一种调控甘蓝型油菜粒色的转基因表达载体及其构建方法和应用与流程
本发明属于基因工程领域,具体涉及一种调控甘蓝型油菜粒色的转基因表达载体及其构建方法和应用。
背景技术:
油菜是世界五大油料作物之一,在世界油料作物中占有重要地位。种子作为油菜的主要收获器官,而种皮约占种子干重的10-20%,占饼粕干重的20-30%,种皮富含色素与多酚物质,还会严重影响油品质及饼粕饲用价值。因此,改良甘蓝型油菜种子的构成和提高单位面积单位含油量是甘蓝型油菜育种的主要目标之一。而甘蓝型黄籽油菜具有种皮薄,蛋白质和含油量高,饼粕蛋白含量高、纤维素和色素等有毒物质含量低等优点成为近年来油菜育种的重要目标之一。
大量研究表明,甘蓝型油菜种皮色泽的变化主要受原花色素、多酚和木质素及其衍生物等重要次生代谢物质的影响,控制类黄酮合成途径相关基因也在种皮色泽形成中扮演着重要角色。在芸薹属作物种子中类黄酮物质主要由山萘酚衍生物、槲皮素衍生物、异鼠李素衍生物和表儿茶素糖苷四类组成,类黄酮也是影响种皮色泽性状的重要因子。由于这些物质成分的复杂性,且不同植物种子中类黄酮物质含量还受基因型及环境条件的影响。作为植物中最大的转录因子之一,myb转录因子广泛参与植物各种生物过程的调节,对于植物的生长发育过程中有着非常重要的作用。其中r2r3-myb类转录因子通过广泛参与植物苯丙烷-类黄酮代谢途径中的多种初生和次生代谢过程,在组织器官最终颜色形成中发挥重要作用。但缺乏天然的黄籽基因资源,且粒色遗传机理复杂,性状不稳定,导致其遗传和分子机理研究进展缓慢,是稳定的黄籽油菜育种的主要难点。
因此,针对以上难点,本发明发现并揭示了基因bnmyb47在调控甘蓝型油菜粒色方面的生物学功能,通过lc-ms方法测定对照与转基因株系不同发育期种皮中类黄酮组分,明确其种皮色泽差异的主要原因;通过转录组测序和基因表达模式分析,揭示bnmyb47参与调控甘蓝型油菜种皮差异的关联。本发明所提供bnmyb47转基因载体和构建方法,对油菜黄籽育种和对基因工程进行油菜品质的遗传改良和育种实践具有重要指导意义。
技术实现要素:
有鉴于此,本发明的目的之一是提供一种调控甘蓝型油菜粒色的rnai-bnmyb47转基因表达载体,对油菜黄籽育种和对基因工程进行油菜品质的遗传改良和育种实践具有重要指导意义。
为实现上述目的,本发明的技术方案为:
一种调控甘蓝型油菜粒色的rnai-bnmyb47转基因表达载体,所述rnai-bnmyb47转基因表达载体的rnai片段的核苷酸序列如seqidno:1所示。
其次,本发明的第二个目的在于提供一种调控甘蓝型油菜粒色的rnai-bnmyb47转基因表达载体的构建方法,通过此构建方法能够准确得到所述rnai-bnmyb47转基因表达载体。
为实现上述目的,本发明的技术方案为:
构建所述的rnai-bnmyb47转基因表达载体的方法,包括以下步骤:
1)克隆bnmyb47基因的rnai片段:根据bnmyb47基因基因设计的rnai引物,以甘蓝型油菜黑籽品系中油821的cdna为模板,进行pcr反应,得阳性克隆子;
2)用限制性内切酶对bnmyb47基因与干扰片段相连的阳性克隆子和pfgc5941m质粒进行完全双酶切,电泳检测后分别回收bnmyb47基因的反义片段和开环的pfgc5941m载体骨架,并用t4dna连接酶将回收的片段连接过夜。获得的重组质粒再次转化大肠杆菌感受态细胞,抽取pcr阳性克隆子菌液质粒;
3)对步骤2)得到的质粒进行完全双酶切,电泳检测后分别回收正义片段和开环的反义片段与pfgc5941m重组的载体骨架,按照同样的方法进行连接转化,并对获得的克隆子菌液进行多重pcr检测,阳性克隆子即为rnai-bnmyb47转基因表达载体。
进一步的,步骤1)中所述正向引物含限制性酶切位点bamhi+aatii,所述反向引物含限制性酶切位点xbai+ncoi。
进一步的,步骤1)所述引物的正向引物的核苷酸序列如seqidno:2所示。
进一步的,步骤1)中,所述反向引物的核苷酸序列如seqidno:3所示。
进一步的,,步骤2)中连接过夜的温度为16℃。
本发明的目的之三还在于提供了一种调控甘蓝型油菜粒色的bnmyb47基因的rnai片段的应用。
为实现上述目的,本发明的技术方案为:
所述的bnmyb47基因的rnai片段在调控甘蓝型油菜粒色中的应用。
本发明的目的之四提供了一种调控甘蓝型油菜粒色的bnmyb47基因的rnai片段的另外一种应用。
为实现上述目的,本发明的技术方案为:
所述的bnmyb47基因的rnai片段在油菜黄籽育种和基因工程进行油菜品质的遗传改良和育种中的应用。
本发明的目的之五在于提供了一种调控甘蓝型油菜粒色的rnai-bnmyb47转基因表达载体的应用。
为实现上述目的,本发明的技术方案为:
所述的rnai-bnmyb47转基因表达载体在调控甘蓝型油菜粒色中的应用。
本发明的目的之六在于提供了一种调控甘蓝型油菜粒色的rnai-bnmyb47转基因表达载体的另外一种应用。
为实现上述目的,本发明的技术方案为:
所述的rnai-bnmyb47转基因表达载体在油菜黄籽育种和基因工程进行油菜品质的遗传改良和育种中的应用。
本发明的有益效果在于:本发明提供的调控甘蓝型油菜粒色的rnai-bnmyb47转基因表达载体可以通过调控脂质代谢,影响种皮中色素和多酚类物质的积累及调控类黄酮代谢,来影响种皮色泽。通过本发明提供的构建rnai-bnmyb47转基因表达载体的方法可准确获得所述的表达载体,所述表达载体克服了甘蓝型油菜粒色性状不稳定,难以获得稳定的黄籽油菜育种的困难,为获得稳定遗传的甘蓝型油菜转基因黄籽新种质材料创造了条件,对油菜黄籽育种和对基因工程进行油菜品质的遗传改良和育种实践具有重要指导意义。
附图说明
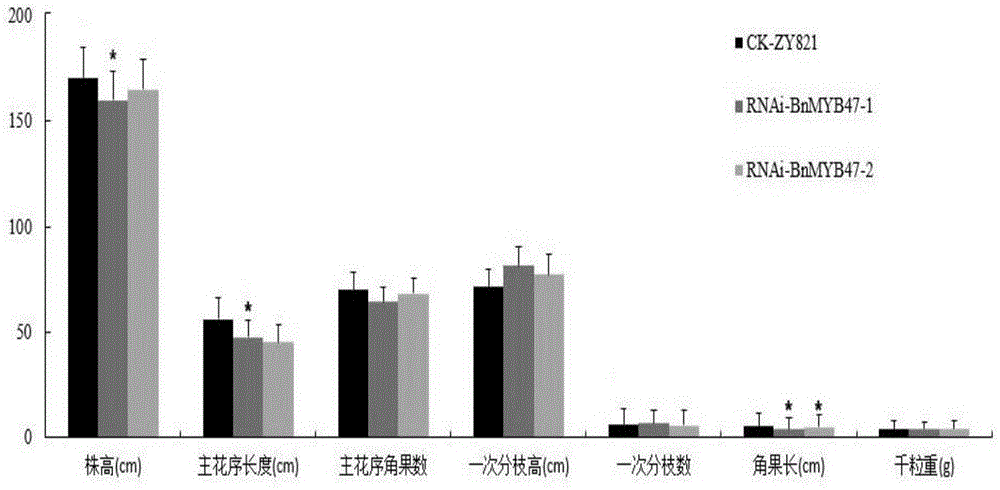
图1对照zy821和rnai-bnmyb47农艺性状分析;
图2对照zy821和rnai-bnmyb47品质性状分析;
图3对照zy821和rnai-bnmyb47种子不同时期发育的表型分析;
图4对照zy821和rnai-bnmyb47成熟种子的表皮结构分析。
具体实施方式
所举实施例是为了更好地对本发明进行说明,但并不是本发明的内容仅局限于所举实施例。所以熟悉本领域的技术人员根据上述发明内容对实施方案进行非本质的改进和调整,仍属于本发明的保护范围。
实施例1bnmyb47基因rnai片段的克隆
根据获得甘蓝型油菜粒色性状bnmyb47基因序列(ganbank的登录号为bnaanng36570d)进行多重比对结果设计rnai引物,正向引物含限制性酶切位点bamhi+aatii,核苷酸序列如seqidno:2所示,反向引物含限制性酶切位点xbai+ncoi,核苷酸序列如seqidno:3所示,以甘蓝型油菜黑籽品系中油821的cdna为模板,用于从甘蓝型油菜中扩增bnmyb47基因干扰片段。
扩增反应体系如表1所示,pcr反应扩增程序为:94℃预变性2min→35个扩增循环(94℃变性45s→58℃退火45s→72℃延伸1min)→72℃延伸10min。反应扩增产物用琼脂糖凝胶电泳检测,将获得的bnmyb47基因的目的片段切胶回收,连接到pmd19-t载体上,转化大肠杆菌jm109感受态细胞,pcr阳性克隆子菌液送样测序,bnmyb47基因干扰片段的核苷酸序列如seqidno:1所示。
表1.bnmyb47基因干扰片段扩增反应体系
实施例2rnai-bnmyb47转基因表达载体的构建和鉴定
用限制性内切酶ncoi+aatii对bnmyb47基因pmd19-t与干扰片段相连的阳性克隆子抽提质粒和pfgc5941m质粒进行完全双酶切,电泳检测后分别回收bnmyb47基因的反义片段和开环的pfgc5941m载体骨架,并用t4dna连接酶将回收的片段于16℃连接过夜。获得的重组质粒再次转化大肠杆菌jm109感受态细胞,抽取pcr阳性克隆子菌液质粒。
用限制性酶bamhi+xbai对bnmyb47基因pmd19-t与干扰片段相连的阳性克隆子抽提质粒和反义片段与pfgc5941重组后的质粒进行完全双酶切,电泳检测后分别回收正义片段和开环的反义片段与pfgc5941m重组的载体骨架,按照同样的方法进行连接转化,并对获得的克隆子菌液进行bar-sf+bar-sr、f35s3nd+rnai-f和rpa3ndr+rnai-r的多重pcr检测,阳性克隆子即为甘蓝型油菜黄籽性状bnmyb47基因的rna干扰载体。
实施例3bnmyb47基因的rnai片段的应用
bnmyb47基因的rnai片段在调控甘蓝型油菜粒色和在油菜黄籽育种和基因工程进行油菜品质的遗传改良和育种中的应用。
实施例4rnai-bnmyb47转基因表达载体的应用
rnai-bnmyb47转基因表达载体在调控甘蓝型油菜粒色和在油菜黄籽育种和基因工程进行油菜品质的遗传改良和育种中的应用。
实施例5rnai-bnmyb47转基因植株对甘蓝型油菜粒色的调控作用
5.1rnai-bnmyb47转基因植株及对照植株农艺性状的考察和转基因家系rnai-bnmyb47及对照种子品质性状的测定
将rnai-bnmyb47转基因的t3代家系和黑籽品系zy821材料同期播种,育苗移栽,栽培管理办法与大田生产相同。成熟后各株系各取10株按照油菜考种方法进行考种。分别记录各株株高(cm)、主花序长度(cm)、主花序角果数(个)、一次分枝高(cm)、一次分枝数(个)、角果长(cm)和千粒重(g)等性状指标,并进行数据分析。
株高是指子叶节至全株最高部分长度,记为“cm”;主花序长度是指主花序顶端至主花序基部的长度,记为“cm”;主花序角果数是指主花序上含有一粒以上饱满或欠饱满种子的角果数,记为“个”;一次分枝高是指第一次有效分枝着生点至子叶节的长度,记为“cm”;一次分枝数是指主茎上具有一个以上有效角果的第一次分枝数,记为“个”;角果长是指从角果底部到角果顶部长度的平均值,记为“cm”;千粒重是指收获完成后经过自然风干的种子(含水量不高于9%)中随机抽取1000粒种子的重量,记为“g”。
近红外反射光谱(nirs)扫描是一种能够快速检测种子品质性状的分析技术,并且对于待测种子无损伤。通过使用nirsystem6500来测量rnai-bnmyb47转基因的t3代和黑籽品系zy821材料种子含油量、蛋白质含量、高油酸、花生烯酸、黄籽r值、芥酸、硫苷、亚麻酸、亚油酸、油酸和棕榈酸等品质性状。每个样品各取5g完整的油菜种子,装入环杯中,转基因rnai-bnmyb47扫描三次,对照zy821扫描五次(反射光谱400-2500nm,2nm一个间隔),分别记录,然后取平均值。其中所用到的矫正模型由重庆市油菜工程中心开发。最后将由该模型得到的表型数据进行统计学分析。
在农艺性状方面,除角果长度外,rnai-bnmyb47转基因植株在一次分枝数、株高、主花序长度、主花序角果数、千粒重、一次分枝高度等农艺性状上差异不显著(图1,表2,表3);通过近红外模型对转基因株系rnai-bnmyb47进行品质性状的分析发现,rnai-bnmyb47株系种子蛋白质含量、硫苷、花生烯酸、亚麻酸含量等无显著差异,而黄籽度、含油量、油酸和亚油酸含量差异显著(图2,表4,表5)。说明抑制bnmyb47基因的表达除了显著影响甘蓝型油菜的黄籽度外,对其含油量、油酸等品质性状也起到一定的调控作用。
表2.rnai-bnmyb47-1农艺性状方差分析
*:在0.05水平相关
表3.rnai-bnmyb47-2农艺性状方差分析
*:在0.05水平相关
表4.rnai-bnmyb47-1种子品质性状方差分析
**:在0.01水平相关
表5.rnai-bnmyb47-2种子品质性状方差分析
**:在0.01水平相关
5.2转基因家系rnai-bnmyb47及对照种子发育的表型分析
通过olympusmvx10体视镜对不同发育期的转基因家系rnai-bnmyb47和黑籽zy821的种子进行了观察并拍照记录。
通过su3500scanningelectronmicroscope扫描电镜对转基因家系rnai-bnmyb47和黑籽zy821成熟种子的种皮结构进行了分析。
利用体视镜对不同发育时期的rnai-bnmyb47种子和zy821种子进行观察发现:rnai-bnmyb47种子较对照zy821种子在早期色泽相近,中后期rnai-bnmyb47种子着色晚于对照zy821种子,而在后期rnai-bnmyb47种子色泽明显浅于zy821种子,zy821种子表现为较深的棕黑色,rnai-bnmyb47种子表现为浅黄色(图3);同时扫描电镜对成熟的rnai-bnmyb47种子和zy821种子的种皮结构进行了分析,结果发现,在成熟的种子表皮上均有许多类似叶片气孔的孔眼,与对照zy821种子相比,rnai-bnmyb47成熟种子表面孔洞直径变小,细胞紧凑(图4)。结合体视镜和扫描电镜的观察结果可以说明bnmyb47基因可能参与调控甘蓝型油菜种皮色泽的形成过程,且在不同时期的调控程度不一致,其可能是直接参与调控甘蓝型油菜种皮色泽的形成过程,也可能是作为调控因子通过调控色素合成过程的一个或几个基因来实现对甘蓝型油菜种皮色泽的调控。除此之外可以看出成熟的rnai-bnmyb47种子表面孔洞直径变小,细胞紧凑,说明抑制bnmyb47基因的表达能够影响种皮的结构。
5.3转基因家系rnai-bnmyb47及对照种皮类黄酮组分lc-ms分析
5.3.1转基因家系rnai-bnmyb47及对照种皮总类黄酮的提取
采用花后20、30、40和50天的种皮材料进行黄酮类化合物的提取和lc-ms分析,每个株系取5份材料,具体方法参照auger等(2010)进行,操作步骤如下:
(1)将不同发育期的新鲜种皮材料充分冷冻干燥,装入含有不锈钢珠的ep管中,于高速振荡仪上充分振荡研磨;
(2)称取5mg充分磨碎的粉末装入5ml玻璃管中,加入1ml的甲醇:丙酮:水:三氟乙酸(tfa)(40:32:28:0.05)的混合溶剂,充分振荡混匀;
(3)4℃下超声波破碎处理15min,然后在19000×g下离心5min,吸取上清液到干净的2mlep管,所得的样品上清液保存于-80度;
(4)加入1ml的甲醇:丙酮:水:三氟乙酸(tfa)(40:32:28:0.05)的混合溶剂,重悬管底沉淀,4℃下超声波破碎处理15min,然后在摇床上4℃200×rpm摇晃4h进一步抽提;
(5)19000×g下离心5min,吸取上清液,合并两次的上清液,保存于-80℃冰箱,等待lc-ms分析,同时将沉淀保存于-80℃,用于后续原花色素含量测定。
5.3.2转基因家系rnai-bnmyb47及对照种皮类黄酮组分lc-ms分析
lc-ms分析在dionexultimatetm3000uhplc系统(thermofisherscientific,usa)上进行,方法可参考(augeretal.,2010)进行,具体方法如下:
(1)分析前所有样品通过尼龙膜(0.22μm,membrana,ger)过滤,然后每个样品吸取200μl到干净的样品管中;
(2)每个样品进样量为5μl,进入质谱前先在acquityuplcbehc18柱(2.1×150mm,1.7μm;waters,usa)上分离,洗脱液a是0.1%甲酸乙腈。洗脱梯度是:5%a~9%a,5min;9%a~16%a,10min;16%a~50%a,25min;50%a~95%a,15min;95%a,5min,然后接着是洗柱和重新平衡;
(3)质谱分析在thermoscientifictmq-exactivetm质谱仪上进行,该质谱仪配备了hesi源,并且在负离子模式运行。源工作参数如下:喷涂电压,3kv;鞘气流速,35单位;辅气流速,10单位;毛细管温度,350℃;辅气体加热器温度,300℃;氮气作为鞘气和辅助气;
(4)首先通过全ms模式分析鉴定主要的黄酮类化合物,然后再对所选的m/z离子进行ms2分析。数据采集和后续处理分析采用thermoxcalibur3.0.63(thermofisherscientific)。黄烷-3-醇和可溶性pas提取物(黄烷3醇构成的原花青素单体)含量表示成表儿茶素当量,相对于表儿茶素的校准曲线,即每mg干重中同等μg的epicatechin(μgecequiv/mgdw)。其它酚类化合物的含量最终表示成相应可用标样校准曲线的相对含量,这些可用标准品包括对香豆酸(p-coumaricacid)、咖啡酸(caffeicacid)、阿魏酸(ferulicacid)、表儿茶素(epicatechin)、槲皮素(quercetin)、山奈酚(kaempferol)和异鼠李素(isorhamnetin)(购买于purechem-standardco.,ltd,chengdu,china),以及芥子酸(sinapicacid)和松柏醛(coniferylaldehyde)(购买于sigma-aldrich)。
利用lc-ms测定了对照zy821与转基因株系rnai-bnmyb47在不同发育时期种皮中类黄酮组分,结果共检测到27种主要的类黄酮组分,主要包括表儿茶素、山奈酚、异鼠李素和花青素苷及其衍生物等次生代谢产物(表6)。说明bnmyb47可能参与了这些代谢产物的合成。
表6.甘蓝型油菜种子发育过程种皮中主要类黄酮组分的lc-ms鉴定
最后说明的是,以上实施例仅用以说明本发明的技术方案而非限制,尽管参照较佳实施例对本发明进行了详细说明,本领域的普通技术人员应当理解,可以对本发明的技术方案进行修改或者等同替换,而不脱离本发明技术方案的宗旨和范围,其均应涵盖在本发明的权利要求范围当中。
序列表
<110>西南大学
<120>一种调控甘蓝型油菜粒色的转基因表达载体及其构建方法和应用
<160>3
<170>siposequencelisting1.0
<210>1
<211>499
<212>rna
<213>人工序列(artificialsequence)
<400>1
cgacgggatgaagaaaggagagtggacggcggaggaagatcaaaaccttgtcgcttacat60
caatgagcatggggtctctgattggcgttctctccccaaaagagctggtttgcagagatg120
tggaaagagctgcaggttaaggtggcttaattacctaaggcctgggattaaaagaggcaa180
attcactcctcaggaagaagaagagatcatcaaacttcatgctgtcctcggaaacaggtg240
ggcagctattgccaaggagatggataatcgaacagacaacgacatcaagaaccattggaa300
ctcttgtctcaagaaaagactgtcgaggaaaggaattgacccgatgacccacgagcctat360
catcaataacctcaccgtcacgatcactaacgaagaatgtggtagctcttcaataactac420
attttctcctacctcatcaccttccggctcggcttgtctcctgaacaagctcgccacagg480
tatctcatctagacaacat499
<210>2
<211>38
<212>rna
<213>人工序列(artificialsequence)
<400>2
ctggatccgacgtccgacgggatgaagaaaggagagtg38
<210>3
<211>41
<212>rna
<213>人工序列(artificialsequence)
<400>3
agtctagaccatggatgttgtctagatgagatacctgtggc41
- 还没有人留言评论。精彩留言会获得点赞!