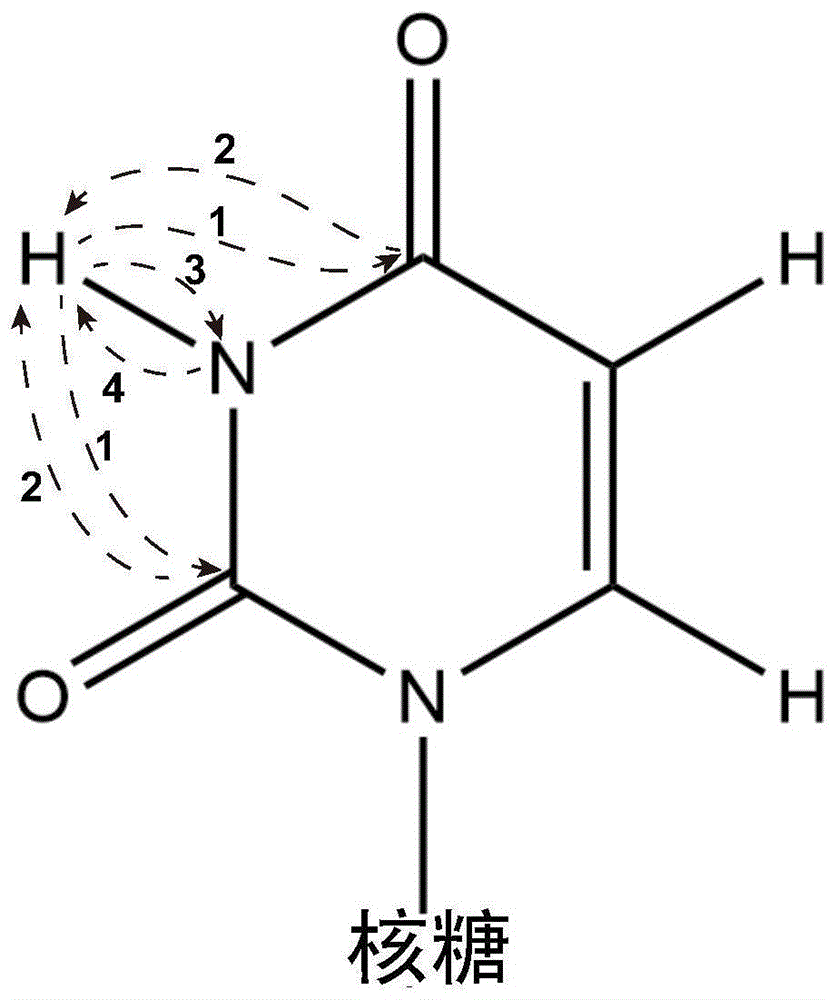
本发明提供了一种基于固体核磁共振技术检测rna结构的方法,通过一种新型二维固体核磁脉冲序列hchnh,实现了1h-13c相关谱的建立,并建立了rna碱基内部原子的长程相关。hchnh脉冲序列利用两个高传递效率的13c-1h和1h-15ncp传递,代替传统的低传递效率的13c-15n传递,实现长程13c-1h相关谱的建立,并用于经典和非经典配对信息的获取。以尿嘧啶(uracil,记做u)为例,信号传递过程如图1所示,第一个1h-13ccp首先实现低灵敏度13c核的激发,随后进行化学位移记录,随后将13c信号传递给1h原子核,再由1h原子将信号传递给临近的15n原子核;最后传递回1h原子,进行1h化学位移纪录。hchnh脉冲序列中,所有的信号传递均通过交叉极化脉冲序列完成。与常用的hcnh序列不同,hchnh利用两个高效率的包含氢原子核的cp传递方法代替13c-15ncp传递,在高转速固体核磁的条件下,灵敏度提高60%以上。基于hchnh脉冲序列的rna结构固体核磁共振技术检测还可以实现区分rna中沃森-克里克碱基配对和非沃森-克里克碱基配对,例如:u-u和g-u配对等,拓展了固体核磁共振技术在解析rna结构方面的应用。
附图说明
图1为以尿嘧啶为例hchnh脉冲序列的信号传递方式;
图2为hchnh脉冲序列示意图;
图中:
图3为本发明一实施例humantrnasec(pdb:3a3a)结构模型;
图4为本发明一实施例u-labeledtrna的二维hchnh谱(4a)和hnch谱(4b);(4c)一维hchnh1h谱与一维hcnh1h谱的比较;(4d)一维hcnh1h谱与一维hnh1h谱的比较。
具体实施方式
以下将结合实施例对本发明做进一步说明,所举实施例并不用于限制本发明。对于本领域技术人员来说,本申请可以有各种更改和变化。凡在本申请的精神和原理之内所作的任何修改、等同替换、改进等,均应包含在本申请的权利要求范围之内。
实施例1
1.rna样品的制备:
本实施例以人源的硒代半胱氨酸转运rna(humantrnasec)为模型进行脉冲序列的测试。该rna由92个核糖核苷酸构成,结构如图3所示。为了简化谱图信息,在体外转录时,本实施例仅使用了15n,13c标记的三磷酸尿苷(utp),其余三种核苷三磷酸(atp、ctp、utp)均为天然丰度。合成的rna记作u-labeledtrna。
(1)dna模板的构建
本实施例通过体外转录方法得到u-labeledtrna,首先需要根据目标rna设计相应的dna模板。dna模板由两条互补的dna单链组成,分别为正向链(r链)和反向链(f链),序列如下:
r链:5’-tggcgcccgaaaggtggaattgaaccactctgtcgctagacagctacaggtttgaagcctgcaccccagaccactgaggatcatccgggccctatagtgagtcgtattaa-3’
f链:5’-ttaatacgactcactatagggcccggatgatcctcagtggtctggggtgcaggcttcaaacctgtagctgtctagcgacagagtggttcaattccacctttcgggcgcca-3’
两条dna单链经过离心、溶解、升温退火等过程,形成线性化的双链dna模板(终浓度为25μm)。
(2)体外转录制备rna样品
本实施例通过10ml的体外转录体系得到rna样品:在50ml离心管中依次加入5.3mldepc处理水,500μl100mm13c,15n同位素标记的utp,100mm天然丰度的atp、gtp、ctp各500μl,1mltrisbuffer(含400mmtris/hcl,ph8.0,10mmspermidine,0.1%tritonx-100),100μl1m二硫苏糖醇(dtt),350μl1mmgcl2,180μl25μmdna模板,1ml2mg/mlt7rna聚合酶。37℃水浴中孵育24小时以保证原料完全反应。
(3)rna的纯化
转录反应完成后通过含7m尿素的8%变性聚丙烯酰胺(page)凝胶电泳进行纯化,电泳结束后在紫外灯下切胶得到目标条带。将含有目标rna的条带浸泡于提取缓冲液中(20mmtris-hcl,300mmsodiumacetate,1mmedta,ph7.4)约12小时,收集溶液,通过超滤的方法将rna样品溶解于含10mmmgcl2,10mmtris–hcl,ph8.0,75%氘代的溶液中。65℃下加热5分钟,冰浴30分钟,使trna正确折叠。
(4)rna固体样品的制备
向一定浓度的u-labeledtrna纯品溶液中先加入1/10体积的2mnacl溶液,再加入2-3倍体积的-20℃预冷的75%氘代无水乙醇,将整个体系放置在-20℃的洁净环境中过夜,以9600g离心5min后,弃去上清,用少量70%的预冷乙醇清洗底部沉淀,即可得到rna沉淀。通过上述方法制备大约5mg含13c,15n同位素标记的u碱基的rna纯品,装进1.9mm固体核磁转子中。
2.脉冲序列的设置
2dhchnhssnmr实验在40khz的魔角转速下完成。直接维(f1:1h),间接维(f2:13c),在t1演化期和t2采集期分别取了574个点和100个点。1h的π/2脉冲为2.5μs,13c的π/2脉冲为3.5μs,15n的π/2脉冲为4.5μs。循环等待时间为2s。在1h-13c交叉极化传递和13c-1h交叉极化传递过程中,13c的连续锁场功率均为60khz,1h的锁场功率为100khz,接触时间为2ms。在13c化学位移演化期,采用tppm脉冲对1h去耦,去耦功率10khz。实验中利用missisippi脉冲序列对水的信号进行压制。1h-15n交叉极化传递和15n-1h交叉极化传递中,cp接触时间设为300μs。1h-15n和15n-1h传递时15n的连续锁场功率设为50khz,1h的锁场功率为90khz。采样叠加200次。
3.谱图收集与结果分析
请参照图4。为了简化谱图信息,本实施例收集了尿嘧啶核糖核苷酸被标记的humantrnasec(u-labeledtrna)的二维hchnh谱图,结果如图4a所示。在图中既观察到了沃森-克里克碱基配对信息,又观察到了非沃森-克里克碱基配对信息,其中包括参与u-a碱基互补配对时尿嘧啶中h3与c4的交叉峰和h3与c2的交叉峰,以及参与非沃森-克里克碱基配对时尿嘧啶中h3与c4的交叉峰和h3与c2的交叉峰。
此外,本实施例还收集了该rna的2dhcnh谱进行对比(图4b),t1演化期和t2采集期也分别取了574和100个点。在谱图中也可以观察到参与沃森-克里克碱基配对的尿嘧啶中h3和c4的交叉峰、参与沃森-克里克碱基配对的尿嘧啶中h3和c2的交叉峰、参与非沃森-克里克碱基配对的尿嘧啶中h3和c4的交叉峰,但与2dhchnh谱对比发现谱峰强度大幅度降低。非沃森-克里克碱基配对的尿嘧啶中h3和c2的交叉峰由于信号强度很弱未能在谱图中观察到。
为了进一步对比hchnh和hcnh两脉冲的传递效率,收集该rna的1dhchnh1h谱和1dhcnh1h谱,并且分别与1dhnh1h谱进行比较。图4c对比了hchnh1h谱和1dhnh1h谱的信号强度,其中hchnh1h谱的信号强度大约为1dhnh1h谱的50%。而图4d中1dhcnh1h谱信号强度仅为1dhnh1h谱的30%。由此可见,hchnh脉冲序列的设计,大幅度提高了长程13c-1h相关谱的灵敏度。
- 还没有人留言评论。精彩留言会获得点赞!