用于生产具有新功能的淀粉的组合物和方法与流程
相关申请的交叉参考本申请是2016年4月5日提交的美国申请号15/090,926的延续,其公开内容出于所有目的经此引用全文并入本文。本发明涉及衍生自植物和谷物的玉米淀粉、用于制造这样的淀粉的组合物和方法。
背景技术:
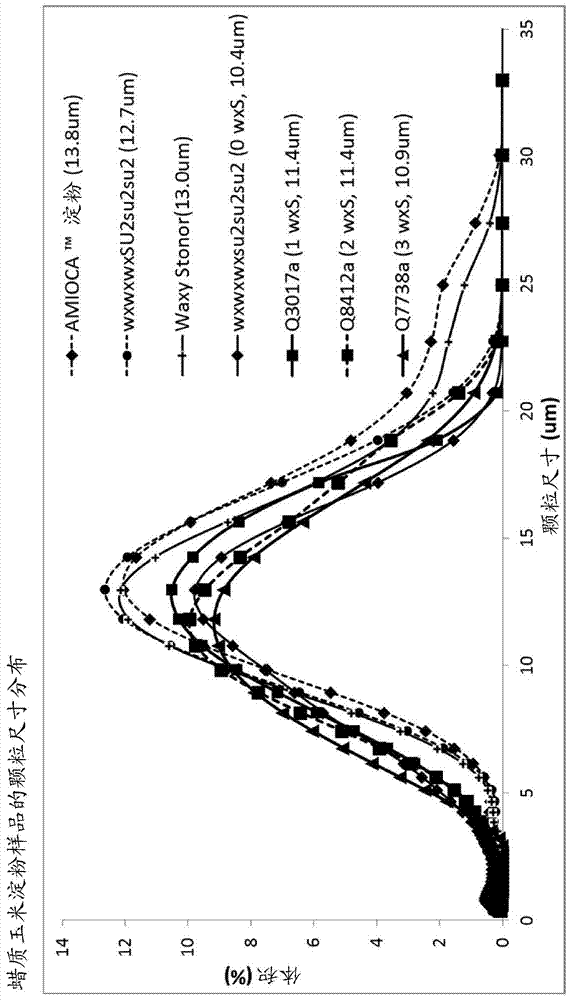
:现有的waxy/sugary-2双突变玉米淀粉显示出非常良好的冻/融稳定性。但是,现有的waxy/sugary-2玉米淀粉的粘度显著低于商业蜡质(waxy)玉米淀粉,如amioca™淀粉。此外,现有的waxy/sugary-2双突变玉米淀粉的糊化温度显著低于amioca™淀粉。因此,在本领域中仍然需要生产具有良好的冻/融稳定性、提高的粘度和/或更高的糊化温度的玉米淀粉。在本发明中,我们通过向现有的waxy/sugary-2双突变玉米中引入低活性的gbssi来开发了具有比商业蜡质玉米淀粉,如amioca™淀粉更高的粘度和/或更高的糊化温度的新型玉米淀粉。技术实现要素:本发明提供了在自然界中从未发现的具有新功能的淀粉。在一些实施方案中,使用直链淀粉/支链淀粉测定试剂盒,如megazyme®,该淀粉包含大约2重量%至大约20重量%的直链淀粉含量。在一些实施方案中,如使用快速粘度分析仪(rva,newportscientific,sydney,australia)测得的,该淀粉具有比对照淀粉的淀粉糊化温度高至少大约5%的水淀粉糊化温度。在一些实施方案中,该淀粉衍生自在胚乳中包含至少一个wx-stonor(wxs)等位基因的waxy/sugary-2双突变玉米植物。在一些实施方案中,该waxy/sugary-2双突变玉米植物包含隐性su2突变等位基因。在一些实施方案中,本发明的淀粉由对于产生所公开和要求保护的淀粉的基因型不是转基因(即非gmo)的植物生产。在一些实施方案中,该淀粉的直链淀粉含量为大约8重量%至大约15重量%。在一些实施方案中,该水糊化温度比对照淀粉的水糊化温度高大约5%至大约15%。在一些实施方案中,该淀粉进一步具有结晶焓变。在一些实施方案中,如使用差示扫描量热计(dsc)测得的,该结晶焓变比对照淀粉的结晶焓变高至少大约30%。在一些实施方案中,如使用dsc测得的,该结晶焓变比对照淀粉的结晶焓变高大约30%至大约200%。在一些实施方案中,该淀粉进一步具有淀粉颗粒尺寸的分布,如使用扫描电子显微镜观察到的,其中淀粉颗粒尺寸的分布包含与对照淀粉中小于5微米的颗粒的比例相比少至少大约40%的小于5微米的颗粒。在一些实施方案中,如使用扫描电子显微镜观察到的,该淀粉具有与衍生自对照玉米植物的淀粉相比表现出提高的完整性的颗粒。在一些实施方案中,该淀粉能够承受至少三次冻融循环。在一些实施方案中,该淀粉能够承受至少三次到至少五次冻融循环。在一些实施方案中,制造本发明的淀粉的植物通过具有wxwxsu2su2的基因型的第一玉米植物与具有wxswxssu2su2的基因型或wxswxsu2su2的基因型的第二玉米植物的异花传粉来产生。在一些实施方案中,该waxy/sugary-2玉米植物的胚乳具有1剂量、2剂量或3剂量的wxs等位基因,并且该胚乳具有wxswxwxsu2su2su2、wxswxswxsu2su2su2或wxswxswxssu2su2su2的基因型。本发明还提供了制造具有改善的糊化曲线的淀粉的方法。在一些实施方案中,该方法包括向蜡质玉米植物的胚乳中引入至少一个wxs等位基因。本发明还提供了新的双突变植物。在一些实施方案中,该植物是玉米植物。在一些实施方案中,该植物对淀粉合成酶iia(su2)基因是纯合隐性的。在一些实施方案中,该植物对突变的颗粒结合淀粉合成酶i(gbssi)基因是纯合或杂合的。在一些实施方案中,该突变的gbssi基因具有比野生型野生型gbssi基因(wx)更低的gbssi活性,但是具有比隐性的功能缺失的gbssi基因(wx)更高的gbssi活性。在一些实施方案中,该突变的gbssi基因具有大约8%至大约10%的野生型gbssi基因的gbssi活性。在一些实施方案中,该突变的gbssi基因是wx-stonor(wxs)等位基因。在一些实施方案中,该突变的gbssi基因是wx-g等位基因、wx-b5等位基因或wx-m等位基因。在一些实施方案中,该双突变植物通过两个亲本植物的异花传粉来产生。在一些实施方案中,该第一植物具有wxwxsu2su2的基因型。在一些实施方案中,该第二植物具有wxswxssu2su2的基因型或wxswxsu2su2的基因型。在一些实施方案中,该植物的胚乳具有1剂量、2剂量或3剂量的wxs等位基因。在一些实施方案中,该植物的胚乳具有wxswxwxsu2su2su2、wxswxswxsu2su2su2或wxswxswxssu2su2su2的基因型。在一些实施方案中,在该植物的胚乳中产生的淀粉与具有wxwxwxsu2su2su2的基因型的双waxy/sugary-2隐性突变植物的胚乳中产生的淀粉相比具有更高的粘度。在一些实施方案中,在该植物的胚乳中产生的淀粉与具有wxwxwxsu2su2su2的基因型的双waxy/sugary-2隐性突变植物的胚乳中产生的淀粉相比具有更高的淀粉糊化温度。本发明还提供了植物部分、植物细胞和植物的组织培养物。在一些实施方案中,该植物部分是胚乳、叶、花、胚珠、花粉、根茎或接穗。在一些实施方案中,该植物部分是植物的种子。本发明还提供了从上述植物中提取的淀粉。本发明还提供了制造淀粉的方法。在一些实施方案中,该方法包括从本发明的植物的胚乳中获得淀粉。在一些实施方案中,制造淀粉的方法包括种植本发明的植物,并从该植物收获种子。本发明还提供了产生种子的方法。在一些实施方案中,该方法包括使第一亲本植物与第二亲本植物杂交并收获所得种子。在一些实施方案中,所述第一亲本植物和/或第二亲本植物是本发明的植物。在一些实施方案中,该方法包括使本发明的植物自花传粉,并收获所得种子。本发明还提供了无性繁殖该植物的方法。在一些实施方案中,该方法包括收集该植物的部分并由所述部分再生植物。本发明还提供了产生衍生自本发明的植物的植物的方法。在一些实施方案中,该方法包括使本发明的植物自花传粉至少一次以制造衍生自该植物的后代植物。在一些实施方案中,该方法包括使本发明的植物与第二植物杂交以制造植物后代。本发明还提供了产生转基因植物的方法。在一些实施方案中,该方法包括将相关转基因引入本发明的植物。所述相关转基因在该植物中赋予一种或多种期望表型。本发明还提供了产生具有期望表型的植物的方法,包括基因编辑以在本发明的植物中制造基因序列。该基因序列在该植物中赋予一种或多种期望表型。本发明还提供了产生具有期望表型的植物的方法,包括在本发明的植物中诱导一个或多个突变以产生基因序列。该基因序列在该植物中赋予一种或多种期望表型。附图说明图1描述了在加热-冷却循环过程中随时间推移通过快速粘度分析仪(rva)测得的amioca™淀粉、wxwxwxsu2su2su2淀粉和具有0、1、2或3剂量的wxs基因的waxy/sugary-2双突变植物的淀粉的糊化曲线。图2描述了amioca™淀粉、wxwxwxsu2su2su2淀粉和具有0、1、2或3剂量的wxs基因的waxy/sugary-2双突变植物的淀粉的糊化性质,包括糊化温度、峰值rvu、保持rvu、最终rvu、崩解rvu和消减rvu。图3描述了具有1、2、3或0剂量的wxs基因的waxy/sugary-2双突变植物的淀粉、来自waxystonor种子的淀粉、wxwxwxsu2su2su2淀粉和amioca™淀粉的冻-融稳定性测试。上述淀粉以行呈现,并在第一次冻融(循环1)之后、在第三次冻融(循环3)之后和在第五次冻融(循环5)之后提供了淀粉凝胶的说明性图像。图4描述了amioca™淀粉、wxwxwxsu2su2su2淀粉和具有0、1、2或3剂量的wxs基因的waxy/sugary-2双突变植物的淀粉的热性质。图5描述了amioca™淀粉、wxwxwxsu2su2su2淀粉和具有0、1、2或3剂量的wxs基因的waxy/sugary-2双突变植物的淀粉的淀粉颗粒尺寸分布。图6描述了具有1、2、3或0剂量的wxs基因的waxy/sugary-2双突变植物的淀粉、wxwxwxsu2su2su2淀粉、amioca™淀粉和来自waxystonor种子的淀粉的颗粒形态。提供了使用扫描电子显微镜捕获的上述淀粉各自的说明性图像。具体实施方式定义本文中所用的动词“包含”当用于本说明书和权利要求及其结合时以其非限制性含义使用,指的是包括该词之后的项目,但是不排除并未具体提及的项目。本发明提供制造植物的组合物和方法。本文中所用的术语“植物”指的是植物界的任何活生物体(即植物王国中的任何属/种)。在一些实施方案中,能够制造淀粉的植物包括但不限于玉米、马铃薯、小麦、木薯、水稻、马铃薯和高粱。在一些实施方案中,该植物是玉米植物。本发明提供植物部分。本文中所用的术语“植物部分”指的是植物的任何部分,包括但不限于胚芽、胚乳、枝条、根、茎、种子、托叶、叶、花瓣、花、胚珠、苞片、分枝、叶柄、节间、树皮、柔毛、分蘖、根茎、复叶、叶片、胚珠、花粉、雄蕊等等。本发明提供了包含分离的(例如野生型,对该生物体是内源性的)、嵌合的、重组的或合成的核酸序列的基因。本文中所用的术语“基因”指的是与生物功能相关的任何dna片段。由此,基因包括但不限于其表达所需的编码序列和/或调节序列。基因还可以包括非表达的dna片段,例如,形成其它蛋白的识别序列。基因可以获自多种来源,包括从相关来源克隆,或由已知或预测的序列信息合成,并且可以包括设计为具有所需参数的序列。本发明提供了与野生型参照序列相比时具有核苷酸变化的多核苷酸。本文中所用的术语“核苷酸变化”指的是例如核苷酸替换、缺失、插入或转座子,如本领域中充分理解的那样。例如,突变包含产生沉默替换、添加、缺失或可变剪接的改变,这会改变或不改变编码的蛋白的性质或活性或是如何制造该蛋白。核苷酸变化还可以是基因中逆转录转座子元件的插入和/或排除。本发明提供了与野生型参照序列相比时具有蛋白修饰的多肽。本文中所用的术语“蛋白修饰”指的是例如氨基酸替换、氨基酸修饰、缺失、插入或蛋白活性变化,如本领域中充分理解的那样。本发明提供了衍生自野生型参照序列的多核苷酸和多肽。本文中所用的术语“衍生自”指的是起源或来源,并可以包括天然存在的、重组的、未纯化的或纯化的分子。衍生自起源或来源的核酸或氨基酸可以具有如本文中其它地方所定义的所有类型的核苷酸变化或蛋白修饰。本发明提供了本发明的核酸序列和多肽序列的生物活性变体或功能变体。本文中所用的短语“生物活性变体”或“功能变体”对于蛋白质而言指的是相对于参照序列被一个或多个氨基酸改变,同时仍保留参照序列的实质性生物活性的氨基酸序列。该变体可以具有“保守”变化,其中替换的氨基酸具有类似的结构或化学性质,例如用异亮氨酸代替亮氨酸。下面的表显示了示例性的保守氨基酸替换。在一些实施方案中,该变体具有一个或多个氨基酸替换。其中一个或多个或全部替换是酸性氨基酸,如天冬氨酸、天冬酰胺、谷氨酸或谷氨酰胺。原始残基极高度保守的替换高度保守的替换(来自blosum90matrix)保守的替换(来自blosum65matrix)alasergly,ser,thrcys,gly,ser,thr,valarglysgln,his,lysasn,gln,glu,his,lysasngln;hisasp,gln,his,lys,ser,thrarg,asp,gln,glu,his,lys,ser,thraspgluasn,gluasn,gln,glu,sercysser无alaglnasnarg,asn,glu,his,lys,metarg,asn,asp,glu,his,lys,met,sergluaspasp,gln,lysarg,asn,asp,gln,his,lys,serglyproalaala,serhisasn;glnarg,asn,gln,tyrarg,asn,gln,glu,tyrileleu;valleu,met,valleu,met,phe,valleuile;valile,met,phe,valile,met,phe,vallysarg;gln;gluarg,asn,gln,gluarg,asn,gln,glu,ser,metleu;ilegln,ile,leu,valgln,ile,leu,phe,valphemet;leu;tyrleu,trp,tyrile,leu,met,trp,tyrserthrala,asn,thrala,asn,asp,gln,glu,gly,lys,thrthrserala,asn,serala,asn,ser,valtrptyrphe,tyrphe,tyrtyrtrp;phehis,phe,trphis,phe,trpvalile;leuile,leu,metala,ile,leu,met,thr或者,变体可具有“非保守”变化,例如用色氨酸替代甘氨酸。类似的微小变化也可以包括氨基酸缺失和/或插入。确定哪些氨基酸残基可以替换、插入或缺失而不会消除生物或免疫活性的指导可以使用本领域公知的计算机程序(例如dnastar软件)来找到。对于多核苷酸而言,变体包括以下多核苷酸:在5'和/或3'末端处具有缺失(即截短);在参照多核苷酸中在一个或多个内部位点处缺失和/或添加一个或多个核苷酸;和/或在参照多核苷酸中在一个或多个位点处替换一个或多个核苷酸。本文中所用的“参照”多核苷酸包含通过本文中公开的方法制得的核苷酸序列。变体多核苷酸还包括合成衍生的多核苷酸,如例如通过使用定向诱变生成但仍包含基因调节元件活性的那些。通常,本发明的特定多核苷酸或核酸分子,或多肽的变体将具有如通过本文中其它地方描述的序列比对程序和参数所确定的与特定多核苷酸/多肽至少大约60%、65%、70%、75%、80%、85%、90%、91%、91.5%、92%、92.5%、93%、93.5%、94%、94.5%、95%、95.5%、96%、96.5%、97%、97.5%、98%、98.5%、99%、99.1%、99.2%、99.3%、99.4%、99.5%、99.6%、99.7%、99.8%、99.9%或更高的序列同一性。变体多核苷酸还包括衍生自诱变和重组程序(如dna改组)的序列。这样的dna改组的策略是本领域已知的。参见例如stemmer(1994)pnas91:10747-10751;stemmer(1994)nature370:389-391;crameri等人(1997)naturebiotech.15:436-438;moore等人(1997)j.mol.biol.272:336-347;zhang等人(1997)pnas94:4504-4509;crameri等人(1998)nature391:288-291;以及美国专利号5,605,793和5,837,458。对于本文中公开的多核苷酸的pcr扩增,寡核苷酸引物可以设计用于pcr反应以扩增来自从任何相关植物中提取的cdna或基因组dna的相应dna序列。设计pcr引物和pcr克隆的方法通常是本领域已知的,并且公开在sambrook等人(1989)molecularcloning:alaboratorymanual(第2版,coldspringharborlaboratorypress,plainview,newyork)中。还参见innis等人主编的(1990)pcrprotocols:aguidetomethodsandapplications(academicpress,newyork);innis和gelfand主编的(1995)pcrstrategies(academicpress,newyork);以及innis和gelfand主编的(1999)pcrmethodsmanual(academicpress,newyork)。pcr的已知方法包括但不限于使用配对引物、巢式引物、单特异性引物、简并引物、基因特异性引物、载体特异性引物、部分错配引物等等的方法。本发明提供转基因植物。本文中所用的术语“转基因”指的是已经通过各种转化方法之一接受外源或修饰的基因的细胞、细胞培养物、生物体(例如植物)和后代,其中该外源或修饰的基因来自与接受该外源或修饰的基因的生物体的物种相同或不同的物种。waxy基因:waxy基因是颗粒结合淀粉合成酶i(gbssi)基因。该基因编码负责在发育的核的三倍体胚乳中和在单倍体花粉和胚囊中的直链淀粉生物合成的淀粉颗粒结合adp-葡萄糖-葡糖基转移酶(ec2.4.1.242)。“wx”指的是野生型等位基因,并且“wx”指的是无效的参照突变等位基因。术语wxs(又名wx-stonor)、wx-g、wx-b5和wx-m指的是在waxy基因中具有逆转录转座子插入的中间等位基因,如在wessler和varagona(pnas,82:4177-4181,1985)以及varagona等人(theplantcell,4:811-820,1992)中所述,其各自经此引用全文并入本文。这些中间等位基因在具有该等位基因的植物中导致与在野生型植物中的蜡质蛋白(waxyprotein)活性相比较低的蜡质蛋白活性。该玉米waxy基因编码具有seqidno:1的序列的蛋白。wx-stonor:玉米植物中的wx-stonor等位基因由该waxy基因的内含子5的剪接供体位点处的逆转录转座子插入所导致。尽管该插入不会影响剪接机制,其导致与野生型等位基因相比更低水平的酶活性(例如仅为野生型的gbssi活性的大约9.5%)。因此,具有wx-stonor等位基因的植物具有比不具有该wx-stonor等位基因的相同植物更高的直链淀粉水平。sugary-2基因:sugary-2基因编码淀粉合成酶同种型iia(ssiia,ec2.4.1.21)。“su2”指的是野生型等位基因,且“su2”指的是隐性的突变等位基因。sugary-2蛋白在玉米中的功能描述在zhang等人(plantmolecularbiology,54(6):865-879,2004)中,其经此引用全文并入本文。该玉米野生型su2基因编码具有seqidno:2的序列的野生型蛋白。玉米中的waxy基因和sugary-2基因的更多细节描述在美国专利号4428972、5954883、6828474、6960703和7678898中,其各自经此引用全文并入本文。amioca™淀粉:该术语指的是由nationalstarchandchemicalcompany(现为ingredionincorporated)制造的amioca™玉米淀粉。wxwxwxsu2su2su2淀粉:该术语指的是从具有wxwxwxsu2su2su2的基因型的玉米胚乳中分离的淀粉。wxwxwxsu2su2su2淀粉:该术语指的是从具有wxwxwxsu2su2su2的基因型的玉米胚乳中分离的淀粉,其为waxy/sugary-2双隐性突变体的胚乳。该突变的waxy基因有助于其良好的冻-融稳定性,并且突变的sugary-2基因导致短支链长度。但是,该淀粉具有低淀粉颗粒完整性。该颗粒容易崩解,并且该淀粉具有低粘度。melojel®淀粉:该术语指的是由nationalstarchandchemicalcompany(现为ingredionincorporated)制造的melojel®玉米淀粉。对照玉米植物:该术语指的是用于比较的适当的对照植物。在一些实施方案中,对照植物可以是野生型植物。在一些实施方案中,该对照植物是不具有本文中描述的中间waxy基因的植物。在一些实施方案中,该对照植物是制造wxwxwxsu2su2su2淀粉或wxwxwxsu2su2su2淀粉的玉米植物。因此,在一些实施方案中,该对照玉米植物是在胚乳中不具有wx-s等位基因的玉米植物。对照淀粉:该术语指的是对照淀粉。wxswxwxsu2su2su2:该术语指的是具有1剂量的wxs的玉米胚乳。wxswxwxsu2su2su2淀粉:该术语指的是从具有1剂量的wxs的玉米胚乳中分离的淀粉,该胚乳具有基因型wxswxwxsu2su2su2。wxswxswxsu2su2su2:该术语指的是具有2剂量的wxs的玉米胚乳。wxswxswxsu2su2su2淀粉:该术语指的是从具有2剂量的wxs的玉米胚乳中分离的淀粉,该胚乳具有基因型wxswxswxsu2su2su2。wxswxswxssu2su2su2:该术语指的是具有3剂量的wxs的玉米胚乳。wxswxswxssu2su2su2淀粉:该术语指的是从具有3剂量的wxs的玉米胚乳中分离的淀粉,该胚乳具有基因型wxswxswxssu2su2su2。下面的实施例举例说明了本发明的各个方面。当然,该实施例应理解为仅举例说明本发明的某些实施方案,并不对本发明的范围构成限制。具有新功能的淀粉在一个方面,本公开提供了衍生自包含中间gbssi等位基因的双突变玉米植物的淀粉,所述中间gbssi等位基因具有比野生型gbssi基因(wx)更低的gbssi活性,但是具有比隐性的功能缺失的gbssi基因(wx)更高的gbssi活性。在一些实施方案中,具有中间gbssi等位基因的双突变玉米植物是在胚乳中包含至少一个wx-stonor(wxs)等位基因的waxy/sugary-2玉米植物。在一些实施方案中,该waxy/sugary-2玉米植物通过具有wxwxsu2su2的基因型的第一玉米植物与具有wxswxssu2su2的基因型或wxswxsu2su2的基因型的第二玉米植物的异花传粉来产生。在进一步的实施方案中,该waxy/sugary-2玉米植物的胚乳具有1剂量、2剂量或3剂量的wxs等位基因,并且该胚乳具有wxswxwxsu2su2su2、wxswxswxsu2su2su2或wxswxswxssu2su2su2的基因型。在一些实施方案中,该淀粉包含如使用megazyme®直链淀粉/支链淀粉测定试剂盒测得的大约2重量%至大约20重量%的直链淀粉含量,以及如使用rva分析测得的比对照淀粉的糊化温度高至少大约5%的水淀粉糊化温度。直链淀粉含量:淀粉由直链淀粉和支链淀粉组成。直链淀粉是由α-d-葡萄糖单元制成并通过α(1→4)糖苷键连接的螺旋形聚合物。支链淀粉是包含α(1→4)和α(1→6)糖苷键的葡萄糖的高度支化聚合物。在α(1→6)键的情况下发生支化,其出现在大约每24至30个葡萄糖单元处。在一些实施方案中,淀粉中的直链淀粉含量可以使用直链淀粉/支链淀粉测定试剂盒,例如由megazyme®制造的直链淀粉/支链淀粉测定试剂盒测得,如实施例部分所述的那样。观察到双突变玉米植物中gbssi基因的中间突变相对于具有无效等位基因gbssi基因的对照玉米植物提高了衍生自其的淀粉中的直链淀粉含量。例如,观察到衍生自在胚乳中包含至少一个wx-stonor(wxs)等位基因的waxy/sugary-2玉米植物的淀粉的直链淀粉含量相对于对照玉米植物得到了提高。在一些实施方案中,衍生自在胚乳中具有至少一个wx-stonor(wxs)等位基因的玉米植物的淀粉具有大约2重量%至大约20重量%、大约4重量%至大约18重量%、大约5重量%至大约15重量%或大约8重量%至大约15重量%的直链淀粉含量,包括其间的所有值和子范围。在其它实施方案中,该直链淀粉含量为大约5重量%、大约6重量%、大约7重量%、大约8重量%、大约9重量%、大约10重量%、大约11重量%、大约12重量%、大约13重量%、大约14重量%、大约15重量%、大约16重量%、大约17重量%或大约18重量%,包括其间的所有值。不受理论的限制,直链淀粉含量的提高预期将降低该淀粉中的支链淀粉含量,由此降低可用于与水性介质相互作用的葡萄糖单元和氢键合基团的总数。氢键合的这种降低应当对应于所述淀粉与水性介质的相互作用的强度的整体降低,并降低该淀粉的糊化性质。但是,在本文中公开的淀粉中,当直链淀粉含量提高时,观察到糊化曲线中的预料不到的改善。不受理论的限制,这种预料不到的结果可能归因于在胚乳中包含至少一个wx-stonor(wxs)等位基因的waxy/sugary-2玉米植物中观察到的支链长度的提高。在该淀粉上较长的支链具有更多葡萄糖单元,这与较短的支链长度相比提高了与水性介质的氢键合相互作用的数量。淀粉的糊化曲线可以通过已知方法来测量,如使用快速粘度分析仪(rva)(newportscientific,sydney,australia)。例如,rva可以测量糊化温度(℃)、峰值粘度(rvu)、保持粘度(rvu)、最终粘度(rvu)、崩解粘度(rvu)和消减粘度(rvu)。在一些实施方案中,改善的糊化曲线允许凝胶化发生,尤其是在与对照淀粉相比较低的淀粉有效量和/或较高的糊化温度下。淀粉凝胶化指的是在水和热的存在下破坏淀粉分子的分子间键,使得氢键合位点(例如羟基基团)能够结合更多水的过程。为了测量淀粉糊化曲线,可以在中性ph下向水溶液中添加有效量的淀粉。例如,淀粉可以8重量%的固体水平添加到水溶液中。糊化温度:本文中所用的糊化温度旨在如实施例部分中所述的那样测量。淀粉的糊化温度是当悬浮在水中时淀粉颗粒发生初始溶胀时的温度。在一些实施方案中,本文中描述的淀粉具有使用如rva分析测得的比对照淀粉的糊化温度高至少大约5%的水糊化温度。在一些实施方案中,该水糊化温度提高了大约1%至大约100%、大约1%至大约50%、大约5%至大约50%、大约5%至大约40%、大约5%至大约30%、大约5%至大约25%、大约5%至大约20%或大约5%至大约14%,包括其间的所有值和子范围。在特定实施方案中,该水糊化温度比对照玉米植物(如双突变waxy/sugary-2玉米植物)高大约5%至大约14%。在其它实施方案中,与对照淀粉的糊化温度相比,该水糊化温度提高了至少大约2%、至少大约3%、至少大约4%、至少大约5%、至少大约6%、至少大约7%、至少大约8%、至少大约9%、至少大约10%、至少大约11%、至少大约12%、至少大约13%、至少大约14%、至少大约15%、至少大约16%、至少大约17%、至少大约18%、至少大约19%或至少大约20%,包括其间的所有值。在还有的其它实施方案中,该水糊化温度提高了大约1℃至大约20℃、或大约5℃至大约10℃,包括其间的所有值和子范围。在其它实施方案中,该水糊化温度与对照淀粉的糊化温度相比提高了至少大约4℃、至少大约5℃、至少大约6℃、至少大约7℃、至少大约8℃、至少大约9℃、至少大约10℃、至少大约11℃、至少大约12℃、至少大约13℃、至少大约14℃或至少大约15℃。在还有的其它实施方案中,本文中描述的淀粉具有至少大约50℃、至少大约55℃、至少大约60℃、至少大约65℃、至少大约70℃、至少大约75℃、至少大约80℃、至少大约85℃、至少大约90℃、至少大约95℃或至少大约100℃的水糊化温度。在特定实施方案中,衍生自在胚乳中包含至少一个wx-stonor(wxs)等位基因的waxy/sugary-2玉米植物的淀粉具有大约55℃至大约68℃、或大约59℃至大约65℃的水糊化温度,包括其间的所有值和子范围。在其它实施方案中,该水糊化温度为大约59℃、大约60℃、大约61℃、大约62℃、大约63℃、大约64℃、大约65℃、大约66℃、大约67℃或大约68℃,包括其间的所有值。结晶焓变:本文中所用的结晶焓变旨在如实施例部分中所述的那样测量。结晶指的是通过沉淀从溶液中形成固体晶体的过程。在一些实施方案中,观察到在waxy/sugary-2双突变玉米植物中在胚乳中引入至少一个wx-stonor(wxs)等位基因相对于来自对照玉米植物的淀粉提高了该结晶焓变。焓变可以使用例如差示扫描量热法(dsc)来测量。在一些实施方案中,该结晶焓变提高了大约10%至大约300%、大约10%至大约200%、大约10%至大约150%、大约10%至大约100%、或大约10%至大约50%,包括其间的所有值和子范围。在一个这样的实施方案中,该结晶焓变如使用dsc测得的比对照淀粉的结晶焓变高大约70%至大约200%。在一些实施方案中,该结晶焓变提高了至少大约10%、至少大约20%、至少大约30%、至少大约40%、至少大约50%、至少大约60%、至少大约70%、至少大约80%、至少大约90%、至少大约100%、至少大约110%、至少大约120%、至少大约130%、至少大约140%、至少大约150%、至少大约160%、至少大约170%、至少大约180%、至少大约190%或至少大约200%,包括其间的所有值。在一些实施方案中,该结晶焓变为大约1j/g至大约20j/g、大约5j/g至大约15j/g、大约6j/g至大约13j/g,包括其间的所有值和子范围。在其它实施方案中,结晶度8重量%的焓变为大约4j/g、大约5j/g、大约6j/g、大约7j/g、大约8j/g、大约9j/g、大约10j/g、大约11j/g、大约12j/g、大约13j/g,包括其间的所有值。在特定实施方案中,该玉米植物在胚乳中包含一个wxs等位基因,且结晶焓变比衍生自对照植物的淀粉的结晶焓变高至少大约150%,所述对照植物具有wxwxwxsu2su2su2的胚乳基因型。在另一特定实施方案中,该玉米植物在胚乳中包含两个wxs等位基因,且结晶焓变比衍生自对照植物的淀粉的结晶焓变高至少大约30%,所述对照植物具有wxwxwxsu2su2su2的胚乳基因型。在又一特定实施方案中,该玉米植物在胚乳中包含三个wxs等位基因,且结晶焓变比衍生自对照植物的淀粉的结晶焓变高至少大约75%,所述对照植物具有wxwxwxsu2su2su2的胚乳基因型。冻融稳定性:本文中所用的冻融稳定性旨在如实施例部分中所述的那样测量。在一些实施方案中,本文中描述的淀粉表现出期望的冻融特性,例如改善的冻-融稳定性。冻-融稳定性可以通过以下方法测得:将有效量的淀粉添加到水中以形成浆料,随后将其烹煮,并在冷冻/冷却循环后测量脱水收缩。在一些实施方案中,向玉米植物的胚乳中引入至少一个wx-stonor(wxs)等位基因与对照淀粉相比不会降低由其衍生的淀粉的冻-融稳定性。在其它实施方案中,向玉米植物的胚乳中引入至少一个wx-stonor(wxs)等位基因改善了该淀粉的冻-融稳定性。在一个这样的实施方案中,该淀粉能够承受总计至少三次到至少五次冻融循环。颗粒分布与完整性:本文中所用的颗粒分布与完整性旨在如实施例部分中所述的那样测量。在一些实施方案中,观察到在玉米植物的胚乳中引入至少一个wx-stonor(wxs)等位基因相对于对照淀粉改变了淀粉颗粒的尺寸分布。在一些这样的实施方案中,如使用扫描电子显微镜观察到的,本文中描述的淀粉的淀粉颗粒尺寸的分布包含与对照淀粉中小于5微米的颗粒的比例相比少至少大约40%的小于5微米的颗粒。本文中所用的颗粒完整性指的是该淀粉在溶于水性介质之后保持为完整颗粒的能力。在一些实施方案中,如使用扫描电子显微镜观察到的,本文中描述的淀粉是与衍生自对照玉米植物的淀粉相比表现出提高的完整性的颗粒。粘度:本文中所用的粘度旨在如实施例部分中所述的那样测量。在一些实施方案中,本文中描述的淀粉具有如使用rva分析测得的比对照淀粉的水峰值粘度高至少大约10%的水峰值粘度。在一些实施方案中,该水峰值粘度提高了大约10%至大约300%、大约10%至大约250%、大约10%至大约200%、大约10%至大约150%、大约10%至大约100%或大约10%至大约50%,包括其间的所有值和子范围。在一些实施方案中,该水峰值粘度提高了至少大约10%、至少大约20%、至少大约30%、至少大约40%、至少大约50%、至少大约60%至少大约70%、至少大约80%、至少大约90%、至少大约100%、至少大约110%、至少大约120%、至少大约130%、至少大约140%、至少大约150%、至少大约160%、至少大约170%、至少大约180%、至少大约190%或至少大约200%,包括其间的所有值。在一些实施方案中,在8重量%固体下该淀粉的水峰值粘度为大约100rvu至大约500rvu、大约100rvu至大约400rvu、大约100rvu至大约300rvu、大约100rvu至大约200rvu或大约100rvu至大约150rvu,包括其间的所有值和子范围。在一些实施方案中,该水峰值粘度为大约150rvu、大约155rvu、大约160rvu、大约165rvu、大约170rvu、大约175rvu、大约180rvu、大约185rvu、大约190rvu、大约195rvu、大约200rvu、大约205rvu、大约210rvu、大约215rvu、大约220rvu、大约225rvu、大约230rvu、大约235rvu、大约240rvu、大约245rvu、大约250rvu、大约255rvu、大约260rvu、大约265rvu、大约270rvu、大约275rvu、大约280rvu、大约285rvu、大约290rvu、大约295rvu、大约300rvu、大约305rvu、大约310rvu、大约315rvu、大约320rvu、大约325rvu、大约330rvu、大约335rvu、大约340rvu、大约345rvu、大约350rvu、大约355rvu、大约360rvu、大约365rvu、大约370rvu、大约375rvu、大约380rvu、大约385rvu、大约390rvu、大约395rvu或大约400rvu,包括其间的所有值。在特定实施方案中,该waxy/sugary-2玉米植物在胚乳中包含一个wx-stonor(wxs)等位基因,且该淀粉具有比具有wxwxwxsu2su2su2的胚乳基因型的对照植物的淀粉高大约110%的峰值粘度。在另一特定实施方案中,该waxy/sugary-2玉米植物在胚乳中包含两个wx-stonor(wxs)等位基因,且该淀粉具有比具有wxwxwxsu2su2su2的胚乳基因型的对照植物的淀粉高大约30%的峰值粘度。在又一特定实施方案中,该waxy/sugary-2玉米植物在胚乳中包含三个wx-stonor(wxs)等位基因,且该淀粉具有比具有wxwxwxsu2su2su2的胚乳基因型的对照植物的淀粉高大约40%的峰值粘度。在甚至另一特定实施方案中,该waxy/sugary-2玉米植物在胚乳中包含两个wx-stonor(wxs)等位基因,且该淀粉具有比具有wxwxwxsu2su2su2的胚乳基因型的对照植物的淀粉高大约20%的峰值粘度。在又一特定实施方案中,该waxy/sugary-2玉米植物在胚乳中包含两个wx-stonor(wxs)等位基因,且该淀粉具有比具有wxwxwxsu2su2su2的胚乳基因型的对照植物的淀粉高至少大约30%的峰值粘度。提取:从本发明的植物的核中提取该淀粉可以通过本领域中公知的湿磨或干磨方法以标准方式来进行,但是不限于这样的方法。在一个典型的湿磨方法中,该植物通过强气流、筛子和电磁铁来清洁以除去不想要的材料。随后将其浸泡在含有少量二氧化硫的温水中。将浸泡水排空,软化的核运行通过磨碎机以将其分开。除去胚芽并将剩余的混合物研磨,洗涤并筛分为浆料。通过离心将淀粉与谷蛋白分离,随后将剩余的浆状淀粉过滤、洗涤、再悬浮和再过滤。从植物中提取面粉或其变体可以通过干磨方法来完成。在适于本文但不排除其它程序的典型程序中,获自本发明的植物的谷物首先彻底清洁并通过脱壳机,随后调合或调理并通过去胚芽机。将来自去胚芽机的原料干燥并随后冷却,通过碎玉米分离器和吸引器,研磨,并最终根据需要整体还是单独部分来进行筛分。改性:可以理解的是,在上述提取方法之一中的一些修改,如使用低于通常所用温度的浸泡水温度可能是期望的,并将容易被淀粉制造业者所识别。如果需要的话,本文中的淀粉可以通过本领域中已知的程序来改性,如通过衍生化以形成醚或酯,如羟丙基醚、乙酸酯、磷酸酯、琥珀酸酯(例如辛烯基琥珀酸酯)、叔胺和季铵醚等等,或通过制造具有本文中定义的特性的淀粉的任何其它改性技术。本文中优选的取代基是羟丙基、磷酸酯或乙酸酯基团。出于商业目的而言,本文中的淀粉的一种改性是交联以针对在制造操作中常常遇到的处理与加工条件来强化该颗粒,并提供能够赋予食品系统期望的流变性质的淀粉。本领域中已知的任何交联剂可用于该目的,对于食品系统包括但不限于表氯醇、直链二羧酸酐、柠檬酸丙烯醛、磷酰氯、己二酸/乙酸混合酸酐和三偏磷酸盐,并且在非食品系统中包括但不限于表氯醇、直链二羧酸酐、柠檬酸丙烯醛、磷酰氯、己二酸/乙酸混合酸酐、三偏磷酸盐、甲醛、氰尿酰氯、二异氰酸酯和二乙烯基砜。使用本领域中已知的技术进行该交联反应,例如在美国专利号2,328,537和2,801,242中描述的那些。用于将淀粉改性的程序描述在m.w.rutenberg的章节“starchanditsmodification”,第22-26至22-47页,handbookofwatersolublegumsandresins,r.l.davidson编著(mcgraw-hill,inc.,newyork,n.y.1980)中。得到合适的产物所需交联剂的量将根据例如使用的交联剂的类型、交联剂的浓度、反应条件、和具有落在所需粘度范围内的交联淀粉的必要性而变化。从业者将认识到可以采用哪些量,因为这些在本领域中是公知的。通常,该量将低至该淀粉重量的大约0.001%,高至对食品用途被视为可接受的。本发明的淀粉还可以物理改性,如通过wo95/04082(1995年2月9日公开)中描述的热抑制。加工:还可以使该淀粉预胶化。制备预胶化淀粉的示例性方法公开在美国专利号4,280,851(pitchon等人)、美国专利号4,465,702(eastman等人)、美国专利号5,037,929(rajagopalan)、美国专利号5,131,953(kasica等人)和美国专利号5,149,799(rubens)中。使淀粉预胶化的常规程序是本领域技术人员公知的,并描述在诸如以下的文献中:第xxii章“productionanduseofpregelatinizedstarch”,starch:chemistryandtechnology,第iii卷industrialaspects,r.l.whistler和e.f.paschall编著,academicpress,newyork1967。本发明的淀粉可以通过本领域中已知的任何方法纯化以除去该淀粉原有的或在淀粉改性过程中产生的异味和颜色。优选用于处理本发明的淀粉的纯化方法公开在kasica等人在1992年2月7日提交的美国专利申请系列号07/832,838中。碱洗技术,对于旨在以颗粒或预胶化形式使用的淀粉而言也是可用的,并描述在美国专利号5,187,272(bertalan等人)所代表的专利族中。衍生自本发明的淀粉的转化产物,包括但不限于通过氧化、酶转化(特别是通过α-淀粉酶)、酸水解、热和/或酸糊精化、热和/或剪切产物制备的流动性或稀糊淀粉也可用于本文中。产生淀粉的植物本发明提供了产生上述淀粉的植物。所述产生淀粉的植物包括但不限于玉米、马铃薯、小麦、木薯、水稻、木薯和高粱。本发明的淀粉可以从玉米植物中,如从玉米植物的胚乳中提取。产生本发明的淀粉的玉米植物是waxy/sugary-2基因型的双突变植物。该waxy基因位于玉米的9号染色体上,而sugary-2基因位于6号染色体上(参见m.g.nueffer,l.jones和m.zuber,“themutantsofmaize"(cropsciencesocietyofamerica,madison,wi,1968),第72和73页)。本发明的植物对淀粉合成酶iia(su2)基因是纯合隐性的,并且对突变的颗粒结合淀粉合成酶i(gbssi)基因是纯合或杂合的。该突变的gbssi基因具有比野生型gbssi基因(wx)更低的gbssi活性,但是具有比隐性的功能缺失的gbssi基因(wx)更高的gbssi活性。如本文中所用,这样的waxy突变基因被称为中间蜡质突变体。在一些实施方案中,该中间蜡质突变体具有野生型gbssi基因表达的低于30%、低于25%、低于20%、低于15%、低于10%或低于5%。在一些实施方案中,该中间蜡质突变体具有野生型gbssi基因表达的大约5%、大约6%、大约7%、大约8%、大约9%、大约10%、大约11%、大约12%、大约13%、大约14%或大约15%。具有比野生型gbssi基因更低的gbssi活性但具有比隐性的功能缺失的gbssi基因更高的gbssi活性的中间gbssi基因可以是本领域中先前已知的那些,或可以通过任何合适的诱变方法来产生。突变等位基因如wxs(又名wx-stonor)、wx-g、wx-b5和wx-m具有中间gbssi活性。如wessler与varagona(pnas,82:4177-4181,1985)和varagona等人(theplantcell,4:811-820,1992)所述,这些突变等位基因是由于gbssi基因中的逆转录转座子插入。这些中间等位基因导致了具有比野生型蜡质蛋白更低的活性的蜡质蛋白。具有中间gbssi活性的附加突变等位基因可以通过合适的方法来产生。例如,逆转录转座子诱变可用于产生用于筛选具有突变的gbssi基因的玉米植物的群体,所述突变的gbssi基因具有中间gbssi活性。在一些实施方案中,wxs(又名wx-stonor)、wx-g、wx-b5和wx-m可以用作原材料以产生附加突变等位基因。可用于产生具有中间gbssi活性的突变等位基因的其它诱变方法包括但不限于化学诱变、辐射诱变、转座子诱变、插入诱变、信号标签诱变、定点诱变和天然诱变。在又一些实施方案中,反义rna、核酶、dsrnai、rna干扰(rnai)可用于制造具有中间gbssi活性的突变等位基因。反义rna技术涉及在细胞中表达或向细胞中引入与细胞中的特定mrna中发现的序列互补或反义的rna分子(或rna衍生物)。通过与该mrna结合,该反义rna可以抑制编码的基因产物的翻译。使用反义技术减少或抑制特定植物基因的表达已经描述在例如欧洲专利公开号271988,smith等人,nature,334:724-726(1988);smith等人,plantmol.biol.,14:369-379(1990)中。核酶是具有催化结构域和与特定mrna互补的序列的rna。核酶通过与该mrna结合(通过核酶的互补结构域)并随后使用催化结构域分裂(降解)信息来起作用。rna干扰(rnai)是在动物和植物中的序列特异性的、转录后基因沉默的或转录基因沉默的过程,其通过在序列中与沉默基因同源的双链rna(dsrna)引发。该rnai技术例如描述在elibashir等人,methodsenzymol.26:199(2002);mcmanus&sharp,naturerev.genetics3:737(2002);pct申请wo01/75164;martinez等人,cell110:563(2002);elbashir等人,supra;lagos-quintana等人,curr.biol.12:735(2002);tuschl等人,naturebiotechnol.20:446(2002);tuschl,chembiochem.2:239(2001);harborth等人,j.cellsci.114:4557(2001);等人,emboj.20:6877(2001);lagos-quintana等人,science294:8538(2001);hutvagner等人,loccit,834;elbashir等人,nature411:494(2001)。同样在本发明的范围内的是由其中突变的wx和/或su2等位基因已经通过易位、倒位或任何其它染色体工程方法来移至该植物基因组另一部分的突变体所产生的淀粉。此外,从由上述基因组成的人工突变和变异生长的植物中提取的淀粉也适用于本文,所述植物可以通过已知的突变育种的标准方法来产生。同样在本发明的范围内的是由玉米植物以外的植物产生的淀粉,只要产生该淀粉的植物包含具有中间gbssi活性的玉米gbssi基因的突变直系同源物和玉米ssiia基因的隐性突变直系同源物。在一些实施方案中,该植物可以是小麦、木薯、水稻、木薯、西米、马铃薯或高粱。在一些实施方案中,除玉米以外的植物中的直系同源gbssi基因编码具有与seqidno:1至少80%、至少81%、至少82%、至少83%、至少84%、至少85%、至少86%、至少87%、至少88%、至少89%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%或更高的同一性的蛋白,并具有该gbssi活性。在一些实施方案中,该直系同源物具有uniprot条目号o82627(antirrhinummajus(金鱼草))、p12615(avenasativa(燕麦))、q42968(oryzaglaberrima(非洲水稻))、q43784(manihotesculenta(木薯))、p0c585(oryzasativa(水稻))、q00775(solanumtuberosum(马铃薯))、p27736(triticumaestivum(小麦))、q43134(sorghumbicolor(高粱))、q43092(pisumsativum(豌豆))、p84766(aegilopstauschiisubsp.strangulata(山羊草))、q0dev5(oryzasativasubsp.japonica(水稻))、q42857(ipomoeabatatas(甘薯))、p84633(fagopyrumesculentum(普通荞麦))、p84765(secalecereale(黑麦))、q9maq0(arabidopsisthaliana)、p09842(hordeumvulgare(大麦))或a2y8x2(oryzasativasubsp.indica(水稻))的序列,其各自经此引用整体并入本文。在一些实施方案中,除玉米以外的植物中的直系同源ssiia基因编码具有与seqidno:2至少80%、至少81%、至少82%、至少83%、至少84%、至少85%、至少86%、至少87%、至少88%、至少89%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%或更高的同一性的蛋白,并具有ssiia活性。在一些实施方案中,该直系同源物具有uniprot条目号v6bpg5(triticumurartu(红色野生单粒小麦))、v6bpn2(triticummonococcumsubsp.monococcum)、c3w8l4(hordeumvulgarevar.distichum(训化大麦))、e2ghr4(oryzasativasubsp.indica(水稻))或e2ghr6(oryzasativasubsp.japonica(水稻))的序列,其各自经此引用整体并入本文。在一些实施方案中,为了产生具有中间wx突变体和隐性su2突变体的双突变植物,可以进行在具有中间wx突变体的植物与具有隐性su2突变体的植物之间的异花传粉。例如,可以在纯合wxs突变体(wxswxssu2su2)或杂合wxs突变体(wxswxsu2su2)与纯合waxy/su2突变体(wxwxsu2su2)或杂合waxy/su2突变体(wxwxsu2su2或wxwxsu2su2)之间进行异花传粉。在杂交后,具有wxswxsu2su2的基因型的f1代的自花传粉可用于产生f2植物。可以相应地分离在胚乳中具有0、1、2或3剂量的wxs基因和3剂量的隐性su2基因的植物(例如具有wxwxwxsu2su2su2、wxswxwxsu2su2su2、wxswxswxsu2su2su2或wxswxswssu2su2su2的基因型的胚乳)。这仅仅是育种方法的一个实例,可以采用其它育种方案,并且其在本发明的范围内。在一些实施方案中,其胚乳具有至少1剂量的中间waxy等位基因和隐性sugary-2等位基因的植物的田间生产是通过使雌性玉米植物与雄性玉米植物杂交来进行。典型的植物布置是一行雄性对几行雌性。雌性行通过本领域已知的各种其它方法(如细胞质或基因手段)去雄或赋予雄性不育。本发明中使用的淀粉可以获自自交系,或者该淀粉可以获自杂交种,该杂交种衍生自含有中间waxy/隐性su2双突变体的自交系。虽然玉米是本文中蜡质淀粉来源的示例性植物,本发明还适用于其它植物物种,例如蜡质大米、蜡质大麦和蜡质高粱,条件是它们具有中间waxy基因和隐性su2基因。育种方法为了制造本发明的植物,可以使用合适的育种或选择方法。常用的选择方法通常包括谱系选择、改良的谱系选择、混合选择、轮回选择和回交育种。开放授粉的种群:这样的作物如黑麦、许多玉米和甜菜、牧草、豆类如苜蓿和三叶草,以及热带树木作物如可可、椰子、油棕榈和一些橡胶的开放授粉种群的改善基本上取决于改变基因的频率,目标是固定有利等位基因同时保持高(但远非最大)杂合度。在这样的种群中的均一性是不可能的,在开放授粉品种中的纯真型是该种群作为整体的统计特征,而非个体植物的特性。因此,开发授粉种群的异质性与自交系、克隆和杂交系的同质性(或几乎同样)形成对比。种群改善方法天然地分成两组,基于纯表型选择的那些(通常称为混合选择),和基于采用后代测试的选择的那些。种群间改善利用了开放育种种群的概念;允许基因从一个种群流向另一个。一个种群(栽培品种、品系、生态型或任何种质源)中的植物与来自其它种群的植物自然地(例如通过风)杂交,或是通过手工或通过蜜蜂(通常为apismelliferal.或megachilerotundataf.)杂交。施加选择以通过分离具有来自两个来源的期望性状的植物来改善一个(或有时为两个)种群。基本上存在两种开放授粉群体改善的主要方法。首先,存在这样的情况,其中通过所选择的选择程序一同改变种群。结果是通过在孤立的自身内随机交配可无限繁殖的改善的种群。第二,合成品种获得了与种群改善相同的最终结果,但是本身不能如此繁殖;其必须由亲本系或克隆重构。这些用于改善开放授粉种群的植物育种程序对本领域技术人员是公知的,并且常规用于改善异花授粉植物的育种程序的全面评述在大量文本和文章中提供,包括:allard,principlesofplantbreeding,johnwiley&sons,inc.(1960);simmonds,principlesofcropimprovement,longmangrouplimited(1979);hallauer和miranda,quantitativegeneticsinmaizebreeding,iowastateuniversitypress(1981);和jensen,plantbreedingmethodology,johnwiley&sons,inc.(1988)。混合选择:在混合选择中,选择期望的个体植物,收获,并且种子在未经后代测试的情况下复合以产生下一代。由于选择仅基于母本,并且不存在对授粉的控制,混合选择相当于具有选择的随机交配形式。如本文中所述,混合选择的目的在于提高种群中优质基因型的比例。合成:通过在所有可能的杂交组合中针对良好配合力所选择的多种基因型相互之间的杂交来产生合成品种,随后通过开放授粉维持该品种。无论亲本是(或多或少自交)种子繁殖系,如在一些甜菜和豆类(vicia)中,或是克隆,如在牧草、三叶草和苜蓿中,原则上没有差异。针对一般配合力选择亲本,有时通过测交或顶交,更通常通过多系杂交。亲本种子系可以故意自交(例如通过自花受精或姐妹种杂交)。但是,即使亲本并非故意自交,在系维护过程中在系内的选择将确保出现一些自交。克隆的亲本当然将保持不变和高度杂合。合成物可以直接从亲本种子生产转移到农民还是必须首先经历一次或两次繁殖周期,这取决于种子生产和对种子的需求的规模。在实践中,牧草和三叶草通常繁殖一次或两次,由此显著从原始合成物中除去。虽然有时采用混合选择,但是对多系杂交而言后代测试通常是优选的,因为它们的操作简便性和与目标的显著相关性,即在合成中利用一般配合力。进入合成的亲本系或克隆的数量变化很大。在实践中,亲本系的数量为10至数百,平均100-200。由100或更多个克隆形成的基础广泛的合成物预期在种子繁殖过程中比基础狭窄的合成物更稳定。纯种品种:纯种品种是通过从分离种群中选出个体植物开发的优良基因型,随后是自花授粉后代的繁殖和种子增加,并对几代仔细测试该基因型。这是一种开放授粉方法,适用于天然自花传粉物种。这种方法可以在物种开发中与混合选择结合使用。纯种和混合选择组合的变体是在自花授粉作物中产生品种的最常见方法。杂交种:杂交种是来自不同基因型的亲本之间的杂交的个体植物。商业杂交种现在广泛用于许多作物,包括玉米(玉蜀黍)、高粱、甜菜、向日葵和花椰菜。杂交种可以多种不同方式形成,包括通过直接使两种亲本杂交(单交种)、通过单交种与另一亲本杂交(三向杂交或三交种)、或通过两种不同杂交种杂交(四向杂交或双交杂交种)。谱系选择:谱系育种通常用于改善自花传粉作物或异花传粉作物的自交系。将具有有利的互补性状的两种亲本杂交以产生f1。通过一个或多个f1的自交或通过两个f1的互交(同胞交配)来产生f2种群。该双单倍体育种方法或其它倍性降低技术也可以使用。通常在f2种群中开始最佳个体的选择;随后,从f3开始,选择最佳家族中的最佳个体。家族或涉及这些家族的个体的杂交组合的复制测试通常跟随f4代以提高对具有低遗传力的性状选择的有效性。在同系交配的晚期(即f6和f7),测试最佳品系或表型相似品系的混合物的新品种的潜在释放。类似地,通过双单倍体系统开发新品种需要选择栽培品种,随后在重复区中进行两年到五年的测试。回交育种:回交育种已经用于将简单遗传的高度可遗传性状的基因转移到作为轮回亲本的期望的纯合栽培品种或自交系中。待转移的性状来源称为供体亲本。所得植物预期具有该轮回亲本(例如栽培品种)的属性和从该供体亲本转移的期望性状。在初始杂交后,选择具有供体亲本的表型的个体,并反复与该轮回亲本杂交(回交)。所得植物预期具有该轮回亲本(例如栽培品种)的属性和从该供体亲本转移的期望性状。群体分离分析(bsa):bsa(又名群分离分析或群组分离分析)是由michelmore等人(michelmore等人,1991,identificationofmarkerslinkedtodisease-resistancegenesbybulkedsegregantanalysis:arapidmethodtodetectmarkersinspecificgenomicregionsbyusingsegregatingpopulations,proceedingsofthenationalacademyofsciences,usa,99:9828-9832)和quarrie等人(quarrie等人,1999,journalofexperimentalbotany,50(337):1299-1306)描述的方法。对相关性状的bsa而言,采用qtl分析选择具有某些不同表型的亲本系并将其杂交以产生f2、加倍单倍体或重组自交系种群。该种群随后进行表型以确定具有性状的高或低表达的个体植物或品系。制备两个dna混合池,一个来自具有一种表型(例如对病毒的抗性)的个体,另一个来自具有反向表型(例如对病毒易感)的个体,并用分子标记分析等位基因频率。如果该标记是显性的(例如rapds),在每个混合池中仅需要几个个体(例如每个混合池10株植物)。当标记是共显性时(例如rflps),需要更多的个体。连接到该表型的标记可以被识别并用于育种或qtl作图。定向诱导基因组局部病变(tilling®):本申请的育种方案可以包括采用tilling®植物栽培品种的杂交。tilling®是分子生物学中的一种方法,其能够在特定基因中定向鉴定突变。tilling®在2000年引入,使用模型植物arabidopsisthaliana。tilling®已经在其它生物体如斑马鱼、玉米、小麦、水稻、大豆、番茄和brassicarapavar.nipposinica中作为反向遗传学方法使用。该方法将采用化学诱变剂(例如甲磺酸乙酯(ems))的标准和有效的诱变技术与识别靶基因中单碱基突变(也称为点突变)的敏感dna筛选技术结合。ecotilling是使用tilling®技术寻找个体中的天然突变的方法,通常用于群体遗传学分析(参见comai等人,2003theplantjournal37,778-786;gilchrist等人,2006mol.ecol.15,1367-1378;mejlhede等人,2006plantbreeding125,461-467;nieto等人,2007bmcplantbiology7,34-42,其各自经此引用出于所有目的并入本文)。decotilling是tilling®和ecotilling的修改,其使用廉价的方法来鉴定片段(garvin等人,2007,deco-tilling:aninexpensivemethodforsnpdiscoverythatreducesascertainmentbias.molecularecologynotes7,735-746)。该tilling®法依赖于多个等位基因(其可以来自杂合子或多个纯合子与杂合子的池)在pcr中扩增、加热并随后缓慢冷却时形成的杂合双链的形成。在两条dna链的错配(在tilling®中的诱导突变或天然突变或在ecotilling中的snp)处形成“泡”,其随后被单链核酸酶分裂。产物随后在几个不同平台上通过尺寸分离。关于tilling®的方法和组合物的更详细描述可以在us5994075、us2004/0053236a1、wo2005/055704和wo2005/048692中找到,其各自经此引用出于所有目的并入本文。由此在一些实施方案中,本公开的育种方法包括用一种或多种具有一个或多个鉴定的突变的tilling®植物系进行育种。突变育种:突变育种是向植物中引入新性状的另一种方法。自发发生或人工诱导的突变对植物育种者而言可以是变异性的可用来源。人工诱变的目的是提高所需特性的突变率。突变率可以通过许多不同的手段或突变试剂来提高,包括温度、长期种子储存、组织培养条件、辐射(如x射线、γ射线、中子、β辐射或紫外辐射)、化学诱变剂(如碱基类似物,如5-溴-尿嘧啶)、抗生素、烷基化剂(如硫芥、氮芥、环氧化物、亚乙基胺、硫酸盐、磺酸盐、砜或内酯)、叠氮化物、羟胺、亚硝酸或吖啶。一旦通过诱变观察到所需性状,随后可以通过传统育种技术将该性状并入现有种质。突变育种的细节可以在w.r.fehr,1993,principlesofcultivardevelopment,macmillanpublishingco.中找到。新的育种技术,如涉及使用锌指核酸酶或寡核苷酸定向诱变的技术也应当用于产生遗传变异性和向品种中引入新的性状。在一些实施方案中,突变育种包括单基因座转换。单基因座可以含有几个基因和/或转基因,如用于抗病性的转基因,其在相同的表达载体中还含有用于除草剂抗性的转基因。用于除草剂抗性的基因可以用作可选标记和/或用作表型性状。位点特异性整合系统的单基因座转换允许在转换的基因座处整合多个基因。单基因座转换还能够对该植物的基因组进行一个或多个位点特异性改变。在一些实施方案中,通过基因组编辑进行单基因座转换,即采用工程化核酸酶的基因组编辑(geen)。在一些实施方案中,该基因组编辑包括使用一种或多种工程化核酸酶。在一些实施方案中,该工程化核酸酶包括但不限于锌指核酸酶(zfn)、转录激活因子样效应子核酸酶(talen)、crispr/cas系统和工程化的大范围核酸酶重新工程化的归位内切酶。在一些实施方案中,单基因座转换改变了该植物基因组的一个或多个核酸。双单倍体和染色体倍增:获得纯合植物而无需杂交两个亲本系并随后长时间选择分离后代和/或多次回交的一种方法是产生单倍体并随后将染色体倍增以形成双单倍体。单倍体植物可以自发地发生,或可以经由化学处理或通过使植物与诱导物系杂交来人工诱导(seymour等人,2012,pnas,第109卷,第4227-4232页;zhang等人,2008plantcellrep.dec27(12)1851-60)。单倍体后代的产生可以经由多种机制发生,其可以在配子形成过程中影响染色体的分布。单倍体的染色体组有时自发地倍增以产生纯合双单倍体(dh)。混倍体(其是含有具有不同倍性的细胞的植物)有时可能出现并可以代表经历染色体倍增以便自发产生双单倍体组织、器官、芽、花部分或植物的植物。另一种常见技术是用染色体倍增处理如秋水仙碱来诱导形成双单倍体植株(el-hennawy等人,2011第56卷,第2期,第63-72页;maluszynski编著的doubledhaploidproductionincropplants2003,isbn1-4020-1544-5)。产生加倍单倍体植株得到了高度一致的栽培品种,并且作为长期作物的性自交的替代品是尤其期望的。通过产生双单倍体后代,遗传性状的可能的基因组合数量更易于管理。由此,有效的双单倍体技术可以显著减少自交和栽培品种开发的时间和成本。原生质体融合:在另一种植物育种方法中,原生质体融合也可用于将赋予性状的基因组材料由供体植物转移至受体植物。原生质体融合是两个或多个原生质体(细胞壁通过酶处理被除去的细胞)之间的诱导性或自发性结合(如体细胞杂交)以产生单个双核或多核细胞。将甚至可以用不能在自然界中混种的植物物种获得的融合细胞组织培养成表现出所需性状的组合的杂交植物。组织培养物:本文中所用的术语“组织培养物”表示包含相同或不同类型的分离细胞或组织成植物部分的这样的细胞的集合的组合物。组织培养物的示例性类型是可以产生在植物或植物部分中完整的组织培养物的原生质体、胼胝体、植物簇和植物细胞,所述植物部分例如胚芽、花粉、花、种子、叶、茎、根、根尖、花药、雌蕊、分生细胞、腋芽、子房、种皮、胚乳、下胚轴、子叶等等。用于制备和维护植物组织培养物的手段在本领域中是公知的。例如,包含器官的组织培养物已经用于产生再生植物。美国专利号5,959,185、5,973,234和5,977,445描述了某些技术,其公开内容经此引用并入本文。通过下面的实施例和实施方案进一步举例描述本发明,所述实施例和实施方案不应解释为限制性的。在整个本申请中引用的所有参考文献、专利和公开的专利申请的内容,以及附图和序列表均经此引用并入本文。实施方案1.淀粉,包含:如使用直链淀粉/支链淀粉测定试剂盒测得的大约2重量%至大约20重量%的直链淀粉含量;和如使用rva分析测得的比对照淀粉的淀粉糊化温度高至少大约5%的水淀粉糊化温度,其中,所述淀粉衍生自在胚乳中包含至少一个wxs等位基因的waxy/sugary-2玉米植物。2.实施方案1的淀粉,其中直链淀粉含量为大约8重量%至大约15重量%。3.实施方案1或2任一项的淀粉,其中水糊化温度比对照淀粉的水糊化温度高大约5%至大约14%。4.实施方案1-3任一项的淀粉,进一步具有如使用rva分析测得的比对照淀粉的结晶焓变高至少大约30%的结晶焓变。5.实施方案4的淀粉,其中如使用差示扫描量热计(dsc)测得的,该结晶焓变比对照淀粉的结晶焓变高大约70%至大约200%。6.实施方案1-4任一项的淀粉,进一步具有淀粉颗粒尺寸的分布,其中如使用扫描电子显微镜观察到的,该淀粉颗粒尺寸的分布包含与对照淀粉中小于5微米的颗粒的比例相比少至少大约40%的小于5微米的颗粒。7.实施方案1-6任一项的淀粉,其中如使用扫描电子显微镜观察到的,该淀粉是与对照淀粉相比表现出提高的完整性的颗粒。8.实施方案1-7任一项的淀粉,其中该淀粉能够承受至少三次冻融循环。9.实施方案8的淀粉,其中该淀粉能够承受至少三次到至少五次冻融循环。10.实施方案1-9任一项的淀粉,其中该waxy/sugary-2玉米植物通过具有wxwxsu2su2的基因型的第一玉米植物与具有wxswxssu2su2的基因型或wxswxsu2su2的基因型的第二玉米植物的异花传粉来产生。11.实施方案1-10任一项的淀粉,其中该waxy/sugary-2玉米植物的胚乳具有1剂量、2剂量或3剂量的wxs等位基因,并且该胚乳具有wxswxwxsu2su2su2、wxswxswxsu2su2su2或wxswxswxssu2su2su2的基因型。12.用于改善淀粉的糊化曲线中的至少一个特性的方法,包括向蜡质玉米植物的胚乳中引入至少一个wxs等位基因,其中衍生自在胚乳中包含至少一个wxs等位基因的蜡质玉米植物的所得淀粉具有如使用rva分析测得的比对照淀粉的淀粉糊化温度高至少大约5%的水淀粉糊化温度,由此与对照淀粉相比改善该淀粉的糊化曲线中的至少一个特性。13.双突变玉米植物,其中该玉米植物对淀粉合成酶iia(su2)基因是纯合隐性的,并且对突变的颗粒结合淀粉合成酶i(gbssi)基因是纯合或杂合的,其中该突变的gbssi基因具有比野生型gbssi基因(wx)更低的gbssi活性,但是具有比隐性的功能缺失的gbssi基因(wx)更高的gbssi活性。14.实施方案13的双突变玉米植物,其中该突变的gbssi基因是wx-stonor(wxs)等位基因。15.实施方案14的双突变玉米植物,其中该植物通过具有wxwxsu2su2的基因型的第一玉米植物与具有wxswxssu2su2的基因型或wxswxsu2su2的基因型的第二玉米植物的异花传粉来产生。16.实施方案15的双突变玉米植物,其中该玉米植物的胚乳具有1剂量、2剂量或3剂量的wxs等位基因,其中所述胚乳具有wxswxwxsu2su2su2、wxswxswxsu2su2su2或wxswxswxssu2su2su2的基因型。17.实施方案13-16任一项的双突变玉米植物,其中在该玉米植物的胚乳中产生的淀粉与具有wxwxwxsu2su2su2的基因型的双waxy/sugary-2隐性突变玉米植物的胚乳中产生的淀粉相比具有更高的糊化温度。18.实施方案13-17任一项的双突变玉米植物的种子。19.实施方案13-18任一项的双突变玉米植物的植物部分、植物细胞或组织培养物。20.实施方案19的植物部分,其中该植物部分选自胚乳、叶、花、胚珠、花粉、根茎和接穗。21.生产淀粉的方法,包括从实施方案13-20任一项的双突变玉米植物的胚乳中获得淀粉。22.生产淀粉的方法,包括种植实施方案13-21任一项的双突变玉米植物,并从该双突变玉米植物收获种子。23.生产玉米种子的方法,包括使第一亲本玉米植物与第二亲本玉米植物杂交并收获所得玉米种子,其中所述第一亲本玉米植物和/或第二亲本玉米植物是实施方案13-22任一项的双突变玉米植物。24.生产玉米种子的方法,包括使实施方案13-23任一项的双突变玉米植物自花传粉,并收获所得玉米种子。25.无性繁殖实施方案13-24任一项的玉米植物的方法,所述方法包括收集实施方案1-11任一项的植物部分并由所述部分再生植物。26.生产衍生自实施方案13-17任一项的双突变玉米植物的玉米植物的方法,该方法包括使自实施方案13-17任一项的双突变植物自花传粉至少一次以产生衍生自实施方案13-17任一项的双突变植物的后代植物。27.生产衍生自实施方案13-17任一项的双突变植物的玉米植物的方法,该方法包括使实施方案13-17任一项的双突变植物与第二玉米植物杂交以产生实施方案13-17任一项的双突变植物的后代植物。28.生产玉米植物的方法,包括向玉米植物中引入转基因,其中该转基因赋予实施方案13-17任一项的双突变植物的期望表型。29.生产玉米植物的方法,包括基因编辑以产生基因序列,所述基因序列赋予实施方案13-17任一项的双突变植物的期望表型。30.生产玉米植物的方法,包括引入突变以产生基因序列,所述基因序列赋予实施方案13-17任一项的双突变植物的期望表型。31.从实施方案13-17任一项的双突变玉米植物中提取的淀粉。32.从实施方案15的双突变玉米植物中提取的淀粉。33.从实施方案16的双突变玉米植物的胚乳中提取的淀粉。实施例1生成具有1、2或3剂量的wxs等位基因的谷物由于玉米胚乳的三倍体性质,其中来自母本的两个极性核通过来自父本的一个精细胞受精,可能存在三种基因型组合。使用wxswxssu2su2植物作为雌性,其通过wxwxsu2su2雄性授粉导致“剂量-2”或wxswxswxsu2su2su2谷物。使用wxswxssu2su2作为雄性,其授粉wxwxsu2su2雌性导致“剂量-1”或wxswxwxsu2su2su2谷物。wxswxssu2su2的自花传粉导致“剂量-3”或wxswxswxssu2su2su2谷物。wxs剂量胚乳基因型(1wxs)wxswxwxsu2su2su2(2wxs)wxswxswxsu2su2su2(3wxs)wxswxswxssu2su2su2来自wxwxwxsu2su2su2种子的淀粉(0wxs)wxwxwxsu2su2su2amioca™淀粉(0wxs)wxwxwxsu2su2su2来自wxwxwxsu2su2su2的淀粉(0wxs)wxwxwxsu2su2su2实施例2使用快速粘度糊化仪(rva)的淀粉的粘度使用快速粘度糊化仪(newportscientific,edenprairie,minn.)测定衍生自在隐性sugary-2突变植物中含有0、1、2和3剂量的wxs等位基因的植物的玉米淀粉的粘度。包括amioca™淀粉和wxwxwxsu2su2su2淀粉作为对照。与所述淀粉分离的胚乳的基因型总结在上表中。使用megazyme®直链淀粉试剂盒测量直链淀粉含量。随着gbssi活性的提高,该淀粉的直链淀粉含量提高。wxwxwxsu2su2su2淀粉(7.4%)、amioca™淀粉(7.1%)和wxwxwxsu2su2su2淀粉(6.1%)的直链淀粉含量低于waxystonor(12.8%)和在胚乳中具有1至3剂量的wxs基因的wxwxwxsu2su2su2淀粉(8.2-14.6%)。wxwxwxsu2su2su2淀粉的直链淀粉含量(7.4%、8.2%、13.8%和14.6%)随着wxs剂量的提高(分别为0、1、2和3剂量的wxs)而提高。结果表明,向wxwxwxsu2su2su2中引入wxs基因略微提高了该淀粉的直链淀粉含量。采用“标准2”轮廓法使用快速粘度分析仪(rva,newportscientific,sydney,australia)分析蜡质玉米淀粉的糊化性质。含有8%淀粉(w/w,干淀粉基,dsb)的悬浮液(28.0克)在50℃下平衡1分钟,以6℃/分钟的速率加热至95℃,在95℃下保持5分钟,随后以6℃/分钟的速率冷却至50℃。除了960rpm用于最初10秒外,桨叶的旋转速度为160rpm。结果显示在图1和图2中。现有的wxwxwxsu2su2su2淀粉具有最低的糊化温度、峰值粘度和最终粘度。在向wxwxwxsu2su2su2淀粉中引入wxs基因后,淀粉样品(q3017a[1wxs]、q8412a[2wxs]和q7738a[3wxs])显示出与现有的wxwxwxsu2su2su2淀粉(0wxs)相比提高的峰值粘度。但是,仅仅具有1剂量的wxs基因的wxswxwxsu2su2su2淀粉显示出远高于waxystonor、amioca™淀粉和wxwxwxsu2su2su2淀粉的峰值粘度。结果表明,糊化温度的改善将允许经由湿磨法进行淀粉提取。实施例3淀粉的冻-融稳定性测试衍生自在隐性sugary-2突变植物中含有0、1、2和3剂量的wxs等位基因的植物的玉米淀粉的冻-融稳定性。包括amioca™淀粉和wxwxwxsu2su2su2淀粉作为对照。8%固体的淀粉样品在水浴中在95℃下烹煮20分钟。该淀粉浆料用玻璃棒混合3分钟,随后在烹煮过程中静态加热17分钟。中性ph下淀粉凝胶(8%,w/w)的冻-融稳定性使用caron循环冷冻器分析五次冻-融循环。在各循环中,将样品冷冻6小时并解冻6小时,每天2次循环。目视检查样品在表面上和在压制时的脱水收缩,以及不透明度和胶凝变化。重复该冻/融循环。结果显示在图3中。在第一次冻-融循环后,仅有waxystonor淀粉凝胶显示出高不透明度,表明高的淀粉凝沉程度。淀粉凝胶的不透明度随着冻/融循环的增加而提高。在第三次冻/融循环时,amioca™淀粉、wxwxwxsu2su2su2淀粉和waxystonor的淀粉凝胶变得非常不透明,而q3017a(1wxs)、q8412a(2wxs)、q7738a(3wxs)和现有的wxwxwxsu2su2su2淀粉的淀粉凝胶仍未不透明。在五次冻/融循环后,q3017a(1wxs)、q8412a(2wxs)、q7738a(3wxs)和现有的wxwxwxsu2su2su2淀粉变得略微不透明。结果表明,淀粉样品中直链淀粉的略微增加对冻/融稳定性几乎没有影响。q3017a(1wxs)、q8412a(2wxs)、q7738a(3wxs)淀粉具有优于wxwxwxsu2su2su2淀粉和amioca™淀粉的冻/融稳定性。实施例4差示扫描量热法(dsc)测试采用差示扫描量热法(dsc)测定衍生自在隐性sugary-2突变植物中含有0、1、2和3剂量的wxs等位基因的植物的玉米淀粉的热性质。包括amioca™淀粉和wxwxwxsu2su2su2淀粉作为对照。差示扫描量热法(dsc)测量在规定温度下在受控气氛中样品所吸收或释放的热量。dsc提供关于样品随着温度升高的比热和潜热的信息,这显示出在非晶与结晶结构中的变化。数据根据热流记录,并以焦耳/克(j/g)表示(cassel,2002)。在淀粉的分析中,收集淀粉糊化参数如峰值开始、峰值温度、峰值终点和糊化焓变信息。结果显示在图4中。wxswxwxsu2su2su2淀粉(q3017a)、wxswxswxsu2su2su2淀粉(q8412a)、wxswxswxssu2su2su2淀粉(q7738a)和wxwxwxsu2su2su2淀粉(0wxs)显示出与amioca™淀粉、waxystonor和wxwxwxsu2su2su2淀粉相比更低的糊化温度,这与从具有1、2和3剂量的wxs的玉米胚乳中分离的淀粉的更好的冻/融稳定性一致。具有1剂量的wxs的wxswxwxsu2su2su2淀粉(q3017a)具有比其它杂交种(q8412a[2wxs]、q7738a[3wxs]和wxwxwxsu2su2su2淀粉)略微更高的开始糊化温度,表明该q3017a具有比其它杂交种更好的湿磨特性。实施例5淀粉颗粒通过激光散射测量衍生自在隐性sugary-2突变植物中含有1、2和3剂量的wxs等位基因的植物的蜡质玉米淀粉的平均粒度。包括amioca™淀粉和wxwxwxsu2su2su2淀粉作为对照。结果显示在图5中。此外,在图6中提供了各样品的扫描电子显微镜照片以显示颗粒形态。q3017a(11.4微米)、q8412a(11.4微米)和q7738a(10.9微米)的平均淀粉颗粒尺寸大于现有的wxwxwxsu2su2su2淀粉(10.4微米),表明向waxyvo中引入wxs基因提高了淀粉颗粒尺寸。该结果与sem图像(图5)一致,表明向wxwxwxsu2su2su2淀粉中引入wxs基因提高了该淀粉颗粒完整性。除非另行规定,本文中所有技术和科学术语具有与本发明所属领域普通技术人员通常理解的相同含义。虽然与本文中描述的那些相似或等同的任何方法和材料可以用于本公开的实践或测试,但是优选的方法和材料描述在本文中。出于所有目的,引用的所有出版物、专利和专利公开经此引用整体并入本文。本文讨论的出版物仅用于其在本申请的申请日之前的公开内容而提供。本文中的任何内容不应被解释为承认本公开没有权利由于在先发明而先于这样的公开。虽然已经结合其具体实施方案对本发明进行了描述,应该理解的是,能够进一步修改,并且本申请旨在覆盖通常遵循本发明的原则的对本发明的任何改变、使用或改编,并且包括对本公开的这样的偏离,如在本发明所属领域内的已知或常规实践内,和如可以应用于上文阐述的基本特征并且如在所附权利要求的范围内。序列表<110>ingredionincorporatedjiang,hongxinostrander,bradlane,chris<120>用于生产具有新功能的淀粉的组合物和方法<130>ingr-036/00us317596-2045<160>2<170>patentinversion3.5<210>1<211>605<212>prt<213>玉米<400>1metalaalaleualathrserglnleuvalalathrargalaglyleu151015glyvalproaspalaserthrpheargargglyalaalaglnglyleu202530argglyalaargalaseralaalaalaaspthrleusermetargthr354045seralaargalaalaproarghisglnglnglnalaargargglygly505560argpheproserleuvalvalcysalaseralaglymetasnvalval65707580phevalglyalaglumetalaprotrpserlysthrglyglyleugly859095aspvalleuglyglyleuproproalametalaalaasnglyhisarg100105110valmetvalvalserproargtyraspglntyrlysaspalatrpasp115120125thrservalvalsergluilelysmetglyaspglytyrgluthrval130135140argphephehiscystyrlysargglyvalaspargvalphevalasp145150155160hisproleupheleugluargvaltrpglylysthrgluglulysile165170175tyrglyprovalalaglythrasptyrargaspasnglnleuargphe180185190serleuleucysglnalaalaleuglualaproargileleuserleu195200205asnasnasnprotyrpheserglyprotyrglygluaspvalvalphe210215220valcysasnasptrphisthrglyproleusercystyrleulysser225230235240asntyrglnserhisglyiletyrargaspalalysthralaphecys245250255ilehisasnilesertyrglnglyargphealapheserasptyrpro260265270gluleuasnleuprogluargphelysserserpheasppheileasp275280285glytyrglulysprovalgluglyarglysileasntrpmetlysala290295300glyileleuglualaaspargvalleuthrvalserprotyrtyrala305310315320glugluleuileserglyilealaargglycysgluleuaspasnile325330335metargleuthrglyilethrglyilevalasnglymetaspvalser340345350glutrpaspproserargasplystyrilealavallystyraspval355360365serthralavalglualalysalaleuasnlysglualaleuglnala370375380gluvalglyleuprovalaspargasnileproleuvalalapheile385390395400glyargleuglugluglnlysglyproaspvalmetalaalaalaile405410415proglnleumetglumetvalgluaspvalglnilevalleuleugly420425430thrglylyslyslysphegluargmetleumetseralagluglulys435440445pheproglylysvalargalavalvallyspheasnalaalaleuala450455460hishisilemetalaglyalaaspvalleualavalthrserargphe465470475480gluprocysglyleuileglnleuglnglymetargtyrglythrpro485490495cysalacysalaserthrglyglyleuvalaspthrileileglugly500505510lysthrglyphehismetglyargleuservalaspcysasnvalval515520525gluproalaaspvallyslysvalalathrthrleuglnargalaile530535540lysvalvalglythrproalatyrgluglumetvalargasncysmet545550555560ileglnaspleusertrplysglyproalalysasntrpgluasnval565570575leuleuserleuglyvalalaglyglygluproglyvalgluglyglu580585590gluilealaproleualalysgluasnvalalaalapro595600605<210>2<211>729<212>prt<213>玉米<400>2metserseralaalavalserserserserserthrphepheleuala151015leualaseralaserproglyglyargargargalaargvalglyser202530serprophehisthrglyalaserleuserphealaphetrpalapro354045proserproproargalaproargaspalaalaleuvalargalaglu505560alaglualaglyglylysaspalaproprogluargserglyaspala65707580alaargleuproargalaargargasnalavalserlysargargasp859095proleuglnprovalglyargtyrglyseralathrglyasnthrala100105110argthrglyalaalasercysglnasnalaalaleualaaspvalglu115120125ilelysserilevalalaalaproprothrserilevallysphepro130135140alaproglytyrargmetileleuproserglyaspilealaproglu145150155160thrvalleuproalaprolysproleuhisgluserproalavalasp165170175glyaspserasnglyilealaproprothrvalgluproleuvalgln180185190glualathrtrpaspphelyslystyrileglypheaspgluproasp195200205glualalysaspaspserargvalglyalaaspaspalaglyserphe210215220gluhistyrglyaspasnaspserglyproleualaglygluasnval225230235240metasnvalilevalvalalaalaglucysserprotrpcyslysthr245250255glyglyleuglyaspvalvalglyalaleuprolysalaleualaarg260265270argglyhisargvalmetvalvalvalproargtyrglyasptyrval275280285glualapheaspmetglyilearglystyrtyrlysalaalaglypro290295300valasntyrphehisalapheileaspglyvalaspphevalpheile305310315320aspalaproleuphearghisargglnaspaspiletyrglyglyser325330335argglngluilemetlysargmetileleuphecyslysvalalaval340345350gluvalprotrphisvalprocysglyglyvalcystyrglyaspgly355360365asnleuvalpheilealaasnasptrphisthralaleuleuproval370375380tyrleulysalatyrtyrargasphisglyleumetglntyrthrarg385390395400servalleuvalilehisasnilealahisglnglyargglyproval405410415aspglupheprotyrmetaspleuprogluhistyrleuglnhisphe420425430gluleutyraspprovalglyglygluhisalaasnilephealaala435440445glyleulysmetalaaspargvalvalthrvalserargglytyrleu450455460trpgluleulysthrvalgluglyglytrpglyleuhisaspileile465470475480argserasnasptrplysileasnglyilevalasnglyileasphis485490495glnglutrpasnprolysvalaspvalhisleuargseraspglytyr500505510thrasntyrserleugluthrleuaspalaglylysargglncyslys515520525alaalaleuglnarggluleuglyleugluvalargaspaspvalpro530535540leuleuglypheileglyargleuaspglyglnlysglyvalaspile545550555560ileglyaspalametprotrpilealaglyglnaspvalglnleuval565570575metleuglythrglyargalaaspleugluargmetleuglnhisleu580585590gluarggluhisproasnlysvalargglytrpvalglypheserval595600605prometalahisargilethralaglyalaaspvalleuvalmetpro610615620serargphegluprocysglyleuasnglnleutyralametalatyr625630635640glythrvalprovalvalhisalavalglyglyleuargaspthrval645650655alapropheasppropheseraspalaglyleuglytrpthrpheasp660665670argalaglualaasnlysleuileglualaleuarghiscysleuasp675680685thrtyrargasntyrgluglusertrplysserleuglnalaarggly690695700metserglnaspleusertrpasphisalaalagluleutyrgluasp705710715720valleuvallysalalystyrglntrp725当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1