一种全人源抗人白介素17A抗体及其应用的制作方法
本发明涉及生物医药领域,具体涉及一种全人源抗人白介素17a抗体及其用途。
背景技术:
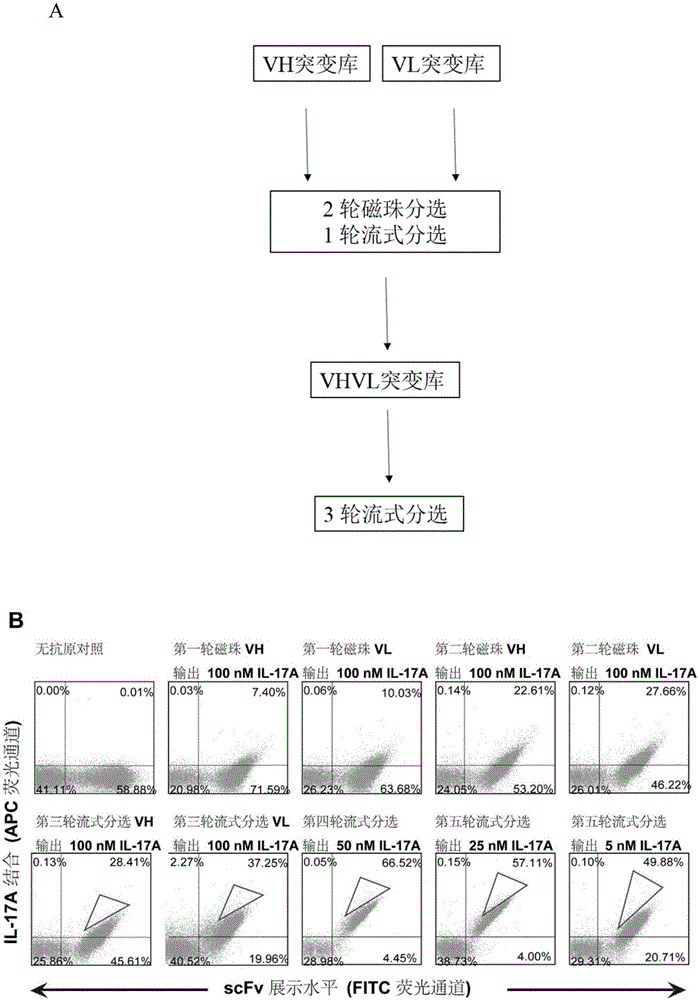
:人白细胞介素17(humaninterleukin-17,hil-17)于1993年从一种激活的t细胞杂交瘤中分离出来,最初在啮齿类动物发现时称为细胞毒性t细胞抗原8(ctla8)。目前已知il-17即il-17a主要由th-17细胞分泌,且在il-17家族中存在6个成员(il-17a-f),它们分别由150~180个氨基酸构成,通常以二聚体的形式存在。il-17a可诱导多种细胞(包括成纤维细胞、内皮细胞、上皮细胞以及巨噬细胞等)产生其他细胞因子如il-6、g-csf、gm-csf、il-1β、tgf-β、tnf-α;化学趋化因子如il-8,gro-α,和单核细胞趋化因子-1(mcp-1);以及前列腺素pge2。il-17诱导化学趋化因子il-8,gro-α以及mcp-1增加,促使中性粒细胞和单核细胞的募集。il-17诱导产生的il-6和pge2加强局部炎症反应;诱导细胞间黏附因子(icam)产生,促使t细胞反应。il-17具有强大的促炎作用,其与多种疾病如类风湿性关节炎(rheumatoidarthritis,ra)、银屑病、气道炎症、慢性炎性疾病、纤维增生疾病以及肿瘤的生长相关。类风湿性关节炎:一种以关节滑膜炎为特征,以慢性多发性关节炎为主要临床表现的一种自身免疫性疾病。研究表面,在病人关节的滑液中可以检测到活性的il-17,il-17刺激软骨细胞产生inos和no及其它与分解代谢相关的酶,并与il-1β,tnf-α一起刺激成骨细胞分泌如gm-csf和il-6等细胞因子,从而抑制软骨细胞增殖和蛋白多糖合成,调节破骨细胞,发生骨破坏。il-17还可以诱导基质金属蛋白酶,直接在疾病进展中起到破坏性作用。因此,阻断il-17/il-17r通路可能是治疗ra的新途径。研究发现,利用抗鼠il-17抗体治疗胶原诱导的关节炎(collagen-inducedarthritis,cia)模型中,关节局部炎症,软骨破坏,骨质侵蚀明显减轻。银屑病(teunissen等,1998)和银屑病关节炎:抗il-17a抗体secukinumabiii期研究结果显示,300mgsecukinumab治疗12周后在消除银屑病皮损方面的疗效显著高于安慰剂组(p<0.0001)2015年1月美国fda批准secukinumab治疗恶性瘢块状银屑病,此抗体在日本也应用于银屑病性关节炎。纤维化疾病:多种慢性纤维化性疾病均发现与il-17信号传导通路相关(mi等,2011)。各种原因引起的慢性病主要以慢性炎症和组织纤维化为基本病理特征。许多慢性病,包括哮喘、支气管扩张、慢阻肺、肺结核、肺癌、间质性肺病等,都伴有纤维增生或以纤维增生为主要病变。因此,纤维增生快慢和纤维化程度决定了这些疾病的临床预后。肺成纤维细胞、肝星型胶质细胞分别是肺、肝纤维化发生发展过程中的关键效应细胞。肺病:如慢性阻塞性肺病(chronicobstructivepulmonarydiseases),是一种具有气流受限特征的疾病,气流受限不完全可逆、呈进行性发展,多与肺部对有害颗粒物或有害气体的异常炎症反应有关。其病程的恶化,与气道中中性粒细胞的数量增多有关。研究表明,在copd急性加重期及稳定期il-17水平显著升高,il-17引起中性粒细胞增多,促进释放il-6、il-8,打破基质金属蛋白酶平衡,参与了copd的发病,是引起肺内炎症细胞浸润及肺实质破坏的主要原因之一。肿瘤:il-17促进多种肿瘤疾病的发生和发展(zeng等,2015),尤其是结肠癌、膀胱癌、肝癌、肺癌、胰腺癌、乳腺癌以及胃癌、脑垂体瘤,此外亦有报道淋巴瘤(maggio等,2002,ferretti等,2015)、前列腺癌(liu等,2016)、子宫内膜癌(ning等,2016)、甲状腺癌(lu等,2016)。il-17对食管鳞状细胞癌以及卵巢癌则呈现保护作用(zeng等,2015)。其他疾病如:急性移植物排斥(antonysamy等,1999;yoshida等,2006;tang等,2001);全身性硬化症(kurasawa等,2000);全身性红斑狼疮(wong等,2000);自身免疫炎性肠炎和克罗恩病(nielsen等,2003;fujino等,2003;yen等,2006);心血管疾病,包括动脉粥样硬化(csiszar和ungvari,2004),川崎病(sohn等,2003),缺血性心脏病(csiszar,2003),非酒精性肝病(giles等,2016)和中风;病毒感染导致的疾病等。综上所诉,此类疾病均出现il-17异常表达,利用il-17拮抗剂治疗此类疾病,具有重要价值。上世纪90年代中期以来,抗体药物在新药中崭露头角。在治疗性应用中,全人抗体可以克服鼠源单克隆抗体在临床应用中的诸多缺点:如诱发人体产生抗鼠抗体(hama)反应,不能有效引起cdc及adcc等。随着对人类抗体基因及结构的研究及分子生物学技术的进展,利用噬菌体抗体库制备人源抗体,已成为获得全人抗体的最主要手段之一。噬菌体抗体库是利用基因重组技术将外源基因的基因型和表型统一在同一个噬菌体颗粒内。更重要的是,该类噬菌体抗体,不仅可以与特异的配体结合,而且保持感染能力。这样又将抗体的选择能力和噬菌体的扩增能力偶联起来,使得噬菌体展示技术成为一种极其有效的抗体筛选系统。经典的筛选方法是将抗原纯化,然后与抗体库孵育,通过数次“吸附—洗涤—洗脱—扩增”的过程(即生物淘选),使得特异克隆得以富集。可以迅速得到针对靶抗原的抗体,通过功能筛选最终获得中和性抗体。针对il-17a的抗体已见报道,wo2006/013107(诺华制药股份有限公司)报道了il-17a的特异性抗体,特别称为ain457(secukinumab),从杂交瘤分离人抗il-17抗体与人il-17a的亲和力为227pm。2014年10月,诺华公司公布的ain457(secukinumab)iii期研究结果显示,300mgain457(secukinumab)治疗12周后在消除银屑病皮损方面的疗效显著高于安慰剂组(p<0.0001)。2015年1月美国fda批准secukinumab治疗恶性瘢块状银屑病,此抗体在日本也应用于银屑病性关节炎。r&d公司已生产鼠抗人il-17单克隆抗体mab317。wo2006/054059(ucb细胞技术公司)描述了il-17a中和性抗体分子,该分子最初从杂交瘤中分离并peg化抗体片段。采用biacore测定该片段对il-17的亲和力为133-365pm。cn200580026569.4(诺瓦提斯公司)报道il-17a的特异性抗体,并应用与治疗il-17介导的相关疾病。cn200680046605.8(伊莱利利公司)报道il-17a的特异性抗体,并应用与治疗il-17介导的相关疾病。cn200780023566.4(阿斯特拉捷利康股份公司),报道利用噬菌体库分离获得抗il-17单链抗体片段。cn200780029886.0(ucb医药有限公司)报道il-17a的特异性抗体,并应用与治疗il-17介导的相关疾病。cn200780003925.x(诺瓦提斯公司)报道了将il-17a特异性抗体用于治疗实体或造血系统性肿瘤。cn200780037662.4(先灵公司)报道il-17a的特异性抗体,并应用与治疗il-17介导的相关疾病。cn201180048098.2(安奈普泰斯生物有限公司)报道il-17a的特异性抗体,并应用与治疗il-17介导的相关疾病。cn201380012452.5(伊莱利利公司)报道il-17a的特异性抗体,并应用与治疗il-17介导的相关疾病。cn201510097117.0(北京百特美博生物科技有限公司)报道il-17a的特异性抗体,并应用与治疗il-17介导的相关疾病。cn104231080a报道一种构建天然噬菌体抗体库的方法以及由该抗体库获得的可与人白介素17a特异结合的单链抗体。该抗体片段构建成全长抗体,通过真核分泌表达亲和纯化后能够抑制il-17a对人纤维肉瘤细胞ht1080释放白介素6(il-6)的反应;并且可用于检测及治疗类风湿性关节炎。然而,此抗体相对有限的亲和力可是临床应用的一个限制。技术实现要素:本发明要解决的技术问题是在cn104231080a公开的抗il-17a抗体基础上,经随机突变、生物学结构改造等方法,获得全新il-17a的抗体分子,特别是人抗体分子,其具有抑制il-17a相关生物学活性,可用于治疗il-17a相关疾病。本发明以cn104231080a公开的抗il-17a抗体的重链可变区(vh)、轻链可变区(vl)为基础,进行随机突变以及生物学结构改造。随机突变后(筛选)获得高亲和力的抗体,经分析验证发现突变位点在轻链可变区和重链可变区的框架区(fr)和抗原互补决定区(cdr)均有分布;发明人进而在该突变抗体的基础上进行人重链框架区和人轻链框架区的不同抗体类型的替换,得到骨架稳定性好的抗体,并进一步在此基础上进行点突变,最终所得的抗体均具有良好的自身稳定性、高表达量以及亲和力。本发明解决上述技术问题的技术方案之一是:一种全人源抗人白介素17a抗体,其包含重链可变区、和/或轻链可变区;所述重链可变区为a):其氨基酸序列如seqidno.1所示、编码其的核苷酸序列优选如seqidno.8所示,或b):所述a)中的人重链框架区改变后的重链可变区;所述轻链可变区为a):其氨基酸序列如seqidno.2所示、编码其的核苷酸序列优选如seqidno.9所示,或b):所述a)中的人轻链框架区改变后的轻链可变区。所述人重链框架区的改变包括不同种类的人重链框架区的替换和/或对重链框架区的氨基酸进行点突变,本发明中优选为人重链框架区的替换,替换后的框架区根据抗体框架分类属于人的vh1、vh2、vh3、vh4和/或vh5;较佳地属于人的vh3;更佳地,重链框架区替换为人vh3后的重链可变区的氨基酸序列如seqidno.3所示;进一步更佳地,编码所述重链可变区的核苷酸序列如seqidno.10所示。所述人轻链框架区的改变包括不同种类的人轻链框架区的替换和/或其点突变体。根据本发明,所述突变体优选是在替换后人轻链框架区基础上进行点突变。所述框架区指的是可变区中除抗原互补决定区之外的区域,通常一个可变区内包含四个框架区,分别命名为fr1、fr2、fr3以及fr4。本发明中,替换后的框架区根据抗体框架分类属于人的vκ1、vκ2、vκ3、vκ4、vλ1、vλ2和/或vλ4;较佳地,属于人的vκ1;更佳地,轻链框架区替换为人的vκ1的轻链可变区的氨基酸序列如seqidno.4所示;进一步更佳地,编码所述的轻链可变区的核苷酸序列如seqidno.11所示。其中,替换后的轻链框架区的点突变体的突变位置位于人轻链框架区的fr3区;较佳地,位于序列seqidno.4的第66位、第85位和/或第86位;更佳地,所述第66位突变后的氨基酸为丙氨酸,第85位突变后的氨基酸为天冬酰胺,第86位突变后的氨基酸为丝氨酸或者苏氨酸。在本发明中,轻链框架区替换为人vκ1点突变体的轻链可变区的氨基酸序列如seqidno.5、seqidno.6或seqidno.7所示;较佳地,编码所述的轻链可变区的核苷酸序列如seqidno.12、seqidno.13或seqidno.14所示。在本发明较佳实施例中,所述抗体包含的:所述重链可变区的氨基酸序列如seqidno.1所示,且所述轻链可变区的氨基酸序列如seqidno.2所示;较佳地,编码所述重链可变区的核苷酸序列如seqidno.8所示,且所述轻链可变区的核苷酸序列如seqidno.9所示;或者所述重链可变区的氨基酸序列如seqidno.3所示,且所述轻链可变区的氨基酸如seqidno.4所示;较佳地,编码所述重链可变区的核苷酸序列如seqidno.10所示,且所述轻链核苷酸序列如seqidno.11所示;或者所述重链可变区的氨基酸序列如seqidno.3所示,且所述轻链可变区的氨基酸如seqidno.5所示;较佳地,编码所述重链可变区的核苷酸序列如seqidno.10所示,且所述轻链可变区的核苷酸序列如seqidno.12所示;或者所述重链可变区的氨基酸序列如seqidno.3所示,且所述轻链可变区的氨基酸如seqidno.6所示;较佳地,编码所述重链可变区的核苷酸序列如seqidno.10所示,且所述轻链可变区的核苷酸序列如seqidno.13所示;或者所述重链可变区的氨基酸序列如seqidno.3所示,且所述轻链可变区的氨基酸如seqidno.7所示;较佳地,编码所述重链可变区的核苷酸序列如seqidno.10所示,且所述轻链可变区的核苷酸序列如seqidno.14所示。具体如下表1所示。表1本发明5种抗体重链可变区以及轻链可变区的氨基酸以及核苷酸序列编号抗体名vh-proteinvh-dnavl-proteinvl-dna9nt.d3seqidno.1seqidno.8seqidno.2seqidno.99nt.c3seqidno.3seqidno.10seqidno.4seqidno.119nt.c53aseqidno.3seqidno.10seqidno.5seqidno.129nt.nsseqidno.3seqidno.10seqidno.6seqidno.139ntseqidno.3seqidno.10seqidno.7seqidno.14本发明中hfr1(重链可变区的fr1)的氨基酸序列如序列表中seqidno.21或者24所示,hfr2的氨基酸序列如序列表中seqidno.22或者25所示,hfr3的氨基酸序列如序列表中seqidno.23或者26所示;lfr1(轻链可变区的fr1)的氨基酸序列如序列表中seqidno.27或者33所示,lfr2的氨基酸序列如序列表中seqidno.28或者34所示,lfr3的氨基酸序列如序列表中seqidno.29、30、31、32或者35所示。本发明的抗体,可以含有上述结合片段的改型片段、抗体基因片段,以及同源性在30%以上的免疫结合片段。所述的全人源抗人白介素17a抗体较佳地为抗体全长蛋白、抗原抗体结合域蛋白质片段、双特异性抗体、多特异性抗体、单链抗体(singlechainantibodyfragment,scfv)、单域抗体(singledomainantibody,sdab)或单区抗体(signle-domainantibody)中的一种或多种。所述的抗体全长蛋白为本领域常规的抗体全长蛋白,其包括重链可变区、轻链可变区、重链恒定区和轻链恒定区。本发明解决上述技术问题的技术方案之二是:一种编码上述全人源抗人白介素17a抗体的核酸。较佳地,其包括编码所述重链可变区的核酸,和/或,编码所述轻链可变区的核酸;所述重链可变区的氨基酸序列如seqidno.1或者seqidno.3所示;较佳地,编码所述重链可变区的核酸的核苷酸序列如序列表中seqidno.8或者seqidno.10所示;和/或,所述的轻链可变区的氨基酸序列如seqidno.2、seqidno.4、seqidno.5、seqidno.6或seqidno.7所示;较佳地,编码所述轻链可变区的核酸的核苷酸序列如seqidno.9、seqidno.11、seqidno.12、seqidno.13或seqidno.14所示。较佳地,所述重链可变区的氨基酸序列如seqidno.1所示,且所述轻链可变区的氨基酸序列如seqidno.2所示;较佳地,编码所述重链可变区的核酸的核苷酸序列如seqidno.8所示,且所述轻链可变区的核苷酸序列如seqidno.9所示;或者所述重链可变区的氨基酸序列如seqidno.3所示,且所述轻链可变区的氨基酸如seqidno.4所示;较佳地,所述重链可变区的核苷酸序列如seqidno.10所示,且所述轻链核苷酸序列如seqidno.11所示;或者所述重链可变区的氨基酸序列如seqidno.3所示,且所述轻链可变区的氨基酸如seqidno.5所示;较佳地,所述重链可变区的核苷酸序列如seqidno.10所示,且所述轻链可变区的核苷酸序列如seqidno.12所示;或者所述重链可变区的氨基酸序列如seqidno.3所示,且所述轻链可变区的氨基酸如seqidno.6所示;较佳地,所述重链可变区的核苷酸序列如seqidno.10所示,且所述轻链可变区的核苷酸序列如seqidno.13所示或者所述重链可变区的氨基酸序列如seqidno.3所示,且所述轻链可变区的氨基酸如seqidno.7所示;较佳地,所述重链可变区的核苷酸序列如seqidno.10所示,且所述轻链可变区的核苷酸序列如seqidno.14所示。本发明解决上述技术问题的技术方案之三是:一种包含上述核酸的重组表达载体。其中,所述重组表达载体可通过本领域常规方法获得,即:将本发明所述的核酸分子连接于各种表达载体上构建而成。所述的表达载体为本领域常规的各种载体,只要其能够容载前述核酸分子即可。所述载体较佳地包括:各种质粒、粘粒、噬菌体或病毒载体等。本发明解决上述技术问题的技术方案之四是:一种包含上述重组表达载体的重组表达转化体。其中,所述重组表达转化体的制备方法为本领域常规的制备方法,较佳地为:将上述重组表达载体转化至宿主细胞中制得。所述的宿主细胞为本领域常规的各种宿主细胞,只要能满足使上述重组表达载体稳定地自行复制,且所携带所述的核酸可被有效表达即可。本发明解决上述技术问题的技术方案之五是:一种全人源抗人白介素17a抗体的制备方法,其包括如下步骤:培养如权利要求11所述的重组表达转化体,从培养物中获得全人源抗人白介素17a抗体。本发明解决上述技术问题的技术方案之六是:一种上述的全人源抗人白介素17a抗体在制备治疗il-17a相关疾病中的应用;较佳地,所述的il-17a相关疾病选自类风湿性关节炎、纤维化疾病、肿瘤或自身免疫病;更佳地,所述纤维化疾病为肺纤维化和/或肝纤维化,所述肿瘤为结肠癌或乳腺癌,所述自身免疫病为银屑病。在符合本领域常识的基础上,上述各优选条件,可任意组合,即得本发明各较佳实例。本发明所用试剂和原料均市售可得。本发明的积极进步效果在于:本发明获得的全新的il-17a的抗体分子,特别是人抗体分子,其具有抑制il-17a相关生物学活性;可以治疗il-17a相关疾病,如风湿性关节炎、纤维化疾病、结肠癌以及乳腺癌等。其中9nt以及9nt.ns抗体都具有nm级的细胞中和活力和温度稳定性。9nt抗体具有中和il-17a诱导cxcl1趋化因子生成的活性。附图说明图1:利用酵母展示系统亲和力成熟抗体。利用酵母展示系统亲和力成熟抗体。a.技术路线图。分别构建重链随机突变库与轻链随机突变,经过两轮磁珠筛选和一轮流式分选获得亲和力提高的酵母克隆,再将这些克隆的重链和轻链重组经过三轮流式分选。b.流式细胞术检测每一轮分选的酵母细胞。横轴(x轴)为单链抗体的展示水平,代表细胞群抗体的表达量,用fitc标记。纵轴(y轴)为结合于抗体的抗原il-17a水平,用apc标记。经过两轮磁珠分选后,流式细胞术分选apc通道荧光强度最强的0.1%-0.3%。图2:利用酵母展示系统筛选与鉴定抗il-17a亲和力成熟抗体。a.第三轮流式分选后筛选获得的抗体克隆与10nmil-17a结合,流式细胞术分析抗体结合抗原的能力,x轴表示结合在单链抗体上抗原的荧光强度。b.利用流式细胞术分析抗il-17a野生型及突变型单链抗体与不同浓度的il-17a结合的平均荧光强度(mfi)。图3:抗体亲和力测定。(a)fortebio测定抗il-17a抗体亲和力;(b)抗il-17a抗体阻断il-17a刺激h1080分泌il-6。图4:抗il-17a抗体体外中和活性的检测。重组蛋白il-17a刺激人成纤维肉瘤细胞(ht1080)分泌il-6,将纯化的浓度梯度抗体9nt与il-17a共孵育后,刺激ht1080细胞,20小时后收集细胞上清elisa检测il-6细胞因子水平计算抑制il-17a生物活性的抑制曲线。纵坐标为il-6最大反应值(%),横坐标为抗体浓度。图a为改造抗体9nt中和活性;图b为改造抗体9nt.ns的中和活性。图5:抗体亲和力测定(biacore)。将抗体固定在芯片表面,从100nm起,2倍梯度稀释抗原。除100nm外,每个浓度重复两次。横坐标为进样时间,纵坐标为ru值。图a为抗体9nt,图b为抗体9nt.ns。图6:elisa方法显示温度稳定性。图a为抗体9nt.ns的稳定性,从55℃开始温度上升,孵育30分钟后取出,做elisa,检测od450,以4℃作为活性100%作图。图中x轴为温度,y轴为相对活性百分比。ec50值为77.06℃。图b为抗体9nt的热稳定性,ec50值为77.16℃。图7:dsc法测定抗il-17a抗体稳定性。(a)9nt.c53afab片段热稳定数值为76.3℃;(b)9nt.nsfab片段热稳定数值为77.4±1.2℃;(c)9ntfab片段热稳定数值为78.1±0.7℃。图8:抗il-17抗体9nt改善风湿性关节炎关节。a.9nt降低类风湿关节炎病人滑膜分泌il-17a水平;b.9nt降低类风湿关节炎病人滑膜分泌il-6水平;c.9nt降低类风湿关节炎病人滑膜产生i型胶原c前肽水平。图9:抗il-17a抗体生物学活性。给c57小鼠皮下注射人源化的il-17a,会导致小鼠体内发生免疫反应,表现在gro-α水平的升高。为了鉴定9nt抗体对il-17a的中和活性,将二者混合置于室温20分钟后,注入小鼠皮下组织。实验共设三组:皮下注射pbs,(n=4);皮下注射il-17a因子1.5mg/ml,(n=5);皮下注射室温预孵育的il-17a因子1.5mg/ml+9nt抗体2mg/ml,(n=5)。两个小时后眼底静脉丛取血,离心取上清。用cxcl1-elisa试剂盒检测血清中gro-α的存在。***p<0.001与pbs组比较,###p<0.001与il-17a处理组比较。图10:抗il-17a抗体抑制肺成纤维细胞迁移以及胶原生成。(a)1e5人肺成纤维细胞(mrc-5)置于transwell小室上层培养,上层及下层培养基均为mem培养基添加neaa,10%胎牛血清。10ng/ml人tgf-β细胞因子刺激人肺成纤维细胞迁移,抗il-17a抗体9nt抑制mrc-5细胞迁移。(b-c)抗il-17a抗体9nt抑制tgfbeta诱导mrc-5细胞胶原生成。(d)抗il-17a抗体9nt抑制tgfbeta诱导mrc-5细胞增殖。图11:抗il-17a抗体抑制肝星形细胞胶原生成。抗il-17a抗体9nt抑制il-17a诱导hhstec生成alpha-sma和胶原i。图12:抗il-17抗体抑制银屑病发展。nod小鼠接种人银屑病组织后第28天,开始给予抗il-17a抗体9nt(每周2次)。4周后取皮肤病理组织进行病理学分析,按照无恢复(1)、部分恢复(2)、部分到全部恢复(3)以及全部恢复(4)进行病理组织学评测。*p<0.05,**p<0.01与对照组(ctrl)比较。图13:抗il-17a抗体抑制结肠癌侵袭。1×105人结肠癌细胞hct-8与人igg对照抗体、抗il-17a抗体9nt或pbs孵育1小时后,加至transwell小室,16小时后计数转移细胞数目。pbs(a)、9nt(b)、人源抗体igg(c)典型图。(d)抗il-17a抗体9nt抑制结肠癌侵袭(n=3)。*p<0.05与对照组比较。图14:抗il-17a抗体抑制乳腺癌体外成球。5×102人乳腺癌mcf7细胞与人igg对照抗体、抗il-17a抗体9nt或pbs孵育1小时后,在96孔板与200lstemxvivotm无血清干细胞成球培养基培养7天后计数。抗il-17a抗体9nt抑制乳腺癌体外成球(n=3)。**p<0.01与igg处理组比较。具体实施方式下面通过实施例的方式进一步说明本发明,但并不因此将本发明限制在所述的实施例范围之中。下列实施例中未注明具体条件的实验方法,按照常规方法和条件,或按照商品说明书选择。实施例1抗体的亲和力成熟1.1酵母电转化感受态的制备1)从新鲜的ypd平板中,挑eby100酵母菌(invitrogentm)单克隆于5mlypd培养基中。30℃过夜培养。2)将过夜培养物接入至50mlypd培养基中,初始浓度为od600=0.1,30℃培养至od600=1.3-1.5,该过程大约需要6小时。3)当细胞培养至od600=1.3-1.5,加入500μltris-dtt缓冲液至培养基中。30℃在摇床中摇15min。4)4℃,2500g离心3min培养物,用25ml冰冷的1m山梨醇柔和重悬清洗细胞沉淀。离心,用1ml冰冷的1m山梨醇柔和重悬清洗细胞沉淀,离心。5)用冰冷的双蒸水重悬细胞沉淀至终体积为300μl,放置冰上待用。1.2利用易错pcr制备随机突变库1)构建随机突变库采用stratagene生产的iirandommutagenesis试剂盒。设计突变引物:for:5’-tgagtcagctcagaggaggcat-3’seqidno.36);rev:5’-gttagggataggcttaccttcgaag-3’(seqidno.37)。配置如下pcr体系:94℃变性3min;94℃变性45s,60℃退火30s,72℃延伸90s,35循环;72℃补平10min。2)琼脂糖凝胶电泳分离纯化突变pcr产物,pcr进一步富集该产物以如下体系:上游引物(10μm)5μl下游引物(10μm)5μl10×taq缓冲液(不含mgcl2)10μlmgcl2(50mm)4μldntp(20nm)2μl突变的pcr产物4μltaqdna聚合酶(5u/μl)1μl加去离子水至100μl。以如下循环扩增dna片段:94℃变性3min;94℃变性45s,60℃退火30s,72℃延伸90s,30循环;72℃补平10min。3)将pcr产物电泳纯化后,切胶试剂盒(购自qiagen)回收,定量。与pyd2酵母展示质粒一同酶切,条件如下:37℃孵育3h后终止反应,1%tae电泳分离酶切片段。1.3酵母-dna连接产物的电转化1)准备4个电转杯,以5μgpcr纯化产物片段与1μg线性化pyd2载体连接,本实验中1μg载体对应一个电转杯。理论上转四次合集库容可达1×107。2)取1.1实验例中制备的50μl感受态细胞与本试验中获得的连接产物混合,并加入电转杯中,该过程始终保持低温,电转前,电转杯放置冰上。3)电转条件:0.54kv,25uf(酵母程序)。电转后向电转杯中迅速加入预热的(30℃)ypd培养基1ml。4)将这1ml电转化产物转至15ml离心管中,再用1ml洗电转杯。四次合并,30℃摇1h。5)4℃,2500g离心5min去上清。用10mlsdcaa培养基重悬细胞沉淀,即获得转化菌。取10ul菌液梯度稀释涂sdcaa平板,计算库容及转化效率。附:sdcaa(1×)培养基的配制:5g/l酪蛋白水解物(casaminoacids)(bd,223120);20g/l葡萄糖;6.7g/lynb(yeastnitrogenbasewithammoniumsulfatew/outaminoacids)(bd,1364756);13.6g/lna2hpo4·12h2o;9.68g/lnah2po4·2h2o;山梨醇182.17g/l(amresco);其中,葡萄糖、na2hpo4·12h2o以及nah2po4·2h2o均购自北京化学试剂公司。1.4构建酵母单链突变抗体库1)将10ml1.3步骤中制得的转化菌以体积比1:100转接入1000ml含有链青霉素(青霉素的工作浓度为100u/ml,链霉素的工作浓度为0.1mg/ml)sdcaa(1×)培养基30℃摇48小时。2)培养物吸光值一般可达od600=6-8,诱导前,细胞需要全新传代。为尽可能减少死细胞,以1×1010个细胞于新鲜的sdcaa(1×)培养基中30℃摇48h,即为传代细胞库。3)再从扩增后的传代细胞库中取1×1010个细胞,用50ml离心管2500g离心5min弃上清,加入sgcaa培养基,至细胞终密度为od600=0.5-1,在20℃震荡培养20小时条件下,含有单链抗体基因的酵母质粒将被诱导表达,抗体片段展示于酵母细胞表面。(备注:sdcaa培养基为扩增培养基,sgcaa培养基为诱导培养基)1.5流式分选亲和力成熟片段1)取上述诱导培养基中诱导后5×107个酵母细胞,15000rpm离心30s将细胞收集于ep管中,去上清,用1mlpbsb缓冲液(pbs溶液,加入0.1%bsa)清洗。2)第一轮筛选加入100μm浓度生物素化il-17a抗原(为peprotech公司的重组il--17a抗原,货号:#200-17)与酵母细胞混合,第二轮加入10μm,第三轮加入1μm,第四轮加入0.1μm生物素化标记il-17a抗原。室温孵育1h。3)14000g4℃离心30s沉淀细胞,用1ml冰冷的pbsb缓冲液清洗细胞。4)200μl重悬细胞,选择alexafluor488羊抗鸡igg(体积比1:100稀释)和链亲和素-藻红蛋白(pe)(1:100稀释),涡旋细胞以重悬。在冰上避光孵育的细胞10-20分钟。5)14000g,4℃离心30s沉淀细胞并用1mlpbsb缓冲液清洗细胞。保持细胞沉淀在冰上。6)设出一个适当的分类门在双阳性四分之一象限以分离scfv阳性表达细胞和抗原结合。在第一轮流式细胞分选中,习惯使用一个非常保守选择设置门大约在细胞群顶端的5%,以避免失去独特的克隆。随着细胞群被富集,可以设置更多的对角分类窗口,收集顶端0.1-1%。7)收集分选的细胞于1mlsdcaa培养基.随后的几轮分选扩增筛选到的酵母。加sdcaa培养基到洗脱细胞悬液,终体积为5ml,加链青霉素(体积比1:100稀释),30℃过夜培养。8)分别构建重链随机突变库与轻链随机突变(chaoetal,natprotoc.2006;1(2):755-68;mayorovetalbmcimmunol.2005jun10;6:10;mcconnelletal,plosone.2012;7(11):e49458),经过两轮磁珠筛选和一轮流式分选获得亲和力提高的酵母克隆,再将这些克隆的重链和轻链重组经过三轮流式分选。9)流式细胞术检测每一轮分选的酵母细胞。经过两轮磁珠分选后,流式细胞术分选apc通道荧光强度最强的0.1%-0.3%。参照图1。10)第三轮流式分选后筛选获得的抗体克隆与10nmil-17a(为peprotech公司的重组il--17a抗原,货号:#200-17)结合,流式细胞术分析抗体结合抗原的能力,fortebio进一步确定抗原抗体结合能力(具体参见下述实施例4),使用ht1080体外筛选系统鉴定生物学活性(具体参见下述实施例3)。结果:经过多轮流式分选获得多个高亲和力的抗体片段b9,d1,d3,d4,d5,d7,d8,h9,2f6。d1、d3(即9nt.d3)和d5亲和力最优(见图2和表2)。d1和d3体外表达量远超过d5。进一步亲和力测定和中和活性研究发现,d3(即9nt.d3)比d1具有更高的亲和力(kd:2.3e-9v.s.6.1e-9)和更好的生物学活性(ic50:11nmv.s.21nm)。9nt.d3(抗体1)经框架移植获得9nt.c3(抗体2),经框架区关键位点突变获得更高亲和力抗体9nt.c53a(抗体3)、9nt.ns(抗体4)以及9nt(抗体5)。表2.亲和力kd测定所获得9nt.d3抗体片段通过测序获得该片段的重链氨基酸序列如:seqidno.1所示;该片段的轻链氨基酸序列如:seqidno.2所示;重链氨基酸序列如:seqidno.1对应的核苷酸序列(即重链核苷酸序列)为:seqidno.8;轻链氨基酸序列对应的核苷酸序列(即轻链核苷酸序列)为:seqidno.9。各抗体片段vh和vl的氨基酸及核酸序列在序列表中的编号如上表1所示。互补决定区(cdrs)是参与抗原结合的抗体可变区的残基。我们采用katat定义标识cdrs区域。根据kabat算法,本发明的重链可变区以及轻链可变区均有三个cdr区,分别命名为hcdr1、hcdr2、hcdr3、lcdr1、lcdr2以及lcdr3;本发明5个抗体的上述cdrs氨基酸序列均相同,其氨基酸序列分别如seqidno.15~20所示。而框架区中hfr4及lfr4与现有抗体(请参见cn104231080a)相同,故不再序列表中列出;而9nt.d3抗体的hfr1、hfr2、hfr3的氨基酸序列分别如seqidno.24、25、26所示,lfr1、lfr2、lfr3的氨基酸序列分别如seqidno.33、34、35所示;9nt.c3抗体的hfr1、hfr2、hfr3的氨基酸序列分别如seqidno.21、22、23所示,lfr1、lfr2、lfr3的氨基酸序列分别如seqidno.27、28、29所示;9nt.c53a抗体的lfr3的氨基酸序列如seqidno.30所示,余同9nt.c3抗体;9nt.ns抗体的lfr3的氨基酸序列如seqidno.31所示,余同9nt.c3抗体;9nt抗体的lfr3的氨基酸序列如seqidno.32所示,余同9nt.c3抗体。具体见下表3所示。表3不同的框架区以及抗原互补决定区的氨基酸序列编号实施例2全长抗体蛋白表达与纯化1)293ft悬浮细胞需要维持4×105至3×106个细胞/ml的浓度范围内,通常情况在125mlerlenmeyer培养瓶中培养25ml,摇床转速120rpm,37℃,8%co2。2)转染细胞24小时前,按照1×106个细胞/ml分细胞并过夜培养,次日细胞浓度应该达到2×106个细胞/ml。3)转染细胞当天,对细胞进行计数,通过离心重悬细胞使其浓度为2.5-3.0×106个细胞/ml之间。4)转移10ml重悬细胞至一个新的125mlerlenmeyer培养瓶中,再次确认细胞浓度,将细胞置于细胞培养箱中培养。5)抗体轻链和重链的基因片段通过xho1和bamh1双酶切,t4酶连接至ptt5表达载体中。测序正确后大提质粒获得抗体轻链和重链。将30μg含有表达抗il-17抗体的质粒混合物(抗体轻链:抗体重链=1:1,质量比,15μg抗体轻链与15μg抗体重链混合)加入到1mlopti-mem培养液,短暂涡旋后,离心将所有液体至于ep管底部,然后加入45μlpei聚醚酰亚胺储存液(1mg/ml),再次短暂涡旋后,室温放置10分钟。6)将上述质粒-pei/opti-mem混合物缓慢滴入293ft悬浮细胞中,将培养瓶放回培养箱,培养3小时。7)在细胞中加入15ml预热好的培养基,继续培养。8)转染后3-4天离心收集第一批上清,在细胞培养瓶中加入等体积新鲜的培养基,7天后收集第二批上清。9)收集到的上清用0.22μm滤膜过滤后进行抗体蛋白纯化,上清通过5mlproteinasepharoseff柱,ph7.4的pbs平衡,未结合组分用pbs缓冲液洗净,目标抗体蛋白用ph3.0缓冲液洗脱并立即中和。10)中和后的抗体用50kd浓缩柱浓缩至2-5mg/ml,并用superdex200层析柱进行分离,只收集目标抗体蛋白,再次用50kd浓缩柱浓缩至所需浓度,0.22μm滤膜过滤后4℃保存。结果:纯化后全长抗体分别在还原性及非还原性sds-page电泳检测,获得纯度约为95%的全长抗体蛋白为9nt-igg。实施例3抗体体外中和实验il-17a可诱导刺激ht1080(atcc)产生细胞因子il-6。本实验通过il-17a诱导ht1080细胞产生il-6活力检测抗il-17a抗体的中和活性。1)胰酶消化人纤维肉瘤细胞系(ht1080),于96孔板中。细胞计数,使得每孔中约有9000个细胞,在含有10%胎牛血清mem培养基中,培养过夜。2)无菌pbs稀释重组蛋白il-17a(peprotech公司重组il-17a抗原,货号:#200-17)为10nm。3)无菌pbs梯度稀释纯化后重组抗体9nt.ns、9nt(实施例1),分别为:1350nm、337.5nm、84.38nm、21.09nm、5.27nm、1.32nm、0.33nm、0.08nm、0.02nm及0.01nm十个梯度,与10nm重组il-17a蛋白混合,去除ht1080细胞培养上清,加入抗体抗原共孵育溶液,每个梯度5个复孔,过夜培养。4)次日收集细胞上清,利用商业化il-6细胞因子检测试剂盒(biolegend),按照试剂盒说明书步骤检测各孔细胞il-6表达的量,利用graphpadsoftware5.0软件绘制量效关系曲线,并在图中得出单链抗体ec50值。结果:通过绘制量效关系曲线,如图4。通过5次独立实验计算各抗体ec50值,如表4。表4.中和性抗体的ec50值结果提示,抗il-17a抗体9nt以及9nt.ns都具有nm级的细胞中和活力。抗il-17a抗体9nt比较9nt.ns具有更高的中和活力。实施例4抗体亲和力的检测4.1biacoret100测定抗体亲和力1)利用biacoret100(ge公司)表面等离子共振技术检测单抗的亲和力。以氨基偶联方式,目标偶联水平为1200ru,将抗体用ph5.0的10mm醋酸钠固定缓冲液稀释到20g/ml,乙醇胺作为洗涤缓冲液。2)再生缓冲液为ph1.5的甘氨酸,注入时间为30s,流速调为30l/min。开始程序将抗体固定在cm5芯片(generalelectric,ge)上。3)hbs-ep缓冲液洗2小时,以1-乙基-3-(3-二甲基氨丙基)-碳化二亚胺(edc)和n-羟基琥珀酰亚胺(nhs)交联剂,活化cmd的羧基。梯度稀释il-17a蛋白,浓度从100g/ml,稀释终浓度为:0nm,0.09765625nm,0.1953125nm,0.390625nm,0.78125nm,1.5625nm,3.125nm,6.25nm,12.5nm,25nm,50nm,100nm,除100nm外,每个浓度重复一次,作为对照。注入时间为500sec,解离时间为600sec,流速调为30l/min。按仪器提示加入各试剂,运行程序。4)程序结束,可见抗体与抗原相结合的动力学图形,根据模型计算单抗的平衡解离常数,如图5。结果说明,抗il-17a抗体9nt、9nt.ns均具有nm级高亲和力,9nt比较9nt.ns具有更高的亲和力。结果:检测数据如表5。表5.biacoret100表面等离子共振技术检测抗体的亲和力4.2fortebio测定抗体亲和力octetred96检测人源抗体与其抗原间亲和力。光线在传播过程中会由于光程差的存在而形成干涉,光程差的变化会导致干涉图样的变化;生物膜干涉技术正是利用干涉原理进行分子间相互作用的检测:生物分子在传感器表面结合形成光程差,当所用光源为复色光如白光时会获得干涉谱,而分子的结合与解离会导致光程差变化,因此引起干涉谱的变化,生物膜干涉技术所检测信号即为生物分子结合或解离所导致的干涉谱的偏移量—亲和力越强,分子结合越快,解离越慢,干涉谱在结合过程中偏移越快,而在解离过程中偏移越慢,由此通过计算干涉光谱偏移快慢可获得分子间亲和力大小。抗体(人)分子量150kda,工作浓度10μg/ml;抗原分子量66kda,工作浓度500nm-7.81nm,2倍稀释;人抗固定化在ahc传感器上,固化浓度为10μg/ml,检测浓度梯度抗原(500nm~7.81nm)。步骤:(1)将传感器(sensor)泡在加有200μlhepes缓冲液的96孔板中,室温放置10min。(2)按照如下顺序,在greinerbio-one的96孔板中各加入200μl:hepes缓冲液、il-17a(设置不同浓度梯度)、甘氨酸(gly)再生液、hepes缓冲液。(3)设置程序:基线,结合抗原,基线,再生,基线。结果如图3和表6所示,抗体9nt.d3(d3)具有较高的亲和力和中和活性。表6.抗il-17a抗体亲和力及中和活性ic50汇总表实施例5抗体稳定性的检测5.1elisa法测il-17a抗体稳定性1、温度稳定性样品处理1)用pbs稀释抗体,终浓度为5μg/ml,40μl分装,使用pcr仪设置温度梯度,孵育30min。2)每管加入160μlpbs,混匀。2、温度稳定性elisa1)包被。用碳酸盐缓冲液配置il-17a,终浓度为80ng/ml。加入到96孔板(康宁,货号为3690)中,25μl/孔。用密封膜封闭,4℃过夜。2)次日拿出96孔板,甩出包被液,拍板。3)pbs(含有1‰tween20)洗板,180μl/孔,2min/次,共5次,在最后一次拍板。4)pbs(含有1%bsa)封闭,每孔加入100μl,密封膜封闭,37℃孵育1小时。5)重复步骤3。6)每孔加入25μl的处理过后的样品,密封膜封闭,37℃孵育1小时。7)重复步骤3。8)用pbs(含有1%bsa)1:8000(体积比)稀释二抗(羊抗人igghrp标记),50μl/孔加入到96孔板中,密封膜封闭,37℃孵育1小时。9)重复步骤3。10)在96孔板中加入显色液,50μl/孔。11)2m硫酸,50μl/孔终止显色,使用酶标仪读读od450与od650。12)用graphpadprism5作图,横坐标为温度,纵坐标为相对活性百分比。如表7及图6。结果说明抗il-17a抗体9nt(77.2±0.6)与9nt.ns(76.1±2.2)相比具有更高的温度稳定性。表7.抗il-17a抗体温度稳定性(elisa)5.2dsc法测il-17a抗体稳定性。nanodsc测量样品加热或冷却时吸收或释放的热量。传统的dsc仪器是为广泛的应用而设计,但是在生物样品的研究中往往缺少灵敏度。像蛋白质这样的大分子在特定的温度下通过去折叠对加热或冷却作出响应;生物聚合物固有的稳定性越高,其去折叠转化的温度变化中点就越高。这些过程往往伴随着微焦耳级热量的转换,nanodsc的灵敏度是成功研究这些反应的关键。nanodsc采用创新的双重毛细管设计,在能量补偿模式下运行。毛细管设计能够延迟蛋白质团聚,直到去折叠过程完成为止,从而使dsc测量具有无与伦比的灵敏度、准确性和精密度。nanodsc采用固态热电元件来精密而准确地控制样品的温度。因为采用同样的元件进行加热和冷却,所以向上和向下的温度扫描均能得到相同的灵度。nanodsc压力控制是通过内置的由电脑控制的精确线性致动器来驱动的高压活塞获得的。dsc实验中采取恒压是为了获得恒压热容(cp)以及防止起泡或沸腾。在压力微扰实验中,可根据用户可选择的模式改变压力,从而获得压缩系数和热膨胀数据。(1)样品:0.5-1mg/ml,至少600μl;(2)打开软件,旋开机器盖子,用400ml超纯水冲洗两个管道;(3)在s管道和r管道充满样品缓冲液,盖好小黑帽,旋紧盖子;(4)用带白套管的蓝枪头吸取700μl溶液,对准c口;用另一个带白套管的蓝枪头对准a口;轻轻吹打,使样品充满整个管道并且没有气泡;用纸巾擦掉多余溶液;盖好c的小黑帽;bd充满缓冲液,方法同上;(5)设置程序:温度范围,升温速度等;(6)运行程序,输入文件名,先加压至3pa,开始实验。缓冲液进行3次实验,以获得良好的空白图像,可根据实验调整;(7)冲洗管道,上待测样品,运行;(8)降压至0.3,打开盖子,冲洗管道,关闭仪器。结果见图7及表8。结果说明,抗il-17a抗体9nt(78.1±0.7)与9nt.ns(77.4±1.2)相比具有更高的温度稳定性。表8.抗il-17a抗体热稳定性实施例6抗体中和人il-17a改善风湿性关节炎关节6.1风湿性关节炎体外模型制备1)收集风湿性关节炎患者关节置换术中获得滑膜样本。2)无菌条件下,样本被切成2立方毫米小块并分成数份置于2块24孔板中。3)样本利用mem(gibco,grandisland,ny,usa)完全培养基(含有2mml-谷氨酰胺,100u/ml青霉素(penicillin),50mg/ml庆大霉素(gentamicin),20mmhepes缓冲液and1%胎牛血清)每孔2ml,37℃,5%co2,95%湿度培养。6.2治疗活性检测1)按照6.1制备风湿性关节炎体外模型,分成抗体处理组及未处理对照组,每组6个复孔。2)抗体处理组中使用9nt抗体处理滑膜培养物,抗体终浓度为10nm,未处理组加等体积生理盐水,培养24h。3)24h后收集各孔细胞培养上清,使用商业化细胞因子检测试剂盒(r&dsystem),按照试剂盒说明书进行操作,双夹心酶联免疫吸附法检测培养液中il-17a、il-6、i型胶原c前肽等含量。4)利用graphpadsoftware5.0软件柱状图,t检验统计分析各组差异,结果如图8。结果显示:9nt与未处理组相比较,9nt可以中和人il-17a,改善风湿性关节炎相关因子。实施例7抗il-17a抗体生物学活性人源il-17a蛋白经皮下注射c57bl/6小鼠可诱导免疫反应,刺激包括gro-α(cxcl1)等趋化因子的生成。本实验通过抗il-17a抗体与il-17a蛋白体外孵育后注射,评价抗il-17a抗体的中和活性。7.1取样。选取5-8周龄的spf级雌性c57小鼠,分成四组,每组5只。配制三组小鼠的注射样品:a组:pbs,(n=4);b组:人源il-17a因子1.5mg/kg,(n=5);c组:人源il-17a因子1.5mg/kg+9nt抗体2mg/kg,(n=5)。室温孵育20分钟,皮下注射c57小鼠,400μl/只。两个小时后眼底静脉丛取血,离心取上清。7.2用cxcl1-elisa试剂盒检测血清中cxcl1(gro-α)的存在:(1)每孔加入300μl的1×洗液静置浸泡30s。弃液,拍板。每孔加入50μl的1×检测缓冲液;15分钟内,再加入血清样品或标准品,50μl/孔;每孔最后加50μl稀释的检测抗体。封板膜封板,300rpm/min振荡,室温孵育2小时;弃液,每孔加入300μl的1×洗液静置浸泡30s。弃液,拍板。重复六次。每孔加入100μl稀释的hrp,新膜封板。300rpm/min振荡,室温孵育45分钟。弃液,洗板6次。每孔加入100μltmb,避光显色。室温孵育5-30分钟。每孔加入100μl终止液。30分钟内扫板,读取od450-od570数值。用graphpadprism5作图。结果见图9。结果说明,抗il-17a抗体9nt具有中和il-17a诱导cxcl1趋化因子生成的活性。实施例8抗il-17a抗体抑制肺纤维化8.1将10000个人肺成纤维细胞(mrc-5)置于transwell小室上层培养,上层及下层培养基均为mem培养基添加neaa,10%胎牛血清。将10ng/ml人促纤维化细胞因子tgf-β添加于上层培养基中,刺激人肺成纤维细胞,同时加入人源igg对照抗体或抗il-17a抗体9nt,24小时候取出transwell小室,用结晶紫进行染色。水洗三次各5min,随后使用棉签轻轻擦除transwell小室内侧细胞。使用bx-51dp72纤维成像系统观察迁移的肺成纤维细胞数量。8.2使用6孔板培养人肺成纤维细胞mrc-5(1×106),培养体系为mem培养基添加neaa,10%胎牛血清,总体积为2ml。在人肺成纤维细胞中使用lipofectamine2000转染i型胶原的荧光素酶质粒(5mg),转染24小时后使用10ng/ml人tgf-b细胞因子刺激成纤维细胞,同时加入对照抗体或9nt抗体,24小时后加入报告基因细胞裂解液(碧云天)。融解萤火虫荧光素酶检测试剂和renilla荧光素酶检测缓冲液,并达到室温。renilla荧光素酶检测底物(100×)置于冰浴或冰盒上备用。按照每个样品100微升的量,取适量renilla荧光素酶检测缓冲液,按照1:100加入renilla荧光素酶检测底物(100×)配制成renilla荧光素酶检测工作。按仪器操作说明书开启荧光测定仪,将测定间隔设为2秒,测定时间设为10秒。8.3使用6孔板培养人肺成纤维细胞mrc-5(1×106),培养体系为mem培养基添加neaa,10%胎牛血清,总体积为2ml。使用10ng/ml人tgf-β细胞因子刺激成纤维细胞,同时加入对照抗体或9nt抗体,24小时后收集细胞培养上清。使用abcam(ab210966)elisa试剂盒检测细胞培养上清中的i型胶原含量。8.4使用6孔板培养人肺成纤维细胞mrc-5(1×104),培养体系为mem培养基添加neaa,10%胎牛血清,总体积为2ml。使用10ng/ml人tgf-β细胞因子刺激成纤维细胞,同时加入对照抗体或9nt抗体,培养后的第1、2、4、6天统计人肺成纤维细胞数量。8.5结果见图10。结果说明抗il-17a抗体9nt具有抑制促纤维化细胞因子tgf-β诱导引起的人肺成纤维细胞的迁移、增殖以及i型胶原的生成,提示该抗体具有抑制纤维化疾病的前景。实施例9抗il-17a抗体抑制肝纤维化人肝星形细胞hhstec细胞肝纤维化形成过程中发挥重要作用。il-17a促进人肝星形细胞hhstecalpha-sma和i型胶原的生成。我们通过alpha-sma和i型胶原生成量评价抗il-17a抗体的中和能力。使用6孔板培养人肝星形细胞hhstec细胞(1×105),培养体系为stecm。使用人il-17a细胞因子刺激星形细胞,同时加入对照抗体或9nt抗体,24小时后收集细胞,ripa缓冲液裂解。western检测alpha-sma和i型胶原表达量。结果见图11。结果说明抗il-17a抗体9nt抑制il-17a引起的alpha-sma和i型胶原生成,提示该抗体具有抑制纤维化疾病的前景。实施例10抗il-17a抗体抑制银屑病发展10.1银屑病病理组织1)收集银屑病患者皮肤样本组织。2)无菌条件下,样本被切成约1×1毫米小组织块。10.2动物模型选取8-10周龄的spf级雌性nod.scid小鼠,分成四组,每组5只。皮下埋植病理组织块。动物分组情况如下:a组:pbs,(n=5);b组:抗il-17a抗体9nt5mg/kg,(n=5)。c组:抗il-17a抗体9nt10mg/kg,(n=5)。皮下接种银屑病病理组织块28天后,开始给予抗il-17a抗体(每周2次),四周后获取组织并进行病理分级:1为银屑病组织无恢复;2为部分恢复;3为部分到全部恢复;4为完全恢复。结果见图12。结果说明抗il-17a抗体9nt抑制il-17a引起的alpha-sma和i型胶原生成,提示该抗体具有抑制纤维化疾病的前景。实施例11抗il-17a抑制结肠癌细胞迁移1、人结肠癌细胞hct-8细胞无血清饥饿处理2小时;配制200ng/l人igg对照抗体、抗人il-17a抗体9nt,置于冰上备用;2、用无血清/无双抗的dmem按1:100倍比(体积比)稀释fibronectin母液(2μg/l),吸取20μl纤连蛋白(fibronectin)工作液,均匀涂于每个小室外侧,静置风干2h;3、将24孔板每孔加入700μl含有10%fbs的完全培养液,并将小室置于孔中,确保小室底部和培养液之间没有气泡;4、调整hct-8细胞浓度为5×105/ml,取200μl无血清的细胞悬液,分别加入5lpbs、5l抗il-17a抗体9nt(200ng/l)、5l人igg对照抗体(200ng/l),置于37℃孵箱中共孵育1小时;5、将孵育完的细胞接种于transwell小室,置于37℃恒温培养箱中培养;6、16小时后,弃去小室内的培养液,将小室取出,用pbs或生理盐水清洗小室底部3次,用4%多聚甲醛固定20min,蒸馏水清洗三次,用棉签轻轻擦去小室内部细胞,用结晶紫室温染色30min,蒸馏水洗3次,于倒置显微镜下观察并计数。7、结果说明抗il-17a抗体9nt阻断il-17a引起结肠癌细胞转移,提示该抗体具有抑制肿瘤转移的前景。实施例12抗il-17a降低乳腺癌干性,抑制体外成球能力。1、stemxvivoserum-freetumorspheremedia置于37℃预热。收集人乳腺癌细胞mcf7细胞细胞,制备成5×103/ml细胞悬液。2、细胞悬液与5g/ml人igg对照抗体、抗il-17a抗体9nt以及pbs置于37℃孵箱中共孵育1小时。3、细胞悬液补充含2u/ml肝素和0.5μg/ml氢化可的松的stemxvivoserum-freetumorspheremedia,混匀并避免产生气泡。4、按照100l/孔加至低吸附的96孔板中,培养7d。用相差显微镜观察拍照计数。5、结果说明抗il-17a抗体9nt阻断mcf7乳腺癌细胞体外成球能力,提示抗il-17a抗体9nt具有抑制肿瘤干性的能力。sequencelisting<110>北京伟峰益民科技有限公司<120>一种全人源抗人白介素17a抗体及其应用<130>p1611309c<160>37<170>patentinversion3.5<210>1<211>121<212>prt<213>artificialsequence<220><223>9nt.d3-vh<400>1glnvalglnleuglnglnserglyproglyleuvallysproprogln151015thrleuserleuthrcysalaileserglyaspservalserserasn202530seralaalatrpasptrpileargglnserproserargglyleuglu354045trpleuglyargthrtyrtyrargserlystrpserhisasptyrala505560valservallysasnargilethrileasnproaspthrserlysasn65707580glnpheserleuglnleuasnservalserprogluaspthralaval859095tyrtyrcysvalarggluglyasptyrglyasptyrglytyrtrpgly100105110glnglythrleuvalthrvalserser115120<210>2<211>109<212>prt<213>artificialsequence<220><223>9nt.d3-vl<400>2sersergluleuthrglnaspproalavalservalalaleuglygln151015thrvalargilethrcysglnglyaspserleuargglytyrtyrala202530sertrptyrglnglnlysproglyglnalaprovalleuvaliletyr354045tyraspaspaspargprothrglyileprogluargpheserglyser505560asnserglyasnseralathrleuthrileserargvalglualagly65707580aspglualaasptyrtyrcysalaalatrpaspvalserleuasngly859095provalpheglyglyglythrlysvalthrvalleuser100105<210>3<211>121<212>prt<213>artificialsequence<220><223>9nt.c3、9nt.c53a、9nt.ns以及9nt共有重链的氨基酸序列<400>3gluvalglnleuglnglnserglyproglyleuvalglnproglygly151015serleuargleusercysalaileserglyaspservalserserasn202530seralaalatrpasptrpileargglnalaproglylysglyleuglu354045trpleuglyargthrtyrtyrargserlystrpserhisasptyrala505560aspservallysglyargilethrileserproaspthrserlysasn65707580thrphetyrleuglnleuasnserleuargalagluaspthralaval859095tyrtyrcysvalarggluglyasptyrglyasptyrglytyrtrpgly100105110glnglythrleuvalthrvalserser115120<210>4<211>109<212>prt<213>artificialsequence<220><223>9nt.c3-vl<400>4aspileglnleuthrglnserproserserleuseralaservalgly151015aspargvalthrilethrcysglnglyaspserleuargglytyrtyr202530alasertrptyrglnglnlysproglylysalaprolysleuleuile354045tyrtyraspaspaspargprothrglyvalproserargphesergly505560serasnserglythraspalathrleuthrileserserleuglnpro65707580gluaspphealathrtyrtyrcysalaalatrpaspvalserleuasn859095glyprovalpheglyglnglythrlysvalgluilelys100105<210>5<211>109<212>prt<213>artificialsequence<220><223>9nt.c53a-vl<400>5aspileglnleuthrglnserproserserleuseralaservalgly151015aspargvalthrilethrcysglnglyaspserleuargglytyrtyr202530alasertrptyrglnglnlysproglylysalaprolysleuleuile354045tyrtyraspaspalaargprothrglyvalproserargphesergly505560serasnserglythraspalathrleuthrileserserleuglnpro65707580gluaspphealathrtyrtyrcysalaalatrpaspvalserleuasn859095glyprovalpheglyglnglythrlysvalgluilelys100105<210>6<211>109<212>prt<213>artificialsequence<220><223>9nt.ns-vl<400>6aspileglnleuthrglnserproserserleuseralaservalgly151015aspargvalthrilethrcysglnglyaspserleuargglytyrtyr202530alasertrptyrglnglnlysproglylysalaprolysleuleuile354045tyrtyraspaspalaargprothrglyvalproserargphesergly505560serasnserglyasnseralathrleuthrileserserleuglnpro65707580gluaspphealathrtyrtyrcysalaalatrpaspvalserleuasn859095glyprovalpheglyglnglythrlysvalgluilelys100105<210>7<211>109<212>prt<213>artificialsequence<220><223>9nt-vl<400>7aspileglnleuthrglnserproserserleuseralaservalgly151015aspargvalthrilethrcysglnglyaspserleuargglytyrtyr202530alasertrptyrglnglnlysproglylysalaprolysleuleuile354045tyrtyraspaspalaargprothrglyvalproserargphesergly505560serasnserglyasnthralathrleuthrileserserleuglnpro65707580gluaspphealathrtyrtyrcysalaalatrpaspvalserleuasn859095glyprovalpheglyglnglythrlysvalgluilelys100105<210>8<211>363<212>dna<213>artificialsequence<220><223>9nt.d3-vhdna<400>8caggtacagctgcagcagtcaggtccaggactggtgaagcccccgcagaccctctcactt60acatgtgccatctccggggacagtgtctctagcaacagtgctgcttgggactggatcagg120cagtccccatcgagaggccttgagtggctgggaaggacatactacaggtccaagtggtct180catgattatgcagtgtctgtgaaaaatcgaataaccatcaacccagacacatccaagaat240cagttttccctgcagttgaactctgtgtctcccgaggacacggctgtgtattactgtgta300agagagggggactacggtgactacgggtactggggccagggaaccctggtcaccgtctcc360tca363<210>9<211>327<212>dna<213>artificialsequence<220><223>9nt.d3-vldna<400>9tcctctgagctgactcaggaccctgctgtgtctgtggccttgggacagacagtcaggatc60acatgccaaggagacagcctcagaggctattatgcaagctggtaccagcagaagcctggc120caggcccctgtactggtcatctattatgatgacgaccggcccacagggatccctgagcga180ttctctggctccaactctgggaactcggccaccctgaccatcagcagggtcgaagccggg240gatgaggctgattattactgtgcagcatgggatgtcagcctgaatggtccggtgttcggc300ggagggaccaaggtcaccgtcctatcc327<210>10<211>363<212>dna<213>artificialsequence<220><223>9nt.c3、9nt.c53a、9nt.ns以及9nt共有重链的核苷酸序列<400>10gaagtacagttgcagcaatctggacccggcctggtgcaacccggtgggagcttgagactc60agttgcgccatctctggcgattcagtctcaagcaacagcgccgcctgggattggatccga120caggcgcccggcaagggtcttgagtggttgggccgcacctactatcggagcaaatggagt180catgattacgcagatagtgtcaaggggagaatcacgatctctcccgatacaagtaaaaac240accttttatcttcagctcaattcactccgcgccgaggacaccgccgtgtactactgtgtg300cgcgaaggcgactacggcgactacggttattggggccagggcaccttggtcaccgtttca360tct363<210>11<211>327<212>dna<213>artificialsequence<220><223>9nt.c3-vldna<400>11gatatccaactcactcaaagcccatcctctctcagcgcctcagtgggcgatcgggtaacg60atcacttgtcagggtgattcccttcgaggctattatgcttcatggtaccagcagaagccc120gggaaggctcccaagctcctcatttattatgatgacgacagacctaccggtgtccctagc180cgctttagcggttccaatagcgggaccgatgccactcttaccatctcttcattgcagcct240gaagatttcgcaacctactattgtgccgcttgggacgtttctctgaacgggcctgtgttc300gggcagggcacaaaagttgagattaaa327<210>12<211>327<212>dna<213>artificialsequence<220><223>9nt.c53a-vldna<400>12gatatccaactcactcaaagcccatcctctctcagcgcctcagtgggcgatcgggtaacg60atcacttgtcagggtgattcccttcgaggctattatgcttcatggtaccagcagaagccc120gggaaggctcccaagctcctcatttattatgatgacgccagacctaccggtgtccctagc180cgctttagcggttccaatagcgggaccgatgccactcttaccatctcttcattgcagcct240gaagatttcgcaacctactattgtgccgcttgggacgtttctctgaacgggcctgtgttc300gggcagggcacaaaagttgagattaaa327<210>13<211>327<212>dna<213>artificialsequence<220><223>9nt.ns-vldna<400>13gatatccaactcactcaaagcccatcctctctcagcgcctcagtgggcgatcgggtaacg60atcacttgtcagggtgattcccttcgaggctattatgcttcatggtaccagcagaagccc120gggaaggctcccaagctcctcatttattatgatgacgccagacctaccggtgtccctagc180cgctttagcggttccaatagcgggaacagcgccactcttaccatctcttcattgcagcct240gaagatttcgcaacctactattgtgccgcttgggacgtttctctgaacgggcctgtgttc300gggcagggcacaaaagttgagattaaa327<210>14<211>327<212>dna<213>artificialsequence<220><223>9nt-vldna<400>14gatatccaactcactcaaagcccatcctctctcagcgcctcagtgggcgatcgggtaacg60atcacttgtcagggtgattcccttcgaggctattatgcttcatggtaccagcagaagccc120gggaaggctcccaagctcctcatttattatgatgacgccagacctaccggtgtccctagc180cgctttagcggttccaatagcgggaacaccgccactcttaccatctcttcattgcagcct240gaagatttcgcaacctactattgtgccgcttgggacgtttctctgaacgggcctgtgttc300gggcagggcacaaaagttgagattaaa327<210>15<211>10<212>prt<213>artificialsequence<220><223>hcdr1<400>15glyaspservalserserasnseralaala1510<210>16<211>9<212>prt<213>artificialsequence<220><223>hcdr2<400>16thrtyrtyrargserlystrpserhis15<210>17<211>12<212>prt<213>artificialsequence<220><223>hcdr3<400>17valarggluglyasptyrglyasptyrglytyrtrp1510<210>18<211>6<212>prt<213>artificialsequence<220><223>lcdr1<400>18serleuargglytyrtyr15<210>19<211>3<212>prt<213>artificialsequence<220><223>lcdr2<400>19tyraspasp1<210>20<211>12<212>prt<213>artificialsequence<220><223>lcdr3<400>20alaalatrpaspvalserleuasnglyprovalphe1510<210>21<211>25<212>prt<213>artificialsequence<220><223>9nt.c3、9nt.c53a、9nt.ns以及9nt共有的hfr1<400>21gluvalglnleuglnglnserglyproglyleuvalglnproglygly151015serleuargleusercysalaileser2025<210>22<211>17<212>prt<213>artificialsequence<220><223>9nt.c3、9nt.c53a、9nt.ns以及9nt共有的hfr2<400>22trpasptrpileargglnalaproglylysglyleuglutrpleugly151015arg<210>23<211>38<212>prt<213>artificialsequence<220><223>9nt.c3、9nt.c53a、9nt.ns以及9nt共有的hfr3<400>23asptyralaaspservallysglyargilethrileserproaspthr151015serlysasnthrphetyrleuglnleuasnserleuargalagluasp202530thralavaltyrtyrcys35<210>24<211>25<212>prt<213>artificialsequence<220><223>hfr1-9nt.d3<400>24glnvalglnleuglnglnserglyproglyleuvallysproprogln151015thrleuserleuthrcysalaileser2025<210>25<211>17<212>prt<213>artificialsequence<220><223>hfr2-9nt.d3<400>25trpasptrpileargglnserproserargglyleuglutrpleugly151015arg<210>26<211>38<212>prt<213>artificialsequence<220><223>hfr3-9nt.d3<400>26asptyralavalservallysasnargilethrileasnproaspthr151015serlysasnglnpheserleuglnleuasnservalserprogluasp202530thralavaltyrtyrcys35<210>27<211>26<212>prt<213>artificialsequence<220><223>9nt.c3、9nt.c53a、9nt.ns以及9nt共有的lfr1<400>27aspileglnleuthrglnserproserserleuseralaservalgly151015aspargvalthrilethrcysglnglyasp2025<210>28<211>16<212>prt<213>artificialsequence<220><223>9nt.c3、9nt.c53a、9nt.ns以及9nt共有的lfr2<400>28alasertrptyrglnglnlysproglylysalaprolysleuleuile151015<210>29<211>36<212>prt<213>artificialsequence<220><223>9nt.c3-lfr3<400>29aspargprothrglyvalproserargpheserglyserasnsergly151015thraspalathrleuthrileserserleuglnprogluasppheala202530thrtyrtyrcys35<210>30<211>36<212>prt<213>artificialsequence<220><223>9nt.c53a-lfr3<400>30alaargprothrglyvalproserargpheserglyserasnsergly151015thraspalathrleuthrileserserleuglnprogluasppheala202530thrtyrtyrcys35<210>31<211>36<212>prt<213>artificialsequence<220><223>9nt.ns-lfr3<400>31alaargprothrglyvalproserargpheserglyserasnsergly151015asnseralathrleuthrileserserleuglnprogluasppheala202530thrtyrtyrcys35<210>32<211>36<212>prt<213>artificialsequence<220><223>9nt-lfr3<400>32alaargprothrglyvalproserargpheserglyserasnsergly151015asnthralathrleuthrileserserleuglnprogluasppheala202530thrtyrtyrcys35<210>33<211>25<212>prt<213>artificialsequence<220><223>9nt.d3-lfr1<400>33sersergluleuthrglnaspproalavalservalalaleuglygln151015thrvalargilethrcysglnglyasp2025<210>34<211>17<212>prt<213>artificialsequence<220><223>9nt.d3-lfr2<400>34alasertrptyrglnglnlysproglyglnalaprovalleuvaliletyr151015<210>35<211>36<212>prt<213>artificialsequence<220><223>9nt.d3-lfr3<400>35aspargprothrglyileprogluargpheserglyserasnsergly151015asnseralathrleuthrileserargvalglualaglyaspgluala202530asptyrtyrcys35<210>36<211>22<212>dna<213>artificialsequence<220><223>突变引物-for<400>36tgagtcagctcagaggaggcat22<210>37<211>25<212>dna<213>artificialsequence<220><223>突变引物-rev<400>37gttagggataggcttaccttcgaag25当前第1页12
相关技术
网友询问留言
已有1条留言
-
 0访客 来自[未知地区] 2019年04月08日 08:46那个正规医院有啊大概多少钱啊
0访客 来自[未知地区] 2019年04月08日 08:46那个正规医院有啊大概多少钱啊
1