一种酶活提高的双果糖酐水解酶突变体C387A的制作方法
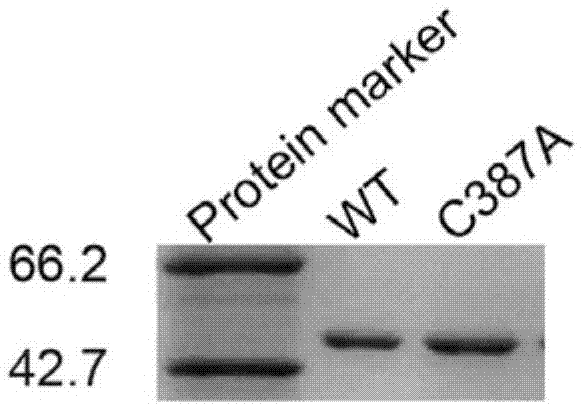
本发明涉及一种酶活提高的双果糖酐水解酶(acdfa-iiiase)突变体c387a,属于基因工程
技术领域:
。
背景技术:
果聚糖(fructan)是一种主要由果糖基连接成的碳水化合物的总称,自然界中普遍存在,其分布就像葡聚糖(gluctan)比如淀粉一样广泛。其中,有一种果聚糖分子,其末端是一分子的葡萄糖,因此这种果聚糖整体不显示还原性。果聚糖广泛存在于自然界中:其不仅存在于大约15%的开花植物中,也大量存在于微生物包括细菌和真菌中。果聚糖可以提高植物的耐旱能力,还可以起到液泡渗透缓冲剂和低温保护剂的作用。这主要是由于植物中的果聚糖主要分布在细胞液泡中,起到了调节渗透压的作用。细菌果聚糖它可以保护细胞和防止失水。由于果糖基连接位置的不同以及物种间聚合度的差异,果聚糖的结构相当复杂。自然界中主要存在两大类:(1)菊糖:主要以β-(2,1)键连接,这一类型的果聚糖主要存在于双子叶植物中。菊糖的生产主要是通过从植物中提取的方式,目前相关技术比较成熟,因此,菊糖的价格便宜。(2)左聚糖(levan)型果聚糖:主要以β-(2,6)键连接,这一类型的果聚糖主要存在于细菌和单子叶植物中;但不像菊糖,levan目前合成研究比较少,市场上没有相关levan纯品,可能与levan合成过程复杂性有关。菊糖的一条代谢途径如下所示,即廉价底物菊糖先通过菊糖果糖转移酶(inulinfructotransferase,iftase)被转化成为附加值比较高的双果糖酐(difructoseanhydrideiii,dfa-iii)。双果糖苷是一种新型的甜味剂和功能性食品添加剂,其相关研究是一个热点。双果糖酐在不被人体系统的消化酶分解,也不被人体肠道吸收,主要是通过肠道中微生物中的双果糖酐水解酶水解成菊二糖(inulobiose),菊二糖进一步被微生物中其它果糖苷酶水解成为果糖而被人体吸收利用。但是目前只有三篇有关双果糖酐水解酶的报道,其中只有一篇报道利用的是纯化出来的双果糖酐水解酶,其它两篇主要利用的是微生物全细胞而并没有分离纯化出双果糖酐水解酶。因此,目前,对于双果糖酐水解酶的研究处于萌芽阶段。在菊糖的这条代谢途径中,菊糖是一种重要的膳食纤维,具有多种益生功能,在医疗上主要是用于肾小球过滤功能检测用试剂;双果糖酐的具有促进钙离子吸收和骨骼生长等多种益生功能;果糖在人体代谢中发挥了重要的作用,是能量的直接来源。但是目前对于菊二糖的生理功能未有相关报道,甚至是对其理化性质也很少有研究。但是鉴于这条代谢途径中其它糖的重要生理功能,菊二糖应该也是一种重要的糖。但是,目前市场上未有菊二糖的纯样品。对其合成的报道也是极少,其合成的研究可能是限制了菊二糖的大量生产和相关性质研究。为了获得更高效的生物催化剂,筛选新的并且可能具有高效率合成菊二糖的双果糖酐水解酶是当前的关键任务。同时,通过定点突变的办法,对来筛选到的双果糖酐水解酶进行分子改造,从而进一步提高其催化活性以期得到更适合于工业应用的双果糖酐水解酶。技术实现要素:本发明的一个目的是提供一种酶活提高的双果糖酐水解酶突变体,氨基酸序列如seqidno.4所示。编码所述突变体c387a的基因的核苷酸序列如seqidno.3所示。本发明还提供了携带所述突变体c387a基因的载体、细胞或质粒。本发明还提供了一种重组大肠杆菌,以大肠杆菌(escherichiacoli)bl21(de3)为表达宿主,以pet-22b(+)为载体,表达权利要求1所述的双果糖酐水解酶(acdfa-iiiase)突变体c387a。本发明的另一个目的是,提供一种获得所述突变体的方法。首先以seqidno.1所示编码acdfa-iiiase的基因的核苷酸序列为模板,以序列如seqidno.5、seqidno.6所示的引物,进行pcr,即得到编码的387位氨基酸由半胱氨酸突变成丙氨酸的c387a突变体基因序列;将上一步得到的重组基因序列,分别连接到pet-28a表达载体中,得到重组质粒pet-22b(+)-c387a,将重组质粒转化到大肠杆菌bl21,挑选阳性单克隆进行发酵培养。本发明通过将突变体重组质粒pet-22b(+)-c387a转化至表达宿主大肠杆菌bl21(de3)中,表达分离纯化后,催化底物双果糖酐为菊二糖等实验表明,突变体c387a与野生酶acdfa-iiiase相比,最适催化条件没有发生改变,即最适反应ph为6.5,最适反应温度为55℃,但酶活提高了85.2%,由原来的101.25umg-1提高到187.51umg-1;催化效率提高了近2倍,由原来的4.61s-1mm-1提高到8.59s-1mm-1。这一发现对于工业化制备菊二糖具有重要的研究价值,为进一步工业应用双果糖酐水解酶提供了有利保障。附图说明图1:野生酶acdfa-iiiase(wt)及其突变体c387a的sds-page;图2:acdfa-iiiase及其突变体c387a催化双果糖酐合成菊二糖hplc检测实验;图3:温度对acdfa-iiiase与其突变体c387a催化反应的影响;图4:ph对acdfa-iiiase与其突变体c387a催化反应的影响。具体实施方式实施例1:acdfa-iiiase的制备和鉴定(1)acdfa-iiiase的制备将来源于微生物arthrobacterchlorophenolicusa6的编码双果糖酐水解酶的基因在大肠杆菌工程菌中进行分子克隆表达。该菌种在genbank中基因组编号是cp001341,而该酶基因的编号为achl_2895,基因全长为1338个核苷酸,见序列表中seqidno:1。该基因所编译的蛋白质编号是acl40859.1,共有445个氨基酸,见序列表中seqidno:2。克隆所用质粒载体为pet-22b(+),构建的重组表达质粒命名为pet-22b(+)-acdfa-iiiase。基因片段的两段分别添加了限制性内切位点ndei和xhoi,c端添加了6个组氨酸标记,用于镍离子亲和色谱分离纯化实验用。克隆宿主胞是e.colidh5α,主要是用于质粒的扩增和抽提。而将质粒pet-22b(+)-acdfa-iiiase转化到e.colibl21(de3)细胞中,主要是用于双果糖酐酶的表达。质粒pet-22b(+)-acdfa-iiiase转化到大肠杆菌bl21(de3)细胞中后,将50μl这些细胞液体涂布在含有100μgml-1的luria-bertani固体平板培养基上,37℃培养16小时后从平板中挑选平板阳性单克隆进行发酵培养;在发酵后,离心获得菌体,重悬后超声破碎,镍离子亲和层析纯化和透析后得到acdfa-iiiase,见附图1,sds-page显示acdfa-iiiase的到完全纯化并已经达到了电泳纯级别。(2)acdfa-iiiase的鉴定将acdfa-iiiase与双果糖酐反应,在1ml的反应体系中,包括终浓度10gl-1底物双果糖酐、终浓度50mm的ph6.5磷酸盐缓冲液和100nomll-1纯酶,在55℃条件下反应10min,沸水浴10min终止反应。反应后的溶液利用高效液相(highperformanceliquidchromatography,hplc)检测其产物。附图2显示在反应24小时后,利用糖柱sugar-paki(4.6mm×250mm,waters,ma,usa)检测出产物峰对应菊二糖出峰时间。对此峰值处的样品进行接样,并进行核磁共振实验,碳13谱见附表1。所产样品的化学位移与报道的参考文献中菊二糖的化学位移一致,目前报道的双果糖酐水解酶所产菊二糖的构象只有呋喃和吡喃中的一种,但是acdfa-iiiase所产菊二糖的还原端果糖分子同时具有呋喃和吡喃这两种构型。实施例2:acdfa-iiiase酶突变体的制备及表达纯化(1)acdfa-iiiase酶的突变体c387a的制备方法①在arthrobacterchlorophenolicusa6的acdfa-iiiase酶模拟结构的基础上确定突变位点;突变位点位于结构的盖子区域,盖子是氨基酸序列在378~402位点的区域。②设计定点突变的引物:以携带acdfa-iiiase酶基因的载体pet-22b(+)-acdfa-iiiase为模板进行定点突变构建突变质粒pet-22b(+)-c387a;突变引物如下所示,下划线为突变点:正向突变引物如seqidno:5所示,其中ggcggaa,下划线为突变碱基;反向突变引物如seqidno:6所示,其中ccgcctt,下划线为突变碱基。③pcr反应:pcr反应体系为:以带有双果糖酐水解酶目的基因的克隆载体pet-22b(+)-acdfa-iiiase为模版。10×pcrbuffer5μldntp(2mmol/l)4μl正向引物(10μm)1μl反向引物(10μm)1μlpet-22b(+)-acdfa-iiiase1μltaqplusdnapolymerase(5u/μl)0.5μlddh2o将体系补足50μlpcr扩增条件为:94℃预变性4min;随后94℃变性1min,56℃退火1min,72℃延伸1min,进行35个循环;最后72℃保温10min。pcr扩增产物经琼脂糖电泳检测,割胶回收纯化。经琼脂糖电泳胶回收纯化后的pcr扩增产物经限制性内切酶ndei和xhoi酶切后连接至载体pet-22b(+),并转化至大肠杆菌dh5α感受态细胞中,在含有50μg/ml氨节青霉素的lb固体培养基培养过夜后,挑单克隆于50μg/ml氨节青霉素的lb液体培养基培养,后提取突变质粒pet-22b(+)-c387a,将突变质粒pet-22b(+)-c387a转化至宿主大肠杆菌bl21(de3)感受态细胞中,突变质粒经测序鉴定为正确的突变。(2)acdfa-iiiase酶突变体的表达纯化将测序验证后的突变质粒pet-22b(+)-c387a转化至大肠杆菌bl21(de3)细胞,挑取阳性转化子在lb培养基中37℃、200rpm摇培过夜,后接入lb培养基37℃培养3~4h至od值为0.6~0.8,降温至30℃,加入iptg终浓度为0.6mm诱导6h。发酵液于4℃、l0000rpm离心20min,取菌体。加入20ml缓冲液(50mmpbs,200mmnacl,调节ph至6.5)充分重悬菌体,然后将离心管置于冰浴中,放入超声波细胞破碎仪中,超声破碎的条件为:工作时间1s,停止时间2s,共计18min。将获得的破碎液进行低温高速离心,4℃、10000rpm离心30min,得粗酶液。用0.45μm微孔滤膜过滤,备用。准备镍离子亲和层析柱,首先,在4℃环境下利用恒流泵,向柱子里泵入去离子水冲洗柱子(约6~12倍柱体积),然后用低盐浓度的缓冲液(500mmol/lnacl,50mmpbs调节ph至6.5)平衡柱子环境。待柱子下端的流出液与泵入柱子的低盐浓度缓冲液ph值一致时(约需5倍柱体积缓冲液),将得到的过膜粗酶液加入到柱子中。先用含有低浓度咪唑的缓冲液(500mmoll-1nacl,50mmoll-1咪唑,50mmpbs调节ph至6.5)冲洗杂蛋白至基线平衡,再用含有高浓度咪唑的洗脱液(500mmoll-1nacl,500mmoll-1咪唑,50mmpbs调节ph至6.5)洗脱。收集吸收峰的洗脱液,并测定其酶活,获得目的蛋白。纯化后acdfa-iiiase的突变体酶c387a达到电泳纯,见附图1。实施例3:acdfa-iiiase与其突变体c387a酶活测定本实施例比较了突变前后酶活变化,酶活测定方法:1ml的反应体系,包括终浓度10gl-1底物双果糖酐、终浓度50mm的ph6.5磷酸盐缓冲液和100nomll-1纯酶,在55℃条件下反应10min,沸水浴10min终止反应。18000×g4℃离心20min后,反应液经过0.22μm滤膜过滤到液相小瓶中,高效液相配备糖柱sugar-paki(4.6mm×250mm,waters,ma,usa)和示差折光显示器用于检测菊二糖。1u酶活定义为在ph6.5,55℃条件下反应,每分钟生成1μmol菊二糖所需要的酶量。酶活比较:野生酶acdfa-iiiase在最适反应条件下的比酶活定义为100%,而突变体相对酶活是基于野生酶活,即用突变体比酶活除以野生酶比酶活来计算。催化效率kcat/km定义:转化数(turnovernumber,kcat)代表代为时间内每个酶分子将底物分子转化成产物的最大值,而米氏常数km是衡量一个酶与底物亲和力大小的动力学参数,这两个参数的比值kcat/km代表着催化效率。催化效率在实际工业生产中是一个重要的参数,代表催化能力的大小,其值越大则表示酶的催化能力越大。结果表明,acdfa-iiiase的酶活为101.25umg-1,其突变体c387a的酶活为187.51umg-1,突变体酶活相较于原始酶活提高了85.2%。催化效率由原来的4.61s-1mm-1提高到8.59s-1mm-1,提高了近2倍。实施例4:acdfa-iiiase与其突变体c387a的最适催化条件(1)温度对acdfa-iiiase与其突变体c387a催化反应的影响在1ml的反应体系中,包括终浓度10gl-1底物双果糖酐、终浓度50mm的ph6.5磷酸盐缓冲液和100nmoll-1纯酶,分别在30℃、40℃、45℃、50℃、55℃、60℃、65℃、70℃、80℃条件下反应10min,沸水浴10min终止反应。18000×g4℃离心20min后,反应液经过0.22μm滤膜过滤到液相小瓶中,高效液相配备糖柱sugar-paki(4.6mm×250mm,waters,ma,usa)和示差折光显示器用于检测菊二糖。1u酶活定义为在ph6.5,55℃条件下反应,每分钟生成1μmol菊二糖所需要的酶量。结果如图4所示,对于acdfa-iiiase与其突变体c387a,均是在温度为55℃时,具有最好的催化活性。(2)ph对acdfa-iiiase与其突变体c387a催化反应的影响在1ml的反应体系中,包括终浓度10gl-1底物双果糖酐、终浓度50mm的磷酸盐缓冲液和100nmoll-1纯酶,在55℃温度条件下,分别设置ph为4.0、4.5、5.0、5.5、6.0、6.5、7.0、7.5、8.0,反应10min,沸水浴10min终止反应。18000×g4℃离心20min后,反应液经过0.22μm滤膜过滤到液相小瓶中,高效液相配备糖柱sugar-paki(4.6mm×250mm,waters,ma,usa)和示差折光显示器用于检测菊二糖。1u酶活定义为在ph6.5,55℃条件下反应,每分钟生成1μmol菊二糖所需要的酶量。结果如图3所示,对于acdfa-iiiase与其突变体c387a,均是在ph为6.5时,具有最好的催化活性。实施例5:acdfa-iiiase与其突变体c387a所产菊二糖的构型特征将acdfa-iiiase突变体c387a分别与双果糖酐反应,反应后的溶液利用高效液相检测其产物。附图2显示在反应24小时后,利用糖柱sugar-paki(4.6mm×250mm,waters,ma,usa)检测出产物峰对应菊二糖出峰时间。对此峰值处的样品进行接样,并进行核磁共振实验,碳13谱见附表1。所产样品的化学位移均与报道的参考文献中菊二糖的化学位移一致,目前报道的双果糖酐水解酶所产菊二糖的构象只有呋喃和吡喃中的一种,但是acdfa-iiiase与其突变体c387a所产菊二糖的还原端果糖分子同时具有呋喃和吡喃这两种构型,这是acdfa-iiiase与其突变体c387a特殊之处。表1acdfa-iiiase与其突变体合成的菊二糖的核磁共振碳13谱化学位移(ppm)a参考文献:shiomi,n.&onodera,s.the13c-nmrspectraofinulooligosaccharides.agric.biol.chem.54,215-216(1990).虽然本发明已以较佳实施例公开如上,但其并非用以限定本发明,任何熟悉此技术的人,在不脱离本发明的精神和范围内,都可做各种的改动与修饰,因此本发明的保护范围应该以权利要求书所界定的为准。sequencelisting<110>江南大学<120>一种酶活提高的双果糖酐水解酶突变体c387a<160>6<170>patentinversion3.3<210>1<211>1338<212>dna<213>arthrobacterchlorophenolicusa6<400>1atgccaagcaacaaccgctacgacgtcacggaatggcccgcgggcaacccggccaaggac60atcggcgaggtcatcaacagcatcatcgccgacatcaaggccaggcagggcgcagcggac120gtggacgacggcgggaagcccggcgccgtgatctacctcccgccgggggactaccacctc180cggacgcaggtgctgatcgacatcagtttcctgcggatcgaaggctcggggcacggtttc240acgtcctccagcatccgcttcaacgtcccggaggaggaatggccggacctgcacgaactg300tggcccggcgggagccgcgtgatcgttgaccttcccgccggcggcgccggggattccgcc360gcgggcgcggcgttcctcgtggcgcgcgagggcagcccccggatcagctcggtggagttt420tccaacttctgcatcgacgggctgcacttcacggccgacggttccggcaggcacccggag480aacacctacgccaacgggaagaccggcatccacgtagccagcgccaacgattccttccgc540gtcacggacatgggtttcgtctacctggagaatgcattgaccatccacaaagcggatgcg600ctgtccatccaccacaacttcatcgccgagtgcggcagctgcatcgagctgcgcggctgg660gggcaggcctcaaagatcactgacaaccttgtgggggcgggcccccgcgggcactccatc720tatgccgaaaaccacggcggactcctggtcaccgcaaacaacgttttcccgcgcggcgcc780agcagcgtccacttcaagggggtcacgcggtccagcgtcaccaacaaccggctgcacgcg840ttctaccccgggatggtgcggctggaggagaacagctcggagaatctcgtggccaccaac900cacttcctccgcgaccacgagccctggacgccgttcttcggcgtggacaacgggctggat960gacctcaccgggctgctgagcatcagcggcaacaacaattcggtcatcgggaaccacttc1020tccgaggtggtggacgcgaacgaaatccggcctgagggtgccacgccggtcatcatccgg1080ctgacggcaggcaccggaaacttcgtctccaccaaccatgtggtggcaatggatgtggac1140gctgcgtcgagtgactcctgcttcgaagcccaggtggacgcactgctggcaacggaggcg1200gcggatctcgccgtgacggctgtcctggtggacccagggtcagcccgcaacaccatcctg1260gactccggaagcgacacccaggtggttgccgacagggcagtcaacgcgatccgggccaca1320cccacggtaggtttctaa1338<210>2<211>445<212>prt<213>arthrobacterchlorophenolicusa6<400>2metproserasnasnargtyraspvalthrglutrpproalaglyasn151015proalalysaspileglygluvalileasnserileilealaaspile202530lysalaargglnglyalaalaaspvalaspaspglyglylysprogly354045alavaliletyrleuproproglyasptyrhisleuargthrglnval505560leuileaspileserpheleuargilegluglyserglyhisglyphe65707580thrserserserileargpheasnvalproglugluglutrpproasp859095leuhisgluleutrpproglyglyserargvalilevalaspleupro100105110alaglyglyalaglyaspseralaalaglyalaalapheleuvalala115120125arggluglyserproargileserservalglupheserasnphecys130135140ileaspglyleuhisphethralaaspglyserglyarghisproglu145150155160asnthrtyralaasnglylysthrglyilehisvalalaseralaasn165170175aspserpheargvalthraspmetglyphevaltyrleugluasnala180185190leuthrilehislysalaaspalaleuserilehishisasnpheile195200205alaglucysglysercysilegluleuargglytrpglyglnalaser210215220lysilethraspasnleuvalglyalaglyproargglyhisserile225230235240tyralagluasnhisglyglyleuleuvalthralaasnasnvalphe245250255proargglyalaserservalhisphelysglyvalthrargserser260265270valthrasnasnargleuhisalaphetyrproglymetvalargleu275280285glugluasnsersergluasnleuvalalathrasnhispheleuarg290295300asphisgluprotrpthrprophepheglyvalaspasnglyleuasp305310315320aspleuthrglyleuleuserileserglyasnasnasnservalile325330335glyasnhisphesergluvalvalaspalaasngluileargproglu340345350glyalathrprovalileileargleuthralaglythrglyasnphe355360365valserthrasnhisvalvalalametaspvalaspalaalaserser370375380aspsercyspheglualaglnvalaspalaleuleualathrgluala385390395400alaaspleualavalthralavalleuvalaspproglyseralaarg405410415asnthrileleuaspserglyseraspthrglnvalvalalaasparg420425430alavalasnalaileargalathrprothrvalglyphe435440445<210>3<211>1338<212>dna<213>人工序列<400>3atgccaagcaacaaccgctacgacgtcacggaatggcccgcgggcaacccggccaaggac60atcggcgaggtcatcaacagcatcatcgccgacatcaaggccaggcagggcgcagcggac120gtggacgacggcgggaagcccggcgccgtgatctacctcccgccgggggactaccacctc180cggacgcaggtgctgatcgacatcagtttcctgcggatcgaaggctcggggcacggtttc240acgtcctccagcatccgcttcaacgtcccggaggaggaatggccggacctgcacgaactg300tggcccggcgggagccgcgtgatcgttgaccttcccgccggcggcgccggggattccgcc360gcgggcgcggcgttcctcgtggcgcgcgagggcagcccccggatcagctcggtggagttt420tccaacttctgcatcgacgggctgcacttcacggccgacggttccggcaggcacccggag480aacacctacgccaacgggaagaccggcatccacgtagccagcgccaacgattccttccgc540gtcacggacatgggtttcgtctacctggagaatgcattgaccatccacaaagcggatgcg600ctgtccatccaccacaacttcatcgccgagtgcggcagctgcatcgagctgcgcggctgg660gggcaggcctcaaagatcactgacaaccttgtgggggcgggcccccgcgggcactccatc720tatgccgaaaaccacggcggactcctggtcaccgcaaacaacgttttcccgcgcggcgcc780agcagcgtccacttcaagggggtcacgcggtccagcgtcaccaacaaccggctgcacgcg840ttctaccccgggatggtgcggctggaggagaacagctcggagaatctcgtggccaccaac900cacttcctccgcgaccacgagccctggacgccgttcttcggcgtggacaacgggctggat960gacctcaccgggctgctgagcatcagcggcaacaacaattcggtcatcgggaaccacttc1020tccgaggtggtggacgcgaacgaaatccggcctgagggtgccacgccggtcatcatccgg1080ctgacggcaggcaccggaaacttcgtctccaccaaccatgtggtggcaatggatgtggac1140gctgcgtcgagtgactccgccttcgaagcccaggtggacgcactgctggcaacggaggcg1200gcggatctcgccgtgacggctgtcctggtggacccagggtcagcccgcaacaccatcctg1260gactccggaagcgacacccaggtggttgccgacagggcagtcaacgcgatccgggccaca1320cccacggtaggtttctaa1338<210>4<211>445<212>prt<213>人工序列<400>4metproserasnasnargtyraspvalthrglutrpproalaglyasn151015proalalysaspileglygluvalileasnserileilealaaspile202530lysalaargglnglyalaalaaspvalaspaspglyglylysprogly354045alavaliletyrleuproproglyasptyrhisleuargthrglnval505560leuileaspileserpheleuargilegluglyserglyhisglyphe65707580thrserserserileargpheasnvalproglugluglutrpproasp859095leuhisgluleutrpproglyglyserargvalilevalaspleupro100105110alaglyglyalaglyaspseralaalaglyalaalapheleuvalala115120125arggluglyserproargileserservalglupheserasnphecys130135140ileaspglyleuhisphethralaaspglyserglyarghisproglu145150155160asnthrtyralaasnglylysthrglyilehisvalalaseralaasn165170175aspserpheargvalthraspmetglyphevaltyrleugluasnala180185190leuthrilehislysalaaspalaleuserilehishisasnpheile195200205alaglucysglysercysilegluleuargglytrpglyglnalaser210215220lysilethraspasnleuvalglyalaglyproargglyhisserile225230235240tyralagluasnhisglyglyleuleuvalthralaasnasnvalphe245250255proargglyalaserservalhisphelysglyvalthrargserser260265270valthrasnasnargleuhisalaphetyrproglymetvalargleu275280285glugluasnsersergluasnleuvalalathrasnhispheleuarg290295300asphisgluprotrpthrprophepheglyvalaspasnglyleuasp305310315320aspleuthrglyleuleuserileserglyasnasnasnservalile325330335glyasnhisphesergluvalvalaspalaasngluileargproglu340345350glyalathrprovalileileargleuthralaglythrglyasnphe355360365valserthrasnhisvalvalalametaspvalaspalaalaserser370375380aspseralapheglualaglnvalaspalaleuleualathrgluala385390395400alaaspleualavalthralavalleuvalaspproglyseralaarg405410415asnthrileleuaspserglyseraspthrglnvalvalalaasparg420425430alavalasnalaileargalathrprothrvalglyphe435440445<210>5<211>33<212>dna<213>人工序列<400>5cgcagctcactgaggcggaagcttcgggtccac33<210>6<211>33<212>dna<213>人工序列<400>6gcgtcgagtgactccgccttcgaagcccaggtg33当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1