采用分选酶的单步骤双多肽缀合酶促一锅反应的制作方法
本文报道用于通过肽键同时酶促缀合三个化合物的方法。
背景技术:
分选酶a(srta)是将蛋白质共价附着至细菌细胞壁的膜结合酶。srta底物上的特异性识别基序是lpxtg,由此该酶在残基苏氨酸和甘氨酸之间切割。肽聚糖上的识别基序是五甘氨酸基序。已显示游离n端上的三甘氨酸和甚至二甘氨酸基序足以支持srta反应(clancy,k.w.等,peptidescience94(2010)385-396)。反应通过硫酯酰基-酶中间体进行,该中间体通过来自寡聚甘氨酸的胺亲核体的攻击而分解,将蛋白质底物共价连接至肽聚糖,并再生srta。srta可以用于将化学合成的肽共价缀合至重组表达的蛋白质。
来自金黄色葡萄球菌(staphylococcusaureus)的分选酶a是含有巯基的转肽酶,其识别多种结构上不相关底物中的lpxtg基序,且耐受寡聚甘氨酸亲核体的c端延伸,允许将多样的功能化亲核体位点特异性地安装至配有lpxtg基序的蛋白质上(popp,w.m.等proc.natl.acad.sci.usa108(2011)3169-3174)。相关的动物酿脓链球菌(streptococcuspyogenes)分选酶接受基于二-丙氨酸的亲核体,金黄色葡萄球菌酶则不然。此分选酶在苏氨酸和丙氨酸之间切割lpxta基序,并允许安装经修饰的基于丙氨酸的亲核体。虽然效率低,但srtastrep也识别和切割lpxtg基序,但是,lpxta基序难以通过srtastaph切割(popp等上文)。
在wo2010/087994中,报道了用于连接的方法及其用途。marvin,j.s.等(actapharmacol.sinica26(2005)649-658)报道了igg样双特异性抗体的重组方法。levary,d.a.等(plosone,6(2011)e18342.1-e18342.6)报道了分选酶a催化的蛋白质-蛋白质融合。在wo2013/003555中,报道了用分选酶安装点击化学把手进行蛋白质连接。
strijbis,k.等(traffic13(2012)780-789)报道了用分选酶在活细胞中进行蛋白质连接。他们已指出,依赖ca2+的金黄色葡萄球菌分选酶a在细胞内没有功能,但不依赖ca2+的动物酿脓链球菌分选酶a在酿酒酵母(saccharomycescerevisiae)和哺乳动物hek293t细胞二者的胞质和内质网(er)腔中都具有功能。
levary,d.a.等报道了分选酶a催化的蛋白质-蛋白质融合(plosone6(2011))。madej,m.p.等(biotechnol.bioeng.109(2012)1461-1470)报道了将抗表皮生长因子受体抗体改造为单链型式,并通过分选酶a介导的蛋白质连接进行标记。ta,h.t.等报道了酶促单链抗体标记作为心血管疾病中靶分子成像和细胞归巢的通用方法(cir.res.109(2011)365-373)。popp,m.等报道了产生和断裂肽键——使用分选酶的蛋白质改造(angew.chem.int.ed.eng.50(2011)5024-5032)。在wo2010/087994中,报道了用于连接的方法及其用途。wo2010/099536中报道了对dota螯合物具有高亲和力的改造蛋白质。
已将缺乏n端膜锚定基序的截短srta用于细胞表面蛋白质标记、供价蛋白质固定化及在蛋白质中掺入新功能。但是,如果使用等摩尔量的底物,srta介导连接的产率总是低于70%,因为反应是可逆的。另一个缺点是反应中间体的水解,其产生lpxt产物而不是预期产物。这在与srta长时间孵育时尤其有问题。这也意味着残留在最终产物中的甚至小量srta可以随时间推移而破坏它;这对质量标准非常高的生物制品而言是个大问题。
已报道了阻断分选酶的逆反应的不同努力。yamamura,y.等(chem.commun.47(2011)4742-4744)报道了通过在底物识别位点附近引入β-发夹,在连接位点附近诱导β-发夹结构来增强分选酶a介导的蛋白质连接。
biswas,t.等(biochem.53(2014)2515-2524)报道了通过原型分选酶分选lpxtg肽、不变底物残基在调节酶动力学中的作用及生产底物的构象特征。
li,y.m.等报道了通过使用分选酶来使蛋白质发生不可逆位点特异性肼解(angew.chem.int.ed.engl.53(2014)2198-2202)。
在wo2014/001324中,报道了用于选择和产生包含至少两个不同结合实体的定制高选择性和多特异性靶向实体的方法及其用途。marraffini,l.a.等(j.biol.chem.279(2004)37763-37770)报道,为了将表面蛋白质锚定至金黄色葡萄球菌细胞壁,分选酶a的有效催化需要保守的精氨酸残基。
发明概述
已发现,通过同时与源自金黄色葡萄球菌的第一分选酶和源自动物酿脓链球菌的第二分选酶孵育,可以在酶促一锅反应(one-potreaction)中形成两个肽键。该第一分选酶对i)分选酶氨基酸基序lpxtg(seqidno:20,其中x可以是任意氨基酸残基)和ii)作为亲核体的寡聚甘氨酸(seqidno:22至25)的组合特异,而该第二分选酶对i)分选酶氨基酸基序lpxta(seqidno:31,其中x可以是任意氨基酸残基)和ii)作为亲核体的寡聚丙氨酸(seqidno:26至29)的组合特异。由于这两种分选酶具有正交特异性,可以作为酶促一锅反应同时进行两个转肽反应。
因此,本文报道的一个方面是用于产生三种多肽的酶促缀合产物的方法,包括以下步骤:
-同时孵育
a)i)含有氨基酸序列lpxtg(seqidno:20,其中x可以是任意氨基酸残基)的第一多肽;
ii)含有氨基酸序列lpxta(seqidno:31,其中x可以是任意氨基酸残基)的第二多肽;
iii)具有两个n端的第三多肽,由此该多肽在其第一n端具有寡聚甘氨酸gm(m=2(seqidno:22)、或3(seqidno:23)、或4(seqidno:24)、或5(seqidno:25))氨基酸序列,在其第二n端具有寡聚丙氨酸am(m=2(seqidno:26)、或3(seqidno:27)、或4(seqidno:28)、或5(seqidno:29))氨基酸序列;
iv)具有分选酶活性的第四多肽,因为该多肽源自金黄色葡萄球菌分选酶a;和
v)具有分选酶活性的第五多肽,因为该多肽源自动物酿脓链球菌分选酶a;
或
b)i)含有氨基酸序列lpxtg(seqidno:20,其中x可以是任意氨基酸残基)的第一多肽;
ii)在其n端具有寡聚甘氨酸gm(m=2(seqidno:22)、或3(seqidno:23)、或4(seqidno:24)、或5(seqidno:25))氨基酸序列且包含氨基酸序列lpxta(seqidno:31,其中x可以是任意氨基酸残基)的第二多肽;
iii)在其n端具有寡聚丙氨酸am(m=2(seqidno:26)、或3(seqidno:27)、或4(seqidno:28)、或5(seqidno:29))氨基酸序列的第三多肽;
iv)具有分选酶活性的第四多肽,因为该多肽源自金黄色葡萄球菌分选酶a;和
v)具有分选酶活性的第五多肽,因为该多肽源自动物酿脓链球菌分选酶a;
或
c)i)含有氨基酸序列lpxta(seqidno:31,其中x可以是任意氨基酸残基)的第一多肽;
ii)在其n端具有寡聚丙氨酸am(m=2(seqidno:26)、或3(seqidno:27)、或4(seqidno:28)、或5(seqidno:29))氨基酸序列且包含氨基酸序列lpxtg(seqidno:20,其中x可以是任意氨基酸残基)的第二多肽;
iii)在其n端具有寡聚甘氨酸gm(m=2(seqidno:22)、或3(seqidno:23)、或4(seqidno:24)、或5(seqidno:25))氨基酸序列的第三多肽;
iv)具有分选酶活性的第四多肽,因为该多肽源自金黄色葡萄球菌分选酶a;和
v)具有分选酶活性的第五多肽,因为该多肽源自动物酿脓链球菌分选酶a;
和
-从反应混合物回收该缀合物,从而产生三种多肽的酶促缀合产物。
在一个实施方案中,该第一、第二和第三多肽大致按等摩尔浓度存在。
在一个实施方案中,该寡聚丙氨酸是二-丙氨酸(seqidno:26)或三-丙氨酸(seqidno:27)。
在一个实施方案中,该寡聚甘氨酸是二-甘氨酸(seqidno:22)或三-甘氨酸(seqidno:23)。
在一个实施方案中,该第四多肽具有氨基酸序列seqidno:21。
在一个实施方案中,该第五氨基酸序列具有氨基酸序列seqidno:34。
在一个实施方案中,该第一多肽、该第二多肽和该第三多肽相互独立地选自抗体可变结构域、抗体重链fab片段、抗体fc区和标记。
在一个实施方案中,该方法进一步包括纯化三种多肽的酶促缀合产物的步骤。
在一个实施方案中,氨基酸序列lpxtg(seqidno:20,其中x可以是任意氨基酸残基)和/或氨基酸序列lpxta(seqidno:31,其中x可以是任意氨基酸残基)后有至少一个氨基酸残基。
发明详述
i.定义
在本说明书和权利要求书中,免疫球蛋白重链fc区中残基的编号是kabat(kabat,e.a.等,sequencesofproteinsofimmunologicalinterest,第5版,publichealthservice,nationalinstitutesofhealth,bethesda,md(1991),nihpublication91-3242,在此明确引入作为参考)的eu指数的编号。
术语“改变”指在例如至少包含fc区的fcrn结合部分的抗体或融合多肽的亲本氨基酸序列中突变、添加或缺失一个或多个氨基酸残基,以获得变体抗体或融合多肽。
术语“氨基酸突变”指亲本氨基酸序列的氨基酸序列中的修饰。示例性修饰包括氨基酸取代、插入和/或缺失。在一个实施方案中,该氨基酸突变是取代。术语“位置上的氨基酸突变”指取代或缺失指定的残基,或在邻近指定残基处插入至少一个氨基酸残基。术语“邻近指定残基的插入”指其一个或两个残基内的插入。插入可以在指定残基的n端或c端。
术语“氨基酸取代”指用不同的“替换”氨基酸残基替换预先确定的亲本氨基酸序列中的至少一个氨基酸残基。该一个或多个替换残基可以是“天然存在的氨基酸残基”(即由遗传密码编码),且选自:丙氨酸(ala);精氨酸(arg);天冬酰胺(asn);天冬氨酸(asp);半胱氨酸(cys);谷氨酰胺(gln);谷氨酸(glu);甘氨酸(gly);组氨酸(his);异亮氨酸(ile);亮氨酸(leu);赖氨酸(lys);甲硫氨酸(met);苯丙氨酸(phe);脯氨酸(pro);丝氨酸(ser);苏氨酸(thr);色氨酸(trp);酪氨酸(tyr);和缬氨酸(val)。在一个实施方案中,该替换残基不是半胱氨酸。一个或多个非天然存在或非蛋白质的氨基酸残基的取代也为本文的氨基酸取代的定义所涵盖。“非天然存在的氨基酸残基”指除上文所列出的那些天然存在的氨基酸残基外的残基,其能够共价结合多肽链中的一个或多个邻近氨基酸残基。非天然存在的氨基酸残基的实例包括正亮氨酸、鸟氨酸、正缬氨酸、高丝氨酸、aib和其他氨基酸残基类似物,如ellman等,meth.enzym.202(1991)301-336中所述的那些。为了产生这类非天然存在的氨基酸残基,可以使用noren等(science244(1989)182)和/或ellman等(上文)的方案。简言之,这些方案涉及用非天然存在的氨基酸残基化学活化阻抑型trna,然后体外转录和翻译rna。非天然存在的氨基酸还可以通过化学肽合成掺入肽,然后使这些肽与重组产生的多肽(如抗体或抗体片段)融合。
术语“氨基酸插入”指在预先确定的亲本氨基酸序列中掺入至少一个附加的氨基酸残基。虽然插入通常将包括插入一个或两个氨基酸残基,但本申请考虑更大的“肽插入”,例如插入约3个至约5个或甚至至多约10个氨基酸残基。一个或多个插入残基可以是上文定义的天然存在或非天然存在的残基。
术语“氨基酸缺失”指在氨基酸序列中预先确定的位置上去除至少一个氨基酸残基。
在本申请内,任何时候提到氨基酸改变时,它是有意的氨基酸改变,而不是随机氨基酸修饰。
术语“标记”指具有特异性结合特性的经肽键相互连接的氨基酸残基序列。在一个实施方案中,该标记是亲和或纯化标记。在一个实施方案中,该标记选自arg标记、his标记、flag标记、3xflag标记、strep标记、nano标记、sbp标记、c-myc标记、s标记、钙调蛋白结合肽、纤维素结合结构域、壳多糖结合结构域、gst标记或mbp标记。在一个实施方案中,该标记选自seqidno:01(rrrrr)、或seqidno:02(rrrrrr)、或seqidno:03(hhhhhh)、或seqidno:04(kdhlihnvhkefhahahnk)、或seqidno:05(dykddddk)、或seqidno:06(dykdhdgdykdhdidykddddk)、或seqidno:07(awrhpqfgg)、或seqidno:08(wshpqfek)、或seqidno:09(mdveawlgar)、或seqidno:10(mdveawlgarvplvet)、或seqidno:11(mdekttgwrgghvveglageleqlrarlehhpqgqrep)、或seqidno:12(eqkliseedl)、或seqidno:13(ketaaakferqhmds)、或seqidno:14(krrwkknfiavsaanrfkkisssgal)、或seqidno:15(纤维素结合结构域)、或seqidno:16(纤维素结合结构域)、或seqidno:17(tnpgvsawqvntaytagqlvtyngktykclqphtslagwepsnvpalwqlq)、或seqidno:18(gst标记)、或seqidno:19(mbp标记)、或seqidno:35(c标记)。
活性剂(agent)(例如药物制剂)的“有效量”指对在必要的剂量下和时间内达到希望得到的治疗或预防结果的有效量。
术语“个体”或“受试体”是哺乳动物。哺乳动物包括但不限于饲养的动物(例如牛、绵羊、猫、狗和马)、灵长类(例如人类和非人灵长类,如猴)、兔和啮齿类(例如小鼠、大鼠和仓鼠)。在某些实施方案中,该个体是人。
术语“药物制剂”指这样的制备物,其以这样的形式存在,使得允许包含在其中的活性成分的生物活性有效,且不包含对将要施用该制剂的个体具有不可接受的毒性的附加成分。
“可药用载体”指药物制剂中除活性成分之外的成分,其对个体无毒性。可药用载体包括但不限于缓冲剂、赋形剂、稳定剂或防腐剂。
术语“位置”指多肽的氨基酸序列中的氨基酸残基的位置。位置可以顺次编号,或按照确定的型式(例如用于抗体编号的kabat的eu指数)编号。
本文所用的“处理”(及其语法变形)指改变所处理的个体的自然过程的尝试中的临床干预,且可以为了预防而进行或在临床病理的过程中进行。希望得到的处理作用包括但不限于防止疾病的发生或复发、减轻症状、减少疾病的任何直接或间接的病理结果、防止转移、降低疾病进展的速率、改善或缓和疾病状态及消退或改善的预后。在一些实施方案中,用本发明的抗体来延迟疾病的发展或减慢疾病的进展。
ii.使用分选酶的酶促缀合
可以通过使用分选酶(尤其是分选酶a)来在体外获得包含两个在体内不共价结合的实体的共价缀合物。
分选酶a(srta)是将蛋白质共价附着至细菌细胞壁的膜结合酶。srta底物上的特异性识别基序是lpxtg,由此该酶在残基苏氨酸和甘氨酸之间切割。肽聚糖上的识别基序是五甘氨酸基序。已显示n端上的三甘氨酸和甚至二甘氨酸基序足以支持srta反应(clancy,k.w.等,peptidescience94(2010)385-396)。反应通过硫酯酰基-酶中间体进行,该中间体通过来自寡聚甘氨酸的胺亲核体的攻击而分解,将蛋白质底物共价连接至肽聚糖,并再生srta。srta可以用于将化学合成的肽共价缀合至重组表达的蛋白质。
许多革兰氏阳性菌用分选酶来将多种表面蛋白质(包括毒力因子)共价锚定至其细胞壁(肽聚糖)。分选酶是膜结合酶。野生型金黄色葡萄球菌分选酶a(srta)是具有n端跨膜区的206个氨基酸的多肽。在第一步中,分选酶a识别含有lpxtg(seqidno:20)氨基酸序列基序的底物蛋白质,并借助活性部位cys切割thr和gly之间的酰胺键。此肽切割反应产生分选酶a-底物硫酯中间体。在第二步中,该硫酯酰基-酶中间体通过含有寡聚甘氨酸的第二底物多肽(对应于金黄色葡萄球菌中肽聚糖的五甘氨酸单位)的(n端)氨基的亲核攻击而分解,产生共价连接的细胞壁蛋白质,并再生分选酶a。在缺乏寡聚甘氨酸的情况下,酰基-酶中间体可以被水分子水解。
分选酶介导的连接/缀合已开始应用于多种蛋白质改造和生物缀合目的。此技术能够在lpxtg标记的重组或化学合成多肽中引入天然和合成的功能性。实例包括共价附着寡聚甘氨酸衍生的聚合物(例如peg)、荧光团、维生素(例如生物素和叶酸)、脂质、糖类、核酸、合成肽和蛋白质(例如gfp)(参见例如tsukiji,s.和nagamune,t.,chembiochem10(2009)787-798;popp,m.w.l.和ploegh,h.l.,angew.chem.int.ed.engl.50(2011)5024-5032)。
对于酶促缀合,可以使用缺乏跨膜区(srta;金黄色葡萄球菌srta的氨基酸残基60-206)的可溶性截短分选酶a(seqidno:21;还参见ton-that,h.等,proc.natl.acad.sci.usa96(1999)12424-12429;ilangovan,h.等,proc.natl.acad.sci.usa98(2001)6056-6061)。
iii.重组方法
可以从真核细胞(例如hek293细胞、cho细胞)表达并从上清纯化在其n端包含寡聚甘氨酸基序(gg(seqidno:22)、ggg(seqidno:23)、gggg(seqidno:24)、ggggg(seqidno:25))的任何多肽结构域(例如单链抗原结合多肽如scfv、scfab或darpin,或多链抗原结合多肽如dsfv或fab)。多肽是分离的多肽还是包含在多聚或异聚实体中没有关系。
适合用于克隆或表达/分泌编码多肽的载体的宿主细胞包括本文所述的原核或真核细胞。例如,尤其是在不需要糖基化时,可以在细菌中产生多肽(参见例如us5,648,237、us5,789,199和us5,840,523,charlton,methodsinmolecularbiology248(2003)245-254(b.k.c.lo,(编辑),humanapress,totowa,nj),描述在大肠杆菌(e.coli.)中表达抗体片段)。表达后,可以从可溶性级分中的细菌细胞糊分离或可以从所谓内含体(且可以溶解并重折叠为生物活性形式)的不溶性级分分离多肽。然后可以进一步纯化多肽。
除原核生物外,诸如丝状真菌或酵母的真核微生物也是适合用于编码多肽的载体的克隆或表达宿主,包括糖基化途径已“人源化”,导致产生具有部分或完全人糖基化模式的多肽的真菌和酵母菌株(参见例如gerngross,nat.biotech.22(2004)1409-1414和li等,nat.biotech.24(2006)210-215)。
适合用于表达糖基化多肽的宿主细胞还源自多细胞生物(无脊椎动物和脊椎动物)。无脊椎动物细胞的实例包括植物细胞和昆虫细胞。已鉴定了许多杆状病毒株,其可以与昆虫细胞结合使用,尤其是用于转染草地夜蛾(spodopterafrugiperda)细胞。
也可以用植物细胞培养物作为宿主(参见例如us5,959,177、us6,040,498、us6,420,548、us7,125,978和us6,417,429(描述用于在转基因植物中产生抗体的plantibodiestm技术))。
可以用脊椎动物细胞作为宿主。例如,可以使用适应悬浮生长的哺乳动物细胞系。有用的哺乳动物宿主细胞系的实例是cos-7细胞系(sv40转化的猴肾cv1细胞);hek293细胞系(人胚肾细胞);bhk细胞系(幼仓鼠肾);tm4小鼠支持细胞系(例如mather,biol.reprod.23(1980)243-251中所述的tm4细胞);cvl细胞系(猴肾细胞);vero-76细胞系(非洲绿猴肾细胞);hela细胞系(人宫颈癌细胞);mdck细胞系(犬肾细胞);brl3a细胞系(buffalo大鼠肝细胞);w138细胞系(人肺细胞);hepg2细胞系(人肝细胞);mmt060562细胞系(小鼠乳腺肿瘤细胞);tri细胞系(例如描述于mather等,anal.n.y.acad.sci.383(1982)44-68中);mrc5细胞系;和fs4细胞系。其他有用的哺乳动物宿主细胞系包括cho细胞系(中国仓鼠卵巢细胞),包括dhfr阴性cho细胞系(参见例如urlaub等,proc.natl.acad.sci.usa77(1980)4216);及骨髓瘤细胞系,如y0、ns0和sp2/0细胞系。某些适合用于多肽产生的哺乳动物宿主细胞系的综述参见例如yazaki和wu,methodsinmolecularbiology,antibodyengineering248(2004)255-268(b.k.c.lo,(编辑),humanapress,totowa,nj)。
iv.本文报道的方法
已发现,通过同时与源自金黄色葡萄球菌的第一分选酶和源自动物酿脓链球菌的第二分选酶孵育,可以在酶促一锅反应中形成两个肽键。该第一分选酶对i)分选酶氨基酸基序lpxtg(seqidno:20,其中x可以是任意氨基酸残基)和ii)作为亲核体的寡聚甘氨酸(seqidno:22至25)的组合特异,而该第二分选酶对i)分选酶氨基酸基序lpxta(seqidno:31,其中x可以是任意氨基酸残基)和ii)作为亲核体的寡聚丙氨酸(seqidno:26至29)的组合特异。由于这两种分选酶具有正交特异性,可以作为酶促一锅反应同时进行两个转肽反应。
因此,本文报道的一个方面是用于产生三种多肽的酶促缀合产物的方法,包括以下步骤:
-同时孵育
a)i)含有氨基酸序列lpxtg(seqidno:20,其中x可以是任意氨基酸残基)的第一多肽;
ii)含有氨基酸序列lpxta(seqidno:31,其中x可以是任意氨基酸残基)的第二多肽;
iii)具有两个n端的第三多肽,由此该多肽在其第一n端具有寡聚甘氨酸gm(m=2(seqidno:22)、或3(seqidno:23)、或4(seqidno:24)、或5(seqidno:25))氨基酸序列,在其第二n端具有寡聚丙氨酸am(m=2(seqidno:26)、或3(seqidno:27)、或4(seqidno:28)、或5(seqidno:29))氨基酸序列;
iv)具有分选酶活性的第四多肽,因为该多肽源自金黄色葡萄球菌分选酶a;和
v)具有分选酶活性的第五多肽,因为该多肽源自动物酿脓链球菌分选酶a;
或
b)i)含有氨基酸序列lpxtg(seqidno:20,其中x可以是任意氨基酸残基)的第一多肽;
ii)在其n端具有寡聚甘氨酸gm(m=2(seqidno:22)、或3(seqidno:23)、或4(seqidno:24)、或5(seqidno:25))氨基酸序列且包含氨基酸序列lpxta(seqidno:31,其中x可以是任意氨基酸残基)的第二多肽;
iii)在其n端具有寡聚丙氨酸am(m=2(seqidno:26)、或3(seqidno:27)、或4(seqidno:28)、或5(seqidno:29))氨基酸序列的第三多肽;
iv)具有分选酶活性的第四多肽,因为该多肽源自金黄色葡萄球菌分选酶a;和
v)具有分选酶活性的第五多肽,因为该多肽源自动物酿脓链球菌分选酶a;
或
c)i)含有氨基酸序列lpxta(seqidno:31,其中x可以是任意氨基酸残基)的第一多肽;
ii)在其n端具有寡聚丙氨酸am(m=2(seqidno:26)、或3(seqidno:27)、或4(seqidno:28)、或5(seqidno:29))氨基酸序列且包含氨基酸序列lpxtg(seqidno:20,其中x可以是任意氨基酸残基)的第二多肽;
iii)在其n端具有寡聚甘氨酸gm(m=2(seqidno:22)、或3(seqidno:23)、或4(seqidno:24)、或5(seqidno:25))氨基酸序列的第三多肽;
iv)具有分选酶活性的第四多肽,因为该多肽源自金黄色葡萄球菌分选酶a;和
v)具有分选酶活性的第五多肽,因为该多肽源自动物酿脓链球菌分选酶a;
和
-从反应混合物回收该缀合物,从而产生三种多肽的酶促缀合产物。
该第一、第二和第三多肽可以相互独立地包含一个或多个非分选酶基序部分。这些部分可以直接或通过接头与分选酶氨基酸序列基序连接。
非分选酶基序部分
如果它不直接包含在这些分子之一中,则可以将分选酶基序氨基酸序列lpxtg缀合至治疗剂(药物)、细胞毒性剂(例如毒素,如阿霉素或百日咳毒素)、荧光团(如荧光染料,如二氢荧光素或罗丹明)、用于成像的螯合剂或放射性治疗金属、肽基或非肽基标记、标记、或清除修饰剂(如聚乙二醇的多种异构体)、结合第三成分的肽、或另一糖类或亲脂剂。缀合可以是直接缀合或经居间接头缀合。
a)治疗部分
药物部分可以是具有治疗作用的任意化合物、部分或基团,如抗体、细胞毒性或细胞抑制化合物。
已知许多抗细胞表面分子及其配体的治疗性抗体,如美罗华(rituxan)/mabthera/利妥昔单抗(rituximab)、2h7/ocrelizumab、zevalin/ibrizumomab、arzerra/ofatumumab(cd20)、hll2/epratuzumab、inotuzomab(cd22)、zenapax/daclizumab、simulect/巴利昔单抗(basiliximab)(cd25)、赫赛汀(herceptin)/曲妥珠单抗(trastuzumab)、帕妥珠单抗(pertuzumab)(her2/erbb2)、mylotarg/吉妥珠单抗(gemtuzumab)(cd33)、raptiva/efalizumab(cd11a)、爱必妥(erbitux)/西妥昔单抗(cetuximab)(egfr,表皮生长因子受体)、imc-1121b(vegf受体2)、tysabri/那他珠单抗(natalizumab)(α4β1和α4β7整联蛋白的α4亚基)、reopro/阿昔单抗(abciximab)(gpiib-gpiia和αvβ3整联蛋白)、orthocloneokt3/muromonab-cd3(cd3)、benlysta/belimumab(baff)、tolerx/oteliximab(cd3)、soliris/eculizumab(c5补体蛋白质)、actemra/tocilizumab(il-6r)、panorex/依决洛单抗(edrecolomab)(epcam,表皮细胞黏附分子)、cea-cam5/labetuzumab(cd66/cea,癌胚抗原)、ct-11(pd-1,程序性死亡-1t细胞抑制受体,cd-d279)、h224g11(c-met受体)、sar3419(cd19)、imc-a12/cixutumumab(igf-1r,胰岛素样生长因子1受体)、medi-575(pdgf-r,血小板衍生生长因子受体)、cp-675、206/tremelimumab(细胞毒性t淋巴细胞抗原4)、ro5323441(胎盘生长因子或pgf)、hgs1012/mapatumumab(trail-r1)、sgn-70(cd70)、vedotin(sgn-35)/brentuximab(cd30)和arh460-16-2(cd44)。
用本文报道的方法获得的缀合物可以用于制备用于治疗例如肿瘤学疾病、心血管疾病、感染性疾病、炎性疾病、自身免疫病、代谢性(例如内分泌)疾病或神经(例如神经变性)疾病的药物。这些疾病的示例性非限制性实例是阿尔茨海默病、非霍奇金淋巴瘤、b细胞急性和慢性淋巴细胞白血病、伯基特淋巴瘤、霍奇金淋巴瘤、多毛细胞白血病、急性和慢性髓细胞白血病、t细胞淋巴瘤和白血病、多发性骨髓瘤、神经胶质瘤、waldenstrom巨球蛋白血症、癌(如口腔、胃肠道、结肠、腹部、肺道、肺、乳腺、卵巢、前列腺、子宫、子宫内膜、宫颈、膀胱、胰腺、骨、肝、胆囊、肾、皮肤和睾丸癌)、黑素瘤、肉瘤、神经胶质瘤、皮肤癌、急性特发性血小板减少性紫癜、慢性特发性血小板减少性紫癜、皮肌炎、sydenham舞蹈症、重症肌无力、系统性红斑狼疮、狼疮肾炎、风湿热、多腺性综合征、大疱性类天疱疮、糖尿病、henoch-schonlein紫癜、链球菌感染后肾炎、结节性红斑、takayasu动脉炎、addison病、类风湿性关节炎、多发性硬化、结节病、溃疡性结肠炎、多形性红斑、iga肾病、结节性多动脉炎、关节强硬性脊髓炎、goodpasture综合征、血栓闭塞性脉管炎、sjogren综合征、原发性胆汁性肝硬化、桥本甲状腺炎、甲状腺毒症、硬皮病、慢性活动性肝炎、多肌炎/皮肌炎、多软骨炎、寻常性天疱疮、wegener肉芽肿病、膜性肾病、肌萎缩性侧索硬化症、脊髓痨、巨细胞性动脉炎/多肌痛、恶性贫血、急进性肾小球肾炎、银屑病或致纤维性肺泡炎。
已知许多细胞表面标志及其配体。例如,已报道癌细胞至少表达以下细胞表面标志和/或配体之一,包括但不限于:碳酸酐酶ix、甲胎蛋白、α-辅肌动蛋白-4、a3(a33抗体特异性抗原)、art-4、b7、ba-733、bage、bre3抗原、ca125、camel、cap-1、casp-8/m、cccl19、cccl21、cd1、cd1a、cd2、cd3、cd4、cds、cd8、cd1-1a、cd14、cd15、cd16、cd18、cd19、cd20、cd21、cd22、cd23、cd25、cd29、cd30、cd32b、cd33、cd37、cd38、cd40、cd40l、cd45、cd46、cd54、cd55、cd59、cd64、cd66a-e、cd67、cd70、cd74、cd79a、cd80、cd83、cd95、cd126、cd133、cd138、cd147、cd154、cdc27、cdk-4/m、cdkn2a、cxcr4、cxcr7、cxcl12、hif-l-α、结肠特异性抗原p(csap)、cea(ceacam5)、ceacam6、c-met、dam、egfr、egfrviii、egp-1、egp-2、elf2-m、ep-cam、flt-1、flt-3、叶酸受体、g250抗原、gage、grob、hla-dr、hm1.24、人绒毛膜促性腺激素(hcg)及其亚基、her2/neu、hmgb-1、低氧诱导因子(hif-1)、hsp70-2m、hst-2或la、igf-1r、ifn-γ、ifn-α、ifn-β、il-2、il-4r、il-6r、il-13r、il-15r、il-17r、il-18r、il-6、il-8、il-12、il-15、il-17、il-18、il-25、胰岛素样生长因子-1(igf-1)、kc4抗原、ks-1抗原、ks1-4、le-y、ldr/fut、巨噬细胞迁移抑制因子(mif)、mage、mage-3、mart-1、mart-2、ny-eso-1、trag-3、mcrp、mcp-1、mip-1a、mip-1b、mif、muc1、muc2、muc3、muc4、muc5、mum-1/2、mum-3、nca66、nca95、nca90、胰腺癌黏蛋白、胎盘生长因子、p53、plagl2、前列腺酸性磷酸酶、psa、prame、psma、p1gf、ilgf、ilgf-1r、il-6、il-25、rs5、rantes、t101、sage、s100、存活蛋白、存活蛋白-2b、tac、tag-72、生腱蛋白、trail受体、tnf-α、tn抗原、thomson-friedenreich抗原、肿瘤坏死抗原、vegfr、ed-b纤连蛋白、wt-1、17-1a抗原、补体因子c3、c3a、c3b、c5a、c5、血管发生标志、bcl-2、bcl-6、kras、cmet、癌基因标志和癌基因产物(参见例如sensi等,clin.cancerres.12(2006)5023-5032;parmiani等,j.immunol.178(2007)1975-1979;novellino等,cancerimmunol.immunother.54(2005)187-207).
因此,识别特异性细胞表面受体(包括其配体)的抗体可以用于特异性和选择性靶向和结合许多/大量疾病相关细胞表面标志。细胞表面标志是例如与信号发放事件或配体结合相关的定位在细胞表面(例如疾病相关细胞)的多肽。
在一个实施方案中,为了治疗癌症/肿瘤,产生靶向肿瘤相关抗原的多特异性结合分子/双特异性抗体,该肿瘤相关抗原如fleisher(编辑),"theclinicalbiochemistryofcancer",第347页(americanassociationofclinicalchemists(1979))中的herberman,"immunodiagnosisofcancer"中及us4,150,149、us4,361,544和us4,444,744中报道的那些。
关于肿瘤相关抗原(taa)的报道包括mizukami等,(naturemed.11(2005)992-997);hatfield等,(curr.cancerdrugtargets5(2005)229-248);vallbohmer等,(jclin.oncol.23(2005)3536-3544);和ren等,(ann.surg.242(2005)55-63),每篇文献在此就所鉴定的taa引入作为参考。
在疾病涉及淋巴瘤、白血病或自身免疫障碍时,靶向的抗原可以选自:cd4、cd5、cd8、cd14、cd15、cd19、cd20、cd21、cd22、cd23、cd25、cd33、cd37、cd38、cd40、cd40l、cd46、cd54、cd67、cd74、cd79a、cd80、cd126、cd138、cd154、cxcr4、b7、muc1或la、hm1.24、hla-dr、生腱蛋白、vegf、p1gf、ed-b纤连蛋白、癌基因、癌基因产物(例如c-met或plagl2)、cd66a-d、坏死抗原、il-2、t101、tag、il-6、mif、trail-r1(dr4)和trail-r2(dr5)。
已知许多抗两种不同靶标的双特异性抗体,该两种不同靶标如bcma/cd3;her家族的不同抗原的组合(egfr、her2、her3);cd19/cd3;il17ra/il7r;il-6/il-23;il-1-β/il-8;il-6或il-6r/il-21或il-21r;第一特异性针对选自lewisx、lewisb和lewisy结构、globoh结构、kh1、tn抗原、tf抗原和黏蛋白的糖类结构、cd44、糖脂和鞘糖脂(如gg3、gb3、gd3、gd2、gb5、gm1、gm2、唾液酰基四糖神经酰胺(sialyltetraosylceramide))的抗原的糖表位,第二特异性针对选自egfr、her2、her3和her4的erbb受体酪氨酸激酶;gd2与选自t淋巴细胞nk细胞、b淋巴细胞、树突细胞、单核细胞、巨噬细胞、嗜中性粒细胞、间充质干细胞、神经干细胞的免疫细胞相关的第二抗原结合部位的组合;ang2/vegf;vegf/pdgfr-β;血管内皮生长因子(vegf)受体2/cd3;psma/cd3;epcam/cd3;选自vegfr-1、vegfr-2、vegfr-3、flt3、c-fms/csf1r、ret、c-met、egfr、her2/neu、her3、her4、igfr、pdgfr、c-kit、bcr、整联蛋白和mmp的抗原与选自vegf、egf、pigf、pdgf、hgf和血管生成素(angiopoietin)的水溶性配体的组合;erbb-3/c-met;erbb-2/c-met;egf受体1/cd3;egfr/her3;psca/cd3;c-met/cd3;endosialin/cd3;epcam/cd3;igf-1r/cd3;fapalpha/cd3;egfr/igf-1r;il17a/f;egf受体1/cd3和cd19/cd16。
毒性药物部分包括:(i)化疗剂,其可以作为微管抑制剂、有丝分裂抑制剂、拓扑异构酶抑制剂或dna嵌入剂发挥作用;(ii)蛋白质毒素,其可以通过酶活性发挥作用;和(iii)放射性同位素。
示例性毒性药物部分包括但不限于美登木素生物碱(maytansinoid)、auristatin、多拉司他汀(dolastatin)、单端孢霉烯(trichothecene)、cc1065、加利车霉素(calicheamicin)及其他烯二炔类(enediyne)抗生素、紫杉烷(taxane)、蒽环类抗生素(anthracycline)、及其立体异构体、等构物、类似物或衍生物。
蛋白质毒素包括白喉a链、白喉毒素的非结合活性片段、外毒素a链(来自铜绿假单胞菌(pseudomonasaeruginosa))、蓖麻毒蛋白a链(vitetta等(1987)science,238:1098)、相思豆毒蛋白a链、蒴莲根毒蛋白a链、α-帚曲霉素、油桐(aleuritesfordii)蛋白质、香石竹毒蛋白蛋白质、美洲商陆(phytolacaamericana)蛋白质(papi、papii和pap-s)、苦瓜(momordicacharantia)抑制剂、麻风树毒蛋白、巴豆毒蛋白、sapaonariaofficinalis抑制剂、多花白树毒蛋白、米托洁林(mitogellin)、局限曲霉素、酚霉素、伊诺霉素和单端孢霉烯(wo93/21232)。
治疗性放射性同位素包括32p、33p、90y、125i、131i、131in、153sm、186re、188re、211at、212b、212pb及lu的放射性同位素。
放射性同位素或其他标记可以以已知方式掺入(fraker等(1978)biochem.biophys.res.commun.80:49-57;"monoclonalantibodiesinimmunoscintigraphy"chatal,crcpress1989)。碳14标记的3-甲基二乙烯三胺五乙酸1-异硫氰基苄酯(mx-dtpa)是用于将放射性核素缀合至复合物的示例性螯合剂(wo94/11026)。
b)标记
非分选酶基序部分可以是标记。可以使用可以共价附着至分选酶氨基酸序列的任何标记部分(参见例如singh等(2002)anal.biochem.304:147-15;harlowe.和lane,d.(1999)usingantibodies:alaboratorymanual,coldspringsharborlaboratorypress,coldspringharbor,n.y.;lundbladr.l.(1991)chemicalreagentsforproteinmodification,第2版crcpress,bocaraton,fla.)。标记可以发挥功能来:(i)提供可检测信号;(ii)与第二标记相互作用来修饰由该第一或第二标记提供的可检测信号,例如以提供fret(荧光共振能量转移);(iii)通过电荷、疏水性、形状或其他物理参数影响迁移率,例如电泳迁移率或细胞通透性;或(iv)提供捕获部分,例如以调节离子络合。
本文报道的包含半抗原化标记的缀合物可以用于诊断测定,例如用于检测目的抗原在具体细胞、组织或血清中的表达。对于诊断应用,将使用双特异性抗体,其中第一结合特异性结合靶标,第二结合特异性结合半抗原化标记。半抗原通常将用可检测部分标记。有许多标记可用,这些标记通常分为以下类别:
(a)放射性同位素(放射性核素),如3h、11c、14c、18f、32p、35s、64cu、68gn、86y、89zr、99tc、111in、123i、124i、125i、131i、133xe、177lu、211at或131bi。放射性同位素标记的缀合物可以用于靶向报道分子的成像实验。可以使用currentprotocolsinimmunology,(1991)1和2卷,coligen等编辑wiley-interscience,newyork,n.y.,pubs中所述的技术,用结合、螯合或以其他方式络合放射性同位素金属的配体试剂标记抗原(半抗原)。可以络合金属离子的螯合配体包括dota、dotp、dotma、dtpa和teta(macrocyclics,dallas,tex.)。放射性核素可以经与本文报道的复合物络合而靶向(wu等,naturebiotechnology23(9)(2005)1137-1146)。用放射性核素标记复合物进行受体靶标成像,可以通过检测和定量复合物或对应的治疗性抗体在肿瘤组织中的进行性累积来提供途径激活的标志(albert等(1998)bioorg.med.chem.lett.8:1207-1210)。
金属-螯合剂复合物适合作为用于成像实验的标志(us2010/0111856;us5,342,606;us5,428,155;us5,316,757;us5,480,990;us5,462,725;us5,428,139;us5,385,893;us5,739,294;us5,750,660;us5,834,456;hnatowich等,j.immunol.methods65(1983)147-157;meares等,anal.biochem.142(1984)68-78;mirzadeh等,bioconjugatechem.1(1990)59-65;meares等,j.cancer(1990),suppl.10:21-26;izard等,bioconjugatechem.3(1992)346-350;nikula等,nucl.med.biol.22(1995)387-90;camera等,nucl.med.biol.20(1993)955-62;kukis等,j.nucl.med.39(1998)2105-2110;verel等.,j.nucl.med.44(2003)1663-1670;camera等,j.nucl.med.21(1994)640-646;ruegg等,cancerres.50(1990)4221-4226;verel等,j.nucl.med.44(2003)1663-1670;lee等,cancerres.61(2001)4474-4482;mitchell等,j.nucl.med.44(2003)1105-1112;kobayashi等bioconjugatechem.10(1999)103-111;miederer等,j.nucl.med.45(2004)129-137;denardo等,clinicalcancerresearch4(1998)2483-90;blend等,cancerbiotherapy&radiopharmaceuticals18(2003)355-363;nikula等j.nucl.med.40(1999)166-76;kobayashi等,j.nucl.med.39(1998)829-36;mardirossian等,nucl.med.biol.20(1993)65-74;roselli等,cancerbiotherapy&radiopharmaceuticals,14(1999)209-20)。
(b)荧光标记,如稀土螯合剂(铕螯合剂);荧光素类型,包括fitc、5-羧基荧光素、6-羧基荧光素;罗丹明类型,包括tamra;丹磺酰;丽丝胺;花菁;藻红蛋白;德克萨斯红;及其类似物。可以用例如currentprotocolsinimmunology,上文中所公开的技术将荧光标记缀合至抗原(半抗原)。荧光染料和荧光标记试剂包括可从invitrogen/molecularprobes(eugene,oregon,usa)和piercebiotechnology,inc.(rockford,ill.)购得的那些。
检测标记如荧光染料和化学发光染料(briggs等"synthesisoffunctionalisedfluorescentdyesandtheircouplingtoaminesandaminoacids,"j.chem.soc.,perkin-trans.1(1997)1051-1058)提供了可检测信号,通常可用于标记,尤其是具有以下特性:(i)标记的缀合物应以低背景产生非常高的信号,使得可以在无细胞和基于细胞的测定二者中选择性检测小量缀合物;和(ii)标记的缀合物应光稳定,使得可以观察、监测和记录荧光信号而无显著的光漂白。对于涉及标记的缀合物与膜或细胞表面(尤其是活细胞)的细胞表面结合的应用,标记应(iii)具有良好的水溶性,以达到有效缀合浓度和检测灵敏度;和(iv)对活细胞无毒性,以便不破坏细胞的正常代谢过程或引起细胞提前死亡。
(c)多种酶-底物标记可用或已公开(参见例如us4,275,149)。酶通常催化可以用多种技术测量的生色底物的化学改变。例如,酶可以在底物中催化可通过分光光度计测量的颜色变化。备选地,酶可以改变底物的荧光或化学发光。化学发光底物通过化学反应变为电子激发态,然后可以发射可以测量(用例如化学发光计)的光或贡献能量给荧光受体。酶标记的实例包括萤光素酶(例如萤火虫萤光素酶和细菌萤光素酶;us4,737,456)、荧光素、2,3-二氢酞嗪二酮、苹果酸脱氢酶、脲酶、过氧化物酶如辣根过氧化物酶(hrp)、碱性磷酸酶(ap)、3-半乳糖苷酶、葡糖淀粉酶、溶菌酶、糖氧化酶(例如葡萄糖氧化酶、半乳糖氧化酶和葡萄糖-6-磷酸脱氢酶)、杂环氧化酶(如尿酸氧化酶和黄嘌呤氧化酶)、乳过氧化物酶、微过氧化物酶体等。methodsinenzym.(j.langone&itvanvunakis编辑),academicpress,newyork,73(1981)147-166中的o'sullivan等"methodsforthepreparationofenzyme-antibodyconjugatesforuseinenzymeimmunoassay"中描述了用于将酶缀合至多肽的技术。
酶-底物组合的实例(us4,275,149;us4,318,980)包括,例如:
(i)以过氧化氢酶为底物的辣根过氧化物酶(hrp),其中过氧化氢酶氧化染料前体(例如邻苯二胺(opd)或3,3',5,5'-四甲基联苯胺盐酸盐(tmb));
(ii)以对硝基苯磷酸酯为生色底物的碱性磷酸酶(ap);和
(iii)具有生色底物(例如对硝基苯基-(3-d-半乳糖苷酶)或荧光底物(4-甲基伞形基-(3-d-半乳糖苷酶))的3-d-半乳糖苷酶((3-d-gal)。
本文报道的标记缀合物可以用于任何已知的测定方法,如elisa、竞争结合测定、直接和间接夹心测定和免疫沉淀测定(zola,monoclonalantibodies:amanualoftechniques(1987)第147-158页,crcpress,inc.)。
本文报道的标记缀合物可用作多种生物医学和分子成像方法和技术的成像生物标志和探针,如:(i)mri(磁共振成像);(ii)microct(计算机化断层照相);(iii)spect(单光子发射计算机化断层照相);(iv)pet(正电子发射断层照相)tinianow,j.等,nuclearmedicineandbiology,37(3)(2010)289-297;chen等,bioconjugatechem.15(2004)41-49;us2010/0111856;(v)生物发光;(vi)荧光;和(vii)超声。免疫闪烁显像是成像方法,对动物或人患者施用放射性物质标记的缀合物,并采集缀合物在体内定位部位的图像(us6,528,624)。成像生物标志可以作为正常生物学过程、致病过程或对治疗干预的药理学反应的指示物客观地测量和评价。生物标志可以属于几种类型:0型标志是疾病的天然历史标志,与已知的临床指数纵向相关,例如类风湿性关节炎中滑液炎症的mri评估;i型标志按照作用机制捕获干预的作用,即使该机制可与临床结果不相关;ii型标志作为替代终点发挥作用,其中生物标志的变化或来自生物标志的信号预测临床收益来“验证”目标反应,如类风湿性关节炎中通过ct测量的骨侵蚀。成像生物标志因此可以提供关于以下的药效学(pd)治疗信息:(i)靶蛋白的表达;(ii)治疗剂与靶蛋白的结合,即选择性;和(iii)清除率和半衰期药代动力学数据。体内成像生物标志相对于基于实验室的生物标志的优势是:非侵入性处理、可定量、整体评估、重复给药和评估(即多个时间点)及从临床前(小动物)至临床(人)结果的潜在可转移效应。对于一些应用,生物成像取代了临床前研究中的动物实验或使临床前研究中的动物实验数目最小化。
肽标记方法众所周知。参见haugland(2003)molecularprobeshandbookoffluorescentprobesandresearchchemicals,molecularprobes,inc.;brinkley(1992)bioconjugatechem.3:2;garman,(1997)non-radioactivelabeling:apracticalapproach,academicpress,london;means(1990)bioconjugatechem.1:2;glazer等chemicalmodificationofproteins.laboratorytechniquesinbiochemistryandmolecularbiology(t.s.work和e.work编辑)americanelsevierpublishingco.,newyork;lundblad,r.l.和noyes,c.m.(1984)chemicalreagentsforproteinmodification,i和ii卷,crcpress,newyork;pfleiderer,g.(1985)"chemicalmodificationofproteins",modernmethodsinproteinchemistry,h.tschesche编辑,walterdegruyter,berlin和newyork;及wong(1991)chemistryofproteinconjugationandcross-linking,crcpress,bocaraton,fla.);deleon-rodriguez等,chem.eur.j.10(2004)1149-1155;lewis等,bioconjugatechem.12(2001)320-324;li等,bioconjugatechem.13(2002)110-115;mier等bioconjugatechem.16(2005)240-237。
接头
术语“接头”指可以用于将第一部分与第二部分缀合(连接)的双功能或多功能部分。可以用具有两种反应性功能的接头方便地制备连接的缀合物。
在一个实施方案中,接头具有这样的反应性部位,该反应性部位具有可与存在于分选酶氨基酸序列中的亲核基团反应的亲电子基团。有用的亲电子基团包括但不限于另一巯基、马来酰亚胺和卤乙酰胺基团(参加例如klussman等,bioconjugatechemistry15(4)(2004)765-773的第766页中的缀合方法)。
巯基反应官能团的实例包括但不限于巯基、马来酰亚胺、α-卤乙酰基、活化酯(如琥珀酰亚胺酯、4-硝基苯酯、五氟苯酯、四氟苯酯)、酐、酰基氯、磺酰氯、异氰酸酯和异硫氰酸酯。
接头可以包含将分选酶氨基酸序列连接至非分选酶基序部分的氨基酸残基。该氨基酸残基可以形成二肽、三肽、四肽、五肽、六肽、七肽、八肽、九肽、十肽、十一肽或十二肽单位。氨基酸残基包括那些天然存在的,以及非天然存在的氨基酸类似物,例如瓜氨酸或β-氨基酸,例如β-丙氨酸,或ω-氨基酸,如4-氨基丁酸。
在另一实施方案中,该接头具有这样的反应性官能团,该官能团具有可与存在于分选酶氨基酸序列中的亲电子基团反应的亲核基团。有用的亲电子基团包括但不限于醛和酮羰基。接头的亲核基团的杂原子可以与分选酶氨基酸序列中的亲电子基团反应,与分选酶氨基酸序列形成共价键。接头上有用的亲核基团包括但不限于酰肼、肟、氨基、肼、缩氨基硫脲、肼羧酸和芳基酰肼。抗原(半抗原)上的亲核基团提供了用于附着至接头的方便位点。
通常,可以通过在两个或多个氨基酸和/或肽片段之间形成肽键来制备肽型接头。这类肽键可以例如按照肽化学领域公知的液相合成法(e.schroder和k.lubke"thepeptides",1卷(1965)76-136,academicpress)制备。
在另一实施方案中,可以用调节溶解性或反应性的基团取代该接头。例如,带电荷的取代基如磺酸(so3-)或铵或聚合物如peg可以提高试剂的水溶解性,便于接头试剂与抗原(半抗原)或药物部分的偶联反应,或便于取决于所利用的合成途径的偶联反应。
本文报道的包含非分选酶基序部分的缀合物明确考虑但不限于用以下接头试剂制备的复合物:bmpeo、bmps、emcs、gmbs、hbvs、lc-smcc、mbs、mpbh、sbap、sia、siab、smcc、smpb、smph、磺基-emcs、磺基-gmbs、磺基-kmus、磺基-mbs、磺基-siab、磺基-smcc、磺基-smpb和svsb(琥珀酰亚胺基-(4-乙烯砜)苯甲酸酯),且包括双马来酰亚胺试剂:dtme、bmb、bmdb、bmh、bmoe、bm(peo)3和bm(peo)4,其可从piercebiotechnology,inc.购得。双马来酰亚胺试剂允许以顺次或同时的方式将例如巯基附着至含巯基的药物部分、标记或接头中间体。除马来酰亚胺外可与例如巯基反应的其他官能团包括碘乙酰胺、溴乙酰胺、乙烯基吡啶、二硫化物、吡啶基二硫化物、异氰酸酯和异硫氰酸酯。
示例性接头包括具有马来酰亚胺延伸基团(stretcher)和对-氨基苄基甲氨酰(pab)自降解间隔基团的缬氨酸-瓜氨酸(val-cit或vc)二肽接头试剂,及具有马来酰亚胺延伸单元和对-氨基苄基自降解间隔基团的phe-lys(mtr)二肽接头试剂。
半胱氨酸巯基是亲核基团,能够与接头试剂和非分选酶基序部分或分选酶氨基酸序列上包括以下的亲电子基团反应形成共价键:(i)活性酯,如nhs酯、hobt酯、卤甲酸和酰基卤;(ii)烷基和苄基卤,如卤乙酰胺;(iii)醛、酮、羧基和马来酰亚胺基团;和(iv)二硫化物,包括吡啶基二硫化物,经硫化物交换。半抗原化化合物上的亲核基团包括但不限于:能够与接头部分和接头试剂上的亲电子基团反应形成共价键的胺、巯基、羟基、酰肼、肟、肼、缩氨基硫脲、肼羧酸和芳基酰肼基团。
附图简述
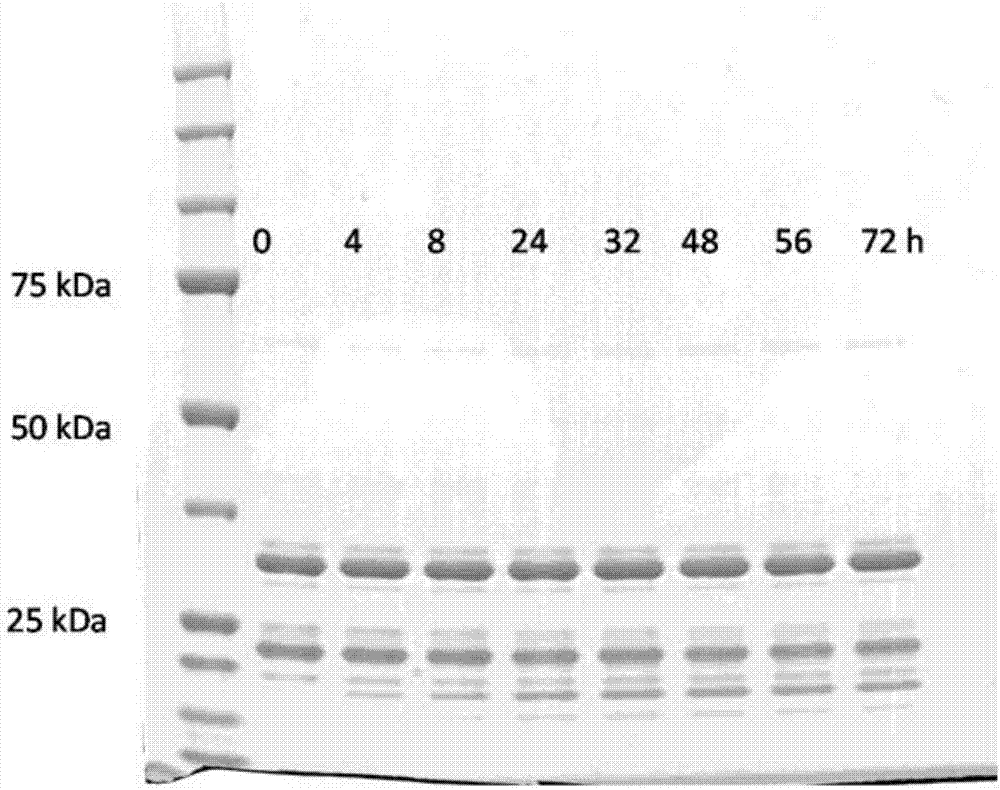
图1(a)至(h):实施例3实验1至8的sds-page分析。
图2实施例1实验1、7和8的包含三种多肽的缀合物的形成时程;棱形:实验1;方形:实验7;三角形:实验8;y轴:按照分选酶量归一化的两次缀合产物的量。
提供以下实施例、附图和序列来辅助理解本发明,本发明的真正范围在所附权利要求书中给出。应理解,可在所示方法中进行修改而不背离本发明的精神。
虽然为了理解的清晰性目的以说明和实例的方式较为详细地描述了前述发明,但这些描述和实例不应解释为限制本发明的范围。
实施例
重组dna技术
按sambrook,j.等,molecularcloning:alaboratorymanual;coldspringharborlaboratorypress,coldspringharbor,纽约,1989中所述用标准方法来操作dna。分子生物学试剂按厂家说明书使用。
基因和寡核苷酸合成
在geneartgmbh(regensburg,德国)通过化学合成制备希望得到的基因区段。将合成的基因片段克隆入大肠杆菌质粒进行繁殖/扩增。通过dna测序验证亚克隆的基因片段的dna序列。备选地,通过退火化学合成的寡核苷酸或通过pcr组装短的合成dna片段。各寡核苷酸由metabiongmbh(planegg-martinsried,德国)制备。
基本/标准哺乳动物表达质粒描述
为了表达希望得到的基因/蛋白质(例如全长抗体重链、全长抗体轻链或在其n端含有寡聚甘氨酸的fc链),使用含有以下功能元件的转录单位:
-包含内含子a的来自人巨细胞病毒的立即早期增强子和启动子(p-cmv);
-人重链免疫球蛋白5’非翻译区(5’utr);
-鼠免疫球蛋白重链信号序列;
-待表达的基因/蛋白质(例如全长抗体重链);和
-牛生长激素多腺苷酸化序列(bghpa)。
除包含所希望的待表达基因的表达单元/盒外,基本/标准哺乳动物表达质粒包含:
-允许此质粒在大肠杆菌中复制的来自载体puc18的复制起点;和
-在大肠杆菌中赋予氨苄青霉素抗性的β-内酰胺酶基因。
蛋白质测定
使用根据多肽氨基酸序列计算的摩尔消光系数,通过测定280nm光密度(od)来测定纯化多肽的蛋白质浓度。
实施例1
可溶性金黄色葡萄球菌分选酶a表达质粒的产生
分选酶基因编码n端截短的金黄色葡萄球菌分选酶a分子(氨基酸序列seqidno:21)。
除可溶性分选酶表达盒外,用于在大肠杆菌细胞中表达可溶性分选酶的表达质粒包含允许此质粒在大肠杆菌中复制的来自载体puc18的复制起点、作为选择标记的ura3基因、及允许用iptg诱导转录的laci基因。
可溶性分选酶的转录单位包含以下功能元件:
-t5启动子;
-纯化标记;
-n端截短的金黄色葡萄球菌分选酶a编码核酸;和
-to和fd终止序列。
源自金黄色葡萄球菌的成熟可溶性分选酶a的氨基酸序列是:
qakpqipkdkskvagyieipdadikepvypgpatpeqlnrgvsfaeeneslddqnisiaghtfidrpnyqftnlkaakkgsmvyfkvgnetrkykmtsirdvkptdvgvldeqkgkdkqltlitcddynektgvwekrkifvatevk(seqidno:21).。
源自动物酿脓链球菌的成熟可溶性分选酶a的氨基酸序列是:
vlqaqmaaqqlpviggiaipelginlpifkglgnteliygagtmkeeqvmggennyslashhifgitgssqmlfspleraqngmsiyltdkekiyeyiikdvftvapervdviddtaglkevtlvtctdieateriivkgelkteydfdkapadvlkafnhsynqvst(seqidno:34)。
纯化标记具有氨基酸序列mrgshhhhhhgs(seqidno:36)。
实施例2
分选酶的表达和分析表征
通过在37℃培养用各分选酶表达质粒转化的大肠杆菌细胞至od578为约0.9来进行可溶性分选酶的重组产生。在od578为约0.9时,通过加入2mmiptg并在28℃再培养细胞24小时来诱导蛋白质表达。然后,通过离心收集细胞,用匀浆器高压裂解。离心细胞裂解物以去除细胞碎片,然后低温(例如-80℃)保存细胞裂解物直至纯化。ni-nta层析后进行大小排阻层析来纯化可溶性分选酶。为了去除内毒素,以流穿模式进行阴离子交换层析。使用根据氨基酸序列计算的摩尔消光系数,通过测量280nm光密度(od)来测定分选酶制备物的蛋白质浓度。通过存在和缺乏还原剂(5mm1,4-二硫苏糖醇)情况下的sds-page及考马斯亮蓝染色来测定分选酶的纯度和完整性。
使用根据氨基酸序列计算的摩尔消光系数,通过测量280nm光密度(od)来测定蛋白质浓度。通过存在和缺乏还原剂(5mm1,4-二硫苏糖醇)情况下的sds-page及考马斯亮蓝染色来分析纯度。
实施例3
c端修饰的f区多肽表达质粒的产生
除fc区表达盒外,用于在hek293细胞中瞬时表达c端修饰的fc区多肽(lpxtg或lpxta)的表达质粒包含允许此质粒在大肠杆菌中复制的来自载体puc18的复制起点,及在大肠杆菌中赋予氨苄青霉素抗性的β-内酰胺酶基因。
fc区多肽的转录单位包含以下功能元件:
-包含内含子a的来自人巨细胞病毒的立即早期增强子和启动子(p-cmv);
-人重链免疫球蛋白5’非翻译区(5’utr);
-鼠免疫球蛋白重链信号序列;
-n端截短的金黄色葡萄球菌分选酶a编码核酸;和
-牛生长激素多腺苷酸化序列(bghpa)。
实施例4
fc区多肽的瞬时表达
通过瞬时转染培养在f17培养基(invitrogencorp.)中的hek293细胞(人胚肾细胞系293衍生)来进行重组产生。对于转染,使用“293-fectin”转染试剂(invitrogen)。转染按照厂家说明书中的说明进行。转染后三至七(3-7)天收集细胞培养上清。将上清保存于低温(例如-80℃)。
meissner,p.等,biotechnol.bioeng.75(2001)197-203中给出了关于在例如hek293细胞中重组表达人免疫球蛋白的一般信息。
使用根据氨基酸序列计算的摩尔消光系数,通过测量280nm光密度(od)来测定蛋白质浓度。通过存在和缺乏还原剂(5mm1,4-二硫苏糖醇)情况下的sds-page和考马斯亮蓝染色来分析纯度。
实施例5
分选酶介导的同时缀合
对于分选酶介导的转肽反应,使用n端截短的金黄色葡萄球菌分选酶a(△α1-59srtastaph)和n端截短的动物酿脓链球菌分选酶a(△1-59srtastep)。
反应在包含50mmtris-hcl、150mmnacl、ph7.5的缓冲液(分选酶缓冲液)中进行。
所利用的多肽是:
(1)在其c端具有氨基酸序列lpetg(seqidno:30)的fc区多肽;
(2)在其c端具有氨基酸序列lpeta(seqidno:32)的fc区多肽;
(3)具有氨基酸序列h2n-ggg-gsgsk(gsgs-aaa-nh2)-cooh(seqidno:33)的具有两个n端的多肽。
测试了以下多肽/酶组合、浓度和序列:
-:未向反应混合物中加入此化合物
(24h):在反应开始24小时后加入此化合物
(*):由于第二srta/第二fc区多肽的体积变化和由于取样造成的总体积减少,浓度随二n端多肽浓度的降低而降低(与其中在开始时混合所有成分的反应相比)。
0、4、8、24、32、48、56和72小时后从各实验取样,并通过sds-page凝胶电泳分析(2μl样品、1μldtt、2.5μllds、4.5μlh2o,70℃10分钟)。预期分子量对srta为约22kda,对fc区多肽和单缀合产物(+1106kda)为约30kda,对包含全部三个部分的缀合产物为约60kda。结果显示在图1中。
可以看到,可以实现两种fc区多肽与二n端多肽的同时缀合。在开始反应时混合所有化合物(多肽和酶)的情况下可获得最佳结果。还可以看到,反应混合物中存在或缺乏钙的影响可忽略。还可以看到,使用等摩尔量的所有三种多肽产生更高的两次缀合产物产率,同时单缀合产物的量降低。还可以看到,如果按等摩尔浓度利用多肽,则浓度越高,产率越高。在实验8中,产率线性提高直至72小时反应时间,且似乎在约100小时达到平台。
实施例6
双缀合物的纯化
使用hiload16/600superdex200制备级柱(gehealthcare,目录号28-9893-35)和含有0.15mnacl的0.05mnapo4缓冲液ph7.2,通过大小排阻层析从反应混合物纯化双缀合物。样品体积0.3ml,上样环体积2.0ml。从20%柱体积后开始0.5μl级分的收集。
- 还没有人留言评论。精彩留言会获得点赞!
- 一种新的多肽——人内皮单细胞活化多肽ii-62和编码这种多肽的多核苷酸的制作方法
- 一种新的多肽——人U4/U6核内小RNA结合蛋白(U4/U6 sn RNP)13.09和编码这种多肽的 ...的制作方法
- 一种新的多肽——人sirtuin type3(SIRT3)蛋白30.91和编码这种多肽的多核苷酸的制作方法
- 一种新的多肽——花生四烯酸环氧酶42.46和编码这种多肽的多核苷酸的制作方法
- 一种新的多肽——人atp酶
- 一种新的多肽——人转录因子MEF2C(hM EF2C)11.11和编码这种多肽的多核苷酸的制作方法
- 一种梅鱼酶解多肽及其应用的制作方法
- 一种新的多肽——人an1相似蛋白16和编码这种多肽的多核苷酸的制作方法
- 突变体羟基苯丙酮酸双加氧酶多肽及其使用方法
- 贻贝酶解多肽及其制备方法和应用的制作方法
- 一种新的多肽——Ras GTP 酶激活蛋白10.34和编码这种多肽的多核苷酸的制作方法
- 一种新的多肽——Ras GTP酶激活蛋白11.33和编码这种多肽的多核苷酸的制作方法
- 一种新的多肽——组氨酰-tRNA合成酶类似蛋白12.21和编码这种多肽的多核苷酸的制作方法
- 一种新的多肽——Ras GTP 酶激活蛋白11.22和编码这种多肽的多核苷酸的制作方法
- 一种新的多肽——ATP合成酶alpha亚基24.97和编码这种多肽的多核苷酸的制作方法
- 一种新的多肽——信号肽酶9.02和编码这种多肽的多核苷酸的制作方法
- 一种新的多肽——Ras GTP酶激活蛋白18和编码这种多肽的多核苷酸的制作方法
- 一种新的多肽——Ras GTP酶激活蛋白10和编码这种多肽的多核苷酸的制作方法
- 一种新的多肽-β内酰胺酶11和编码这种多肽的多核苷酸的制作方法
- 蜈蚣酶解物抗血栓性多肽的制作方法