一种耐高温酸性的果胶酶TsPec及基因与应用的制作方法
本发明属于基因工程技术领域,更具体地,涉及一种耐高温酸性的果胶酶tspec及基因序列、以及重组表达载体、重组表达菌株、该果胶酶tspec的应用和一种水解果胶的方法。
背景技术:
果胶是存在于植物的果实等器官中的一种天然高分子产物。它是植物细胞壁的组成成分,是相邻细胞间的粘结物,使植物细胞黏结在一起。天然果胶类物质通常以原果胶、果胶和果胶酸的形态存在于植物组织和器官中。果胶是一种线形的糖类高聚物,其主链主要由半乳糖醛酸聚糖、半乳聚糖和阿拉伯聚糖半乳糖醛酸残基组成,分子量约为50000~150000。作为一种天然高分子化合物,果胶具有良好的胶凝化和乳化稳定作用。广泛用于食品、医药、日化及纺织行业。另一方面,果胶对果汁、果酒等饮料的加工、饲料的营养利用以及造纸和麻类脱胶等均有显著影响。
果胶酶是指分解果胶质的酶类的总称。通常包括果胶酯酶、聚半乳糖醛酸、果胶裂解酶等。它们能催化果胶解聚或者催化果胶分子中的酯水解。果胶酶可根据其最适应的作用ph值分为酸性果胶酶和碱性果胶酶。
酸性果胶酶在食品工业应用较为广泛,主要涉及果蔬汁的榨取和澄清、天然产物提取以及饲料加工等。果胶酶可以降解果胶,在很多行业均有应用。果胶酶在果蔬汁澄清、果酒酿造、天然产物提取、榨油、饲料、造纸、纺织、环境保护等领域中均有涉及。果胶酶最先应用于果蔬汁加工工艺中。由于果汁一般为酸性,所以应用的主要为酸性果胶酶。加入果胶酶可以提高出汁率、降低果蔬汁的黏度、增加果蔬汁的透光率。另外将果胶酶与淀粉酶混合处理澄清果汁,可减少50%的过滤时间。在果酒酿造领域,葡萄浆用果胶酶处理后可改善酒的澄清度和成色。果胶酶可以一定程度破坏葡萄的细胞壁,降低果汁的粘度,促进色素的溶解,改善葡萄酒的品质。在榨油加工领域,如果在榨油之前用果胶酶进行前处理,使细胞壁通透,可以提高出油率,增加酚类物含量,改善油的浊度、色泽。在饲料领域,果胶酶加入饲料当中可降低抗营养因子,增加饲料利用率,降低食物糜的黏度,减少家禽粪便量。果胶酶是一种绿色饲料添加剂,在饲料的生产应用中日益普遍。
尽管果胶酶在食品、饲料、农产品加工等领域具有广泛的应用前景,但仍然存在需要解决的关键问题:1)酶的比活力低。天然存在的果胶酶活性普遍较低,对果胶的水解效率低。2)果胶酶在工业生产中产量较低,生产成本较高。3)果胶酶的高温稳定性较差,特别是在饲料工业中,高温制粒工艺易使不耐热的果胶酶失去活性。
因此,需要对果胶酶进行改造,从而提高酶对温度的稳定性,提高其表达水平。
技术实现要素:
本发明的目的是提供一种高水平表达的耐高温酸性果胶酶tspec及基因序列、以及重组表达载体、重组表达菌株、该果胶酶tspec的应用和一种水解果胶的方法。本发明的果胶酶基因tspec表达出的果胶酶tspec的最适宜温度为75℃,最适ph值为5.0。
为了实现上述目的,本发明提供一种耐高温酸性的果胶酶tspec,所述果胶酶tspec的氨基酸序列含有seqidno:1所示的氨基酸序列,并且,seqidno:1所示的氨基酸序列中77位和81位的半胱氨酸形成二硫键,194位和210位的半胱氨酸形成二硫键,272位和317位的半胱氨酸形成二硫键。根据本发明,所述果胶酶tspec的氨基酸序列含有seqidno:1所示的氨基酸序列即可实现本发明的效果。最优选情况下,所述果胶酶tspec的氨基酸序列为seqidno:1所示的氨基酸序列,并且,seqidno:1所示的氨基酸序列中77位和81位的半胱氨酸形成二硫键,194位和210位的半胱氨酸形成二硫键,272位和317位的半胱氨酸形成二硫键。
根据本发明,所述果胶酶tspec由来自篮状菌(talaromycessp.)果胶酶pec(genbank登录号:xm_002484792.1)通过蛋白质设计的方式引入了三个二硫键,所述果胶酶pec的氨基酸序列如seqidno:2所示。具体的设计包括:将原始果胶酶pec序列中77位亮氨酸和81位的苏氨酸(t81)均突变为半胱氨酸(l77c和t81c),引入一对二硫键;将194位的丙氨酸和210位的缬氨酸均突变为半胱氨酸(a194c和v210c);将272位的苏氨酸和317位的缬氨酸(t272c和v317c),引入一对二硫键。重新设计的氨基酸所述可采用本领域常规的各种方法实现,其具体操作步骤为本领域技术人员公知,在此不再赘述。
本发明的第二方面提供一种耐高温酸性的果胶酶基因tspec,用于编码果胶酶tspec。具体地,所述果胶酶基因tspec的碱基序列与seqidno:3所示的碱基序列具有95%以上的同一性。优选地,根据本发明,所述的果胶酶基因tspec,含有seqidno:3所示的碱基序列即可实现本发明的效果。在最优选情况下,本发明所述的果胶酶tspec的碱基序列为seqidno:3所示的碱基序列。
本发明提供的果胶酶基因tspec,系基于果胶酶氨基酸序列人工设计。采用了在毕赤酵母中高频使用的密码子,从而显著提高了人工设计的果胶酶基因tspec在毕赤酵母细胞中的表达量。本发明根据果胶酶的氨基酸序列,人工设计出一条全新的果胶酶基因序列,并通过人工合成的方法获得所述果胶酶基因片段,其碱基序列如seqidno:3所示,命名为tspec,所述果胶酶基因tspec编码出的果胶酶的氨基酸序列如seqidno:1所示。本发明的果胶酶基因tspec表达出的果胶酶tspec的适宜温度为65-75℃,最适宜温度为75℃,适宜ph值为4.5-5.5,最适ph值为5.0。
本发明的第三方面提供一种重组表达载体,其包括上述果胶酶基因tspec,还可包括其他功能单元。在果胶酶tspec的氨基酸序列和果胶酶基因tspec的碱基序列确定的情况下,本领域技术人员能够选择合适的重组表达载体以及其他功能单元,例如,毕赤酵母表达载体。
本发明对所述重组表达载体的制备方法没有特别限定,可根据本领域常规技术手段确定。
根据本发明一种具体实施方式,所述重组表达载体的制备方法包括以下步骤:将合成的果胶酶基因tspec连接至表达载体中。在本发明的一些实施例中,所述表达载体为毕赤酵母表达载体ppiczαa。当然,在本发明其他实施例中,也可选用其他毕赤酵母表达载体。
根据本发明一种更加具体的实施方式,所述重组表达载体的制备方法包括如下步骤:将合成的果胶酶基因tspec通过限制性内切酶ecori和xbai酶切,获得具有粘性末端的果胶酶基因tspec片段;将毕赤酵母表达载体ppiczαa通过限制性内切酶ecori和xbai酶切,获得具有粘性末端的ppiczαa片段;将果胶酶基因tspec片段和ppiczαa片段通过t4dna连接酶连接,获得ppic-tspec重组表达质粒。
本发明的第四方面提供一种重组表达菌株,其表达产物为上述果胶酶tspec。在果胶酶tspec的氨基酸序列确定的情况下,本领域技术人员能够获得合适的重组表达菌株。
通常来说,所述宿主细胞可以是大肠杆菌、芽孢杆菌、曲霉,也可以是酵母等细胞,或者是其他类型的细胞,例如动物细胞等。优选地,所述重组表达菌株的宿主细胞为毕赤酵母。同时,本发明提供的果胶酶基因tspec经过密码子优化,采用了适合于毕赤酵母进行转录翻译表达出果胶酶的高频密码子,提高了果胶酶基因tspec在毕赤酵母细胞中的表达量。
本发明对所述重组表达菌株的制备方法没有特别限定,可根据本领域常规技术手段确定。
根据本发明一种具体实施方式,所述重组表达菌株的制备方法包括如下步骤:将重组表达载体通过限制性内切酶线性化;将上述线性化片段导入毕赤酵母中,获得重组表达菌株。可选地,通过电转将重组表达载体导入毕赤酵母宿主细胞中,获得重组表达菌株。在本发明的一些实施例中,所述毕赤酵母为巴斯德毕赤酵母x-33,当然,在本发明其他实施例中,也可选用其他毕赤酵母菌株。
根据本发明一种更加具体的实施方式,所述重组表达菌株的制备方法包括如下步骤:将ppic-tspec重组表达质粒通过bglii限制性内切酶线性化;通过电转化法将线性化的ppic-tspec导入毕赤酵母宿主细胞中,获得重组表达菌株。
本发明中,重组表达菌株制备果胶酶的方法可以采用本领域常规的方法。根据本发明一种具体实施方式,生产果胶酶的方法包括:通过发酵培养上述重组表达菌株,从发酵上清液中提纯获得果胶酶。在一些实施方式中,也可以不经过提纯处理,直接将发酵上清液视为果胶酶产物。
根据本发明一种更加具体的实施方式,由上述重组表达菌株制备果胶酶的方法包括如下步骤:挑取重组表达菌株至培养基中培养约24小时,并收获菌体;将菌体转入新鲜的培养基中,每24小时加入约1%的甲醇,诱导果胶酶tspec的表达。诱导表达约96小时后,离心收集上清液,获得含果胶酶tspec的溶液。
本发明的第五方面提供上述果胶酶tspec的应用。
具体地,本发明的第六方面提供一种水解果胶的方法,该方法包括:将上述果胶酶tspec与含果胶物料接触。所述含果胶物料包括各种类型的果胶,也包括含有果胶的其他物料。
本发明提供的果胶酶tspec系由原始果胶酶突变而来,基因tspec依照tspec的氨基酸序列进行人工设计,根据毕赤酵母密码子的偏好性,选用毕赤酵母中高频使用的密码子,降低了tspec基因mrna二级结构的自由能,从而显著提高了人工合成的果胶酶基因tspec在毕赤酵母细胞中的表达量,显著提高了果胶酶的耐高温特性和表达水平,获得了在毕赤酵母中高水平表达的、高酶活力、耐高温的果胶酶及基因。
本发明的其它特征和优点将在随后具体实施方式部分予以详细说明。
附图说明
通过结合附图对本发明示例性实施方式进行更详细的描述,本发明的上述以及其它目的、特征和优势将变得更加明显。
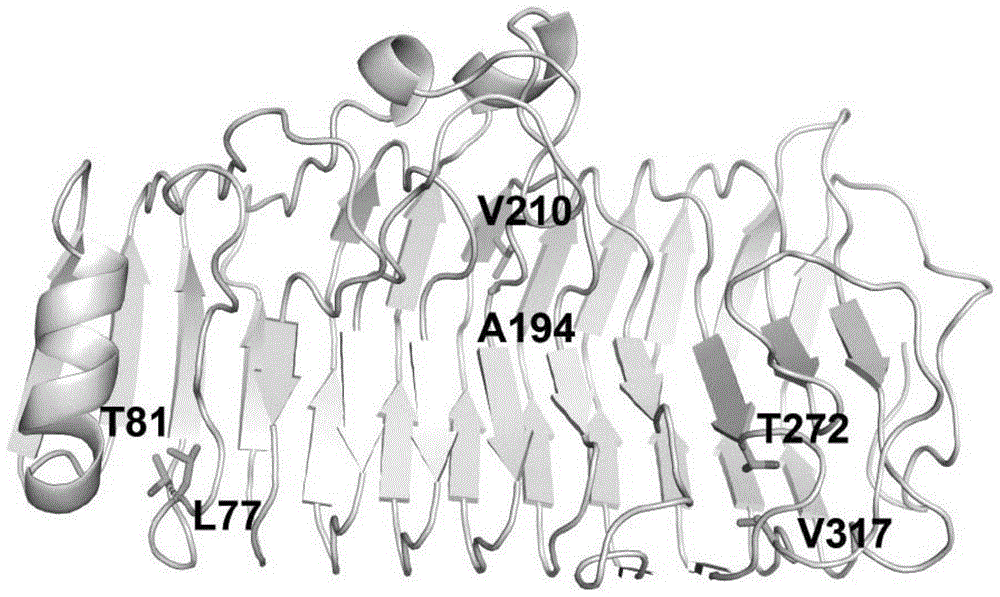
图1a和图1b为本发明实施例1中原始果胶酶和重新设计后引入三对二硫键的结构图。
图2a-图2b为本发明实施例2中将果胶酶克隆入重组表达载体ppiczαa的酶切检验结果。
图3示出了本发明实施例3中产果胶酶菌株在摇瓶条件下表达的结果。
图4a和4b为果胶酶pec和本发明实施例3重新设计的果胶酶tspec的温度曲线。
图5a和5b为果胶酶pec和本发明实施例3重新设计的果胶酶tspec的ph曲线。
图6a和6b为原始果胶酶基因pec和优化后的果胶酶基因tspec的二级结构图。
图7a和7b为本发明实施例6中果胶酶在发酵罐中表达的蛋白质和酶活变化情况。
图8为实施例7中果胶酶tspec水解果胶后产物的薄层层析检测结果。
具体实施方式
下面将更详细地描述本发明的优选实施方式。虽然以下描述了本发明的优选实施方式,然而应该理解,可以以各种形式实现本发明而不应被这里阐述的实施方式所限制。实施例中未注明具体条件者,按照常规条件或制造商建议的条件进行。所用试剂或仪器未注明生产厂商者,均为可以通过市售购买获得的常规产品。
实施例1
本实施例用于说明向果胶酶pec中引入三对二硫键的基因设计过程。具体实施过程如下:
根据果胶酶pec的三维结构,所述果胶酶tspec由果胶酶pec通过蛋白质设计的方式引入了三个二硫键,所述果胶酶pec的氨基酸序列如seqidno:2所示。具体的设计包括:将原始果胶酶pec序列中77位亮氨酸和81位的苏氨酸均突变为半胱氨酸(l77c和t81c),引入一对二硫键;将194位的丙氨酸和210位的缬氨酸均突变为半胱氨酸(a194c和v210c),引入第二对二硫键;将272位的苏氨酸和317位的缬氨酸均突变为半胱氨酸(t272c和v317c),引入第三对二硫键。果胶酶tspec氨基酸序列所对应的基因序列如seqidno:1所示。该基因采用全基因合成的方法合成。
图1a所示,列出了果胶酶pec中用于改造的77位亮氨酸(l77c),81位的苏氨酸(t81),194位的丙氨酸(a194),210位的缬氨酸(v210),272位的苏氨酸(t272)和317位的缬氨酸(v317);图1b所示为新引入的三对二硫键的具体位置。
实施例2
本实施例用于说明果胶酶基因tspec重组表达菌株的构建。
(1)在果胶酶基因tspec两端加上酶切位点ecori和酶切位点noti,经ecori和noti酶切;同样载体ppiczαa经ecorⅰ和noti酶切。其中,酶切体系为:3mgdna、10uecori、10unoti、20μl的bufferh加水补齐至300μl体积,在37℃条件下酶切4h。结果如图2a所示(图中:m为dl5000dnamarker,1泳道为酶切后的ppiczαa重组表达载体;2泳道为酶切后的tspec片段;由图可知,ppiczαa和tspec片段的大小分别为3.7kb和1.0kb左右,与实际相符。
(2)将果胶酶基因tspec酶切片段和ppiczαa酶切片段通过t4dna连接酶连接,获得ppic-tspec重组表达载体;其中,连接体系为:150ng的基因片段、50ng的ppiczαa片段、1μl的t4buffer、5u的t4dna连接酶、加水补齐至10μl;在16℃条件下连接8h后,将连接体系转化至大肠杆菌(dh5α)。转化过程为:将体系与dh5α菌株混匀后于冰上放置10min,然后置于42℃处理1min,再加入800μl培养基后,置于37℃培养40min;最后,涂布于筛选平板上筛选阳性克隆,提取质粒,即得到具有果胶酶基因tspec的重组表达载体ppic-tspec。提取重组表达载体ppic-tspec,进行双酶切验证(ecori、noti),结果如图2b所示(图中:m为dl5000dnamarker,1泳道为双酶切后的ppic-tspec重组表达载体,2泳道为单酶切后的ppic-tspec;3泳道为双酶切后的ppic-tspec)。由图可知,说明果胶酶基因tspec成功连接至ppiczαa重组表达载体上。将重组表达载体ppic-tspec转化入巴斯德毕赤酵母x-33中,得到含果胶酶基因tspec的重组表达菌株。本发明同步也构建了原始果胶酶基因pec的毕赤酵母重组表达菌株。原始果胶酶基因pec的序列如seqidno:4所示。构建方法和步骤同上。
实施例3
本实施例用于说明果胶酶tspec在摇瓶中的发酵生产。
(1)将实施例2中含tspec基因的毕赤基因工程菌接种到5mlypd培养基中,培养20h,获得种子液;
(2)将上述种子液接种至含30mlbmgy发酵培养基的摇瓶中进行发酵培养,其中,bmgy发酵培养基包括:1%酵母粉、2%蛋白胨、100mm磷酸缓冲液(ph6.0)、1.34%酵母氮碱、0.004%生物素、1%甘油。培养24h后,收集菌体并转移到30mlbmmy培养基中继续培养。其中bmmy培养基包括:1%酵母粉、2%蛋白胨、100mm磷酸缓冲液(ph6.0)、1.34%酵母氮碱、0.004%生物素、0.5%甲醇。每24h补充0.15ml甲醇,至96h时停止发酵。发酵液采用sds-page进行检测。如图3所示为发酵液的检测结果(图中:m为蛋白质标准,1泳道为非转基因ppiczαa;2泳道为ppic-pec表达产物;3泳道为ppic-tspec表达产物。图3说明果胶酶基因pec和重新设计的果胶酶基因tspec表达成功。
实施例4
本实施例用于测试果胶酶tspec最适温度和最适ph。
(1)果胶酶活性采用还原糖法测定。步骤如下:
a、取0.5ml稀释500倍的发酵上清液,加入到0.5ml的果胶溶液(0.5%)和1ml的醋酸钠-醋酸缓冲液(ph值为5.0)的混合液中,于特定的温度下水浴反应10min,随后,用2ml的dns(3,5-二硝基水杨酸)终止反应,并置于沸水浴中5min让还原糖充分显色;冷却至室温后用蒸馏水定容至25ml,混合均匀后测试520nm处的吸光度;
b、根据所测得的吸光度,参照酶活公式计算出果胶酶酶活。酶活(u/ml)计算公式为:u=r×df×1000/150.14/t,其中,df为发酵上清液稀释倍数;r为根据od520吸光度值,得到的产物还原糖的摩尔浓度(μmol/ml);t为酶解反应时间(min)。酶活单位定义为:每分钟果胶形成1μmol还原糖所需的酶量为1个酶活力单位(u)。
(2)原始果胶酶pec和重新设计的果胶酶tspec最适温度的测定体系为:果胶0.25g,用50mmolph5.0的naac-hac溶液配制成0.5%浓度的果胶溶液,加入0.1ml适量稀释后的果胶酶溶液。分别设置40℃、50℃、60℃、70℃、75℃、80℃等温度梯度,将上述反应体系置入所设置的不同温度中,反应10分钟,反应结束后,采用如实施例3中的dns法测定反应产生的还原糖含量并计算剩余的酶活性。根据不同温度下测定的酶活绘制出果胶酶活与反应温度的曲线,获得最适反应温度。结果如图4a所示,原始的果胶酶pec的最适温度为60℃。图4b所示,引入了三个二硫键的果胶酶tspec在75℃时活性最高,因此最适温度为75℃。并且,温度为80℃时,仍保留90%左右的酶活。因此,本发明的果胶酶tspec具有良好的耐高温特性。
(3)原始果胶酶pec和重新设计的果胶酶tspec最适ph的测定体系为:果胶0.25g,分别设置ph3.0、ph4.0、ph5.0、ph6.0、ph7.0的naac-hac溶液配制成0.5%浓度的果胶溶液,加入0.1ml适量稀释后的果胶酶溶液。将上述反应体系于75℃反应10分钟,反应结束后,采用如实施例3中的dns法测定反应产生的还原糖含量并计算剩余的酶活性。根据不同ph下测定的酶活绘制出果胶酶活与ph的曲线,获得最适反应ph。结果如图5a和5b所示,图5a为果胶酶pec的曲线,图5b为果胶酶tspec的曲线,可见,果胶酶pec和tspec的最适ph均为5。两者间的酶活与ph关系曲线变化趋势一致。因此,本发明的果胶酶tspec为酸性果胶酶。
实施例5
本实施例用于说明果胶酶基因密码子的优化获得基因tspec。
本实施例基于重新设计引入新的二硫键后的果胶酶tspec的氨基酸序列,人工重新设计了果胶酶基因的核苷酸序列。具体包括:采用高频率的密码子替代低频率的密码子;降低了基因编码的mrna的二级结构的复杂度和最小自由能(图6)。计算机辅助下设计果胶酶基因tspec序列,通过人工合成的方法获得果胶酶基因tspec片段。
本实施例根据果胶酶的氨基酸序列,人工设计出一条全新的果胶酶基因序列,并通过人工合成的方法获得所述果胶酶基因片段,其碱基序列如seqidno:3所示,命名为tspec,所述果胶酶基因tspec编码出的果胶酶的氨基酸序列如seqidno:1所示。图6a为本发明果胶酶基因pec前180个核苷酸的二级结构,其最小自由能为-65.00kcal/mol。图6b为优化后基因tspec中前180个核苷酸的二级结构,其最小自由能为-59.20kcal/mol。因此,本发明所获得的果胶酶基因tspec的稳定性有明显的优化。
实施例6
本实施例用于说明优化后果胶酶基因tspec在发酵罐中的发酵生产。
(1)将实施例5中含tspec基因的毕赤基因工程菌接种到400mlypd培养基中,培养20h,获得种子液;
(2)将上述种子液接种至含发酵培养基的发酵罐中进行发酵培养,其中,发酵培养基包括:350gkh2po4、7gcaso4、40g(nh4)2so4、50gmgso4、120gk2so4和560g甘油。
(3)发酵培养可分为营养生长阶段和甲醇诱导表达阶段。在营养生长阶段,发酵参数控制为:培养温度28℃,ph值为5.5,转速为300-500rpm,通气量3.0-5.5l/min。本阶段持续约30h。营养生长结束后,即进入甲醇诱导表达阶段;在甲醇诱导阶段,溶氧维持在20-30%,温度25℃,ph值为5.0-5.5,甲醇流速为2.5-4.0ml/l/h,流加时间为105h。每隔15h取样,并测定发酵液的果胶酶活性。发酵液中蛋白质组份的检测采用sds-page法检测,检测结果如图7a所示。随着发酵时间的延长,发酵液中tspec的含量逐渐增加。由图7b可知,果胶酶tspec生产水平随着甲醇诱导时间的延长,发酵液中果胶酶的活力逐渐增高,在培养时间为96h时,发酵液中果胶酶的活力最高为13,500u/ml发酵液。
实施例7
本实施例用于说明采用果胶酶tspec水解果胶。
(1)果胶酶tspec水解果胶:取1ml实施例6制备的发酵上清液,用蒸馏水稀释100倍,取0.5ml加入到0.5ml浓度为1%的果胶溶液和1ml醋酸钠-醋酸缓冲液(ph为5.0)的混合溶液中,并75℃水浴反应10min。
(2)采用薄层层析法检测果胶水解产物:取上述步骤(1)中的水解产物10μl点于硅胶板上,放入展开剂中展层60min;取出硅胶板,干燥,并均匀喷涂显色剂,放入108℃的烘箱中烘干显色。结果如图8所示(1泳道为未水解的果胶,2泳道为半乳糖,3泳道为果胶水解0.5h,4泳道为果胶水解1.0h,5泳道为果胶水解1.5h,6泳道为果胶水解2.0h)。由图8可知,本发明提供的果胶酶可以将果胶完全水解为聚合度在1-3之间的低聚糖。
综上所述,本发明通过酶分子设计向原始果胶酶中引入三对二硫键,显著提高了其耐高温的特性,并根据毕赤酵母的偏好性对果胶酶基因进行优化设计后,合成出耐高温的酸性果胶酶基因tspec。该基因与表达载体ppiczαa连接后,转入毕赤酵母gs115中表达,从而获得了高效表达的、高产、耐高温酸性的果胶酶生产菌株;其在发酵罐中诱导表达96小时后,上清液中果胶酶的酶活可达13,500u/ml。本发明获得的耐高温果胶酶的最适温度为75℃,且对果胶具有高效的水解能力。
以上已经描述了本发明的各实施例,上述说明是示例性的,并非穷尽性的,并且也不限于所披露的各实施例。在不偏离所说明的各实施例的范围和精神的情况下,对于本技术领域的普通技术人员来说许多修改和变更都是显而易见的。
序列表
<110>武汉轻工大学
中国大洋矿产资源研究开发协会(中国大洋事务管理局)
<120>一种耐高温酸性的果胶酶tspec及基因与应用
<130>bji1900305whpu
<160>4
<170>siposequencelisting1.0
<210>1
<211>375
<212>prt
<213>人工序列(artificialsequence)
<400>1
metargserpheileglyleualaglyleualacysleuserserval
151015
valleualavalproalaproalaprothralaproalalysleuglu
202530
aspargalaalasercysthrpheserglyserasnglyalaalaala
354045
alaserlysserlysthrsercysserthrilevalleuserserval
505560
alavalproserglythrthrleuaspleuthrasncysasnaspgly
65707580
cyslysvalilephegluglythrthrthrpheglytyralaglutrp
859095
serglyproleuvalservalserglyasnaspilethrvalglugly
100105110
alaserglyalavalleuaspglyaspglyserargtrptrpaspgly
115120125
lysglyserasnglyglylysthrlysprolysphephetyralahis
130135140
asnleulysserserserileserasnilelysileleuasnserpro
145150155160
valglnvalpheserileaspglyalaglnthrleuthrleuasphis
165170175
ilethrileaspasnseralaglyaspserglnglyglyhisasnthr
180185190
aspcyspheaspvalglyaspserasnglyvalthrileserasnala
195200205
ilecysglnasnglnaspaspcysleualavalasnserglythrasn
210215220
ileilephethrglyglythrcysserglyglyhisglyleuserile
225230235240
glyservalglyglyargseraspasnthrvalglnthrvalhisile
245250255
gluasnserthrilelysasnserglnasnglyvalargilelyscys
260265270
valtyrglyalathrglyservalserglyvalthrtyrlysaspile
275280285
thrleuserglyilethrlystyrglyilevalilegluglnasptyr
290295300
gluasnglyserprothrglythrprothrasnglycysproilethr
305310315320
glyleuthrileaspglyvalthrglyservalthrserseralathr
325330335
gluvaltyrileleucysglyserglysercysserasntrpthrtrp
340345350
glnglyvalasnilethrglyglylyslyssersersercyslysasn
355360365
valproserglyalasercys
370375
<210>2
<211>375
<212>prt
<213>talaromycessp.
<400>2
metargserpheileglyleualaglyleualacysleuserserval
151015
valleualavalproalaproalaprothralaproalalysleuglu
202530
aspargalaalasercysthrpheserglyserasnglyalaalaala
354045
alaserlysserlysthrsercysserthrilevalleuserserval
505560
alavalproserglythrthrleuaspleuthrasnleuasnaspgly
65707580
thrlysvalilephegluglythrthrthrpheglytyralaglutrp
859095
serglyproleuvalservalserglyasnaspilethrvalglugly
100105110
alaserglyalavalleuaspglyaspglyserargtrptrpaspgly
115120125
lysglyserasnglyglylysthrlysprolysphephetyralahis
130135140
asnleulysserserserileserasnilelysileleuasnserpro
145150155160
valglnvalpheserileaspglyalaglnthrleuthrleuasphis
165170175
ilethrileaspasnseralaglyaspserglnglyglyhisasnthr
180185190
aspalapheaspvalglyaspserasnglyvalthrileserasnala
195200205
ilevalglnasnglnaspaspcysleualavalasnserglythrasn
210215220
ileilephethrglyglythrcysserglyglyhisglyleuserile
225230235240
glyservalglyglyargseraspasnthrvalglnthrvalhisile
245250255
gluasnserthrilelysasnserglnasnglyvalargilelysthr
260265270
valtyrglyalathrglyservalserglyvalthrtyrlysaspile
275280285
thrleuserglyilethrlystyrglyilevalilegluglnasptyr
290295300
gluasnglyserprothrglythrprothrasnglyvalproilethr
305310315320
glyleuthrileaspglyvalthrglyservalthrserseralathr
325330335
gluvaltyrileleucysglyserglysercysserasntrpthrtrp
340345350
glnglyvalasnilethrglyglylyslyssersersercyslysasn
355360365
valproserglyalasercys
370375
<210>3
<211>1128
<212>dna
<213>人工序列(artificialsequence)
<400>3
atgcgttctttcatcggtctggcgggtctggcgtgcctgtcttctgttgttctggcggtt60
ccggcgccggcgccgaccgcgccggcgaaactggaagaccgtgcggcgtcttgcaccttc120
tctggttctaacggtgcggcggcggcgtctaaatctaaaacctcttgctctaccatcgtt180
ctgtcttctgttgcggttccgtctggtaccaccctggacctgaccaacctgaacgacggt240
accaaagttatcttcgaaggtaccaccaccttcggttacgcggaatggtctggtccgctg300
gtttctgtttctggtaacgacatcaccgttgaaggtgcgtctggtgcggttctggacggt360
gacggttctcgttggtgggacggtaaaggttctaacggtggtaaaaccaaaccgaaattc420
ttctacgcgcacaacctgaaatcttcttctatctctaacatcaaaatcctgaactctccg480
gttcaggttttctctatcgacggtgcgcagaccctgaccctggaccacatcaccatcgac540
aactctgcgggtgactctcagggtggtcacaacaccgacgcgttcgacgttggtgactct600
aacggtgttaccatctctaacgcgatcgttcagaaccaggacgactgcctggcggttaac660
tctggtaccaacatcatcttcaccggtggtacctgctctggtggtcacggtctgtctatc720
ggttctgttggtggtcgttctgacaacaccgttcagaccgttcacatcgaaaactctacc780
atcaaaaactctcagaacggtgttcgtatcaaaaccgtttacggtgcgaccggttctgtt840
tctggtgttacctacaaagacatcaccctgtctggtatcaccaaatacggtatcgttatc900
gaacaggactacgaaaacggttctccgaccggtaccccgaccaacggtgttccgatcacc960
ggtctgaccatcgacggtgttaccggttctgttacctcttctgcgaccgaagtttacatc1020
ctgtgcggttctggttcttgctctaactggacctggcagggtgttaacatcaccggtggt1080
aaaaagtcttcttcttgcaaaaacgttccgtctggtgcgtcttgctaa1128
<210>4
<211>1128
<212>dna
<213>talaromycessp.
<400>4
atgcgttccttcattggcctggccgggctggcctgcttgtcatcagtggtgctggccgtc60
ccagcgccagctcctacagcacctgctaagttagaagatcgtgccgcgtcttgcacattc120
tctggctccaacggtgcagcagcagctagcaaatccaagacttcgtgctcaaccattgta180
ttgtcgagtgttgctgtgccgtcgggaacgacacttgacctaactaatcttaacgatgga240
acgaaggtcatcttcgaaggcacaaccactttcggctacgcagagtggtcaggtcccctt300
gtctccgtctcaggaaacgacatcaccgtcgaaggagcctctggagctgtgctcgacggt360
gacggatctcgttggtgggacggcaagggcagcaacggaggcaagacgaagcccaagttc420
ttttatgctcacaacctcaagtcctcgtctatcagcaacatcaagatcctgaactcgcct480
gtgcaggtcttcagcattgatggtgctcagacactgactttggaccatatcacaattgac540
aattcggctggtgactctcaaggtggccataatacagatgcgtttgatgttggtgattcg600
aatggggttactatctcgaatgctattgttcagaatcaggatgattgtttggcagtcaac660
tctggcactaacatcattttcactggcggtacctgctccggtggtcatggtttgtccatt720
ggctctgttggcggccgttctgataacacggtccagaccgttcacattgagaactctact780
atcaagaactctcaaaacggcgtccgcatcaaaaccgtctatggcgccaccggctccgtc840
tccggcgtaacctacaaggacatcacactctccggcatcaccaaatacggcatcgtcatt900
gagcaggactacgagaacggcagccctacaggaacacctactaacggcgtcccaattact960
ggccttaccattgatggtgtaaccggtagcgtcactagcagcgctacagaagtctatatc1020
ctctgcggcagtggtagctgctccaactggacttggcagggcgttaatattaccggtggt1080
aagaagagtagcagttgtaagaatgttcctagtggtgctagctgctaa1128
- 还没有人留言评论。精彩留言会获得点赞!