新型鱼致病病毒的制作方法
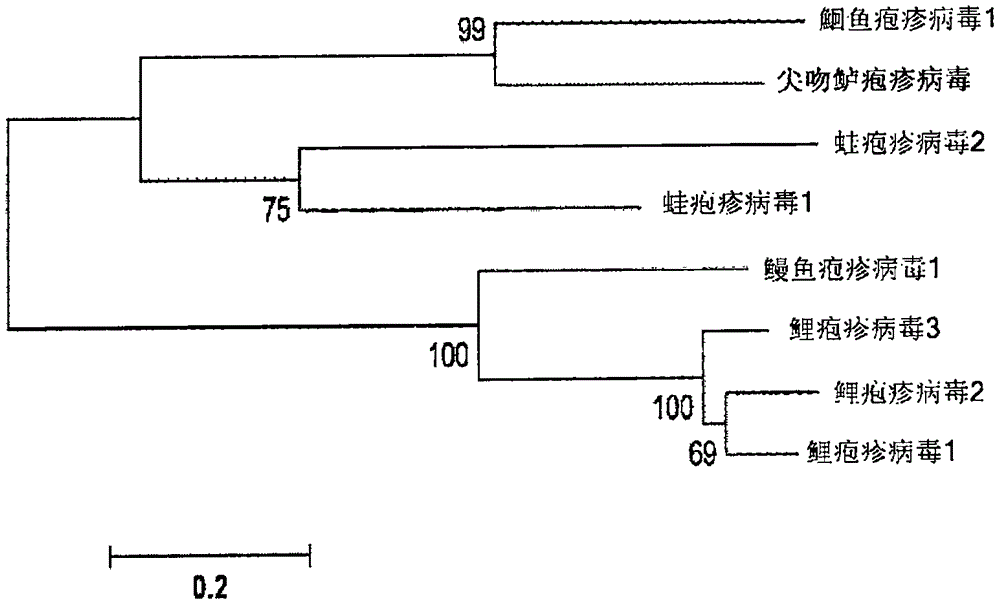
本发明涉及一种引起鱼的疾病的新型鱼致病病毒,包含所述病毒的细胞培养物、所述病毒的dna片段和相应的蛋白质,基于所述病毒、dna和/或蛋白质的疫苗以及与所述病毒反应的抗体和检测所述病毒的诊断检测试剂盒。一般背景在最近的数十年中,观察到世界范围内鱼类消费的迅猛增加。这对于冷水鱼(诸如鲑鱼、大菱鲆、大比目鱼和鳕鱼),以及热带鱼(诸如亚洲海鲈(澳洲肺鱼)、罗非鱼、遮目鱼、黄尾鱼、琥珀鱼、石斑鱼和军曹鱼)的消费,是均等的。因此,观察到养鱼场的数目和大小的增加,以便满足日益增长的市场需求。如从例如畜牧业已知的,密集地生活在一起的大量动物易发所有类型的疾病,甚至是在大规模商业畜牧时代之前几乎不曾知道或见到或甚至未知的疾病。鱼类养殖的情况同样如此。在2015年,在养鱼场(尤其是在越南和新加坡)中报道了亚洲海鲈(尖吻鲈(latescalcarifer))中一种新疾病的爆发。观察到鱼经历脱鳞病-样症状。脱鳞病(scaledropdisease)是由最近分离并在wo2014/191445中描述的虹彩病毒科的病毒引起的。然而,使用脱鳞病病毒dna特异性pcr引物测试,这些病例为脱鳞病阴性。当在田间(例如在养鱼场,因此不是在受控的实验室环境中)发现时,通常与这种新疾病相关的主要临床体征(不一定在每一条患病的鱼中看到所有这些这些症状)可被描述为如下。存在急性发作的临床体征与高百分比的发病率,其中许多鱼患病,并且在临床体征发作4-7天内死亡率高达60%(通常为30-70%)。这些临床体征可描述如下:患病的鱼具有全身性皮肤病变,变得嗜睡,并表现出明显的食欲不振。随着疾病的进展,皮肤病变变得更加严重,导致皮肤变黑及鳍和尾巴的淡白色斑块和糜烂,使鱼呈现幽灵般的外观。眼睛变得肿胀且稍微浑浊。经常发现的疾病的内部体征是脾脏和肾脏肿大。肾脏变得易碎并且易于剥离。可以经常观察到一些苍白的肝脏。随着疾病的进展,鳃变得苍白。发明目的本发明的目的是提供这种疾病的病原体。这使得能够提供旨在对抗所述疾病的检测方法和疫苗。发明概述已经发现,如上文所述的疾病的病原体是疱疹病毒和异疱疹病毒科(alloherpesviridae)的成员。它是属于双链dna病毒的二十面体病毒。该病毒与异疱疹病毒科的病毒具有一定但低水平的相似性,所述异疱疹病毒科是对鱼类或两栖动物具有致病性的一个疱疹病毒科。异疱疹病毒科的病毒具有包膜,且具有二十面体和球形到多形的几何形状。它们的直径约为150-200nm。基因组是线性且非分段的,长度为约100-250kbp。本发明的病毒具有约130kbp的基因组。图1显示系统树,其指示已知的异疱疹病毒和新发现的病毒之间的关系。所述新病毒在该树中被称为尖吻鲈疱疹病毒(latescalcariferherpesvirus,lchv)。该树是使用mega程序第5版和使用标准设置制作的。(mega5:molecularevolutionarygeneticsanalysisusingmaximumlikelihood,evolutionarydistance,andmaximumparsimonymethods.koichirotamura,danielpeterson,nicholaspeterson,glenstecher,masatoshineiandsudhirkumar.mol.biol.evol.28(10):2731-2739.2011doi.10.1093/molbev/msr121advanceaccess公开2011年5月4日)。在hanson,l等人的综述论文(viruses3:2160-2191,2011)中概述了异疱疹病毒科,更具体而言在鱼中发现的异疱疹病毒科。根据本发明的疱疹病毒在亚洲海鲈(尖吻鲈)中发现。到目前为止,尚未描述对亚洲海鲈具有致病性的疱疹病毒。不能排除该病毒对其他(亚)热带鱼类也具有致病性。在已知的异疱疹病毒科中,鮰鱼疱疹病毒1(ictaluridherpesvirus1,ihv1)是最接近的已知科成员。这种病毒对斑点叉尾鮰(channelcatfish)是致病的。然而,根据本发明的病毒与鮰鱼疱疹病毒1之间在核苷酸水平上的总体序列同一性远低于60%。与更远距离相关的鳗鱼疱疹病毒(anguilidherpesvirus,ahv)和锦鲤疱疹病毒(koiherpesvirus,khv)的序列同一性更低。表1显示了在新的疱疹病毒中鉴定的几种基因与ihv1中的同源基因之间的比较。该表显示了鉴定的最重要的orf(左栏),条件是具有少于300个碱基对的鉴定的orf在序列编号中没有被采用(为了该表的目的而是被简单忽略),在表中已经显示了dna(对于病毒的一个代表性实例,保藏在巴斯德研究所,见下文)上的位置、ihv1中的相应orf以及与这些已知orf的序列在氨基酸水平上的同一性水平。所述病毒的代表性实例已经保藏在国家微生物菌种保藏中心(collectionnationaledeculturesdemicroorganisms(cncm),巴斯德研究所,25ruedudocteurroux,f-75724pariscedex15,法国),保藏号为cncmi-5118(尖吻鲈疱疹病毒)。表1:新型疱疹病毒与ihv1之间的比较表1的说明orf:lchv中的开放阅读框(编号)框:基因组上的阅读框(1、2、3、负数指相反链)长度:以碱基对计的lchvorf的长度aa长度:blast搜索中氨基酸命中的长度ihvid.:lchvorf与相应的ihvorf之间的百分比氨基酸同一性ihv-1:鮰鱼疱疹病毒-1同源物:基于与已知蛋白质的同源性,由lhcvorf编码的蛋白质的推定功能。定义当前病毒的野生型形式是具有复制能力形式的病毒,如可以从患病鱼、特别是亚洲海鲈中分离,并且能够在分离出其野生型形式的病毒的鱼的品种的健康鱼中诱导相同的疾病。根据定义,灭活或减毒后的野生型病毒能够以其野生型形式诱导疾病。分离的病毒是这样的病毒,所述病毒从在自然界的患病宿主中通常与该病毒相关的组织中释放,并且在没有其他病毒和细菌的情况下转移到容器,诸如皿、烧瓶或生物反应器中。分离的病毒的实例是存在于生物反应器中特定细胞系的细胞培养物中的病毒。分离的dna片段是从如存在于相应的、天然存在的能够复制的生物中的其天然(完整)dna中取出的dna片段。此类dna片段可以如此存在于稳定液中,或者可以重组转移到另一种生物的dna中。在每种情况下,dna片段在本发明的意义上仍然是分离的。分离的蛋白质是从其天然环境取出的蛋白质,即脱离其与相应的天然存在的能够复制的生物的天然结合。疫苗是药物组合物,其对主体动物施用是安全的,并且能够在该动物中诱导针对致病微生物的保护性免疫,即诱导成功的预防性治疗。在这种意义上,成功的预防性治疗是这样的治疗,其帮助预防或减轻该病原体的感染或由该感染引起、由致病病原体的治疗后攻击导致的病症,特别是在减少在此类攻击后其在宿主中的负荷,并任选地帮助预防或减轻由病原体的治疗后感染引起的一种或多种临床表现。开放阅读框(orf)是具有编码蛋白质或肽的潜力的阅读框的一部分。orf是连续的一段密码子,其不含终止密码子。实施方案在第一个实施方案中,该新型病毒的特征在于分离的疱疹病毒,所述疱疹病毒是异疱疹病毒科的成员,并且其以野生型形式引起亚洲海鲈中的疾病,所述疾病特征在于以下体征(在实验室环境中用所述病毒腹膜内攻击后):攻击后3天左右出现临床体征,导致皮肤变黑及苍白色斑块和鳍糜烂的全身性皮肤病变(以显著量的病例出现,通常超过50%),嗜睡且观察到丧失游泳平衡(“观察到”意味着它可以见到,但并非在所有病例中,只发生在极度嗜睡的鱼中),(几乎完全)食欲不振,鳃盖呼吸率增加,并且在攻击后约2周出现死亡。在另一个实施方案中,所述病毒具有dna,其对应于具有根据seqidno:1的序列的dna。在实践中,这意味着该dna全长上的同一性水平超过70%,优选超过71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98%,超过99%或甚至高达100%。值得注意的是,目前认为(在130k碱基对中)在80,000位附近,所述病毒具有内部重复。基于与其他疱疹病毒的同源性,因此,所述病毒可能会以四种基因组异构体存在(如mahiet等人于以下中所解释:structuralvariabilityoftheherpessimplexvirus1genomeinvitroandinvivo;journalofvirology,2012年8月,volume86,number16,pp8592-8601)。除了bp80,000附近的重复之外,新型病毒可能具有一个或多个额外的内部重复。但是,这还没有确定。seqidno:1对应于新型病毒的可能基因组异构体之一。在病毒的又一个实施方案中,在其dna中,至少95%的开放阅读框(特别是包含至少300个碱基对的orf),例如至少96%、97%、98%、99%或100%,与具有根据seqidno:1的序列的dna的相应开放阅读框具有至少80%的序列同一性,例如至少81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99%或甚至高达100%同一性。值得注意的是,为了限定本发明的目的,用于确定同一性水平的合适程序是ncbi的基本的局部比对搜索工具(basiclocalalignmentsearchtool)的核苷酸blast程序(blastn),使用“比对2个或更多序列(aligntwoormoresequences)”选项和标准设置(http://blast.ncbi.nlm.nih.gov/blast.cgi)。在另一个实施方案中,作为异疱疹病毒科的成员,分离的疱疹病毒具有保藏在法国巴黎巴斯德研究所,国家微生物菌种保藏中心(cncm),保藏号为cncmi-5118的病毒的至少一种鉴定特征。这意味着所述病毒可以根据本发明被鉴定为的新型异疱疹病毒,即尖吻鲈疱疹病毒。在另一个实施方案中,鉴定特征选自:1)病毒具有这样的dna序列,所述dna序列在其全长上与根据seqidno:1的序列具有至少70%(或至少71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99%或100%)的同一性,且附带上述关于内部重复的条件;或2)处于其野生型形式的病毒在亚洲海鲈中引起疾病,所述疾病特征在于以下体征:攻击后3天左右出现临床体征,导致皮肤变黑及苍白色斑块和鳍糜烂的全身性皮肤病变(以显著量的病例出现,通常超过50%),嗜睡且观察到丧失游泳平衡(“观察到”意味着它可以见到,但并非在所有病例中,只发生在极度嗜睡的鱼中),(几乎完全)食欲不振,鳃盖呼吸率增加,并且在攻击后约2周出现死亡的发生;或3)病毒包含主要包膜蛋白(mep)基因,所述基因与seqidno:2中所示的核苷酸序列具有至少80%的同一性水平;或4)病毒包含dutp酶基因,所述dutp酶基因与seqidno:4中所示的核苷酸序列具有至少80%的同一性水平;或病毒包含终止酶基因,所述终止酶基因与seqidno:6中所示的核苷酸序列具有至少80%的同一性水平;或病毒包含聚合酶基因,所述聚合酶基因与seqidno:8中所示的核苷酸序列具有至少80%的同一性水平。在新型疱疹病毒(其是异疱疹病毒科的成员,且以其野生型形式在亚洲海鲈中引起疾病)的仍另一个实施方案中,所述病毒的特征在于该病毒具有dna,其对应于具有根据seqidno:1的序列的dna。这意味着病毒具有这样的dna,所述dna在其全长上与根据seqidno.1的dna具有至少70%的同一性(附带上述关于内部重复的条件)。同一性水平可以更高,例如71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99%或甚至100%。在新型疱疹病毒(其是异疱疹病毒科的成员,且以其野生型形式在亚洲海鲈中引起疾病)的又另一个实施方案中,所述病毒的特征在于具有dna,其中至少95%的开放阅读框(特别是包含至少300个碱基对的orf),例如至少96%、97%、98%、99%或100%,与具有根据seqidno:1的序列的dna的相应开放阅读框具有至少80%的序列同一性,例如至少81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99%或甚至高达100%的同一性。在其他实施方案中,基于其主要包膜蛋白(orf28)及其dutp酶(orf1)的编码dna序列,可以将新型病毒与异疱疹病毒科的已知成员区分开。事实证明,所述病毒的主要包膜蛋白与异疱疹病毒科的其他种的甚至最接近的mep仅具有30%的序列同一性水平。dutp酶与异疱疹病毒科的其他种的最接近的dutp酶仅具有45%的序列同一性水平。编码mep和dutp酶的dna序列的典型实例分别显示在seqidno:2和4中。它们各自的氨基酸序列,即由根据seqidno:2和no:4的dna片段编码的蛋白质(术语“由……编码”不排除其他dna导致相同的蛋白质,或换句话说:“由特定的dna序列编码”意指蛋白质可以基于特定序列合成,但也可能通过使用另一种序列合成”)显示于seqidno:3和5。应当理解,对于本文所涵盖的具体蛋白质,病原体的个体代表之间可存在自然变异。导致例如主要包膜蛋白序列中的微小变化的遗传变异确实存在。对于dutp酶也是如此。首先,存在所谓的“第二和第三碱基中的摇摆”,从而解释了可能发生核苷酸变化,所述变化在它们编码的氨基酸序列中保持不被注意到:例如,三联体tta、ttg、tca、tct、tcg和tcc均编码亮氨酸。此外,可以在氨基酸序列中看到根据本发明的新病毒的代表之间的微小变异。这些变异可以通过整个序列中的一个或多个氨基酸差异,或通过所述序列中的一个或多个氨基酸的缺失、取代、插入、倒置或添加来反映。例如,neurath等人在“theproteins”academicpressnewyork(1979)中已经描述了没有实质改变生物学和免疫学活性的氨基酸取代。相关氨基酸之间的氨基酸替换,或在进化中经常发生的替换尤其是ser/ala、ser/gly、asp/gly、asp/asn、ile/val(参见dayhof,m.d.,atlasofproteinsequenceandstructure,nat.biomed.res.found.,washingtond.c.,1978,vol.5,suppl.3)。其他氨基酸取代包括asp/glu、thr/ser、ala/gly、ala/thr、ser/asn、ala/val、thr/phe、ala/pro、lys/arg、leu/ile、leu/val和ala/glu。基于这些信息,lipman和pearson开发了一种用于快速和灵敏的蛋白质比较(science227,1435-1441,1985)并确定同源蛋白质之间的功能相似性的方法。本发明示例性实施方案的这种氨基酸取代,以及具有缺失和/或插入的变化都在本发明的范围内。这解释了为何例如当从根据本发明的病毒的不同代表中分离时,mep和dutp酶可具有显著低于100%的同源性水平,同时仍然代表根据本发明的病毒的mep或dutp酶。通常,作为根据本发明的mep或dutp酶的蛋白质分别与seqidno:3和seqidno:5的氨基酸序列具有至少70%的序列同一性,因此与这些序列具有70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99%或甚至100%的序列同一性。因此,涉及这些蛋白质(即mep和dutp酶)的实施方案(a-g)具体如下:a:包含主要包膜蛋白(mep)基因的分离的疱疹病毒,其特征在于该病毒是异疱疹病毒科的成员,该病毒引起亚洲海鲈中的疾病,并且mep基因的核苷酸序列与seqidno:2中所示的核苷酸序列具有至少80%(例如81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99%或100%)的同一性水平。b:包含dutp酶基因的分离的疱疹病毒,其特征在于该病毒是异疱疹病毒科的成员,该病毒引起亚洲海鲈中的疾病,并且dutp酶基因的核苷酸序列与如seqidno:4中所示的核苷酸序列具有至少80%(例如81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99%或100%)的同一性水平。c:具有mep基因和dutp酶基因两者的分离的疱疹病毒,其特征在于,mep基因的核苷酸序列与如seqidno:2中所示的核苷酸序列具有至少80%的同一性水平,并且dutp酶基因的核苷酸序列与如seqidno:4中所示的核苷酸序列具有至少80%(例如81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99%或100%)的同一性水平。d:包含主要包膜蛋白(mep)基因的分离的疱疹病毒,其特征在于该病毒是异疱疹病毒科的成员,该病毒引起亚洲海鲈中的疾病,并且mep基因在pcr反应中与如seqidno:21和22中所示的引物组反应以产生277+/-10个碱基对的pcr产物。e:包含dutp酶基因的分离的疱疹病毒,其特征在于该病毒是异疱疹病毒科的成员,该病毒引起亚洲海鲈中的疾病,并且dutp酶基因在pcr反应中与如seqidno:23和24中所示的引物组反应以产生346+/-10个碱基对的pcr产物。f:包含mep基因和dutp酶基因的分离的疱疹病毒,其特征在于mep基因在pcr反应中与如seqidno:21和22中所示的引物组反应以产生277+/-10个碱基对的pcr产物,并且dutp酶基因在pcr反应中与如seqidno:23和24中所示的引物组反应以产生346+/-10个碱基对的pcr产物。g:分离的疱疹病毒,其特征在于mep基因的核苷酸序列与如seqidno:2中所示的核苷酸序列具有至少80%(或至少81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99%或100%)的同一性水平,并且dutp酶基因的核苷酸序列与如seqidno:4中所示的核苷酸序列具有至少80%(或至少81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99%或100%)的同一性水平,并且特征在于病毒dna在pcr反应中与如seqidno:21和22中所示的引物组反应以产生277+/-10个碱基对的pcr产物,并在pcr反应中与如seqidno:23和24中所示的引物组反应以产生346+/-10个碱基对的pcr产物。实施方案d至g利用pcr-测试,所述pcr-测试利用用于根据本发明的病毒的主要包膜蛋白基因序列或dutp酶基因序列的引物组。选择两个不同的引物组,其序列描述于seqidno:21-22和seqidno:23-24。使用与病毒的主要包膜蛋白基因反应的第一引物组(seqidno:21-22)的pcr-测试使用两个引物lchvmepfw和lchvmeprev(参见实施例部分中的表2b)。使用与病毒的dutp酶基因反应第二引物组(seqidno:23-24)的pcr-测试使用两个引物lchvdutpfw和lchvdutprev(参见实施例部分中的表2c)。在实施例部分中更详细描述的测试是标准pcr测试。如果对第一引物组的pcr产物的分析显示大约277个碱基对的pcr产物,或者如果第二引物组的pcr产物的分析显示大约346个碱基对的pcr产物,并且病毒是异疱疹病毒科的成员并引起亚洲海鲈中的疾病,则这明确地证实所分析的病毒是根据本发明的病毒。出于本发明的目的,大约277个碱基对的pcr产物是具有277+10和277-10个碱基对之间的长度的pcr产物。大约346个碱基对的pcr产物是具有346+10和346-10个碱基对之间的长度的pcr产物。涉及对新型病毒特异性的其他蛋白质(即终止酶和聚合酶)的其他实施方案(h-k)具体如下:h:包含终止酶基因的分离的疱疹病毒,其特征在于该病毒是异疱疹病毒科的成员,该病毒引起亚洲海鲈中的疾病,并且终止酶基因在pcr反应中与如seqidno:25和26中所示的引物组反应以产生585+/-10个碱基对的pcr产物。i:包含聚合酶基因的分离的疱疹病毒,其特征在于该病毒是异疱疹病毒科的成员,该病毒引起亚洲海鲈中的疾病,并且聚合酶基因在pcr反应中与如seqidno:27和28中所示的引物组反应以产生314+/-10个碱基对的pcr产物。j:包含终止酶基因和聚合酶基因的分离的疱疹病毒,其特征在于终止酶基因在pcr反应中与如seqidno:25和26中所示的引物组反应以产生585+/-10个碱基对的pcr产物,并且聚合酶基因在pcr反应中与如seqidno:27和28中所示的引物组反应以产生314+/-10个碱基对的pcr产物。k:分离的疱疹病毒,其特征在于终止酶基因的核苷酸序列与如seqidno:6中所示的核苷酸序列具有至少80%(或至少81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99%或100%)的同一性水平,并且聚合酶基因的核苷酸序列与如seqidno:8中所示的核苷酸序列具有至少80%(或至少81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99%或100%)的同一性水平,并且特征在于病毒dna在pcr反应中与如seqidno:25和26中所示的引物组反应以产生585+/-10个碱基对的pcr产物,并在pcr反应中与如seqidno:27和28中所示的引物组反应以产生314+/-10个碱基对的pcr产物。实施方案h至k利用pcr-测试,所述pcr-测试利用用于根据本发明的病毒的终止酶基因序列或聚合酶基因序列的引物组。选择两个不同的引物组,其序列描述于seqidno:25-26和seqidno:27-28。使用与病毒的末端酶基因反应的第一引物组(seqidno:25-26)的pcr-测试使用两个引物lchvterfw和lchvterrev(参见实施例部分中的表2d)。使用与病毒的聚合酶基因反应第二引物组(seqidno:27-28)的pcr-测试使用两个引物lchvpolfw和lchvpolrev(参见实施例部分中的表2e)。在实施例部分中更详细描述的测试是标准pcr测试。如果对第一引物组的pcr产物的分析显示大约585个碱基对的pcr产物,或者如果第二引物组的pcr产物的分析显示大约314个碱基对的pcr产物,并且病毒是异疱疹病毒科的成员并引起亚洲海鲈中的疾病,则这明确地证实所分析的病毒是根据本发明的病毒。基于上述新型病毒的主要包膜蛋白和dutp酶的dna编码序列,还提供了本发明的以下实施方案l-o:l:包含编码主要包膜蛋白的基因的(分离的)dna片段,其特征在于所述基因与如seqidno:2中所示的mep基因的核苷酸序列具有至少80%(或至少81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99%或100%)的同一性水平,和m:由该dna片段编码的(分离的)主要包膜蛋白。n:包含编码dutp酶的基因的(分离的)dna片段,其特征在于所述基因与如seqidno:4中所示的dutp酶基因的核苷酸序列具有至少80%(或至少81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99%或100%)的同一性水平,和o:由该dna片段编码的(分离的)dutp酶。目前认为,几种其他基因也可用于鉴定根据本发明的新型病毒。这些基因之一是对应于orf224(seqidno:10)的基因,其编码膜(糖)蛋白(seqidno:11)。还另一个基因是与鮰鱼疱疹病毒1(ihv1)tk(胸苷激酶)基因同源的基因。在ihv-1中,该基因对应于orf5(hanson等,virology.1994aug1;202(2):659-64)。该新型病毒具有“脱氧核糖核苷激酶”保守结构域,其与ihv-1的orf5在氨基酸水平上具有33%的同一性。用于鉴定新型病毒的另一基因(seqidno:12)是包含orf206的基因,其编码主要衣壳蛋白(seqidno:13)。根据本发明的仍另一个实施方案涉及具有长度为至少100个核苷酸的开放阅读框的任何(分离的)dna片段,其中所述dna与具有根据seqidno:1的序列的dna的开放阅读框具有至少80%(或至少81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99%或100%)的序列同一性。尽管已发现30-40个核苷酸的长度足以区分本发明的病毒dna与任何公众已知病毒的dna,但实际相关的长度、特别是用于基于相应蛋白质的相应亚单位疫苗的实际相关长度为至少100个核苷酸(或至少150、200、250或甚至至少300个核苷酸),以对应于具有对应于病毒蛋白的相关免疫原性表位的相关和区别性3d一致性的蛋白质。因此,在另一个实施方案中,本发明还涉及由这种dna片段编码的(分离的)蛋白质。在一个实施方案中,本发明还涉及细胞培养物(即,在人工培养容器中的限制数目的类型的细胞(也称为细胞系)的人工培养物;还描述了细胞在其自然环境之外的受控条件下生长的过程),其包含能够复制形式的根据本发明的新型病毒。几种鱼细胞系可能可用于支持根据本发明的病毒的复制。可用于生长根据本发明的病毒的细胞系的实例是来自亚洲海鲈脑细胞的细胞系。分离这种细胞系的方法已尤其描述于hasoon等人的invitrocell.dev.biol.-animal47:16-25(2011)。另一个可能可用于支持病毒复制的细胞系的实例由chi等人公开:“persistentinfectionofbetanodavirusinanovelcelllinederivedfromthebraintissueofbarramundilatescalcarifer”,chisc,wuyc,chengtm,disaquatorgan.2005jun;65(2):91-8.pmid:16060261。还已经确定,使用常规方法的来自海鲈鳍的原代细胞培养物可用于支持病毒的复制。可能可用于生长病毒的其他细胞系是来自皮肤、脑、心脏、即其中病毒可能复制的器官的细胞。在又另一个实施方案中,本发明涉及用于对抗鱼中的疱疹病毒疾病的疫苗,其中所述疫苗包含根据本发明的疱疹病毒、或如上文所述的免疫原性蛋白质和药学上可接受的载体。这种载体可以像水一样简单,只要它适合于以临床相关量施用材料而不引起不可接受的副作用。典型的载体是油和水的乳液、不溶性佐剂(通常是铝盐或其他盐、或大的免疫刺激聚合物分子)在水中的悬浮液,或可溶性佐剂(如皂苷、pamp、卡波姆或其他免疫刺激分子)的溶液。通常,载体包括本领域公知的稳定剂和防腐剂。在进一步的实施方案中,疫苗包含以活减毒(即能够复制,但不再能够诱导如由起源野生型病原体所诱导的全套症状)或灭活形式的根据本发明的疱疹病毒。已经用不同的技术设计单纯疱疹病毒的实验疫苗。疫苗由以下组成:肽、(重组)病毒蛋白、病毒蛋白的混合物、完整和分级的杀死的病毒(参见例如yasumoto,s.等人于fishpathology41:141-145(2006),其描述了包含捕获在脂质体区室内的灭活的完整锦鲤疱疹病毒)、复制缺陷型病毒和减毒复制型病毒(如由koelle,d.m.和l.corey.2003:recentprogressinherpessimplexvirusimmunobiologyandvaccineresearch.clinmicrobiolrev.16(1):96-113所概述)。每种方法都有特定的优点和缺点,已由stanberry在2000年讨论过(stanberry,l.r.,a.l.cunningham,a.mindel,l.l.scott,s.l.spruance,f.y.aoki和c.j.lacey.2000:prospectsforcontrolofherpessimplexvirusdiseasethroughimmunization.clin.infect.dis.30:549-566.)。已知毒性疱疹病毒毒株在细胞培养物中的体外连续传代导致产生减毒后代,或者换句话说,产生引起保护性免疫应答而不引起疾病临床症状的非毒性复制型毒株。例如,对于马立克氏病病毒(mdv)通过体外连续传代毒性病毒直至所得分离物变得无毒实现了减毒,并获得名为rispens或cvi988的保护性疫苗(rispensbh,vlotenh,mastenbroekn,maashj,schatka.1972:controlofmarek′sdiseaseinthenetherlands.1.isolationofanavirulentmarek′sdiseasevirus(straincvi988)anditsuseinlaboratoryvaccinationtrials.aviandis.16:108-125)。相应地,描述了通常在涉及dna复制和转录调节的途径中的多个基因参与mdv的重新减毒,并为未来md和因此相应的疱疹病毒疫苗的合理设计提供靶标。还已经描述了鱼疱疹病毒通过连续传代来减毒(尤其参见noga,e.j.等人,can.j.fish.aquat.sci.38:925-929,1981)。如通常已知的,病毒减毒可以是自发的,或者可以通过药物诱导(诱变或其他类型,诸如例如uv光;参见例如mutationresearch768,2016,53-67andj.gen.virol,1985,66,2271-2277)。减毒背后的潜在遗传机制通常仍然知之甚少,但病毒基因组中的遗传变化(突变、缺失等)和/或其积累是疱疹病毒毒株减毒的基础。突变可能涉及多种病毒机制,包括复制能力、病毒传播等。对于病毒复制和体内感染性必需的某些分子途径可能对于复制培养不是必需的,并且参与此类途径的那些基因可能更倾向于在长期培养中发生遗传改变。如果表征基因组中的这种随机突变和缺失,并且随着下一代测序技术的发展,对大病毒诸如疱疹病毒的整个基因组进行测序变得相对简单,这使得能够合理地设计减毒。从关于疱疹病毒减毒的文献中,多个基因已作为可能的靶标出现,其中基因功能障碍导致病毒的功能性减毒。roizman和knipe在2001年给出了可能已在活减毒单纯疱疹疫苗中发生突变的参与病毒复制的基因的详细概述(roizman,b.,和d.m.knipe:herpessimplexvirusesandtheirreplication,p.2399-2459.于d.m.knipe,p.m.howley,和d.e.griffin(编辑),fieldsvirology,4thed.,vol.2.lippincott,philadelphia,pa)。此外,这种突变可用于建立具有不连续复制病毒的疫苗方法。突变的病毒在基因工程化细胞系中生长,该细胞系以反式提供所需的非突变基因。例如,当缺失编码gh的晚期基因ul22的单纯疱疹病毒感染非互补细胞时,后代病毒粒子可以离开细胞但不能感染次级细胞(koelle和corey,2003,如上所引用)。下面是疱疹病毒基因的列表,已经描述了所述基因功能障碍或缺失导致锦鲤疱疹病毒、异疱疹病毒或其他疱疹病毒的功能性减毒。这些基因是用于根据本发明的疱疹病毒的减毒的靶基因:1)锦鲤疱疹病毒的胸苷激酶(tk)基因。还已经在斑点叉尾鮰疱疹病毒(ccv)中描述了tk基因,ccv是与本发明中描述的病毒相对密切相关的病毒(hansonla,kousoulaskg,andrlthune.1994:channelcatfishherpesvirus(ccv)encodesafunctionalthymidinekinasegene:elucidationofapointmutationthatconfersresistancetoara-t.virology202(2):659-64)。2)锦鲤疱疹病毒的d-utp酶基因。3)编码锦鲤疱疹病毒orf57的基因,如genbank登录号nc_009127中给出,其中orf57起始和终止密码子位于位置99382和100803(boutierm,ronsmansm,ouyangp,fournierg,reschnera,等人(2015):rationaldevelopmentofanattenuatedrecombinantcyprinidherpesvirus3vaccineusingprokaryoticmutagenesisandinvivobioluminescentimaging.plospathog11(2):e1004690)。4)如在牛疱疹病毒、马疱疹病毒和伪狂犬病毒(伪狂犬病)中发现的gd(ehvenbhv)/gp50(prv)基因。该基因编码可以针对其产生中和抗体的糖蛋白。本发明还涉及用如上所述的疫苗预防性治疗动物(即用于治疗动物以防止被相应的野生型病原体治疗后感染)的方法,其包括将疫苗全身性施用于动物。全身性施用疫苗是指施用所述疫苗,使得它到达身体的循环系统(其包含心血管和淋巴系统),从而影响整个身体而不是特定部位诸如胃肠道。全身性施用可以如下进行:例如将抗原施用进肌肉组织中(肌肉内)、真皮中(真皮内)、皮肤下面(皮下)、粘膜下面(粘膜下)、静脉中(静脉内)、体腔内(腹膜内)等。本发明还体现在与根据本发明的病毒反应的抗体或抗血清,以及用于检测与根据本发明的病毒或其抗原物质反应的抗体的诊断测试试剂盒,其中所述测试试剂盒包含根据本发明的病毒或其抗原物质。本发明还体现在用于检测根据本发明的疱疹病毒或其抗原物质的诊断测试试剂盒,其中所述测试试剂盒包含与根据本发明的病毒或其抗原物质反应的抗体或如上所述的pcr引物组。现将使用以下实施例进一步解释本发明。实施例实施例1:发现尖吻鲈疱疹病毒收集血清和组织样品用以分离感染因子在新加坡的亚洲海鲈(尖吻鲈)养鱼场观察到患病鱼。患病鱼出现的临床症状对于渔民而言似乎与脱鳞病相似(gibson-kueh等人,jfishdis.2012jan;35(1):19-27.doi:10.1111/j.1365-2761.2011.01319.x.pmid:22103767;degroof等人,plospathog.2015aug7;11(8):e1005074.doi:10.1371/journal.ppat.1005074.pmid:26252390)。然而,当更密切地研究疾病症状时,看起来患病鱼显示出更加急性的感染(3-10天而不是超过15天)且具有更高的发病率。与脱鳞病相比,皮肤病变严重性较低,但更加全身性的。皮肤损伤与脱鳞病损伤不同之处在于整条鱼变得颜色更深且更迟钝,并具有苍白色粘液的斑块。观察到一些鳞片丧失,但这并不显著,也不是主要的临床体征。这不同于由脱鳞病病毒引起的鳞片丧失,其具有局部性、斑片状的损伤,所述损伤更严重地受到坏死的影响并具有严重的鳞片丧失。脱鳞病病毒通常也会引起较为慢性的爆发。基于观察到的差异以及脱鳞病病毒pcr给出阴性结果的事实,基于对受影响鱼场的临床观察怀疑存在不同的感染性病毒因子。决定进行目的在于病毒分离的后续研究,并发现一种新的病毒。除了鳞片丧失之外,对患病鱼的其他观察结果是嗜睡、严重食欲不振、眼睛混浊/肿胀和高死亡率(高达30-70%的患病鱼)的急性发作。从患病鱼中取样(血清、肾、脾)。将合并的血清样品在-70℃保存,直至进一步分析。将合并的肾脏样品保持在+4℃直至第二天匀浆。组织样品的组织匀浆用于分离感染因子通过在svdb(标准疫苗稀释缓冲液=pbs)中使用匀浆棒手动研磨将肾样品匀浆,并且最后将样品在svdb中以1∶9(w/v)稀释,且然后在庆大霉素中预处理1小时。将匀浆的样品在5,500rpm、+4℃下离心10分钟,且随后收集无细胞上清液。在接种单层的前一天,将海鲈脑(sbb)细胞以2×104个细胞/cm2接种于t75烧瓶中的emem+10%fbs+庆大霉素+两性霉素中。对于该实验,细胞在培养基中生长,所述培养基中还加入hepes和碳酸氢钠,并优化细胞以在没有co2的这些条件下生长。第二天,将培养基(15ml)更换为新鲜培养基,向所述培养基中加入0.1ml或0.3ml未稀释的无细胞上清液。将烧瓶在28℃温育。3天后在其中加入未稀释的无细胞上清液(0.1和0.3ml二者)的烧瓶中观察到cpe,并在第5天收获(第1代)。将假定的感染因子命名为v511。根据上述方案,将引起cpe的因子在sbb细胞上再传代3次。引起cpe的因子被命名为v511/sbb_4p。对患病鱼的细胞和血清中引起cpe的因子的培养上清液的病毒检测使用由devries等人(2011)plosone6(1):e16118描述的vidisca-454技术分析患病鱼的血清样品和引起cpe的因子的第4代收获物v511/sbb_4p的培养物上清液。在两种类型的样品中,都获得了怀疑源自新型鱼病原体的序列。将这些序列用于衍生用于常规pcr和定量pcr的pcr引物(参见表2a-e)。对新序列进行blast表明,培养上清液中的引起cpe的因子和血清中怀疑的感染因子显示出与异疱疹病毒科的病毒的一定水平的同源性。这种新病毒因此被称为尖吻鲈疱疹病毒(lchv)。全基因组测序将病毒培养上清液样品以10,000×g离心10分钟,并如所述用turbodnase(thermofisher)处理(devriesm,等人plosone.2011;6(1):e16118.doi.10.1371/journal.pone.0016118),之后通过boom提取法提取核酸(boomr,等人jclinmicrobiol.1990;28(3):495-503)。使用dsdna片断化酶(newenglandbiolabs)剪切样品。剪切的样品用ampurexp珠粒(agencourtampurexppcr,beckmancoulter)以1∶1.8的比例(样品∶珠粒)纯化以除去酶。纯化后,用dna聚合酶i,大(klenow)片段(newenglandbiolabs)将样品进行末端修复。用ampurexp珠粒(agencourtampurexppcr,beckmancoulter)以1∶1.8的比例(样品∶珠粒)纯化末端修复的样品以除去酶,然后使用klenow片段(3’□5’exo-)(newenglandbiolabs)将样品加a尾。用ampurexp珠粒(agencourtampurexppcr,beckmancoulter)以1:1.8的比例(样品:珠粒)纯化样品以除去聚合酶。通过使用t4连接酶(thermofisher),将来自nebnextmultiplexoligosforillumina(newenglandbiolabs)的泡状衔接子(bubbleadaptors)连接到加a尾的样品上。通过使用ampurexp珠粒(agencourtampurexppcr,beckmancoulter)进行大小选择,其中首先以1∶0.5的比例(样品∶珠粒),以确保大多数大小超过400bp的片段被去除,然后通过将另外的ampurexp珠粒(agencourtampurexppcr,beckmancoulter)添加至上清液达到1∶0.85(样品∶珠粒)的最终比例,以结合200-400bp之间的dna片段并去除小于200bp的片段。在大小选择后,通过使用来自nebnextmultiplexoligosforillumina(newenglandbiolabs)的user酶打开泡状衔接子。接下来,用来自nebnextmultiplexoligosforillumina(newenglandbiolabs)的衔接子特异性引物和q5热启动主混合物(newenglandbiolabs)进行28个循环的pcr;30秒98℃,和10秒98℃和75秒65℃的循环,然后为5分钟65℃。pcr后,通过使用ampurexp珠粒(agencourtampurexppcr,beckmancoulter),以1∶0.5的比例(样品∶珠粒),对样品进行大小选择,以去除大小超过400bp的片段,并向上清液中添加另外的ampurexp珠粒(agencourtampurexppcr,beckmancoulter)以达到1∶0.85的最终比例(样品∶珠粒)以结合200-400bp之间的dna片段并除去小于200bp的片段。接下来,通过qubitdsdnahs测定试剂盒(thermofisher)测量dna浓度,用高灵敏度dna分析试剂盒在生物分析仪上检查大小。将dna稀释至2.49ng/μl的浓度。通过使用配对末端测序和v2试剂盒(illumina),使用miseq(illumina)对它们进行测序。系统进化分析初始系统进化分析基于在疾病暴发样本中发现的尖吻鲈疱疹病毒的208bp的dna片段(seqidno:14)。该片段显示在翻译的核苷酸水平上与鮰鱼疱疹病毒1np_041153.2和其他鱼病毒的orf62的同源性。使用blast基本的局部比对搜索工具(http://blast.ncbi.nlm.nih.gov/blast.cgi)和多序列比对工具clustalw生成核苷酸和蛋白质序列比对。用mega5软件使用邻接法创建系统进化树,在缺口或插入的情况下具有部分删除(mega5:molecularevolutionarygeneticsanalysisusingmaximumlikelihood,evolutionarydistance,andmaximumparsimonymethods.koichirotamura,danielpeterson,nicholaspeterson,glenstecher.masatoshineiandsudhirkumar.mol.biol.evol.201128(10):2731-2739.2011doi:10.1093/molbev/msr121)。该系统进化分析的结果如图1所示,其描绘了基于lchv的208bpdna片段与ihv1orf62的同源性分析的lchv的系统进化树。系统进化分析证实lchv是异疱疹病毒科中的新发现的病毒。实施例2:使用pcr、qpcr分析检测尖吻鲈疱疹病毒引物设计在疾病暴发样本中发现的尖吻鲈疱疹病毒的208bp的dna片段上设计pcr引物。该片段显示在翻译的核苷酸水平上与鮰鱼疱疹病毒1np_041153.2的orf62的同源性。还在mep、dutp酶、终止酶和聚合酶基因上设计引物。表2a:在尖吻鲈疱疹病毒的208bpdna片段上设计的引物表2b:在尖吻鲈疱疹病毒的mep上设计的引物表2c:在尖吻鲈疱疹病毒dutp酶上设计的引物表2d:在尖吻鲈疱疹病毒终止酶上设计的引物表2e:在尖吻鲈疱疹病毒聚合酶上设计的引物pcr和凝胶电泳使用veriti96孔热循环仪(appliedbiosystems)进行常规pcr。制备包含1xsupertaq缓冲液、0.02u/μlsupertaq酶、0.2mm脱氧核糖核苷三磷酸(dntp)、1μm正向和1μm反向引物的主混合物。对于每个样品,在48μlpcr混合物中加入2.0μldna模板,将2μl无菌水用作阴性对照。设计pcr程序,以95℃初始化60秒开始,然后分别在95℃、基于引物的tm的引物组特异性退火温度和72℃下进行变性、退火和延伸,重复40次。该程序以72℃下最终延伸10分钟结束。将样品与1x溴化乙锭一起上样到1.5%琼脂糖凝胶和1xtae缓冲液中,以115伏持续60分钟。qpcr分析定量聚合酶链反应使用bioradcfx96系统进行。使用probefastmastermix和序列特异性探针进行qpcr实验。每个反应存在18μl主混合物,其含有1xprobefastq-pcrmastermix(kapa)、200nm正向引物、200nm反向引物和200nm探针。引物对2用于qpcr分析,且优化的tm为60.7℃。探针dna序列是cgcgggatgacctcttctcg(seqidno:31),使用的标记是5′6fam和3′tamra。向18μl主混合物中加入2μldna模板。每个反应一式两份进行,并且所有板在2,200×g下离心4分钟,然后插入cfx系统。使用如下程序进行扩增,以95.0℃持续3分钟开始,然后在95.0℃下持续3秒和在60.7℃下持续30秒,重复40次。标准线将含有具有尖吻鲈疱疹病毒-同一性-序列构建体的puc57载体(通过genscript合成[合成如上所述的208bp片段并克隆在质粒载体中])的稀释系列用作阳性对照、用于效率和准确度以及用于样品定量的指示物。将载体溶解并稀释于水中,范围为每个qpcr反应1.0×101拷贝/2μl至1.0×109拷贝/2μl。稀释系列包括在所有qpcr实验中并储存在-20℃。通过支持软件(cfx-manager版本3.1),完成样品中的尖吻鲈疱疹病毒dna的定量,该软件使用来自稀释系列的获得数据来创建标准线。进行的所有qpcr实验显示效率在98.0%和100.2%之间,且准确度在0.997和0.999之间,强调了设计用于定量lchvdna的qpcr方法的稳健性。实施例3:亚洲海鲈中新型感染因子的实验感染亚洲海鲈(尖吻鲈)中v511/sbb_4p的实验感染在该实验中,用v511进行鱼的腹膜内和共栖感染,以研究v511是否是田间疾病爆发的原因。感染后在不同时间点对不同器官取样以确定感染过程。表3:处理组和水槽分配水槽组剂量(ip)鱼数目1av511/sbb_4p-未掺入0.3ml251b共栖-25一组25条亚洲海鲈通过腹膜内(ip)注射0.3ml在sbb细胞系v511/sbb_4p上生长的未稀释的v511无细胞上清液(cfs)进行攻击。csf的病毒滴度为3.2×106tcid50/ml(滴定方案见下文)。第二组25条鱼在由网隔开的同一个水槽中共栖(表3)。该网允许两个水槽半部之间的水自由移动并允许紧密接近,但不允许两个处理组的鱼之间直接接触。在实验开始时,鱼的平均重量为18克。在攻击前将鱼饥饿约24小时,以确保排空胃肠道以降低受伤风险。在攻击前按照标准程序用aqui-s镇静对鱼进行麻醉。将ip攻击组中的鱼网捕,镇静并在腹鳍的基部和尖端之间的中线进行ip注射。攻击后,将鱼放入指定的水槽中恢复和观察。未感染的共栖鱼被引入通过网隔开的相邻的水槽分区。记录死亡率和临床体征。在攻击后第4天、第7天、第11天、第14天和第18天,将共栖组中的3条鱼随机取样并进行尸检以收集鱼组织用于进一步测试。取样包括脾、心、脑、血清、肝脏、肠、鳃、皮肤和肾脏,以便更好地了解新因子通过自然途径感染后的感染过程。在攻击后第4天、第7天、第11天和第14天,随机取样ip感染组中的3条鱼并进行尸检以收集肾组织用于进一步测试。在第17天,从3条实验感染的鱼中获取脾、心、脑、血清、肝脏、肠、鳃、皮肤和肾脏样品。在攻击后第18天,所有鱼进行取样或已死于感染(见表6)。样品收集总结在表4和表5中。表4:实验ip感染组中鱼组织的取样ip感染后的天数ip感染。合并的器官(3条鱼/合并物)4、7、11、14肾脏17肾脏、脾、心脑、血清、肝脏、肠、鳃、皮肤表5:共栖组中鱼组织的取样表6:ip和共栖攻击处理组中观察到的临床体征表6中提供的结果显示ip和共栖攻击途径均能够产生与在标志亚洲海鲈鱼场观察到的那些相似的临床体征。观察到的总体临床体征是嗜睡、食欲不振、皮肤、鳍和眼部病变,其在标志鱼场的患病养殖鱼类中也类似地观察到。这表明在受控的实验室环境中(当与田间环境如养鱼场相比时,其中不存在其他病原体且存在较少的应激物),(ip)攻击后的典型症状是1)攻击后3天左右临床体征发作,2)全身皮肤病变,其可能导致皮肤变黑并伴有苍白色斑块和鳍糜烂,3)由于极度嗜睡而失去游泳平衡,4)几乎完全食欲不振,5)鳃盖呼吸率增加,和6)攻击后约2周出现死亡的发生。在一些鱼中可观察到眼睛肿胀和混浊。死亡率和百分比累积死亡率记录显示于表7。表7:每日死亡率和百分比累积死亡率记录**不包括取样用于收集组织的鱼。v511/sbb_4p病毒上清液从ip攻击的鱼传播至原初共栖鱼。两组都经历类似的临床症状。临床症状也与标志鱼场(初次爆发)观察到的那些相似。与共栖组相比,ip感染组经历更急性和严重的疾病,其中感染后3天内出现疾病体征。相比之下,共栖攻击的鱼在感染后第7天显示出初始临床体征。使用来源自在田间疾病调查期间收集的组织样本的传染性材料,以及随后的体外传代物,我们能够通过ip和共栖感染途径复制在爆发期间观察到的临床症状。还证实了疾病从ip感染的鱼传播到同一水域中的共栖的原初鱼,确认了这种病原体的传染性。实施例4:感染实验(实施例3)期间收集的组织样品的样品制备、组织匀浆、dna分离器官样品的匀浆使用precellys24匀浆器仪器,将在上述实验中收集的鱼器官样品(表4、5)匀浆。使用存在6,500rpm20秒和间隔10秒的两个循环的程序,制备磷酸盐缓冲盐水(pbs)中10%器官匀浆。心、脾、肾脏、脑、肠和肝脏样品的匀浆在一个循环中完成,将皮肤和鳃样品匀浆两次。在匀浆过程中将所有样品保持在冰上并储存在-80℃。dna提取使用magnapure96系统以及magnapure96dna和viralna试剂盒进行dna提取。为了提取,将250μlmagnapure96外部裂解缓冲液加入到200μl样品中。用预先安装的外部裂解方案分离dna,并在50μlmilli-q水中洗脱。dna在-20℃保存,直至进一步使用。实施例5:病毒培养和病毒滴定海鲈脑(sbb)细胞系的建立和培养:细胞系sbb最初在intervetnorbiosingaporepteltd(msdah的一部分)来源自亚洲海鲈脑细胞的胰蛋白酶消化的悬浮液。已经由hasoon等人在invitrocell.dev.biol.-animal47:16-25(2011)和由chi等人,disaquatorgan.2005jun;65(2):91-8描述了海鲈脑细胞的衍生程序。sbb培养基由899ml补充有2mml-谷氨酰胺和110mg/l丙酮酸钠的e-mem、100mlfcs(10%)和(任选)1ml新霉素多粘菌素抗生素溶液1000x储液组成。细胞通常在28℃和5%co2下生长。在培养启动之前将培养基保持在4℃。使用一安瓿冷冻储液sbb开始培养。通过在28℃的水中温热安瓿来快速解冻来自液氮的细胞。将细胞悬浮液加入管中并用9体积的培养基缓慢稀释。随后,计数细胞。将悬浮液分配到合适的培养瓶或滚瓶中,并在28℃和5%co2下孵育。烧瓶或滚瓶中的接种密度约为3×104个细胞/cm2。在6-24小时或细胞完全附着后,更新培养基以除去剩余的dmso(冷冻培养基由90%培养基和10%dmso组成)。将细胞进一步孵育3-7天或直至达到汇合。对于滚瓶,需要0.2-0.5rpm的辊转速。滚瓶可具有480、960和1750cm2的不同表面积。一旦达到汇合,将细胞进行传代。可以每3-4天进行一次传代,且初始接种密度为3.0×104个细胞/cm2。或者,当以1.0×104个细胞/cm2的密度铺板时,可以每7天进行一次传代。将用于细胞传代的试剂(培养基、pbs、胰蛋白酶/edta)预热至28℃。弃去培养基,并用适当体积的pbs(3ml用于t25烧瓶)洗涤汇合的单层一次。随后弃去pbs,并将细胞在相同体积的pbs中在28℃下孵育15分钟,所述pbs补充有1%(vol/vol)的2.5%胰蛋白酶溶液和1%(vol/vol)的2%edta溶液。脱离后,加入相同体积的新鲜培养基,并重悬细胞并计数。在适合培养瓶或滚瓶的培养体积中,以所需的细胞密度设置新的烧瓶。对于冷冻细胞,在程序之前,将培养基和2x浓缩的冷冻培养基(80%(vol/vo1)培养基加20%(vol/vol)dmso)保持在4℃。如上所述处理汇合细胞培养物直至并包括胰蛋白酶消化。将细胞重悬,计数,进一步重悬于适量的培养基中,并逐滴加入等体积的2×冷冻培养基,同时涡旋悬浮液。用于液氮储存的安瓿用每安瓿5.25×106个细胞填充以启动t175或用2.25×106个细胞填充以启动t75。用尖吻鲈疱疹病毒接种sbb细胞在建立接种实验之前,将从液氮储存培养的细胞传代至少一次。在以3.0×104个细胞/cm2接种在组织培养瓶中之前,将细胞传代并培养24小时。接种物由来自病毒先前传代物的新鲜或冻融培养物的未稀释上清液组成。从烧瓶中取出培养基。将烧瓶随后在28℃下接种至少60分钟。当在培养基中用lchv病毒的先前传代物的收获物接种细胞时,优选使用每细胞0.001-0.01tcid50的moi。除去接种物后(60分钟后,尽管这不是绝对要求),加入新鲜培养基并培养细胞直至使用倒置光学显微镜观察到完全cpe(通常在2-4天后)。通过收集培养上清液收获病毒,将所述培养上清液以800×g离心5分钟以除去碎片。或者,可以通过过滤使上清液澄清。将澄清的上清液用于随后的传代或pcr/dna/em分析或在-70℃冷冻。可以使用(定量)pcr分析和/或收获物滴定来确认病毒的复制。将dna测序技术用于确认病毒的特征以及em。使用magnapure96系统和magnapure96dna和viralna试剂盒(实施例4)从组织培养基中分离用于(定量)pcr的dna。在sbb细胞上滴定病毒如上所述培养sbb细胞。在测试前一天,制备在培养基(emem+10%fcs+l-glu+napyr)中含有6.0×104个细胞/ml的sbb细胞悬浮液。用100μl该细胞悬浮液接种微量滴定板的96个孔。将板在28℃和5%co2下孵育24小时。在该孵育期后,单层为约50%汇合。在测试当天,通过以下制备每个病毒样品的直至10-7的10倍系列稀释液:将0.5ml样品转移到含有4.5ml冰冷(0-20℃)滴定培养基(具有减少的fcs的培养基;50%emem+l-glu+napyr+50%培养基)的管中,混合并转移0.5ml在含有4.5ml滴定培养基的下一个管中,然后仔细混合,转移等。第1列和第12列以及a和h行用作阴性对照,并用100μl/孔新鲜滴定培养基接种。用100μl/孔的病毒稀释液(10-2、10-3、10-4、10-5、10-6、10-7)在b至g行(10个孔/稀释液)接种微量滴定板。在处理期间,病毒稀释液的温度保持在0℃至20℃之间。将板在28℃和5%co2下孵育6天。在6天病毒孵育期后,用倒置光学显微镜筛选平板的lchv特异性cpe。cpe的特征在于细胞的聚拢,然后是细胞脱离/裂解(图2)。显示lchv特异性cpe的每个孔被评为阳性。tcid5o根据由以下描述的方法和计算来测定:reed和muench,am.j.epidemiol.(1938)27(3):493-497。在滴定测定中从阳性孔分离的dna样品的qpcr分析证实了病毒的存在和复制。结果将组织培养瓶以3.0x104个细胞/cm2接种并培养24小时。24小时后,在胰蛋白酶消化后计数一个烧瓶中的细胞以确定烧瓶中存在的实际细胞数。从随后接种的其他烧瓶中取出培养基。将由来自lchv病毒在培养基中的先前传代物(病毒的传代数在4-8之间)的未稀释的培养上清液组成的0.001tcid5o/细胞的接种物施加至单层,并孵育60分钟。除去接种物,并将新鲜培养基加入细胞培养瓶中。培养细胞直至使用倒置光学显微镜观察到完全cpe。通过收集培养上清液收获病毒,将所述培养上清液以800×g离心5分钟以除去碎片。样品取自(1)用于感染单层的未稀释的接种物、(2)在用新鲜培养基替换接种物后1小时收获的培养瓶的培养上清液、(3)50%cpe的单层和(4)100%cpe的单层。对样品(1)、(3)和(4)进行滴定,并对样品(1)、(2)、(3)和(4)进行qpcr分析。结果示于表8中。使用olympusckx41倒置光学显微镜以40x放大率拍摄照片。这些显示于图2中。图2a显示90%汇合的sbb细胞(p18)的形态和图2b显示在培养瓶中50%cpe的sbb细胞(p18)的形态。表8:在sbb细胞上lchv生长的详细结果*na:未分析实施例6:电子显微镜术电子显微镜术将带有纯碳膜的400目铜网暴露于空气中的辉光放电20秒,从而使膜表面变得亲水。将病毒样品置于碳包覆的网格上,体积为10μl,并放置孵育约2分钟。用滤纸吸去过量的样品,并将10μl水置于网格上并通过吸干立即再次除去。然后将10μl的1%乙酸铀酰放置在网格上用于染色。30秒后,通过吸干除去过量的染料,并将样本放置以干燥几分钟,然后在电子显微镜下观察。在以80kv运行的jeol1011透射电子显微镜中观察样本。使用sisveleta2kx2k照相机记录图像。使用jeol1011透射电子显微镜捕获lchv培养物样品的照片,以确认鉴定的病毒是疱疹病毒并排除任何其他病毒的存在。以0.01的moi用lchv第5代接种sbb(p9)细胞。收获后,将病毒在-80℃的培养基中保存,并得到滴度为104.43tcid5o/ml。制备1μl样品用于电子显微镜术,其中两张图片显示在图3中。在图a中,可观察到两个近似直径为100nm的暗点,其与普通疱疹病毒衣壳的平均直径(115-130nm)相匹配。图b显示了放大图,其中可以清楚地观察到该颗粒的二十面体轮廓。样品中没有发现包膜的病毒。图3显示了用电子显微镜捕获的lchv病毒。在图3a中,两种疱疹病毒(暗点)是可识别的。比例尺为500nm。图3b显示了所述斑点之一的放大图,其清楚地显示了二十面体轮廓。上述实施例描述了从患病鱼中检测和分离新病原体。在用分离的病原体实验感染健康鱼后,可以再现相同的疾病症状。从实验感染的鱼中分离的感染因子与最初分离的病原体相同。这证明上述疾病症状仅仅归因于发现的病原体,即尖吻鲈疱疹病毒。实施例7:原代细胞培养物海鲈鳍细胞建立了来自海鲈鳍细胞(sbf)的原代细胞培养物。将细胞在培养物中培养至少5代。培养物建立如下。将鱼麻醉。修剪尾鳍(尾部)并在pbs+庆大霉素0.3%+恩诺沙星0.002%+两性霉素0.5%中洗涤三次。使用手术刀片将鳍切成组织碎片。将碎片转移到含有l15培养基的25cm2组织培养瓶中,所述l15培养基补充有20%fcs和庆大霉素0.3%+恩诺沙星0.002%+两性霉素0.5%。将烧瓶在28℃下于加湿的培养箱(无co2)中孵育。基于存在碎片和ph,根据需要更换培养基(l15)。在pbs中用0.125%胰蛋白酶通过胰蛋白酶消化传代细胞直至细胞脱离,并且取决于细胞密度,以1∶1-3的低比例分开。在初始传代期间,将细胞在补充有20%fcs的l15培养基中培养。在随后的传代中,fcs百分比降低至10%。使用如实施例5中所述的程序进行尖吻鲈疱疹病毒的接种。感染后第4天观察到100%cpe。实施例8:新型病毒的额外毒株病鱼样品来自不同于标志鱼场的鱼场,在所述鱼场中观察到尖吻鲈疱疹病毒感染的临床症状。然而,使用实施例2中描述的引物组2(seqidno:17和seqidno:18)的pcr分析产生了阴性pcr结果。作为替代方案,将基于聚合酶(orf21)和终止酶(orf37)序列(与其他异疱疹病毒科相比具有相对高水平的氨基酸保守性的orf)设计的引物组用于pcr。用于pcr的引物呈现于表2d和2e中。这些pcr产生阳性结果。对终止酶pcr片段进行测序,并显示与seqidno:1中所示的尖吻鲈疱疹病毒的序列97%的同一性。seqidno:29呈现了如seqidno:1中所示的lchv的终止酶pcr的pcr产物。seqidno:30显示了从不同于标志鱼场的鱼场获得的lchv的终止酶pcr的pcr产物。结论是疾病的爆发是由lchv的另一毒株引起。pct仅由受理局使用仅由国际局使用当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1