抗PD-1/抗HER2天然抗体结构的异源二聚双特异性抗体及其制备方法与流程
本发明涉及抗pd-1/抗her2天然抗体结构的异源二聚双特异性抗体及其制备方法,更具体地,涉及一种具有天然igg特征而在重链和轻链之间没有错配的高度稳定的异源二聚抗pd-1/抗her2双特异性抗体及其制备方法。
背景技术:
单克隆抗体是仅作用于单一抗原表位的高度特异性抗体,已被广泛用于许多疾病的治疗,例如癌症、炎性疾症、自身免疫性疾病和感染性疾病。但是,这种治疗分子在单独使用时,该治疗分子不能显示出足够的药效。这可能是由于疾病的复杂性。例如,癌症或炎性疾病通常涉及多种疾病介导的分子通路以及信号通路之间的相叉作用。在这些情况下,靶向单一靶标的分子不能提供最佳的治疗效果。通过同时阻断多个靶标或者阻断同一靶标的多个位点处的分子能够改善治疗效果。由于多特异性分子、例如双特异性分子是单一分子,多特异性分子可以使得进行双靶向治疗可以简化新药的开发过程。与使用多个单特异性分子组合相比,使用多特异性分子对于患者和医疗服务提供者都更方便。
许多不同形式的双特异抗体或双功能分子已在该领域被报道。最初的双特异抗体是通过应用化学方法使用双功能偶联试剂将igg分子、fab’或(fab’)2片段连接而制备的。但是,这种化学偶联的双特异性抗体具有很多的局限性,例如生产的劳动强度、异源偶联物和同源偶联物的纯化、以及去除初始单特异性抗体或其片段的复杂性;以及低收率。
用于产生双特异性抗体的另一种方法是使用杂交-杂交瘤(或四源杂交瘤)技术,其是采用体细胞融合两种分泌不同抗体的杂交瘤细胞系。由于免疫球蛋白重链和轻链的随机配对,仅有抗体的1/10是所需的功能性双特异性抗体,这因而可能使纯化过程复杂化并降低产率。
wo2013/060867公开了一种大规模生产异源二聚双特异性抗体的方法。在该方法中,首先还原两种混合的同源二聚抗体。然后,在这两种同源二聚抗体的ch3区中引入不对称氨基酸突变,从而促使不同抗体的fab臂交换。最后,通过氧化铰链区的链间二硫键来形成稳定的双特异性抗体。
wo2009/089004公开了一种用于制备异源二聚蛋白质的方法。在该方法中,在ch3-ch3界面处的氨基酸突变为带电荷氨基酸,从而通过静电作用促进异源二聚体的形成。然而,该方法不利于同源二聚体的形成。
us5,731,168公开了一种利用“突起-腔”策略来制备异源二聚igg的方法。在该方法中,通过用较大的氨基酸侧链替换来自第一多肽的ch3结构域的界面的小氨基酸侧链来构建“突起”,并且在该多肽的ch3结构域的界面中产生补偿性“腔”。突起和腔之间的相互作用有利于异源二聚igg的形成,并且在同源二聚体的形成中无效。
wo2012/058768描述了一种制备高度特异性稳定的异源二聚igg的方法。该方法组合了阴性和阳性设计以及结构和计算机模型指导的蛋白工程技术,并且允许待设计的igg1ch3结构域中的多个突变的新型组合,从而形成同源二聚体杂质含量低的稳定异源二聚igg。
程序性死亡受体-1(pd-1)近期作为免疫检查点受到关注,其参与t细胞活化的调控,并且调节免疫应答的强度和持续时间。在正常情况下,pd-1可以介导和维持机体组织的自身免疫耐受,并防止在炎性反应过程中免疫系统的过度活化及其自身组织的伤害,因此提供了积极作用。然而,在病理情况下,pd-1参与各种肿瘤和自身免疫疫病的发生和发展(anticanceragentsmedchem.2015;15(3):307-13.hematoloncolstemcellther.2014年3月;7(1):1-17.trendsmolmed.2015年1月;21(1):24-33.immunity.2013年7月25日;39(1):61-73.jclinoncol.2015年6月10日;33(17):1974-82.)。
pd-1属于cd28家族,但不同于cd28家族的其它成员、例如ctla4,ctla4能以二硫键形成共价二聚体,而pd-1以单体形式存在。pd-1的结构主要包括胞外免疫球蛋白可变区样结构域、疏水的跨膜结构域和胞内结构域,并且胞内结构域含有两个独立的磷酸化位点,分别为免疫受体酪氨酸基抑制基序(itim)和免疫受体酪氨酸基转移基序(itsm)。pd-1主要诱导为在活化的t细胞表面上表达,并且也在b细胞、nk细胞、单核细胞和dc细胞中表达。pd-1的配体包括程序性死亡配体1(pd-l1)和程序性死亡配体2(pd-l2),并且该配体属于b7家族,其中pd-l1诱导为在包括t细胞、b细胞、单核细胞、巨噬细胞、dc细胞、内皮细胞和表皮细胞等的多种免疫细胞的表面上表达,而pd-l2仅诱导为在包括巨噬细胞、dc细胞和b细胞等的一些免疫细胞中表达(autoimmunrev,2013,12(11):1091-1100.frontimmunol,2013,4:481.natrevcancer,2012,12(4):252-264.trendsmolmed.2015jan;21(1):24-33.)。
在二十世纪80年代,denisslamon首次在189个原发性乳腺癌病例的30%中发现her2(人表皮生长因子受体2)基因过度扩增,并发现her2与总存活率和复发时间密切相关(salmandj等人,science,235:177-182,1985)。目前的研究表明,大约25百分比(%)至30%的乳腺癌患者中的her2过表达(revillionf等人,eurjcancer,34:791-808,1998),且这些研究与肿瘤的恶性生长程度相关(wrightc等人,cancerres,49:2087-2090,1989)。
曲妥珠单抗是一种人源化单克隆抗体的抗her2胞外结构域(carterp等人,pnas,89(10):4285-4289,1992)。然而,曲妥珠单抗在临床应用中的抗癌效果往往没有临床前实验那么好。因此,曲妥珠单抗通常需要与化疗药等组合使用(slamondj等人,nengljmed,344:783-792,2001)。
设计一种可以募集效应细胞的双功能抗体在提高抗体效能中是有效的。目前为止,最多数量的研究是利用cd3分子的功能。cd3分子通过激活杀伤性t细胞而可以有效地清除目标肿瘤(haasc等人,immunobiology,214:441-453,2009)。micormet开发的重组双特异性t细胞衔接子(bite)具有良好的前景;但是,最大的问题在于血浆半衰期非常短,在人体中半衰期仅为1小时(lofflera等人,blood,95:2098-2103)。这是由bite本身的结构所导致的,其由两个单链抗体片段组成,分子量仅为60千道尔顿(kda),并且缺失了抗体分子中对延长半衰期有重要作用的fc片段。
卡妥索单抗是另一种具有前景的多功能抗体,是一种靶向cd3和epcam的杂合ig分子。卡妥索单抗已被批准用于腹水癌的治疗(jagerm,等人,cancerres,72:24-32,2012)。另一种处于临床二期的多功能抗体为厄妥索单抗,其靶向cd3和her2。该杂合抗体的重链和轻链的一个分支来源于大鼠igg并靶向cd3;而其重链和轻链的另一个分支来源于小鼠igg并靶向her2。厄妥索单抗的问题就是其生产非常困难。这个的原因是因为为了获得表达双功能厄妥索抗体的克隆,首先要获得表达cd3双特异性抗体的二倍体杂交瘤,以及表达her2双特异性抗体的二倍体杂交瘤,然后将两个杂交瘤再次杂交,以获得可以表达抗cd3和抗her2双特异性的四倍体杂交瘤。生产常规的单靶标抗体只需要一个二倍体杂交瘤。相反地,双功能抗体的生产更复杂,而且四倍体杂交瘤的生产更困难,因为其来源于大鼠,可能导致其免疫原性高。
另外,抗cd3抗体的最明显副反应是短时间内增加体内的细胞因子,也称为细胞因子风暴。因此需要开发一种将免疫细胞招募至肿瘤细胞的新双功能抗体。
联合施用需要依次注射两种或更多种抗体或将抗体形成在同一剂型中。然而,一方面,依次注射抗体会降低患者的治疗配合性并增加疼痛。另一方面,由于不同抗体的物化性质存在差异,很难或几乎不可能将不同抗体配制在同一剂型中。
鉴于此,仍然有必要研究一种阻断pd-1和her2信号通路的新型治疗药物。
技术实现要素:
技术问题
本公开涉及一种具有天然igg结构特征而在重链和轻链之间没有错配、并且能够同时防止pd-1和her2的高度稳定的异源二聚双功能抗体以及制备该双功能抗体的方法,该双功能抗体可能倾向于选择性结合同时高表达pd-1和her2的肿瘤细胞,从而发挥高效力和特异性的杀伤效果,而具有较低的毒副作用。
本发明的第一方面涉及一种异源二聚双特异性抗体。该双特异性抗体可以包含:能够与pd-1特异性结合的第一抗原结合位点和能够与her2特异性结合的第二抗原结合位点,其中该双特异性抗体可以包含可通过至少一个二硫键链彼此链接的第一fc链和第二fc链,其中该第一fc链和第二fc链可以各自通过共价键或连接基团分别连接到pd-1抗原结合位点和her2抗原结合位点,或者该第一fc链和第二fc链可以各自通过共价键或连接基团分别连接到her2抗原结合位点和pd-1抗原结合位点,并且pd-1抗原结合位点中的免疫球蛋白轻链可变区的氨基酸序列可以为seqidno.10,pd-1抗原结合位点中的免疫球蛋白重链可变区的氨基酸序列为seqidno.12,并且第一fc链和第二fc链可以在如下位置处包含5个氨基酸的替换:
第一fc链可以包含366位氨基酸和399位氨基酸的氨基酸替换;并且第二fc链可以包含351位氨基酸、407位氨基酸和409位氨基酸的氨基酸替换,
其中各自包含氨基酸替换的第一fc链与第二fc链可以各自倾向于一起形成异源二聚体而不倾向于形成同源二聚体,
其中氨基酸位置可以根据kabateu索引编号系统编号。
在一些实施方式中,第一fc链和第二fc链的氨基酸替换可以如下:
a)l351g、l351y、l351v、l351p、l351d、l351e、l351k或l351w;
b)t366l、t366p、t366w或t366v;
c)d399c、d399n、d399i、d399g、d399r、d399t或d399a;
d)y407l、y407a、y407p、y407f、y407t或y407h;和
e)k409c、k409p、k409s、k409f、k409v、k409q或k409r。
在一些实施方式中,氨基酸替换可以包括:
a)第一fc链的t366l和d399r替换,以及第二fc链的l351e、y407l和k409v替换;
b)第一fc链的t366l和d399c替换,以及第二fc链的l351g、y407l和k409c替换;
c)第一fc链的t366l和d399c替换,以及第二fc链的l351y、y407a和k409p替换;
d)第一fc链的t366p和d399n替换,以及第二fc链的l351v、y407p和k409s替换;
e)第一fc链的t366w及d399g替换,以及第二fc链的l351d、y407p和k409s替换;
f)第一fc链的t366p和d399i替换,以及第二fc链的l351p、y407f和k409f替换;
g)第一fc链的t366v和d399t替换,以及第二fc链的l351k、y407t和k409q替换;以及
h)第一fc链的t366l和d399a替换,以及第二fc链的l351w、y407h和k409r替换。
在一些实施方式中,第一fc链的氨基酸替换为t366l和d399r,并且第二fc链的氨基酸替换为l351e、y407l和k409v。
在一些实施方式中,fc链可以来源于igg。
在一些实施方式中,pd-1抗原结合位点和her2抗原结合位点可以各自为fab片段或scfv片段。
在一些实施方式中,pd-1抗原结合位点和her2抗原结合位点可以各自为fab片段。
在一些实施方式中,选自pd-1抗原结合位点和her2抗原结合位点中的一个可以为fab片段,而另一个可以为scfv片段。
在一些实施方式中,fab片段可以包含不同的第一重链可变区和第二重链可变区,以及不同的第一轻链可变区和第二轻链可变区。
在一些实施方式中,当第一fc链和连接到第一fc链的pd-1抗原结合位点以及第二fc链和连接到第二fc链的her2抗原结合位点;或者第一fc链和连接到第一fc链的her2抗原结合位点以及第二fc链和连接到第二fc链的pd-1抗原结合位点在或不在还原剂的存在下,组成同源二聚体的重量比小于50%。
在一些实施方式中,双特异性抗体的氨基酸序列选自seqidno.2、4、6、8、10、12和14。在一些实施方式中,双特异性抗体的氨基酸序列选自seqidno.2、4、6、8、10、12和14的相应组合。
本公开的第二方面涉及一种分离的多核苷酸,其编码第一方面的异源二聚双特异性抗体。
在一些实施方式中,分离的多核苷酸的序列选自seqidno:1、3、5、7、9、11和13。在一些实施方式中,分离的多核苷酸的序列选自seqidno.1、3、5、7、9、11和13的相应组合。
本公开的第三方面涉及一种重组质粒,其包含第二方面的分离的多核苷酸。
在一些实施方式中,重组表达载体可以为从pcdna修饰的质粒载体x0gc。
本公开的第四方面涉及一种宿主细胞,其包含第二方面的分离的多核苷酸,或第三方面所述的重组表达载体。
在一些实施方式中,宿主细胞可以是人胚肾细胞hek293或从hek293细胞修饰的hek293t、hek293f或hek293e;或仓鼠卵巢细胞cho或从cho细胞修饰的cho-s、cho-dhfr-、cho/dg44或expicho。
本公开的第五方面涉及一种组合物,其包含第一方面的异源二聚双特异性抗体或第二方面的分离的多核苷酸或第三方面的重组表达载体或第四方面的宿主细胞,以及药学上可接受的载体。
本公开的第六方面涉及一种生产如第一方面的异源二聚双特异性抗体的方法,其包括:
1)在宿主细胞中表达第二方面的分离的多核苷酸或第三方面的重组表达载体;
2)还原在宿主细胞中各自表达的蛋白;以及
3)混合还原的蛋白并氧化钙混合物。
在一些实施方式中,宿主细胞可以选自人胚肾细胞hek293或从hek293细胞修饰的hek293t、hek293f或hek293f;以及仓鼠卵巢细胞cho或从cho细胞修饰的cho-s、cho-dhfr-、cho/dg44或expicho。
在一些实施方式中,还原可以包括:1)使用还原剂进行还原反应,所述还原剂包括2-巯基乙胺、二硫苏糖醇、三(2-羧乙基)膦或其化学衍生物,或其组合;以及2)去除还原剂。
在一些实施方式中,氧化可以在空气中进行,并且包括在氧化剂存在下进行的氧化,所述氧化剂选自l-脱氢抗坏血酸或其他化学衍生物。
在一些实施方式中,所述方法还包括分离和纯化。
本公开的第七方面涉及第一方面的异源二聚双特异性抗体、第二方面的分离的多核苷酸、第三方面的重组表达载体、第四方面的宿主细胞、和/或第五方面的组合物在制备用于预防和/或治疗受试者疾病的药物中的用途。
本公开的第八方面涉及第一方面的异源二聚双特异性抗体、第二方面的分离的多核苷酸、第三方面的重组表达载体、第四方面的宿主细胞、和/或第五方面的组合物,其用于预防和/或治疗受试者疾病。
本公开的第九方面涉及一种预防和/或治疗疾病的方法,该方法包括:将第一方面的异源二聚双特异性抗体、第二方面的分离的多核苷酸、第三方面的重组表达载体、第四方面的宿主细胞、和/或第五方面的组合物施用于有需要的受试者。
在一些实施方式中,受试者可以是哺乳动物,优选为人。
在一些实施方式中,疾病选自包括白血病、淋巴瘤、骨髓瘤、脑肿瘤、头颈部鳞状细胞癌、非小细胞肺癌、鼻咽癌、食道癌、胃癌、胰腺癌、胆囊癌、肝癌、结直肠癌、乳腺癌、卵巢癌、宫颈癌、子宫内膜癌、子宫肉瘤、前列腺癌、膀胱癌、肾细胞癌和黑色素瘤的疾病。
本公开涉及一种抗pd-1/抗her2天然抗体结构的异源二聚双特异性抗体,其中该抗pd-1/抗her2结构的异源二聚双特异性抗体是高度稳定的并且具有天然igg特征,而在重链和轻链之间没有错配。该双特异性抗体可以能够同时结合靶分子pd-1和her2并且在治疗复杂疾病方面更有效。
附图说明
图1示出了抗pd-1表达产物的洗脱峰色谱图;
图2示出了抗pd-1/抗her2异源二聚抗体分子的结构;
图3示出了重链和轻链的不完全抗体的结构;
图4a示出了对重链和轻链的抗pd-1不完全抗体进行的尺寸排阻色谱-高效液相色谱(sec-hplc)的分析结果;图4b是对重链和轻链的抗pd-1不完全抗体进行sec-hplc的分析结果;
图5示出了对抗pd-1/抗her2异源二聚抗体分子进行sec-hplc的分析结果;
图6示出了对抗pd-1/抗her2异源二聚抗体分子进行sec-hplc的纯度分析结果;
图7a示出了抗pd-1/抗her2异源二聚抗体对pd-1的亲和力;图7b示出了抗pd-1/抗her2异源二聚抗体对her2的亲和力;
图8示出了pd-1单克隆抗体和her2单克隆抗体的组合不能结合pd-1和her2,并且抗pd-1/抗her2异源二聚抗体具有同时结合两种抗原的活性;
图9示出了抗pd-1/抗her2异源二聚抗体引发sk-br-3和cho/pd-1细胞的相互靠近;
图10a和图10b各自示出了抗pd-1/抗her2异源二聚抗体阻断pd-1/pd-l1结合和pd-1/pd-l2结合,并且较好地保持双价单克隆抗体的阻断活性;
图11示出了抗pd-1/抗her2异源二聚抗体显示出与pd-1单克隆抗体相当的t细胞调控活性,并且显著促进细胞因子ifn-γ的分泌;
图12示出了抗pd-1/抗her2异源二聚抗体显示出与her2单克隆抗体相当的肿瘤细胞杀伤抑制活性;并且
图13示出了抗pd-1/抗her2异源二聚抗体显示出比pd-1单克隆抗体或her2单克隆抗体更强的抗肿瘤效力,且在停止药物施用后依然维持良好的肿瘤调控作用。
具体实施方式
定义:
共价连接是指异源二聚双特异性抗体中两个fc链中的一个fc链及与其结合的抗原结合功能位点之间通过共价键结合,从而使fc链与抗原结合功能位点结合以形成分子。fc链可以包含通过至少一个共价连接(例如二硫键链)连接的第一抗原结合位点和第二抗原结合位点。第一fc链和第二fc链各自通过共价连接(例如亚胺键或酰胺键)连接到抗原结合位点。抗原结合位点是指可以与目标分子例如抗原特异性相互作用的位点。其作用具有高度选择性,并因此识别一种目标分子的序列通常不识别其他分子序列。
代表性的抗原结合位点包括:抗体的可变区、抗体的结构变构可变区、受体的结合结构域、配体结合结构域或酶结合结构域。
至少一个链间二硫键是指第一fc链和第二fc链通过至少一个二硫键连接以形成异源二聚片段。在本公开中,至少一个二硫键的形成可以是在第一fc链和第二fc链之间;或者在同一细胞中合成的第一fc链、第二fc链以及与其结合的抗原结合位点之间。另外,第一fc链和第二fc链;或者第一fc链、第二fc链以及与其结合的抗原结合位点可以各自单独地在不同细胞中合成,并然后通过体外还原氧化而形成。
第一fc链和第二fc链可以通过共价连接而形成结合片段,其中共价连接可以包括二硫键,每条链包含免疫球蛋白(ig)的重链恒定区的至少一部分;并且第一fc链和第二fc链在氨基酸序列上可以是不同的,且包括至少一个氨基酸差异。在本公开的第一fc链和第二fc链中,可以在相同链中发生强烈的相互排斥,且可以在不同链间发生吸引。因此当在细胞内共同表达时,第一fc链和第二fc链,或者第一fc链、第二fc链以及与其结合的抗原结合位点更倾向于形成异源二聚体。当第一fc链和第二fc链,或者第一fc链、第二fc链以及与其结合的抗原结合位点分别在两个宿主细胞内各自表达时,第一fc链或与第一fc链结合的抗原结合位点不倾向于形成同源二聚体,且第二fc链或与第二fc链结合的抗原结合位点不倾向于形成同源二聚体。在本公开中,当第一fc链和第二fc链,或者第一fc链、第二fc链以及与其结合的抗原结合位点在存在还原剂下分别在两个宿主细胞内各自表达时,同源二聚体的比例可以为50%或更低,也就是说,fc链的单链或者fc链以及与其结合的抗原结合位点的单链的比例大于50%。
免疫球蛋白是具有四多肽链的对称结构。四多肽链中的两条是具有相对较大分子量及450至550个氨基酸残基的相同重链,且其相对分子量可以在55000道尔顿(da)至70000da的范围内。四多肽链中的另两条具有相对较小分子量及约210个氨基酸残基的相同轻链(l链),且其相对分子量可以为约24000da。免疫球蛋白的不同重链和轻链中的靠近n端的约110个氨基酸的序列可以变化很大。因此,该序列可以称为可变区(v区)。靠近c端的其余氨基酸序列相对稳定。因此,该其余序列称为恒定区(c区)。重链中的可变区占重链长度的约1/4,恒定区占重链长度的约3/4。已知五种ig包括igg(γ)、iga(α)、igd(δ)、igm(μ)和ige(ε)。igg(γ)、iga(α)和igd(δ)各自的重链包含三个恒定区,即ch1、ch2和ch3。igm(μ)和ige(ε)的重链包含一个可变重链(vh)区和四个恒定区,即ch1、ch2、ch3和ch4。恒定区既是免疫球蛋白分子的骨架,又是激活免疫反应的位点之一。
本公开中的恒定区可以包括第一fc链和第二fc链的至少一个相互作用区,且该相互作用区可以位于igg中ch3区域的一部分氨基酸中,包括至少gln347、tyr349、thr350、leu351、ser354、arg355、asp356、glu357、lys360、ser364、thr366、leu368、lys370、asn390、lys392、thr394、pro395、val397、asp399、ser400、phe405、tyr407、lys409和lys439。
第一fc链和第二fc链分别通过共价键或接头连接至抗原结合位点是指第一fc链和第二fc链分别通过共价键或接头连接至抗体的抗原结合片段,其中抗原结合片段识别可识别抗原的单链抗体,识别配体的受体,或识别受体的配体。共价键是一种化学键,其中两个或更多个原子一起共享外层电子,在理想情况下达到电子饱和状态,从而形成相对稳定的化学结构,称为共价键。共价键是原子间通过共享电子对而形成的相互作用。相同或不同元素的原子可以通过共价键结合。本公开的第一fc链和第二fc链之间的共价键包括但不限于氨基酸分子的氨基与另一个氨基酸分子的羧基脱水形成的肽键,或者乙二醇或聚乙二醇或其他化合物或其多聚物的醛基与氨基酸分子的氨基之间的肽键或亚胺键。接头可以是氨基酸序列、化合物或化合物的多聚物,其中两条多肽链通过共价键连接,其中氨基酸序列可以包括但不限于小肽,例如ggggsggggsggggs,其中接头可以通过肽键将第一fc链或第二fc链、与可识别抗原的单链抗体或可识别抗原的其他抗体片段的结构变构变体连接。
第一fc链和第二fc链可能倾向于形成异源二聚体并且可能不倾向于形成同源二聚体的事实意味着在相同的多肽链之间可能发生强烈的相互排斥,并且在第一fc链和第二fc链多肽链的不同多肽链之间可能发生吸引,因此当在细胞中共表达时,第一fc链和第二fc链;或者第一fc链、第二fc链和与其结合的抗原结合位点更倾向于形成异源二聚体。当第一fc链和第二fc链;或者第一fc链、第二fc链和与其结合的抗原结合位点分别在两个宿主细胞中各自表达,第一fc链或与第一fc链结合的抗原结合位点可能不倾向于形成同源二聚体,并且第二fc链或与第二fc链结合的抗原结合位点也可能不倾向于形成同源二聚体。
kabateu索引编号系统意味着kabat使用对抗体序列的每个氨基酸编号的方法,并且该编号每个残基的方法已成为本领域的标准方法。kabat方案可扩展到可能尚未研究的其他抗体,并且基于保守的氨基酸,靶抗体可以与由kabat鉴定的共有序列之一进行比对。
fc片段对应于ig的可结晶片段(fc)或ch2和ch3结合结构域,其中ig与效应分子或细胞发生相互作用。
igg即免疫球蛋白g的缩写,是血清中的主要抗体成分,根据igg分子中的r链抗原性差异,人igg具有igg1、igg2、igg3和igg4的四种亚型。
不完全抗体分子是指由抗体的重链和轻链形成的结构,其中重链和轻链可以通过共价键连接或可以不通过共价键连接,并且是可识别抗原的单克隆抗体结构。
fab片段是分子识别序列和抗原结合片段(fab),其对应于抗体分子的两个臂,并由轻链和重链的完整可变重链(vh)区和ch1区组成。scfv是分子识别序列,其是通过抗体的轻链可变区和重链可变区的基因工程获得的抗体片段的结构异构体。质膜受体的细胞外区域是分子识别序列,质膜受体包括通常位于细胞外侧并与相应的抗原或配体识别并结合的细胞外区域;可以将受体锚定到细胞表面的跨膜区;可能具有激酶活性或可能具有在细胞内传输信号的通道的细胞内区域。质膜受体的配体是指可以被质膜受体的细胞外区域识别并结合的蛋白质、多肽或化合物。细胞因子是由免疫原、促细胞分裂剂或其他刺激物诱导的各种细胞产生的低分子量可溶性蛋白。细胞因子执行各种功能,例如在先天性和适应性免疫、造血、细胞生长、成人多能干细胞(apsc)调节以及修复受损组织中。细胞因子可分为白介素、干扰素、肿瘤坏死因子超家族、集落刺激因子、趋化因子和生长因子等。蛋白质表达标签是指在靶蛋白的n末端或c末端添加的氨基酸序列,其可以是小肽或长氨基酸。蛋白质表达标签的添加可以促进蛋白质的正确折叠、蛋白质的分离和纯化以及减少细胞中蛋白质的降解。常用的标签包括但不限于ha、sumo、his、gst、gfp和flag。
用作本公开的异源二聚双特异性抗体的抗体类型没有特别限制。优选地,在本领域中可以使用在相关领域中已知的可用于治疗和/或预防疾病的抗体。
本公开的异源二聚双特异性抗体可以具有至少一个替换、缺失、添加和/或插入。例如,某些氨基酸可以替代蛋白质结构中的其他氨基酸,而不会显著丧失与其他多肽(例如抗原)或细胞结合的能力。由于蛋白质的结合能力和性质决定蛋白质的生物学功能活性,因此可以在蛋白质序列中替换某些氨基酸序列而不会明显丧失生物学用途或活性。
在许多情况下,多肽变体可以包含至少一个保守替换。术语“保守替换”可以指用具有相似性质的另一氨基酸替换氨基酸。肽化学领域的普通技术人员可以预期,多肽的二级结构和亲水性基本上不会发生变化。
氨基酸的替换通常可以基于氨基酸的侧链取代基的相对相似性,例如疏水性、亲水性、电荷和大小等。具有各种前述特征的示例性替换是本领域普通技术人员众所周知的,并且包括:精氨酸和赖氨酸;谷氨酸和天冬氨酸;丝氨酸和苏氨酸;谷氨酰胺和天冬酰胺;以及缬氨酸、亮氨酸和异亮氨酸。
如本公开中使用的术语“同一性”具有本领域已知的含义,并且本领域的普通技术人员还可以熟悉用于确定不同序列之间的同一性的规则和标准,是指在序列比对和引入缺口之后,在多核苷酸或多肽的变体与其非变体的序列之间的相同残基的同源性百分比(如果需要,在获得最大同源性百分比后)。在本公开中,在满足同一性定义的情况下,获得的变体序列可以具有与亲本序列相同的生物学活性。使用活性筛选变异序列的方法和手段是本领域普通技术人员众所周知的。根据本公开的教导,本领域的普通技术人员可以容易地获得这样的变体序列。在一个示例性实施方式中,与多核苷酸或多肽相比,多核苷酸或多肽的变体可以分别具有至少约70%、至少约75%、至少约80%、至少约90%、至少约95%、至少约98%、至少约99%、至少约99.1%、99.2%、99.3%、99.4%、99.5%、99.6%、99.7%、99.8%或99.9%的多核苷酸或多肽同一性。由于遗传密码的冗余,可能存在编码相同氨基酸序列的这些序列的变体。
本公开的另一个实施方式包括一种多核苷酸组合物,其中根据本公开的多核苷酸序列、其片段或其互补序列在中等至高度严格条件下杂交。杂交技术在分子生物学领域是众所周知的。为了说明的目的,用于测试本公开的多核苷酸与另一多核苷酸的杂交的适当中等严格条件包括:在5×柠檬酸钠(ssc)、0.5%sds和1.0mmol(mm)edta(ph8.0)的溶液中预洗涤;在5×ssc中在50℃至60℃的温度范围内过夜杂交;然后在65℃的温度下分别在2倍、0.5倍和0.2倍的含0.1%sds的ssc中各自洗涤两次,每次20分钟。本领域普通技术人员可以理解,可以通过例如改变杂交溶液的盐含量和/或发生杂交的温度来容易地操纵杂交的严格性。例如,在另一个实施方式中,除了例如在60℃至65℃或65℃至70℃范围内的升高杂交温度外,合适的高度严格杂交条件包括上述条件。
本公开的宿主细胞可以是可用于外源基因表达的任何细胞。宿主细胞可包括但不限于大肠杆菌(e.coli)、酵母、昆虫细胞、植物细胞和哺乳动物细胞。
本公开的载体包括可以在任何类型的细胞或生物中执行复制的载体,例如包括质粒、噬菌体、粘粒和微型染色体。在一些实施方式中,包含本公开的多核苷酸的载体可以是适合于多核苷酸的繁殖或复制的载体或适用于表达本公开的多肽的载体。这样的载体是本领域已知的并且可商购。
“载体”可以包括穿梭载体和表达载体。通常,质粒构建体还可分别包含复制起点(例如cole1的复制起点)和选择标记(例如氨苄青霉素或四环素抗性)用于细菌中的质粒复制和选择。“表达载体”是指包含表达本公开的抗体所需的调控序列或调控元件并且包括细菌或真核细胞中的抗体片段的载体。
本公开的载体可以是用于外源基因表达的任何载体,包括但不限于质粒载体。质粒载体可以包括复制起点、启动子、靶基因、多克隆位点和选择标记基因中的至少一种。优选地,本公开的载体可以包括但不限于从pcdna载体修饰的质粒载体,例如x0gc载体。
本公开的受试者包括鸟类、爬行动物和哺乳动物等。优选地,哺乳动物包括啮齿动物、灵长类动物,并且优选为人。
本公开中的疾病包括但不限于肿瘤,并且优选地,肿瘤可以包括诸如白血病、淋巴瘤、骨髓瘤、脑肿瘤、头颈部鳞状细胞癌、非小细胞肺癌、鼻咽癌、食道癌、胃癌、胰腺癌、胆囊癌、肝癌、大肠癌、乳腺癌、卵巢癌、子宫颈癌、子宫内膜癌、子宫肉瘤、前列腺癌、膀胱癌、肾细胞癌和黑色素瘤的疾病。
药学上可接受的载体是指通常在药学领域中使用的药学上的载体,例如稀释剂;赋形剂和水;填充剂如淀粉;蔗糖、乳糖或微晶纤维素;粘合剂如纤维素衍生物、藻酸盐、明胶和聚乙烯吡咯烷酮;润湿剂如甘油;崩解剂如羧甲基淀粉钠、羟丙基纤维素、交联羧甲基纤维素、琼脂、碳酸钙和碳酸氢钠;吸收增强剂如季铵化合物;表面活性剂如鲸蜡醇和十二烷基硫酸钠;吸附载体如老化土壤和肥皂粘土;润滑剂如滑石粉、硬脂酸钙和硬脂酸镁、微粉化硅胶和聚乙二醇等。佐剂例如调味剂和甜味剂等也可以添加到组合物中。
本公开将通过以下非限制性实施例进一步阐明,这是本领域普通技术人员已知的,并且在不脱离本公开的精神和范围的情况下可以对其进行许多修改。
除非另有说明,否则以下实验方法是通用方法,并且除非另有说明,否则可以容易地从商业公司获得所使用的实验材料。在本公开的以下实施例中使用的各种抗体均源自商业途径的标准抗体。
实施例1、抗pd-1/抗her2异源二聚抗体分子的载体的构建
获得抗人pd-1的重链和轻链的x0gc表达载体,其中轻链可变区的核苷酸序列如seqidno.9所示,氨基酸序列如seqidno.10所示。轻链恒定区的核苷酸序列如seqidno.3所示,氨基酸序列如seqidno.4所示。重链可变区的核苷酸序列如seqidno.11所示,氨基酸序列如seqidno.12所示。重链恒定区的核苷酸序列如seqidno.13所示,氨基酸序列如seqidno.14所示。使用聚合酶链反应(pcr)方法分别扩增轻链可变区、轻链恒定区、重链可变区和重链恒定区。在本公开中,所有pcr反应均使用phusion高保真dna聚合酶(f-530l,可从newenglandbiolabs(neb)获得)进行。根据碱基互补原理和限制性位点的需要设计pcr引物。反应系统包括8.9微升(μl)的h2o、4μl的5×phusion高保真dna聚合酶缓冲溶液、4μl的1mmdntp、1μl的正向引物、1μl的反向引物、0.1μl的phusion高保真dna聚合酶和1μl模板。在1.5%琼脂糖凝胶上对可变区和恒定区的pcr产物进行电泳,并使用dna回收试剂盒(产品编号a9282,可从promega获得)回收相应的片段。将回收的可变区片段和恒定区片段用作模板,并且使用可变区的正向引物和恒定区的反向引物再次进行pcr反应。然后,回收相应的片段,从而获得重链和轻链的全长片段。使用限制酶ecori(产品编号r3101l,可从neb获得)和hindiii(产品编号r3104l,可从neb获得)切割x0gc载体和全长片段,限制酶系统包括:32μl的10×缓冲溶液、各0.5μl的ecori和hindiii、3μl通过凝胶回收获得的全长片段和14.5μlh2o。将限制酶体系在37℃的温度下进行反应3小时。用t4dna连接酶(neb,产品编号m0202v)连接限制性酶产物,且反应体系包括2μl10×连接酶缓冲液、0.5μl连接酶、3μl通过凝胶回收获得的全长片段、3μl通过凝胶回收获得的x0gc载体和11.5μlh2o。使反应在室温下反应12小时。将连接产物分别转化到大肠杆菌感受态细胞dh5α(产物cb104,可从tiangen获得)中,从而获得抗体的重链和轻链的x0gc表达载体,用于在真核细胞中表达抗体的重链和轻链。
获得抗人her2抗体的重链和轻链的x0gc表达载体。抗体可变区的序列来源于http://www.drugbank.ca/drugs/db00072。轻链可变区的核苷酸序列如seqidno.1所示,氨基酸序列如seqidno.2所示。轻链恒定区的核苷酸序列如seqidno.3所示,氨基酸序列如seqidno,4所示。重链可变区的核苷酸序列如seqidno.5所示,氨基酸序列如seqidno.6所示。重链恒定区的核苷酸序列如seqidno.7所示,氨基酸序列如seqidno.8所示。根据前述方法,获得抗体的重链和轻链的x0gc表达载体,用于在真核细胞中表达抗体的重链和轻链。
实施例2、抗pd-1/抗her2异源二聚抗体分子的表达
将包含抗人pd-1的抗体的重链和轻链的表达载体转染到293f细胞(freestyletm293-f细胞,产品编号r79007,可从invitrogen获得)中,并且还将包含抗人her2的抗体的重链和轻链的表达载体转染到293f细胞中。转染前一天接种细胞。转染当天通过离心收集细胞。将细胞以200×105细胞/ml的细胞密度重悬于新鲜的freestyletm293表达培养基(freestyletm293表达培养基,产品号123380001,可获自gibco)。根据转染体积,以36.67μg/ml的终浓度向其中添加质粒,将混合物轻轻且均匀地混合。随后,以55μg/ml的最终浓度向其中添加线性聚乙烯亚胺(pei,线性,分子量(mw)为25000,产品编号43896,可从alfaaesar获得),并且将混合物轻轻且均匀地混合。此后,将细胞置于细胞培养箱中,并在振动器中以每分钟120转(rpm)的速度在37℃的温度下培养1小时。然后向其中加入19倍转染体积的新鲜培养基。随后,将细胞在摇床上以37℃的温度以120rpm的速率培养。通过离心收集转染5天至6天的细胞培养上清液。
表达量通过酶联免疫吸附测定(elisa)确定。在将结果应用于柱纯化之前,使用0.2μm滤膜过滤除去沉淀物。该过程在4℃的温度下进行。
实施例3、抗pd-1/抗her2异源二聚抗体分子的表达产物的纯化
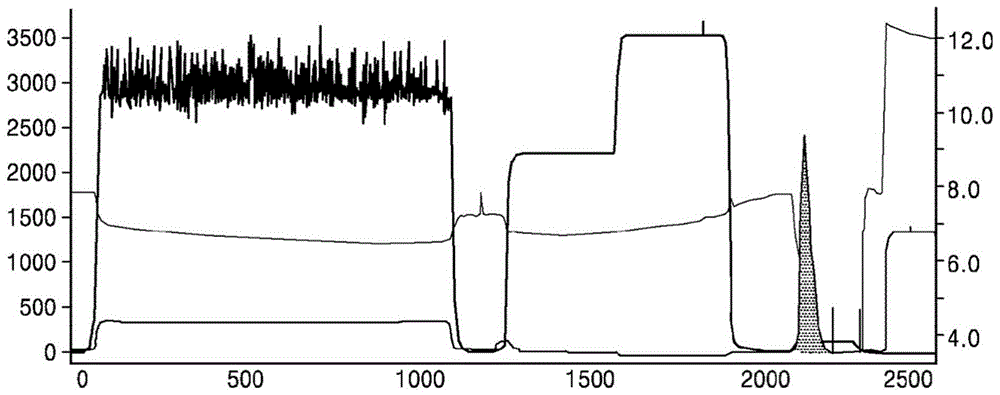
使用aktaexplorer100型蛋白质纯化系统(可从gehealthcare获得)和亲和色谱柱rproteinasepharosefastflow(16mmi.d.,10ml,可从gehealthcare获得),在4℃的温度下纯化。首先用流动相a(20mm磷酸钠缓冲溶液,150mm氯化钠,ph7.4)平衡色谱柱。基线稳定后,将样品以5ml/min的流速上样至处理过的细胞的上清液中。加载样品后,使用流动相a进行平衡。样品分别为抗pd-1表达产物和抗her2表达产物。随后,使用流动相b1(含有0.5m精氨酸的流动相a)洗涤5-柱体积。然后,使用流动相b2(100mm柠檬酸,ph3.0)洗脱5-柱体积,以最大程度地收集目标蛋白质,即洗脱峰。洗脱流速为5ml/min。抗pd-1表达产物的洗脱峰色谱图如图1所示。抗her2表达产物的洗脱峰与抗pd-1表达产物的洗脱峰相似(其结果在本文中未描述),并且收集洗脱峰(图1中的灰色区域),并通过逐滴添加1m乙酸钠溶液将ph值调节至5.0。
实施例4、抗pd-1/抗her2异源二聚抗体分子的制备和纯化
抗pd-1/抗her2异源二聚抗体分子的结构如图2所示。
将通过上述rproteinasepharosefastflow(16mmi.d.,10ml,可从gehealthcare获得)获得的抗pd-1表达产物和抗her2表达产物在体外重组以获得异源二聚体。首先,将纯化并收集的蛋白质溶液通过超滤管(标准分子量截止值为10千道尔顿(kda))通过超滤进行浓缩,然后用磷酸盐缓冲液(pbs)溶液(ph=7.4)代替溶液。将浓度为1mg/ml的pbs和最终体积为1/200的1mdtt加入到获得的抗pd-1和抗her2纯化的表达产物溶液中。dtt的最终浓度为5mm。在4℃的温度下进行还原(3至8小时),并且通过还原打开二硫键,并且抗pd-1和抗her2表达产物中包含的少量抗体同源二聚分子的铰链区的二硫键也被打开,从而形成包含重链和轻链的不完全抗体分子。其结构如图3所示。通过使用体积排阻色谱-高效液相色谱(sec-hplc)(tskgelsupersw3000,可得自tosoh),在流动相缓冲溶液中使用1mm的dtt还原剂来分析还原的样品。其结果示于图4a和4b中。抗pd-1不完全抗体分子的比例为100%,抗her2不完全抗体分子的比例为89.3%,其中剩余的10.7%为聚集体,但没有其中二硫键打开的同源二聚体存在。
将还原的抗pd-1和抗her2不完全抗体分子根据摩尔比在4℃的条件下混合24小时以进行重组。在重组中,抗pd-1和抗her2不完全抗体分子通过ch2和ch3之间的非共价相互作用形成包括抗pd-1和抗her2不完全抗体分子的异源二聚双特异性抗体。随后,通过超滤浓缩器(标准分子量截止值为10kda)对蛋白溶液进行超滤,并用pbs溶液(ph=7.4)替换以完成还原。然后通过空气或使用氧化剂进行氧化以重新形成异源二聚双特异性抗体的二硫键。氧化条件包括添加100mml-脱氢抗坏血酸作为氧化剂,蛋白质的终浓度为1mg/ml,氧化剂的终浓度为1mm。在该条件下,在4℃下进行氧化24小时。将通过氧化获得的样品进行sec-hplc分析,结果示于图5中。
将通过还原和氧化抗pd-1和抗her2不完全抗体分子而获得的异源二聚抗体分子通过超滤浓缩器(标准分子量截止值为10kda)进行超滤,并用10mmpbs溶液代替,ph为5.8。使用aktaexplorer100型蛋白质纯化系统(可从gehealthcare获得)和离子色谱柱source15s(16mmi.d.,17ml,可从gehealthcare获得),在4℃的温度下进行纯化。首先用流动相a(10mm磷酸钠缓冲溶液,ph7.0)平衡色谱柱。基线稳定后,将样品以3ml/min的流速上样至处理过的蛋白质溶液中。上样后,使用流动相a进行平衡。随后,将20-柱体积(0%b至100%b,170分钟,流速为2ml/min)从a(10mm磷酸钠,ph5.8)洗脱至b(10mm磷酸钠,ph5.8)以收集洗脱峰。收集的蛋白质溶液通过超滤浓缩器(标准分子量截止值为10kda)进行超滤,用pbs溶液代替(ph=7.4),并进行过滤灭菌。温度保持在4℃。对纯化的产物进行sec-hplc进行纯度分析。其结果在图6中示出,纯度为99.96%。
实施例5、抗pd-1/抗her2异源二聚抗体分子的稳定性
将完全密封的1mg/ml抗pd-1/抗her2异源二聚体样品置于温度为40℃的气候箱(kbf240,可从binder获得)中。在相应的时间点(基线(第一天)、两周和四周),收集20μg样品以进行sec-hplc,从而进行分离。sec-hplc的条件如下:(1)尺寸排阻色谱法:tskgelg3000swxl(购自tosohbioscience),5μm,7.8mm×30cm;(2)流动相:5mmpbs,150mmnacl,ph6.7;(3)流速:0.6ml/min;(4)紫外线检测波长:280纳米(nm);(5)采集时间:30分钟。使用的仪器为agilent1200infinity色谱仪,其使用agilentchemstation进行记录,并计算剩余单体的比例。如表1所示,在40℃的实验条件下,二聚体没有明显的聚集。因此,抗pd-1/抗her2异源二聚体被认为具有相对优异的热稳定性。
[表1]
抗pd-1/抗her2异源二聚抗体分子的稳定性
实施例6、抗pd-1/抗her2异源二聚抗体的体外靶结合活性
通过使用elisa确定抗pd-1/抗her2异源二聚抗体结合单一抗原的能力。
elisa如下进行:将重组人pd-1(产品号10377-h08h,可从北京义翘神州获得)或人her2(产品号10004-h08h,可从北京义翘神州获得)用ph9.6碳酸盐缓冲溶液(0.05m)涂布在96孔的高吸附elisa板(产品编号42592,可从costar获得)上。涂布浓度为1μg/ml。涂布量为每孔100μl,并且涂布在4℃下进行过夜。进行五次pbst洗涤。将结果使用含有1%bsa的pbst以300μl/孔进行密封,并在25℃下培养1小时。进行五次pbst洗涤。将在含1%bsa的pbst中连续稀释的100μl异源二聚抗体样品和对照加入到每个孔中,并在25℃下培养1小时。进行五次pbst洗涤。随后,将在含1%bsa的pbst中以1:2000稀释的100μl辣根过氧化物酶标记的抗人igg抗体(产品编号ap309p,可从chemicon获得)然后添加到每个孔中,并将细胞25℃下培养1小时。进行五次pbst洗涤。将100μl的比色底物tmb添加到每个孔中,并在室温下进行显色10分钟。通过向每个孔中添加100μl1mh2so4停止显色。在酶标仪上读取450nm处的吸光度。
结果,如图7a和7b所示,发现抗pd-1/抗her2异源二聚抗体对pd-1和her2具有高亲和力,从而保持二价单克隆抗体的抗原亲和活性。
实施例7、抗pd-1/抗her2异源二聚抗体的同时双重靶向结合活性
通过使用elisa测量抗pd-1/抗her2异源二聚抗体对两种不同抗原的同时结合能力。
elisa如下进行:将重组人her2(产品号10004-h08h,可从北京义翘神州获得)用ph9.6碳酸盐缓冲溶液涂布在96孔的高吸收elisa板上。涂布浓度为1μg/ml。涂布量为每孔100μl,并且涂布在4℃下进行过夜。进行五次pbst洗涤。将结果使用含1%bsa的pbst以300μl/孔进行密封,并在25℃下培养1小时。进行五次pbst洗涤。将在含1%bsa的pbst中连续稀释的100μl异源二聚抗体样品和对照加入到每个孔中,并在25℃下培养1小时。进行五次pbst洗涤。随后,将稀释于含1%bsa的pbst中的100μl生物素标记的pd-1-fc(可从北京韩美药品有限公司购买)以0.5μg/ml的浓度添加到各孔中,并将细胞在25℃下培养1小时。然后将在含有1%bsa的pbst中以1:1000稀释的100μl链霉亲和素-辣根过氧化物酶结合物(产品编号554066,可从bdpharmingen获得)加入每个孔中,并将细胞在25℃下培养1小时。进行五次pbst洗涤。将100μl的比色底物tmb添加到每个孔中,并在室温下进行显色10分钟。通过向每个孔中添加100μl1mh2so4停止显色。在酶标仪上读取450nm处的吸光度。
结果,如图8所示,pd-1单克隆抗体(重链可变区的序列和轻链可变区的序列与抗pd-1/抗her2异源二聚抗体中pd-1结合位点的对应序列相同)和her2单克隆抗体(曲妥珠单抗)的组合无法同时与pd-1和her2结合,而抗pd-1/抗her2异源二聚抗体仅具有同时与两种抗原结合的活性。
通过使用流式细胞仪(fcm,facscalibur,可从bdbiosicences获得),使用表达高pd-1的cho/pd-1细胞(产品编号为m00529,可从genscript获得)和高表达her2的sk-br-3细胞来测定抗pd-1/抗her2异源二聚抗体对双重靶向抗原的同时结合能力。
根据pkh26试剂盒(产品号slbh4568v,可从sigma获得)的说明书对cho/pd-1细胞染色。简而言之,收集cho/pd-1细胞,并在无血清培养基中洗涤一次。然后,使用pkh26试剂盒稀释液c将cho/pd-1制成2×107/ml细胞悬液。然后,将pkh26染料稀释至4μm,并与细胞悬液以1:1的比例混合。混合物悬浮液的细胞密度为1×107/ml,pkh26的浓度为2μm。将混合物悬浮液在室温下培养1小时。随后,使用等体积的胎牛血清(fbs)培养1分钟,从而完成染色。将结果以400g的离心力离心10分钟,用完全培养基洗涤两次,然后重悬在完全培养基中以备后用。根据cfse试剂盒(产品号c34554,可从lifetechnology获得)的说明书对sk-br-3细胞染色。简而言之,将cfse用工作浓度为0.5的pbs稀释,并在37℃的温度下预热,然后通过以1000rpm的速度离心5分钟收集sk-br-3细胞。将sk-br-3细胞重悬于预热的cfse工作溶液中,并在37℃的温度下培养15分钟,然后,以1000rpm的速率离心5分钟收集细胞。接下来,将细胞重悬于完全培养基中并培养30分钟。此后,使用完全培养基洗涤细胞,并重悬以完成制备。将染色的细胞离心并收集,并用含有2%fbs的冷pbs洗涤一次。将细胞重悬于含有2%fbs的冷pbs中,使得细胞密度为5×106/ml。将sk-br-3细胞和cho/pd-1细胞以1:1的比例混合,并分别从流量管中收集每种细胞100μl(即2.5×105个sk-br-3细胞和2.5×105个cho/pd-1细胞)。接下来,将100μl的稀释的异源二聚抗体样品、对照和同种型对照(人免疫球蛋白,可从江西博雅生物药业,国家食品药品监督管理局(sfda)批准号s19993012获得)添加到最终浓度为5nm的含有2%fbs的冷pbs中。将流量管在冰上培养30分钟。然后用含有2%fbs的pbs洗涤细胞。将细胞重悬于500μl的冷pbs中,并将细胞悬浮在流式细胞仪中以进行测量分析。
结果,如图9所示,发现异源二聚抗体同时与表达高pd-1的cho/pd-1细胞和表达高her2的sk-br-3细胞结合。因此,抗pd-1/抗her2异源二聚抗体能够将sk-br-3细胞吸引至cho/pd-1细胞。
实施例8、抗pd-1/抗her2异源二聚抗体对pd-1与配体pd-l1或pd-l2之间结合的阻断活性
通过elisa测定抗pd-1/抗her2异源二聚抗体对pd-1和pd-l1之间的结合以及pd-1和pd-l2之间的结合的阻断活性。
将重组人pd-1-fc用ph9.6的pbs涂布在96孔的高吸附elisa板上。涂布浓度为1μg/ml。涂布量为每孔100μl,并且涂布在4℃下进行过夜。进行五次pbst洗涤。将结果使用含有1%bsa的pbst以300μl/孔进行密封,并在25℃下培养1小时。进行五次pbst洗涤。将在含1%bsa的pbst中连续稀释的异源二聚体样品和对照加入其中,然后向每个孔中加入100μl终浓度为1μg/ml的生物素标记的pd-l1-fc或终浓度为4μg/ml的生物素标记的pd-l2。然后,将细胞在25℃下培养1小时。进行五次pbst洗涤。随后,在每个孔中加入在含1%bsa的pbst中以1:1000稀释的100μl辣根过氧化物酶标记的链霉亲和素(产品编号554066,可从bdpharmingen获得),然后将细胞在25℃下培养1小时。进行五次pbst洗涤。将100μl的比色底物tmb添加到每个孔中,并在室温下进行显色10分钟。通过向每个孔中添加100μl1mh2so4停止显色。在酶标仪上读取450nm处的吸光度。
结果,如图10a和10b所示,发现抗pd-1/抗her2异源二聚抗体阻断pd-1和pd-l1与pd-1和pd-l2之间的结合,从而维持二价单克隆抗体的阻断活性具有相对优秀的水平。
实施例9、抗pd-1/抗her2异源二聚抗体分子的t细胞调节活性
通过使用混合淋巴细胞反应(mlr)测量抗pd-1/抗her2异源二聚抗体对t细胞免疫反应的调节活性。
人树突状细胞(dc)的获得:通过繁殖收集人外周血单核细胞(pbmc)(产品号cc-2702,可获自lonza)。将人pbmc以5×106/ml的细胞密度重悬于无血清的rpmi1640培养基中,并接种在细胞培养瓶中,随后在37℃的二氧化碳培养箱中培养90分钟。弃去上清液和培养液的重悬细胞,并将贴壁细胞在完全培养基(含有10%fbs的rpmi1640)中培养。随后,将100ng/mlgm-csf(产品编号10016-hnah,可从北京义翘神州获得)和100ng/mlil-4(产品编号11846-hnae,可从北京义翘神州获得)添加到细胞中。将细胞培养三天,然后更换溶液。然后,将细胞再次培养三天。接下来,将培养基更换为完全培养基(含有10%fbs的rpmi1640),其中含有100ng/mlgm-csf、100ng/mlil-4和20ng/mltnf-α,然后培养一天。因此,获得dc。
人t细胞的获得:通过繁殖收集人pbmc(产品号cc-2702,可获自lonza)。pbmc和生成dc的pbmc各来自不同的受试者。根据pant细胞分离试剂盒(产品号5150414820,可从miltenyibiotech获得)的说明书分离人t细胞。简而言之,pbmc用pbs洗涤一次,并将pbmc重悬于107个细胞/40μl的分离缓冲溶液(2mm乙二胺四乙酸(edta),ph7.2pbs,含0.5%bsa)中。然后,向其中加入10μlpant细胞生物素抗体混合物,然后在4℃的温度下培养5分钟。随后,向其中添加30μl分离缓冲溶液和20μlpant细胞microbead混合物,然后在4℃下培养10分钟。然后通过磁性活化细胞分选仪(macs)分离柱获得t细胞。
将收集的人dc和人t细胞重悬在完全培养基(含有10%fbs的rpmi1640)中,并接种在96孔板上。将接种的dc和t细胞混合并分别以1×104个/孔、1×105个/孔的浓度培养。向其中加入在完全培养基中连续稀释的异源二聚抗体样品和对照。将培养板置于37℃的二氧化碳培养箱中并培养5天。培养完成后,根据试剂盒手册将孔中的上清液用于检测细胞因子ifn-γ(产品号elh-ifng,可从raybiotech获得)。
如图11所示,人t细胞在同种异体dc的刺激下激活ifn-γ的分泌。当添加pd-1单克隆抗体时,可以增强t细胞的活化,并且可以促进细胞因子的分泌。但是,her2单克隆抗体不具有这种活性。抗pd-1/抗her2异源二聚抗体具有与pd-1单克隆抗体相当的t细胞调节活性,并显著促进细胞因子ifn-γ的分泌
实施例10、抗pd-1/抗her2异源二聚抗体分子的肿瘤细胞抑制活性
测试抗pd-1/抗her2异源二聚抗体在人pbmc存在下对人乳腺癌细胞即sk-br-3细胞的杀伤活性。
将sk-br-3细胞在含有10%fbs的rpmi1640培养基(即完全培养基)中培养。收集sk-br-3细胞并重悬于完全培养基中。然后将100μl细胞以5×104/ml的细胞密度接种到96孔培养板的每个孔中。也就是说,每个孔中接种5000个细胞。通过繁殖收集人pbmc(产品号cc-2702,可获自lonza)。将人pbmc以5×105/ml的细胞密度重悬于rpmi1640完全培养基中。然后在96孔培养板的每个孔中接种50μl细胞。也就是说,每个孔中接种了25000个细胞。有效靶细胞的比例为5:1。将在完全培养基中连续稀释的50μl每种异源二聚抗体样品分别添加到每个孔中。将培养板置于37℃和5%二氧化碳的培养箱中培养三天。培养完成后,将细胞培养板中的pbmc洗涤并使用培养基除去。然后,向其中加入100μl完全培养基和20μlmts(celltiter96水溶液,产品编号g358b,可从promega获得),以检测sk-br-3细胞。将细胞培养板在培养箱中进一步培养3至4小时。之后,在酶标仪上读取490nm处的吸光度。
如图12所示,加入her2单克隆抗体后,her2单克隆抗体杀死并抑制sk-br-3细胞;但是,pd-1单克隆抗体在体外没有这种活性。抗pd-1/抗her2异源二聚抗体还表现出与her2单克隆抗体相当的抑制肿瘤细胞的杀伤活性。
实施例11、抗pd-1/抗her2异源二聚抗体分子在动物中的抗肿瘤功效的研究
实验材料选自6至8周龄的雌性免疫缺陷的ncg小鼠(可从南京大学的南京生物医学研究所获得)。在小鼠适应环境后一周,在每只小鼠的右背侧皮下接种5×106个人乳腺癌细胞。一旦肿瘤体积的长度达到约100立方毫米(mm3),就根据肿瘤体积将小鼠分组,每组包括8只小鼠。首先,通过施用人pbmc将小鼠的免疫系统部分人源化,并在每只小鼠中静脉内接种5×106个细胞。随后,每周两次施用各溶剂(pbs)、35纳摩尔每千克(nmol/kg,5mg/kg)pd-1单克隆抗体,35nmol/kg(5mg/kg)her2单克隆抗体,35nmol/kgpd-1单克隆抗体和35nmol/kgher2单克隆抗体的组合,以及35nmol/kg(5mg/kg)抗pd-1/抗her2异源二聚抗体,连续施用2周。通过腹膜内注射进行施用。从施用开始之日起,每周测量三次肿瘤体积。测量相对长的直径(a)和相对短的直径(b),并且如下计算肿瘤体积:肿瘤体积(mm3)=(a×b2)/2。观察肿瘤体积3周,即停止施用后再观察肿瘤体积变化1周。
结果,如图13所示,发现抗pd-1/抗her2异源二聚抗体比pd-1单克隆抗体和her2单克隆抗体具有更强的抗肿瘤功效。即使停止施用后,抗pd-1/抗her2异源二聚抗体也显示出中等的肿瘤抑制作用。
序列表
<110>北京韩美药品有限公司
信达生物制药(苏州)有限公司
<120>抗pd-1/抗her2天然抗体结构的异源二聚双特异性抗体及其制备方法
<130>lz1800230cn07
<160>14
<170>patentin版本3.3
<210>1
<211>321
<212>dna
<213>人工序列
<220>
<223>抗人her2抗体轻链可变区
<400>1
gacattcagatgactcagagcccttcttcactgtcagcttccgtgggcgacagagtcact60
atcacctgccgcgcaagtcaggatgtgaacaccgcagtcgcctggtaccagcagaagcct120
ggcaaagctccaaagctgctgatctacagcgcatctttcctgtattctggagtgcccagt180
aggtttagtgggtcacggtccggtaccgacttcacactgactatctccagcctgcagcct240
gaggattttgccacatactattgccagcagcactataccacaccccctactttcggccag300
ggaaccaaagtggagatcaag321
<210>2
<211>107
<212>prt
<213>人工序列
<220>
<223>抗人her2抗体轻链可变区
<400>2
aspileglnmetthrglnserproserserleuseralaservalgly
151015
aspargvalthrilethrcysargalaserglnaspvalasnthrala
202530
valalatrptyrglnglnlysproglylysalaprolysleuleuile
354045
tyrseralaserpheleutyrserglyvalproserargphesergly
505560
serargserglythraspphethrleuthrileserserleuglnpro
65707580
gluaspphealathrtyrtyrcysglnglnhistyrthrthrpropro
859095
thrpheglyglnglythrlysvalgluilelys
100105
<210>3
<211>321
<212>dna
<213>人工序列
<220>
<223>抗人her2抗体轻链可变区
<400>3
cgaactgtggccgctccaagcgtcttcatttttccaccctctgacgaacagctgaagtca60
gggacagcttccgtggtctgtctgctgaacaatttttaccccagggaggccaaagtgcag120
tggaaggtcgataacgctctgcagagcggaaattctcaggagagtgtgacagaacaggac180
tcaaaagattccacttatagcctgtctagtaccctgacactgtccaaggcagactacgaa240
aagcataaagtgtatgcctgtgaggtcacacatcagggtctgtcaagccccgtcactaag300
tccttcaatcgtggcgaatgc321
<210>4
<211>107
<212>prt
<213>人工序列
<220>
<223>抗人her2抗体轻链可变区
<400>4
argthrvalalaalaproservalpheilepheproproseraspglu
151015
glnleulysserglythralaservalvalcysleuleuasnasnphe
202530
tyrproargglualalysvalglntrplysvalaspasnalaleugln
354045
serglyasnserglngluservalthrgluglnaspserlysaspser
505560
thrtyrserleuserserthrleuthrleuserlysalaasptyrglu
65707580
lyshislysvaltyralacysgluvalthrhisglnglyleuserser
859095
provalthrlysserpheasnargglyglucys
100105
<210>5
<211>360
<212>dna
<213>人工序列
<220>
<223>抗人her2抗体重链可变区
<400>5
gaggtgcagctggtcgaaagtgggggtgggctggtgcagccaggcggatcactgaggctg60
tcctgcgccgctagcggcttcaacatcaaagacacctatattcactgggtccgacaggca120
ccagggaagggtctggaatgggtggctcgtatctaccctacaaatggttacactagatat180
gccgactccgtgaaaggccggtttactatttctgctgataccagtaagaacacagcatac240
ctgcagatgaatagcctgagggctgaggataccgcagtgtactattgctctcggtggggg300
ggtgacggcttctacgctatggattattggggccagggaactctggtcaccgtgtccagc360
<210>6
<211>120
<212>prt
<213>人工序列
<220>
<223>抗人her2抗体重链可变区
<400>6
gluvalglnleuvalgluserglyglyglyleuvalglnproglygly
151015
serleuargleusercysalaalaserglypheasnilelysaspthr
202530
tyrilehistrpvalargglnalaproglylysglyleuglutrpval
354045
alaargiletyrprothrasnglytyrthrargtyralaaspserval
505560
lysglyargphethrileseralaaspthrserlysasnthralatyr
65707580
leuglnmetasnserleuargalagluaspthralavaltyrtyrcys
859095
serargtrpglyglyaspglyphetyralametasptyrtrpglygln
100105110
glythrleuvalthrvalserser
115120
<210>7
<211>990
<212>dna
<213>人工序列
<220>
<223>抗人her2抗体重链恒定区
<400>7
gcttcaacaaaaggaccttccgtgtttccactggcaccctctagtaagagtacttcagga60
ggaaccgcagcactgggatgtctggtgaaggactacttcccagagcccgtcaccgtgtct120
tggaacagtggagcactgacctccggggtccatacatttcctgccgtgctgcagtcatcc180
ggtctgtatagcctgagctctgtggtcacagtcccaagttcatccctgggcacccagaca240
tacatctgcaacgtgaatcacaaaccttccaatactaaggtcgacaagaaagtggaacca300
aaaagctgtgataagactcatacctgcccaccttgtcctgcaccagagctgctgggaggt360
ccaagcgtgttcctgtttccacccaagcccaaagacacactgatgatttctcgcacaccc420
gaagtcacttgtgtggtcgtggacgtgtcccacgaggatcctgaagtcaagttcaactgg480
tacgtggatggcgtcgaggtgcataatgctaagaccaaacccagagaggaacagtacaac540
agcacctatcgcgtcgtgtctgtcctgacagtgctgcaccaggattggctgaacggaaag600
gagtacaagtgcaaagtgagcaacaaggccctgcccgctcctatcgagaagaccatttct660
aaggctaaaggccagcctagagaaccacaggtgtatacagagcctccaagtcgcgacgag720
ctgacaaaaaaccaggtctccctgacttgtctggtgaagggattctaccctagcgatatc780
gcagtggagtgggaatctaatgggcagccagaaaacaattataagaccacaccccctgtg840
ctggactcagatggttccttctttctgctgagtgtgctgaccgtggacaagtccaggtgg900
cagcaggggaacgtcttttcctgcagcgtgatgcatgaggccctgcacaatcattacaca960
cagaaatctctgagtctgtcaccaggaaag990
<210>8
<211>330
<212>prt
<213>人工序列
<220>
<223>抗人her2抗体重链恒定区
<400>8
alaserthrlysglyproservalpheproleualaproserserlys
151015
serthrserglyglythralaalaleuglycysleuvallysasptyr
202530
pheprogluprovalthrvalsertrpasnserglyalaleuthrser
354045
glyvalhisthrpheproalavalleuglnserserglyleutyrser
505560
leuserservalvalthrvalproserserserleuglythrglnthr
65707580
tyrilecysasnvalasnhislysproserasnthrlysvalasplys
859095
lysvalgluprolyssercysasplysthrhisthrcysproprocys
100105110
proalaprogluleuleuglyglyproservalpheleuphepropro
115120125
lysprolysaspthrleumetileserargthrprogluvalthrcys
130135140
valvalvalaspvalserhisgluaspprogluvallyspheasntrp
145150155160
tyrvalaspglyvalgluvalhisasnalalysthrlysproargglu
165170175
gluglntyrasnserthrtyrargvalvalservalleuthrvalleu
180185190
hisglnasptrpleuasnglylysglutyrlyscyslysvalserasn
195200205
lysalaleuproalaproileglulysthrileserlysalalysgly
210215220
glnproarggluproglnvaltyrthrgluproproserargaspglu
225230235240
leuthrlysasnglnvalserleuthrcysleuvallysglyphetyr
245250255
proseraspilealavalglutrpgluserasnglyglnprogluasn
260265270
asntyrlysthrthrproprovalleuaspseraspglyserphephe
275280285
leuleuservalleuthrvalasplysserargtrpglnglnglyasn
290295300
valphesercysservalmethisglualaleuhisasnhistyrthr
305310315320
glnlysserleuserleuserproglylys
325330
<210>9
<211>321
<212>dna
<213>人工序列
<220>
<223>抗人her2抗体轻链可变区
<400>9
gacatccagatgacccagtcccctagcagcgtgagcgcttccgtgggcgacagggtgacc60
atcacctgcagggcctcccagggcatctcctcctggctggcctggtatcaacagaagccc120
ggcaaggcccccaagctgctgatctccgctgcctcctccctgcagtccggagtgccttcc180
aggttcagcggttctggctccggcaccgacttcaccctgaccatctcctccctgcagccc240
gaggacttcgccacctactactgccagcaggccaaccacctgcctttcaccttcggcggc300
ggcaccaaggtggagatcaag321
<210>10
<211>107
<212>prt
<213>人工序列
<220>
<223>抗人her2抗体轻链可变区
<400>10
aspileglnmetthrglnserproserservalseralaservalgly
151015
aspargvalthrilethrcysargalaserglnglyilesersertrp
202530
leualatrptyrglnglnlysproglylysalaprolysleuleuile
354045
seralaalaserserleuglnserglyvalproserargphesergly
505560
serglyserglythraspphethrleuthrileserserleuglnpro
65707580
gluaspphealathrtyrtyrcysglnglnalaasnhisleuprophe
859095
thrpheglyglyglythrlysvalgluilelys
100105
<210>11
<211>360
<212>dna
<213>人工序列
<220>
<223>抗人her2抗体重链可变区
<400>11
caggtgcagctggtgcagtccggagccgaggtgaagaagcccggctcctccgtgaaggtg60
tcctgcaaggcctccggcggcaccttctcctccaccgccatctcctgggtgaggcaggct120
cctggccagggactggagtggatgggaggcatctggccctccttcggcacagcctcctac180
gcccagaagttccagggcagggtgaccatcaccgccgacgagagcacctccaccgcctac240
atggagctgagctccctgaggtccgaggacaccgccgtgtactactgtgccagggccgag300
tactcctccaccggcatcttcgactactggggccagggcaccctggtgacagtgtcctcc360
<210>12
<211>120
<212>prt
<213>人工序列
<220>
<223>抗人her2抗体重链可变区
<400>12
glnvalglnleuvalglnserglyalagluvallyslysproglyser
151015
servallysvalsercyslysalaserglyglythrpheserserthr
202530
alailesertrpvalargglnalaproglyglnglyleuglutrpmet
354045
glyglyiletrpproserpheglythralasertyralaglnlysphe
505560
glnglyargvalthrilethralaaspgluserthrserthralatyr
65707580
metgluleuserserleuargsergluaspthralavaltyrtyrcys
859095
alaargalaglutyrserserthrglyilepheasptyrtrpglygln
100105110
glythrleuvalthrvalserser
115120
<210>13
<211>990
<212>dna
<213>人工序列
<220>
<223>抗人her2抗体重链恒定区
<400>13
gcttcaacaaaaggaccttccgtgtttccactggcaccctctagtaagagtacttcagga60
ggaaccgcagcactgggatgtctggtgaaggactacttcccagagcccgtcaccgtgtct120
tggaacagtggagcactgacctccggggtccatacatttcctgccgtgctgcagtcatcc180
ggtctgtatagcctgagctctgtggtcacagtcccaagttcatccctgggcacccagaca240
tacatctgcaacgtgaatcacaaaccttccaatactaaggtcgacaagaaagtggaacca300
aaaagctgtgataagactcatacctgcccaccttgtcctgcaccagagctgctgggaggt360
ccaagcgtgttcctgtttccacccaagcccaaagacacactgatgatttctcgcacaccc420
gaagtcacttgtgtggtcgtggacgtgtcccacgaggatcctgaagtcaagttcaactgg480
tacgtggatggcgtcgaggtgcataatgctaagaccaaacccagagaggaacagtacaac540
agcacctatcgcgtcgtgtctgtcctgacagtgctgcaccaggattggctgaacggaaag600
gagtacaagtgcaaagtgagcaacaaggccctgcccgctcctatcgagaagaccatttct660
aaggctaaaggccagcctagagaaccacaggtgtatacactgcctccaagtcgcgacgag720
ctgacaaaaaaccaggtctccctgctgtgtctggtgaagggattctaccctagcgatatc780
gcagtggagtgggaatctaatgggcagccagaaaacaattataagaccacaccccctgtg840
ctgcggtcagatggttccttctttctgtacagtaaactgaccgtggacaagtccaggtgg900
cagcaggggaacgtcttttcctgcagcgtgatgcatgaggccctgcacaatcattacaca960
cagaaatctctgagtctgtcaccaggaaag990
<210>14
<211>330
<212>prt
<213>人工序列
<220>
<223>抗人her2抗体重链恒定区
<400>14
alaserthrlysglyproservalpheproleualaproserserlys
151015
serthrserglyglythralaalaleuglycysleuvallysasptyr
202530
pheprogluprovalthrvalsertrpasnserglyalaleuthrser
354045
glyvalhisthrpheproalavalleuglnserserglyleutyrser
505560
leuserservalvalthrvalproserserserleuglythrglnthr
65707580
tyrilecysasnvalasnhislysproserasnthrlysvalasplys
859095
lysvalgluprolyssercysasplysthrhisthrcysproprocys
100105110
proalaprogluleuleuglyglyproservalpheleuphepropro
115120125
lysprolysaspthrleumetileserargthrprogluvalthrcys
130135140
valvalvalaspvalserhisgluaspprogluvallyspheasntrp
145150155160
tyrvalaspglyvalgluvalhisasnalalysthrlysproargglu
165170175
gluglntyrasnserthrtyrargvalvalservalleuthrvalleu
180185190
hisglnasptrpleuasnglylysglutyrlyscyslysvalserasn
195200205
lysalaleuproalaproileglulysthrileserlysalalysgly
210215220
glnproarggluproglnvaltyrthrleuproproserargaspglu
225230235240
leuthrlysasnglnvalserleuleucysleuvallysglyphetyr
245250255
proseraspilealavalglutrpgluserasnglyglnprogluasn
260265270
asntyrlysthrthrproprovalleuargseraspglyserphephe
275280285
leutyrserlysleuthrvalasplysserargtrpglnglnglyasn
290295300
valphesercysservalmethisglualaleuhisasnhistyrthr
305310315320
glnlysserleuserleuserproglylys
325330
- 还没有人留言评论。精彩留言会获得点赞!