结合并抑制过氧化氢酶和/或超氧化物歧化酶的抗原结合构建体即单域VHH片段以及用于肿瘤疗法的含有它们的药物组合物的制作方法
本发明基于意想不到的发现,即抗原结合构建体,特别是中和人过氧化氢酶或人sod1的单域vhh片段(纳米抗体(nanobodies))引起比均由轻链和重链部分组成并且也都分别表现出对过氧化氢酶或sod1的中和效应的经典重组产生的fab片段或单克隆抗体在导致肿瘤凋亡的细胞内ros强制信号通路(rosforcedsignalpaths)再活化中大超过100倍的效力。针对过氧化氢酶或sod的单域vhh片段的应用再激活了细胞间ros信号传导并导致细胞凋亡。尽管过氧化氢酶和sod的保护作用部分是冗余的和相互关联的,针对两种保护性酶之一的单域vhh片段足以消除肿瘤细胞免受ros信号传导的保护。这是基于这一事实,即sod仅实现部分保护作用,但是抑制sod导致间接对过氧化氢酶的抑制,这通过现在以增加的浓度存在的超氧化物阴离子实现。由抑制sod或过氧化氢酶引起的信号通路的质量是不同的。过氧化氢酶的抑制允许no/过氧亚硝酸盐(peroxynitrite)和hocl通路的顺序激活,而抑制sod仅导致再激活no/过氧亚硝酸盐路径。下述发现是高度治疗性意义的,即针对sod和过氧化氢酶的单域vhh片段协同合作,以及在优选实施方案中的所述协同作用可以集中在杂合分子中。在此基础上,公开了用结合sod或过氧化氢酶或这两种抗原的抗原结合构建体的肿瘤疗法的有利形式。
发明背景
仅在几年前才发现,活性氧和氮种类(氮类物质)(一起简称为活性氧种类=“ros”)除了它们的非定向诱变作用之外也可以发挥特定的信号功能(baueretal.,chimica,62,1-9,2008)。所述发现的看法尤其由于下述实情而变得复杂,即通过某些ros,经常可以引起相反的生物学效应。
ep11170076.1公开了抑制肿瘤细胞的保护性膜过氧化氢酶导致细胞内ros信号传导的再激活,并因此导至肿瘤细胞的选择性细胞死亡。由于在sod的抑制之后,由相邻的膜nadph氧化酶产生的超氧化物阴离子也不再转化为h2o2,也可以通过抑制膜sod来间接实现对保护性过氧化氢酶的抑制。以这种方式,现在存在足够高局部浓度的游离超氧化物阴离子,其将过氧化氢酶的活性中间体化合物i(catfeiv=o.+)通过单电子转移而转化为无活性中间体化合物ii(catfeiv=o),并另外将活性过氧化氢酶(catfeiii)转化为无活性化合物iii(catfeiiio2.-)。通过这两个反应,有效地抑制了过氧化氢酶的活性(konoyandfridovichi:superoxideradicalinhibitscatalase.jbiolchem257:5751-5754,1982.shimizun,kobayashikandhayashik:thereactionofsuperoxideradicalwithcatalase.mechanismoftheinhibitionofcatalasebysuperoxideradical.jbiolchem259:4414-4418,1984)。
然而,对于ros在多阶段肿瘤发生中的作用,现在可以画出一个相当清晰的图画,其知识可以显示建立新的选择性肿瘤治疗方法的前进之路(综述于bauerg.tumorcellprotectivecatalaseasanoveltargetforrationaltherapeuticapproachesbasedonspecificintercellularrossignaling.anticancerres.32:2599–2624,2012;bauerg.targetingextracellularrossignalingoftumorcells.anticancerres.34:1467-1482,2014;bauerg.increasingtheendogenousnolevelcausescatalaseinactivationandreactivationofintercellularapoptosissignalingspecificallyintumorcells.redoxbiol.6:353-371.2015):
1)ros的诱变作用有助于“肿瘤起始”的经典步骤。
2)ros对“肿瘤促进”的步骤至关重要,没有详细解释根本的机制。因此,大多数已知的肿瘤促进剂诱导增加的ros水平,并且ros的特异性清除抑制大多数肿瘤促进剂的作用。
3)细胞的恶性转化经常导致膜nadph氧化酶(nox1)的表达。该酶的连续活性对于恶性细胞的增殖和维持转化状态至关重要。nox1产生细胞外超氧化物阴离子,其歧化产物h2o2代表细胞的自分泌增殖刺激剂。
4)然而,细胞外超氧化物阴离子的产生也代表了形成选择性诱导转化细胞凋亡的细胞内ros控制信号通路的基础。这里,下面详细说明的“hoc1信号路径”和“no/过氧亚硝酸盐信号路径”是特别重要的。
因此,将转化细胞中特异性ros依赖性细胞凋亡诱导作为恶性细胞的潜在消除机制加以讨论。
5)肿瘤的成功发展需要产生ros的恶性细胞保护其自身免受ros信号传导通路的破坏作用,从而并不影响其增殖所必需的自分泌ros产生。这导致了规律以及迄今为止所研究的所有肿瘤系统中发现的肿瘤细胞的表型,并且其特征在于膜过氧化氢酶的组成型nox1活性和平行活性。另外,通过膜超氧化物歧化酶(sod)发生进一步的ros信号传导的调节。
6)这些膜保护酶的选择性抑制允许在肿瘤细胞中的ros依赖性选择性凋亡诱导,并因此具有很大的治疗潜力。
考虑到由于相互作用而总体来说相当复杂和难以预测的上面简要概述并在下面进一步解释的机制,本发明公开了具有广泛意义的抗原结合构建体,其特别可用于肿瘤疗法。因此,本发明的目的是抗原结合构建体(即,单域vhh片段,还称为纳米抗体),其特异性结合并优选抑制超氧化物歧化酶或过氧化氢酶或前述两种酶,即超氧化物歧化酶和过氧化氢酶。
wo2014/191493公开了针对sod1的单域抗体以及它们在治疗als(肌萎缩性侧索硬化)中的用途。这里,als患者中致病性sod1的水平由针对sod1的单域抗体降低以达到积极效果。
根据本发明的抗原结合构建体是针对过氧化氢酶或sod的单域vhh片段。这里,尽管由于vhh和fab片段的不同结构(一条链对两条链)(预期只有50%的浓度差异),相同的凋亡引起效果是用相当的常规fab片段浓度的不到百分之一来实现的。对于所有过氧化氢酶或sod中和单域vhh片段发现功效的此显著差异,并因此代表其特征性特点。这指示常规fab片段和单域vhh片段的键动力学与键可逆性之间在片段效果以及因此还在其极大改善的治疗性采用可能性上一定存在到目前为止还未认识到并且无法预测的基本差异以及深远的后果。作为解释该发现的基础,可以使用通过经典fab片段(由免疫球蛋白分子的重链和轻链的抗原结合部分组成)和通过单域抗体的vhh片段(其仅由重链的抗原结合部分组成)的表位鉴定之间的差异。虽然经典的fab片段通过由fab片段的空间结构和表面电荷定义的键结合特异性表位,并且在一定程度上包括它,单域vhh片段具有插入形状结构(plug-shapedstructure),其适合于靶分子的就形状和电荷而言互补的凹部(muyldermanss.nanobodies:naturalsingledomainantibodies.ann.rev.biochem.82:775-797,2013)。假设有效抑制过氧化氢酶或sod的本申请中公开的单域vhh片段结合到这些酶特征性的漏斗(funnel)中,并将底物分子传递到活性中心。可以想象,在此类结合之后,诱导酶的构象变化,这有效地防止了逆反应,即终止单域vhh片段与抗原之间的键。通过这种方式,由于没有发生逆反应,由直接和逆反应的反应常数确定的生物物理参数“亲和力”大大增加。在应用实践中,通过这种方式,单域vhh片段比传统的fab片段实现了更有效的中和效果,尽管它们的结合应该以相同的效率进行。单域vhh片段在抑制特异性酶中的这个基本优势尚未被描述,并且是令人惊奇的。
不能排除某些单域vhh片段通过它们的结合发生在酶中不同于活性中心的位置的事实来实现其对过氧化氢酶或sod的抑制作用,但是以这种方式通过蛋白质中已知的变构的远距离效应实现对酶活性的抑制作用。由于根据本发明的结果,也对于一系列效果的这种变化,可以假设单域vhh片段对引起变构抑制的酶上的区域的亲和力必须显著高于通过经典fab片段可以实现的亲和力。因此,本发明不限于直接结合酶的活性中心的单域vhh片段。
关于人造产生的抗体片段的结构,应该指出,同时存在着庞大的各种不同的结构。在holligeretal.(2005),naturebiotechnology,vol.23,no.9,第1126-1136页的文章中,已经说明和解释了抗原结合分子的各种结构。除了根据本发明使用的vhh单域片段,还有许多其他的结构,如fab片段、单链fv、双价抗体(diabodies)、微型抗体(minibodies)、三价抗体(triabodies)等。
根据本发明使用的单域抗体片段衍生自骆驼科(骆驼,羊驼(llama),羊驼(alpaca))的抗体分子。天然表型中的这些抗体具有fc部分(具有ch2和ch3),但由两条重链组成,每条重链又具有含cdr的vhh部分。轻链不具有骆驼科的这些抗体。这些vhh部分被称为单域vhh片段或还称为纳米抗体。
根据本发明的单域vhh片段可以原样使用或也可以化学修饰以实现特定的目的。通过共价化学键或通过其它化学相互作用(例如离子相互作用或范德华力)的根据本发明的单域vhh片段可以与具有有利的作用的其他成分连接。这些可以是与可以结合特定细胞的生物活性分子如配体和/或受体的缀合物。根据本发明的单域vhh片段也可能与成像材料连接,如放射性核素、产生颜色的酶等。根据本发明的分子也可以与导致体内延长的保留时间的分子相连接;这可以例如通过将它们连接到较大的聚合物(如例如聚乙二醇)来实现。在eyeretal.,veterinarnimedicina(2012),9,第439-531页的综述文章中公开了单域vhh片段的各种可能的应用选项和修饰。明确引用该参考文献。
在另一优选实施方案中,有代表性的杂合分子,其“自身浓缩”协同效应,尽管由于结构,可以排除同一个杂合分子同时结合过氧化氢酶和sod。这将这些杂合分子从经典的双特异性抗体的效应分开,所述双特异性抗体通常必须结合两个靶结构以达到预期的效果。
本发明的主要方面在于抗原结合构建体不仅特异性地结合各自的靶分子(超氧化物歧化酶和过氧化氢酶),而且还抑制它们。所述方面通过本领域技术人员熟知的标准方法进行测试。在合适的测试模型中,例如在细胞表面表达期望的靶分子(过氧化氢酶或超氧化物歧化酶)的肿瘤细胞,或在加入了纯化的sod或过氧化氢酶的测试系统中,添加用于各种酶的底物或由细胞产生。然后,通过适当的测量方法测量离析物(educts)的消耗或产物的产生。这里,还可以有使用生物过程(如例如肿瘤细胞的ros依赖性凋亡的诱导)作为参数的应用方法。所述一般相当灵敏的生物学方法的意义的先决条件在于被测试酶降解的底物在可控和定量的情况下具有测量的生物学效应。对于sod和过氧化氢酶尤其如此。例如,sod将细胞内ros信号传导所需的超氧化物阴离子代谢为h2o2,并因此防止hocl依赖性凋亡诱导所需的超氧化物阴离子和hocl之间的相互作用。
因此,可以通过这一事实,即通过采用外源添加hocl的肿瘤细胞的凋亡诱导作为测试系统来确定单域vhh或fab片段(比较)介导的sod抑制。hocl以基于hocl和超氧化物阴离子之间的相互作用的浓度依赖性方式诱导凋亡,所述相互作用导致形成攻击性羟基自由基(bauerg.hocl-dependentsingletoxygenandhydroxylradicalgenerationmodulateandinduceapoptosisofmalignantcells.anticancerres33:3589-3602,2013)。所述凋亡诱导被以适当浓度外源性加入的sod完全抑制。即,可以通过检查凋亡的抑制是否被待测单域vhh或fab片段再次消除来确定单域vhh或fab片段的中和作用。所述测试系统的定量使用需要连接技术人员熟悉的几个简单的步骤。在第一步中,确定在合理的试验时间(1-2小时)内产生显著可定量的细胞凋亡信号的市售hocl(或在培养基中充分形成hocl的次氯酸钠)的浓度。经验显示,0.1-1mm的hocl浓度是非常适合的。适当地,参数是凋亡细胞的百分比(通过相差显微镜直接测定)或(3-(4,5-二甲基噻唑-2-基)-2,5-二苯基四唑溴化物(mtt)测试,该测试是市售的并且通过测量线粒体代谢活性来确定细胞活力。在第二步中,测定sod的浓度,所述浓度仅仅足以完全抑制所选择的hocl浓度的影响。如经验所示,对此,sod浓度在5至50u/ml之间是合适的。所述测定的准确度是特别重要的,因为cu/zn-sod的hocl效应的抑制表示其自身作为最佳曲线,并且达到最佳抑制效果对测试的意义至关重要。一旦前两步已经令人满意地进行,就可以进行单域vhh或fab片段的抑制作用的实际测试。对此,将待测试的片段的所选浓度与在步骤#2中预先测定的sod的最佳浓度预孵育20分钟。随后,将所述混合物加入到测试细胞中。此后,加入hocl并测定凋亡诱导。如果将对sod的抑制效果消除了至少90%,则获得sod的最佳抑制,从而获得中和效果的明确结果,即实现了没有添加sod的凋亡诱导仅仅低10%的凋亡诱导。如果实现了特征在于实现无sod的比较值的凋亡诱导的至少50%的对sod的抑制作用,则对sod有显著的抑制作用。通过在随后的试验中增加单域vhh或fab片段浓度可以进一步检查此类发现的含义,因为在单域vhh或fab片段的浓度增加时sod显著抑制的情况下,预期增加凋亡诱导的恢复。
用于确定针对过氧化氢酶的单域vhh或fab片段的中和作用,基于h2o2的细胞凋亡诱导作用和过氧化氢酶h2o2降解的试验系统是合适的。这里,可以使用非恶性和恶性细胞两者作为细胞试验系统。仅需要在选择的细胞系统中凋亡诱导是可定量测定的。作为h2o2源,h2o2生成性葡萄糖氧化酶(gox)优于直接加入h2o2,因为通过gox连续产生h2o2(通过使用来自细胞的培养基的葡萄糖)允许非常精确调节h2o2通量。测试系统基于抑制通过由过氧化氢酶介导的gox的凋亡诱导以及通过过氧化氢酶-中和单域vhh或fab片段恢复凋亡诱导。在第一步中,测定了在适当的时间(1-2小时)内显著抑制凋亡的gox的浓度。对此,一般取决于细胞系统,需要0.1至40mu/ml的gox浓度。在第二步中,测定了人过氧化氢酶的浓度,所述浓度仅仅足以消除gox实现的效果。经验显示,其所需的浓度范围在小于20u/ml的过氧化氢酶的范围内。成功测定合适浓度的gox和过氧化氢酶后,然后可以进行用于测定单域vhh或fab片段的过氧化氢酶-中和效应的实际实验。对此,将所选择的过氧化氢浓度与所测试的单域vhh或fab片段的选定浓度预孵育20分钟。随后,将所述混合物与确定为合适的gox浓度一起加入到细胞中,并测量凋亡诱导。如果过氧化氢酶对gox介导的凋亡诱导的抑制作用被消除至少90%(即,如果实现不加入过氧化氢酶的对照制剂所示的凋亡诱导值的90%),则获得单区vhh或fab片段的可靠的中和作用。如果消除了过氧化氢酶的抑制作用至少50%,则可假设测试的单域vhh或fab片段具有显著的抑制作用。通过在随后的试验中增加所述片段的浓度,可以实现进一步检查这一发现。
作为此处列出的测试方法的对照,采用了结合完全不同抗原的抗原结合构建体。然后,在实际测试中,如上述详细所述,将每个待研究的构建体添加到反应混合物中,并且测定过氧化氢酶或超氧化物歧化酶的酶活性是否被抑制。以此目的,可以使用本领域的技术人员很容易熟悉的各种测试模型。该发现对于能够确定抗原结合构建体以何种方式结合靶酶(分别为过氧化物酶或sod)是重要的。
假设在大多数情况下,其中抗原结合构建体不结合催化中心或在其附近结合,酶活性不被或基本上不被抑制。然而,如果抗原结合构建体直接结合催化中心或在其空间附近结合,或者尽管结合远离催化中心,但的确通过变构效应改变靶酶的构象,使得酶的正常反应不能再自由顺其自然进行,反应物不能再与酶反应,并且不能再自由发生转化。由于生化系统绝对值的确定总是有差异的,优选在一个测试制剂中进行此类测试,其中处于相同状态和相同量的细胞经受不同的测试条件以能做出叙述(statement)。在本发明的含义中,如果通过加入适量的抗原结合构建体,相应酶的催化活性(或相应酶群体的总活性)降低至少50%,优选降低至少70%,和特别地优选降低至少90%,则获得超氧化物歧化酶和/或过氧化氢酶的抑制作用。
在本发明的另一实施方案中,提供了抗原结合构建体(优选纳米抗体),其结合过氧化氢酶和/或超氧化物歧化酶并且其不抑制或仅部分抑制这些酶。此外,可以有利地使用这些抗原结合构建体,所述构建体例如被配置为使得它们可以连接到或连接到用作标志物的部件。通过这种方式,例如可以标记肿瘤细胞,如果这是在合适的检测系统中提供相应信号的标志物的话。或者,这些构建体也可以与细胞毒剂连接或配置为使得它们可以与所述细胞毒剂连接或者任选地在目标生物体内将其自身与所述细胞毒性效应物连接。
在另一优选实施方案中,使用了混合物,其中一些抗原结合构建体抑制过氧化氢酶和/或超氧化物歧化酶并且其他构建体不抑制或仅部分抑制超氧化物歧化酶和/或过氧化氢酶。这些也可以是包含两种不同抗原结合构建体的杂合分子。
根据本发明的抗原结合单域vhh片段优选通过基因工程生产,并且不具有像完全抗体一样的fc部分。它们不同于经典的fab片段之处在于缺少轻链。即,它们仅是抗体重链的抗原结合部分。
在本发明的范围内,通过遗传工程制备并测序了各种优选的抗原结合片段。序列在本申请中公开。对于抗原结合序列特别重要的是构建体的cdr区。所谓的“互补决定区”(以下简称cdr)是免疫球蛋白可变链的非常特定的部分。所述cdr区嵌入免疫球蛋白的框架序列内,决定其特异性,并建立与免疫球蛋白结合的特异性抗原的接触。cdr区域是免疫球蛋白最可变的部分并且基本上有助于这些分子的种类。在具有重链和轻链的免疫球蛋白中有六个cdr区。然而,如果免疫球蛋白仅由一条链组成(如在根据本发明优选的单域vhh片段的情况下),则有三个cdr区域。
通常,重要的是如果要保持结合特异性,则cdr区几乎不变存在。然而,可能的是,轻微的突变不会不利地影响抗体结合构建体的功能。如果cdr的结构不受氨基酸交换的不利影响,则尤其如此。如果新插入的氨基酸与替代的氨基酸非常相似,则可以进行此类氨基酸交换。因此,在优选的实施方案中,抗原结合片段具有在本申请的范围内公开的cdr区或者它们最多在来自分别公开的cdr序列的较少数量的氨基酸上不同,其不显著改变或降低可结合性和结合特异性。
在本发明的范围内,公开了来源于此类构建体的抗原结合单域vhh片段的cdr区,所述构建体抑制或不抑制过氧化氢酶和/或超氧化物歧化酶,但的确结合它们。在优选的实施方案中,根据本发明的单域vhh片段(纳米抗体)含有至少一个,优选至少两个并最优选至少三个cdr区,其中特别优选那些来自抑制过氧化氢酶和/或超氧化物歧化酶的构建体的cdr区。
当它们倾向于治疗应用时,根据本发明的构建体可优选是人源化的。这里,框架序列由人框架物质代替或者将非人序列通过突变变成人序列,但保持结合特性。
对于开发治疗上可用的生物分子(这里指纳米抗体),修饰氨基酸序列经常是不可避免的。由于分子不来源于人而源自骆驼科,有可能或可能的是,产生针对外源表位的抗体。此类抗体反应将中和用于疗法中的抗体片段的效果。为避免这些困难,治疗上应用的分子是人源化的。抗体或抗体片段的人源化是在这一特殊领域中众所周知的技术。通常,寻找与原始分子具有最高可能相似性的人框架序列(主链)。然后,从原始纳米抗体上切出cdr区并移植入人序列中。这里,不排除必须进行氨基酸序列的某些改变。
重要的方面是,抗原上的与抗体的结合部分结合的位点由cdr序列定义。人源化过程中,可能需要稍微修饰三个cdr序列中的一个或两个以维持抗原结合部分的有利性质。因此,根据本发明的单域vhh片段的特征在于它们含有如本申请中公开的至少一个cdr序列,优选地两个和特别优选地三个cdr序列。
因此,本发明的目标是单域vhh片段,其具有至少一个,优选地至少两个和特别优选地至少三个以下以seqid编号为特征的cdr区。这些是具有seqid编号19、20、21、22、23、24、25、26、27、28、29,以及30的序列。进一步优选的源自结合sod的克隆的cdr序列是具有seqid编号31、32、33、34、35、36、37、38,和39的cdr。还优先的是具有seqid编号40、41、42、43、44、45、46、47、48、49、50,和51的cdr序列。
在另一优选实施方案中,根据本发明的构建体还可以提供有标志物。对于该应用,不必要的是构建体在其功能上抑制靶酶。相反,具有足够亲和力的特定键就足够了。此类结合可以在经典的elisa系统中测量,其中人过氧化氢酶或sod与合适的载体结合,并且通过常用检测方法检测酶和单域vhh片段的复合物。一方面,标志物可用于标记所需的肿瘤细胞,以便在此处得到诊断报告。另一方面,标志物也可以用于标记其他效应物的肿瘤细胞。例如,作为标志物,可以使用此类构建体,所述构建体将其它们自身与诱导肿瘤细胞凋亡的效应物连接。此类标志物的一个例子是与细胞毒性t细胞的cd3受体反应的抗cd3部分。例如此类抗cd3部分可以是针对cd3受体的抗体的抗原结合部分。
在本发明的另外的实施方案中,还可以配置本发明的抗原结合构建体使得其可以连接细胞毒剂。作为细胞毒剂,已知例如源自各种来源的各种毒素,例如细菌、真菌,或植物。优选地,这可以是霍乱毒素、肉毒毒素,或链球菌溶血素等。
在另外的实施方案中,根据本发明的抗原结合构建体可以通过键合的不同类型和可能性与细胞毒剂连接。这些还可以是化学键,如共价键以及离子相互作用或范德华力。
在另外的实施方案中,也可能的是,根据本发明的抗原结合构建体可以是放射性同位素或与放射性同位素紧密地连接。放射性同位素可用于肿瘤细胞的诊断检测或增强根据本发明的抗原结合构建体的细胞毒性作用,通过使放射性同位素与肿瘤细胞接近来实现。优选地,在治疗应用中,使用此类同位素,其仅相对较短辐射,以保持副作用尽可能低。此外,同位素的半衰期应相对较短以将身体和环境的负担保持在可接受的范围内。优选使用的是钇-90、铼-186或铒-169。
在另外的实施方案中,根据本发明的抗原结合构建体可以通过键合的不同类型和可能性与着色剂连接。这些可以是固体化学键如共价键以及离子相互作用或范德华力。可以通过不同的常用方法检测着色剂。此类构建体对于诊断目的应当是可用的。
在另外的实施方案中,公开了药物组合物,其含有根据本发明的至少一个抗原结合构建体。这些药物制备物优选用于治疗肿瘤疾病,特别是治疗胃癌(gastriccarcinoma)。
根据本发明的药物组合物含有至少一个特异性结合并抑制过氧化氢酶的抗原结合构建体。在另一个实施方案中,根据本发明的药物组合物含有至少一个特异性结合并抑制超氧化物歧化酶的抗原结合构建体。在另一个实施方案中,药物组合物含有结合过氧化氢酶和超氧化物歧化酶并抑制这两种靶酶的杂合分子。
令人惊讶并预测不到的是,杂合分子的协同效应专门再激活no/过氧亚硝酸盐通路,而单独的抗sod也再激活该通路,但抗过氧化氢酶诱导hocl通路。通过杂合分子的no/过氧亚硝酸盐通路的专门再激活代表未来应用的优势,因为应当通过额外的no代谢修饰来进一步优化不太复杂的no/过氧亚硝酸盐通路,特别是鉴于剂量效应关系的平台期的期望宽度。
通过抗sod或来自抗sod和抗cat的杂合分子的no/过氧亚硝酸盐通路的专门再激活的另一个优点在于,由于在no/过氧亚硝酸盐通路过程中的信号化学(图1-4),不应存在游离h2o2。这是期望的,因为h2o2对存活的肿瘤细胞具有不利于治疗效果的增殖刺激作用。ep11170076.1中没有公开这里公开的抗sod或来自抗过氧化氢酶和抗sod的杂合分子的治疗用途的优点的原理,并且具有令人惊讶的效果。该效果表示存在创造性。
在本发明的另一个优选实施方案中,本发明的至少一个抗原结合构建体与具有抗肿瘤活性的活性成分一起使用。已知具有抗肿瘤作用的各种活性成分。特别是,作为也可以存在于本发明组合物中的化学治疗剂,可以提到物质诸如泰素(taxol)、顺铂、内皮抑素、奥沙利铂、依托泊苷或秋水仙碱,仅举几例活性成分为例。优选使用泰素。
根据本发明的抗原结合构建体是可以通过遗传工程制备的单域vhh片段。
抗原结合构建体中个体片段的精确分子排列具有次要的意义,只要期望的功能(即结合和抑制过氧化氢酶和/或超氧化物歧化酶)得到保留即可。
用分子生物学手段制备根据本发明的抗原结合构建体。这些不是天然存在的抗体或通过简单(酶促)切割从中制备的fab片段。为了更好地了解,可以简要概述抗体的结构。例如igg型的抗体由两个fab片段和一个fc片段组成。每个fab片段由重链和轻链组成,其中重链可以分为可变部分(vh)和恒定部分(ch1)并且轻链可以分为可变部分(vl)和恒定部分(cl)。特别感兴趣的是可变部分vh和vl,其又含有与抗原结合有关的cdr(互补决定区)1-6。fab片段的结合特性由嵌入在空间上排列个体cdr区的框架结构中的cdr区决定。
抗原结合构建体是单域vhh片段(纳米抗体)。纳米抗体仅含有与结合相关的抗体的重链的部分,并且可以在细菌细胞中非常好地表达(muyldermanss.nanobodies:naturalsingle-domainantibodies.ann.rev.biochem.82:775-797,2013)。
在各种遗传工程方法的帮助下,可以制备一些抗原结合构建体。用于此的方法是相当多样的并且是本领域普通技术人员熟知的。通常,这里,进行以使得用所需的抗原免疫实验室动物(小鼠、大鼠、兔、鸡或骆驼、羊驼等)。由于除常规抗体外,骆驼和羊驼天然还具有专门由重链构成的igg,这些动物与已建立的选择性筛选方法结合使用导致获得编码单域抗体的核酸序列。然后,从合适的免疫细胞(如b细胞),可以分离使用适当方法(例如用所谓的噬菌体展示)进一步优化的核酸序列。然后,用这些方法获得特异性结合所需抗原的抗原结合构建体分子。在该语境中,特异性意指构建体尽可能地仅所寻求的结合分子,更优选地仅结合所述分子(sod或过氧化氢酶)的表位。非特异性交叉反应通常是不期望的。
所述抗原结合构建体的另一重要特性在于它们灵敏地结合所需的抗原。灵敏地意指已经在抗原结合构建体的非常低的浓度下,发生特异性结合所需抗原或所需表位。简言之,抗原结合构建体与目标抗原结合的越好则它越灵敏。由于单域vhh片段不包含像fab片段那样的某些表位,但由于它们的分子结构结合抗原的空间凹槽,这导致关于通过这些两个类型的单域vhh片段的某些表位的可检测性的实质差异。
根据本发明的抗原结合构建体特异性结合超氧化物歧化酶并抑制该酶。靶分子超氧化物歧化酶的抑制受到以下事实的影响:抗原结合构建体结合超氧化物歧化酶(sod)的催化活性中心或该催化中心的附近结合,从而实现酶的空间抑制。然后,超氧化物阴离子(sod的典型和特异性底物)不再能与酶结合且不能通过它被酶促转化为h2o2。
这适用于过氧化氢酶。根据本发明的抗原结合构建体特异性地并灵敏地结合过氧化氢酶并抑制它,使得h2o2向h2o+1/2o2,或者过氧化亚硝酸盐向no2-和1/2o2的酶促转化被抑制,并且防止了通过过氧化氢酶的活性中间体“化合物i”的no氧化。
在本申请中,使用以下列表中给出的缩写:
aebsf4-(2-氨基乙基)-苯磺酰氟、
(nadph氧化酶的抑制剂)
3-at3-氨基三唑
(过氧化氢酶抑制剂)
抗cat针对抗过氧化氢酶的抗体
抗sod针对sod的抗体
(为了图片中的空间,用acat和asod替代抗cat和抗sod的名称)
cat过氧化氢酶
化合物i公式catfeiv=o+.的过氧化氢酶和公式podfeiv=o.+的过氧化物酶两者的活化中间阶段。化合物i是在过氧化氢酶或过氧化物酶与一分子过氧化氢的反应中形成的。过氧化氢酶也可以与一分子过氧亚硝酸盐形成化合物i。
duox双重氧化酶
(由nadph氧化酶和过氧化物酶结构域组成的膜酶。过氧化物酶结构域在蛋白酶的帮助下被切除。)
fbs胎牛血清
fetpps5-,10-,15-,20-四(4-磺酸基苯基)卟啉铁(iii)氯化物
(过氧亚硝酸盐分解催化剂)
no一氧化氮
nod一氧化氮双加氧酶(将no氧化成硝酸盐)
nosno合酶
noxnadph氧化酶(这里,特别是膜nox-1)
pod过氧化物酶
(在本文中特别是某些过氧化物酶的能力发挥作用,其在过氧化氢存在下,它们能够将氯化物氧化成hocl)
pon过氧化亚硝酸盐
por细胞色素p450氧化还原酶
ras,rac癌基因
ros活性氧和氮种类
(自由基种类和非自由基种类,诸如超氧化物阴离子、羟基自由基、一氧化氮、过氧化氢、hocl、过氧亚硝酸盐等)
sirna小干扰rna
(特异性下调定义的基因产物合成的试剂)
sod超氧化物歧化酶
(这里,特别是sod-1(肿瘤细胞的活性中心中的cu++和来自细菌的mnsod,用于分析目的)
tgf-beta转化生长因子beta型
以下图解表示首先将转化的细胞中凋亡诱导的ros信号传导通路联合起来(方案1:hocl通路;方案2:no/过氧化亚硝酸盐通路)然后,显示膜过氧化氢酶和sod对这些信号通路的肿瘤细胞的特异性作用(方案3和4)。
附图简述
图1:方案1(hocl信号通路)显示恶性转化的以nox1和duox(由nox1相关nox结构域和过氧化物酶结构域组成)表达为特征的细胞的膜。这里,nox1的表达代表恶性细胞的特定特征,而duox也可以在正常细胞中检测到。通过基质金属蛋白酶(mmp)的作用,释放了过氧化物酶结构域①。由nox1生成的超氧化物阴离子②歧化为h2o2③,其由过氧化物酶用作底物④。这里从天然过氧化物酶(podfeiii)形成了能够将氯离子氧化成hocl⑤的反应性中间物“化合物i”(podfeiv=o+)。存在于微摩尔浓度范围内的hocl在它与超氧化物阴离子反应形成凋亡诱导的羟基自由基时会产生毒性作用⑥。这里,决定性的步骤是通过羟基自由基的脂质过氧化⑦,其经由形成神经酰胺,导致诱导细胞凋亡的线粒体通路(未示出)。如果相对于过氧化物酶有相对过量的h2o2,通过消耗hocl⑧或阻止其合成⑨的示于⑧和⑨中的反应可能导致hocl信号通路的终止。
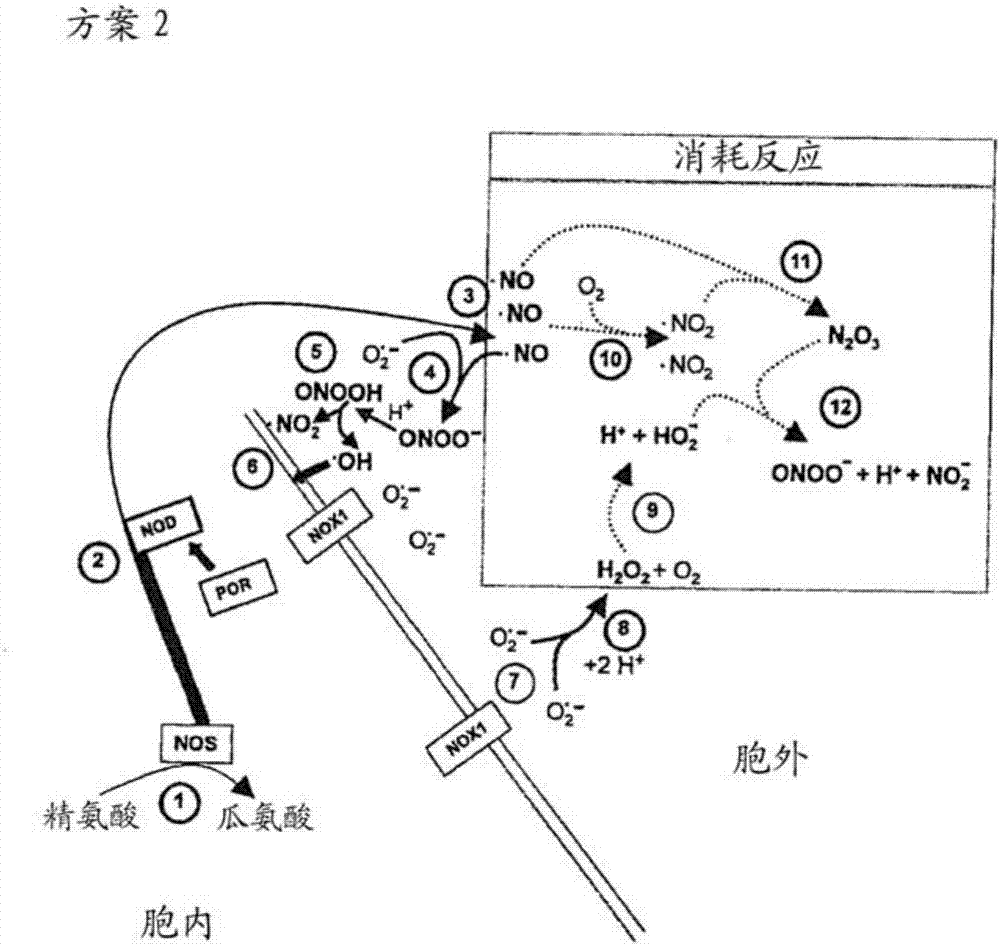
图2:方案2显示了转化细胞的no/过氧亚硝酸盐信号通路。no合酶(nos)产生一氧化氮(no)①。相当大的一部分的no由no双加氧酶(nod)转化为硝酸盐②。这里通过细胞色素p450依赖的氧化还原酶(por)来控制nod。no展示出高膜渗透性③并能够与转化细胞外部产生的超氧化物阴离子反应生成过氧化亚硝酸盐(onoo-)④。由过氧亚硝酸盐质子化形成的过氧亚硝酸(onooh)⑤极快地分解为no2和凋亡诱导羟基自由基⑥。反应顺序⑦-
图3:方案3显示了肿瘤细胞通过表达膜过氧化氢酶(cat),通过抑制hocl合成,通过破坏h2o2③来有效抑制hocl信号通路①,②,④-⑥。当然,膜sod促进超氧化物阴离子对h2o2的歧化,这可以影响hocl通路,但该效应不会带来由降解所述h2o2的过氧化氢酶的效果得到的结果。然而,sod介导的超氧化物阴离子浓度的降低抑制hocl和超氧化物阴离子之间的相互作用⑤,其对于信号通路是必不可少的,并且通过这种方式增强针对ros信号传导对肿瘤细胞的保护。抑制作用的分析表明,肿瘤细胞膜上的过氧化氢酶足以完全且明显抑制hocl合成,而当然单独的sod效果提供了明显的可测量的、但仅仅部分的保护。
图4:方案4总结了在保护肿瘤细胞免受no/过氧亚硝酸盐信号通路中膜过氧化氢酶和sod的复杂相互作用。过氧化氢酶通过将no氧化成no2来防止过氧亚硝酸盐的形成⑤并且通过降解来破坏可能形成的过氧亚硝酸盐⑥。sod通过清除超氧化物阴离子来防止过氧亚硝酸盐的形成④并且还能够将过氧化亚硝酸盐破坏
在下面的概略图(diagrammaticrepresentations)5(图5)和6(图6)中,总结了了解保护性酶过氧化氢酶和sod的多重作用所需的酶学细节。
图5:方案5显示了通过过氧化氢酶的hocl信号通路①的抑制由两阶段机制②,③实现,其中过氧化氢酶中间体“化合物i”(catfeiv=o+)是至关重要的,并重新形成为天然酶。一方面,no/过氧亚硝酸盐通路④的抑制通过经由在化合物i的参与下的两阶段机制的过氧亚硝酸盐的降解而发生,另一方面通过no的氧化⑧而发生。no的氧化与过氧化氢酶的no依赖性抑制相平衡,然而其仅在较高的no浓度下才起作用。
图6:方案6显示了sod介导的超氧化物阴离子对h2o2的歧化①,②由两阶段机制实现,其中酶结合的cu++的还原以及cu+的氧化起关键作用。超氧化物阴离子的反应抵消了hocl和超氧化物阴离子之间的相互作用③,其是hocl通路和过氧亚硝酸盐形成④所必需的。此外,sod具有通过通过反应步骤⑦-⑨来破坏过氧亚硝酸盐的潜力。
方案5和6显示了过氧化氢酶和sod(与教科书的知识相悖)不以高选择性反应为特征,而是能够执行多个、部分重叠的功能。总而言之,这导致出色的可塑性和复杂的生物学效应。
图7:方案7呈现了作用于肿瘤细胞外部的酶之间的总体背景。膜nadph氧化酶(nox)产生对这两个信号通路至关重要的超氧化物阴离子。膜sod将大部分超氧化物阴离子转化为h2o2①,②。这导致过氧亚硝酸盐形成的部分抑制③以及对超氧化物阴离子和hocl之间的相互作用的部分抑制④,其中hocl通过过氧化物酶(pod)形成⑤。此外,sod还分解过氧亚硝酸盐(反应⑥-⑧的顺序)。过氧化氢酶通过h2o2降解⑨来阻止hocl合成,降解过氧化亚硝酸盐⑩,并通过no的氧化阻止过氧亚硝酸盐的形成
总结在概略图7中的事实得出了在概略图8(图8)中呈现的结论,根据概略图8,仅抑制膜过氧化氢酶⑨,⑩能够导致hocl信号通路和no/过氧亚硝酸盐通路的再激活,因为sod仅实现了步骤③和④的部分保护。
另一方面,首先不会假设sod的抑制应当对于凋亡诱导信号通路的再激活是足够的,因为过氧化氢酶的主要抑制作用应与之相反。然而,如方案9(图9)中所揭示的,这是不正确的。理解这一初步意料不到的发现的关键是超氧化物阴离子对过氧化氢酶的抑制作用。由于在sod的抑制之后,观察到超氧化物阴离子的浓度局部增加(这仅被自发的歧化反应所抵消),如果仅直接抑制sod,则实现对过氧化氢酶的间接抑制作用。
在方案10(图10)中阐述了由超氧化物阴离子介导的过氧化氢酶抑制的酶学基础。一方面,超氧化物阴离子能够将天然过氧化氢转化成无活性化合物iii(catfeiiio2.–)①,另一方面,超氧化物阴离子能够通过单电子跃迁将化合物i(catfeiv=o.+)转化为无活性化合物ii(catfeiv=o)并且以这种方式基本上抑制酶的作用。因此,应理解,sod介导的超氧化物阴离子浓度的降低低于抑制作用所需的浓度有助于建立过氧化氢酶活性(参见图11,方案11)。另一方面,立即认识到,sod的直接抑制必然涉及过氧化氢酶的间接抑制(图12,方案12)。
通过以下实施例详细说明本发明。根据本发明的实验结果通常在图中示出。公开了特别优选实施方案中的序列。
实施例1:构建体和材料的提供
使用以下抗体,fab片段或单域vhh片段进行以下实施例:
1)针对人sod-1的单克隆抗体(小鼠,igg1)(克隆sd-g6)(收费号码035k4823)。制造商sigmaaldrich,schnelldorf,德国(作为对照)。
2)针对人过氧化氢酶的重组人fab片段,形式fab-v5sx2,自
3)针对人sod的重组人fab片段,形式fab-v5sx2,自
4)针对人过氧化氢酶的重组单域vhh片段(根据本发明),与商业供应商合作准备。
通过如下获得制备物:用人过氧化氢酶(过氧化氢酶[ec1.11.16],其纯化自人红细胞,获得自sigma(schnelldorf),目录号c3556)在兽医监督下免疫羊驼,获得来自动物b细胞的rna,反转录,克隆进大肠杆菌并经由噬菌体展示技术分离。通过在合适的elisa中检测上清液来选择编码结合人过氧化氢酶的单域vhh片段的克隆。在第二次运行中,通过采用heinzelmann和bauer(heinzelmanns.andbauerg.multipleprotectivefunctionsofcatalaseagainstintercellularapoptosis-inducingrossignalingofhumantumorcells,biol.chem.391,675-693,2010)描述的细胞培养系统,检查结合过氧化氢酶的单域vhh片段中的哪一个实际上导致过氧化氢酶的抑制,这在细胞中表示为ros依赖性细胞凋亡诱导。使用单域vhh片段acatcb0972,acatcb0974(两者均结合人过氧化氢酶并中和它),以及acatcb0973和acatcb0975(结合人过氧化氢酶,但不中和它)。通过标准方法对作为单域vhh片段的基础的克隆测序,并且由此确定氨基酸序列。
本发明的优选实施方案具有以下序列。
针对dna和蛋白质水平分析抗原结合片段的序列,并且测定了抗原结合区(cdr)。在下文中仅描述了氨基酸序列。这些抗原结合区域对于抗原结合片段的特异性是重要的。
在本发明的优选实施方案中,抗原结合构建体,特别是单域vhh片段或者纳米抗体,含有至少一个cdr序列,优选至少两个和更优选三个cdr序列。
下面,描述了蛋白质水平的cdr序列。在完整的序列中,各个位置由下划线给出。
cb0972(结合并中和过氧化氢酶):
蛋白质序列(seqidno:10):
maqvqlvesggglvqaggslrlscaasertfntygmgwfrqapgkerefvatiswsgdstyyadsvkgrftisrdnakntmylqmnslkpedtavyycnanseygdsywgqgtqvtvsskkkhhhhhh
蛋白质水平的cdr序列:
cdr1:rtfntygmg(seqidno:19)
cdr2:tiswsgdstyyadsvkg(seqidno:20)
cdr3:nseygdsy(seqidno:21)
cb0973(结合过氧化氢酶,但不中和):
蛋白质序列(seqidno:11):
maevqlvesggglvqpggslrlscavsgfifntysmrwgrqapgkglewvssistggystyadsvkgrftisrdnaknlvylqmnslkpedtavyycgwgafvrgerpqgqgtqvtvsskkkhhhhhh
蛋白质水平的cdr序列:
cdr1:fifntysmr(seqidno:22)
cdr2:sistggystyadsvkg(seqidno:23)
cdr3:gafvrgerp(seqidno:24)
cb0974(结合并中和过氧化氢酶):
蛋白质序列(seqidno:12):
maqvqlvesggglvqpggslrlscaasgsifsiasmgwyrqapgkqrdlvatitsdgstkyadsvkgrftisrdnakntmylqmnsvkpedaavyycnadaddlepgsydydywgqgtqvtvsskkkhhhhhh
蛋白质水平的cdr序列:
cdr1:sifsiasmg(seqidno:25)
cdr2:titsdgstkyadsvkg(seqidno:26)
cdr3:daddlepgsydydy(seqidno:27)
cb0975(结合过氧化氢酶,但不中和):
蛋白质序列(seqidno:13):
maqvqlvesggglvqpggslrlscaasasifsiyvmawyrqapgkqrelvatvtsggatnyansvkgrftisrdnakntmdlqmnslkpedtavyycnaedyydyglsrskiywgqgtqvtvsskkkhhhhhh
蛋白质水平的cdr序列:
cdr1:sifsiyvma(seqidno:28)
cdr2:tvtsggatnyansvkg(seqidno:29)
cdr3:edyydyglsrskiy(seqidno:30)
5)针对人sod1的重组单域vhh片段,与商业供应商合作准备。
通过以下获得制备物:用人sod1(sod1=cu/znsod[ec1.15.1.1],其纯化自人红细胞,获得自sigma(schnelldorf),目录号s9636)在兽医监督下,免疫羊驼,获得来自动物b细胞的rna,反转录,克隆进大肠杆菌并经由噬菌体展示技术分离。通过在合适的elisa中检测上清液来选择编码结合人sod1的单域vhh片段的克隆。在第二次运行中,通过采用heinzelmann和bauer(heinzelmanns.andbauerg.multipleprotectivefunctionsofcatalaseagainstintercellularapoptosis-inducingrossignalingofhumantumorcells,biol.chem.391,675-693,2010)描述的细胞培养系统,检查结合sod的单域vhh片段中的哪一个实际上导致sod的抑制,这在使用的肿瘤系统中表示为ros依赖性细胞凋亡诱导,因为通过抑制sod,游离超氧化物阴离子的浓度显著增加(由于缺乏酶促歧化),并导致过氧化氢酶的平行间接抑制。以下,这表示为ros依赖性细胞凋亡诱导。在进一步的对照试验中,如bauer2013(hocl-dependentsingletoxygenandhydroxylradicalgenerationmodulateandinduceapoptosisofmalignantcells.anticancerres33:3589-3602,2013)中描述的,通过以下事实验证了通过重组单域vhh片段的sod的特异性抑制:检查了这些片段对通过外源性添加的hocl的凋亡诱导的增加作用。使用结合并中和人sod1的单域vhh片段asodcb0989,以及结合人sod1但不中和它的片段asodcb0987和asodcb0991
通过标准方法对作为单域vhh片段基础的克隆测序,并且由此确定氨基酸序列。本发明的优选实施方案具有以下序列:
抗sodvhh:
cb0987(结合sod,但不中和):
蛋白质序列(seqidno:14):
maqvqlvesgggivqpggslrlscvasesiseidamywhrqapgkerelvagitndgtryyadsvkgrftisrdnakstlylqmnslkfedtamyycaalpnpppgywgqgtqvtvsskkkhhhhhh
蛋白质水平的cdr序列:
cdr1:siseidamy(seqidno:31)
cdr2:gitndgtryyadsvkg(seqidno:32)
cdr3:lpnpppgy(seqidno:33)
cb0989(结合并中和sod):
蛋白质序列(seqidno:15):
maqvqlvesggglvqsggsltlsctasgftisnypmtwvrqapgkglewvsrinsggdrtlyadsvkgrftvsrdnarntmylqmnnlkpedtglyfcadsgagwrywgqgtqvtvsskkkhhhhhh
蛋白质水平的cdr序列:
cdr1:ftisnypmt(seqidno:34)
cdr2:rinsggdrtlyadsvkg(seqidno:35)
cdr3:sgagwry(seqidno:36)
cb0991(结合sod,但不中和):
蛋白质序列(seqidno:16):
maqvqlvesgggivqpggslrlscvasesisdidamywhrqapgkrrelvagitndgteyfadsvkgrfaisrdntksslylqmnslkledtamyycatlpnpppgywgqgtqvtvsskkkhhhhhh
蛋白质水平的cdr序列:
cdr1:sisdidamy(seqidno:37)
cdr2:gitndgteyfadsvkg(seqidno:38)
cdr3:lpnpppgy(seqidno:39)
通过经由接头来连接克隆cb0972(中和过氧化氢酶)和cb0989(中和sod),通过遗传工程化制备了双特异性杂合单域vhh片段cb1081(抗cat抗sod)和cb1082(抗sod抗cat)。
双特异性抗过氧化氢酶-sodvhh
cb1081:
蛋白质序列(seqidno:17):
maqvqlvesggglvqaggslrlscaasertfntygmgwfrqapgkerefvatiswsgdstyyadsvkgrftisrdnakntmylqmnslkpedtavyycnanseygdsywgqgtqvtvssggggsggggsggggsaqvqlvesggglvqsggsltlsctasgftisnypmtwvrqapgkglewvsrinsggdrtlyadsvkgrftvsrdnarntmylqmnnlkpedtglyfcadsgagwrywgqgtqvtvsskkkhhhhhh
蛋白质水平的cdr序列:
catcdr1:rtfntygmg(seqidno:40)
catcdr2:tiswsgdstyyadsvkg(seqidno:41)
catcdr3:nseygdsy(seqidno:42)
sodcdr1:ftisnypmt(seqidno:43)
sodcdr2:rinsggdrtlyadsvkg(seqidno:44)
sodcdr3:sgagwry(seqidno:45)
双特异性抗sod-过氧化氢酶vhh
cb1082
蛋白质序列(seqidno:18):
maqvqlvesggglvqsggsltlsctasgftisnypmtwvrqapgkglewvsrinsggdrtlyadsvkgrftvsrdnarntmylqmnnlkpedtglyfcadsgagwrywgqgtqvtvssggggsggggsggggsaqvqlvesggglvqaggslrlscaasertfntygmgwfrqapgkerefvatiswsgdstyyadsvkgrftisrdnakntmylqmnslkpedtavyycnanseygdsywgqgtqvtvsskkkhhhhhh
蛋白质水平的cdr序列:
sodcdr1:ftisnypmt(seqidno:46)
sodcdr2:rinsggdrtlyadsvkg(seqidno:47)
sodcdr3:sgagwry(seqidno:48)
catcdr1:rtfntygmg(seqidno:49)
catcdr2:tiswsgdstyyadsvkg(seqidno:50)
catcdr3:nseygdsy(seqidno:51)
nadph氧化酶抑制剂4-(2-氨乙基-苯磺酰氟(aebsf)、过氧化氢酶抑制剂3-氨基三唑(3-at)、hocl清除剂牛磺酸、单线态氧清除剂组氨酸、葡萄糖氧化酶(gox)获得自sigma(schnelldorf,德国)。
过氧亚硝酸盐和“过氧亚硝酸盐分解催化剂”(功能性过氧化亚硝酸盐清除剂)5-,10-,15-,20-四(4-磺酰基苯基)卟啉铁(iii)氯化物(5-,10-,15-,20-tetrakis(4-sulfonatophenyl)porphyrinatoiron(iii)chloride,fetpps)获得自calbiochem(merckbiosciencesgmbh,schwalbach/ts,德国)。
这些活性成分的精确描述见出版物heinzelmann和bauer(2010,multipleprotectivefunctionsofcatalaseagainstintercellularapoptosis-inducingrossignalingofhumantumorcells,biol.chem.391,675-693),以及bechtel和bauer(2009,catalaseprotectstumorcellsagainstapoptosisinductionbyintercellularrossignaling,anticancerres29:4541-4557)。
实施例2:用sirna的基因敲除
对于通过利用通过sirna技术的基因的特异性敲除的描述于图22中的分析,使用以下sirna(获得自qiagen,hilden,德国):
a.对照sirna(“sico”),(目录号1022076;序列:
r(uucuccgaacgugucacgu)dtdt(正义)(seqidno:1)
acgugacacguucggagaa)dtdt(反义)(seqidno:2)。
制造商发现sico不影响任何已知基因的表达。
b.对于以下敲除的“高性能验证的sirna(high-performancevalidatedsirna)”
fas受体(“sirnafasr.”)(hs_fas_7_hp验证的sirna,目录号si02654463;目标序列:aaggagtacacagacaaagcc)(seqidno:3);
caspase-8(“sirnacasp8”)(hs_casp8_11_hp验证的sirna;目录号si02661946,目标序列:aagagtctgtgcccaaatcaa)(seqidno:4);
caspase-9(“sirnacasp-9”)(hs_casp9_7_hp验证的sirna,目录号si02654610,目标序列:cagtgacatctttgtgtccta)(seqidno:5);
c:针对人nox1的hpsirna(“sirnanox1”);
目标序列:ccgacaaatactactacacaa(seqidno:6)
d:针对人inos2的sirna(siinos)
目标序列:ctgggccgtgcaaaccttcaa(seqidno:7)
对于示于图27中的对照测试,除了对照sirna以外,还使用了针对人过氧化氢酶的sirna(对于mkn-45细胞)
hs_cat_4_hpsirna,目录no.si00027713
目标序列:ccggatctcacttggcggcaa(seqidno:8)
和针对鼠过氧化氢酶的sirna(对于208f和208fsrc3细胞)
hpmm_cat_4_hpsirna,目录号si00941976
目标序列:cccaataggagataaacttaa(seqidno:9)。
heinzelmann和bauer(同上),2010中详细描述了通过sirna的转染技术。转染效率超过95%。转染后24小时,进行各基因功能的对照检查以确保完成“功能性敲除”。这意味着特定的sirna有效地抑制了分析的基因产物的从头合成,以及意味着在施用sirna之前基因产物的浓度的自然降解发生,直至低于检测限。
将人胃癌细胞系mkn-45保持在补充有10%灭活的胎牛血清和40u/ml青霉素、50μg/ml链霉素、10μg/ml新霉素、10u/mlmoronal(抗真菌抗生素)和280μg/ml谷氨酰胺的rpmi1640培养基中。将人神经母细胞瘤系shep以及正常大鼠成纤维细胞(208f)及它们由src癌基因(208fsrc3)转化的后代保持在补充有5%灭活的胎牛血清和40u/ml青霉素、50μg/ml链霉素、10μg/ml新霉素、10u/mlmoronal和280μg/ml谷氨酰胺的eagle最低必需培养基(emem)中。有关细胞系及它们的培养基的细节见于heinzelmann和bauer,2010以及的bechtel和bauer,2009的工作。
实施例3:用于分析肿瘤细胞中自分泌ros控制的凋亡诱导的标准制备
用基于在heinzelmann和bauer2010中进行的方法的凋亡诱导的标准制备来进行示于图13-24中的实验。使用的肿瘤细胞取自最佳生长的半密封培养物,离心并置于新鲜培养基中。该试验在具有12500个mkn-45细胞/100μl培养基或10000个shep细胞/100μl的培养基的96孔细胞培养板中进行。mkn-45细胞在悬浮液中生长,shep细胞是粘附的。一旦细胞生长,shep细胞的测试就开始了。通过添加增加浓度的抑制过氧化氢酶或sod的单域vhh片段,使肿瘤细胞免于其自身的ros信号传导的保护被消除,使得基于细胞拥有的细胞外超氧化物阴离子生成,no/过氧亚硝酸盐信号通路和hocl通路(用mkn-45细胞)或者在shep细胞的情况下仅no/过氧亚硝酸盐通路可以当细胞在37℃下温育时开始。信号通路的分析是通过应用nadph氧化酶抑制剂(aebsf)、hocl清除剂牛磺酸、过氧化亚硝酸盐清除剂fetpps以及单线态氧清除剂组氨酸来实施的。在一些用于比较的实验中,代替单域vhh片段,应用了重组fab片段或单克隆抗体。这在相应的图示中指出。
在本文所示的时间,通过用于凋亡细胞百分比的相差倒置显微术来检测双重制备物。这里,使用了heinzelmann和bauer2010中描述和记录了经典凋亡标准,如细胞核凝聚,细胞核的片段化,以及质膜出泡(membraneblebbing)。每一个制备物,检测了至少250个随机选择的细胞的凋亡特征的存在。
平行对照检测,例如如bauer等人的工作(bauerg,bereswills,aichelepandglockere.helicobacterpyloriprotectsoncogenicallytransformedcellsfromreactiveoxygenspecies-mediatedintercellularinductionofapoptosis,carcinogenesis35:1582-1591,2014,增刊)中记录的,确保应用的形态学标准与凋亡标准(如dna片段化(通过tunel反应测量))或对膜联蛋白v结合的阳性相关。
实施例4:用于通过针对过氧化氢酶的单域vhh片段的凋亡诱导ros信号传导的肿瘤细胞特异性致敏
在本发明的范围内,已经进行了许多试验,其中试验结果已经在图13至图30中进行了总结。
图13显示了mkn-45胃癌细胞中通过针对过氧化氢酶的单域vhh片段的特异性凋亡诱导。
将在用于自分泌凋亡诱导的标准条件下mkn-45细胞与给定浓度的结合但不抑制人过氧化氢酶的单域vhh片段(acatcb0973,acatcb0975)以及结合并抑制人过氧化氢酶的单域vhh片段(acatcb0972;acatcb9074)混合,并进一步在37℃,5%co2下温育3.5小时。在相同条件下的平行中,应用了针对并中和人过氧化氢酶的重组fab片段(由轻链和重链组成)(abdacat15562)。此后,根据上述经典凋亡标准测定凋亡细胞(双重制备物)的百分比。
图13a显示了仅抑制过氧化氢酶,而非使得结合酶而不抑制它的单域vhh片段诱导肿瘤细胞中的凋亡。这里,效果以最佳曲线的形式表示,正如heinzelmann和bauer,2010,对于过氧化氢酶抑制剂3-氨基三唑的工作所描述的。最佳曲线下降后,凋亡诱导第二次增加。由轻链和重链组成的重组fab片段abdacat15562的添加也导致最佳曲线形式的凋亡诱导及随后第二次增加,但是为了实现作用最佳值,需要与过氧化氢酶抑制单域vhh片段相比高约250倍的abdacat15562的摩尔浓度。这通过根据本发明的构建体的优异功效而明显。
胃癌细胞系mkn-45中抑制人过氧化氢酶的单域vhh片段和经典fab片段的添加导致相对于抗体浓度的最佳曲线形式的诱导凋亡。诱导过程的特异性由这一事实而明显,即结合过氧化氢酶但不中和它的单域vhh片段不导致凋亡诱导(图13)。即,对于致敏,抗体仅与过氧化氢酶结合是不够的。对过氧化氢酶的功能的特异性抑制似乎对致敏作用至关重要。值得注意的是和意想不到的是发现单域vhh片段比经典的fab片段具有更强的细胞凋亡诱导作用。由于这两组片段的结构不同,可以预期为了达到类似的效果,与单域vhh片段相比,经典的fab片段将以两倍浓度来应用。然而,该差异构成了约500倍(以摩尔计为250)。
实施例5:结合过氧化氢酶并抑制的单域vhh片段对胃癌细胞的影响
图14显示了mkn-45系的胃癌细胞中过氧化氢酶中和单域vhh片段acatcb0972经由no/过氧化亚硝酸盐和hocl通路诱导特异性ros信号传导。
对用于诱导凋亡的标准制备物,在100μmnox1抑制剂aebsf、50mmhocl清除剂牛磺酸(tau)、25μm过氧化亚硝酸盐清除剂fetpps或者2mm单线态氧清除剂组氨酸(his)的存在下,添加给定浓度的单域vhh片段acatcb0972。在没有抑制剂的情况下平行进行对照制备物。在37℃,5%co23.5小时后,测定了凋亡细胞的百分比。
图14显示了单域vhh片段acatcb0972(结合并抑制过氧化氢酶)以最佳曲线及随后第二次增加的形式诱导凋亡。这里,在单域vhh片段的整个浓度范围内,凋亡诱导被nox1抑制剂aebsf所抑制。hocl清除剂牛磺酸不会在最佳曲线左侧的浓度范围内抑制,但然后导致在最佳曲线的整个进一步范围内的凋亡的强烈抑制。在单域vhh片段的6.2pg/ml浓度范围内的凋亡的第二次增加不被牛磺酸抑制。过氧亚硝酸盐清除剂fetpps仅在左侧部分的最佳曲线中导致强烈的抑制作用,然后将凋亡诱导的最佳曲线变为平台曲线。单线态氧清除剂组氨酸不会导致细胞凋亡的抑制,但将最佳曲线变为平台曲线。
即,图14证明了单域vhh片段acatcb0972的凋亡诱导效果实际上由ros信号传导的再激活引起,因为在所有应用浓度下,当通过aebsf抑制超氧化物阴离子产生时,发生凋亡诱导的完全抑制。此外,图14中证明的结果显示了凋亡诱导的最佳范围(0-6.2pg/ml)的特征在于一系列no/过氧亚硝酸盐通路和hocl通路。在acatcb0972的0.09和0.18pg/ml的范围内,存在由过氧亚硝酸盐清除剂fetpps的抑制,而非被hocl清除剂牛磺酸的抑制,这说明no/过氧亚硝酸盐通路的过程。从0.75pg/ml的fab片段,没有fetpps的实质抑制,但是牛磺酸的抑制非常强烈,这证明了hocl通路的过程。如预期的,这两种信号通路在0.37pg/ml重叠。单域vhh片段在更高浓度的凋亡诱导的新的增加依赖于
i)超氧化物阴离子的产生,
ii)过氧化氢酶抑制程度以及不受hocl清除剂和过氧亚硝酸盐清除剂的影响。
因此,它是h2o2的唯一影响,而不是特定ros信号传导通路的影响。
通过单线态氧清除剂组氨酸和过氧亚硝酸盐清除剂fetpps将凋亡诱导的最佳曲线改变为平台曲线说明在单域vhh片段acatcb0872>0.75pg/ml的浓度下,单线态氧似乎也起作用。单线态氧可以由h2o2与过氧亚硝酸盐的反应产生。也已知单线态氧可以使过氧化氢酶无活性。然后,由此引起的h2o2的增加的可用性可以引起示于图1中的副反应⑧和⑨,其中hocl被消耗或较少的hocl被合成。以这种方式,引起最佳曲线右侧的凋亡诱导下降。
实施例6:用于通过针对sod的单域vhh片段的凋亡诱导ros信号传导的肿瘤细胞的特异性致敏
图15中示出了mkn-45胃癌细胞中通过针对sod的单域vhh片段的特异性凋亡诱导。凋亡细胞的百分比绘制在x轴上。
将在用于自分泌凋亡诱导的标准条件下的mkn-45细胞与给定浓度的结合但不抑制人sod1的单域vhh片段(asodcb0987,asodcb0991)以及结合并抑制人sod的单域vhh片段(asodcb0989)混合,并进一步在37℃,5%co2下温育3.5小时。在相同条件下的平行中,应用了针对并中和人过氧化氢酶的重组fab片段(由轻链和重链组成)(abdasod15660)。此后,根据上述经典凋亡标准测定凋亡细胞(双重制备物)的百分比。
图15a显示了抑制sod,而非使得仅与酶结合的单域vhh片段在肿瘤细胞中诱导凋亡。这里,该效果以宽泛的最佳曲线的形式表示。凋亡诱导的最佳曲线也通过经典fab片段abdasod15660实现(图15b),然而,对此,为了达到同样的效果,与单域vhh片段的浓度相比,需要高250倍的浓度。
即,图15显示了抗体结合sod不足以致敏。对sod的功能的特异性抑制似乎对致敏是至关重要的。值得注意的是,膜sod的特异性抑制足以再激活导致细胞凋亡的ros信号传导。值得注意和意想不到的是这一发现,即中和单域vhh片段具有比经典中和fab片段更强的细胞凋亡诱导作用。由于这两组片段的结构不同,可以预期的是,为了达到同样的效果,与单域vhh片段相比,将应用两倍浓度的经典fab。然而,差异是约250倍。
图16显示了mkn-45系胃癌细胞中sod中和单域vhh片段asodcb0989通过no/过氧亚硝酸盐通路仅诱导特定的ros信号传导。
对用于诱导凋亡的标准制备物,在100μmnox1抑制剂aebsf、25μm过氧化亚硝酸盐清除剂fetpps、50mmhocl清除剂牛磺酸(tau)或者2mm单线态氧清除剂组氨酸(his)的存在下,添加给定浓度的单域vhh片段asodcb0989。在没有抑制剂的情况下平行进行对照制备物。在37℃,5%co23.5小时后,测定了凋亡细胞的百分比。
图16显示了在mkn-45系胃癌细胞中,sod中和单域vhh片段以宽泛最佳曲线的形式诱导凋亡,然后是指定的第二次增加。aebsf在整个浓度范围内抑制凋亡诱导。整个最佳范围由fetpps抑制,但不被牛磺酸抑制。组氨酸导致最佳曲线右侧下降的可察觉的部分消除。
即,图16证明了肿瘤细胞中由asodcb0989诱导的凋亡确实经由超氧化物阴离子依赖性过程连续诱导,但这(与acatcb0972相反)仅是no/过氧亚硝酸盐通路且hocl信号没有明显的影响。通过fetpps方式的完全抑制和牛磺酸抑制的消失也证明了这一点。这一结果表明,鉴于缺乏sod效应,显然没有足够可用于hocl通路的h2o2。由于no/过氧亚硝酸盐通路被膜过氧化氢酶非常有效地抑制,可以从图16所示的结果得出结论,即通过单域vhh片段抑制sod也必然导致过氧化氢酶的间接抑制。由asodcb0989和acatcb0972再激活的信号通路的不同质量允许排除asodcb0989的作用可以是与过氧化氢酶的交叉反应的理论假设并因此也证实了asodcb0989的作用的特异性。
实施例7:单域vhh片段与目标细胞之间的关系
图17显示了针对过氧化氢酶或者sod的单域vhh片段对靶细胞的密度做出不同的要求。
除了标准细胞密度(12500细胞/100μl)外,还在低细胞密度(4000细胞/100μl)和增加浓度的单域vhh片段acatcb0972或者asodcb0989的情况下制备用于诱导凋亡的制备物,并且在37℃,5%co2温育4小时。此后,测定凋亡细胞的百分数。
图17显示了在最佳方式中由仅具有更高浓度运行的针对过氧化氢酶的单域vhh片段诱导的凋亡,其中在针对sod的单域vhh片段的应用中,当目标细胞以较低的密度存在时,效果的减弱要小得多。这种差异为再激活信号通路的不同质量提供了证据并且特别是与sod的抑制优选再激活no/过氧亚硝酸盐通路的事实相一致,因为这与hocl信号通路在相同程度上不依赖于高细胞密度。
实施例8:针对过氧化氢酶和sod的单域vhh片段的协同效应
图18显示了同时应用针对过氧化氢酶和sod的单域vhh片段的显著的协同效应。
将用于用mkn-45细胞的凋亡诱导的标准制备物与单独的以及与0.005pg/m的结合sod但不中和它的sod中和性单域vhh片段asodcb0989或者单域vhh片段asodcb0987组合的增加浓度的过氧化氢酶中和性单域vhh片段acatcb0972相混合(a)。在互补实验(b)中,用单独的或者与0.005pg/ml的过氧化氢酶中和性单域vhh片段acatcb0972组合的增加浓度的asodcb0989实现混合。所有制备物在37℃,5%co2下温育3.5小时。然后,测定凋亡细胞的百分数。
图18显示了可以通过acatcb0972和asodcb0989两者来确定已经在初步实验中测量的浓度依赖凋亡诱导效果。与单独不足以诱导凋亡的分别互补的单域vhh片段的微小(minor)浓度的组合(即asod在增加浓度的acat中以及acat在增加浓度的asod中)导致细胞凋亡诱导中的显著协同作用。所述协同效应的特异性由以下事实证明,即它在结合但不中和sod的单域vhh片段的施用的情况下尚未出现(图18a)。
即,图18显示了针对过氧化氢酶或者sod的单域vhh片段的组合导致非常强烈的协同作用,当最佳地使用这种效应时,单域vhh片段的所需浓度可以大大降低。
实施例9:与化学治疗剂的协同作用
图19显示了分别中和过氧化氢酶或者sod的单域vhh片段与建立的化学治疗剂泰素引起强烈的协同作用。
将用于用mkn-45细胞的凋亡诱导的标准制备物与单独的或者与过氧化氢酶中和性acatcb0972,sod中和性asodcb0989以及sod结合而非中和性asodcb0987组合的增加浓度的泰素混合并在37℃,5%co2温育4小时。然后,测定凋亡细胞的百分数。
图19显示了在肿瘤细胞中泰素以浓度依赖方式诱导凋亡。
这里,在1100ng/ml浓度范围内发现了最佳效果。这种效果是预期的。与可以抑制过氧化氢酶或者sod的单域vhh片段相组合,泰素依赖性最佳曲线出人意料地被迅速置换到较低的浓度范围。现在,效果最佳值为0.17ng/ml至0.5ng/ml之间。所需浓度至小于千分之一的这种令人印象深刻的置换只能通过还中和各自目标结构(过氧化氢酶或者sod)的单域vhh片段实现,而纯粹的结合没有引起任何增强的效果。
示于图19中的效果起初是非常感兴趣的,因为其证明了泰素对肿瘤细胞和ros信号传导的作用之间的以前未知的功能联系。与针对sod或者过氧化氢酶的单域vhh片段的协同效应的出现提供了通过适当地组合活性成分来保存总共资源的机会,从而获得治疗成本的降低,并且还开启减少或甚至避免由泰素引起的副作用并且避免了单域vhh片段可能的副作用的机会。
实施例10:通过来自过氧化氢酶或sod中和性单域vhh片段的杂合分子对肿瘤细胞的特异性致敏以实现凋亡诱导ros信号传导
图20显示了单域vhh片段acat和asod之间的协同效应,acat和asod可以浓缩为一个杂合分子。
将用于用mkn-45细胞的凋亡诱导的标准制备物与增加浓度的过氧化氢酶中和性acatcb0972、sod中和性asodcb0989、以两种可能排列的来自acatcb0972和asodcb0989的杂合分子以及作为对照的针对过氧化氢酶的中和性常规单克隆抗体(sigma)相混合,并在37℃,5%co2温育3.5小时。然后,测定凋亡细胞的百分数。
图20显示了用单独添加的单域vhh片段acatcb0972和asodcb0989的组合观察到的协同效应可以实际上浓缩为杂合分子。这里,排列acatasod似乎逐渐优于反向排列。虽然单独施用的单域vhh片段的最佳效果在约170fg/ml的浓度范围内,杂合分子的最佳效果已经在0.24fg/ml实现。这些值得注意的活性与在111ng/ml时形成最佳效果的常规单克隆抗体的显著降低的功效相冲突。在例示中,为了清楚起见,没有包括中和sod或过氧化氢酶的经典重组fab片段的数据。它们的最佳效果在0.1-0.3ng/ml的范围内。
即,图20证实了
i)单区vhh片段的效果出乎意料地远远大于常规单克隆抗体或常规重组fab片段的效果,以及
ii)当从单域vhh片段产生杂合分子时,可以获得效果上进一步非常清晰的增加。
实施例11:
图21显示了在mkn-45系的胃癌细胞中来自单域vhh片段acatcb0972和asodcb0989的杂合分子经由no/过氧亚硝酸盐通路仅诱导特异性ros信号传导。
对用于凋亡诱导的标准制备物,在100μm的nox1抑制剂aebsf、25μm的过氧化亚硝酸盐清除剂fetpps、50mm的hocl清除剂牛磺酸(tau)或者2mm的单线态氧清除剂组氨酸(his)的存在下添加给定浓度的杂合分子acatasod。在不含抑制剂的情况下平行进行对照制备物。在37℃,5%co2下3小时后,测定凋亡细胞的百分数。
图21证实了肿瘤细胞中杂合分子acatasod经由ros信号传导的再激活来诱导凋亡,因为通过抑制超氧化物阴离子的合成(通过aebsf的方式)完全阻止了该过程。此外,明显的是通过杂合分子再激活的凋亡诱导仅是no/过氧亚硝酸盐通路,因为存在有通过fetpps的完全抑制作用,但无受牛磺酸的抑制。单线态氧清除剂继而对最佳曲线的右侧下降具有减弱作用。no/过氧亚硝酸盐通路的主导性是从以前的结果看出人意料的和不可预测的。
实施例12:
图22通过基于sirna的分析确认了由杂合分子诱导的ros信号传导的特异性并导致凋亡。
用24nm针对nox1、inos2、caspase-9、fas受体或者caspase-8的sirna转染mkn-45细胞。用不相关的对照sirna转染对照制备物。37℃24小时后,将细胞在新鲜培养基中继续并与给定浓度的杂合分子acatasod相混合。4小时后测定凋亡细胞的百分比。
图22显示了仅当通过线粒体通路对凋亡过程至关重要的完整的nadph氧化酶(nox1)、no合酶(inos2)和caspase-9可用时才能发生以由杂合分子acatasod引起的最佳曲线形式的凋亡诱导。sirna介导的对这些酶的敲低各自完全阻止由杂合分子诱导的凋亡诱导,并因此证明了经由no/过氧亚硝酸盐通路的根本的ros信号传导及随后的线粒体凋亡信号传导。另一方面,由杂合分子诱导的凋亡不需要fas受体和其下游caspase-8,强调了线粒体凋亡通路的专门作用并且排除了通过用于在选择的条件下凋亡诱导的死亡受体apo/fas的凋亡通路发挥作用。然而,fas受体和caspase-8的敲低防止细胞凋亡诱导曲线的超最佳(supra-optimum)下降。这可以通过如bauer,2012中所述的fas受体和caspase-8和单线态氧相关过程的参与来解释,并且与图21中建立的关于组氨酸的发现一致。
从图13、图15和图20,确定了肿瘤细胞中最适凋亡诱导所必需的抗体和单域vhh片段的浓度(表1)。该表解释了与经典的重组fab片段和单克隆抗体相比,单域vhh片段的优异功效。表2强调通过使用杂合分子可以实现令人印象深刻的协同效应。单域vhh片段对经典重组fab片段的优越作用是不可预测的,并因此是预料不到的。相反,由于目前的知识状况,预期中和性经典fab片段和中和性单域vhh片段当考虑摩尔浓度时应当对凋亡诱导实现相同的效果,在考虑浓度(pg/ml)时,从而仅应当必须实现2倍的差异。这种不可预测的显著的效力差异是令人惊讶的。这种效果最适合治疗用途。
表1
表2
数值取自图13、图15和图20。首先,确定常规重组fab片段和单克隆抗体的哪个浓度量是更高的,以实现与重组单域vhh片段相同的效果。通过包括各种fab片段和抗体的摩尔质量,然后在摩尔浓度(molarity)的基础上确定关系。在该校正中,采用了有效的假设,即经典的fab片段占完整igg分子摩尔质量的三分之一,并且单域vhh片段的摩尔质量占经典fab片段的摩尔质量的大约一半。
实施例13:具有过氧化氢酶中和效果或者sod中和效果的单域vhh片段对仅能够建立no/过氧亚硝酸盐通路的人肿瘤细胞的效果
虽然用于之前实施例中的胃癌系mkn-45的特征在于当其膜过氧化氢酶被抑制时,其能够表达已知的细胞内ros信号传导(hocl和no/过氧亚硝酸盐通路作为主要通路,硝酰氯(nitrylchloride)通路作为次要通路)的整个谱(spectrum),在一系列其他人肿瘤细胞系中,显示出对no/过氧亚硝酸盐通路的限制(heinzelmann和bauer,2010;bauer,2012)。因此,在之前的检查中,发现特定类型的肿瘤各自显示一致的ros信号系统。迄今为止,我们仅对神经母细胞瘤、尤因氏肉瘤、乳腺癌、卵巢癌和小细胞肺癌观察到对no/过氧亚硝酸盐信号传导的限制。
图23显示了单域vhh片段也能够在仅可以形成no/过氧亚硝酸信号传导的肿瘤细胞中再激活凋亡。
将每100μl培养基10000个人成神经母细胞瘤系shep细胞与给定浓度的过氧化氢酶中和性单域vhh片段acatcb0972、sod中和性单域vhh片段asod0989、sod结合性而非中和性单域vhh片段asodcb991以及杂合分子acatasod相混合,在37℃,5%co2温育5小时,之后测定凋亡细胞的百分数。
图23显示了过氧化氢酶中和性单域vhh片段和sod中和性单域vhh片段以及杂合分子acatasod能够在神经母细胞瘤细胞系shep中诱导浓度依赖性凋亡。如所预期的,这里,杂合分子证明比每个仅针对一个目标结构的单域vhh片段有效得多。高浓度的sod结合性但非中和性单域vhh片段asodcb0991也实现了凋亡诱导效果,但对此,需要比中和性单域vhh片段高106倍的浓度。
即,图23也确认了过氧化氢酶抑制后仅有no/过氧亚硝酸盐信号传导能力的细胞系可以通过针对过氧化氢酶或sod的单域vhh片段引起细胞凋亡。此外,这里出现了acat和asos之间的协同效应。值得注意的是用该细胞系观察到的浓度-效应曲线,所述细胞系与mkn-45系相比,仅在右侧具有非常平坦的持久的超级最佳下降。
可以用以下事实最佳解释由仅能结合但不能中和的单域vhh片段asodcb0991实现的弱但显著的凋亡诱导:在单域vhh片段结合后,有sod的内化,并因此其在表面上的浓度是减少的,这将导致与抑制类似的效果。
图24显示了当另外使用no供体snp时,可以进一步优化asodcb0989对shep细胞的效果。
将每100μl培养基10000个人成神经母细胞瘤系shep细胞与给定浓度的sod中和性单域vhh片段asod0989相混合。在没有其他添加剂的情况下温育接受20μm或者100μmno供体硝普钠的平行制备物。37℃,5%co25小时后,测定凋亡细胞的百分数。
图24显示了额外施用no供体(单独不能诱导凋亡)导致关于凋亡诱导再激活的致敏并有效地抵消最佳曲线的超最佳右侧下降。从这之中,应当建立将凋亡诱导的最佳曲线转化为平台曲线的调节方法,这应该导致更大的治疗用途的确定性。
实施例14:单域vhh片段仅抑制膜过氧化氢酶(其对肿瘤细胞是特征性的和至关重要的)并且未到达细胞内过氧化氢酶(在正常细胞中也起作用)。
图25和图26显示了过氧化氢酶中和性单域vhh片段仅中和肿瘤细胞的膜过氧化氢酶并不影响正常细胞的细胞内过氧化氢酶。
分别将6000个未转化的208f细胞、转化的208fsrc3细胞,以及mkn-45肿瘤细胞接种在100μl培养基中,并与0.1或1pg/ml过氧化氢酶中和性acatcb0972或者仅过氧化氢酶结合性acatcb0973相混合。使对照制备物不添加单域vhh片段。随后,加入指定浓度的葡萄糖氧化酶并且在1.5小时后测量凋亡诱导。葡萄糖氧化酶(gox)产生h2o2,其是细胞可渗透的并因此可以通过细胞内和膜过氧化氢酶降解。在足够的浓度下,就细胞的恶性状态而言,h2o2非选择性地诱导凋亡(ivanovasetal.,selectiveandnonselectiveapoptosisinductionintransformedandnontransformedfibroblastsbyexogenousreactiveoxygenandnitrogenspecies.anticancerresearch,anticancerres.22:841-856,2002)。
图25a显示了与正常细胞和转化的细胞系相比,好得多地保护肿瘤细胞系mkn-45免于h2o2。在过氧化氢酶中和性单域vhh片段的存在下,对h2o2的效果而言非常清楚地敏化肿瘤细胞,而仅结合过氧化氢酶的单域vhh片段不导致致敏。图26显示了中和性单域vhh片段导致对转化的细胞系的致敏作用,但不能影响正常细胞。
图25和26确认了单域vhh片段对恶性细胞的具体效果,而正常细胞不受影响。这里,用肿瘤细胞观察到最大的效果,因为这些(首先)受膜过氧化氢酶保护。已知转化的细胞在表面上携带可检测量的过氧化氢酶,然而,比在肿瘤细胞中以更低的局部浓度存在,并因此不足以提供保护而免受ros信号传导。正常细胞中反应的缺乏证明了单域vhh片段不穿过细胞并且不能影响存在于那里的过氧化氢酶。即,它们特异性地对膜过氧化氢酶(其尤其对于肿瘤细胞是特征性的)起作用。然而,关于单域vhh片段的反应位点的这个非常重要的陈述仅当同时能够证明正常细胞的细胞内过氧化氢酶的抑制实际上将影响它们针对h2o2的灵敏性时是有意义的。
在图27中处理该对照方面。
图27显示了正常细胞的细胞内过氧化氢酶的sirna介导的敲低增加了它们对h2o2的灵敏性。
用对照sirna(sico)和针对过氧化氢酶的sirna(sicat)转染正常细胞(208f)、转化的细胞(208fsrc3)以及肿瘤细胞(mkn-45),并在37℃和5%co2维持24小时。此后,将细胞在新鲜培养基中继续,并且以6000个细胞/100μl培养基的细胞密度继续。随后,用增加浓度的gox(a-c)或者过氧化亚硝酸盐(pon)处理制备物(d-f)。37℃和5%co2温育1.5小时后,测定凋亡细胞的百分数。为了评估该实验,概括了gox产生h2o2,其具有非常好的细胞渗透性,并因此可被膜过氧化氢酶和细胞内过氧化氢酶两者降解。另一方面,当外源添加的过氧化亚硝酸盐与细胞接触时,其与细胞膜反应。因此,针对过氧化亚硝酸盐的保护仅可以通过位于膜外部的过氧化氢酶实现。
首先,图27确认了与正常或转化细胞相比,好得多地保护肿瘤细胞免于h2o2和过氧亚硝酸盐。此外,该图显示了sirna介导的过氧化氢酶的降解导致针对h2o2的影响非常强烈地致敏正常细胞、转化的细胞,以及肿瘤细胞。用过氧亚硝酸盐处理后,出现了完全不同的画面:现在,正常细胞中sirna介导的过氧化氢酶降解不导致致敏,而在转化的细胞和肿瘤细胞中出现强烈的致敏。这显示出仅在恶性细胞(即转化的细胞和肿瘤细胞)表面上,存在有过氧化氢酶,其针对外源过氧化亚硝酸盐提供保护。在正常细胞中不能检测出保护性膜过氧化氢酶作用。通过h2o2生成性gox做出的发现表明,正常细胞具有功能性保护性过氧化氢酶,该酶的作用可以例如通过sirna介导的敲低检测。然而由于在示于图26中的实验中,不能通过过氧化氢酶中和性单域vhh片段方式检测针对h2o2致敏正常细胞,证明它们不能在细胞内起作用。
实施例15:对单域vhh片段所显示的体外效应与肿瘤生长的体内抑制相关。
图28和图29显示了免疫受损小鼠的人结肠癌异种移植物的生长受单域vhh片段asodcb0989重复施用所抑制。
图28和图29中进行的实验由商业认证供应商(oncotestgmbh,freiburg)进行。这里,将人结肠癌的异种移植物植入合适的小鼠中。肿瘤生长并达到一定的最小尺寸后,开始治疗试验。每周两次静脉施用指定剂量(mg/kg体重)的asodcb0989或缓冲液。此外,依靠卡尺测径(caliber)每周两次测量肿瘤大小。
首先,图28显示了在对照动物中的肿瘤生长以非常清晰的扩散为特征。当施用0.03mg/kg的asodcb0989时,不产生与对照的可识别的差异(图28a)。当施用0.3mg/kg的asodcb0989时,单域vhh片段处理组非常清晰地区别于对照组,尽管这两组受到非常强烈的扩散。所说明的对照组和处理组之间肿瘤体积上的差异是由asodcb0989的清晰增殖抑制的表达。当进一步将asodcb0989的剂量增加到0.9mg/kg(图29)时,在一些动物中,增强了生长抑制,而在其他动物中导致相反结果。这显示,还呈现了最佳值形式的体内剂量效应曲线,并且示于图29中的实验条件已达到超最佳值抑制的浓度限。
<110>乔治.鲍尔
曼弗莱德.莫兹
<120>抗原结合构建体
<130>pwo00251bau
<160>51
<170>patentinversion3.5
<210>1
<211>19
<212>rna
<213>人工序列
<220>
<223>对照sirna
<400>1
uucuccgaacgugucacgu19
<210>2
<211>19
<212>rna
<213>人工序列
<220>
<223>对照sirna反义
<400>2
acgugacacguucggagaa19
<210>3
<211>21
<212>dna
<213>人工序列
<220>
<223>sirnafasr
<400>3
aaggagtacacagacaaagcc21
<210>4
<211>21
<212>dna
<213>人工序列
<220>
<223>sirnacasp8
<400>4
aagagtctgtgcccaaatcaa21
<210>5
<211>21
<212>dna
<213>人工序列
<220>
<223>sirnacasp9
<400>5
cagtgacatctttgtgtccta21
<210>6
<211>21
<212>dna
<213>人工序列
<220>
<223>sirnanox
<400>6
ccgacaaatactactacacaa21
<210>7
<211>21
<212>dna
<213>人工序列
<220>
<223>sirnanos2
<400>7
ctgggccgtgcaaaccttcaa21
<210>8
<211>21
<212>dna
<213>人工序列
<220>
<223>sirna过氧化氢酶
<400>8
ccggatctcacttggcggcaa21
<210>9
<211>21
<212>dna
<213>人工序列
<220>
<223>sirna小鼠过氧化氢酶
<400>9
cccaataggagataaacttaa21
<210>10
<211>128
<212>prt
<213>人工序列
<220>
<223>过氧化氢酶结合
<400>10
metalaglnvalglnleuvalgluserglyglyglyleuvalglnala
151015
glyglyserleuargleusercysalaalasergluargthrpheasn
202530
thrtyrglymetglytrppheargglnalaproglylysgluargglu
354045
phevalalathrilesertrpserglyaspserthrtyrtyralaasp
505560
servallysglyargphethrileserargaspasnalalysasnthr
65707580
mettyrleuglnmetasnserleulysprogluaspthralavaltyr
859095
tyrcysasnalaasnserglutyrglyaspsertyrtrpglyglngly
100105110
thrglnvalthrvalserserlyslyslyshishishishishishis
115120125
<210>11
<211>128
<212>prt
<213>人工序列
<220>
<223>过氧化氢酶结合
<400>11
metalagluvalglnleuvalgluserglyglyglyleuvalglnpro
151015
glyglyserleuargleusercysalavalserglypheilepheasn
202530
thrtyrsermetargtrpglyargglnalaproglylysglyleuglu
354045
trpvalserserileserthrglyglytyrserthrtyralaaspser
505560
vallysglyargphethrileserargaspasnalalysasnleuval
65707580
tyrleuglnmetasnserleulysprogluaspthralavaltyrtyr
859095
cysglytrpglyalaphevalargglygluargproglnglyglngly
100105110
thrglnvalthrvalserserlyslyslyshishishishishishis
115120125
<210>12
<211>133
<212>prt
<213>人工序列
<220>
<223>过氧化氢酶结合
<400>12
metalaglnvalglnleuvalgluserglyglyglyleuvalglnpro
151015
glyglyserleuargleusercysalaalaserglyserilepheser
202530
ilealasermetglytrptyrargglnalaproglylysglnargasp
354045
leuvalalathrilethrseraspglyserthrlystyralaaspser
505560
vallysglyargphethrileserargaspasnalalysasnthrmet
65707580
tyrleuglnmetasnservallysprogluaspalaalavaltyrtyr
859095
cysasnalaaspalaaspaspleugluproglysertyrasptyrasp
100105110
tyrtrpglyglnglythrglnvalthrvalserserlyslyslyshis
115120125
hishishishishis
130
<210>13
<211>133
<212>prt
<213>人工序列
<220>
<223>过氧化氢酶结合
<400>13
metalaglnvalglnleuvalgluserglyglyglyleuvalglnpro
151015
glyglyserleuargleusercysalaalaseralaserilepheser
202530
iletyrvalmetalatrptyrargglnalaproglylysglnargglu
354045
leuvalalathrvalthrserglyglyalathrasntyralaasnser
505560
vallysglyargphethrileserargaspasnalalysasnthrmet
65707580
aspleuglnmetasnserleulysprogluaspthralavaltyrtyr
859095
cysasnalagluasptyrtyrasptyrglyleuserargserlysile
100105110
tyrtrpglyglnglythrglnvalthrvalserserlyslyslyshis
115120125
hishishishishis
130
<210>14
<211>127
<212>prt
<213>人工序列
<220>
<223>sod结合
<400>14
metalaglnvalglnleuvalgluserglyglyglyilevalglnpro
151015
glyglyserleuargleusercysvalalasergluserileserglu
202530
ileaspalamettyrtrphisargglnalaproglylysgluargglu
354045
leuvalalaglyilethrasnaspglythrargtyrtyralaaspser
505560
vallysglyargphethrileserargaspasnalalysserthrleu
65707580
tyrleuglnmetasnserleulysphegluaspthralamettyrtyr
859095
cysalaalaleuproasnproproproglytyrtrpglyglnglythr
100105110
glnvalthrvalserserlyslyslyshishishishishishis
115120125
<210>15
<211>127
<212>prt
<213>人工序列
<220>
<223>sod结合
<400>15
metalaglnvalglnleuvalgluserglyglyglyleuvalglnser
151015
glyglyserleuthrleusercysthralaserglyphethrileser
202530
asntyrprometthrtrpvalargglnalaproglylysglyleuglu
354045
trpvalserargileasnserglyglyaspargthrleutyralaasp
505560
servallysglyargphethrvalserargaspasnalaargasnthr
65707580
mettyrleuglnmetasnasnleulysprogluaspthrglyleutyr
859095
phecysalaaspserglyalaglytrpargtyrtrpglyglnglythr
100105110
glnvalthrvalserserlyslyslyshishishishishishis
115120125
<210>16
<211>127
<212>prt
<213>人工序列
<220>
<223>sod结合
<400>16
metalaglnvalglnleuvalgluserglyglyglyilevalglnpro
151015
glyglyserleuargleusercysvalalasergluserileserasp
202530
ileaspalamettyrtrphisargglnalaproglylysargargglu
354045
leuvalalaglyilethrasnaspglythrglutyrphealaaspser
505560
vallysglyargphealaileserargaspasnthrlysserserleu
65707580
tyrleuglnmetasnserleulysleugluaspthralamettyrtyr
859095
cysalathrleuproasnproproproglytyrtrpglyglnglythr
100105110
glnvalthrvalserserlyslyslyshishishishishishis
115120125
<210>17
<211>260
<212>prt
<213>人工序列
<220>
<223>双特异性
<400>17
metalaglnvalglnleuvalgluserglyglyglyleuvalglnala
151015
glyglyserleuargleusercysalaalasergluargthrpheasn
202530
thrtyrglymetglytrppheargglnalaproglylysgluargglu
354045
phevalalathrilesertrpserglyaspserthrtyrtyralaasp
505560
servallysglyargphethrileserargaspasnalalysasnthr
65707580
mettyrleuglnmetasnserleulysprogluaspthralavaltyr
859095
tyrcysasnalaasnserglutyrglyaspsertyrtrpglyglngly
100105110
thrglnvalthrvalserserglyglyglyglyserglyglyglygly
115120125
serglyglyglyglyseralaglnvalglnleuvalgluserglygly
130135140
glyleuvalglnserglyglyserleuthrleusercysthralaser
145150155160
glyphethrileserasntyrprometthrtrpvalargglnalapro
165170175
glylysglyleuglutrpvalserargileasnserglyglyasparg
180185190
thrleutyralaaspservallysglyargphethrvalserargasp
195200205
asnalaargasnthrmettyrleuglnmetasnasnleulysproglu
210215220
aspthrglyleutyrphecysalaaspserglyalaglytrpargtyr
225230235240
trpglyglnglythrglnvalthrvalserserlyslyslyshishis
245250255
hishishishis
260
<210>18
<211>260
<212>prt
<213>人工序列
<220>
<223>双特异性
<400>18
metalaglnvalglnleuvalgluserglyglyglyleuvalglnser
151015
glyglyserleuthrleusercysthralaserglyphethrileser
202530
asntyrprometthrtrpvalargglnalaproglylysglyleuglu
354045
trpvalserargileasnserglyglyaspargthrleutyralaasp
505560
servallysglyargphethrvalserargaspasnalaargasnthr
65707580
mettyrleuglnmetasnasnleulysprogluaspthrglyleutyr
859095
phecysalaaspserglyalaglytrpargtyrtrpglyglnglythr
100105110
glnvalthrvalserserglyglyglyglyserglyglyglyglyser
115120125
glyglyglyglyseralaglnvalglnleuvalgluserglyglygly
130135140
leuvalglnalaglyglyserleuargleusercysalaalaserglu
145150155160
argthrpheasnthrtyrglymetglytrppheargglnalaprogly
165170175
lysgluarggluphevalalathrilesertrpserglyaspserthr
180185190
tyrtyralaaspservallysglyargphethrileserargaspasn
195200205
alalysasnthrmettyrleuglnmetasnserleulysprogluasp
210215220
thralavaltyrtyrcysasnalaasnserglutyrglyaspsertyr
225230235240
trpglyglnglythrglnvalthrvalserserlyslyslyshishis
245250255
hishishishis
260
<210>19
<211>9
<212>prt
<213>人工序列
<220>
<223>cdr
<400>19
argthrpheasnthrtyrglymetgly
15
<210>20
<211>17
<212>prt
<213>人工序列
<220>
<223>cdr
<400>20
thrilesertrpserglyaspserthrtyrtyralaaspservallys
151015
gly
<210>21
<211>8
<212>prt
<213>人工序列
<220>
<223>cdr
<400>21
asnserglutyrglyaspsertyr
15
<210>22
<211>9
<212>prt
<213>人工序列
<220>
<223>cdr
<400>22
pheilepheasnthrtyrsermetarg
15
<210>23
<211>16
<212>prt
<213>人工序列
<220>
<223>cdr
<400>23
serileserthrglyglytyrserthrtyralaaspservallysgly
151015
<210>24
<211>9
<212>prt
<213>人工序列
<220>
<223>cdr
<400>24
glyalaphevalargglygluargpro
15
<210>25
<211>9
<212>prt
<213>人工序列
<220>
<223>cdr
<400>25
serilepheserilealasermetgly
15
<210>26
<211>16
<212>prt
<213>人工序列
<220>
<223>cdr
<400>26
thrilethrseraspglyserthrlystyralaaspservallysgly
151015
<210>27
<211>14
<212>prt
<213>人工序列
<220>
<223>cdr
<400>27
aspalaaspaspleugluproglysertyrasptyrasptyr
1510
<210>28
<211>9
<212>prt
<213>人工序列
<220>
<223>cdr
<400>28
serilepheseriletyrvalmetala
15
<210>29
<211>16
<212>prt
<213>人工序列
<220>
<223>cdr
<400>29
thrvalthrserglyglyalathrasntyralaasnservallysgly
151015
<210>30
<211>14
<212>prt
<213>人工序列
<220>
<223>cdr
<400>30
gluasptyrtyrasptyrglyleuserargserlysiletyr
1510
<210>31
<211>9
<212>prt
<213>人工序列
<220>
<223>cdr
<400>31
serilesergluileaspalamettyr
15
<210>32
<211>16
<212>prt
<213>人工序列
<220>
<223>cdr
<400>32
glyilethrasnaspglythrargtyrtyralaaspservallysgly
151015
<210>33
<211>8
<212>prt
<213>人工序列
<220>
<223>cdr
<400>33
leuproasnproproproglytyr
15
<210>34
<211>9
<212>prt
<213>人工序列
<220>
<223>cdr
<400>34
phethrileserasntyrprometthr
15
<210>35
<211>17
<212>prt
<213>人工序列
<220>
<223>cdr
<400>35
argileasnserglyglyaspargthrleutyralaaspservallys
151015
gly
<210>36
<211>7
<212>prt
<213>人工序列
<220>
<223>cdr
<400>36
serglyalaglytrpargtyr
15
<210>37
<211>9
<212>prt
<213>人工序列
<220>
<223>cdr
<400>37
serileseraspileaspalamettyr
15
<210>38
<211>16
<212>prt
<213>人工序列
<220>
<223>cdr
<400>38
glyilethrasnaspglythrglutyrphealaaspservallysgly
151015
<210>39
<211>8
<212>prt
<213>人工序列
<220>
<223>cdr
<400>39
leuproasnproproproglytyr
15
<210>40
<211>9
<212>prt
<213>人工序列
<220>
<223>cdr
<400>40
argthrpheasnthrtyrglymetgly
15
<210>41
<211>17
<212>prt
<213>人工序列
<220>
<223>cdr
<400>41
thrilesertrpserglyaspserthrtyrtyralaaspservallys
151015
gly
<210>42
<211>8
<212>prt
<213>人工序列
<220>
<223>cdr
<400>42
asnserglutyrglyaspsertyr
15
<210>43
<211>9
<212>prt
<213>人工序列
<220>
<223>cdr
<400>43
phethrileserasntyrprometthr
15
<210>44
<211>17
<212>prt
<213>人工序列
<220>
<223>cdr
<400>44
argileasnserglyglyaspargthrleutyralaaspservallys
151015
gly
<210>45
<211>7
<212>prt
<213>人工序列
<220>
<223>cdr
<400>45
serglyalaglytrpargtyr
15
<210>46
<211>9
<212>prt
<213>人工序列
<220>
<223>cdr
<400>46
phethrileserasntyrprometthr
15
<210>47
<211>17
<212>prt
<213>人工序列
<220>
<223>cdr
<400>47
argileasnserglyglyaspargthrleutyralaaspservallys
151015
gly
<210>48
<211>7
<212>prt
<213>人工序列
<220>
<223>cdr
<400>48
serglyalaglytrpargtyr
15
<210>49
<211>9
<212>prt
<213>人工序列
<220>
<223>cdr
<400>49
argthrpheasnthrtyrglymetgly
15
<210>50
<211>17
<212>prt
<213>人工序列
<220>
<223>cdr
<400>50
thrilesertrpserglyaspserthrtyrtyralaaspservallys
151015
gly
<210>51
<211>8
<212>prt
<213>人工序列
<220>
<223>cdr
<400>51
asnserglutyrglyaspsertyr
15
- 还没有人留言评论。精彩留言会获得点赞!
- 桑树过氧化物酶MmPOD.12及其应用
- Marinamoeba thermophila生产耐热超氧化物歧化酶(SOD)的方法
- Corynascus thermophilus 生产耐热超氧化物歧化酶(SOD)的方法
- Thermoanaerobacter thermocopriae 生产耐热超氧化物歧化酶(SOD)的方法
- Stilbella thermophila生产耐热超氧化物歧化酶(SOD)的方法
- Thermoflavimicrobium dichotomicum生产耐热超氧化物歧化酶(SOD)的方法
- 普通嗜热放线菌生产耐热超氧化物歧化酶(sod)的方法
- 中间型嗜热放线菌生产耐热超氧化物歧化酶(sod)的方法
- Spirochaeta thermophila生产耐热超氧化物歧化酶(SOD)的方法
- Platyamoeba thermophila生产耐热超氧化物歧化酶(SOD)的方法
- 一种储片盒换气歧管的制作方法
- 一种能增加氧化反应速度的化工试剂反应釜结构的制作方法
- 标志物及其应用、试剂盒、该标志物的检测方法
- Marinamoeba thermophila生产耐热超氧化物歧化酶(SOD)的方法
- Corynascus thermophilus 生产耐热超氧化物歧化酶(SOD)的方法
- Thermoanaerobacter thermocopriae 生产耐热超氧化物歧化酶(SOD)的方法
- Stilbella thermophila生产耐热超氧化物歧化酶(SOD)的方法
- 普通嗜热放线菌生产耐热超氧化物歧化酶(sod)的方法
- 中间型嗜热放线菌生产耐热超氧化物歧化酶(sod)的方法
- Spirochaeta thermophila生产耐热超氧化物歧化酶(SOD)的方法
- 月桂酸修饰超氧化物歧化酶的制备方法
- Marinamoeba thermophila生产耐热超氧化物歧化酶(SOD)的方法
- Corynascus thermophilus 生产耐热超氧化物歧化酶(SOD)的方法
- Thermoanaerobacter thermocopriae 生产耐热超氧化物歧化酶(SOD)的方法
- Stilbella thermophila生产耐热超氧化物歧化酶(SOD)的方法
- Thermoflavimicrobium dichotomicum生产耐热超氧化物歧化酶(SOD)的方法
- 普通嗜热放线菌生产耐热超氧化物歧化酶(sod)的方法
- 中间型嗜热放线菌生产耐热超氧化物歧化酶(sod)的方法
- Spirochaeta thermophila生产耐热超氧化物歧化酶(SOD)的方法
- Platyamoeba thermophila生产耐热超氧化物歧化酶(SOD)的方法
- Thermoleophilum minutum生产耐热超氧化物歧化酶(SOD)的方法
- Cryptococcus thermophilus生产耐热超氧化物歧化酶(SOD)的方法
- Thermobispora bispora生产耐热超氧化物歧化酶(SOD)的方法
- Thermoleophilum album生产耐热超氧化物歧化酶(SOD)的方法
- Talaromyces thermo-citrinum 生产耐热超氧化物歧化酶(SOD)的方法
- Moorella thermoautotrophica 生产耐热超氧化物歧化酶(SOD)的方法
- Lagenidium thermophilum生产耐热超氧化物歧化酶(SOD)的方法
- Thermoanaerobacterium thermosulfurigenes生产耐热超氧化物歧化酶(SOD)的方法
- Calcarisporiella thermophila生产耐热超氧化物歧化酶(SOD)的方法
- Crithidia luciliae thermophila生产耐热超氧化物歧化酶(SOD)的方法