重组酮酸还原酶突变体、基因、工程菌及其应用的制作方法
本发明涉及一种重组酮酸还原酶突变体、基因、重组载体、工程菌及其在制备(r)-2羟基-4-苯基丁酸中的应用。
(二)
背景技术:
光学纯α-羟酸是一类在羧酸侧c2位有羟基取代的活性化合物,其种类繁多、性质活泼,是重要的药物和精细化工手性中间体,在医药化工等领域具有重要的应用价值。其中,较为重要的是光学纯芳香族的α-羟酸及其衍生物,(r)-2-羟基-4-苯基丁酸是合成众多血管紧张素转化酶抑制剂(acei)类治疗高血压和充血性心力衰竭药物的关键手性中间体,目前已经上市的普利类药物已达到数十种。
正是由于(r)-2-羟基-4-苯基丁酸在医药行业的重要应用,是得其合成方法一直以来受到了诸多的关注。主要可以分为二类,即化学法和生物法。
化学法分为化学拆分法和化学合成法。化学拆分法主要利用手性拆分试剂将(r,s)-2-羟基-4-苯基丁酸((r,s)-hpba)形成非对映体盐,从而增加原对映异构体物理性质差异,然后再进行结晶分离;该法具有产物的纯度、产率不高(最高50%)、手性拆分试剂昂贵、反应有一定的毒性及环境污染等缺点。同时,化学合成法过程中需要昂贵的手性辅助试剂和手性起始原料,整个的合成过程复杂而冗长,并且反应条件苛刻;反应过程使用的大量有机溶剂无法回收,手性试剂昂贵。化学法的上述缺点严重的阻碍了其在普利类药物的合成中的应用。
生物法中主要有酶法拆分、不对称还原法、腈水解酶法。总体而言,生物法所得产品纯度较高,反应条件温和、环境友好。
酶法拆分,酶法拆分法制备光学纯的2-羟酸通常是利用生物酶选择性的衍生外消旋-羟酸中的一种构型而得到另外一种单一构型的目标构型,主要是利用蛋白酶、脂肪酶。该方法保留了生物酶法的优势,但是最大的缺点是理论收率仅有50%。同时,也有利用脂肪酶选择性拆分外消旋体2-羟酸酯得到单一构型的酯化物,但是该方法目前的水平较低,缺乏高效的催化剂。
腈水解酶法,腈水解酶是一类重要的水解酶,可以催化腈类底物一步转化为相应的光学纯羟酸,具有较好的催化特性,目前关于腈水解酶法制备(r)-2-羟基-4-苯基丁酸的研究较少。此外,由于该方法在催化过程中会产生等量剧毒的氢氰酸(hcn),从而增加了操作的危险性和操作难度。
不对称还原法,性的生物合成和转化是利用酶或微生物催化反应对底物立体选择性地将潜手性化合物转化成单一光学活性产物。该法的优点是理论产率高,操作简单。生物不对称还原在总收率和对映选择性方面比生物拆分更有优势。同时,相对于外消旋化合物,潜手性的2-酮酸底物不易获得、价格昂贵,限制了该方法的应用。
在应用生物法不对称还原的策略制备光学纯2-羟酸时,发现酮酸还原酶的活性严重限制了整体的催化效率,导致了底物装载量和产率相对与工业化的方法较低。因此,寻找具有高活性和底物耐受性的酮酸还原酶用于潜手性2-酮酸的不对称还原,对实现潜手性的2-酮酸的高效不对称还原酶具有重要的意义。
酮酸还原酶是一类重要的氧化还原酶,能够催化潜手性酮酸不对称还原为单一构型的2-羟酸,同时需要nadh(烟酰胺腺嘌呤二核苷酸)或nadph(烟酰胺腺嘌呤二核苷酸磷酸)做辅因子参与还原反应。目前,已经发现的酮酸还原酶特定的底物谱。已经发现的酮酸还原酶对有c3、c4支链的酮酸底物具有很低的催化活力,甚至没有催化活力。本研究从实验室已有的酮酸还原酶中筛选到了一株对c3、c4支链的酮酸底物有较高活力的酮酸还原酶(llkar),该酶属于d-扁桃酸脱氢酶;同时,以2-氧代-4-苯基丁酸作为模式底物,对llkar进行了分子改造,提高催化活力;这对于实现潜手性芳香族2-酮酸底物的高效不对称还原具有重要的应用价值。
(三)
技术实现要素:
本发明目的是提供一种重组酮酸还原酶突变体、编码基因、含有该突变体基因的重组载体、该重组载体转化得到的重组基因工程菌,以及其在制备(r)-2-羟基-4-苯基丁酸中的应用。本发明提供的重组酮酸还原酶突变体具有较高的底物耐受性、较高的催化活力,反应条件温和,催化效率提高,生产成本降低且对环境友好。
本发明采用的技术方案是:
本发明提供一种重组酮酸还原酶突变体,所述重组酮酸还原酶突变体是将seqidno:2所示氨基酸序列第127位、第244位、第245位或第250位进行单位点或多位点突变获得的,所述突变包括定点饱和突变、组合突变或者迭代饱和突变。
进一步,所述酮酸还原酶突变体为下列之一:seqidno.2氨基酸序列127位的亮氨酸突变为异亮氨酸(即突变体llkar-l127i(mut-l127i))、seqidno.2氨基酸序列第244位亮氨酸突变为甘氨酸(llkar-l244g(即mut-l244g))、seqidno.2氨基酸序列第250位的丙氨酸突变为甘氨酸(llkar-a250g(mut-a250g))、seqidno.2氨基酸序列第127位亮氨酸突变为异亮氨酸且第244位亮氨酸突变为甘氨酸(llkar-l127i/l244g(即mut-l127i/l244g))、seqidno.2氨基酸序列127位的亮氨酸突变为异亮氨酸且第250位的丙氨酸突变为甘氨酸(llkar-l127i/a250g(即mut-l127i/a250g))、seqidno.2氨基酸序列第244位亮氨酸突变为甘氨酸且第250位的丙氨酸突变为甘氨酸(llkar-l244g/a250g(即mut-l244g/a250g))、seqidno.2氨基酸序列第244位亮氨酸突变为甘氨酸+第250位的丙氨酸突变为甘氨酸+第245位亮氨酸突变为精氨酸(llkar-l244g/a250g/l245r(即mut-l244g/a250g/l245r))。
由于氨基酸序列的特殊性,任何含有该发明所示氨基酸序列的肽蛋白片段或其突变体,如其保守性突变体、生物活性片段或衍生物,只要该肽蛋白的片段或肽蛋白突变体与前述氨基酸序列同源性在90%以上,均属于该发明保护范围之列。具体所述改变氨基酸序列中氨基酸的缺失、插入或替换;其中,对于突变体的保守性改变,所提还的氨基酸具有与原氨基酸相似的结构或化学性质,如用亮氨酸替换异亮氨酸,突变体也可具有非保守性改变,如用色氨酸替换甘氨酸。
由于核苷酸序列的特殊性,任何本发明所示多核苷酸的突变体,只要其与前述多核苷酸具有90%以上同源性,均属于本发明保护范围之列。所述多核苷酸的突变体是指一种具有一个或多个核苷酸改变的多核苷酸序列。此多核苷酸的变体包括取代变异体、缺失变异体和插入变异体。如本领域所知的,等位变异体是一个多核苷酸的替换形式,他可能是一个多核苷酸的取代、确实或插入,但不会从实质上改变其编码的肽蛋白的功能。
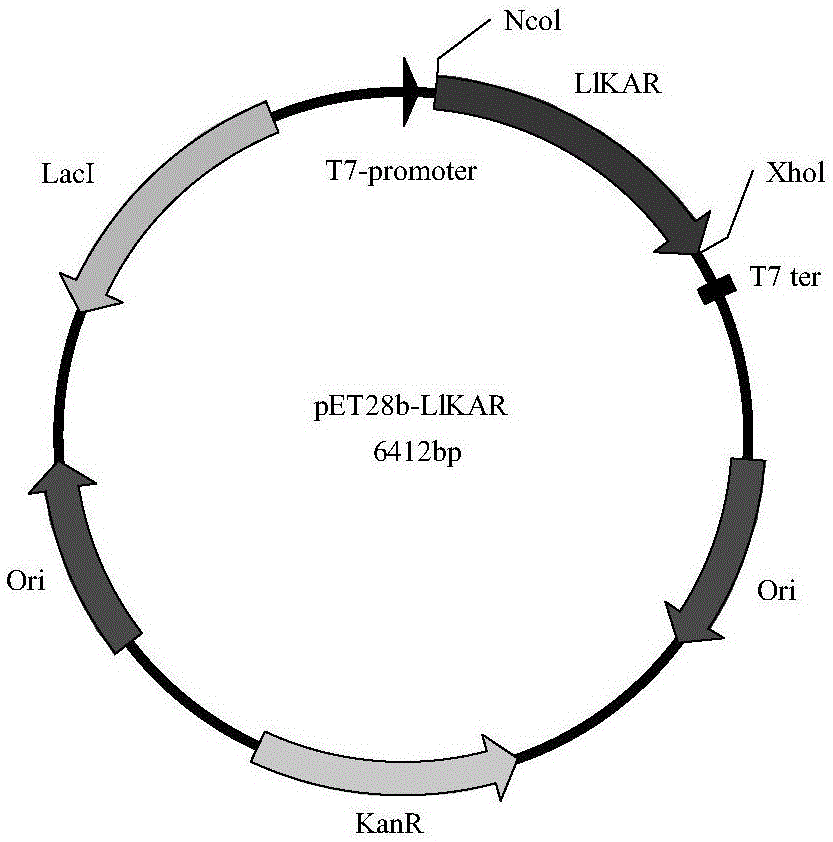
本发明还涉及含所述重组酮酸还原酶突变体编码基因的重组载体。所述重组载体包含与适合指导在宿主细胞中表达的控制序列可操作地连接的多核苷酸。优选该表达载体为pet-28b。
本发明涉及含所述重组酮酸还原酶突变体编码基因和葡萄糖脱氢酶(gdh)的共表达载体。所述共表达载体包含与适合指导在宿主细胞中表达的控制序列可操作地连接的多核苷酸。优选该共表达表达载体为pcdf-duet。所述葡萄糖脱氢酶(gdh)编码基因核苷酸序列为seqidno.3所示。
本发明涉及利用所述重组载体转化得到的重组基因工程菌,具体为:将重组酮酸还原酶突变体编码基因同表达载体pet-28b连接,构建了含有酮酸还原酶突变体编码基因的异源表达重组质粒。将表达重组质粒转化至宿主菌中,获得含有重组质粒的重组基因工程菌。
本发明涉及利用所述共表达载体转化得到的共表达基因工程菌,具体为:以重组酮酸还原酶突变体编码基因和葡萄糖脱氢酶编码基因先后连接到共表达载体pcdf-duet为模板,构建了含有酮酸还原酶突变体编码基因和葡萄糖脱氢酶编码基因的异源共表达质粒。将共表达质粒转化至宿主菌中,获得含有共表达质粒的基因工程菌。
本发明涉及酮酸还原酶突变体编码基因在制备重组酮酸还原酶突变体中的应用,具体为:以含有所述酮酸还原酶编码基因的重组载体pet-28b为模板,构建含有所述酮酸还原酶突变体编码基因的重组载体,将所述重组载体转化至宿主菌(优选大肠杆菌e.colibl21(de3))中,对获得的重组基因工程菌进行诱导培养,培养液分离得到含有重组酮酸还原酶突变体的菌体细胞,破碎后获得的酮酸还原酶粗酶液进行纯化,获得酮酸还原酶突变体纯酶。
本发明还提供一种酮酸还原酶突变体催化2-氧代-4-苯基丁酸制备(r)-2-羟基-4-苯基丁酸的应用,所述的应用方法为:以含重组酮酸还原酶突变体编码基因和葡萄糖脱氢酶(gdh)编码基因的工程菌经发酵培养获得的湿菌体或湿菌体分离纯化获得的纯酶为催化剂,以2-氧代-4-苯基丁酸为底物,以葡萄糖为辅底物,以ph6.0-8.0的缓冲液(优选ph7.0的kh2po4-k2ho4缓冲液)为反应介质构成反应体系,在25-60℃(优选35-40℃)、200-700rpm条件下反应完全后,得到含有芳香(r)-2-羟基-4-苯基丁酸的转化液,分离纯化,获得(r)-2-羟基-4-苯基丁酸。所述催化剂用量以湿菌体干重计为2.5~20dcw/l(优选10dcw/l,以干重计),所述底物终浓度为1~250mm(优选100mm),所述辅助底物与底物的物质的量之比为0.5-3:1(优选2:1)。
所述底物终浓度为250mm时分批加入,所述分批加入是指先加入终浓度100mm底物,待其反应完全后再加入终浓度100mm底物,待第二个100mm底物反应完全后再加入终浓度50mm底物至反应完全。
本发明所述含重组酮酸还原酶突变体编码基因的工程菌经发酵培养获得的湿菌体按如下方法制备:将含有重组酮酸还原酶突变体编码基因的工程菌接种至含有终浓度50μg/ml链霉素的lb液体培养基,37℃,180rpm下培养至菌体od600nm达到0.6-0.8(优选0.6),加入终浓度为0.1mm的iptg,28℃下诱导培养12h后,4℃、8000rpm离心10min,弃去上清液,收集湿菌体并用生理盐水洗涤两次,并将其重悬于kh2po4-k2hpo4缓冲液(100mm,ph7.0)中,即得湿菌体。
所述含重组酮酸还原酶突变体编码基因和葡萄糖脱氢酶(gdh)编码基因的工程菌经发酵培养获得的湿菌体的制备方法:将含有酮酸还原酶突变体编码基因和葡萄糖脱氢酶编码基因的共表达工程菌接种到含有50μg/ml链霉素的lb液体培养基中,于37℃、180rpm条件下振荡培养8~10h,获得种子液;将种子液按2%(体积浓度)的接种量接入到50μg/ml链霉素的lb液体培养基中,于37℃、180rpm条件下振荡培养至od600达到0.4~0.8(优选0.6),加入iptg至终浓度为0.1mm,于28℃、180rpm条件下振荡培养10~12h,离心收集湿菌体并用生理盐水洗涤二次,即得湿菌体、静息细胞。
本发明所述湿菌体分离纯化提取纯酶的方法为:将湿菌体以结合缓冲液(50mm,ph8.0磷酸缓冲液,含有500mmnacl)重悬后,于4℃冰浴中,进行超声破碎(40w,工作1s,停1s),12000rpm离心10min,得到的上清为粗酶液;将上清于结合缓冲液(50mm,ph8.0磷酸钠缓冲液,含500mmnacl)平衡过的ni亲和层析树脂孵育后,再用缓冲液(50mm,ph8.0磷酸钠缓冲液,含500mmnacl,25mm咪唑)冲洗至无杂蛋白,随后以洗脱缓冲液(50mm,ph8.0磷酸钠缓冲液,含500mmnacl,500mm咪唑)洗脱并收集目的蛋白,以透析缓冲液(100mm,ph6.5磷酸缓冲液)透析24h,取截留液即获得所述纯酶。
本发明所述含酮酸还原酶突变体、葡萄糖脱氢酶(gdh)编码基因的共表达工程菌按如下步骤构建:
(1)构建含醛酮还原酶突变体基因的菌株
以酮酸还原酶因序列连入到表达质粒pet-28b中的得到重组质粒,通过pcr方式,获得含有酮酸还原酶突变体的pet-28b重组质粒并将其转化到e.colibl21(de3)中,构建重组菌;
(2)构建共表达菌株
以来源于exiguobacteriumsibiricum的葡萄糖脱氢酶(gdh)基因(核苷酸序列seqidno:3,氨基酸序列seqidno:4)和酮酸还原酶基因连入pcdfduet-1(其中:酮酸还原酶突变体连接于第一个多克隆位点,葡萄糖脱氢酶(gdh)基因连接于第二个多克隆位点)的共表达质粒为模板,通过pcr的方式,获得含有酮酸还原酶突变体的pcdfduet-1并将其转化到e.colibl21(de3)中,构建共表达重组菌。
所述单菌单质粒双酶共表达体系的反应机制为:利用具有(r)-选择性的酮酸还原酶(优选突变体l244g/a250g/l245r)将潜手性的2-酮酸不对称还原为(r)-2-羟酸,其中的gdh可以实现辅酶nadh有效的循环再生,无需添加外源性辅酶。
与现有技术相比,本发明的有益效果主要体现在:
本发明提供一种来源于乳酸明串珠菌(leuconostoclactis)的高效酮酸还原酶突变体(llkar/l127i、llkar/l244g、llkar/l250g、llkar/l127i/l244g、llkar/l127i/l250g、llkar/l244g/l250g,优选llkar/l244g/a250g/l245r),这些突变体相比于野生型酮酸还原酶催化活力大大提高,具体:野生型酮酸还原酶比酶活18.37±0.15u·mg-1,llkar/l127i比酶活20.65±0.17u·mg-1,llkar/l244g比酶活49.02±0.23u·mg-1,llkar/l250g比酶活30.93±0.43u·mg-1,llkar/l127i/l244g比酶活25.86±0.24u·mg-1,llkar/l127i/l250g比酶活28.57±0.19u·mg-1,llkar/l244g/l250g比酶活123.48±0.31u·mg-1,llkar/l244g/a250g/l245r比酶活128.84±0.22u·mg-1。利用酮酸还原酶(优选llkar/l244g/a250g/l245r)和葡萄糖脱氢酶(gdh)建立的单菌单质粒双酶偶联体系,相对于野生型酮酸还原酶与葡萄糖脱氢酶的共表达菌,突变体酮酸还原酶与葡萄糖脱氢酶的共表达菌催化活力大大提高、反应进程耗时明显缩短,具体:各共表达突变体llkar/l127i、llkar/l244g、llkar/l250g、llkar/l127i/l244g、llkar/l127i/l250g、llkar/l244g/l250g,llkar/l244g/a250g/l245r菌相对野生型酮酸还原酶的共表达菌的相对酶活分别达到144.32%、163.14%、135.17%、152.70%、168.23%、193.63%、277.51%;同时,优选突变体llkar/l244g/a250g/l245r与葡萄糖脱氢酶的共表达菌完全转化20mm的2-氧代-4-苯基丁酸需要2小时,相同条件下野生型酮酸还原酶与葡萄糖脱氢酶的共表达菌需要6小时;优选突变体llkar/l244g/a250g/l245r与葡萄糖脱氢酶的共表达菌完全转化40mm的2-氧代-4-苯基丁酸需要4小时,相同条件下野生型酮酸还原酶与葡萄糖脱氢酶的共表达菌需要10小时。以2-氧代-4-苯基丁酸为模式底物时,底物装载量由提升至250mm,ee值均大于99%。最终应用于2-氧代-4-苯基丁酸不对称还原制备光学纯(r)-2-羟基-4-苯基丁酸。
利用酮酸还原酶(优选llkar/l244g/a250g/l245r)和gdh构建的单菌单质粒双酶偶联体系,可用于潜手性芳香族2-酮酸的高效不对称还原,制备具有较高利用价值的光学纯芳香(r)-2-羟酸。该反应具有成本低、绿色环保、工艺简单、催化效率高、无需添加外源性辅酶等优势,工业化前景广阔。
(四)附图说明
图1为酮酸还原酶llkar表达载体示意图。
图2为酮酸还原酶llkar与gdh共表达载体示意图。
图3酮酸还原酶和葡萄糖脱氢酶共表达菌不对称还原2-氧代-4-苯基丁酸的示意图。
图4突变体的初筛原理。
图5饱和突变、组合突变及迭代饱和突变的筛选sds-page图;泳道m为marker;泳道2为空载宿主菌e.colibl21(de3)破碎上清样;泳道3为llkjar纯化样;泳道4为mut-l127i纯化样;泳道5为mut-l244g纯化样;泳道6为mut-a250g纯化样;泳道7为mut-l127i/l244g纯化样;泳道8为mut-l127i/a250g纯化样;泳道9为mut-l244g/a250g纯化样;泳道10为mut-l244g/a250g/l245r纯化样;泳道11为pcdf-duet-llkar-gdh破碎上清样;泳道12为pet28b-gdh破碎上清样。
图6为重酮酸还原酶野生酮酸还原酶共表达菌、mut-l127i表达菌、mut-l244g表达菌、mut-a250g表达菌、mut-l127i/l244g表达菌、mut-l127i/a250g表达菌、mut-l244g/a250g表达菌、mut-l244g/a250g/l245r表达菌相对活力的测定结果。
图7mut-l244g/a250g/l245r共表达菌体催化2-氧代-4-苯基丁酸不对称还原的条件优化,a为ph值对反应的影响,b为最适温度的筛选,c为菌体用量对反应的影响,d为操作温度对反应的影响。
图8为辅酶循环体系中葡萄糖与底物比例优化结果示意图。
图9mut-l244g/a250g/l245r共表达菌体催化2-氧代-4-苯基丁酸不对称还原(一次性投料)
图10mut-l244g/a250g/l245r共表达菌体催化2-氧代-4-苯基丁酸不对称还原(分批补料)。
(五)具体实施方式
下面结合具体实施例对本发明进行进一步描述,但本发明的保护范围并不仅限于此:
实施例1:共表达酮酸还原酶突变体的获得
以含有表达载体pcdfduet-llkar-gdh的共表达菌(e.colibl21(de3)/pcdfduet-llkar-gdh,参见cn201810239718.4)为出发菌株(催化示意图如图3),通过定点饱和突变、组合突变和迭代饱和突变技术,进一步提高酮酸还原酶对底物2-氧代-4-苯基丁酸的催化活力和底物耐受性。
(1)第一轮突变(定点饱和突变)
表1第一轮突变突变引物
注:突变的碱基用下划线标出
本发明以llkar基因(核苷酸序列为seqidno.1所示,氨基酸序列seqidno.2所示)所在的质粒dna模板进行pcr扩增,扩增条件如下所示:95℃5min;95℃30s,合适退火温度30s,72℃7min,重复30循环;72℃继续延伸10min。pcr产物用0.9%的琼脂糖凝胶电泳检测。将pcr产物用dpnⅰ于37℃处理3h,灭活后转化至e.colibl21(de3)受体菌中,涂布于含终浓度50mg/l链霉素抗性的lb固体平板上,37℃培养12h后。
(2)定点饱和突变阳性克隆子的初筛
随机挑取步骤(1)平板上的克隆子于96孔板中,加入1ml含终浓度50μg/ml卡那霉素的lb培养基,37℃,180rpm培养8-10小时,得到种子液,即为一级板。转接该种子液200μl于另一新的每孔加有800μl含终浓度50μg/ml卡那霉素+0.1mmiptg的新鲜lb的96孔板中,37℃、180rpm培养隔夜培养12小时;余下的种子液加入600μl30%甘油后,保存待用。所得的培养液体通过96孔板离心机,1500rpm离心30min后,弃上清得到湿菌体;将该湿菌体用ph7.0的磷酸盐缓冲液(100mm)洗涤一次后,加入500μlph7.0的磷酸盐缓冲液保存,制成菌悬液,将该菌悬液用于初筛,初筛的方法如下所述。
在新96孔板中,每孔含有10μl的200mm葡萄糖溶液(溶于100mm、ph7.0的磷酸盐缓冲液)、50μl的20mm2-氧代-4-苯基丁酸溶液(溶于100mm的磷酸盐缓冲液,ph7.0)、250μl上述菌悬液、最后ph7.0的磷酸盐缓冲液补齐体积至975μl;在35℃,200rpm条件下反应3-4小时(3.5小时优先)后,加入25μl6m的盐酸终止共表达细胞催化的反应,1500rpm离心15min;取75μl反应液上清于新的96孔板中,加入每孔75μl2.5mm的2,4-二硝基苯肼(溶于1m的盐酸),于25℃、200rpm条件下反应30min后,先加入75μl6m的氢氧化钠水溶液,再加入超纯水补齐至1000μl,所得即为显色液。取200μl的显色液于445nm酶标仪中,检测吸光值。吸光值越小,对应一级板中酮酸还原酶的活力越高,反之则反,具体的反应如图4所示。
(3)定点饱和突变阳性克隆子的复筛
通过步骤(2)定点饱和突变体初筛,我们获得一系列正向突变体leu127,leu244和a250。通过比较不同突变体在催化底物时的转化率的高低来确定每个点的最优突变体。转化反应在2ml的ep管中进行,底物(2-氧代-4-苯基丁酸)浓度为5mm,葡萄糖10mm,湿菌体5g/l,35℃,700rpm反应10min,反应液通过hplc分析来确认反应的转化率从而确定最优的突变体;并将确定的正向突变体送样测序,确定具体突变情况,具体的突变情况是l127i,leu244g和a250g。定点饱和突变各阳性突变体相对活力筛选结果如图6所示。
(4)第二轮突变组合突变阳性克隆子的确定
将l127i,leu244g和a250g三点定点饱和突变获得的阳性突变子作为组合突变的模板,从而获得二、三组合突变(引物见表2),并进行转化率的测定。具体的实验:以突变体l127i所在的质粒为模板,以l244g-f和l244g-r为引物,进行pcr(pcr条件和转化都与(1)相同),获得双突变体l127i/l244g;以突变体l127i所在的质粒为模板,以a250g-f和a250g-r为引物,进行pcr(pcr条件和转化都与(1)相同),获得双突变体l127i/a250g;以突变体a250g所在的质粒为模板,以l244g-f和l244g-r为引物,进行pcr(pcr条件和转化都与(1)相同),获得双突变体l244g/a250g;以突变体l244g/a250g所在的质粒为模板,以l127i-f和l127i-r为引物,进行pcr(pcr条件和转化都与(1)相同),获得三突变体l127i/l244g/a250g。转化率测定的体系和反应条件与(3)完全相同。组合突变各阳性突变体相对活力筛选结果如图6所示。
表2第二轮突变(组合突变)引物
注:突变的碱基用下划线标出
(6)第三轮突变(迭代饱和突变)阳性克隆子的确定
取(5)获得活力最高的突变株为模板,以(2)中的序列作为引物,进行迭代饱和突变(引物见表3);按照(2)(3)进行阳性突变子的筛选及复筛。具体为:以双突变体l244g/a250g所在的质粒为模板,以l245-f和l245-r为引物,进行pcr(pcr条件和转化都与(1)相同),获得迭代饱和突变子,按照(2)(3)进行阳性突变子的初筛及复筛;筛选迭代饱和突变各阳性突变体相对活力筛选结果如图6所示。
表3第三轮突变(迭代饱和突变)引物
注:突变的碱基用下划线标出
结果表明,通过定点饱和突变方法、组合突变和迭代饱和突变,获得的优势突变体为llkar-l127i(即mut-l127i,seqidno.2氨基酸序列127位的亮氨酸突变为异亮氨酸)、llkar-l244g(即mut-l244g,seqidno.2氨基酸序列第244位亮氨酸突变为甘氨酸)、llkar-a250g(即mut-a250g,seqidno.2氨基酸序列第250位的丙氨酸突变为甘氨酸)、llkar-l127i/l244g(即mut-l127i/l244g,seqidno.2氨基酸序列第亮氨酸突变为异亮氨酸且第244位亮氨酸突变为甘氨酸)、llkar-l127i/a250g(即mut-l127i/a250g,seqidno.2氨基酸序列127位的亮氨酸突变为异亮氨酸且第250位的丙氨酸突变为甘氨酸)、llkar-l244g/a250g(即mut-l244g/a250g,seqidno.2氨基酸序列第244位亮氨酸突变为甘氨酸且第250位的丙氨酸突变为甘氨酸)、llkar-l244g/a250g/l245r(即mut-l244g/a250g/l245r,seqidno.2氨基酸序列第244位亮氨酸突变为甘氨酸且第250位的丙氨酸突变为甘氨酸,第245位亮氨酸突变为精氨酸)。
实施例2:重组酮酸还原酶突变体湿菌体的制备
将实施例1确定的共表达酮酸还原酶的阳性突变位点重新引入到重组酮酸还原酶基因中(引物见表2和表3),并通过pcr加上组氨酸标签转入大肠杆菌bl21(de3),分别获得重组大肠杆菌,即重组大肠杆菌bl21(de3)/pet28b-llkar/l127i、重组大肠杆菌bl21(de3)/pet28b-llkar/l244g、重组大肠杆菌bl21(de3)/pet28b-llkar/a250g、重组大肠杆菌bl21(de3)/pet28b-llkar/l127i/l244g、重组大肠杆菌bl21(de3)/pet28b-llkar/l127i/a250g、重组大肠杆菌bl21(de3)/pet28b-llkar/l244g/a250g和重组大肠杆菌bl21(de3)/pet28b-llkar/l244g/a250g/l245r。将前述重组共表达大肠杆菌分别接种至含有终浓度50μg/ml卡那霉素的lb液体培养基,37℃,180rpm下培养至菌体od600nm达到0.6-0.8,加入终浓度为0.1mm的iptg,28℃下诱导培养12h后,4℃、8000rpm离心10min,弃去上清液,收集湿菌体并用生理盐水洗涤两次,即得静息细胞。
实施例3:重组酮酸还原酶突变体纯酶的制备
将实施例2各个静息细胞以结合缓冲液(50mm,ph8.0磷酸缓冲液,含有500mmnacl)重悬后,于4℃冰浴中,进行超声破碎(40w,工作1s,停1s),12000rpm离心10min,得到的上清为粗酶液;将上清液于结合缓冲液(50mm,ph8.0磷酸缓冲液,含有300mmnacl)平衡过的ni亲和层析树脂孵育后,再用缓冲液(50mm,ph8.0磷酸钠缓冲液,含500mmnacl,25mm咪唑)冲洗至无杂蛋白,随后以洗脱缓冲液(50mm,ph8.0磷酸钠缓冲液,含500mmnacl,500mm咪唑)洗脱并收集目的蛋白,以透析缓冲液(100mm,ph6.5磷酸缓冲液)透析24h,取截留液即获得重组酮酸还原酶突变体的纯酶,分别获得重组酮酸还原酶纯酶野生酶、mut-l127i、mut-l244g、mut-a250g、mut-l127i/l244g、mut-l127i/a250g、mut-l244g/a250g、mut-l244g/a250g/l245r及出发酶。采用bca试剂盒测定蛋白含量,并冻存于-80摄氏度中备用。酮酸还原酶的sds-page图见图5,同时:泳道1为不含质粒的大肠杆菌破碎上清样;泳道10为共表达菌(pcdf-duet-llkar-gdh)破碎上清样;泳道11为pet28b-gdh破碎上清样。
实施例4:重组酮酸还原酶活力和表观动力学参数的测定
将实施例3挑取得到的重组酮酸还原酶突变体纯酶mut-l127i、mut-l244g、mut-a250g、mut-l127i/l244g、mut-l127i/a250g、mut-l244g/a250g、mut-l127i/l244g/a250g、mut-l244g/a250g/l245r及出发酶llkar用于催化底物2-氧代-4-苯基丁酸。
酶活测定体系组成及催化条件如下:50μl的重组酮酸还原酶纯酶,nadh终浓度10mm(溶于超纯水,100mm),2-氧代-4-苯基丁酸终浓度5mm,100mm、ph6.5磷酸盐缓冲液为反应介质补齐至1ml,于45℃,700rpm条件下预热10min,反应3分钟。加入25μl6m的盐酸终止反应,混合均匀后取样,利用高效液相色谱检测酶活,结果见表4。
表观动力学测定体系组成及催化条件如下:50μl的重组酮酸还原酶纯酶,nadh(终浓度0.1mm、0.15mm、0.2mm、0.3mm和0.4mm),2-氧代-4-苯基丁酸终浓度0.2mm,100mm、ph6.5磷酸盐缓冲液为反应介质补齐至1ml,于45℃,700rpm条件下预热10min,反应5分钟;50μl的重组酮酸还原酶纯酶,nadh(终浓度0.1mm、0.15mm、0.2mm、0.3mm和0.4mm),2-氧代-4-苯基丁酸终浓度0.3mm,100mm、ph6.5磷酸盐缓冲液为反应介质补齐至1ml,于45℃,700rpm条件下预热10min,反应5分钟;50μl的重组酮酸还原酶纯酶,nadh(终浓度0.1mm、0.15mm、0.2mm、0.3mm和0.4mm),2-氧代-4-苯基丁酸终浓度0.4mm,100mm、ph6.5磷酸盐缓冲液为反应介质补齐至1ml,于45℃,700rpm条件下预热10min,反应5分钟;50μl的重组酮酸还原酶纯酶,nadh(终浓度0.1mm、0.15mm、0.2mm、0.3mm和0.4mm),2-氧代-4-苯基丁酸终浓度0.5mm,100mm、ph6.5磷酸盐缓冲液为反应介质补齐至1ml,于45℃,700rpm条件下预热10min,反应5分钟;50μl的重组酮酸还原酶纯酶,nadh(终浓度0.1mm、0.15mm、0.2mm、0.3mm和0.4mm),2-氧代-4-苯基丁酸终浓度0.6mm,100mm、ph6.5磷酸盐缓冲液为反应介质补齐至1ml,于45℃,700rpm条件下预热10min,反应5分钟;加入25μl6m的盐酸终止反应,混合均匀后取样,利用高效液相色谱检测反应速度,结果见表5。
酶活单位(u)定义:在40℃、ph6.5条件下,1min内产生1μmol产物(r)-2-羟基-4-苯基丁酸所需的酶量定义为1u。产物的生成量通过hplc检测确定。比酶活测定结果见表4。
2-氧代-4-苯基丁酸、(r)-2-羟基-4-苯基丁酸及(s)-2-羟基-4-苯基丁酸的液相检测方法(hplc):
高效液相色谱仪器:shimadzulc-20ad系统-spd-20a紫外检测器
检测转化率室采用色谱柱:welchromrc18(4.6×250mm,5μm),流动相:乙腈:高氯酸水溶液=76:24(其中高氯酸水溶液:1.83g/l高氯酸钠,0.56g/l磷酸二氢铵,ph3.5),流速1ml/min,检测波长210nm,进样量5μl。
检测e.e时采用正相手性色谱柱大赛璐ad-h(4.6×250mm,5μm),流动相:正己烷:异丙醇:三氟乙酸=90:10:0.1,流速1ml/min,检测波长210nm。
表4llkar及其突变体酶活测定结果
表5llkar及其突变体表观动力学参数结果
实施例5:共表达酮酸还原酶辅酶再生体系建立及反应条件优化
将实施例1获得的共表达重组酮酸还原酶突变体共表达工程菌(pcdfduet-llkar/l244g/a250g/l245r-gdh)培养获得湿菌体为生物催化剂,以2-氧代-4-苯基丁酸为底物。
具体的湿菌体的获得方法:将前述重组酮酸还原酶突变体共表达菌(pcdfduet-llkar/l244g/a250g/l245r-gdh)接种至含有50μg/ml链霉素的lb液体培养基中,于37℃、180rpm条件下振荡培养8~10h,获得种子液;将种子液按2%(体积浓度)的接种量接入到含有终浓度50μg/ml链霉素的lb液体培养基,37℃,180rpm下培养至菌体od600nm达到0.6-0.8,加入终浓度为0.1mm的iptg,28℃下诱导培养12h后,4℃、8000rpm离心10min,弃去上清液,收集湿菌体并用生理盐水洗涤两次,即得湿菌体。
将获得的湿菌体用于2-氧代-4-苯基丁酸的不对称还原酶的体系优化。反应体系:2-氧代-4-苯基丁酸的终浓度为10mm、湿菌体浓度2.5dcw/l(以干重计)、辅底物葡萄糖浓度20mm、以100mm、ph7.0磷酸盐缓冲液为反应介质补齐至10ml,反应条件35℃、600rpm,反应5分钟。考察菌体浓度2.5dcw/l、5dcw/l、10dcw/l、20dcw/l(以干重计),辅底物葡萄糖浓度5mm、10mm、15mm、20mm、25mm、30mm,反应介质kh2po4-k2po4缓冲液ph6.0、6.5、7.0、7.5、8.0,tris-hcl缓冲液ph8.0、8.5、9.0,温度25℃、30℃、35℃、40℃、45℃、50℃、55℃、60℃及操作温度(30℃、35℃、40℃、45℃、50℃)等因素对反应的影响。反应结束取样用盐酸(6.0m)终止反应,样品采用hplc方法进行分析检测。优化结果如图7所示、葡萄糖与底物优化摩尔比例结果如图8所示。
结果表明共表达菌(pcdfduet-llkar/l244g/a250g/l245r-gdh)催化2-氧代-4-苯基丁酸不对称还原的最适温度40℃、ph7.0、底物与辅底物葡萄糖的最适摩尔比例是1:1;同时,在催化10mm2-氧代-4-苯基丁酸,最适的菌体浓度为10dcw/l(以干重计)。
实施例6:共表达酮酸还原酶llkar在制备(r)-2-羟基-4-苯基丁酸中的应用
以实施例5中获得的共表达突变酮酸还原酶共表达工程菌(pcdfduet-llkar/l244g/a250g/l245r-gdh)和原始的共表达酮酸还原酶(pcdfduet-llkar-gdh)湿菌体作为生物催化剂,以2-氧代-4-苯基丁酸为底物,进行生物转化反应制备(r)-2-羟基-4-苯基丁酸。
催化体系(10ml)组成及催化条件如下:以磷酸缓冲液(100mm、ph7.0)为反应介质,共表达酮酸还原酶湿菌体(以干重计),2-氧代-4-苯基丁酸(20mm,40mm),葡萄糖(与底物的摩尔比例为2:1),40℃、600rpm条件下反应;定时取样,通过实施例4hplc分析测定转化率,结果如表6所示。
表6酮酸还原酶(pcdfduet-llkar-gdh)及突变体
(pcdfduet-llkar/l244g/a250g/l245r-gdh)不对称还原2-氧代-4-苯基丁酸
其中:wild-type代表原始的酮酸还原酶共表达工程菌(pcdfduet-llkar-gdh),bm代表突变体酮酸还原酶共表达工程菌
(pcdfduet-llkar/l244g/a250g/l245r-gdh)。
实施例7:共表达酮酸还原酶突变体和葡萄糖脱氢酶
(pcdfduet-llkar/l244g/a250g/l245r-gdh)在制备(r)-2-羟基-4-苯基丁酸中的应用
(1)反应在10ml的转化体系中进行,体系组成:2-氧代-4-苯基丁酸(终浓度100mm、125mm、150mm)、葡萄糖(与底物的摩尔比例2:1)、通过实施例6获取的共表达菌体(用量以菌种干重计20dcw/l),以kh2po4-k2po4缓冲液(100mm,ph7.0)为反应介质。反应在40℃、200rpm条件下反应,每隔一定时间取样,样品用hcl(6.0m)终止反应,经离心(12000rpm、2min)、稀释、过膜后采用hplc方法进行监测分析。反应过程中用6.0m的naoh自动调控ph在7.0,反应后得到含有(r)-2-羟基-4-苯基丁酸,催化反应结果图9所示。
(2)共表达酮酸还原酶突变体mut-l244g/a250g/l245r催化分批补料2-氧代-4-苯基丁酸的不对称还原
反应在10ml的转化体系中进行,体系组成:2-氧代-4-苯基丁酸(终浓度250mm)、葡萄糖(与底物的摩尔比例2:1)、通过实施例6获取的共表达菌体(用量以菌种干重计,初浓度20dcw/l),以kh2po4-k2po4缓冲液(100mm,ph7.0)为反应介质。具体补料情况:先加入终浓度100mm2-氧代-4-苯基丁酸,待其反应完全后再加入终浓度为100mm2-氧代-4-苯基丁酸,待其反应完全后加入终浓度50mm2-氧代-4-苯基丁酸。反应在40℃、200rpm条件下反应,每隔一定时间取样,样品用hcl(6.0m)终止反应,经离心(12000rpm、2min)、稀释、过膜后采用hplc方法,检测(r)-2-羟基-4-苯基丁酸的浓度和eep值,进行监测分析,整个反应进程图10所示。
序列表
<110>浙江工业大学
<120>重组酮酸还原酶突变体、基因、工程菌及其应用
<160>4
<170>siposequencelisting1.0
<210>1
<211>954
<212>dna
<213>未知(unknown)
<400>1
ttagatttcgaagttttctttttctttagccagtttggtacgaacgatgtcttctttagc60
gtggatcagacgggtaaccagagcgttgaacggggtcgggataccttcctgctgacccag120
acgagcgaagtaaccgttcaggaagtcgatttcggtacgtttaccgttctggatgtcctg180
gtacatagacgggtagtggttaccagcgttttccggtttcagcagacgagacaggtcagc240
gatgataccagcaacgtcaactttgataccctgagaagcaccaacctgtttcatttcgtc300
caggatgttcagagccatgtccatagcgttaccagcggtaccgaattcagcgatgttagc360
gtccagcagaacagacagcgggttcagaacagcgttgataccagctttgtgccagatagc420
ggtcataacgtccggaaccagttcagcgttcagaccagcttcgttcagagcttcaacgat480
cggctgagcgtcaccaccagccagagcctgcagtttgatagaaccagaaccggtggtgtg540
gatttcacccggtttaaccagagaagaggtccacagggtaacaccagcaacgatctgaga600
tttagcaacgtgtttagccataacttcgatgttacccagaccgttagacaggatcagcag660
ctgggtctggtcggtgatgatcggagcgatgtcggtcagcatacggtccagctgcggggt720
tttggtcagcaggatgatcaggtcgaaggtaccggtaacttcctgcggggtcataaccgg780
caggtagtaaactttcgggtcctggtcgtcgtgaacaacggtcagacctttggtgttgat840
agcagcgatgtgagccggccaaccgtcgataccggtaacgtcgtgaccagcagcctgcag900
catgatacccagacgagcacccagagcaccgaaaccagcgatagcgattttcat954
<210>2
<211>317
<212>prt
<213>未知(unknown)
<400>2
metlysilealailealaglypheglyalaleuglyalaargleugly
151015
ilemetleuglnalaalaglyhisaspvalthrglyileaspglytrp
202530
proalahisilealaalaileasnthrlysglyleuthrvalvalhis
354045
aspaspglnaspprolysvaltyrtyrleuprovalmetthrprogln
505560
gluvalthrglythrpheaspleuileileleuleuthrlysthrpro
65707580
glnleuaspargmetleuthraspilealaproileilethraspgln
859095
thrglnleuleuileleuserasnglyleuglyasnilegluvalmet
100105110
alalyshisvalalalysserglnilevalalaglyvalthrleutrp
115120125
thrserserleuvallysproglygluilehisthrthrglysergly
130135140
serilelysleuglnalaleualaglyglyaspalaglnproileval
145150155160
glualaleuasnglualaglyleuasnalagluleuvalproaspval
165170175
metthralailetrphislysalaglyileasnalavalleuasnpro
180185190
leuservalleuleuaspalaasnilealaglupheglythralagly
195200205
asnalametaspmetalaleuasnileleuaspglumetlysglnval
210215220
glyalaserglnglyilelysvalaspvalalaglyileilealaasp
225230235240
leuserargleuleulysprogluasnalaglyasnhistyrproser
245250255
mettyrglnaspileglnasnglylysargthrgluileasppheleu
260265270
asnglytyrphealaargleuglyglnglngluglyileprothrpro
275280285
pheasnalaleuvalthrargleuilehisalalysgluaspileval
290295300
argthrlysleualalysglulysgluasnphegluile
305310315
<210>3
<211>786
<212>dna
<213>未知(unknown)
<400>3
atgtataattctctgaaaggcaaagtcgcgattgttactggtggtagcatgggcattggc60
gaagcgatcatccgtcgctatgcagaagaaggcatgcgcgttgttatcaactatcgtagc120
catccggaggaagccaaaaagatcgccgaagatattaaacaggcaggtggtgaagccctg180
accgtccagggtgacgtttctaaagaggaagacatgatcaacctggtgaaacagactgtt240
gatcacttcggtcagctggacgtctttgtgaacaacgctggcgttgagatgccttctccg300
tcccacgaaatgtccctggaagactggcagaaagtgatcgatgttaatctgacgggtgcg360
ttcctgggcgctcgtgaagctctgaaatacttcgttgaacataacgtgaaaggcaacatt420
atcaatatgtctagcgtccacgaaatcatcccgtggcctactttcgtacattacgctgct480
tctaagggtggcgttaaactgatgacccagactctggctatggaatatgcaccgaaaggt540
atccgcattaacgctatcggtccaggcgcgatcaacactccaattaatgcagaaaaattc600
gaggatccgaaacagcgtgcagacgtggaaagcatgatcccgatgggcaacatcggcaag660
ccagaggagatttccgctgtcgcggcatggctggcttctgacgaagcgtcttacgttacc720
ggcatcaccctgttcgcagatggtggcatgaccctgtacccgagctttcaggctggccgt780
ggttga786
<210>4
<211>261
<212>prt
<213>未知(unknown)
<400>4
mettyrasnserleulysglylysvalalailevalthrglyglyser
151015
metglyileglyglualaileileargargtyralaglugluglymet
202530
argvalvalileasntyrargserhisprogluglualalyslysile
354045
alagluaspilelysglnalaglyglyglualaleuthrvalglngly
505560
aspvalserlysglugluaspmetileasnleuvallysglnthrval
65707580
asphispheglyglnleuaspvalphevalasnasnalaglyvalglu
859095
metproserproserhisglumetserleugluasptrpglnlysval
100105110
ileaspvalasnleuthrglyalapheleuglyalaargglualaleu
115120125
lystyrphevalgluhisasnvallysglyasnileileasnmetser
130135140
servalhisgluileileprotrpprothrphevalhistyralaala
145150155160
serlysglyglyvallysleumetthrglnthrleualametglutyr
165170175
alaprolysglyileargileasnalaileglyproglyalaileasn
180185190
thrproileasnalaglulysphegluaspprolysglnargalaasp
195200205
valglusermetileprometglyasnileglylysproglugluile
210215220
seralavalalaalatrpleualaseraspglualasertyrvalthr
225230235240
glyilethrleuphealaaspglyglymetthrleutyrproserphe
245250255
glnalaglyarggly
260
- 还没有人留言评论。精彩留言会获得点赞!