多通道神经刺激装置及其应用的制作方法
背景技术:
通过对大脑的神经元进行电脉冲刺激,来影响神经元的活动,从而达到治疗某些脑疾病的目的,是神经医学领域的现有技术。但申请人发现,现有技术大多只是对某些位置的神经元进行孤立的刺激,影响这些神经元的活动,比如用于刺激某些神经核团,使其神经功能产生增强或减弱的变化,或者用于干预神经核团的信号传递,比如阻隔疼痛信号的传递达到电子止痛的目的。而若要用于启动某种具体的神经活动,尤其是要产生或改变神经核团之间具有信号定向传递关系的可持续的动作电位发放,由于大脑中枢神经的各个神经链路的神经元之间是具有神经投射和信号传递关系的,所以,现有孤立的脑神经刺激技术,便难以产生这样的技术效果,而申请人发现在某些有意义的特定应用场合,往往便需要具有这样的技术效果。而且,现有技术对电刺激的具体位置、施加方式、和电脉冲的细节等无针对性,采取的技术往往跟用于外周神经刺激时是一样的,但实际上,对中枢神经元的刺激具有其特殊性,比如,根据对大脑一些神经回路的放电研究,施加到神经元的电脉冲,频率和脉冲幅度过低时无法触发动作电位放电,而频率或脉冲幅度过高时则会对神经元信号传递形成抑制,甚至使其失活。
技术实现要素:
本发明目的是公开一种多通道的神经刺激装置,适合于对某些神经通路的不同位置的神经元同时进行脉冲刺激,促使神经通路产生具有信号定向传递关系的动作电位发放。
本发明还公开对输出信号进行优化,使其能更有效地诱发神经元发放动作电位的技术。
本发明同时公开上述的神经刺激装置的一些应用,包括用于增强某些神经传递通路的传递效能,用于唤醒深度昏迷病人,以及用于对脑死亡在短时间内的激活。
本发明的神经刺激装置,包括主机和连接主机的电极,电极包括有接地电极(或称基准电极)和输出电极,主机包括有用于产生刺激脉冲信号的脉冲产生模块,连接各输出电极的用于将刺激脉冲信号输出到输出电极的各个输出模块,其特征在于:所述的输出电极包括有二组或二组以上的输出电极,(比如三组输出电极);主机还包括有输出时序控制电路,用于控制各输出模块输出的刺激脉冲信号,使各输出电极所输出的相对于基准电极的刺激脉冲具有前后依次输出的时序关系。
脉冲产生模块属于现有技术,所产生的脉冲可以采用方波、三角波、梯形波、或者模仿神经元动作电位脉冲的波形。脉冲宽度T0为0.5—10毫秒,更合适的是1—5毫秒,相当于神经元动作电位的脉冲宽度。各输出电极输出的脉冲信号,相对于基准电极(也即接地电极)的电位,脉冲信号的峰值电压幅度,在20—1000毫伏之间进行调节,根据电极接触位置及电极进入神经元的状态而有所不同。
本发明的主机还包括脉冲周期调节电路,用于调节同一组输出电极(第一、第二和第三输出电极)所输出的前后两个脉冲的脉冲周期T。对于不同的应用,脉冲周期T是不同的,一般在25毫秒至1000毫秒之间,对应于频率是1—40赫兹,见后面应用部分所述。
本发明主机还包括延迟时间调节电路,用于调节在同一个脉冲输出周期中,前后两个电极(比如第一输出电极和第二输出电极)输出的前后两个脉冲之间的延迟时间Ts。
输出时序控制电路用于控制各输出模块的工作,使得各组输出电极输出的刺激脉冲的时序关系是:在一个脉冲周期T中,前一输出电极输出一个刺激脉冲后,经过设定的延迟时间Ts后,后一输出电极才输出一个刺激脉冲。具体的控制方法可以是:脉冲产生模块产生一个脉冲信号,然后经过一个延迟时间Ts后,再产生一个脉冲信号,然后再经过一个延迟时间Ts后,再产生一个脉冲信号,时序控制电路对三个输出模块进行依次的选通输出,将这三个脉冲信号依次从三个输出模块进行输出,从而在三组输出电极上依次各输出一个脉冲信号。(注意这时脉冲产生模块的输出脉冲频率是输出电极输出脉冲频率的3倍),这样的控制属于普通的电子技术,可以通过简单的MCU或者数字电路来实现。
本发明的神经刺激装置具有多组输出电极,各输出电极能够输出相对于基准电极的刺激脉冲信号,且各组输出电极所输出的刺激脉冲信号具有前后依次输出的时序关系。使用本发明能够通过各组输出电极分别对某一神经通路上不同节点的神经元同时进行具有前后时序关系的脉冲刺激,强制神经通路产生具有信号定向传递关系的可持续的动作电位发放,从而可应用于激发和恢复某些具有生理功能的神经活动。(见后面公开的本发明的应用)。
作为一种重要的优化,本发明还对这种输出脉冲的时序关系进行精准的设置。其前后两组输出电极(比如第一与第二输出电极,第二与第三输出电极)输出的两个脉冲的延迟时间Ts,与脉冲周期T,脉冲宽度T0,三者具有如下关系:0<Ts<1/2T-T0;表述为:前后两组输出电极输出脉冲的延迟时间大于0,且小于“脉冲周期的一半减去一个脉冲宽度”的值。这种设置基于脑神经的STDP突触可塑性原理,(Spike Timing-Dependent Plasticity,即时序依赖的突触可塑性),其意义是:以具有这样时序关系的刺激脉冲,分别对某一神经通路上前后不同节点的神经核团进行脉冲刺激,不但在发放刺激脉冲的过程中能强制该神经通路产生定向激活动作,从而强制产生信号传递的神经活动,而且,这种脉冲刺激具有特定的时序关系,能够使所刺激的前后两组神经元之间的突触传递产生增强效应,提高突触传递效能。所以,这样的脉冲刺激,同时具有刺激神经元和定向提高神经元之间突触可塑性的作用,有利于恢复神经通路自身的信号传递效能,从而恢复其生理功能,所以可以用于一些工作异常的神经通路的康复。
在本发明上述的刺激方法和装置中,其进行神经元刺激的电脉冲串的输出特性是很重要的。所以,申请人仔细根据神经元爆发动作电位的具体工作原理,公开三种对输出电极的输出信号进行优化的方案,使得输出脉冲信号能够更准确地诱发神经元爆发动作电位,并且这种动作电位是能够持续发放的,更有利于受刺激的中枢神经元形成可持续的信号传递。
本发明的第一种输出优化方案,包括:所述的脉冲产生模块,带有正脉冲输出端和负脉冲输出端,能够分别输出相对于接地电极是正电位和负电位的两种脉冲输出信号;(这可以通过正负电源来产生,是普通的电子技术);正脉冲和负脉冲两个输出端通过切换开关进行切换,切换开关的输出再作为脉冲产生模块的输出端,连接到各个输出模块。切换开关可以是机械开关或电子开关。如果在实施脑刺激时是采用开颅的方式,并且采用专用于脑神经刺激的膜片钳或神经微电极,能够将输出电极刺入所要刺激的神经元膜内,则可将切换开关切换到正脉冲输出端,使电极输出端连接到正脉冲输出端,这时输出电极是输出正电位的脉冲信号。这种方式是目前实验室进行神经元激活的惯用方式,能够可靠的刺激诱发神经元爆发动作电位,但要将微电极准确刺入目标神经元膜内,显然需要进行开颅手术,极其麻烦。所以,如果将本发明的切换开关切换于负脉冲输出端,使电极输出端连接到负脉冲输出端,这时输出电极输出的是负电位的脉冲信号,可直接施加于神经元的膜外,通过拉低膜外电位,来相对降低膜静息电位,并诱发动作电位。这时可采用普通的电极,并在脑颅上开小孔、或者通过鼻腔或喉后颚开小孔,然后在神经内窥镜的配合辅助下将输出电极插入并置于所需要刺激的目标位置上,只需使输出电极接触或靠近神经细胞的膜外,而无需准确刺入神经元,所以操作更加方便。
本发明的第二种输出优化方案,主机上用于连接各个输出电极的电极输出端,与各个输出模块(包括第一输出模块、第二输出模块和第三输出模块)的输出端之间,加有输出控制电路,(更合适的,在输出模块的输出端与输出控制电路之间还可串接有一电阻器R,用于缓冲和产生一定的电压差);电极输出端还连接到一电压检测电路的输入端,电压检测电路的输出端连接到一电压比较电路,与来自输出模块的输出脉冲信号进行比较;电压比较电路的输出端连接到输出控制电路的控制端,通过输出控制电路对脉冲输出信号进行控制。当电压检测电路检测到的电极输出端的电压信号,与输出模块输出的电压信号之间的差值大于设定数值时,则关闭输出控制电路,在该脉冲周期余下的时间关闭输出模块到电极输出端的输出信号,直到下一脉冲周期才重新打开。各个输出模块到其连接的各个输出电极,其控制电路是一样的。
按照这样的输出控制方案,工作时,在各个输出模块的每一个输出脉冲周期,输出控制电路首先被打开,脉冲输出信号通过电极输出端和输出电极施加于神经元,同时电压检测电路检测电极输出端(相当于输出电极)上的电压信号,并与输出模块的脉冲输出信号进行比较。当输出电极的脉冲信号对神经元的刺激达到一定程度,神经元爆发动作电位,产生离子超射现象,引起膜内外的电位出现急剧变化,通过输出电极传递到电极输出端,造成电极输出端上的电压出现较大波动,而且是与输出模块输出的脉冲输出信号明显的变化不同步。这一电压波动被电压检测电路所检测到,并通过电压比较电路与脉冲输出信号进行比较,识别为神经元已经爆发动作电位,于是电压比较电路的输出关闭了输出控制电路,在该脉冲周期余下的时间关闭输出模块到电极输出端的输出,直到下一个脉冲周期才重新打开。这样可使神经元在每一次动作电位爆发后不再受到外部脉冲的刺激,使其能够恢复膜极化状态,以便准备对下一次刺激进行整合和再次爆发动作电位,形成可持续进行的动作电位发放。这一技术方案,更符合中枢神经细胞的工作机制,既能够诱发神经元爆发动作电位,又不会过度刺激神经元以致使神经元受到抑制失活或受到损伤,能够持续诱发动作电位,形成连续的脉冲发放,并使神经元的离子运动和生理功能逐渐恢复正常。
本发明的第三种输出优化方案,主机上用于连接各个输出电极的电极输出端连接有一电压检测电路,(可以与前面其他技术的电压检测电路共用),电压检测电路用于检测电极输出端的背景电压,所以也可称为背景电压检测电路,电压检测电路的输出连接有一电压保持电路,对电压检测电路输出的背景电压值进行保持记忆,(相当于电压记忆电路,如果电路是采用模拟电路,则可采用电容配合运放电路来构成,如果电路是数字电路,则可直接记忆其电压值),电压保持电路输出的电压值连接到一个电压叠加电路,各个输出模块输出的脉冲输出信号,在电压叠加电路中与背景电压进行叠加,再连接到电极输出端。电压保持电路、电压叠加电路和各个输出模块的工作,受到一个状态切换电路的控制,(也可由主机总的控制电路进行控制),具体工作过程是:在每次进行脑神经刺激、将接地电极和输出电极置于目标区域后,或工作中重新调整输出电极和接地电极的位置时,状态切换电路将电路工作切换在“检测”状态,关闭输出模块的输出,先由电压检测电路检测电极输出端上的电压,(这一电压也即当前输出电极与接地电极之间的电位差,也即背景电压,可能为正也可能为负),其输出电压值由电压保持电路进行保持(也即记忆),并送到电压叠加电路上;之后,由状态切换电路将电路工作切换到“输出”状态,关闭电压保持电路的输入端,(使其输出电压保持之前输入的电压值),打开输出模块的工作,输出模块输出的脉冲信号,在电压叠加电路中与背景电压进行叠加,再连接到电极输出端。这样,输出电极所输出的脉冲信号,便已经将两电极之间的电位差进行抵销,可消除背景电压对脉冲信号刺激效应的影响。
本发明的神经刺激装置具有多组输出电极,各组输出电极输出相对于基准电极的刺激脉冲,且各组输出电极所输出的刺激脉冲具有前后依次输出的时序关系。本发明可以用于对大脑中枢神经受损而工作不正常的某些神经投射通路,比如听觉、触觉的感觉输入通路,或某些运动神经的输出通路,用于对这些神经通路的前后节点的不同神经核团同时进行具有前后时序关系的脉冲刺激,强制神经通路产生具有信号定向传递关系的动作电位发放,并增强其突触传递效能,从而恢复这些受损神经通路的信号传递功能。
本发明也可用于对受损的周围神经的康复。周围神经包括躯体运动神经和躯体感觉神经,当受到骨折、机械创伤、过度压迫、拉挤时,可受损而导致失去感觉或运动障碍。周围神经的再生能力较强,即使不进行手术大多也能自我恢复,但一般需要数月时间才能逐渐恢复。使用本发明具有两组输出电极的装置,将两组输出电极顺着神经信号传递的方向跨越在周围神经受损位置的两边,接地电极接触皮肤,对受损神经的前后两段进行具有前后时序关系的脉冲刺激,能够模拟神经信号的传递过动作,诱发加速其神经再生,大大加快康复速度,且比采用单一脉冲进行刺激的效果更好。
以下是本发明在中枢神经的特殊应用。当然,在使用这些技术进行神经刺激之前,必须先消除各种病因,修复创伤、恢复正常供血供氧、改善微循环和营养神经等护理。
1、本发明用于对深度昏迷病人的唤醒。昏迷大多是由于大脑缺血(缺氧)所致,也有部分是由于脑创伤造成,或异常的化学物质进入脑屏障影响神经活动引起,在消除各种病因后,仍有部分病人处于昏迷状态而无法恢复自主意识。这时病人具有呼吸、循环等生命体征,但处于深度昏迷状态,对外界的刺激没有高级神经反应。典型的如果是长期深度昏迷,也就是俗称的植物人。目前有采用电脉冲和交变磁场对脑部进行刺激的技术,但由于对人脑的思维意识活动的工作机制不清楚,没有理论依据,所以对电磁刺激的具体位置、施加方式、和脉冲的频率等细节也无针对性,效果仍然非常有限。
本发明应用于对昏迷病人进行脑部脉冲刺激时,需要采用三组输出电极,其方法是:
⑴、将第一输出电极置于脑干网状结构内侧区(也即中脑网状结构)的位置,并接触或刺入该部位的神经元;(更合适的是接触或刺入中脑网状结构向丘脑板内核投射的神经元);将第二输出电极置于丘脑板内核,并接触或刺入该部位的神经元;(更合适的是接触或刺入丘脑板内核中发侧枝向丘脑网状核投射的神经元);将第三输出电极置于丘脑网状核的位置,并接触或刺入该部位的神经元;(更合适的是接触或刺入丘脑网状核向中脑网状结构投射的神经元);将接地电极与人体相接触;(比较合适的接触位置是脑部的椎底动脉、脑脊液,更合适的位置是输出电极所在的脑组织周围的脑液处);
⑵、通过三组输出电极向三个位置的神经元进行具有时序性的脉冲刺激。其脉冲周期可选125—25毫秒,(对应的脉冲频率为8—40赫兹,是大脑处于清醒和思维状态时的脑电节律)。
⑶、脉冲刺激最好是间隔性的,即进行1—3秒的脉冲刺激后,停止刺激1—3秒。在停止刺激的间隔期中,同时检测昏迷病人的脑电波(EEG),如果持续检测到脉冲频率大于8赫兹的脑电波信号,(比如频率8—13赫兹的α波或频率14赫兹以上的β波),或观察到病人出现自主意识的反应,则停止发放脉冲刺激,否则继续进行下一次脉冲刺激。
根据研究,思维的本质是大脑皮质区(尤其是端脑联合皮质)的中间神经元的链式激活活动,而意识的本质是大脑对思维活动(也即中间神经元的链式激活活动)的自我感知,(联合皮质上链式激活的神经元的轴突侧枝向感觉皮质的联合感觉区进行投射)。但是,虽然思维和意识的神经元活动是主要发生在脑皮质,其活动却是受到丘脑(主要是丘脑板内核)向皮质投射的动作电位同步激励脉冲的控制。进一步研究发现,丘脑向脑皮质发放的同步激励脉冲,其起源在于“中脑网状结构—丘脑”之间往返发放同步激励脉冲所形成的振荡环路产生的,申请人将其称为“上环振荡环路”或简称“上环路”,其具体的投射关系也即脉冲发放关系为“中脑网状结构→丘脑板内核→丘脑网状核→中脑网状结构”,并同时由丘脑板内核向广泛的皮质区进行同步激励脉冲的发放,来控制皮质的神经活动也即思维活动。(见图5的脑干网状结构与丘脑及下丘脑之间的上下两个神经环路的示意图,思维系统涉及的是其中的上环振荡环路)。当大脑因为缺血缺氧或受伤时,这一振荡环路的神经元受损,对兴奋信号的整合和触发动作较慢,造成其发放的振荡节律过低,使得丘脑板内核向皮质发放的同步激励脉冲的发放节律过低,(低于4赫兹,即深度睡眠和昏迷时的脑电节律),无法使皮质的中间神经元进行突触整合而触发动作电位并以此形成链式激活,所以也就无法产生有意识的思维活动,处于类似于深度睡眠的状态。所以,本发明按照其同步激励脉冲发放的次序,采用具有时序性的电脉冲对这一环路的三个节点的神经元进行依次刺激,强制提高和恢复这一环路的同步激励脉冲的发放节律(大于8赫兹),使皮质的中间神经元能够形成链式激活,也即恢复有意识的思维活动。而由于本发明的输出脉冲具有特定的时序关系,还能够增强这一环路的神经元之间的突触传递效能,有利于恢复这一环路的自主的神经发放动作,从而最终能够不依赖本发明的脉冲刺激而形成自主的正常的神经思维活动。
2、本发明用于对脑死亡的大脑的激活。这里所说的脑死亡并非整个人体的衰老死亡,而是意外死亡。人脑对血氧高度敏感,在心脏停跳、溺水、窒息、外伤导致脑供血中断等情况下,完全性脑缺氧两分钟即可导致大脑生物电活动和脑功能完全停止,不单大脑思维活动完全停止,而且内脏神经系统的活动完全停止,机体丧失自主呼吸和心跳,如果脑缺氧超过四、五分钟,即使再给予心脏起搏和人工呼吸,也很难抢救过来。这时神经活动、自主呼吸和心跳完全停止,没有生命体征,即是法律意义和医学意义上的人体死亡。按照现有技术,脑死亡是不可逆转的,是生命的终结,这时的人体已经不称为病人,而是尸体。
对于脑死亡,这时大脑不单是控制思维活动的“中脑网状结构←→丘脑”的上环振荡环路的神经元停止动作电位发放,更主要的是控制内脏活动的内脏神经(或称自主神经、植物神经)系统也停止活动。根据分析,控制内脏活动的内脏神经系统包括两大环路:(见图5),一是由“内脏(感觉)神经→孤束核→迷走神经背核→内脏(运动)神经”构成的在脑干层面的低级反射环路,由简单的直接的反射环路分别对各个内脏器官的活动进行感觉和反射;二是由“脑干网状结构外侧区←→下丘脑”构成的联合反射环路,接受各种内脏器官的感觉传入,并接受大脑其他神经系统(思维神经系统、运动神经系统、情绪神经系统、内分泌神经系统)的调制输入,经过联合处理后,对脑干的低级反射环路的活动进行激励和调制,从而对内脏器官的工作进行平衡控制,比如,调节呼吸的频率和幅度,调节心跳的速率,调节血压等等。“脑干网状结构外侧区←→下丘脑”构成的这一反射环路控制和维持着生命的基本活动,其中由脑干网状结构外侧区(尤其包括臂旁内侧核、外侧核)的胆碱能神经向下丘脑(尤其包括下丘脑外侧区、后区、室旁核等)发出激励脉冲来形成往返振荡的脉冲发放,申请人把它称为“下环振荡环路”或简称“下环路”。当脑干的低级反射环路的神经元停止活动,则内脏器官也停止活动,当“下环振荡环路”的神经元停止活动,则内脏器官工作紊乱,甚至也会使低级反射环路因得不到激励而停止活动,同样导致内脏器官停止活动,呼吸和心跳停止,人体死亡。
所以,要对脑死亡的大脑进行脑激活使其恢复神经活动,可以对这两个反射环路的相关神经元进行联合的电刺激,使其激活并恢复动作电位发放,激励内脏器官恢复活动,从而恢复心跳和自主呼吸。需要注意的是,出于治疗风险和法律层面的考虑,应该对病人尽量进行传统抢救,在确认传统抢救无效,确认病人已经心跳停止、自主呼吸停止和脑死亡,也即确认病人已经处于法律意义和医学意义上的死亡状态,属于一具尸体后,再采用本发明的脑激活方法进行抢救。
本发明应用于对脑死亡的大脑进行脑部脉冲刺激时,也需要采用三组输出电极,其方法是:
⑴、将第一输出电极置于脑干网状结构外侧区(尤其包括臂旁核)的位置,并接触或刺入该部位的神经元;(更合适是接触或刺入脑干网状结构外侧区包括臂旁核向下丘脑投射的神经元);将第二输出电极置于下丘脑外侧区或后区,并接触或刺入该部位的神经元;(更合适是接触或刺入下丘脑外侧区或后区向迷走神经背核投射的神经元);将第三输出电极置于迷走神经背核的位置,并接触或刺入该部位的神经元;(更合适是接触或刺入迷走神经背核向节后神经元投射的神经元);将接地电极与人体相接触;(比较合适的接触位置是脑部的椎底动脉、脑脊液,更合适的位置是输出电极所在的脑组织周围的脑液处);
⑵、通过三组输出电极向三个位置的神经元进行具有时序性的脉冲刺激。其脉冲周期T为250—1000毫秒,对应的脉冲频率为1—4赫兹。(见下面对这一脉冲周期的分析和确定)。
⑶、脉冲刺激最好是间隔性的,即在发放脉冲刺激一段时间后,停止一段间隔期的时间,再进行下一段时间的脉冲刺激。具体可以是:进行3秒左右的脉冲刺激后,停止刺激2秒左右。间隔性发放的目的是:在停止刺激的间隔期中,同时检测受刺激大脑的脑电波(EEG),如果没有检测到脑电信号,则继续进行下一次脉冲刺激,如果在间隔期检测到有自发的脑电信号,说明大脑已经开始有自主的脑电活动,可停止发放脉冲刺激,观察其活动情况是否是稳定持续的生命特征,再决定是否需要继续刺激。
关于对下环振荡环路神经元进行脉冲刺激的脉冲周期的确定:在现有神经生物学的各种实验和神经刺激应用技术中,按照现有理论,由于脉冲刺激的频率过低时不容易形成兴奋整合而爆发动作电位,而且低频的脉冲刺激容易产生突触传递抑制,所以必然地要选择高频脉冲刺激,即强直刺激。但申请人发现,对于本发明的应用场合,脉冲刺激的频率过高时,在神经元被刺激并爆发动作电位后,仍会不断受到脉冲刺激,则会影响神经元爆发动作电位后的离子回流运动,使其无法重新形成正常的膜极化等生理活动,所以,这时神经元只是完全被动地被强直刺激触发而产生偶发的无序的动作电位,而无法形成可持续的有节律的动作电位发放,更加无法使神经元恢复生理功能并最终自发产生正常的动作电位发放,甚至可能更加损害其生理功能并导致其失活。只有电脉冲刺激的频率跟振荡环路原有的工作节律相接近时,才可能诱发恢复神经元原有的生理功能,形成持续的有节律的动作电位发放,并使神经元最终出现正常的自发的节律性的动作电位发放。而且,申请人分析认为,突触传递抑制一般是产生于兴奋传递通路的氨基酸能的神经元工作,其生理意义是依赖于突触可塑性而产生记忆效能,而对于本发明所涉及的下环振荡环路,属于激励脉冲发放,其生理意义是控制和维持内脏神经系统和内分泌神经系统的工作,并与上环振荡环路相互调制,这一环路的神经投射主要是胆碱能神经元,并不会产生突触传递抑制的问题。由于现有理论从来没有揭示存在这个下环振荡环路,更加没有资料显示其振荡工作的节律,(也即频率,即“脑干网状结构←→下丘脑”之间往返发放动作电位脉冲的节奏),但由于这一动作电位是作为内脏神经系统神经活动的激励信号,其节律要快于内脏神经系统的任何神经活动的节律,申请人根据:⑴、内脏神经系统最快的神经活动,也即心跳的心律正常是在1—2赫兹左右;⑵、大脑在昏迷或深度睡眠时,还能够检测得到频率小于4赫兹的脑电波(δ波),而下环振荡环路和上环振荡环路存在互相影响。所以,这一脑电波的频率是或者接近于下环振荡环路的工作节律,(申请人将这一节律称为生命基本节律),可以采用1—4赫兹的频率范围的电脉冲对下环路的神经元进行电刺激,对应的脉冲周期T为250—1000毫秒。
如果脑死亡是由于意外事故,比如外创伤、溺水、窒息等原因使大脑完全缺血缺氧所导致,且缺血缺氧的的时间不是很长,(最好十几分钟之内),神经细胞尚未失去生理活性,则通过采用上述方法对下环振荡环路的神经元进行电刺激,(并同步进行心脏起搏和呼吸机维持呼吸,或采用机械泵进行临时供血供氧),便能够诱发其神经元产生持续的有节律的动作电位发放,并最终使神经元恢复生理功能,使下环振荡环路产生自主的有节律的动作电位往返发放,实现大脑的重新激活。当然,如果大脑缺血缺氧超过一定时间,神经细胞已经失活,比如失去膜电位,失去生物泵的作用,则脑刺激便完全无意义了。
还有,如果仅仅是恢复下环振荡环路的脉冲发放,人体只是恢复内脏神经系统和内分泌系统的工作,如果这时上环振荡环路没有正常工作,则大脑还是处于深度昏迷状态,这显然不是我们所希望的,所以还需要同时恢复大脑的有意识思维。而且,根据分析,上下两个振荡环路的神经元活动在“脑干网状结构内侧区与脑干网状结构外侧区”之间存在相互调制,所以,在对下环振荡环路进行电脉冲刺激的过程中,同时对上环振荡环路的神经元进行刺激,有利于使两个环路互相调制,更快的恢复正常的工作。所以,在进行上述对脑死亡的激活时,可使用另外一台本发明的神经刺激装置,(或者在一台脉冲刺激装置中同时设置两套本发明的神经刺激装置),按照前面所述的用于对深度昏迷病人的唤醒的方法,同时对上环振荡环路进行脉冲刺激,在恢复生命基本活动时,还能够恢复有意识思维。
本发明根据大脑的工作机制及控制环路,分析了大脑昏迷和脑死亡的实质,并有的放矢地针对大脑特定位置的神经元,施加特定信号的脉冲刺激,能够对昏迷或脑死亡的大脑进行神经刺激,使其恢复思维活动或实现脑激活。另外,申请人希望通过专利申请文件公开的这种方式,将申请人关于人脑的思维、意识和注意的本质,人脑在神经元层面上的工作及控制机制,以及睡眠、做梦、梦游和精神病的一些研究结果进行公开,有利于社会对此进行进一步研究,并以此研发更多的医疗应用技术。
本申请文件所述的将各个输出电极置于大脑各个目标位置的方法,可以采用现有的脑电刺激的技术,比如:开颅直接将电极穿刺目标神经元,或在脑内目标位置预埋电极,更合适的方案,是在脑颅上开小孔、或者通过鼻腔或喉后颚开小孔,然后在神经内窥镜的配合辅助下,将输出电极插入脑内并置于目标位置上。而所使用的电极,可以采用现有用于脑神经的电刺激和电信号检测的电极,刺入神经元时采用脑神经专用的微电极,或膜片钳,只接触神经元膜外时采用普通电极。
本申请文件所述的电极,包括第一输出电极、第二输出电极,第三输出电极等,都可以是一只电极,也可以是一组(多只)电极,视工作需要而定。一般来说,对于周围神经,只需采用单只电极,而对于中枢神经,更合适的是采用多只电极作为同一输出电极,同时对同一位置的相邻的多个神经元进行脉冲刺激,有利于形成信号整合,产生神经活动。
附图说明 图1是本发明的神经刺激装置工作原理的结构示意图。图2是各个输出电极所输出的刺激脉冲的时序图。图3是一种对输出脉冲信号进行更合理控制的工作原理方框图。图4是另一种对输出脉冲信号进行更合理控制的工作原理方框图。图5是脑干网状结构与丘脑及下丘脑之间的上下两个振荡环路的神经投射示意图。
图6是生物体信息处理总结构图。图7是生物体内外信息处理系统示意图。图8是人脑内外感觉信息处理系统示意图。图9是思维通道的中间神经元连接结构示意图。图10是“思维→意识”的第一种神经元投射方式的示意图。图11是“思维→意识”的第二种神经元投射方式的示意图。图12是人脑视觉通道的信号投射结构示意图。图13是人脑听觉通道的信号投射结构示意图。图14是思维系统的振荡环路的工作原理示意图。图15是思维系统振荡环路的神经元投射示意图。图16是思维系统振荡环路的信号投射结构示意图。图17是“海马”中间信息处理环路的信号投射示意图。图18是运动神经系统控制环路的信号投射示意图。图19是内脏神经系统的信号投射示意图。图20是情绪系统的信号投射示意图。图21是隔区和基底前脑的胆碱能神经的信号投射示意图。
具体实施方式 以下为对本发明的原理和具体实施的叙述。
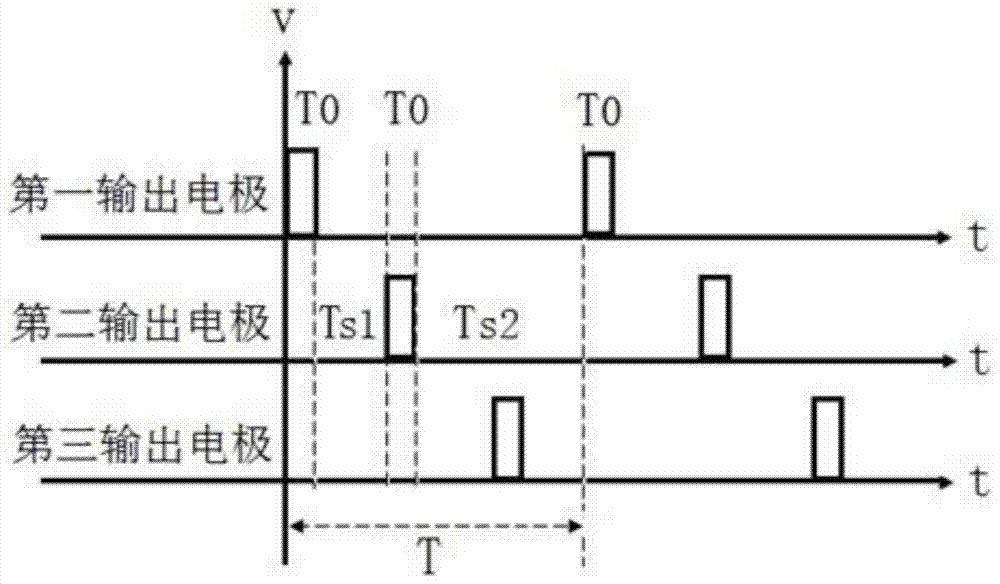
图1是本发明的神经刺激装置的结构示意图。包括主机和连接主机的电极,电极包括有基准电极(或称接地电极)和输出电极,主机包括有用于产生刺激脉冲信号的脉冲产生模块,连接各输出电极的用于将刺激脉冲信号输出到输出电极的各个输出模块,即第一输出模块、第二输出模块、第三输出模块,其特征在于:所述的输出电极包括有二组或二组以上的输出电极,图1为第一、第二、和第三输出电极,共三组输出电极;主机还包括有输出时序控制电路;输出时序控制电路用于控制各输出模块输出的刺激脉冲信号,使各输出电极所输出的相对于基准电极的刺激脉冲具有前后依次输出的时序关系。如图2。
脉冲产生模块属于现有技术,所产生的脉冲可以采用方波、三角波、梯形波、或者模仿神经元动作电位脉冲的波形。脉冲宽度T0为0.5—10毫秒,更合适的是1—5毫秒,相当于神经元动作电位的脉冲宽度。输出模块也属于现有技术,用于接收脉冲产生模块的脉冲信号,并输出到输出电极,其作用是隔离缓冲和调节输出脉冲的幅度。各输出电极输出的脉冲信号,相对于基准电极(也即接地电极),其峰值电压幅度,可在20—1000毫伏之间进行调节,根据电极接触位置及电极进入神经元的状态而有所不同。
本发明的主机还包括脉冲周期调节电路,用于调节同一组输出电极(第一、第二和第三输出电极)所输出的前后两个脉冲的脉冲周期T。对于不同的应用,脉冲周期T是不同的,一般在25毫秒至1000毫秒之间,见后面应用部分所述。
本发明主机还包括延迟时间调节电路,用于调节在同一脉冲输出周期中,前后两个电极(比如第一输出电极和第二输出电极)输出的前后两个脉冲之间的延迟时间Ts。
输出时序控制电路用于控制各输出模块的工作,使得各组输出电极输出的刺激脉冲的时序关系是:在一个脉冲周期T中,前一输出电极输出一个刺激脉冲后,经过设定的延迟时间Ts后,后一输出电极才输出一个刺激脉冲。比如:对于图1这样具有三组输出电极的装置,在一个脉冲周期中,第一输出电极输出一个刺激脉冲后,经过一个延迟时间Ts后,第二输出电极再输出一个刺激脉冲,然后再经过一个延迟时间Ts后,第三输出电极再输出一个刺激脉冲,依次类推。具体的控制方法可以是:脉冲产生模块产生一个脉冲信号,然后经过一个延迟时间Ts后,再产生一个脉冲信号,然后再经过一个延迟时间Ts后,再产生一个脉冲信号,而输出时序控制电路对三个输出模块进行依次的选通输出,将这三个脉冲信号依次从三个输出模块进行输出,从而在三组输出电极上依次各输出一个脉冲信号。(注意这时脉冲产生模块的输出脉冲频率是输出电极输出脉冲频率的3倍),这样的控制属于普通的电子技术,可以通过简单的MCU或者数字电路来实现。
本发明的神经刺激装置具有多组输出电极,各输出电极能够输出相对于基准电极的输出脉冲,且各组输出电极所输出的刺激脉冲信号具有前后依次输出的时序关系。使用本发明能够通过各组输出电极分别对某一神经通路上不同节点的神经元同时进行具有前后时序关系的脉冲刺激,强制神经通路产生具有信号定向传递关系的可持续的动作电位发放,从而可应用于激发和恢复某些具有生理功能的神经活动。(见后面公开的本发明的应用)。
作为一种重要的优化,本发明还对这种输出脉冲的时序关系进行精准的设置。其前后两组输出电极(比如第一与第二输出电极,第二与第三输出电极)输出的两个脉冲的延迟时间Ts,与脉冲周期T,脉冲宽度T0,三者具有如下关系:0<Ts<1/2T-T0;表述为:前后两组输出电极输出脉冲的延迟时间大于0,且小于“脉冲周期的一半减去一个脉冲宽度”的值。这种精准的设置基于神经元的STDP突触可塑性原理,(Spike Timing-Dependent Plasticity,即时序依赖的突触可塑性),根据突触传递的STDP可塑性,突触可塑性具有严格的时间依赖性,其突触传递效能与突触前后膜的脉冲峰电位的时序具有密切关系:如果突触前膜的锋电位早于突触后膜的锋电位,则产生突触传递增强现象,而且锋电位的延迟越小则突触传递增强效应越大;如果突触前膜的锋电位慢于突触后膜的锋电位,则产生突触传递抑制现象,而且锋电位的延迟越小则突触传递减弱效应越大。为了使本发明刺激的神经通路的前后神经元之间的突触产生传递效能增强,则需要保证在同一个脉冲周期(即同一次脉冲刺激动作)中,前一输出电极与后一输出电极所输出的脉冲的延迟时间,大于后一输出电极与前一输出电极所输出的脉冲的延迟时间。
参照图2的输出脉冲时序图,其推算过程如下:对于有两组输出电极的情况,则在一个脉冲周期T中,包含有两个脉冲宽度T0,和两段延迟时间Ts1和Ts2,为了确保第一输出电极输出的第一个脉冲与第二输出电极输出的第一个脉冲之间的延迟时间Ts1,小于第二输出电极输出的第一个脉冲与第一输出电极输出的第二个脉冲的延迟时间Ts2,则需要有2T0+2Ts1<T,依此推出有Ts1<1/2T-2/2T0,也即要求延迟时间Ts<1/2T-T0。对于具有三组及更多输出电极的情况,由于各输出电极是依次按顺序输出的,只要保证前后两个电极的输出信号满足上面所分析的情况即可,所以一样需要有Ts<1/2T-T0,又因为Ts需要大于0,所以有0<Ts<1/2T-T0。比如,脉冲周期T为50毫秒,脉冲宽度T0为5毫秒,则延迟时间0<Ts<20毫秒,比如可选10毫秒。
这样设置的意义是:以具有这样时序关系的刺激脉冲,分别对某一神经通路上前后不同环节的神经核团进行脉冲刺激,不单在发放刺激脉冲的过程中能强制该神经通路产生定向激活动作,从而强制产生信号传递的神经活动,而且,这种脉冲刺激具有特定的时序关系,能够使所刺激的前后两组神经元之间的突触产生STDP可塑性,尤其是LTP突触增强效应,提高突触传递效能。所以,这样的脉冲刺激,同时具有刺激神经元和定向提高神经元之间突触可塑性的作用,有利于恢复神经通路自身的信号传递效能,从而恢复其生理功能,所以可以用于一些工作异常的神经通路的康复。
在本发明的神经刺激装置中,其电脉冲串的输出特性也是很重要的。现有用于进行脑刺激的技术,其输出信号往往是采用跟其他人体电脉冲刺激或治疗技术(比如各种中高低频的电疗技术,电针技术,心脏起搏技术)一样,其脉冲波形是设定输出,也就是由脉冲发生模块调节一定频率、波形、占空比和电压幅度的正脉冲或正负交变脉冲,然后直接作用于目标刺激部位。现有这些电脉冲刺激技术,如果是用于刺激肌细胞或肌神经或心肌细胞上,只要具有足够的脉冲宽度和电压幅度,便能够有效刺激这些细胞以产生反应。但是,如果要用于对中枢神经的中间神经元的进行刺激并诱发连续的动作电位的发放,那就不一样了,这时便不仅仅是要求对神经元进行足够脉冲宽度和电压幅度的刺激便可以,不恰当的脉冲刺激不仅未能够诱发神经元发放动作电位,甚至还会抑制其生理活动,得到相反的结果。
根据神经生物学,由于离子运动的作用,会在神经细胞的膜的内外形成稳定的电位差也即静息电位,人脑的中间神经元的静息电位大约是-70至-90mV,也即神经元膜内电位比膜外电位低70—90mV,形成膜极化。要使神经元爆发动作电位,需要是其去极化,降低膜内外的电位差,当去极化达到或超过某一阈值,便可打开膜上的大量电压门控型离子通道,使正离子大量内流并形成正反馈,从而爆发动作电位。动作电位的发放时间一般只有几毫秒,之后,在动作电位发放完成之后,需要有时间供正离子外流重新恢复到原来的极化状态,才能够在下一次刺激时再次爆发动作电位。
根据动作电位的爆发机制,如果要通过外部电脉冲来刺激诱发神经元爆发动作电位,可以有二种方式:1、是将电极刺入神经元的细胞膜内(且不能与细胞膜外相连接),在膜内施加正电位的电刺激,当这一刺激产生的膜内电位的上升值大于触发阈值时,便能够使膜去极化并爆发动作电位。2、是将电极置于神经元膜外,但需要在膜外施加负电位的电刺激,这一负电位的电刺激拉低了膜外的电位,从而相对地也拉低了膜内相对膜外的电位差,当这一电位差的变化值大于阈值时,同样能够使膜去极化并爆发动作电位。所以,现有技术常常将电极置于某一脑部位,然后对电极施加强烈的正脉冲或正负交变脉冲,其导致的结果实际上并非诱发神经元产生正常的有序的动作电位发放,而是由于强烈的电位波动刺激神经元产生偶发性的动作电位,但之后往往会抑制神经元的离子正常运动,使其无法产生持续的动作电位发放,更加无法使其恢复自发的正常的神经活动。典型的,之前在脑刺激实验中,当对丘脑上的疼痛传递通路的神经元施加电脉冲刺激时,往往不是产生疼痛的感觉,而是抑制了疼痛,这也常被应用于作为脑刺激止痛(SPA),虽然并不知道其具体的止痛机制。
所以,申请人仔细根据神经元爆发动作电位的具体工作原理,对输出电极的输出信号进行优化,使得能够更准确地诱发神经元爆发动作电位,并且这种动作电位是能够持续发放的,更有利于受刺激的中枢神经元形成可持续的信号传递。
本发明的第一种输出脉冲信号优化方案,包括:所述的脉冲产生模块,带有正脉冲输出端和负脉冲输出端,能够分别输出相对于接地电极是正电位和负电位的两种脉冲信号;(这可以通过正负电源来产生,是普通的电子技术);正脉冲和负脉冲两个输出端通过切换开关进行切换,切换开关的输出再作为脉冲产生模块的输出端,连接到各个输出模块。切换开关可以是机械开关或电子开关。如果在实施脑刺激时是采用开颅的方式,并且采用专用于脑神经刺激的膜片钳或神经微电极,(这种微电极常用于神经解剖学的实验室中,用于检测神经元的电信号,采用金属丝或微玻璃管制成,尖端只有1微米或更小,且只有电极尖端处能够导电,可直接刺入神经元的胞体或轴突的膜内),能够将输出电极刺入所要刺激的神经元膜内,则可将切换开关切换到正脉冲输出端,使电极输出端连接到正脉冲输出端,这时输出电极是输出正电位的脉冲信号。这种方式是目前实验室进行神经元激活的惯用方式,能够可靠的刺激诱发神经元爆发动作电位,但要将微电极准确刺入目标神经元膜内,显然需要进行开颅手术,极其麻烦。所以,如果将本发明的切换开关切换于负脉冲输出端,使电极输出端连接到负脉冲输出端,这时输出电极输出的是负电位的脉冲信号,可直接施加于神经元的膜外,通过拉低膜外电位,来相对降低膜静息电位,并诱发动作电位。这时可采用普通的电极,并在脑颅上开小孔、或者通过鼻腔或喉后颚开小孔,然后在神经内窥镜的配合辅助下将输出电极插入并置于所需要刺激的目标位置上,只需使输出电极接触或靠近神经细胞的膜外,而无需准确刺入神经元,所以操作更加方便,创伤也较小。
另外,如果施加于神经元上的电脉冲刺激过于强烈,特别是当神经元已经被诱发动作电位后,仍然受到强烈的电刺激,则会严重影响神经元膜内外的正常离子运动,使神经元爆发动作电位后无法恢复正常的膜极化,所以也就无法进行下一次动作电位的触发。所以这样的电脉冲刺激反而会反过来抑制了神经元的活动,甚至使其失活,无法进行持续的整合和激活。(之前常常对疼痛传递通路的脑区域施加强的电脉冲刺激,用于进行止痛,实际上便是这个原因,因为这时其疼痛传递通路的神经元被强刺激所抑制甚至被损伤,无法传递疼痛的感觉)。所以,现有的电脉冲刺激技术输出的信号,可用于脑部刺激,但往往是神经元由于受到强烈的刺激而产生简单无序的放电,而无法产生可持续多次的正常动作电位发放,甚至是起抑制作用,可用于止痛或治疗各种神经元异常放电的疾病,比如癫痫,但要用于这种希望诱发神经元持续多次发放动作电位的应用场合,则并不合适,或者说要调整得到合适的刺激宽度和幅度,是比较困难的。
申请人对此进行改进,本发明的第二种输出信号优化方案,如图3,主机上用于连接各个输出电极(包括第一、第二和第三输出电极)的电极输出端,(电极输出端也即是主机上设置的用于连接主机外部各输出电极的输出端子,一般采用电气接插件),与各个输出模块(包括第一、第二和第三输出模块)的输出端之间,加有一输出控制电路,(相当于电子开关电路,更加合适的方案,在输出模块的输出端与输出控制电路之间还串接有一电阻器R,用于缓冲和产生一定的电压差);电极输出端还连接到一电压检测电路的输入端,电压检测电路的输出端连接到一电压比较电路,与来自输出模块的输出脉冲信号进行比较;电压比较电路的输出端连接到输出控制电路的控制端,通过输出控制电路对脉冲输出信号进行控制。当电压检测电路检测到的电极输出端的电压信号,与输出模块输出的电压信号之间的差值大于设定数值时,则关闭输出控制电路,在该脉冲周期余下的时间关闭输出模块到电极输出端的输出信号,直到下一脉冲周期才重新打开。由于第一输出模块到第一输出电极,第二输出模块到第二输出电极,第三输出模块到第三输出电极的控制电路是一样的,所以图3只画出一路。
按照这样的输出控制方案,工作时,在各个输出模块的每一个输出脉冲周期(即单个脉冲输出的周期),输出控制电路首先被打开,输出脉冲信号通过电极输出端和输出电极施加于神经元,同时电压检测电路检测电极输出端(相当于输出电极)上的电压信号,并与输出模块的输出脉冲信号进行比较,(允许两者具有一定的差值)。当输出电极的脉冲信号对神经元的刺激达到一定程度,神经元出现去极化,打开膜上的离子通道,大量离子内流并形成正反馈,使神经元爆发动作电位。这时由于大量离子急剧内流,形成超射现象,使神经元膜内电位由负电位变成正电位,膜外由正电位变成负电位,这一急剧的电位变化,会通过输出电极传递到电极输出端,造成电极输出端上的电压出现较大波动,而且是与输出模块输出的脉冲输出信号明显的变化不同步。这一电压波动被电压检测电路所检测到,并通过电压比较电路与脉冲输出信号进行比较,当两者的差别大于设定值(比如10%)时,可识别为神经元已经爆发动作电位,于是,电压比较电路的输出关闭了输出控制电路,在该脉冲周期余下的时间关闭输出模块到电极输出端的输出信号,直到下一个脉冲周期才重新打开。这样可使神经元在每一次动作电位爆发后不再受到外部电脉冲的刺激,使其膜内外的正常离子运动不受影响,能够恢复膜极化状态,以便对下一次刺激进行整合和再次爆发动作电位,形成可持续的多次的动作电位发放。这一技术方案,更符合中枢神经细胞的工作机制,既能够诱发神经元爆发动作电位,又不会过度刺激神经元以致使神经元受到抑制失活或受到损伤,用于刺激丘脑、网状结构、下丘脑等部位的神经元,能够持续诱发动作电位,形成连续的脉冲发放,并使神经元的离子运动和生理功能逐渐恢复正常,最终出现自发的同步脉冲发放。
还有,正如前面所述,跟其他脉冲电疗技术不同,在对中枢神经进行电刺激以诱发动作电位的可持续发放时,对电脉冲信号的刺激幅度是比较敏感的,既要足以使其去极化,但又不能太大以致抑制了其生理活性。申请人还注意到这里还存在一个问题:当将输出电极和接地电极置于不同脑部位时,由于大脑神经元周围的液体存在不同性质和不同浓度的正负离子,在未工作时输出电极与接地电极之间便存在一定的电位差,(可称为背景电压),而且这一电位差随两个电极的相对位置不同、以及所处的脑部位的不同,其电位差也不同。当在输出电极与接地电极之间发放电脉冲信号时,如果以一种设定的脉冲幅度进行输出,则这一原本便存在的电位差,便会对脉冲信号的幅度形成叠加影响,干扰了脉冲信号刺激强度的精确性。
所以,本发明对输出脉冲信号的第三种优化方案,如图4。其电极输出端连接有一电压检测电路,(可以与前面其他技术的电压检测电路共用),电压检测电路用于检测电极输出端的背景电压,所以也可称为背景电压检测电路,电压检测电路的输出连接有一电压保持电路,对电压检测电路输出的背景电压值进行保持记忆,(相当于电压记忆电路,如果电路是采用模拟电路,则可采用电容配合运放电路来构成,如果电路是数字电路,则可直接记忆其电压值),电压保持电路输出的电压值连接到一个电压叠加电路,脉冲产生模块输出的脉冲输出信号,在电压叠加电路中与背景电压进行叠加,(如果电路采用模拟电路,可采用运放电路,或者采用虚拟地的技术,对两者进行叠加,如果是采用数字电路,则将两者数值进行加减运算),再连接到电极输出端。电压保持电路、电压叠加电路和脉冲输出模块(包括第一输出模块和第二输出模块)的工作,受到一个状态切换电路的控制,(也可由主机总的控制电路进行控制),具体工作过程是:在每次进行脑神经刺激、将接地电极和输出电极置于目标区域后,或工作中重新调整输出电极和接地电极的位置时,状态切换电路将电路工作切换在“检测”状态,关闭输出模块的输出,先由电压检测电路检测电极输出端上的电压,(这一电压也即当前的输出电极与接地电极之间的电位差,也即背景电压,可能为正也可能为负),其输出电压值由电压保持电路进行保持(也即记忆),并送到电压叠加电路上;之后,由状态切换电路将电路工作切换到“输出”状态,关闭电压保持电路的输入端,(使其输出电压保持之前输入的电压值),打开输出模块的工作,输出模块输出的脉冲信号,在电压叠加电路中与背景电压进行叠加,再连接到电极输出端。这样,输出电极所输出的脉冲信号,便已经将两电极之间的电位差进行抵销,可消除背景电压对脉冲信号刺激效应的影响。
本发明的应用,包括用于对昏迷病人的唤醒和用于对脑死亡的激活,已经在“发明内容”部分进行充分和详细的叙述,(可参照图5),这里不再重复。其理论依据可参考下面的附带资料。
附带资料:关于思维和意识的本质及人脑工作机制的研究资料。以下部分是申请人对于大脑思维、意识、注意的本质及其神经元层面上的工作机制的研究、分析和描述。这些内容可能跟本发明技术方案没有直接关系,但有助于理解本发明及其应用的工作原理和设计依据。
在大脑的神经活动中,最基本的动作就是神经元的动作电位的发放。某个或某组神经元的活动,通过突触传递对另一个神经元形成兴奋性刺激,当这些兴奋的累积超过某一阈值,也即神经元的膜电位整合超过其动作电位的触发阈值,则该神经元便激活爆发动作电位,并通过轴突向下一神经元形成刺激,这是大脑活动的基本动作,大脑也是以此来实现各种信息处理的功能。问题在于:大脑如何通过这种神经元的基本活动来实现各种脑功能?比如记忆、思维、注意、意识?也即是,如何用神经元的活动动作来描述各种脑功能的工作机制?
⒈大脑的起源及进化。要了解大脑的工作机制,我们首先需要了解大脑思维活动的来源,了解大脑是如何随着生物进化而出现和进化形成的。
⒈⒈生物进化的“简单原则”。不管大脑如何复杂和神秘,如果我们承认人和动物都是在地球的自然环境中进化而来,这一进化过程没有受到非自然力量(神)的操纵,那么,我们似乎应该认定这一点:大自然没有设计师进行事先设计,大自然的一切物体都是物质在自然环境下产生变化,并在漫长的时间中慢慢累积这些变化而形成的。而且造成这些变化和累积的原理都应该是简单和自然的。大自然遵循这种“简单原则”,利用简单的道理,通过漫长的积累,来形成各种复杂和巧妙的物体。
比如雪花,虽然精美和巧妙,可我们知道那只是水分在自然环境的冷与热变化中不经意地自然形成的。比如自然风光,不管如何壮观和精致,可我们知道那只是岩石和土壤通过大自然长时间的风化溶解而形成的。比如花朵,不管如何美轮美奂,可我们知道那也只是多种植物细胞经过分裂生长形成的。同样的,人类的大脑不管结构和功能如何复杂,那也应该只是神经细胞根据某种简单原理进行连结,并在漫长的进化中慢慢累积,而进化并形成目前这种结构和功能。
⒈⒉从生物进化进程来看,大脑(人脑或动物脑)的形成和进化是跟随生物体的进化过程而相应变化的,它是动物处理各种外界信息及机体内部信息、并对自身各种活动进行控制的神经元的组织集合。
当生命从一些分子团进化为单细胞生物,如细菌,这时没有也无需反射系统的,只是按照DNA所固化的程序进行分裂复制。
当生命进化为具有简单结构的生物体,具有最基本的吞食和吸收功能,也具有了生物生存所必须的移动功能(为了进食和逃避被食)。这时需要对这些消化和移动的生命活动进行有效控制,于是进化出某种中间细胞来连接两种不同功能的器官组织,使得一种器官组织的受激,能够引起另一种器官组织的应激反应,达到吞食或移动的动作。这种连结不同器官组织的中间细胞,便是神经细胞。这时的生物依靠简单和直接的反射弧和反射动作来实现对刺激输入的反应控制,这是最原始和简单的神经系统。
随着生物进化出各种生命系统,包括消化系统、循环系统、呼吸系统、内分泌系统等,这时生物体也进化出相应的神经组织来对这些系统的工作进行感知和反射处理,接收它们工作时的各种信息即机体内部信息,并通过长期进化形成的记忆结构(记录在基因的、可遗传的)来控制它们的工作。这些神经反射组织聚集在一起,并且彼此之间通过互相影响和调制,来协调控制各个系统的工作,形成一个机体内部信息的处理系统,它们大多处在脊髓与大脑的下部,是大脑早期形成的部分。
而随着生物对各种外部信息(图像、声音、机械刺激、气味等)的感知及反应的需要,生物体进化出视觉、听觉、触觉等、嗅觉等各个外部信息的感知系统,进化出相应的神经组织来对这些外部信息进行接收、设别、记忆和反射,这些处理外部信息的神经组织各自集中在一起,构成了对应于各种感觉的反射区,包括视区(optic zone),听区(acoustic area),躯体感觉区(somatic sensory area),等等。同时,外部环境和信息的复杂,也使生物需要更复杂的活动来适应环境,从游动到爬行到飞行到奔跑,到各种更复杂更精准的机体动作,生物也进化出相应的神经组织来对机体的肌肉动作进行协调控制,这些对肌肉动作进行协调控制的神经组织聚集在一起,形成了包括小脑等的运动神经系统,而如何根据各种外部信息,来反应及控制机体运动的神经组织的聚集,便构成了皮质上的各个运动分区。各种感觉系统及运动神经系统,共同构成了生物体的外部信息处理系统,用于接收各种外部信息,并通过长期习惯化的记忆结构来反应并控制生物体的运动动作。
机体的内部信息处理系统与外部信息处理系统彼此之间显然需要和存在联系和相互影响。比如当机体外部信息处理系统进行更剧烈的运动时,需要内部信息处理系统提供更多的氧气和营养;当内部信息处理系统感知到饥饿时,需要外部信息处理系统进行觅食的行动。所以从总的方面看,大脑实际就是生物体的内部信息处理系统和外部信息处理系统两个系统的集合,并存在一种相互调制的通道,来对两个系统进行相互影响和调制。于是构成如图6所示的生物体信息处理的总结构图。
随着各种信息的丰富和复杂,生物体需要同时接收多种内外信息的输入,对这些信息的反应也不能只是简单反射,而必需整合各种信息并根据长期形成的适应性习惯(即记忆)来做出一个平衡各种信息的反应。比如,食物的气味信息让生物要前往,而食物附近的视觉的危险信息让生物需要逃避,那么生物需要对前往和逃避的信号强度进行比较和整合,最终只能输出一种行动——前往或逃避。所以,生物体进化出对各种输入信息和中间信息进行比较、切换、整合的联合处理部分,其中对外部信息的联合处理部分便形成了所谓的思维系统。如图7所示的生物体内外信息处理系统示意图。(为了简洁,图7只标出内外各两条信息处理通道)。
⒈⒊于是,生物进化的结果,对信息进行反射的各种神经组织的集合,便构成了脑。由于各种内部和外部信息的复杂甚至矛盾,大脑进化出大量的中间神经元来连接各种感觉输入的神经组织,通过这些中间神经元来比较、切换、协调和整合各种信息,这一部分形成了大脑的脑干网状结构、丘脑、海马、底丘脑、纹状体、小脑、上下丘脑等脑组织。之后,随着各种信息进一步复杂,部分核团进一步延伸和膨大,形成大脑皮质,包括端脑。如图8所示。(为了简洁,图8只画出部分信息通道)。
而随着动物的群居生活和生存环境的复杂化,为了更好的生存和生活,动物进化产生了用于交流信息的语言。这里的语音是广义的,包括声音的语言、肢体语言,和各种生物信息的语言如气味。对于人类,还比较特殊地产生了丰富的语言以及对应的文字。对于这些语言和文字信息的输入输出和处理,大脑也是通过进化出了一系列的中间神经元的集合来实现。包括:位于颞上回后部的听觉性语言中枢(听话中枢),用于对听区所接收的声音信息中的语言信息进行识别。位于额下回后部的运动性语言中枢(说话中枢),控制口腔和咽喉的肌肉实现说话。位于顶下小叶的角回上的视觉性语言中枢(阅读中枢),对视区所接收的视觉信息中的文字信息进行识别。位于额中回后部的运动性文字中枢(书写中枢),通过控制手指肌肉实现文字的书写。
由于语言信息和文字信息是如此之丰富,它们构成的信息量十分巨大和复杂,对听话、说话、阅读、书写各个中枢的输入输出和信息重组的工作,也在皮质上形成了大量的中间神经元,用于连结这几个中枢,对这些中枢的神经元所输入或输出的语言和文字信息进行连接、记忆、重组、反应、输出,等等,形成成串成段的语言化的信息链。比如,将听话中枢输入的语言,或阅读中枢输入的文字,保存记忆在皮质的中间神经元中,并建立起它们之间的关联关系,在下次听到这些语言或看到这些文字时,能够激活皮质原有保存记忆的那些神经元,以此来识别输入的语言和文字的意思(词义)。另一方面,当大脑联合皮质的中间神经元在活动(即思维)时,也可通过回馈投射到说话中枢或书写中枢的神经元,通过这些神经元的激活输出,来将大脑思维的信息串进行输出,形成说话或书写。
⒈⒋所以,大脑的工作,便是采用神经元来感知、记忆和反射各种输入和输出信息,以及采用中间神经元来对各种输入输出信息和中间信息进行整合处理,以便对机体的各种输出进行协调控制。随着机体与环境关系的发展和机体自身构造趋向复杂和精细,大脑也出现相应的复杂和精细的进化,并由于各部分之间产生交互调制而使其信息处理的链路变得复杂。其中随着各种感觉信息和运动动作的复杂化和精细化,尤其是语言化的感觉输入和运动输出,以及对这些信息的整合处理也即思维和记忆,使大脑皮质尤其是端脑的神经元数量相应地快速增加,体积膨大成为人脑最大的部分。
描述大脑进化过程这些现有知识,目的在于表达这一点:从生物进化的角度看,人脑在进入哺乳动物、爬行动物甚至在进入鱼类时所形成的各个神经核团及其信息处理机制(类似于图8所示),便已基本不变。人脑中在早期便形成存在的部位如脑干网状结构,在大脑的系统工作中仍然是核心和主导地位,而一些后期进化形成的部位如皮质尤其是端脑皮质,虽然体积巨大,但只是之前的小核团由于信息增加导致神经元增加而膨大形成,其在人脑的系统工作中仍然是从属的地位。(正如我们不能因为大腿粗大发达便认为双腿在人体进化过程中取代大脑成为人体生理活动的核心和主导)。所以,虽然信息的识别和认知、思维和记忆、运动输出等大脑高级功能的神经活动大多发生在皮质尤其是端脑,但对这些神经活动的控制,也即整个大脑系统工作的控制机制的核心,不在于皮质,不在于端脑,而在于生物进化最早期便存在的脑干和间脑一带,包括脑干网状结构、丘脑、上下丘脑等。(大脑的控制机制是重点,将在后面第3部分,即“⒊大脑思维活动的控制机制”进行详细描述)。
⒉思维和意识的本质。要分析思维和意识,也需要了解认知和记忆,这是思维的两个基础和必要前提。认知是大脑对各种刺激的感知和识别,并通过记忆将经过认知的信息保存下来,之后,大脑对经过认知的信息和经过记忆的中间信息进行反应和重新整合的过程,才形成思维。
⒉⒈记忆的本质和工作机制。申请人在2014年10月30日提交的申请号2014106066977的中国专利申请“神经网络的模拟装置及方法”的说明书中,对记忆的本质、形成、短期记忆与长期记忆的不同及转化,进行描述。在脑神经元网络中,记忆的本质便是神经元的“唯一性反射”。当一个神经网络得到某一个信息(最小最基本的信息元)输入的刺激,也即得到一个或一组神经元激活的动作电位的传入刺激时,该神经网络的连接结构,使得它有且只有另一个或另一组神经元得到充分的兴奋整合,激活爆发动作电位并输出新的信息元,这便是记忆的基本的神经元活动。在脑,外界信息的识别和记忆存在于大脑皮质,而对各种中间信息的记忆先发生于信息传递的中间通道,(如海马),经过多次刺激后才在大脑皮质形成长期记忆。具体工作可参考之前的申请文件,这里不再详细描述。
⒉⒉认知的工作机制。认知也是思维的基础和前提,或者说是思维的前一道工序。传统上认知与思维有时会被混为一起,但实际上认知与思维是两个不同的过程,并采用有差别的工作方式来实现。认知活动存在于各种感觉神经元和初级感觉皮质上,采用并行的方式进行信息处理,而思维是皮质尤其是联合皮质的中间神经元,对认知的输出信息、以及思维自身产生的中间信息进行发射、反应和重新整合,并输出控制信息或新的中间信息的过程,采用串行的方式进行工作。
关于认知尤其是关于视觉、听觉和触觉的认知过程,有较多的相关文献对其神经链路和工作进行公开,这里只简述认知的并行工作方式。认知是大脑对外界的认知对象进行感知和识别的过程,由于认知对象具有复杂多样的信息,为了实现对它们的识别,需要同时感知它们的多个信息,来共同限定一个认知对象,所以,认知过程的感知和识别采用的是信息的并行工作方式,然后通过多对一的投射来限定并识别认知对象。比如,对于一个物体的视觉认知,由于物体的图像包含有大小、形状、颜色、亮度等多种信息,要根据能够实现识别的需要,同时对其中一种或多种信息进行感知,而每种信息又具有不同的值,还可能需要对其值的大小进行感知,最后再根据这些感知的信息来共同限定识别出这个物体。具体的流程大概是:⑴、大脑通过眼睛的几种感觉神经元,来感知图像的形状和大小,颜色信息,亮度信息,并通过动作电位发放的快慢和时序,也即频率编码和时间编码来输出传递这几种信息的值,这种编码方式有利于减少和简化信息传递的神经通路。⑵、视觉输出信息被投射到外侧膝状体和初级视皮质上。这种投射是多种信息通路都进行投射,但在每一种信息通路上是一对多的投射关系。初级视皮质具有非常严密和有序排列的陈列结构,能够根据每一视觉通路输入的神经元动作电位的发放快慢和时序,来使排列结构中不同数量和不同位置的神经元得到激活,也即是将每一种视觉输入信息的频率编码和时间编码,转化为空间上不同数量和不同位置的一组神经元活动的空间编码。⑶、初级视皮质投射到联合视皮质。这种投射是空间位置上多对一的关系,形成高度概括的空间编码。即初级视皮质的多个(或多组)神经元的共同激活发放动作电位,共同对联合视皮质上的一个(或一组)神经元进行协同刺激,通过兴奋的膜整合使这一个(或一组)神经元得到激活发放。从而实现将对应着多种信息以及不同值的多个(多组)神经元,来共同映射和限定一个(一组)神经元,而这一个(或一组)神经元,便唯一性的对应着原有的认知对象,从而实现对认知对象的识别。⑷、如果视觉信号涉及文字,则联合视皮质上的神经元还向视觉性语言中枢(阅读中枢,阅读中枢同样属于联合视皮质)进行投射,对文字进行识别。⑸、联合视皮质神经元的发放活动,成为经过认知的信息输出,通过中间神经元尤其是联合皮质的中间神经元的活动,进行思维和各种反射反应活动。
至于听觉、触觉及其他感觉的认知过程,跟视觉是类似的。其中听觉信息由耳蜗毛细胞通过听神经进入脑干,在丘脑内侧膝状体中继,向初级听皮质投射,再由初级听皮质向联合听皮质投射,实现对声音的识别。如果听觉信息涉及语言,则联合听皮质还向听觉性语言中枢(听话中枢)投射,对语言进行识别。不过,从脑功能来看,联合视皮质应该有直接向运动皮质的神经投射通路,而联合听皮质没有这一投射,所以大脑能够无需注意和思维而直接根据视觉信息来控制机体的运动动作,但听觉不能。另外,包括视觉和听觉的各种感觉皮质似乎都有一个共同投射的区域,使同一个认知对象的各种感觉信息得到交叉对应。比如,对某个人的形象识别信息、跟名字和声音进行对应,当听到声音时便可忆起其名字和形象。而阅读中枢和听话中枢的交叉投射的区域,也即是所谓的语义皮质区。
⒉⒊思维的本质。而皮质尤其是联合皮质的中间神经元的活动,如果在各种调制条件下,形成了受控的、有序的、依次的、逐个(或逐组)步进式的激活,也即链式激活,(激活也即是爆发动作电位),便是思维的神经元活动的基本形式。具体说,如图9所示,某一神经元比如神经元a的轴突可以与数以万计的其他神经元的树突或胞体建立突触联系,另一神经元比如神经元b的树突也可与数以万计的其他神经元的轴突建立有突触联系,这些突触结构是之前通过各种信息刺激所建立起来的,(包括直接的信息记忆和其他的间接联系),使得当神经元a被激活而产生动作电位时,通过突触传递给神经元b施加兴奋,而神经元b在其他神经元协同的兴奋激励下,(也即兴奋的空间整合,包括调制性神经元的调制性作用,和同步脉冲的协同激励,在后面有详细叙述),也被激活而爆发动作电位,即有神经元a→神经元b。同样的,神经元b的激活,其动作电位发放会对其他神经元比如神经元c,神经元d等形成兴奋刺激,使得后面的神经元c,神经元d也被依次激活,形成神经元a→b→c→d→……等的链式激活。由于每一个或每一组神经元对应着某一最基本的信息也即信息元,比如神经元a对应信息元A,神经元b对应信息元B,神经元c对应信息元C,等等,则神经元a→b→c→d→……这一激活链所对应的信息元,便形成一串依次输出的信息链A→B→C→D→……,这种在神经网络中神经元产生受控的、有序的、依次步进的链式激活动作,便是思维最基本的神经活动形式,也即是在神经元层面上的思维的本质。这里所述的神经元,可以是一个或一组,是对应和限定一个信息元的基本神经单位。所述的激活,是神经元爆发一次或连续多次的动作电位,由于突触易化的原因,很多神经元的激活发放可能是连续多次的。
⒉⒋思维的工作方式。思维的工作方式,也即是神经元如何以兴奋信号的传递来进行链式激活以实现信息处理的方式。由于神经元之间的突触联系是数量巨大且结构复杂的,神经元a激活而导致神经元b激活的原因是多方面的,这可以是以下几种。
⒉⒋⒈因为信息记忆而建立的突触联系。即之前曾经多次输入过A→B的信息,而在神经元a和神经元b之间产生建立了直接且传递效能足够强大的突触联系,(突触传递效能一般是由于彼此之间形成的突触数量,和每个突触的传递效能决定的),使得神经元a被激活并对神经元b进行刺激时,神经元b由于得到足够的兴奋整合而触发产生动作电位,产生神经元a→b→的反射链,这种形式的思维输出,实质也是对原有所记忆的信息的忆起过程,其更详细的工作可参考2014106066977号中国专利申请的叙述。
⒉⒋⒉调制性神经元的调制作用。来自情绪系统、运动神经系统等其他神经活动,其所产生的调制性神经元对神经元b的调制作用,包括增强性调制的增强,或抑制性调制的减弱,使得当神经元a激活时,神经元b由于突触整合而被激活,产生神经元a→b→的反射链。(具体工作后面另有叙述)。
⒉⒋⒊来自同步激励脉冲的协同激励作用。特别是来自思维系统“注意”控制环路的作用。神经元a的激活,通过轴突向大量的后神经元发放刺激,但显然这些后神经元并非都被激活,只有当思维“注意”控制环路的同步脉冲发放指向神经元b所在的区域时,神经元b才被同步脉冲的协同刺激整合而激活,产生神经元a→b→的反射链。而如果“注意”控制环路的同步脉冲是发放向其他神经元的区域,则可能是另一神经元比如神经元f被激活,产生神经元a→f→的反射链。应该说,在同一情绪和同一生理状态下,大脑的思维活动,更主要地便是受到“注意”控制环路的同步激励脉冲的控制,而也正是同步脉冲使神经元的活动得以形成同一通路中有序的“步进”式的链式激活而不会混乱,同步脉冲起到类似于电子技术领域的中央处理器中时钟信号的时序控制作用。“注意”的工作机制是大脑工作的重点,将在后面第3部分,即“⒊大脑思维活动的控制机制”进行详细描述。
⒉⒋⒋各种兴奋激励的综合作用。由于神经元之间的突触联系是庞大且结构复杂的,神经元链式激活的产生条件和途径,可以无需彼此之间建立有直接的固定的反射联系,而是因为其他间接联系的中间神经元的共同整合而触发激活。比如,神经元e与神经元f之间本来并没有直接且固定的反射关系,(也即没有信息记忆关系),所以,虽然神经元e与神经元f之间建立有传递效能较弱的一般突触联系,(相关的神经元之间由于受间接的信息的刺激,彼此之间建立有广泛的一般的突触联系,而如果彼此之间建立的突触数量较少,和每个突触的传递性能较低,则这种突触的传递效能便较弱),这种弱的突触联系所能传递的兴奋作用较弱,本来神经元e的激活不足以导致神经元f也产生激活,但是,如果在神经元e激活之前,已经有神经元a、b、c、d的激活,(也即相关信息的神经元激活),多次给神经元f以刺激,这些刺激多次使神经元f发生膜兴奋,并形成膜整合,(这也是工作记忆的产生原因之一,工作记忆是由于突触传递的膜整合和突触短时程可塑性而产生的),且同时还有调制性神经元的调制作用,以致于当神经元e激活发放、且同时伴随着“注意”控制通路的同步脉冲的激励时,神经元e传递给神经元f的弱的兴奋性刺激,已经足以使神经元f突触整合后的兴奋值超过动作电位的阈值,从而触发动作电位,产生神经元e→f→的反射链。在这种情况下,神经元e的兴奋刺激起到最后的关键作用,(最后一根稻草)。这种方式所导致的链式激活,是大脑思维的主要形式,也是大脑能够进行自由思维的原因,它无需前后信息在之前建立有直接的记忆关系,而是通过整个信息结构来整合产生,所以,它并非信息的再次忆起,而是可产生之前没有的新的信息链的输出,产生“自由”的思维。当然,所谓“自由”的思维是相对的,是指能够产生之前没有的新的信息链,但其产生依然是受到其他各种间接信息的限定。在面对信息输入时,链式激活的进行方向也即思维的思路,受到相关神经元的连接结构和多个调制通路以及“注意”控制通路的共同影响。其中相关神经元的连接结构也即是原有相关信息的记忆结构,包括知识、事件、经验等,这是形成智力的基础;而“注意”控制通路和各个调制通路的作用强弱在不同人也会有差异,这是形成不同“性格”的主要原因。
⒉⒋⒌神经元的自发放电。大脑皮质的神经元,由于彼此之间的突触联系是广泛的,所以,在思维通道的激活链之外的其他神经元,在某一时刻,也可能有部分会由于不断受到各种兴奋刺激而产生膜整合并触发动作电位,产生所谓的自发放电。但申请人推测,由于这种自发放电是偶发性的,又未能与包括同步脉冲等其他兴奋激励形成协同整合,一般未能再次激励下一神经元并形成链式激活,所以形成不了思维活动,而只是孤立的自发放电。但这种自发放电也是有其生物学意义的:由于神经元偶尔的自发放电,能够使神经元的胞体及突触保持一定的生理活动,维持其生理活性。而且由于接收具有突触联系的相关神经元的兴奋刺激,而其自发放电又对这些相关神经元形成刺激,所以能够“刷新”与这些神经元的突触联系,提高其突触联系的有效性,也即是增强了依赖这种突触联系建立起来的记忆结构。这种机制应该是存在大脑的其他系统比如运动神经系统。而且,这种机制似乎在睡眠时也可以发生作用,所以,有时我们会发现,经过反复学习或练习的信息或技能,即使过后没有再去学习或联系,有些在几天后会变得更清晰或更熟练。
⒉⒌意识的本质。意识是大脑最神秘的功能之一。申请人根据大脑在信息处理中的链路和流程,分析认为:意识是大脑对思维活动的自我感知。当大脑进行思维时,也即大脑联合皮质的中间神经元网络在进行思维活动时,思维通道的神经元的激活,其轴突一方面向前面其他神经元形成兴奋传递以继续激励思维活动,另一方面还同时向皮质的联合感觉区的神经元形成投射,使得思维通道的神经元的活动被皮质感觉区所感知,于是,大脑便感知和意识到“我”的思维和存在。这种联合皮质的神经元活动向感觉皮质神经元的投射,使感觉皮质能够对联合皮质的思维活动产生自我感知,这便是意识的本质。在正常人,以及在正常的清醒状态下,这种接受投射的感觉皮质似乎主要是听觉性联合感觉区,尤其是语言中枢,所以我们日常“意识”到的“思维”大多是以语言进行。(对于先天性聋哑人,由于其无法形成声音感知,而神经网络又具有可塑性和代偿性,则可以视觉性的手语来进行,这种手语也属于语言)。当然,我们所说的听觉性感觉区和视觉性感觉区,都是人为定义的,实际上都是以中枢神经的中间神经元来实现的。这一思维过程向感觉皮质的投射和自我感知的生物学意义,应该是将思维过程产生的各种中间信息,作为新的信息,投射到感觉区进行感知和输入,以便对这种中间信息进行记忆或与原有信息进行重组,(这种记忆首先表现为工作记忆,之后在某些情况下再转化为长时程记忆和长期记忆),以此不断完善其记忆的信息结构。但正是由于这一重新投射,使得思维过程能够被大脑的感觉皮质所感知,使我们“知道”和“意识”到思维的内容和过程,才使我们产生了这种奇妙的“意识”。如果没有这种重新投射感知的通路,则大脑便只是对输入信息进行反射和反应输出,而没有“知道”和“意识”,也不会自我完善信息结构,便只是属于初等生物的神经反射机制,或者说更像是一台自动化机器。显然,这也是目前的人工智能与人脑的本质差别之所在:目前的人工智能只具备对外部输入信息的感知、识别、处理、反应和输出的能力,而缺乏对信息处理过程的自我感知、和将信息处理过程所产生的中间信息实时地整合到原有信息结构的能力,也即缺乏“自我意识”和“自我完善”能力。显然,没有对人脑“意识”的本质的认识和理解,要设计出真正具有“意识”的人工智能就根本无从谈起。申请人按照上述人脑“思维”和“意识”的本质和工作原理,还有后面将描述的关于思维的“注意”控制机制,并以巧妙的方式解决了其中最困难的将中间信息实时整合到原有信息结构中的“自我完善”的技术,设计了全新工作原理的人工智能系统,将在之后提出的“具有自我意识的人工智能系统”的专利申请中进行公开。
申请人还认为:大脑存在来自下丘脑或脑干的调制通路,即睡眠与觉醒的调制通路,使得正常人在清醒状态下,联合皮质的中间神经元在进行思维活动时,其激活链的神经元主要是向联合听觉皮质进行投射,而向视觉及其他感觉皮质的投射被抑制,所以大脑对思维的自我感知和意识,主要是以听觉性感觉来实现的。这主要是因为视觉皮质还存在有向运动皮质的直接投射以进行无意识的日常活动,而听觉没有,如果思维活动的视觉信息还向视觉皮质进行投射,则可能会造成“视觉—运动”反应的混乱。只有在非清醒状态下(睡眠做梦,或者某种精神病如精神分裂症),对视觉皮质的抑制被异常地解除,思维活动所产生的信息才会向视觉皮质进行投射,使大脑产生可感知的图像幻觉,就像看到的一样。(关于做梦和精神分裂症,后面另有描述)。
⒉⒍意识的产生方式。意识的产生方式,也即思维过程中联合皮质思维通路的中间神经元向皮质感觉区的回馈投射方式,这可能有两种方式:
⒉⒍⒈第一种方式是思维通路的中间神经元直接进行链式激活,比如神经元a1→b1→c1→的激活链,其活动不受皮质感觉区的影响,只是在这一过程中,神经元a1、b1、c1同时发侧枝向皮质感觉区进行投射,使其对应的神经元比如神经元a2、b2、c2感知这一思维过程,即产生a2→b2→c2→的自我感知,从而产生自我“意识”。在这种方式中,思维是由联合皮质的中间神经元直接产生的,感觉皮质只是被动地感知和意识到这种思维活动。其神经元投射方式如图10所示。
⒉⒍⒈第二种方式是思维活动的链式激活是在联合皮质神经元与感觉皮质神经元之间往返交叉进行的,其神经元的投射方式如图11所示。其过程包括:⑴、思维通路的神经元的每一步激活,比如神经元a1激活,便同时向下一神经元b1和感觉皮质的对应神经元a2进行投射激励;⑵、感觉皮质的神经元a2受触发激活感知这一信息;⑶、感觉皮质神经元a2的激活,又反过来向联合皮质的神经元(包括有神经元a2)进行投射激励;⑷、联合皮质的神经元b1同时接受a1和a2的激励,触发激活;⑸、神经元b1的激活,又同时向下一神经元c1和感觉皮质的对应神经元b2进行投射激励;⑹、感觉皮质神经元b2激活感知这一信息;⑺、感觉皮质神经元b2的激活,又反过来向联合皮质的神经元(包括有神经元c1)进行投射激励;⑻、神经元c1同时接受b1和b2的激励,触发激活……;最终,联合皮质思维通路同样出现了神经元a1→b1→c1→的激活链,而感觉皮质的神经元也同样产生了a2→b2→c2→的自我感知而形成意识,但这一过程是以a1→a2→b1→b2→c1→c2→的往返投射的方式进行的。在这种方式中,感觉神经元是实时感知思维活动的每一步,并参与并限定了思维活动的下一个激活动作。
⒉⒍⒉由于缺乏详细和可靠的解剖资料,申请人无法直接确定意识感知是以上述哪一种方式进行,但申请人通过一种特殊的思维,即语言化的思维来对此进行分析和推测。在人类,思维大多时候是以语言化进行的,只有少数特殊情况才有非语言化的思维。人类的语言化思维一般是以第一母语进行的,但也能够以外语包括第二母语来进行,这是申请人认为意识是思维通道向联合感觉皮质投射的原因,而且这为我们分析思维向意识的投射方式提供一种途径:如果思维向意识的投射是第一种方式,即思维活动是联合皮质的中间神经元独立进行,联合感觉皮质(语言的感觉识别也是属于感觉皮质的联合感觉区)只是被动感知这种活动,那么,不管是以母语还是以外语进行思维,思维活动都应该能够正常顺畅地进行并产生思维结果,只是思维有时不一定能够被可靠感知和意识到;相反,如果思维向意识的投射是第二种方式,即思维是在联合皮质与感觉皮质的神经元之间往返进行,那么,在以不熟练的外语进行思维时,思维活动必然会不顺畅,进行到不认识的外语词语时会受阻,无法产生思维结果。实际情况是后者,大脑在以不熟练的外语进行思维时,常常不连贯,受阻而难以进行下去,所以申请人推测,大脑的思维,至少是语言化的思维,是在联合皮质思维通路的中间神经元,与感觉皮质的神经元之间,往返进行链式激活的。当然,这一工作过程还同时受到同步脉冲和其他调制信息的调制影响。(见“⒉⒋⒊来自同步脉冲的协同作用”。还有后面第3部分“大脑思维活动的控制机制”)。
⒉⒍⒊如果思维与意识是以这种方式进行,即在思维通路和感觉皮质的神经元之间往返投射来实现,由于思维活动需要“注意”控制环路的同步脉冲的协同激励,那么,在语言化的思维活动中,神经元活动的每一个步进,便需要更多次的同步脉冲来协同激励,思维通路的神经元激活至少需要一次,感觉皮质的神经元激活至少需要一次,(这里说至少,是因为神经元有时需要不止一次的同步脉冲激励才产生一次激活发放)。而我们知道,大脑活动的同步脉冲,可以被检测得到,也即脑电波。所以,在语言化的思维活动中,即使是在最紧张最快的思维中,思维的速度,即一定时间内能够意识到的思维活动的词语的数量,最快不会超过脑电波节律的一半。这可以通过设计实验来进行验证。
⒉⒍⒋申请人认为,思维和意识以这种往返交叉投射来进行的方式也有其生物学意义和合理性。思维过程的每一步活动都实时投射到感觉皮质,使大脑及时感知和整合这一信息,感觉皮质的感知再重新投射回到思维通路中,对思维活动进行重新影响和限定,促使思维在相关信息中更精准地进行。而且,从大脑的解剖结构来看,大脑皮质具有数量巨大的密集排列的神经元,(尤其是椎体细胞和颗粒细胞),而大脑中任何直接的神经投射链路的神经元的数量(换元数)都不多,大多只是2个到5个神经元的投射链路,但我们的思维活动却能够输入、输出和整合长度惊人的串行的信息链,比如,记忆或背诵长达上千字的文章!显然,大脑正是通过在神经元数量巨大的感觉皮质与联合皮质之间进行往返投射的活动方式,来记忆和输出这种字节数巨大的成串的文章的。其中,感觉皮质记忆了语言和文字的单个的词语,而联合皮质记忆了它们之间的联结关系。当我们阅读并记忆一段文章时,是通过改变联合皮质(包括海马结构的皮质)的中间神经元的连接结构来记忆词语之间具有前后时序的联结关系;而当我们回忆这段文章时,是在同步激励脉冲的控制下,以步进的方式依次激活联合皮质这些具有时序联结关系的中间神经元,并实时与感觉皮质的阅读中枢进行往返投射,于是感觉皮质逐个感知这些词语,产生成串的语言感知,实现对文章的忆起。(这种对原有已记忆的文章的忆起,也属于思维活动的一种形式,见“⒉⒋⒈因为信息记忆而建立的突触联系”)。
⒉⒍⒌在上述这种语言化的思维过程中,皮质上语言感觉区的神经元激活的同时,还可以向皮质上语言相关的运动区,包括说话中枢和书写中枢进行投射。但这两个中枢在平时是被关闭输出的,(受调制性神经元的抑制,或/和没有同步脉冲的协同激励),只有在需要说话或写字时,这两个运动中枢才被打开,(调制性神经元抑制解除,或/和同步脉冲的有效发放激励),于是,语言化思维活动的信息同时可进行运动输出,即进行说话或书写。
如果以跟⒉⒍⒉同样道理的方式进行分析,其结果是:说话中枢或书写中枢只是被动地接受思维活动的信息投射并下行向运动神经进行输出,而没有参与思维的过程。所以,发音不正,字词不会写,都不会影响思维活动的正常进行,甚至有时还会出现大脑“想的跟说出来的不一样”的情况。
⒉⒍⒍当然上面分析的是思维系统“有意识”的思维活动,而对于其他系统,比如运动系统,由于皮质运动区既可以接受联合皮质的输出投射,也可以直接接受皮质感觉区的投射,所以运动系统的运动动作既可以是受思维系统的输出控制,也可以是无需思维系统的“注意”和“意识”而直接进行的,其宏观表现便是,面对外部信息刺激,机体直接产生反应动作,然后才被思维系统所注意和感知。
⒉⒎思维、意识与注意之间的关系。思维是联合皮质的中间神经元有序的依次进行的链式激活,而意识是感觉皮质对思维活动的自我感知,所以,意识依赖于思维而产生。而根据大脑“注意”的工作机制,(见后面第3部分所述内容),“注意”是思维系统的控制环路接受各种感觉刺激,经过比较、竞争和互相抑制,整合出对多个信息处理通道进行协调控制的一种控制方式。“注意”的指向决定了在某一时刻,大脑的思维系统的哪部分神经元接受感知输入并进行思维活动,而这些神经元的思维活动又回馈投射到感觉区,被感觉区神经元所感知,产生了“意识”。所以,“注意”是一种控制方式,“思维”是一种反射活动,“意识”是一种感知过程。“注意”控制着“思维”,而“思维”产生了“意识”。
在一般情况下,这三者常是互相依存的,大脑所“注意”的地方,便是思维活动的内容,同时也会被大脑所“意识”到。尤其是当思维是以语言化的形式在进行时,思维活动回馈投射到进行语言识别的感觉区(应该是听觉性语言中枢),使我们强烈地感受到这种语言化的思考过程,强烈地感受到“意识”的存在。但在其他一些非语言化的思维活动中,比如前面所举的操作性工作的思维过程中,由于动作既可以是受思维系统有“注意”和有“意识”所控制,也可以是无需思维系统的“注意”和“意识”而直接进行的,甚至还可以是依赖于运动系统的程序性记忆而进行的,所以容易使我们弄不清“注意”、“思维”和“意识”之间的关联和区别。
⒊大脑思维活动的控制机制。大脑的信号处理和控制机制是大脑最重要的内容。之前,我们能够通过解剖学了解大脑的组成和结构,了解大脑各神经链路的大概的投射关系,甚至能够了解单个神经元及突触的结构及工作原理,但仍然无法明白大脑的记忆、思维和意识的本质及工作,其主要原因,便是对整个大脑(至少是对其中的思维系统)整体的工作机制、尤其是其信号处理和控制的机制,缺乏一个系统性的认识,这导致我们对大脑的认识如同盲人摸象般的局限性。而一旦弄清楚这种信号的传递和控制机制,很多关于大脑的问题,甚至一些精神病的本质和起因,便可迎刃而解。
⒊⒈关于思维的几个问题。前面分析了思维活动的最基本的神经元动作,即神经元的链式激活是脑思维的基本形式,也是思维的本质。神经元由于有序的依次激活,而形成链式激活,这种链式激活的神经元所对应的信息元的依次输出,形成信息链,便是思维。
可是,如果仅以神经元的链式激活来形成思维,那至少还存在几个问题:
⒊⒈⒈如果没有其他方面的协调作用,那么一个神经元的激活,可能无法单独导致下一个神经元激活,也可能导致多个后神经元同时激活,这时便不是出现有序的链式激活,而是出现辐射状的大范围的神经元激活,这显然不符合我们思维的串行且有序的特点。
⒊⒈⒉大脑在同一时刻的各种外部信息刺激(视觉、听觉等各种感觉)的输入,都可能引发大脑相关神经元的激活,如果没有某种控制机制,那么这些外部刺激都可能同时引发多条链式激活,形成多个思维同时进行,显然这也不符合我们思维在同一时间只是单一“思路”在进行的特点。(这里指的是有“意识”的思维,不包括身体对外部刺激无意识的反射反应,那是另外的反射系统,后面另有分析)。
⒊⒈⒊如果思维仅仅是通过中间神经元的链式激活来进行,那么,思维的速度应该是基本一致的,因为在“输入—触发动作电位—输出”这一过程中,神经元的动作速度基本一致,即使考虑兴奋的膜整合的时间,也相差不会很大。可是,实际上,大脑的思维速度和信息输出速度是可变的,甚至是可“有意识”的自由控制的。比如,在用语言进行思考时,或者用语言默读背诵一段话时,速度既可以很慢,慢至每秒1个字词以下,也可以很快,最快似乎可达到每秒10几个字词。显然,这不是链式激活的联合皮质的那部分神经元的活动决定的,而是需要另外一个其他的信号进行控制。
⒊⒈⒋如果思维仅仅是通过中间神经元的链式激活来进行,那么,思维一旦启动,便只能在有关联的相关神经元(也即相关信息)中进行,而无法将思维向其他方面的内容进行转向。但实际上,大脑在思维过程中是可以转向其他内容的。比如:思维过程中可以联想或切换到其他内容,将“思路”在不同信息内容之间进行切换,(当然,在同一时刻仍然只有单一思路在进行)。再比如,思维过程中当有其他感觉(视觉、听觉、触觉等)输入时,大脑既可以马上切换去“注意”并处理这些感觉输入,也可以忽视这些感觉而继续进行原有的思维。显然,这也需要有一个控制机制来对此进行判断和切换。
⒊⒈⒌神经元动作电位的脉冲宽度只有几毫秒,而思维活动和运动动作的时程大多在秒的数量级上,并可构成更长时程的活动周期,显然,难以依赖毫秒级的动作电位来直接形成秒级的信息活动。
⒊⒈⒍上述这些问题,其实质便是思维的控制机制。大脑在进行思维,也即在进行神经元链式激活的活动中,必然受到某种控制机制的控制,使得在某一时刻只能有某个或某组特定的神经元在激活,使得神经活动是以“步进”的方式有序进行的,保证思维活动是在某一通道以单一思路有序进行。而且,这种控制机制还控制思维能够在各种不同的思路、以及不同的外部感觉输入之间,进行合适的注意、判断和切换,即形成“注意”的控制机制。
申请人分析发现,大脑思维系统的这一控制机制,是由用于产生同步脉冲的“网状结构—丘脑”的思维振荡环路和几个调制通路共同构成的。(同步脉冲是按以往习惯对丘脑向皮质发放动作电位脉冲的称呼,但其实质并非用于同步控制,而是在“网状结构—丘脑”之间往返发放动作电位脉冲,形成振荡节律,并通过丘脑向皮质发放脉冲、以激励皮质的中间神经元依次进行链式激活,所以似乎称为激励脉冲更为合适,但出于习惯仍称为同步脉冲,或同步激励脉冲)。这一振荡环路及几个调制通路作用不同,共同构成思维的控制机制,而且也应该是运动及其他处理系统的控制机制。下面对此进行叙述。
⒊⒉“注意”的控制通路,同步脉冲振荡环路的工作机制。我们知道,丘脑在不停地向皮质发放动作电位,(这也是产生脑电波的原因),按目前的理解,这似乎是在控制皮质神经元的工作,而且,丘脑同步脉冲的发放似乎还跟网状结构有关联。那么,其具体的工作过程和控制机制是如何的?申请人经过分析,对此进行详细描述。
⒊⒉⒈首先,丘脑和脑干网状结构形成了一个往返循环发放动作电位的振荡环路。脑干网状结构内侧区,更准确说是内侧区的中脑网状结构向丘脑板内核投射发放动作电位,这种发放不包含具体的信息内容,但具有一定特异性。目前神经解剖学没有对中脑网状结构进行更详细的分区,但申请人认为中脑网状结构至少是具有分组结构的,其每一组神经元对应并连接着某一类信息输入,(比如视觉、听觉、触觉等输入,以及来自联合皮质思维活动的神经元的各条下行投射通路)。中脑网状结构的每一组神经元,上行投射向丘脑板内核中的一个特定分区,中脑网状结构的某一分组中的某一神经元,上行投射并控制着丘脑板内核中的对应分区中的一组神经元。中脑网状结构向丘脑板内核投射的神经元,属于胆碱能神经元,其突触传递反应速度快,引发的兴奋效应强大,使得中脑网状结构的某一组神经元激活发放时,所投射的丘脑板内核的该组神经元很多被兴奋激活而发放动作电位,但应该不会产生突触可塑性,所以没有记忆效应,其投射关系是固定的。
⒊⒉⒉丘脑板内核的神经元轴突同时向皮质和丘脑网状核进行投射;皮质也同时向板内核和网状核进行下行投射。(板内核向皮质的投射和皮质对板内核的投射在后另有叙述)。板内核向网状核的投射,和皮质向网状核的投射,两者存在投射位置相对应的关系,使得两者的脉冲发放能够进行兴奋整合,共同激励网状核神经元的激活。
⒊⒉⒊丘脑网状核的神经元,向中脑网状结构进行下行投射。这一投射不携带具体信息,主要是给中脑网状结构的神经元提供回馈激励,以维持振荡环路的往返发放能够持续进行。但可能具有分组定位的特异性,即板内核向网状核投射的某一组神经元,与网状核所投射向中脑网状结构的一组神经元,属于同一信息通路。
⒊⒉⒋这样,大脑工作时,中脑网状结构与丘脑之间的往返的脉冲发放过程是:中脑网状结构的某个或某组神经元激活,向丘脑板内核发放,使其所投射到的一组神经元激活;板内核的神经元向网状核发放,使被投射的神经元激活;(板内核同时向皮质发放同步脉冲);网状核的神经元向中脑网状结构的对应的神经元发放,再次激活中脑网状结构的神经元。于是,中脑网状结构与丘脑之间,按照“中脑网状结构→丘脑板内核→丘脑网状核→中脑网状结构”的投射关系,形成一个往返循环发放动作电位脉冲的“中脑网状结构←→丘脑”的闭环环路,如图12所示。这一环路,按其工作形式而言是一种往返发放的振荡环路,按其功能而言又是一种协调控制的控制环路,所以申请人将其称为“思维系统同步脉冲振荡环路”,简称“思维振荡环路”,或者“思维系统控制环路”,简称“思维控制环路”。
⒊⒉⒌丘脑板内核向网状核投射的同时,还向皮质进行上行投射,即皮质的“非特异性传入”投射。这种投射是发散性的,即少对多的投射,但实际是具有位置对应关系的,丘脑板内核的每一个分区,上行投射向皮质上的一个对应的大分区,(视觉感觉区、听觉感觉区、触觉感觉区等等,以及进行思维的多个联合皮质区等等);板内核某一分区中的一组神经元,上行投射向皮质该对应大区中的某一部分神经元;板内核的某一神经元,上行投射向皮质该部分中的一组神经元。丘脑板内核的特定神经元在受到中脑网状结构上行发放而激活时,向皮质所投射的神经元进行脉冲发放,形成了皮质的神经元思维活动的同步激励信号,使皮质神经元的活动有序地逐个依次激活,形成步进式的链式激活即思维,(同步脉冲起到类似于电子技术领域的中央处理器中时钟信号的时序控制作用)。同时,通过板内核不同神经元向皮质不同位置不同神经元的脉冲发放,也控制了思维活动在皮质不同神经元之间进行和切换,形成了思维活动中“注意”在同一类信息通道中的指向转移。(见“⒉思维和意识的本质”部分的叙述)。
同时,皮质也向板内核形成下行投射,并同时向网状核进行投射,这种下行投射是聚合性的,即多对少的收敛投射,跟上行投射相反,但在位置上其投射关系相同。皮质向板内核和网状核的下行投射发放,对板内核和网状核的神经元形成兴奋激励,虽然不是决定其激活的主要因素,但影响其兴奋整合的速度,从而影响了整个环路的发放节律。当皮质产生思维活动时,皮质向板内核和网状核进行回馈发放,通过兴奋整合加快了板内核和网状核神经元的激活发放,从而提高了振荡环路的振荡节律。
丘脑板内核与皮质之间往返投射的神经元属于氨基酸能神经元,其单一神经元动作电位脉冲的发放不足以使所投射的皮质神经元直接被兴奋激活而触发动作电位,只能产生协同的兴奋性激励,皮质的神经元需要有其他兴奋刺激的协同激励,(也即思维的链式激活通路之前激活的其他神经元的兴奋刺激,及其他调制性神经元的兴奋刺激),才能通过空间整合和时间整合来触发动作电位。但氨基酸能神经元能够在发放动作中产生突触可塑性,其投射关系(也即联结结构)可随着神经元活动而产生修饰,所以,丘脑板内核与皮质之间的神经元,可能不只是同步脉冲发放的作用,而是参与了信息的记忆和整合。
⒊⒉⒍丘脑板内核向皮质的投射和发放,和皮质向板内核的投射发放,虽然也存在往返的脉冲发放,但其实质并不构成振荡环路,而只是“中脑网状结构←→丘脑”的思维振荡环路在板内核节点上的对外延伸。如果切断丘脑跟皮质的投射联系,“中脑网状结构←→丘脑”的振荡环路仍然能够维持振荡,反过来,如果切断“中脑网状结构←→丘脑”之间的投射联系,则丘脑跟皮质之间必然无法形成往返发放。其中,板内核向皮质的脉冲发放,形成了皮质神经元思维活动的同步激励信号,而皮质向板内核和网状核的回馈发放,也对思维振荡环路的工作形成回馈,并可能对其振荡节律形成影响。
⒊⒉⒎中脑网状结构的神经元工作,决定着思维振荡环路的振荡节律,也控制着中脑网状结构的哪一组神经元向丘脑板内核进行脉冲发放。由于缺乏更详细的解剖资料,申请人目前无法确定中脑网状结构中,向丘脑板内核上行投射的神经元,和接收网状核下行投射的神经元,是同一组神经元或者是前后两组神经元?申请人偏向于是具有前后两组神经元。但无论是同一组神经元还是前后两组神经元,中脑网状结构这部分神经元,需要同时接收来自几方面的兴奋及调制信号,包括:⑴、网状核下行投射的兴奋激励,用于维持整合振荡环路的振荡继续进行;⑵、来自其他调制性神经核团及来自其他控制环路的调制作用,用于调制神经元的兴奋整合的速度,从而调制整个振荡环路的振荡节律;⑶、来自各个输入信息通路及皮质下行投射通路的兴奋信号,用于根据各个信息通路的兴奋程度(可能还有时序),来控制某一信息通路的某组神经元首先激活,向丘脑板内核发放;⑷、来自中脑网状结构不同信息通道的神经元的抑制性调制,用于使中脑网状结构某组神经元首先激活向丘脑板内核发放脉冲时,同时抑制网状结构的其他信息通道的神经元在该脉冲周期内不再会激活发放。这几方面的兴奋信号和调制信号进行整合比较和协同作用的结果,使得思维振荡环路在同一时刻,中脑网状结构只能有某一信息通道中的一组神经元向丘脑板内核进行同步脉冲发放,从而使得在同一时刻,丘脑板内核只能有一组神经元激活并向皮质的某一分区中的某一部分神经元进行同步脉冲发放,激励其产生思维活动,并同时回馈投射到感觉皮质,对思维进行自我感知,产生“意识”。所以,大脑通过“中脑网状结构—丘脑—皮质”的分级控制,形成了在同一时刻只对皮质思维系统的某一部分神经元发放同步脉冲,以激励这一部分的神经元进行思维活动,以此形成思维系统的单一性的“注意”指向。这也是思维系统“注意”指向的形成机制。
⒊⒊信息的输入传递通道。而要分析大脑如何对各种信息进行“注意”指向的控制及切换,还需要了解各种信息输入通道的传递通路。大脑的信息处理包括对外部感觉信息和对机体内部信息的感知处理,跟思维和“注意”有关的主要是外部信息处理。而对外部信息的感知处理又包括二方面:一是外界的各种感觉信息,包括视觉、听觉、嗅觉等等,来自于各种感觉器官的输入,不同感觉具有不同的输入传递通道;二是大脑在思维过程产生的中间信息,来自于皮质的中间神经元在思维活动中产生的下行投射,不同区域的皮质有各自的下行投射通路。对于各种感觉的输入传递通道,之前的研究比较多,这里只以其中比较复杂的视觉和听觉输入通道为例,分析其与思维和“注意”的关联关系。
⒊⒊⒈视觉信息的输入传递通道。根据目前的解剖学研究,视网膜的多种视觉感觉神经元的输出即视神经,在视交叉之后分为两大传递通路。(参考图13的人脑视觉通道信号投射结构示意图)。
第一条视觉传递通路的神经元很多,形成较大的视束,传递的是包含具体内容的视觉信息,通过丘脑外侧膝状体的中转再投射到初级视皮质,属于皮质的“特异性传入”投射,视觉信息由时间编码和频率编码转换为空间位置编码,完成对各种视觉信息(形状、大小、位置、颜色、亮度等)的初步识别,(参考上面“⒉⒉认知的工作机制”),再以多对一的方式投射到联合视皮质,完成对视觉对象的识别。之后,对于直接的视觉信息(具体的物体、图像、画面等),直接投射到联合皮质进行记忆、思维和反应活动,而对于涉及文字的视觉信息,则投射到“阅读中枢”的皮质,完成对文字的识别,再投射到联合皮质进行语言化的信息记忆和思维活动。联合视皮质还可以直接投射到运动皮质。运动皮质与小脑(似乎还有纹状体)等构成运动系统,运动系统不属于思维系统,既可以受思维系统的输出控制,也可以无需思维系统控制而直接根据视觉信息来反射和控制肌肉运动,也能够进行联合的学习记忆和反应处理,(这里的记忆也即是所谓的“程序性记忆”),完成我们日常生活中无意识进行的大多数动作。
视交叉之后的第二条视觉传递通路的神经元较少,但实际却是更重要的,这一传递通路实际上至少包含三方面:⑴、投射向下丘脑。该投射传递的只是视觉信息中的亮与暗的信息,由下丘脑的视上核、结节核、乳头核等形成反射回路,再上行向丘脑和皮质的广泛区域、下行向脑干和脊髓等进行调制性的投射,(属于组胺能神经元),用以形成日夜节律并进行睡眠和觉醒的控制。⑵、投射向顶盖前区,再投射向中脑网状结构。这一投射没有包含具体的视觉信息,也即非特异性投射,只是传递视觉信息的有无和强度,(应该是通过动作电位的发放频率来实现信息强度的传递),所以申请人把这一神经投射称为“报告”式投射。这一“报告”信号投射到中脑网状结构的视觉上行通道,并与同样投射到网状结构的其他不同类感觉(听觉、嗅觉等)的“报告”信号进行“竞争”,如果能够激活网状结构向丘脑投射的视觉上行通道,便是“引起”网状结构的“注意”。顶盖前区同时有神经元下行投射向眼睛,用以反馈调节晶状体,通过瞳孔反射来调节进入眼睛的光强度。⑶、投射向上丘。上丘跟初级视皮质具有神经元的相互联系调制,同时上丘还有神经元下行投射向眼睛的动眼神经,进行眼动控制。所以,申请人推测上丘的作用是当视觉信息受到“注意”时,控制将视觉的具体“注意点”放在视觉信息的哪一部分,也即视野中的哪个位置,而上丘对眼动的控制,正是将眼睛晶状体的焦点投射在视网膜的哪个位置上。
显然,第⑵和⑶两方面便跟思维的“注意”有关,其中投射向顶盖前区的第⑵方面是引起网状结构对视觉信息的“注意”,而投射向上丘的第⑶方面是将视觉的“注意点”投向视野的哪个位置。值得一提的是:上丘向动眼神经的投射不需要经过皮质运动区和小脑,而直接控制着眼睛的眼动动作。在做梦时,思维系统出现神经元活动,但运动皮质和小脑等运动神经系统的输出受到抑制,不会产生机体动作,只有动眼神经既可受运动系统的控制,又可直接受网状结构和上丘的控制,所以能够跟随思维系统的活动(梦境)出现眼动动作,这也即是做梦时出现眼动动作的原因。(后面另有对做梦进行叙述)。
⒊⒊⒉听觉的信息输入通道跟这类似,如图14所示:第一条听觉传递通路(外侧丘系传导通路)从耳蜗通过内侧膝状体中继,向初级听皮质投射,用于传递包含具体内容的听觉信息;第二条听觉传递通路(丘系外通路)从上橄榄核分支,投射向脑干网状结构,该传递通路并不包含具体的听觉内容,只是传递听觉的有无和强度,也属于“报告”式投射。这一“报告”信号在网状结构跟其他输入的“报告”信号进行“竞争”,以引起网状结构对听觉信息的“注意”。两条听觉传递通路在下丘进行相互联系,下丘同时与初级听皮质有神经元进行相互联络,所以,申请人推测下丘的作用是当听觉信息受到“注意”时,控制将听觉的主要“注意点”放在听觉信息的哪一部分,也即哪个位置和哪个频率段的声音。其他信息(触觉、嗅觉及躯体感觉等)的输入通道跟视觉通道和听觉通道类似,这里不再描述。
⒊⒊⒊而对于大脑在进行思维时,联合皮质的中间神经元活动所产生的中间信息的投射处理,目前没有这方面的相关研究。申请人研究分析,这种中间信息除了向其他中间神经元投射以继续思维活动,还存在两方面的投射:一方面向皮质的联合感觉区进行回馈投射,使思维活动中产生的中间信息被重新感知,(在一些情况下还被进行记忆和整合),形成了大脑对思维活动的自我感知也即形成“意识”,(见第2部分,“思维和意识的本质”部分的内容);另一方面进行下行投射,(氨基酸能神经),这种下行投射是多对少的聚合性投射,即皮质上多个中间神经元的活动共同激励一个下行神经元,而且应该是具有分组特异性的,即皮质的不同分组具有各自的投射通路,(这种分组不局限于解剖学上定义的皮质分区,而是神经元长期活动可塑性形成的,所以称为分组更合适)。聚合性投射的结果,是这种投射信号没有包含神经元活动的具体信息,而只是传递对应分组的神经元是否在活动,以及活动的活跃程度,(通过动作电位发放频率的快慢来传递)。这些皮质下行的各个投射通路,在经过中继后,(中继是进行信号整合及再次聚合),进入到脑干网状结构,成为“报告”式投射,跟其他报告信号进行“竞争”,以引起网状结构对该分组的皮质神经元活动的“注意”。思维系统的这种皮质向网状结构的下行投射通路的意义是,当该分组的皮质在进行思维活动时,这种“注意”能够维持中脑网状结构和丘脑继续对该分组的皮质进行同步脉冲发放,以继续维持该分组的思维活动的进行。
⒊⒋“注意”的控制、维持和切换。根据申请人的研究和分析,思维活动中“注意”指向的控制和切换,并非以往所认为的是在信息处理顶层的端脑皮质产生,再去控制大脑的其他部分,而是在中脑网状结构上产生和切换,再上行控制丘脑和皮质等其他区域的神经元活动。思维系统“注意”指向的切换在两个层面上进行:中脑网状结构负责的是将“注意”指向切换到哪一个信息通道,即“注意”哪一类信息;丘脑负责的是将“注意”指向该信息通道中的哪个具体位置,即“注意”该类信息的哪一部分;而皮质尤其是端脑联合皮质只是在“注意”的控制下进行具体的信息处理。
⒊⒋⒈网状结构对“注意”指向的控制。如前述,各类信息通道,包括外界输入信息通道(视觉、听觉、嗅觉等通道)和皮质思维通道的神经元活动,都有经过聚合性(多对少)整合后的神经输出向中脑网状结构进行投射,这些投射通路只传递信息刺激的有无和强度(通过动作电位发放频率来传递强度),而不包含具体的信息内容,属于“报告”式投射。中脑网状结构具有多个并列的向丘脑进行投射的上行激励通路,对应接收这些来自各个信息通道的“报告”式投射,并且这些并列的上行激励通路彼此之间交互投射进行互相抑制,出现类似“竞争”的现象:当某一时刻有某组神经元受到兴奋激活并上行向丘脑进行脉冲发放时,其输出同时对其他通路的神经元产生抑制,使得其他通路的神经元在同一脉冲周期内无法再激活输出,形成了中脑网状结构层面上的单一性的“注意”指向。
至于在一个脉冲周期内,哪组神经元能够得到激活并上行发放同步脉冲,则取决于几个信号的共同整合:⑴、来自各信息输入通路的非特异性的“报告”式投射的信号,这一信号是主要的,其信号强弱起决定性作用;⑵、来自丘脑网状核下行投射的同步脉冲发放,用于维持振荡环路的振荡继续进行,也因此形成了每一个脉冲周期;⑶、来自脑干和下丘脑等其他神经活动的调制作用,用于调制整个振荡环路的振荡节律;⑷、来自网状结构其他神经元的抑制性调制,用于进行“竞争”;这些信号共同进行兴奋整合,造成在振荡环路的一个脉冲周期内,总有某组神经元的兴奋整合首先触发动作电位而激活,相当于其“报告”竞争成功得到应答而引起“注意”,于是其激活输出一方面向板内核上行发放同步脉冲,另一方面立即对其他通路的神经元产生抑制,使它们无法再激活输出。之后,在丘脑网状核下行发放的下一个脉冲的脉冲周期,上述整合和竞争过程又再次进行。周而复始,使得在每一个脉冲周期内,中脑网状结构有且只有一个信息通路的“报告”竞争成功而引起“注意”,并上行向丘脑板内核进行激励脉冲发放,将信息处理所需要的同步脉冲投射向这一信息通路,实现了对思维活动的“注意”指向“哪一类”信息的控制。
⒊⒋⒉丘脑对“注意”指向的控制。在人,丘脑的主要功能是中继,包括信息中继和同步脉冲中继。负责信息中继的是“特异性中继核团”,主要包括外侧核群、腹侧核群和膝状体核群,用于对多种感觉(视觉、听觉等)的信息输入进行中继和初级处理;还有腹前核和腹外侧核,用于传递运动系统的中间信息。这些特异性中继核团向皮质的投射属于“特异性传入”,而负责同步脉冲中继的是所谓的“非特异性核团”,实际上同步脉冲的发放是具有特定投射关系的,所以也具有特异性,只不过是同步脉冲发放的特异性而非信息内容的特异性。同步脉冲的中继核团接收网状结构上行投射的同步脉冲发放,(胆碱能神经投射),其输出一方面发侧枝投射到网状核,再下行投射回到脑干网状结构形成闭环振荡环路;另一方面这些中继核团的输出也向大脑皮质和基底核、边缘系统和小脑等多个区域投射,(即所谓的非特异性传入),进行同步激励脉冲的发放,激励和协调控制这些区域的神经元进行思维和运动等信息处理。
根据现有的解剖资料进行分析,在丘脑,跟同步脉冲控制相关的中继核团主要包括丘脑前核群、部分内侧核群和板内核群。其中前核群和内侧核群接受脑干脚桥被盖网状核和外侧背核的同步脉冲的发放,其输出向部分大脑皮质和边缘系统(尤其是海马和杏仁核)皮质进行投射,主要用于控制思维系统和情绪系统的中间信息的记忆和整合;而板内核,接受中脑网状结构的同步脉冲发放,其输出投射到端脑基底核、纹状体、和端脑皮质的广泛区域,用于协同控制思维系统的信息处理也即思维活动,还有能够受思维系统控制的那部分运动皮质的运动处理。其中主要以丘脑板内核参与了思维系统的“注意”控制通路:当中脑网状结构的某一信息上行投射通路的神经元激活而向板内核对应位置的神经元发放同步脉冲时,板内核的这部分神经元被上行的同步脉冲发放所激活,其轴突输出以发散投射的方式上行向皮质对应神经元发放同步脉冲,使被投射发放的该部分神经元能够得到同步脉冲的协同激励,形成兴奋整合而进行链式激活,即进行思维活动。(板内核向皮质的相互发放参见⒊⒉⒌部分的描述)。而丘脑网状核接收板内核的侧枝投射和皮质思维通道中间神经元的回馈投射,再下行向中脑网状结构进行回馈投射。
所以,中脑网状结构、丘脑板内核和丘脑网状核构成了思维系统的“注意”控制环路,接收各个信息通道的“报告”式投射的信号,再对思维系统的哪一信息通道的哪部分神经元进行同步激励脉冲发放,以进行思维“注意”指向的控制。其具体的神经元投射如图15所示,各通道的信号投射如图16所示。其中中脑网状结构的上行发放决定着丘脑板内核的哪组神经元能够激活输出,而丘脑板内核这组神经元的激活输出又决定着皮质的哪部分神经元能够进行信息处理。中脑网状结构负责的是将“注意”指向切换到哪一个信息通道,即“注意”哪一类信息;而丘脑负责的是将“注意”指向该信息通道中的哪部分神经元,即“注意”该类信息的哪一部分;而大脑联合皮质是负责具体的信息处理(思维和记忆),并通过感觉皮质自我感知这一处理过程,也即产生自我“意识”。比如,中脑网状结构控制将“注意”指向视觉信息,丘脑控制在视觉画面中“注意”哪一个物体,而联合皮质则是负责识别这个物体,并同时通过感觉皮质感知联合皮质的活动,“意识”到这个物体的存在。
⒊⒋⒊“注意”的维持。当一个思维过程在进行时,该思维通道的皮质神经元活动处于兴奋状态,这些神经元的激活动作,除了继续向前发放以进行链式激活之外,还同时通过聚合式的投射,产生两路下行的投射信号:第一路下行信号回馈发放到丘脑板内核,通过兴奋整合使板内核将下一个同步脉冲继续向该通道进行发放,以维持该通道的神经元能够继续进行链式激活,第一路下行信号还同时发侧枝投射到丘脑网状核,促使振荡环路能够继续进行,(尤其是高节律振荡时);第二路下行信号是向中脑网状结构进行“报告”式的投射,通过竞争维持中脑网状结构对该通道的继续“注意”。这两路下行投射信号共同维持思维控制环路对该通道的“注意”指向,这也即是思维活动的“注意”维持的工作机制。
⒊⒋⒋“注意”的切换。导致“注意”指向出现切换的,大概有以下这几种情况。
⑴、思维活动的内容发生变化。在思维过程中,原有思路的信息内容联想或转换到其他方面的信息,也即原有思维通路的神经元活动由于整合而反射到其他方面的神经元,引起该部分的神经元兴奋激活,而这部分神经元的激活,按照同样的机制产生两路下行投射信号,一方面回馈投射到丘脑板内核,使得板内核将下一个同步脉冲继续向该部分神经元进行发放,维持该部分神经元能够继续活动;另一方面下行向中脑网状结构进行“报告”投射,如果该部分神经元跟原有思路不属于同一思维通道,则吸引了中脑网状结构对新通道的“注意”。至此,使思维的“注意”指向,在不同思路之间进行切换。
⑵、外部输入信息的竞争。在思维过程中,如果有外部信息的输入,(看到什么,听到什么,感觉到什么,等等),其输入通道会通过非特异性传递向中脑网状结构进行“报告”,至于能否引起“注意”,则要看其信号强度。如果输入信息强烈,(比如看到意外事情,听到刺耳声音,受到刺疼,等等),这会反映在其“报告”信号的动作电位发放频率上,所以能够使该信息通道神经元的兴奋整合更快并触发激活,向丘脑上行发放同步脉冲,并同时对之前的“注意”通道进行抑制,也即“竞争”成功并获得中脑网状结构对该信息输入的“注意”。至此,将思维的“注意”指向,切换到对该外部输入信息的处理上。
⑶、其他调制通路的影响。在思维过程中,其他调制通路的调制信号的影响,比如运动神经系统的状态,情绪系统的情绪影响,内脏和内分泌系统的突发变化,酒精、药物、毒品及其他化学物质的作用,等等,都可能对各个信息通道的神经元活动造成影响,从而影响了中脑网状结构对各种信息通道的“注意”控制,造成“注意”指向的切换。
⑷、“注意”控制通路的神经元工作异常。特别是中脑网状结构的各个信息上行投射通路之间进行互相抑制的作用出现异常。在这种情况下,当某一信息通路成功引起“注意”而在进行信息处理的过程中,如果这时无法维持对其他信息通路进行有效的抑制,便可能导致其他信息通路出现意外的激活而引起“注意”,这时中脑网状结构无法将“注意”持续维持在某一信息通路上,而是在不同信息通道中进行非正常的切换。这在宏观上,表现为注意集中困难和注意持续时间短暂,大脑无法专注于某件事情,成为一种精神异常,即“注意”缺陷。(见后面的精神疾病部分,关于“注意”缺陷(ADD)的描述)。
⒊⒋⒌综述之,思维控制环路对思维系统的“注意”指向进行控制、维持和切换。其中中脑网状结构负责的是将信息处理的“注意”指向“哪一类”信息:通过“中脑网状结构←→丘脑”之间上下往返发放的同步脉冲、跟各种外部信息和中间信息的输入信号强度(动作电位发放频率)进行比较整合,来使网状结构的上行激励脉冲向丘脑板内核发放的分区和分组位置发生变化,以选择转向“哪一类”信息通道,(比如,指向视觉、听觉、触觉等,或指向皮质正在进行思维的中间信息通道)。而丘脑负责将信息处理的“注意”指向同一类信息通道中的“哪一部分”:当丘脑某一分区的某组神经元受到网状结构上行发放的脉冲激励时,其神经元激活并向皮质上所投射的那部分神经元发放同步激励脉冲,通过兴奋整合使皮质上该部分神经元进行链式激活即思维活动,从而控制思维活动的“注意”指向这部分神经元所对应的信息上,(比如,在视觉信息中“注意”哪一个人,在众多声音中“注意”哪一个声音,在思维活动中“注意”哪一方面的内容)。而皮质负责具体的信息处理:联合皮质上对应着各个信息元的中间神经元,在丘脑同步脉冲的发放激励下,进行着有序的依次逐个或逐组的步进激活,形成链式激活,实现大脑对信息的识别、思维、反应和记忆功能,(比如,那个人是谁,那个声音说什么,那个问题是什么,等等);并同时投射到感觉皮质,通过感觉皮质感知联合皮质的活动,形成对思维活动的自我感知也即形成“意识”,(比如,意识到那个人的存在,意识到那个声音,意识到“我”在进行什么思维,等等)。
⒊⒋⒍“注意”的切换速度与脑电“N400”电位。对于正常的思维活动,由于是在联合皮质的相关神经元之间进行链式激活,依赖的是丘脑向皮质发放的同步脉冲的激励,所以思维能够以较快的速度进行,在具有增强性调制的情况下(处于兴奋或紧张思维),思维的链式激活的步进速度可以达到每秒十几个步进以上,也即每个步进耗时不到一百毫秒。但是,对于“注意”指向的切换,不管是在来自外界的无关联性的不同感觉信息之间、或者是在来自思维过程的无关联性的不同思维内容之间、或者是在内外不同信息之间进行切换,某一信息要得到新的“注意”,需要该信息通道的神经元先激活发放,再经过聚合投射到中脑网状结构进行“报告”,再在中脑网状结构进行兴奋整合并跟之前的“注意”通道竞争,竞争成功后中脑网状结构该通道的上行神经再向丘脑板内核进行投射发放,板内核再向皮质上跟该信息相关的神经元进行同步脉冲发放,这时才能产生跟该信息相关的感知或思维,引起了“注意”。所以,“注意”切换的速度是比较慢的,其耗时估计在几百毫秒左右。
申请人推测,大脑在没有关联性的不同信息之间进行“注意”指向切换时引起的控制环路神经活动的变化和迟延,正是脑电研究中事件诱发电位“N400”的产生原因:在默读一段句子且句末是语义没有关联性的词语时,或者在呈现两幅完全不相关的图片时,大脑在无法联想到的也即没有关联性的两种不同信息之间,进行“注意”的切换,引发了思维控制环路的同步脉冲发放的迟延和变化,产生了脑电诱发电位的“N400”现象。由于思维系统的“注意”指向发生在信息的输入感知、识别和中间处理环节,而与运动输出无关,所以Wernicke失语也即感觉性失语患者的“N400”不会产生,而Broca失语也即运动性失语患者的“N400”能够产生。
⒊⒌思维振荡环路的节律变化及对大脑工作的影响。
⒊⒌⒈思维振荡环路的发放节律的变化。思维振荡环路通过“中脑网状结构→丘脑板内核→丘脑网状核→中脑网状结构”的循环发放来形成可持续的同步脉冲,这一振荡环路的发放节律,或者说同步脉冲的发放频率,受到多个信号的影响和调制,以此形成振荡环路发放节律的被动改变和主动调整。这些调制信号包括:⑴、来自皮质神经元的下行投射。当丘脑板内核向皮质发放同步脉冲时,如果皮质的神经元在进行思维活动,则皮质会向丘脑板内核和网状核进行回馈发放,加快环路的发放节律。⑵、来自其他脑核团的神经元活动的交互调制。比如,来自“情绪”处理系统(大脑中的另一个信息处理系统)的调制,当情绪激动时,通过对振荡环路神经元活动的影响,会加快环路的发放节律。⑶、来自“脑干网状结构(正中区)←→下丘脑”的“下环控制环路”的交互调制。“下环控制环路”负责控制和调节人体的内分泌和内脏器官的工作,其工作状态形成了人体的身体状况,下环控制环路的工作状态能够对思维控制环路进行调制,所以,身体状况的变化也会对思维路的振荡节律形成影响。⑷、神经调质、药物的影响。比如,酒精、药物等能够影响环路神经元的兴奋传递和整合,从而影响环路的振荡节律。
⒊⒌⒉思维振荡环路振荡节律与大脑工作状态的关系。思维振荡环路的振荡节律,也即丘脑向皮质发放同步脉冲的节律,决定着大脑思维系统的工作状态。
⑴、当同步脉冲发放节律大于14赫兹时,皮质神经元兴奋整合和激活很快,神经活动活跃,大脑处于紧张激动或快速思维的状态。如果还同时受到其他调制通路的调制信号的协同作用,还可以使氨基酸能神经元产生记忆效应。(在海马结构的皮质,高节律的同步脉冲激励可使氨基酸能神经元爆发高阈值的V1.2亚型的动作电位,使神经元产生长时程突触可塑性,形成长时程记忆。见申请人2014106066977号专利申请的说明书中关于神经元不同亚型动作电位的爆发机制方面的描述)。
⑵、当同步脉冲发放节律为8—13赫兹时,这时神经元的膜整合和活动速度中等,大脑处于清醒、安静、或闭眼的状态,没有具体的“注意”点,没有进行紧张思维活动,可能产生思维片段,但对思维过程的内容大多没有进行记忆。
⑶、当同步脉冲发放节律为4—7赫兹时,虽然同步脉冲的兴奋激励尚能够使皮质的部分神经元进行兴奋整合并激活,但兴奋整合缓慢,神经元活动不活跃,无法形成连贯的激活链,所以这时大脑虽然还存在“意识”,但对各种刺激反应迟钝,思维模糊,且对思维过程的内容无法记忆。在大脑临睡觉前,或者非常困倦,或者身体健康状况很差时,属于这种情况。
⑷、当同步脉冲发放节律为0.5—3赫兹时,这时虽然丘脑仍然向皮质发放同步脉冲,但由于脉冲发放节律很低,脉冲之间的间隔周期太长,(几百毫秒到上千毫秒),使得皮质神经元只能产生膜兴奋,但兴奋的膜整合始终无法达到动作电位的触发阈值,所以皮质的神经元没有出现持续激活和传递的情况,所以这时大脑失去“注意”和“思维”活动,也就没有了“意识”。这种情况出现在大脑深度睡眠(即慢波睡眠)、或昏迷、或麻醉的状态下。
⑸、同步脉冲发放节律为0,也即大脑的思维振荡环路的动作电位发放完全停止,这时中脑、丘脑及皮质的神经活动停止,对刺激没有反应,大脑失去高级功能,属于目前医学上定义的脑死亡。值得注意的是,这种情况下虽然脑电图为等电位直线,即没有接收到脑电波,但实质上只是思维振荡环路停止了同步脉冲发放,如果脑死亡主要是由于脑缺血缺氧而导致,而且脑死亡时间不是很长,这种情况下大脑的大部分神经元仍然没有丧失细胞活性,所以,这时生物体仍然没有彻底失去生物活性,如果能够恢复对脑的正常供血供氧,同时对振荡环路的关键环节的神经元施加合适的电脉冲刺激,仍然还有可能重新激励神经元的动作电位发放,恢复振荡环路的往返发放,重新激活大脑的工作。
需要说明的是,丘脑向皮质发放同步脉冲的节律,虽然决定着皮质思维活动的速度,但却不等于思维的神经元链式激活的步进速度,这是因为神经元的动作电位触发可能需要同时几个同步脉冲的激励才能完成兴奋整合(时间总和的整合),而神经元一旦触发发放,由于突触易化的原因,则容易产生连续多个动作电位发放,所以两者并非一一对应的。
⒊⒍多种调制通路对思维的影响。思维是皮质神经元的活动,这一过程受到相关神经元的连接结构的限定,(这些连接结构也即信息记忆的结构,包括知识、事件、经验等,这是形成记忆的基础),同时受到思维系统“注意”控制环路的同步激励脉冲的实时控制,形成单一通路的步进式的链式激活,而且,这种活动还受到其他一些调制通路的调制作用。这些调制通路所产生的调制作用,不像同步脉冲那样进行实时控制,而是缓慢和持续的,通过影响链式激活通路的神经元的递质的释放、吸收和离子通道的活动,来影响膜整合和动作电位发放,从而对思维活动产生影响。
⒊⒍⒈情绪系统对思维的影响。大脑情绪系统接收多种输入信息,包括视觉、嗅觉、听觉、触觉等外显的感觉信息,也包括思维系统的中间信息,还包括来自机体内部的非外显的感觉信息,产生各种感觉,并通过各种单胺能神经核团输出各种调制信号,作用于思维系统,对思维(思路)进行影响。思维系统的中间神经元在进行链式激活时,即使接受同样的来自前面的兴奋信号、和同样的来自丘脑的同步脉冲信号,但由于后级神经元的所受的调制作用不同,造成膜整合和动作电位触发条件不同,导致会触发不同的后级神经元,使链式激活向着不同的方向进行,(比如:a→b→c或a→b→d),从而产生出不同的思路。所以,面对同样的事情,不同的情绪下大脑会产生不同的思维结果。(情绪系统除了对思维形成影响,还广泛地对运动神经系统和内分泌、内脏神经系统形成影响,这将在后面另行叙述)。
⒊⒍⒉睡眠与觉醒的调制作用。控制大脑进行睡眠和觉醒的调制信号,主要来自于下丘脑的组胺神经核团。来自视网膜的传递光照信息的视觉传入投射到下丘脑的视上核,与结节核、乳头核等形成反射回路,产生日夜节律变化的信号,再与网状结构振荡节律的信号互相调制,输出控制睡眠和觉醒的调制信号。这一调制信号上行向丘脑、小脑、边缘系统和皮质(包括感觉皮质、联合皮质、运动皮质等),下行向脑干和脊髓进行广泛的投射,对神经元的活动进行调制,用以进行睡眠和觉醒的控制。其作用大多为抑制性调制,以降低神经元的活动程度。比如,通过投射向感觉皮质的通路来抑制外界和机体的各种感觉信息的输入,通过投射向丘脑和皮质的通路来抑制思维活动,通过投射向小脑和脊髓的通路来抑制各种运动输出,通过投射向下丘脑和脑干的通路来调节机体的内脏和内分泌的活动,等等。当这些调制通路能够正常地一起产生或停止调制作用时,大脑能够正常地在睡眠或觉醒状态进行转换,而当这些调制通路的调制作用不协调不一致时,则会导致大脑出现各种异常而有趣的表现,比如,做梦,梦游,失眠。(在后面另外对这些现象的产生机制进行叙述)。
⒊⒍⒊大脑生化环境对思维的影响。大脑里面的多种生化物质的存在和浓度构成了大脑的生化环境。这些生化物质包括两大类:一类生化物质是机体自身所分泌的,用于控制调节机体的各种器官和系统的工作,即内分泌系统,(实际也包括各种神经递质和神经调质)。另一类生化物质不是机体自身产生的,但机体感知这些生化物质的存在和含量,通过下丘脑一带的神经核团的反应处理,去控制调节机体的各个器官和系统的工作。比如血液中的氧、二氧化碳、血糖、酒精、各种药物和毒品,等等。如果把这两类生化物质的反应处理合并起来,也可统称为大脑的生化反应系统。
大脑生化环境对思维的调制和影响,主要通过两大途径:一是通过对网状结构神经元活动的影响,影响了思维振荡环路的往返振荡节律,从而影响了思维系统的神经元活动状态。(详见“⒊⒌⒉思维振荡环路振荡节律与大脑工作状态的关系”)。二是通过下丘脑、中脑、间脑上的各个单胺能神经元核团,向丘脑、皮质(包括海马皮质,实际还包括运动系统的小脑皮质)投射各种调制性神经元,来直接调制影响思维系统的神经元活动。大脑生化环境对思维的影响,由于生化物质的多样性和复杂性,其影响往往不是单一途径孤立的发生作用,而是多种途径综合作用,既影响了思维振荡环路的工作状态,又通过调制神经元对相关区域进行调制,协同产生作用。
⒊⒍⒋内脏神经系统对思维的影响。大脑的内脏神经系统,通过内脏感觉神经来对各个内脏的工作状态进行感知,通过脑干、下丘脑等的一些神经核团进行反射处理,再通过交感神经和副交感神经去控制调节内脏的工作。内脏神经系统的工作,通过两条途径对思维活动进行影响:一个也是在脑干网状结构的环节,对思维系统的思维振荡环路进行影响,从而影响了思维系统的思维活动。二是通过影响了大脑的生化环境,再通过生化环境对思维系统的思维进行影响。(见上面⒊⒍⒊的内容)。
⒊⒎思维系统的“海马”中间信息处理环路。“中脑网状结构—丘脑板内核和网状核—皮质”是丘脑最主要的涉及最广的控制环路,进行思维系统的“注意”指向及工作状态的控制。丘脑还存在另外一个非常重要的控制环路,即是“脑干网状结构—丘脑前核和内侧背核—丘脑网状核—脑干网状结构”的同步脉冲控制环路,并与皮质的扣带回和海马相关结构一起构成思维系统的“海马”中间信息处理环路。这一处理环路既相对独立,但也可以说是属于思维控制环路的一个分支,起到细化和补充作用,其不直接涉及对各种外部感觉信息的识别和认知,而只涉及联合感觉皮质之后的思维系统的中间信息的中间处理和整合,尤其是涉及中间信息的短期记忆(短时程和长时程记忆)。
⒊⒎⒈这一环路的神经投射结构如图17。其中脑干网状结构(似乎是中脑被盖的核团)向丘脑前核(和内侧背核)进行投射(胆碱能神经元),丘脑前核向丘脑网状核投射,网状核向脑干网状结构进行下行回馈投射,以此构成“脑干网状结构—丘脑前核—丘脑网状核—脑干网状结构”的循环振荡环路,形成激励脉冲的往返发放。这一振荡环路在丘脑前核(和内侧背核)向皮质扣带回进行同步激励脉冲发放(氨基酸能神经),扣带回有向丘脑前核和丘脑网状核的下行投射,其同步激励脉冲的发放和控制机制跟中脑网状结构—丘脑板内核等构成的“注意”控制环路是类似的。在信息传递处理方面,大脑其他皮质区域尤其是联合皮质与扣带回具有广泛的双向投射,而扣带回跟海马结构的内嗅区具有双向投射,以此实现信息的双向传递处理。在海马结构内部,输入海马结构的信息通过“内嗅区—齿状回—CA3—CA1—下托皮质—内嗅区”的环路进行整合处理,重新投射到内嗅区再输出到扣带回,以此实现对皮质的信息的中间整合处理。海马的下托具有直接向脑干网状结构的下行投射,和通过下丘脑乳头核间接的下行投射,这些起到类似于“报告式”投射的作用。在思维系统的“注意”控制环路将“注意”指向切换到“海马中间信息处理通道”时,也即思维系统的神经活动集中在海马结构相关的中间信息整合处理时,海马这一处理通道用于对思维过程的细节性的“注意”控制和对中间信息即陈述性信息的整合处理。
⒊⒎⒉尤其重要的是,海马结构的信息处理还同时受到隔区的内侧隔核和斜角带核的神经投射的控制,而内侧隔核和斜角带核的神经投射属于胆碱能神经,其兴奋传递具有快速和强烈的特性,能够使被控制的神经元形成强烈的膜兴奋,并爆发V1.2亚型的高阈值动作电位,从而产生神经元的突触可塑性,(STDP可塑性),形成记忆效应。(这一点,详见后面的“⒋⒍不同神经递质的神经元在大脑信息处理的不同作用”部分的内容)。所以,在得到“注意”的控制下,海马结构的神经元在进行中间信息的整合处理时,能够直接产生突触可塑性而形成记忆,即所谓的海马记忆。注意这一点跟思维系统在联合皮质上的神经元工作是不同的:在联合皮质的信息处理通道,其中间神经元的信息传递整合没有直接接受中脑网状结构的胆碱能神经的直接投射,而是受丘脑板内核发放的同步激励脉冲的控制,而丘脑板内核向皮质投射的是氨基酸能神经元,与信息处理通道的神经元一样,其兴奋传递不够快速和强烈,在大多情况下难以触发V1.2亚型的高阈值动作电位,而只是触发V1.6亚型的低阈值动作电位,动作电位只是向轴突进行单向传递形成思维活动,而无法向胞体和树突进行反向传递,所以无法产生突触可塑性。这些神经元的记忆效应需要依赖于神经元连接之间的突触发生结构性变化,产生突触重构甚至生成新突触等来改变神经元的连接结构,这种改变需要更长的时间和更多次的信息刺激,但一旦形成其稳定性更高,所以,思维系统在联合皮质上形成的记忆可称为长期记忆或永久记忆。而海马结构形成的记忆依赖于突触的长时程可塑性,能快速形成,但稳定性较低,有效期限较短,称为长时程记忆或短期记忆。
⒊⒎⒊大脑形成陈述性记忆的过程是:当新的陈述性信息输入大脑联合皮质时,在“注意”指向的控制下,信息先是通过扣带回、内嗅区向海马投射,海马结构上的氨基酸能神经在内侧隔核和斜角带核胆碱能神经发放的同步脉冲激励下,依赖STDP突触可塑性形成信息处理的连接通道,也即产生短期记忆;如果这种信息被多次输入或忆起,则联合皮质上的氨基酸能神经元反复受到兴奋刺激,产生突触连接的结构性改变,甚至生成新突触,形成信息处理的直接通道,也即产生长期记忆。而皮质神经元一旦形成直接通道,信息传递便不再通过海马结构的间接的连接通道,则海马结构上的间接连接通道便由于没有信息传递而弱化,最终失效,相关神经元释放出来用于建立新的短期记忆。所以,海马结构在形成短期记忆和转化为长期记忆时起到关键和不可缺少的过渡桥梁的作用,但长期记忆最终却是保存在大脑皮质上。这一海马短期记忆向皮质长期记忆的转换需要一个过程,跟信息的相关性、信息刺激的时间和频繁程度有关。当隔区这些胆碱能神经核团发放功能不足时,便会影响海马结构形成和维持记忆的能力,引起健忘症,而海马摘除时,比如著名的遗忘症患者H.M.,其大脑无法形成新的长期记忆,之前的记忆出现时间分层性的逆行失忆。(关于海马记忆和皮质记忆更详细的描述,可参考申请人之前提交的申请号2014106066977的中国专利申请“神经网络的模拟装置及方法”的说明书中,对记忆的本质和形成、短期记忆与长期记忆的不同及转化部分的内容)。
需要注意的是,内侧隔核、斜角带核、还有Meynert基底核也对感觉皮质和运动皮质进行胆碱能神经投射,所以大脑皮质能够直接对外界感觉信息和运动输出形成记忆,而与海马结构无关。
⒊⒎⒋海马结构的工作,也同时受到来自情绪系统杏仁核的神经投射的调制,以及来自脑干的蓝斑核、中脑中缝背核的去甲肾上腺能神经元,5-HT能神经元的调制。这一点,跟上述“⒊⒍多种调制通路对思维的影响”是一样的。
⒋大脑的其他控制系统。除了思维系统,大脑还有其他几个神经系统,尤其是运动神经系统。严格来说,运动神经系统是动物更早形成的神经系统,用于根据环境信息来控制动物的动作,以实现觅食、避险等活动,而思维系统实质是在运动神经系统的基础上,进化出来的一个用于对各种外部信息尤其是中间信息进行整合处理,以便协调控制运动输出的信息联合处理系统。只是在人,由于语言的出现,使这一中间信息处理系统进化得极其复杂和细致,显得更加重要。
⒋⒈运动神经系统。运动皮质—小脑—纹状体—脊髓等构成大脑的运动神经系统,用于根据输入信息来控制机体运动动作,并对这一过程进行学习记忆。其中运动皮质根据输入信息进行反射或反应,输出空间位置编码的动作信息,在小脑的协同整合下,输出时间编码和频率编码的控制信号,通过锥体系下行投射到脊髓,控制肌肉完成各种具体动作。
⒋⒈⒈运动神经系统的输入信息包括两方面来源:一是来源于感觉皮质的直接投射,包括视觉、听觉、触觉、嗅觉等等,运动皮质根据感觉皮质投射的感觉信息,进行直接反射和反应,包括非条件反射、条件反射、和通过学习记忆的联合反应,控制肌肉进行运动动作,这种控制跟思维系统无关,所以是不需“注意”和“意识”的控制。我们在日常生活中不知不觉便完成的大多数动作,便属于这个。二是来源于联合皮质的中间神经元的输出投射,是大脑思维活动的输出结果。我们在有“意识”有“注意”的状态下,根据思维的需要或结果,主动性地“想”要做什么,便属于这个。
⒋⒈⒉不管是有“意识”做动作还是无“意识”的反应动作,运动过程的动作集合的信息是可以学习和记忆的,这也即是现有理论所称的“程序性记忆”。程序性记忆似乎需要纹状体的参与,动作集合的信息先记忆在纹状体中,形成短期记忆,在多次的记忆或反射活动后,动作信息在运动皮质上形成结构性的长期记忆(或叫永久记忆)。这种动作信息的记忆形成机制,跟思维系统中,通过海马与联合皮质来对中间信息(即所谓陈述性信息)形成短期记忆及转化为长期记忆的机制,是一样的。
⒋⒈⒊运动神经系统的同步脉冲控制环路。作用于运动神经系统的同步脉冲控制环路在以往的脑神经研究中没有提及,但申请人分析认为,运动神经系统一样需要同步脉冲来激励和协调控制神经元的活动,而且其同步脉冲控制环路独立于思维系统而存在。两者具有类似的结构和工作机制,而且存在互相交叉,因为思维系统输出的“有意识”的动作需要投射到运动系统来执行,而运动神经系统的部分输出动作也投射到思维系统被其“注意”。
思维系统的控制环路是由“中脑网状结构←→丘脑”构成振荡环路进行往返发放,该环路再由丘脑板内核向外延伸与大脑皮质进行同步脉冲的往返发放,以激励和控制大脑皮质神经元的活动。申请人分析,由于运动神经系统包括了运动皮质和小脑两大部分,其同步脉冲控制也分为两部分,但两部分存在互相联系:⑴、运动系统在运动皮质部分的同步脉冲控制环路由“被盖网状结构←→底丘脑”构成振荡环路进行往返发放,(胆碱能神经);再由底丘脑核(其作用类似于丘脑的板内核)向外延伸与纹状体(包括苍白球)进行同步激励脉冲的往返发放,(氨基酸能神经),以激励和控制纹状体的神经元活动,(纹状体属于运动皮质的中间神经元);纹状体向底丘脑的网状结构核群(似乎是红核或脚内核,其作用类似于丘脑的网状核)进行聚合性的回馈投射,再下行投射到被盖网状结构,维持环路的振荡。⑵、运动系统在小脑部分的同步脉冲控制环路,由中脑前庭或被盖网状结构与某个核团(似乎是下橄榄核)构成“被盖网状结构←→下橄榄核”的振荡环路进行往返发放,(胆碱能神经),下橄榄核再通过爬行纤维向小脑皮质的浦肯野细胞进行同步激励脉冲往返发放,(氨基酸能神经),以激励和控制小脑皮质神经元的活动。而来自脑桥核团的苔状纤维通过颗粒细胞与浦肯野细胞进行的交叉连接,才是运动信息的传递和整合包括学习记忆的通路。(参考图18的运动神经系统控制环路的信号投射结构示意图)。
运动系统的两部分同步脉冲控制,虽然有不同的投射通道,但两者在同步脉冲发放的起源即被盖网状结构上是统一的(至少是密切联系的),使得两者的脉冲发放存在协调关系,以实现运动皮质与小脑的动作输出的信号保持协调。(或者说,运动皮质的同步脉冲控制与小脑的同步脉冲控制,实质只是运动系统同一个同步脉冲控制环路的两个发放方向?)。
运动神经系统同步脉冲控制环路向运动皮质和小脑投射发放的同步脉冲,申请人推测其作用是:⑴、运动皮质输出的空间编码动作信号,投射到小脑严密排列的神经元,在同步脉冲的协同作用下,转换为时间编码和频率编码的输出信号,以控制肌肉进行精准的运动动作。⑵、通过类似于思维系统的“注意”形成和切换的工作机制,来协调控制运动皮质和小脑的动作输出,避免出现动作输出的矛盾和冲突。⑶、通过同步脉冲的发放节律,控制运动皮质和小脑神经元输出的脉冲发放节律,从而控制机体运动动作的快慢。(类似于思维系统通过同步脉冲发放节律来控制思维活动的步进速率)。
⒋⒈⒋运动皮质是运动神经系统的输出区域,通过锥体系的下行输出纤维输出运动信号,控制机体进行各种运动,但对于每个运动动作具体如何完成的程序和细节,则由小脑来协调完成。运动皮质处理的信息仍然是空间位置编码的,即根据空间结构的不同位置的神经元活动来传递和反射各种动作信息,运动皮质的输出同时投射输入到小脑,小脑在同步脉冲的协同作用下,将这种空间编码的信息转化为发放时间和发放频率不同的动作电位脉冲发放,(类似于电子技术的并行信号转换为串行信号。这一转化过程跟感觉输入比如视觉输入从外侧膝状体投射到视皮质的串行转并行的处理过程,似乎刚好相反),并作为小脑的输出信号,重新投射到皮质(或脊髓),与皮质的输出信号进行协同整合,最终整合为各条运动神经的输出信号,通过向所控制的肌肉发放动作电位脉冲的时间先后(时间编码)和发放频率(频率编码),来控制该肌肉在运动动作中的收缩或松弛的时间和力度,实现对运动动作的协调和精确控制。所以,由运动皮质和纹状体构成的这部分运动系统,主要负责的是“做什么”动作,是由多个动作构成的程序化的动作组合,(尤其包括跟思维系统的输出相关的动作组合),以及这些动作的学习记忆(即程序化记忆)。而小脑负责的是“如何做”,通过对肌肉的精确控制来完成各种动作,(可能包括跟思维系统没有关联的直接的机体反射和反应动作)。申请人推测,从动物进化到人,由于小脑储存记录了实现语言输出的动作(包括说话和书写)的大量程序和细节,所以才导致了小脑体积和神经元数量的急剧增加。
⒋⒈⒌运动神经系统也一样受到下丘脑和脑干中一些神经核团的调制信号的控制。包括来自黑质的多巴胺能的神经投射,来自蓝斑的去甲肾上腺能的神经投射,来自网状结构内侧区的5-HT能的神经投射,等等,这些调制性信号的作用是控制运动神经系统的工作状态,以便与思维系统和内脏内分泌系统的工作状态相配合和相适应。
⒋⒉内脏神经系统。通过内脏感觉神经来对各个内脏工作状态进行感知,通过脑干、下丘脑等的一些神经核团进行反射处理,再通过交感神经和副交感神经去控制调节内脏的工作。按照现有解剖资料,内脏神经系统的主要神经核团在下丘脑,包括控制交感反应的下丘脑外侧区及后区,刺激这个区域可引起心跳加快、血压升高、呼吸增加等反应;控制副交感反应的下丘脑前区及内侧区,刺激这个区域可引起心跳减缓、外周血管扩张等反应。见下面⒋⒋⒈部分的内容。
⒋⒊生化反应系统(包括免疫—内分泌系统)。通过感知机体内各种内分泌物质和非内分泌生化物质的含量,将信息投射到上下丘脑一带,通过其神经核团的反应处理,再反馈去控制各种内分泌物质的分泌或者调节内脏的工作。按照目前的解剖资料,生化反应系统的主要神经核团在上丘脑(比如松果体)和下丘脑部分区域(视上核、室旁核等)。由于缺乏详细解剖资料,无法对具体工作过程进行详细描述。
⒋⒋外部信息处理环路和内部信息处理环路。申请人根据生物体对外部信息和内部信息的控制机制,将大脑各种信息处理归纳为两大环路,即“外部信息控制环路”和“内部信息控制环路”。前者包括了思维系统和运动神经系统,对各种外部信息(视觉、听觉、嗅觉、触觉等)进行感知和整合处理,并输出控制信号对机体运动动作进行控制,(包括说话和书写这种特殊的运动);后者包括内脏神经系统和生化反应系统(包括免疫—内分泌系统),对各种机体内的信息(内脏工作状态、内分泌物质、非内分泌的生化物质)进行感知和整合处理,并输出控制信号对内脏、免疫—内分泌系统进行控制。前者的活动由于能够被思维系统所感知,所以是可以有知觉和意识的,而后者则大多无法被思维系统所感知,所以是无知觉和无意识的;前者的神经核团主要集中在大脑的物理位置的相对上部,所以也可称为“上环处理环路”,简称“上环路”,后者的神经核团主要集中在大脑相对下部,所以也可称为“下环处理环路”,简称“下环路”。在同一环路的内部,各系统之间的兴奋影响和调制作用比较直接而明显,而在内外两个环路之间,也存在相互调制的作用。其中上环路的工作在前面已经有描述,下面对下环路进行描述。
⒋⒋⒈内部信息控制环路(下环路)的构成。主要包括内脏神经系统、生化反应系统(含免疫—内分泌系统)、以及它们之间的控制环路。由于机体各种内部信息的信息量较少,信息变化缓慢,所以其信息的感知输入、识别、记忆、反应、输出控制的各种环节的神经元数量都较少,大多没有形成独立的神经核团,而是聚集在同一核团中。
其中内脏神经系统的信号投射如图19所示。一方面,在脊髓位置已经形成部分内脏的简单的反射弧,对内脏活动进行反馈控制,另一方面,内脏感觉纤维集中在迷走神经通过脊髓上行进入脑干并主要投射到孤束核。孤束核是内脏感觉信息的主要中继核团,其输出一路投射到迷走神经背核,形成内脏神经系统的在脑干层面的低级反射环路,由迷走神经背核发内脏运动神经去控制内脏器官的工作;另一路直接投射到下丘脑视前核、室旁核、背内侧核等区域,传递内脏感觉信息;还有一路投射到脑干网状结构外侧区,尤其包括内侧与外侧臂旁核,内外侧臂旁核发胆碱能神经向下丘脑进行投射。下丘脑的输出则通过迷走神经背核,发内脏运动神经去协调控制内脏的工作。申请人推测内脏神经系统同样具有同步脉冲控制环路,其环路主要由“脑干网状结构外侧区←→下丘脑”构成,其中孤束核直接向下丘脑投射的通路传递的是内脏感觉的具体信息内容,而向脑干网状结构外侧区进行的投射是不包含具体信息的“报告”式投射;而脑干网状结构外侧区(尤其包括内外侧臂旁核)向下丘脑投射的胆碱能神经便是控制环路的同步激励脉冲;同时,下丘脑也向脑干网状结构外侧区进行下行投射;以此形成“脑干网状结构外侧区←→下丘脑”的往返发放的振荡环路也即“下环振荡环路”。
所以,在脑内,控制内脏活动的内脏神经系统包括两大环路:一是由“内脏(感觉)神经→孤束核→迷走神经背核→内脏(运动)神经”构成的在脑干层面的低级反射环路,由简单的直接的反射环路分别对各个内脏器官的活动进行感觉和反射;二是由“脑干网状结构外侧区←→下丘脑”构成的联合反射环路,接受各种内脏器官的感觉传入,并接受大脑其他神经系统(思维神经系统、运动神经系统、情绪神经系统、内分泌神经系统)的调制输入,经过联合处理后,对脑干的低级反射环路的活动进行激励和调制,从而对内脏器官的工作进行平衡控制,比如,调节呼吸的频率和幅度,调节心跳的速率,调节血压等等。由于缺乏资料,申请人无法推断这一振荡环路的振荡节律,但从心跳的节律和思维振荡环路的节律来判断,估计是比较低的,(可能低于3至4赫兹),而且,也无法推断这一控制环路是否跟思维系统的控制环路是一样的工作机制。内脏神经系统也接收脑干一些调制性神经核团的调制性神经投射,如发自蓝斑的去甲肾上腺素能神经投射,发自中缝背核和中央上核的5-HT能神经投射。
而生化反应系统的信息处理核心在上丘脑及下丘脑部分区域,并推测同样拥有一个控制环路,由“脑干网状结构(正中区)—上丘脑(及下丘脑部分区域)”构成。由于生化反应系统(包括内分泌系统)这方面的解剖资料很少,所以无法进行更详细的描述。
⒋⒋⒉下环路的相互调制。下环路包括内脏神经系统和生化反应系统(含内分泌系统),这两个系统之间密切联系互相影响,共同构成对机体内脏器官和内分泌工作的协同控制。由于这两个系统跟机体的健康状况密切相关,如果能够揭示这两个系统的具体工作过程和机制,以及它们之间的相互调制关系,便能够对各种内脏和内分泌异常疾病的成因和治疗,带来有的放矢的指导意义。这方面申请人拟在另外的专利申请中公开,这里不再阐述。
⒋⒋⒊上下环路的互相影响。其相互影响和调制主要通过以下两方面来进行。
⑴、上下环路的神经元活动的相互影响。由于两个控制环路的核心环节都在脑干网状结构上,如果某一环路的网状结构神经元活动更活跃,会造成对营养物质(血氧、血糖、各种神经递质和调质及其合成原料等)的争夺而影响另一环路的网状结构神经元活动,并可能通过调制性神经核团对另一环路进行调制,从而影响了另一环路的工作。其中下环路对上环路的调制和影响,可参见前面⒊⒉⒎,⒊⒌⒈,⒊⒍⒊,⒊⒍⒋部分的内容。而上环路对下环路的影响,由于涉及的是身体的内部活动且反应非常缓慢,所以比较隐蔽。比如,当大脑在紧张思维,思维振荡环路的振荡节律加快,在脑干网状结构这一环节的神经元激活发放动作增加,会影响下环路的工作不稳定,从而对内脏器官和内分泌系统造成负面影响。而在思维系统神经活动下降时,比如,深度睡眠时,思维振荡环路振荡节律很低,在网状结构环节的神经元活动缓慢,有利于下环路的工作,使内脏器官和内分泌系统得到恢复。
⑵、通过情绪系统进行双向调制。申请人认为,大脑的情绪系统直接连接内外两个环路,广泛接收两个环路的信息,反过来对它们的工作进行双向调制,以便使机体能够更协调的适应和应对各种内外信息的变化。这将在下面进行独立描述。
⒋⒌情绪系统。申请人曾经反复犹豫要将情绪系统划分在上环路还是下环路,因为情绪系统既能够直接接收各种外部信息,(包括视觉、听觉、嗅觉等外界信息和中间信息比如“事件”),也能够直接接收各种内部信息,(包括内脏工作状况和体内生化物质的信息,尽管这时我们大脑的思维系统不“知道”),既能够对思维系统和运动系统产生明显的影响,(比如激动时的胡言乱语,兴奋时动作力量大增),也能够对内脏和内分泌系统产生明显的影响,(比如激动时心跳加快,恐惧时肾上腺激素增加,毛骨悚然),后来在分析内外两个环路的相互调制作用时,才明白生物体正是通过情绪系统来联结两个环路,用于对它们的工作进行协调和双向调制。
按照现有解剖资料,情绪系统的主要神经核团是杏仁核。杏仁核的输入和输出的信号投射如图20。其中输入投射包括:⑴、来自广泛的大脑皮质和海马的中间信息的输入,比如来自语言或文字信息的内容可以使人感动或气愤;⑵、来自嗅觉和味觉的外部感觉信息的直接输入,比如特殊的嗅觉或味觉可以使人感觉愉快或恶心;(视觉信息无法直接影响情绪,而需要经过识别后的内容才能够影响情绪);⑶、来自下环路的、主要通过下丘脑和脑干投射的内部信息的输入,比如来自内脏或体内生化物质的异常可以使人感觉痛苦或难受;⑷、来自脑干调制性神经核团的信号输入。而杏仁核的输出和调制作用包括:⑴、向纹体的投射,以影响运动神经系统的神经活动,比如兴奋或恐惧时能够增强运动的速度或力度;⑵、向皮质和海马的投射,以影响思维系统的神经活动,比如激动时思维加快或容易出错;⑶、向下丘脑、孤束核和迷走神经背核的投射,以影响内脏神经系统和内分泌系统的活动,比如激动时心跳加快,恐惧时肾上腺激素增加,毛骨悚然;⑷、向脑干的投射,以影响多个调制性神经核团的活动,进而进一步影响大脑其他系统的工作。从杏仁核的这些输入输出关系及其影响可看出,以杏仁核为核心的情绪系统密切联系内外两个信息处理环路,对它们的神经活动进行协调和双向调制,从而影响它们的活动和输出,以更好地应对信息变化。情绪系统对信息的输入和反应同样是可以学习记忆的,其工作机制跟思维系统的海马类似。参照大脑其他神经反射系统的工作机制,申请人推测,杏仁核似乎也应该跟丘脑以及某个投射胆碱能神经的核团构成一个控制环路,但由于缺乏解剖资料,无法进一步描述。
⒋⒍不同神经递质的神经元在大脑信息处理的不同作用。大脑存在多种不同的神经递质及其膜受体,也存在释放这些神经递质的不同神经元,包括胆碱能神经元、氨基酸能神经元、单胺能神经元、和神经肽类神经元等等。大脑的很多神经核团往往同时存在多种不同的神经元,并形成不同的神经投射,而这些不同的神经投射既可能共同投射到同一脑区或脑核团,也可能彼此交叉投射,而且,不同神经递质的神经元工作异常有时却会造成同一种脑功能异常或疾病,这些都容易给分析大脑的信息传递通路及工作机制造成困扰和误导。根据各种不同神经递质的传递特性和神经投射通路,并结合前面所述的大脑对于信息的感知输入、传递、记忆、反射输出的工作机制和控制机制,分析不同递质的神经元在大脑信息处理中的不同作用,以此能够更好的分析和理解大脑的工作机制。
⒋⒍⒈氨基酸能神经元。其神经递质包括兴奋性氨基酸的谷氨酸(Glu)和天冬氨酸(Asp),以及抑制性氨基酸的γ—氨基丁酸(GABA)和甘氨酸(Gly)。氨基酸作为神经递质,其传递通道容易受其他递质的影响和调制,并且能够形成突触可塑性(STDP可塑性),所以,氨基酸能神经元是大脑构成信息处理通道的最基本和最主要的神经元。中枢神经系统几乎所有信息传递和处理通路都主要以兴奋性氨基酸能和抑制性氨基酸能神经元互相配合来构成,以此来实现信息的输入、传递、整合、反射和输出,并形成记忆、思维和意识的脑功能。氨基酸能神经元也构成部分调制性神经通路。
⒋⒍⒉胆碱能神经元。其神经递质为乙酰胆碱(ACh)。在中枢神经,胆碱能神经递质的门控通道反应极快,无需细胞膜预先存在去极化,在静息电位下和超极化的情况下,一旦配体与受体结合便可直接引起门控通道开放,钙离子快速内流,引发强烈的膜兴奋,大多能直接引发动作电位的爆发。而且,胆碱能神经元的轴突释放ACh是快速又短暂的,释放后ACh除了跟受体结合,也能从突触间隙扩散出去并被胆碱酯酶迅速降解,也即释放后突触间隙的ACh能够被快速清除。所以,胆碱能神经元爆发动作电位后其发生作用和停止作用的时间都极其短暂,而且能够使所投射的后级神经元产生强烈的膜兴奋并爆发动作电位,这一强烈而快速的传递特性,使胆碱能神经元在中枢神经活动中主要用于同步激励,尤其是构成同步脉冲振荡环路,能够无需其他神经通路的协同作用便可自行形成往返循环的脉冲发放,并以此作为同步激励信号从时序上对其他神经活动进行控制,使大脑信息处理的神经活动能够有序进行,在此同步脉冲起到类似于电子计算机的时钟信号的时序控制作用。而如果所投射的神经元为氨基酸能神经,胆碱能神经快速而强烈的兴奋激励,还能够使其爆发高阈值的动作电位,从而产生突触可塑性,形成记忆效应。胆碱能神经元的传递特性也使其适用于运动神经输出、神经—肌肉接头、心肌、平滑肌、和部分感觉信息的输入和传递,但一般不适用于信息记忆处理的直接通道。
⒋⒍⒊单胺能神经元。其神经递质是含有单胺基团的生物胺。单胺能神经元包括多巴胺能神经元、5—羟色胺能神经元、去甲肾上腺能神经元和肾上腺能神经元、以及组胺。单胺能神经元的作用是对神经活动进行调制。尤其是参与情绪系统,对思维系统、运动神经系统和内脏神经系统的信息处理进行调制。其中:⑴、去甲肾上腺素(NE)能神经主要存在延髓和脑桥的神经核团(比如蓝斑核),向大脑皮质、丘脑、下丘脑、边缘系统等广泛区域进行投射,其作用主要是抑制性调制,对所投射区域的神经元活动进行抑制。当NE的作用过强时,可过度抑制思维系统的神经元活动,导致神经元活动异常下降,如出现抑郁症。⑵、5-羟色胺(5-HT)能神经主要存在脑桥中央上核和中脑中缝背核等核团,向丘脑、大脑皮质等广泛区域进行投射,其投射区域跟NE类似,但其作用主要是增强性调制,对所投射区域的神经元活动起到增强兴奋整合的作用。当5-HT过少时容易出现抑郁症,而过多时容易出现躁狂症。⑶、多巴胺(DA)能神经主要存在中脑和间脑的神经核团(比如黑质),向大脑皮质和边缘系统等区域进行投射,其作用主要也是增强性调制。⑷、组胺能神经主要存在于下丘脑后部的结节乳头核,上行向前脑的广泛区域、下行向脑干和脊髓进行投射,其作用主要是进行睡眠和觉醒的调制。调制性神经元的所谓增强性和抑制性调制作用并非绝对的,因为信息处理通路的神经元活动本身就存在互相抑制的现象,氨基酸能神经也分为兴奋性和抑制性两种,对抑制性神经元进行抑制性调制,实质上便相当于对所抑制的兴奋性神经元进行增强性调制。正是单胺能神经元这些复杂而相互交叉的调制通路,使大脑的神经投射通路显得非常复杂,也正是这些复杂而相互影响的调制作用,使大脑的思维活动产生复杂的各种变化,并产生了情绪、情感、性格、欲望、动机等高级脑功能,还有做梦以及各种精神病等现象。
⒋⒍⒋神经肽能神经元。其神经递质为大分子多肽即神经肽(SP)。神经肽种类繁多。其作用产生缓慢而持久,主要是对大脑自身神经元的生理活动,以及对机体的内脏和内分泌的工作进行调节。
另外还有嘌呤类物质,一氧化氮,一氧化碳等,也参与神经元的活动,对神经活动形成影响。
⒋⒍⒌大脑的其他胆碱能神经投射。胆碱能神经元的兴奋传递具有快速且强大的特点,在中枢神经活动中被用于作为同步激励脉冲。除了主要存在脑干网状结构与丘脑、上下丘脑等构成同步脉冲振荡环路,大脑还有其他发出胆碱能神经投射的主要核团,便是边缘系统的隔区的内侧隔核和斜角带核、还有基底前脑的Meynert基底核,(基底前脑复合体)。其相关的信号投射如图21。这些胆碱能神经核团的投射和作用包括:⑴、上行向皮质尤其是感觉皮质和运动皮质进行投射,对感觉信息和运动信息的整合处理进行协同激励,尤其是协同产生突触可塑性以形成认知记忆和运动记忆;⑵、向扣带回和海马结构进行投射,对思维系统的中间信息(即陈述性信息)的整合处理进行协同激励,尤其是协同产生突触可塑性以形成海马记忆;⑶、与杏仁核和下丘脑进行双向的相互投射,以对彼此的工作进行相互调制,并协同形成情绪记忆;⑷、向中脑一些核团的投射。另一方面,这些胆碱能神经核团也同样受到来自蓝斑核、中脑中缝背核、中脑被盖腹侧的去甲肾上腺素能、5-HT能、多巴胺能的神经调制。
隔区和基底前脑这些胆碱能神经核团,与脑干网状结构胆碱能神经核团之间,通过前脑内侧束进行往返联系。由于缺乏更详细的解剖资料,申请人弄不明白的是,到底是隔区和基底前脑这些神经核团受脑干网状结构的控制,还是脑干网状结构受到这些核团的控制?以谁为主?还是彼此相对独立工作只是互相影响?从涉及对整个大脑信息处理的控制的广泛性来看,似乎脑干网状结构更为重要。
另外,在小脑、大脑皮质、纹状体、海马等内部,也有胆碱能神经的短的局部投射,推测其作用是对所接受的胆碱能神经的长投射的延伸,同样起到协同激励的作用。
⒋⒎简单的神经活动方式和强大的大脑功能。综合上面所述,从大脑的整体结构来看:大脑是由思维系统、运动神经系统、情绪系统、内脏神经系统、内分泌神经系统等多个信息处理系统集合构成,这些系统交叉影响,实现对各种外部信息和机体内部信息进行感知和处理。从神经元的类别来看:大脑主要采用氨基酸能神经来构成信息处理的主通道,采用胆碱能神经产生激励脉冲来对信息处理进行激励和时序控制,采用单(组)胺能神经及神经肽能神经来对信息处理进行各种调制,以此共同实现对信息的联合处理。从神经元的活动方式来看:大脑普遍采用由2至5级的神经元往返投射来构成闭环的神经小环路,以兴奋整合和往返发放动作电位的方式来进行属于自己的某种信息处理。对于某一神经小环路,一方面环路中某些节点的神经元接收其他神经环路的投射来实现信息输入或调制输入,另一方面环路中某些节点的神经元也向其他神经环路进行投射来实现信息输出或调制输出,“环环相扣”形成了庞大的神经网络。
这种往返投射的神经小环路只需按照自己固定的简单的活动方式来进行“属于自己的”信号整合处理,而不必“理会”整个神经网络的信号如何复杂,所以有利于保持环路自身神经活动的状态稳定,而又能通过环与环之间纵横交错的相互投射来进行输入和输出各种信息。而且,虽然每个神经元都只是在进行简单的兴奋整合和动作电位发放,每个神经小环路也都只是以同样的简单的反射动作在对信息进行传递和反射,但由于:⑴、神经小环路数量极其巨大,⑵、神经元具有兴奋整合的功能,能够同时对多个其他神经环路的信息输入进行整合处理,而且这种整合处理是具有精确的模拟量处理功能的,⑶、神经小环路内部和神经小环路之间的神经投射,其突触具有传递效能的可塑性,⑷、各个神经小环路之间,能够通过突触重构来取消或形成新的神经投射联系,⑸、具有多种不同的神经元,以不同的神经递质和调质、不同的受体和不同的离子通道来进行不同的神经动作,所以,这些神经小环路构成的庞大神经网络能够对各种复杂信息进行功能强大的联合处理。大脑正是以数量巨大的神经小环路,通过这种“环环相扣”、“无限延伸”的方式,来实现对各种复杂的海量的信息的处理。
而且,在生物进化过程中,各种神经小环路能够根据信息量的增加而相应增加数量,并通过这种“环环相扣”的结构,继续保持相互投射,构成几乎可无限扩展的神经网络,而在扩展的过程中其神经投射的基本架构却能够保持稳定,能够保持生物进化的延续性和稳定性。所以,人与老鼠的大脑,其信息传递处理的基本架构是一样的,不同的只是各部位的神经组织的神经元数量。
也正因为这种互相投射的结构,所以,对于同一种神经功能异常或疾病,既可能是由于其相关的神经环路的神经元工作异常,而直接造成信息处理异常;也可能是由于对该神经环路进行激励和同步控制的神经元工作异常,使其无法形成正常有序的工作;还可能是由于对该神经环路进行调制的各种调制通路的神经元工作异常,而使其受到非正常调制而工作异常。在治疗这些疾病时,需要对此进行分析,才能得到更合适的治疗方案。
⒌对大脑几种现象的解释。申请人发现,根据前面对“思维”、“注意”和“意识”的本质,以及对“注意”控制机制的分析,可以很容易又很好地解释一些脑现象,包括一些目前难以解释的精神异常的疾病,下面对其中一些进行简单描述。
⒌⒈脑电波。以往认为,脑电波是皮质上大量神经元在丘脑同步脉冲的控制下,同时发放动作电位所产生的,所检测到的信号便是大量的皮质神经元同步爆发动作电位的突触后电位的变化信号。因此,也认为思维是依赖于皮质的大量中间神经元进行同步激活发放所进行的。
⒌⒈⒈申请人认同脑电波是丘脑同步脉冲的发放所导致的,但分析认为,脑电波并非皮质的大量神经元同步激活所产生,其理由如下。
⑴.皮质的神经元活动是大脑思维、记忆及各种反应的基础,那么,如果在同一时刻皮质有大量的神经元一起同步发放动作电位,则会导致大量神经元的连接结构在同一时刻发生变化或更新,这必然会导致大脑的思维、记忆和反射活动的混乱和不稳定。
⑵.不符合生物进化的尽量节省耗能的原则。神经元的动作电位发放是一个耗能较大的活动,从生物进化的角度看,生物体不应采用大量的皮质神经元的同步活动来产生一个最小单位的信息变化,而更会选择进化到采用少量(最好是一个)神经元的活动来实现这一结果,以尽量节省非必要的耗能。
⑶.无法解释脑电波的不同波形的振幅变化。如果脑电波是皮质大量神经元动作电位发放所产生的,那么当大脑处于兴奋和快速思维时,(出现β波),这时脑皮质有更多的神经元参与活动,按说脑电波振幅便应该更大,而大脑在深度睡眠或昏迷时,(出现δ波),这时脑皮质只有很少神经元在活动,按说脑电波的振幅应该很小。但实际情况刚好相反,出现β波时脑电波的振幅更小,只有十几到几十微伏,而出现δ波时脑电波振幅最大,超过50微伏,甚至上百微伏,呈现振荡节律与振幅的负相关现象。
⑷.脑电波上没有动作电位脉冲的信息。如果脑电波是皮质大量神经元同步发放动作电位所产生的,即使是这些神经元发放动作电位后的突触后电位产生的,那么,在检测到这些突触后电位的变化信息(也即现有所得到的脑电图)的基础上,还应该检测到很尖很窄但振幅很高的动作电位在轴突传递的脉冲信息,目前的检测技术完全能够做到,但实际上没有,没有检测到动作电位爆发时的脉冲信息。
⒌⒈⒉申请人根据思维以及思维振荡环路对思维进行控制的工作机制,分析认为:脑电波并非皮质上大量神经元同步激活发放动作电位所产生的,而是丘脑向皮质进行同步脉冲发放后,皮质上的大量神经元由于仅依靠同步脉冲的刺激未能整合触发动作电位,而只能出现衰减性的膜局部兴奋,这些大量神经元的膜局部兴奋同步地进行衰减性的扩布,其引起的电场同步波动才是产生脑电波的真正原因。
丘脑向皮质的神经投射是扩散性的,即丘脑的每一个神经元的轴突投射与皮质上的一组(N个)神经元建立有突触联系。某一时刻,当丘脑的一组(M个)神经元受网状结构的上行发放而激活时,这一组神经元同时向皮质上的相关分区域的大量(M×N个)神经元进行同步脉冲发放,但同步脉冲只是形成一方面的兴奋刺激,大多情况下尚不足以单独使皮质受刺激的神经元触发动作电位,只有那些同时(或短时间之前)受到其他兴奋激励的个别神经元,(也即链式激活链路上的相关神经元),才能由于具有足够的兴奋整合而触发动作电位,(并向丘脑回馈发放脉冲,引发下一个同步脉冲,以维持链式激活的继续进行)。而其他的那些大量(M×N个)的神经元,由于缺乏兴奋整合而无法触发同步脉冲,丘脑的同步脉冲只能使它们产生突触后的膜局部兴奋,并在膜上进行扩布并衰减。于是,皮质上这些大量的神经元,一齐受到同步脉冲的兴奋激励,又同样没有触发动作电位,其局部兴奋在膜上进行同步扩布,形成电场的同步波动,便形成了脑电波。所以,脑电波并非是皮质大量神经元“一起同步激活(触发动作电位)”后产生的,反倒是皮质上的大量神经元由于“一起受刺激”后“一起未能激活”产生的。
⒌⒈⒊这样的工作机制更符合生物进化的尽量节省耗能的原则,也可以解释脑电波振荡节律与振幅负相关的现象。当大脑处于兴奋、激动和快速思维时,(出现β波),这时大脑思维活动的“注意”指向非常集中,丘脑只向投射范围比较小的皮质进行快速的同步脉冲发放,所以,受同步脉冲刺激后未能触发动作电位的神经元数量较少,它们产生的膜局部兴奋的同步扩布所产生的脑电波的幅度自然便较小。而大脑在深度睡眠或昏迷时,(出现δ波),这时大脑没有思维活动,也没有具体的“注意”指向,丘脑与网状结构进行缓慢的振荡发放,并对皮质广泛区域的神经元进行同步脉冲发放,但由于同步脉冲间隔期过大而无法激励这些神经元的激活,所以,皮质上受同步脉冲刺激后未能触发动作电位的神经元数量非常多,它们的膜局部兴奋同步扩布所产生的脑电波的幅度自然便更大。
⒌⒈⒋按照这样的解释,在某一时刻,不管丘脑的同步脉冲发放是向皮质小范围还是广泛区域进行投射,皮质上绝大部分神经元都是受激励后未能触发动作电位的,能够激活触发动作电位的只是极少量神经元,这极少量的神经元的动作电位信号虽然幅度更高,但毕竟数量极少,而且被掩盖在大量未能激活的神经元的局部兴奋同步扩散的信号之中,所以我们在脑电图中未能检测到这些动作电位的信号。只有在大脑工作异常时(某些神经病),丘脑异常地同时向皮质多个区域进行高节律脉冲发放,引起皮质多个位置的神经元同时触发动作电位,这时我们才有可能检测得到这种既高频又高振幅的脑电波。
当然,皮质神经元也可能出现非思维活动的激活放电,即所谓的自发放电,但申请人认为自发放电应该是零散的偶发的,所以也不会在脑电图上造成很大影响,可能只是微弱的噪音。见“⒉⒋⒌神经元的自发放电”部分。另外根据解剖资料,皮质上似乎也存在少量的胆碱能神经元,申请人推测这些胆碱能神经元不属于信息处理通路,而是同步脉冲的接续,它们可能会跟随同步脉冲而激活放电,起到类似同步脉冲的作用,但数量较少。
⒌⒈⒌以往认为脑电波是皮质大量神经元同步激活发放动作电位的理论,也使以往对大脑思维的工作机制的理解造成很大的误导,以为脑思维的每一次动作都需要依赖于皮质的大量神经元的同步活动来实现,所以认为皮质对信息的记忆和反射是采用大量神经元通过时间编码或频率编码,在同步脉冲的控制下通过精确的同步发放来实现。但实际上,时间编码和频率编码主要存在于感觉传入和运动输出的通路中:在感觉传入方面,感觉神经元采用时间编码和频率编码来记载和传递各种感觉刺激的性质和强度;在运动输出方面,采用时间编码和频率编码来实现对肌肉运动方向和力度的精确控制;所以,大脑在传入和输出方面是并行工作的,我们可以同时感受多方面的感觉输入,也可以同时进行多种肌肉动作。而大脑在对中间信息的记忆和思维上,采用的是空间位置编码来实现的,就是特定的一个或一组神经元来对应着一个特定的信息,这些单个或单组神经元有序的依次活动(链式激活),便形成了信息流,形成了思维或信息的忆起输出。所以,大脑的思维是串行工作的,在同一时刻“思路”只有一个,当思维采用语言进行时,思维只能逐字逐词进行。
⒌⒉睡眠的本质、启动与维持。现有研究揭示睡眠可能由下丘脑某些核团触发,视觉由下丘脑的视上核、乳头体等形成反射回路,产生日夜节律(即生物钟)控制信号,再上行向丘脑和皮质、下行向脑干和脊髓等进行调制性的投射,(组胺能神经元),对睡眠和觉醒进行控制。但对其具体控制机制尚不清楚。而且,这样的理论难以解释大脑可以由主观意识来控制睡眠,包括偶尔的熬夜和睡懒觉等行为。
⒌⒉⒈睡眠的本质。申请人分析认为,睡眠的本质是大脑外部信息控制环路的一种低节律工作状态:当“网状结构—丘脑”振荡环路的振荡节律较低,低于3赫兹时,丘脑向皮质发放的同步脉冲的间隔期过大,无法使皮质神经元得到连续的兴奋并整合触发动作电位,链式激活无法进行,于是思维系统失去注意、思维和意识,进入睡眠状态。
⒌⒉⒉睡眠的启动机制。根据“⒊⒌思维振荡环路的节律变化及对大脑工作的影响。”的内容,申请人认为,使大脑进入睡眠状态,也即使思维系统振荡环路的振荡节律降低至3、4赫兹以下,有两种主要的启动方式:⑴、通过下环控制环路对思维控制环路的调制作用。这包括来自下丘脑乳头体的日夜节律信号的调制,(生物钟的影响),也包括来自内脏神经系统和内分泌系统的调制,(疲劳、不舒服等状态的影响),这些调制信号直接对思维控制环路各个环节(尤其是中脑网状结构)的神经元活动进行抑制,使其往返发放的振荡节律逐渐下降,最终进入睡眠。这一方式是身体内部的需要,是被动地启动睡眠。⑵、通过皮质思维活动对思维控制环路的自我抑制。由于皮质的神经元活动对丘脑板内核和网状核的下行投射能够影响其神经元兴奋整合的速度,皮质神经元活动对网状结构的下行投射能够影响网状结构的“注意”通路的兴奋,从而影响整个环路的发放节律,所以,通过减少思维活动,(使自己平静,不思考),降低皮质神经活动的下行激励信号,从而逐渐降低思维控制环路的振荡节律,最终进入睡眠。这是主观上想要睡眠,是主动启动睡眠。
这两种启动方式,每一种都能够启动睡眠,但其中一种方式的启动又往往伴随另一种的响应,两者形成正反馈而共同使大脑进入睡眠。当两者出现不协调时,是否能启动睡眠便看哪一方面的作用更强大:当身体不舒服时,虽然主观上你不想睡眠但身体使你睡得昏昏沉沉,比如嗜睡;或者主观上想睡眠但身体又使你睡不着,比如失眠。当身体需要睡眠,你也能够在一定时间内继续工作而坚持不睡眠,比如熬夜;或者当身体没有需要,但无聊时你能够使自己继续睡,比如睡懒觉。另外,根据“⒊⒌思维振荡环路的节律变化及对大脑工作的影响”,还有两种方式可以影响睡眠:⑶、情绪系统的调制。情绪低落容易进入睡眠,情绪激动兴奋则不容易进入睡眠。⑷、药物的作用。比如麻醉,直接抑制神经活动,进入一种特殊的睡眠。
⒌⒉⒊睡眠时大脑的活动状态。进入睡眠的过程,思维环路振荡节律是逐渐降低的,当振荡节律降到7—4赫兹范围时,属于临界状态,这时同步脉冲的激励尚能够使皮质部分神经元进行兴奋整合并激活,但兴奋整合缓慢,神经元活动不活跃,无法形成连贯清晰的激活链,只有某些偶发的孤立的思维片段。这时大脑虽然还存在“意识”,但对各种刺激反应迟钝,思维模糊,且一般不易形成记忆。这便是大脑临睡觉前(包括非常困倦时)的状况。
在深度睡眠状态下:⑴、大脑思维系统被抑制,没有产生思维活动;⑵、大脑的外部感觉输入被抑制,对外界的刺激无反应或反应迟钝;⑶、大脑的运动神经系统也被抑制,没有形成动作输出。但如果这三者没有同时被抑制,某些方面被抑制而某些方面没有被抑制,便不是深度睡眠,这时大脑会出现有趣的状况,包括做梦和梦游。(后面另有描述)。
⒌⒉⒋睡眠的维持。睡眠一旦触发,也即思维控制环路振荡节律低于3、4赫兹,这时丘脑以这种低节律向皮质发放激励脉冲,由于这种低节律的激励脉冲无法使皮质神经元完成兴奋整合而激活,所以皮质的神经元也就没有向丘脑和网状结构进行下行的回馈发放,而这又使得板内核、网状核和网状结构都无法以更快的速度进行兴奋整合,从而使得整个思维振荡环路继续维持在低节律振荡状态,同时,通过相互之间的调制关系,也使下环路的调制性神经核团保持对思维系统的抑制,共同维持睡眠状态。
⒌⒉⒌失眠。根据睡眠的启动机制,导致失眠也即睡眠无法启动也有几种因素:⑴、下环路对思维振荡环路的调制不足,无法降低其发放节律,所以无法启动睡眠。由于下环路与上环路更多的是互相抑制作用,当年纪较大,下环路活动不足时,对思维振荡环路的抑制不足,所以更容易失眠。⑵、思维活跃,思维通路的神经元活动活跃,无法降低思维振荡环路的发放节律,所以无法启动睡眠。⑶、情绪过于兴奋,同样地影响思维通路的神经元活动过于活跃,无法启动睡眠。⑷、药物的影响,直接使神经活动活跃而无法睡眠。根据导致失眠的这些因素,可以研发出治疗失眠的相关应用技术。
⒌⒊觉醒与思维的启动。觉醒是睡眠的逆过程,当思维振荡环路从低节律重新转化为较高节律(高于8赫兹)振荡时,丘脑向皮质发放的激励脉冲间隔期较短,所以又足以激励皮质神经元进行兴奋整合而触发激活,于是又开始思维活动,进入清醒状态,并通过网状结构的影响和调制神经元的调制作用,也使运动神经系统解除抑制,恢复正常工作状态。
导致从睡眠转换为觉醒、也即导致思维振荡环路振荡节律升高的因素主要包括:⑴、来自下环路的调制信号的改变。在环境光线变亮(或睡眠时间足够)时,下丘脑的调制信号(生物钟信号)作用于思维振荡环路,使得振荡环路的发放节律提高。⑵、来自思维环路自身的神经活动的刺激。包括外界感觉信息的刺激,比如较强烈的听觉或触觉刺激,也包括做梦时皮质神经活动过于强烈时的下行刺激,这些强的刺激投射到中脑网状结构,激励其神经元活动而引发振荡环路发放节律升高。不管以哪种因素,都是使下环路的抑制减弱和思维环路的振荡节律升高,并形成正反馈而进一步加快这一过程。而一旦振荡节律升高至7、8赫兹以上,则丘脑向皮质发放的同步脉冲,又足以激励皮质神经元产生连续的链式激活,思维活动便随之自然地产生,(思维的自涌)。而皮质的思维活动一旦产生,其神经元活动一方面下行激励丘脑和网状结构维持高节律振荡,并引发网状结构的“注意”,另一方面向感觉皮质投射产生“意识”,于是“注意”、“思维”和“意识”便都恢复。觉醒时的思维活动可以是从各种偶然信息开始的,比如听觉触觉的信息、梦中的内容、睡眠前的思维内容等等,但一旦形成思维,便回归到思维系统“注意”机制的控制。
⒌⒋做梦。申请人分析认为:做梦是睡眠状态下,大脑各个通道受到的抑制作用不同步所产生的。大脑皮质的神经活动受到来自丘脑同步脉冲发放、下丘脑、脑干调制性神经核团、边缘系统和Meynert基底核的多种控制,在清醒状态下,感觉输入、思维活动、和运动神经输出三者都不会被抑制,感觉输入信息可以投射到思维系统产生听觉视觉等感觉,思维系统的输出也可以投射到运动系统产生机体动作或说话书写,思维系统的活动还同时投射到联合听觉皮质(尤其是听觉性语言中枢)产生自我感知也即产生意识,但思维系统向联合视觉皮质的投射被抑制,(其生物学意义是避免与视觉输入信息互相干扰,以免使运动输出混乱),所以大脑思维活动包括想象都不会产生图像视觉感知。而在深度睡眠时,思维系统、感觉输入、运动输出三者都被同时抑制,所以没有思维活动,感觉迟钝,肌肉松弛无动作。
但如果这三者没有被同时抑制,其中感觉输入和运动输出的神经活动被抑制,而思维系统没有被抑制或抑制不足,便导致思维控制环路的振荡节律升高,丘脑向皮质进行高节律的同步脉冲发放,(所以做梦时脑电图显现高节律的脑电波),于是皮质神经元产生连贯的链式激活,也即产生思维活动。由于这时感觉输入被抑制,思维活动不会接收真实的感觉信息,而只是在联合皮质的中间神经元之间进行,所以做梦呈现的更多是一种类似于想象的思维活动,但这时思维系统向联合感觉皮质包括联合视皮质的投射也没有被抑制,于是思维过程所产生的各种想象的信息,都投射到联合感觉皮质,产生类似于看到或听到的感觉,也即做梦。由于做梦时运动神经输出也被抑制,所以梦境所产生的动作一般不会被运动系统所执行,四肢肌肉依然松弛,只有动眼神经既受运动神经系统控制,又受上丘与网状结构的控制,所以做梦时会出现眼动动作。对于梦的启动、梦的停止、梦境的忆起,申请人将在另外跟做梦相关的发明中进行详细描述,这里不再叙述。
⒌⒌梦游。申请人分析认为:梦游也是睡眠状态下,大脑各个通道受到的调制作用不同步所产生的,但跟做梦不同,梦游是思维系统被抑制,而感觉输入尤其是视觉输入和运动神经输出没有被抑制或抑制不足。由于视觉输入通路和运动输出没有被抑制,所以视觉信息能够被投射到联合视皮质,梦游者能够看到。而由于联合视皮质有投射通道不需经过思维系统的控制而向运动皮质进行直接投射,并引起运动皮质的反应而输出程序性动作,(这种投射和反应是机体成长过程所学习获得的程序性记忆,保存在运动神经系统中,无需受思维系统的控制,大脑正是依靠这种直接投射和程序性记忆来完成各种无需“注意”也无“意识”的行动,如吃饭、行走、规避障碍物等日常动作),所以,运动皮质接收联合视皮质的投射信号,在没有思维系统参与的情况下,根据之前记忆存在的程序性的反应动作,输出运动信号,控制机体进行运动动作,包括起床、行走、开关门、上下楼梯等动作,也即出现梦游的情况。
梦游时,眼睛受运动皮质的控制而张开,梦游者能够看到,但由于思维系统受抑制,所以这时其视觉是没有“注意”的,表现为眼睛无神,没有眼动或眼动不正常。梦游时没有意识,梦游发生的内容也不会被记忆在思维系统中,即没有陈述性记忆,但如果多次进行某一动作,则该动作可被记忆在运动神经系统中,成为程序性记忆。梦游时思维系统是被抑制的,但联合皮质一些睡眠前比较活跃的神经元可能会影响到感觉皮质对运动皮质的投射关系,所以梦游发生的事情可以跟之前的思维内容有一定关系。
从上述做梦和梦游的产生机制可看到,在睡眠时,如果大脑的外部信息环路的外部感觉输入、思维系统(中间信息处理系统)、和运动神经输出,这三者受到的抑制不同步时,便可能产生异常的睡眠,其中单独思维系统抑制不足时产生做梦,外部感觉和运动神经输出抑制不足时产生梦游。申请人以此推测:单独外部感觉输入抑制不足时,睡眠容易被唤醒;单独运动神经输出抑制不足时,出现睡眠时翻身等无意识动作和肌肉紧张,(由于这时没有外部信息和思维系统的投射,运动系统自身无法产生更多的动作);外部感觉输入和思维系统抑制不足时,外部感觉信息会影响梦境内容,或者睡眠容易被唤醒;思维系统和运动神经输出抑制不足时,做梦会产生动作输出,这时往往会导致做梦和睡眠被惊醒。
⒌⒍精神病。大脑的工作异常,可导致各种脑相关的疾病,以往习惯根据是否有器质性病变而把这些疾病分为神经病和精神病,(甚至还有心理疾病),实际上所有的脑疾病归根结底都是脑神经的工作异常,只不过异常的位置和方式不同而已。其中癫痫、帕金森、阿尔茨海默病等这类病的现有研究已经很多。这里简单分析的,是几种跟思维系统相关的精神病,即清醒状态(也即有“意识”状态)下的思维异常,是思维系统的“注意”控制环路和神经调制通路的协同工作异常所导致的。
如前述,思维活动是思维系统的神经元进行链式激活,并受到“注意”控制通路和多个调制通路的协同调制。在面对信息输入的刺激时,链式激活的进行方向也即思维的思路,受到相关神经元的连接结构(也即记忆,包括知识、事件、经验等)和多个调制通路以及“注意”控制通路的共同影响。在不同人,这些控制通路和调制通路的作用强弱会略有差异,因此造成不同人在面对同样的信息输入时其思路和输出结果不同,形成不同的认知偏好和不同的性格。但如果这些调制作用与正常人的差异超过一定程度,成为异常,便会导致思维的异常。具体的,如果在清醒状态下,这些“注意”控制和调制通路的作用及其强弱产生异常,这时患者的思维系统依然是能够在这些异常的调制作用下进行思维活动,但由于此时的思维活动受到非正常的控制或调制,其思维的思路和结果也即行为表现便与正常人大不一样,这种思路和行为表现也会被正常人所无法理解,所以便被称为精神病。
⒌⒍⒈注意缺陷症。在“⒊⒋注意的控制、维持和切换”部分,申请人分析了思维系统“注意”控制、维持和切换的工作机制,即大脑通过中脑网状结构—丘脑—皮质的分级控制,形成对思维“注意”指向的控制,其中中脑网状结构负责的是将“注意”指向哪一信息通道,丘脑负责的是将“注意”指向同一信息通道中的哪一部分。各类信息通道,包括外界输入信息通道(视觉、听觉、嗅觉等)和思维通道的神经元活动,都有经过聚合性整合的神经输出向中脑网状结构进行“报告式”投射,中脑网状结构通过对各类信息输入的强度和时间进行比较整合,来选择“注意”哪一信息通道,而一旦产生对该信息通道的“注意”,则会通过该通道的皮质神经活动下行投射的回馈、和对其他信息通道的抑制来实现对“注意”的维持,直至有其他信息通道更强烈的信息输入,产生“注意”切换。
根据“注意”维持的工作机制,如果皮质神经活动向中脑网状结构下行投射的回馈信号的强度不足,或者中脑网状结构各信息上行通道相互之间的互相抑制作用不足,便会导致某信息通道受“注意”后,无法正常进行“注意”的维持,使得“注意”的指向异常地很容易在各信息通道之间进行切换。这在宏观上,便表现为大脑无法对某一事情进行持续关注,容易分心走神等等,也即出现了“注意”缺陷。
“注意缺陷”是思维系统的“注意”控制机制、也即用于对思维活动发放同步激励脉冲的控制环路出现异常,使“注意”指向无法正常维持所导致,涉及思维系统方面。而由于在运动神经系统也同样存在这种发放同步激励脉冲的控制环路和控制机制,当运动神经系统出现类似的异常,便是多动症。多动症患者会出现异常的多动和冲动的行为,而且涉及细节性的动作的控制和协调性不良。目前的研究一般把注意缺陷(ADD)和多动症(HKS)归为一种疾病,即“注意缺陷多动症”(ADHD),而把ADHD又分为注意缺陷型、多动型和混合型。
而且,由于运动神经系统包括有受思维系统控制部分(有意识的动作)和不受思维系统控制部分(无意识的动作),当这种控制机制的异常发生在运动神经系统不受思维系统控制的部分时,便会导致抽搐症(Tics,或称妥瑞氏症)。患者会产生不受意识控制的不自主的运动输出,包括不由自主地在脸部或身体做出各种动作,即动作型(Mot or Tic),或不由自主地发出各种诡异的声音,即声音型(Vocal Tic),(声音实质上也是一种动作输出)。
⒌⒍⒉自闭症。根据分析,自闭症也是大脑思维系统的“注意”控制机制工作异常所导致,但跟注意缺陷症相反,自闭症不是“注意”无法维持,自闭症是大脑思维系统的“注意”指向无法在各信息通道之间进行正常的“注意”切换,而异常地过度集中和维持在对内部中间信息进行处理的某些神经通道上,从而也相对地抑制了对其他神经通道特别是抑制了对视觉、听觉、触觉等外界信息输入通道的“注意”,造成患者对外界信息能够感知,但却不易引起“注意”,于是这些外界信息便无法得到思维系统的正常处理。由于患者能够感知外界信息,所以能够进行行走等日常活动和避险等本能的动作(也即无需思维系统控制的动作),也不会出现幻觉,但各种外界输入信息(视觉、听觉、触觉等)不易引起中脑网状结构“注意”控制通路的“注意”,也就不易得到思维系统的处理(也即缺乏同步脉冲的发放激励)。所以,患者对各种外界输入信息包括外界的事件、别人的谈话内容等信息不容易“注意”,反应较弱甚至迟钝,而异常的专心“注意”于自己的某些思维活动。而且,由于语言交流的过程往往需要在不同信息内容之间进行随意和频繁的切换,而患者的思维系统却难以进行正常的“注意”切换,所以患者跟他人进行语言交流时会出现困难和障碍,久之便会由于困难而回避这种交流。而由于对外界信息的“注意”较弱,患者也不容易“注意”到他人的表情、目光、语气、情绪等这些微弱的细节性的信息,更不会“注意”到自己的情绪表现以及这些表现引起他人的什么反应,所以患者无法形成对自己和他人的情绪和表情的正常感知和反应,长期以往,便导致患者缺乏正常的情绪表现和情感交流。也就是说,自闭症的语言交流障碍和情感缺乏这些症状,其实是由于其思维系统的“注意”指向无法进行正常切换所导致的结果。根据“注意”切换异常的不同程度,自闭症也可有程度不同的多种表现,严重时患者可将“注意”指向长期维持在某些中间信息的神经活动上而完全忽视各种外部信息,造成语言交流退化甚至最终丧失,甚至由于失去对其他事情的“注意”而失去对自己行为的抑制,产生冲动性或攻击性行为。由于自闭症是“注意”异常集中,不容易进行“注意”切换,所以,自闭症患者应该不容易被诱发脑电的“N400”电位,或者说需要更强烈或更持久的刺激才能被诱发N400电位。
从大脑“注意”指向的控制机制来看,(见“⒊⒊信息的输入传递通道”和“⒊⒋注意的控制、维持和切换”部分的内容)。这种“注意”切换的异常,可能是:⑴、外界输入通道向中脑网状结构的投射信号本身过弱,无法引发“注意”;⑵、皮质的思维通道向中脑网状结构的下行投射信号过强,从而更多更强地引发中脑网状结构“注意”控制通路的“注意”指向;⑶、中脑网状结构的各上行通道之间互相抑制的作用过强,某通道一旦得到“注意”便异常强烈地抑制其他通道的竞争信号。从自闭症的表现来看患者对外界信息不容易注意,似乎是第⑴,但申请人更偏向于推断主要还是后面两种尤其是第⑶种,理由包括:⑴、如果是外界输入通道的投射信息本身过弱,便应该只是某个特定信息通道(视觉、听觉、触觉等通道之一)过弱,而不应该是对所有的外界输入信息都不容易“注意”;⑵、患者并非无法“注意”外界信息,只是难以进行正常的切换,患者有时也会长时间地专注于某些外部信息,比如长时间玩耍一件物品或长时间专注于做某一事情,说明自闭症更主要的还是“注意”切换的异常。⑶、另外有研究发现,有些自闭症患者对某些声音很敏感,常常会觉得很吵,但对交谈却存在障碍,说明患者并非对听觉感知不足,而是能够感知但无法“注意”,无法正常处理这些声音信息,而只是把它当做噪音,(视觉可以通过“不看”来回避,但听觉无法通过“不听”来回避)。所以推测自闭症是联合皮质思维通道向中脑网状结构的下行投射信号过强,或者中脑网状结构的各上行通道之间互相抑制的作用过强,使中脑网状结构的“注意”指向过度维持在某些思维通道,而难以进行正常的“注意”切换。而且,由于大脑的思维通道也是存在多个不同分组和不同通道的,各个思维通道通过各自的下行投射通路向中脑网状结构进行投射,自闭症应该只是其中某些通道的投射过强,所以自闭症患者所专注的思维内容也是片面的,并可能因此造成某些方面的思维能力过弱,而在其“注意”指向过强的通道,由于长期在这些通道进行信息处理,可能会造成该方面的思维能力超强,以致成为畸形的偏重某一方面的天才。
自闭症这种“注意”控制机制的异常,可能大部分是遗传性的,即大脑在婴幼儿生长时便形成神经通道的结构性的发育异常。后天因素也可能是由于某些神经递质或调质异常,或者来自脑干调制性神经核团的调制作用异常,导致下行投射信号过强或互相抑制的作用过强;甚至,由于皮质的下行投射通道属于氨基酸能神经,其信号传递是具有可塑性的,所以也不排除这种“注意”异常是习得性的。由于大脑的各个神经通道普通存在交互调制的现象,所以,导致自闭症产生的原因,必然还会对其他神经活动产生影响,导致某些患者出现其他方面的脑功能异常,使自闭症显得更加复杂。在治疗自闭症时,需要注意其感知异常、交谈障碍和情感缺乏等特征症状实际上只是其思维系统的“注意”指向切换异常长期导致的结果,治疗需要针对其本质才能更有效。
⒌⒍⒊精神分裂症。神经分裂症具有多种临床表现,阳线症状、阴性症状、认知症状、焦虑症状等,医学上都归为精神分裂症,由于找不到神经病理基础,目前被认为属于功能性疾病。申请人认为,精神分裂症也是大脑调制性通路的调制作用异常引发的,而且不同的症状具有不同的调制异常。下面分析其中最典型的阳性症状的神经活动异常的本质。
前面分析到,大脑皮质的神经元活动受到来自下丘脑或/和脑干的调制信号(包括睡眠/觉醒的调制信号)的调制,在清醒状态下,感觉输入、思维活动、和运动神经输出三者都不会被抑制,感觉输入信息可以投射到思维系统产生听觉视觉等感觉,思维系统的输出也可以投射到运动系统产生机体动作或说话书写,思维系统的活动还同时投射到联合听觉皮质尤其是听觉性语言中枢产生自我感知也即产生意识,但思维系统向联合视觉皮质的投射被抑制,避免与真实的视觉输入信息互相干扰,以免造成思维和运动输出混乱,所以大脑的思维活动包括想象都不会产生图像的视觉感知。但如果在清醒状态下,思维系统向联合视觉皮质的投射没有被抑制或抑制不足,则思维过程所产生的涉及视觉的信息尤其是想象出来的幻觉,便会投射到联合视觉皮质而被感知,当成一种真实的视觉图像,并与眼睛输入的真实的外界图像进行叠加,形成复合的图像。这时患者所感知的,也即患者所“看到”的,其实是他眼睛所看到的外界图像与他自己想象出来的幻觉叠加的复合图像,而且,患者的思维系统便对这一复合图像进行反应,进行相应的思维和动作输出。而由于这种思维和动作输出是建立在真实视觉与幻觉叠加的基础上,所以从他人看来,患者的行为既有正常部分,也即对真实视觉的反应部分,包括行走、规避障碍物等行为;也有不可理喻部分,也即对幻觉的反应部分,包括胡言乱语,(实际是在跟幻觉的交谈)。
更麻烦的是,这些幻觉和因为幻觉而产生的异常思维信息,(比如一个幻觉的人,以及跟幻觉人的交流对话),同样会被当成正常的信息和事件而被记忆下来,在下一次出现相关幻觉或想到相关内容时,这些异常信息会被激活,(幻觉人再次出现,并记得之前跟对方的交流对话),再次进入患者的感知和思维活动中,(再次交流对话),从而进一步强化这些幻觉及异常信息。这样反复强化之后,这些幻觉及异常信息逐渐在患者的思维系统中占主导地位,并对患者的正常感知造成影响,患者相信幻觉更多于相信真实的感知,于是患者便更多地生活在幻觉中。当然,如果外界的视觉信息还是能够被输入,则患者一般还能够进行常规的行走等日常活动,因为视觉信息向运动皮质的投射可以不经过思维系统。
上面分析的是导致精神分裂症阳性症状的神经活动异常的本质。造成这种异常的原因是调制性神经核团对思维系统的调制作用异常所导致。而且,既然这种调制作用异常能够产生这样严重的结果,那么便还会导致其他方面的神经元活动的异常,包括注意的控制机制、情绪系统都可能受影响,并引发其他异常表现或症状。而当调制作用的异常被去除,(包括药物治疗),则思维系统向联合视觉皮质的投射又将得到正常抑制,于是幻觉不会产生,患者能够像正常人进行思维和活动。
⒌⒎神经系统与身体健康。大脑的内部信息处理系统也即下环路控制系统,控制和调节着机体各种内脏和内分泌的活动,因此也对机体的体质和健康造成直接影响。人体的体质强弱和很多非器质性疾病比如三高,以及各种内脏非器质性的功能异常,还有中医所说的体质的阴、阳、盛、虚、热、寒,等等,实质上就是内脏神经系统及内分泌系统的工作状态不同及异常的表现。由于大脑的植物神经系统不受思维系统所直接控制,所以人无法依靠意识来调整内脏的工作,只能依靠药物。其中西医是直接将药物作用于异常的器官,优点是效果直接明显,缺点是对有些疾病没有从根源入手,往往没能根除。中医倒好像是希望通过药物来调节植物神经和内分泌系统,从而来调节体质和防治疾病,问题是它说不清楚其中的理论依据,用药也是大杂烩,没弄清各种药是什么成分在如何起作用,无法形成系统的理论。
人体如同一个复杂的社会,通过细胞、组织、器官、系统构成一个复杂的机体,各个器官和系统拥有相对独立的工作而又互相调制,而且,大多通过神经系统的神经元活动来实现信息整合和协调控制,构成一个有机整体,所以,神经系统与机体的健康密切相关。对于身体保健和对功能性疾病的防治,关键还应该研究分析大脑的下环路控制系统对身体健康的影响机制,包括影响内脏和内分泌工作状态的各个信息控制环路的具体的信息传递链路,这些链路不外都包括有信息的感知输入、识别和反射、调节输出这三大环节,研究分析这些环节是哪部分神经元、如何实现感觉和识别、如何进行反射反馈、输出如何实现调节、采用什么递质工作、工作受哪些生化物质的影响,等等,便能够揭示各种疾病的本质和产生原因,以及如何防治这些功能性疾病。
人类在长期生活中,还得到一些通过自我意识来影响植物神经系统、从而用于强身益体的方法,(当然其创始者甚至目前的使用者可能也不知道这种方法的原理)。这就是中国的气功,印度的瑜伽,佛教的禅坐,还有西方的冥思。这些方法的形式不同,可实质都是一样的,是一种意识的自我抑制行为。人的下环控制环路有其天然的工作节律,在这种自然节律下工作状态是最好的。下环路虽然不会受思维系统的直接控制,但却受思维控制环路工作的影响,如果思维控制环路振荡节律快而且波动频繁,则会频繁影响到下环路的工作节律,从而对内脏和内分泌系统的工作状况造成负面影响。而意识自我抑制行为的练习者通过长时间地将意识集中在某一简单的关注点,去深度抑制大脑思维系统的神经活动,使得思维控制环路的往返发放节律变得非常缓慢而均匀,并长时间地处在这种均匀而低节律的状态,因此而减少对下环控制环路的干扰和抑制,使得下环控制环路能够在自然节律下工作,从而使内脏和内分泌系统能够工作得更好,达到增强体质的作用。这种意识的深度抑制状态类似于深度睡眠而优于普通睡眠,因为普通睡眠的过程会周期性地在各个振荡状态之间循环进行,其深度睡眠时间并不多,包括会做梦,做梦时思维系统会处在激烈活动中,使得思维控制环路的振荡节律很高且剧烈波动,从而会干扰下环控制环路的工作。而像太极、健身操等这些动作轻松而又有规律有节奏的运动,似乎也有类似的作用。
- 还没有人留言评论。精彩留言会获得点赞!