用于癌症治疗的精氨酸酶抑制的制作方法
本发明涉及用于治疗癌症的免疫细胞、试剂盒、方法和组合物,特别是在癌症免疫疗法和/或过继细胞转移疗法领域。本发明还涉及用于生产细胞、试剂盒和组合物的方法。
背景技术:
免疫系统在抑制恶性肿瘤的发生、抑制肿瘤进展和促进肿瘤消除方面发挥根本作用。为了逃逸免疫监视,肿瘤倾向于执行多种逃逸机制。免疫逃逸策略包括产生抗炎细胞因子、募集包含调节性t细胞(tregs)和髓样来源的抑制性细胞(mdsc)的抑制性细胞、负性t细胞共刺激分子的表达以及t细胞免疫抑制代谢途径的活化。通过抑制这种逃逸机制增强靶向肿瘤的免疫应答是具有重大治疗前景的策略。抗体介导的负性免疫检查点调节剂阻断是迄今为止最有希望和最成功的免疫治疗方法之一。此类免疫检查点途径通常通过终止或减弱过度t细胞应答以预防或减轻免疫病理。肿瘤细胞利用这种机制防止t细胞攻击。通过治疗干预的方式释放这些“阻碍”可因此恢复有效的抗肿瘤免疫应答。然而,尽管通过干扰pdl1-pd1和b7-ctla4抑制途径获得了空前的成功,但目前的检查点阻断疗法并未在所有患者中引发有效的抗肿瘤免疫应答,并未对所有类型的癌症均有作用,仅具有部分疗效,或肿瘤对检查点阻断产生耐受性。
癌症免疫疗法的另一种形式包括利用患者的免疫细胞通过过继细胞疗法根除癌症,其形式为患者肿瘤浸润淋巴细胞(til)的离体扩增和对肿瘤抗原产生应答的活化t淋巴细胞群的回输,或取出患者的外周血t淋巴细胞,然后进行离体基因修饰,以表达转基因t细胞受体(tcr)或嵌合抗原受体(car)靶向已知的肿瘤抗原,然后回输至患者体内以破坏癌细胞。cart细胞免疫疗法已在血液学肿瘤中显示出空前的成功,但到目前为止,使用cart细胞治疗实体瘤在很大程度上是不成功的,部分原因是免疫抑制性肿瘤的微环境。另外,在体外产生过程中需要予以相当的关注以防止cart细胞过度耗竭或分化。
因此,迫切需要开发旨在单独使用或与现有疗法联合使用以诱导强抗肿瘤应答的新策略。特别地,涉及不同免疫治疗剂的协同治疗联合以及其他互补疗法可能构成将来成功抗癌治疗的基础。
值得进一步研究的一种治疗策略是通过抑制抗肿瘤免疫应答来干扰有利于肿瘤生长的代谢途径。在这些证据中,越来越多的证据表明,l-精氨酸代谢的治疗性操纵可有助于增强抗肿瘤免疫应答。l-精氨酸是半必需氨基酸,需要某些细胞特别是免疫系统的某些细胞(包括t细胞和巨噬细胞)从细胞外环境中吸收。充分的l-精氨酸可获得性对于这些细胞的最佳功能至关重要。在t细胞的情况下,l-精氨酸消耗导致cd3-ζ链表达减少,经t细胞受体(tcr)-cd3复合物的信号传导受损以及抗原特异性t细胞活化、增殖和细胞毒性的抑制。
l-精氨酸是精氨酸酶(arg)和一氧化氮合酶(nos)的底物。nos的l-精氨酸分解代谢导致no和其他反应性氮中间体的产生,这是导致巨噬细胞抗病原体细胞毒活性的关键机制。arg将l-精氨酸转化为尿素和l-鸟氨酸,该反应因其在尿素循环中对肝脏中氨的解毒作用而闻名。l-鸟氨酸被进一步加工成多胺和脯氨酸。已经发现,arg和nos酶在肿瘤中的表达增加,无论是在肿瘤细胞本身中还是在肿瘤浸润细胞中,诸如巨噬细胞和髓样来源的抑制细胞(mdsc)。在肿瘤微环境(tme)中,nos介导的no产生促进肿瘤血管生成和转移,而arg介导的脯氨酸可获得性增加和多胺合成促进肿瘤细胞增殖。此外,肿瘤中由arg和nos表达诱导的l-精氨酸消耗有助于通过上述机制建立免疫抑制性微环境,这种微环境损害抗肿瘤t细胞应答。
哺乳动物有两种arg酶,分别是由两个不同的基因编码的精氨酸酶1(arg1)和精氨酸酶2(arg2)。两种酶催化相同的生化应答,在其催化位点显示100%同源性,并且均被目前可获得的arg抑制剂抑制。arg1和arg2在细胞内定位和表达方式方面有所差异。arg1是一种胞质酶,主要在肝脏中表达,但也可以在几种非肝组织和细胞类型中表达。arg2位于线粒体中,并在多种组织中广泛表达。
在dunand-sauthieri.etal,"repressionofarginase-2expressionindendriticcellsbymicrorna-155iscriticalforpromotingtcellproliferation",jimmunol.193:1690-1700(2014)的出版物中发现arg2是树突细胞(dc)中表达的主要精氨酸酶,并且在dc成熟过程中,mir155抑制其相应的mrna。在活化的mir155缺失的dc中观察到arg2表达和活性异常升高。相反,mir155的过表达抑制dc中arg2的表达。众所周知,mir155可控制不同细胞类型中的不同基因,而且尚未报道mir155是否也可控制其他细胞类型(包括t细胞)中arg2的表达。
wo2014059248教导了通过将编码mir155转录物的核酸分子离体引入从受试者分离的t细胞中以增加t细胞介导的免疫的方法。us2015/275209类似地教导了分离的或纯化的cd8+t细胞,其包括抗原特异性t细胞受体(tcr)和编码微小rna分子的外源核酸。wo2014059248和us2015/275209均未公开mir-155在t细胞中的活性方式。在graciasd.etal,“microrna-155controlscd8+tcellresponsesbyregulatinginterferonsignaling”,natimmunol.14:593-602(2013)中发现mir155调节cd8+t细胞中845个基因的表达,其中大多数显示小于2倍的差异表达,这表明mir155适度影响大量转录本,而不是对单独靶标具有强影响。graciasetal.在mir155调节其表达的845个基因中未识别arg2。因此,这些参考文献并未表明arg2可能是t细胞中mir155的靶标且mir155的作用是通过抑制t细胞中的arg2介导的。
若干专利文献涉及精氨酸酶抑制剂。ep2768491b1公开了用于治疗多种病症的小分子精氨酸酶抑制剂,所述病症诸如心血管疾病、性疾病、伤口愈合疾病、胃肠疾病、自身免疫疾病、免疫疾病、感染、肺疾病、纤维化疾病和溶血性疾病。ep2083812b1公开了用于治疗过敏性和非过敏性哮喘以及过敏性鼻炎的小分子精氨酸酶抑制剂,诸如6-硼诺-l-正亮氨酸(abh)。wo2007/111626公开了用于治疗动脉粥样硬化疾病的arg2活性调节,包括用于沉默arg2的sirna。us9789169公开了用于消耗血浆精氨酸水平的重组arg1蛋白,其目的是调节免疫系统,特别是防止移植器官的排斥。
本发明的目的是提供一种治疗癌症的方法,其优选地有效抗造血肿瘤和/或实体瘤。
本发明的目的是改善基于嵌合抗原受体(car)t细胞和/或转基因t细胞受体t细胞的疗法和/或基于与免疫检查点调节剂的相互作用和/或阻断的疗法的疗效。例如,目的是提供改善现有癌症疗法或现有治疗概念的方法。
本发明的目的是提供可以与其他癌症治疗(包括免疫治疗剂)协同使用的癌症治疗。
本发明的目的是通过促进癌症定向免疫应答来治疗癌症。特别地,本发明的目的是抑制癌细胞利用其逃逸癌症定向免疫应答的机制。
本发明解决了上述需求和目的。
技术实现要素:
值得称道的是,本发明人开展的实验表明,具有受损的精氨酸酶活性和/或表达的免疫细胞对于治疗癌症(包括实体瘤)更有效。
一方面,本发明提供了具有受损的精氨酸酶活性和/或表达的免疫细胞,用于治疗癌症,包括实体瘤。
一方面,本发明提供表达嵌合抗原受体(car)的免疫细胞,所述免疫细胞进一步具有受损的精氨酸酶活性和/或表达。
一方面,本发明提供了具有受损的精氨酸酶活性和/或表达的肿瘤浸润淋巴细胞(til)。优选地,所述til已经从患有癌症的人或动物分离。
一方面,本发明提供表现出组成性或诱导性精氨酸酶活性和/或表达受损的免疫细胞,用于通过过继细胞转移治疗癌症。
一方面,本发明提供了一种治疗癌症的方法,所述方法包括损害免疫细胞中的精氨酸酶活性和/或表达。
一方面,本发明提供了一种治疗癌症的方法,所述方法包括向有需要的受试者施用精氨酸酶活性和/或表达受损的免疫细胞。优选地,所述方法是通过过继细胞转移治疗癌症的方法。
一个方面,本发明提供了一种治疗癌症的方法,所述方法包括施用肿瘤浸润淋巴细胞(til),所述til还具有受损的精氨酸酶活性和/或表达。
一方面,本发明提供了一种治疗癌症的方法,所述方法包括施用表达car的免疫细胞,所述免疫细胞具有受损的精氨酸酶活性和/或表达。
一方面,本发明提供了一种治疗癌症的方法,所述方法包括:提供表达car和/或til的细胞,处理所述细胞,优选在体外和/或离体进行,以降低精氨酸酶活性和/或在所述细胞中的表达,以及向有需要的受试者,优选为患有癌症的受试者,施用所述细胞。
一方面,本发明提供了一种用于改善免疫细胞的抗癌活性的方法,所述方法包括:损害所述免疫细胞中的精氨酸酶活性和/或表达,优选地离体损害所述免疫细胞中的精氨酸酶活性和/或表达。
一方面,本发明提供了一种改善用于过继细胞转移的免疫细胞抗癌活性的方法,所述方法包括:离体损害所述免疫细胞中的精氨酸酶活性和/或表达。
一方面,本发明提供了一种用于准备和/或产生抗癌治疗,特别是用于过继细胞转移的抗癌治疗的方法,所述方法包括:提供免疫细胞,以及离体损害所述免疫细胞中的精氨酸酶活性和/或表达。
一方面,本发明提供了能够降低免疫细胞中的精氨酸酶活性以治疗癌症的制剂。
一方面,本发明提供了用于治疗癌症的试剂盒,所述试剂盒包括适合于损害免疫细胞中的精氨酸酶活性和/或表达的制剂。
一方面,本发明提供了一种联合治疗,其包括施用具有受损的精氨酸酶活性和/或表达的免疫细胞和阻断免疫抑制途径的制剂,诸如pdl1-pd1和b7-ctla4抑制途径。优选地,所述制剂是抗体(ab),例如,抗pd1ab或抗ctla4ab。
一方面,本发明提供了一种组合物,其包括免疫细胞,特别是本发明的免疫细胞。
一方面,本发明提供了具有受损的精氨酸酶活性和/或表达的免疫细胞,用于减小患有肿瘤和/或癌症的受试者的肿瘤大小和/或体积。一方面,本发明提供了具有受损的精氨酸酶活性和/或表达的免疫细胞,用于增加患有癌症的受试者的生存率和/或时间。
一方面,本发明提供了一种用于产生癌症治疗的方法,所述方法包括提供分离的免疫细胞并对所述免疫细胞进行处理以使精氨酸酶活性和/或表达受损。
本发明的其他方面和优选实施例在下文和所附权利要求书中定义。通过以下提供的优选实施例的描述,本发明的其他特征和优势对于技术人员将变得显而易见。
附图说明
在附图中,为了说明本发明的实施例和/或实例,示出了实验部分的结果。
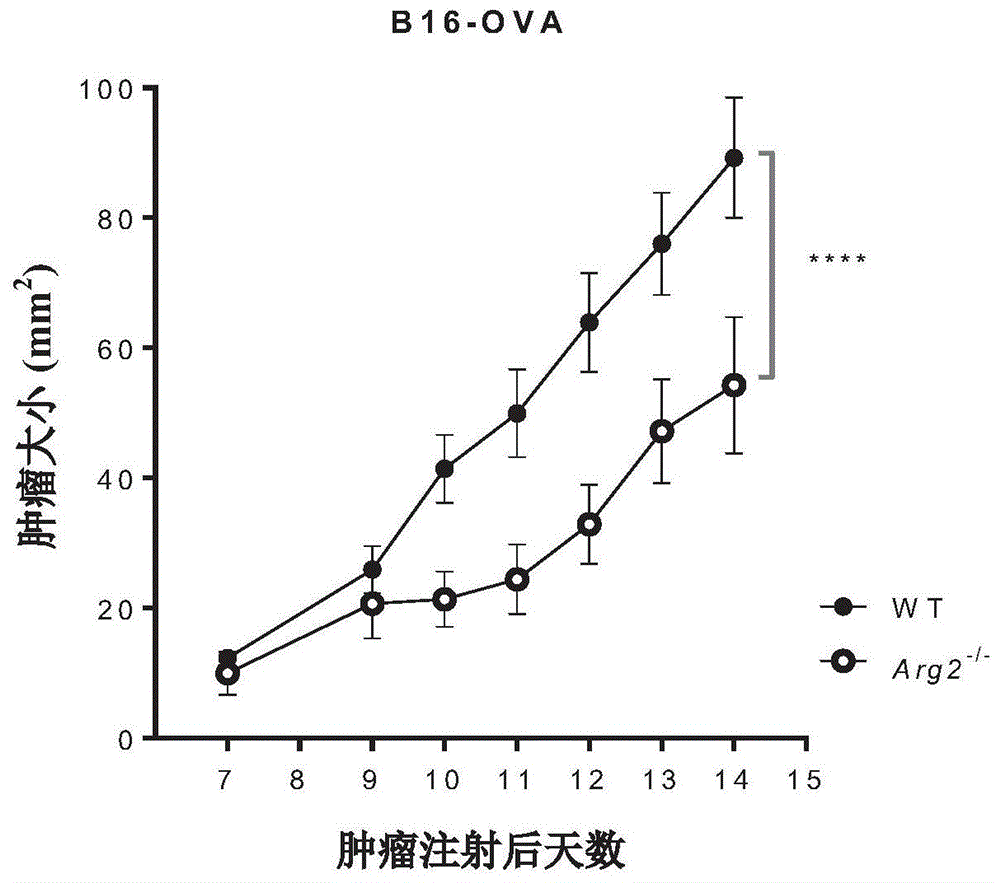
图1a和1b示出了在植入b16-ova或mc38-ova肿瘤细胞的wt和arg2-/-小鼠中肿瘤的生长情况。这些图示出了arg2-/-小鼠的肿瘤生长受损。
图2a和2b示出了在如图1a和1b所述小鼠体内的抗肿瘤免疫力。这些图示出了在arg2-/-小鼠中抗肿瘤免疫力增强。
图3a示出了在wt(圆形)和arg2-/-(方形)小鼠中mc38-ova肿瘤的生长,这些小鼠消耗了(空心圆形和方形)或未消耗(实心圆形和方形)cd8+t细胞。在这些实验中,cd8+细胞消耗使肿瘤生长增加。
图3b示出了图3a中所述的消耗cd8+细胞的小鼠的动物生存率降低。
图4a、4b和4c分别示出了接受(图4a:空心圆形和方形)或不接受(图4a:实心圆形和方形)抗pd1或同种型对照ab的wt(图4a:圆形)和arg2-/-(图4a:方形)小鼠的mc38-ova肿瘤生长、肿瘤清除率和存活率。结果显示抗pd1ab治疗和arg2缺陷对肿瘤大小(图4a:空心方形),肿瘤清除率和存活率具有强联合作用。
图5示出了在嵌合小鼠中mc38-ova肿瘤的生长,该小鼠是通过用全部四种配对联合使用的源自wt或arg2-/-小鼠的骨髓(bm)细胞重建亚致死剂量照射的wt或arg2-/-小鼠而产生的。结果表明减少肿瘤生长主要归因于源自bm的细胞中的arg2缺陷。因此,接受来自arg2-/-小鼠的bm细胞的小鼠(空心圆形和方形)的肿瘤大小较小。
图6a和6b分别比较了从otiwt小鼠和双重纯合子arg2-/-oti小鼠分离的t细胞的体外活化(图6a)和增殖(图6b)。结果表明,与arg2+/+otit细胞相比,arg2-/-otit细胞表现出增强的活化和增殖。
图7a示出了根据本发明的实施例的用于细胞疗法的体内系统的产生。
图7b示出了按图7a所示处理的小鼠中的肿瘤生长。接受arg2-/-otit细胞的wt小鼠表现出明显较慢的肿瘤生长。
图7c示出了如图7a所示处理的小鼠的动物存活率。接受arg2-/-otit细胞的wt小鼠表现出明显增加的存活率。
图8a、8b和8c示出了在携带mc38-ova的wt受体中,arg2-缺陷的过继转移cd8+t细胞产生更多ifn(图8a),耗竭变少(图8b)并持续更长的时间(图8c)。
图9a、9b和9c分别示出了在通过i.p.注射t细胞转移(箭头)后第8、11和14天,接受wtotit细胞(图9a:圆形)或arg2-缺陷的otit细胞(图9a:方形)以及接受(图9a:实心圆形和方形)或未接受(图9a:空心圆形和方形)抗pd1或同种型对照ab的wt小鼠中,在d60时mc-38-ova的肿瘤体积、存活率和肿瘤清除率。结果显示抗pd1ab治疗和过继cd8+t细胞中arg2缺陷对肿瘤体积、存活率和肿瘤清除率的强联合作用。
图10a和10b示出了arg抑制在体外增加人t细胞活化。从pbmc纯化的人t细胞未经处理(-)或在体外用抗cd3和抗cd28ab(+)活化,在存在或不存在arg抑制剂的情况下在96孔板的rpmi中培养,并评估其活化24h后的活化情况(cd69染色)。这些图示出了在存在指示的arg抑制剂的情况下,cd4+(图10a)和cd8+t细胞(图10b)中cd69+细胞的频率增加。
在下文中,为了说明本发明而描述了本发明的优选实施例,但并不旨在限制本发明的范围。
具体实施方式
在一些方面,本发明涉及具有受损的精氨酸酶活性和/或表达的免疫细胞,用于治疗癌症。所述细胞优选用于癌症免疫治疗。
表述“受损的精氨酸酶活性和/或表达”中的术语“受损的”是指与未处理、修饰和/或工程改造的相应野生型免疫细胞相比具有降低的精氨酸酶活性和/或表达,例如,如本说明书中所公开的,以使所述精氨酸酶活性和/或表达受损。在优选的实施例中,使所述精氨酸酶活性和/或表达受损至无法检测到精氨酸酶活性和/或表达的程度。优选地,所述精氨酸酶活性完全不存在。
表述“受损的精氨酸酶活性和/或表达”涵盖其中由于编码精氨酸酶的mrna转录和/或翻译受损而使精氨酸酶活性降低的情况。此外,尽管在本说明书中经常同时提及受损的精氨酸酶“活性”和/或“表达”,但应理解的是,精氨酸酶表达受损导致精氨酸酶活性受损,因此表达“受损的精氨酸酶活性”涵盖了由于精氨酸酶表达降低而使活性受损的情况。在优选的实施例中,免疫细胞产生活性精氨酸酶的能力受损,优选地通过对免疫细胞的基因表达过程的技术干扰而受损,所述过程包括诸如转录、rna剪接、翻译、翻译后修饰的过程,特别是一种或多种编码精氨酸酶的基因。
所述精氨酸酶可以是精氨酸酶1(arg1)和/或精氨酸酶2(arg2)。在优选的实施例中,所述受损的精氨酸酶活性和/或表达是受损的精氨酸酶2(arg2)活性和/或表达。如上所述,arg1和arg2在细胞内定位和表达方式方面有所差异。arg2位于线粒体中,并在多种组织中广泛表达。在实施例中,arg1和arg2的活性均受损。
在优选的实施例中,所述免疫细胞选自t细胞、til、天然杀伤细胞(nk细胞)、先天性淋巴样细胞(ilc)和树突细胞。例如,所述ilc可以选自ilc-1和ilc-2细胞。在优选的实施例中,所述免疫细胞是t细胞,其优选选自cd3+和/或cd4+和/或cd8+t细胞。也可以使用包含两种或更多种不同类型的具有受损的精氨酸酶活性和/或表达的免疫细胞的组合。
在实施例中,所述免疫细胞是树突细胞。在可选的实施例中,所述免疫细胞不是树突细胞和/或不包括树突细胞。
为了本说明书的目的,术语“包括”及其各种语法形式旨在表示“包括其中其他”。其并非旨在表示“仅由……组成”。
在优选的实施例中,向有需要的受试者,特别是患有癌症的受试者,施用免疫细胞。在实施例中,本发明涉及用于治疗癌症的细胞疗法。优选地,所述免疫细胞通过过继细胞转移(act)或在act治疗的框架中施用。act是癌症免疫疗法,诸如自体癌症免疫疗法。术语“过继”通常被理解为表示act是将细胞转移至患者。例如,这些细胞可能源自患者或另一个体。在优选的实施例中,act是指通常通过基因上的和/或在表达蛋白方面已经选择、工程改造和/或修饰的细胞的转移,从而使细胞更有效或易于治疗所考虑的病症,通常是癌症。
在优选的实施例中,所述免疫细胞是分离和/或纯化的。
所述免疫细胞可以源自待治疗的患者个体。在这种情况下,是指自体免疫疗法。换句话说,改善患者免疫细胞的功能和特性,以提高其抵抗患者所患癌症的能力。
在实施例中,所述免疫细胞获自患者的肿瘤。以这种方式可以获得til。
在另一实施例中,免疫细胞取自供体。在这种情况下,一种情况可以参考同种异体免疫疗法。
在又另一实施例中,所述免疫细胞获自干细胞和/或前体免疫细胞。
当免疫细胞源自患者和/或供体时,本发明的方法可以包括从所述患者或所述供体,特别是从患者或供体的血液,或例如,如果在til的情况下,则是供体的肿瘤,收集和/或提取所述免疫细胞。
在另一实施例中,所述细胞预先从个体(例如,待治疗的受试者或供体)中收集和/或提取。
如果免疫细胞源自供体,则优选地进行选择或修饰,以免攻击患者的健康组织。例如,供体的免疫细胞优选与患者相容。例如,供体可以是患者的家庭成员。
在一些实施例中,包括供体是否是家族成员,供体的细胞具有失活的天然t细胞受体(在免疫细胞是t细胞的情况下),从而防止免疫细胞攻击健康的患者组织。例如,优选对细胞进行修饰和/或工程改造以不表达天然t细胞受体或不表达其无活性形式。
如所述,在一些实施例中,免疫细胞可以源自干细胞,例如,多能干细胞。因此,免疫细胞优选从现成的细胞制备,并根据本说明书进一步工程改造。例如,可以基于人工胸腺类器官(ato)系统获得免疫细胞。ato系统是人工模仿胸腺环境以概括人t细胞发育的体外模型。ato系统支持使用各种来源的造血干细胞,以及多能干细胞,如胚胎干细胞和诱导性多能干细胞,对正常t细胞进行有效分化和阳性选择。例如,该技术还为进一步的基因工程改造提供了灵活性,以产生用于治疗用途的现成异基因工程改造的t细胞产品。例如,在wo2016/187459和wo2017/075389中公开了这种技术。
根据本发明,免疫细胞的精氨酸酶活性和/或表达受损。所述免疫细胞中的精氨酸酶活性和/或表达可能以任何合适的方式受损。在实施例中,通过将所述免疫细胞暴露于用于损害所述精氨酸酶活性和/或表达的治疗和/或工程改造来损害所述精氨酸酶活性。优选地,所述处理是离体处理。例如,一旦从个体(诸如患者或供体)中提取细胞,则将细胞暴露于处理。
在一些实施例中,本发明包括将细胞暴露于精氨酸酶抑制剂,从而直接阻断精氨酸酶。例如,抑制剂可以例如通过与蛋白的相关部分(诸如活性位点)共价应答来永久灭活精氨酸酶。精氨酸酶抑制剂先前已经公开,例如,在本申请的引言中引用的文献中。
优选地,通过基因手段和/或在基因或其表达的水平上修饰所述细胞,以在基因上防止精氨酸酶表达。
在优选的实施例中,所述精氨酸酶活性通过损害所述精氨酸酶的表达而使所述精氨酸酶活性受损,例如,通过
-突变、截短或删除编码所述精氨酸酶的基因,
-施用、突变、截短或删除编码转录因子的基因,所述转录因子是所述编码精氨酸酶的基因的转录因子,
-施用对能够与编码所述精氨酸酶的mrna结合的核苷酸序列进行编码的核苷酸序列,或施用将能够与编码所述精氨酸酶的mrna结合的核苷酸序列包括在内的核苷酸序列(administeringanucleotidesequenceencodingorcomprisinganucleotidesequencecapableofbindingtoanmrnaencodingsaidarginase)。例如,该步骤可以包括施用核酸分子,所述核酸分子包括对编码所述精氨酸酶的mrna进行编码的核苷酸序列,或所述核酸分子包括与所述mrna结合的核苷酸序列(forexample,thisstepmaycompriseadministratinganucleicacidmoleculecomprisinganucleotidesequenceencodinganmrnaencodingsaidarginaseorbindingtosaidmrna)。
为了本说明书的目的,术语“突变(mutation)”及其各种语法形式(诸如“突变(mutating)”)涵盖基因的截短和缺失。例如,术语“突变”还包括点突变、点缺失(基因编码区中单个核苷酸的缺失)以及基因编码区中一段核苷酸的插入或缺失。因此,更通常第,术语“突变”涵盖导致基因表达与不携带突变的细胞中的表达不同的任何基因改变。仅出于清楚的原因,提到了某些突变可能未导致基因表达发生改变,并且此类沉默突变通常不被认为是适合于损害精氨酸酶表达的突变。
例如,可以通过删除对精氨酸酶、特别是精氨酸酶2进行编码的一个或多个基因来修饰细胞。例如,可以对细胞进行处理以使其相对于精氨酸酶1和/或2(arg1-/-和/或arg2-/-)双敲除。例如,可以通过定点突变进行编码精氨酸酶的基因或其他与精氨酸酶表达的控制有关的基因(诸如转录因子)中的突变。用于定点突变可用的方法很多,诸如kunkel法、盒突变或pcr定点突变。在实施例中,可以通过基因编辑的方式,例如,通过使用crispr/cas9技术,来修饰精氨酸酶编码基因或转录因子。
精氨酸酶表达可以通过促进精氨酸酶基因的阻遏蛋白的表达而受损,例如,通过用构建为在插入细胞时产生这种阻遏蛋白的载体转染免疫细胞,或通过基因编辑技术插入编码合适阻遏蛋白的基因。
在优选的实施例中,所述精氨酸酶表达通过rna干扰(rnai)降低。在实施例中,所述受损的精氨酸酶活性是由于在所述免疫细胞中施用能够与编码所述精氨酸酶的mrna结合的核酸分子,或施用编码此类核酸分子的载体。所述核酸分子或所述载体的施用优选为离体施用,即在分离的细胞,例如,从个体提取的细胞中。
在实施例中,本发明的方法包括施用或转录抑制基因表达或中和精氨酸酶mrna的rna。例如,可以将小干扰rna(sirna)、微小rna(mirna)或短发夹rna(shrna)施用于细胞或编码选自此类干扰rna的任一种的载体。这些分子可以通过合适的递送系统和/或载体递送至细胞中,所述递送系统和/或载体能够在细胞内递送干扰rna,或者当转移至细胞中时能够转录此类干扰rna。
在实施例中,本发明的方法包括施用或转录特异性抑制基因表达或中和精氨酸酶mrna的核酸分子,诸如rna。例如,可以将特别定制的小干扰rna(sirna)、微小rna(mirna)或短发夹rna(shrna)施用于细胞,或编码选自这种抑制性rna的任一种的载体。这些分子可以通过合适的递送系统和/或载体递送至细胞中,所述递送系统和/或载体能够在细胞内递送抑制性rna,或者当转移至细胞中时能够转录这种抑制性rna。
在实施例中,通过仅将干扰和/或抑制核酸分子与编码精氨酸酶的mrna互补碱基配对,而不与编码其他蛋白的其他mrna分子互补碱基配对,和/或仅与调节精氨酸酶表达的rna分子互补配对,而不与调节其他蛋白表达的rna分子互补碱基配对,特别是在所述免疫细胞中,而提供特异性。
在实施例中,所述抑制和/或干扰核酸分子,优选为rna,不是microrna-155和/或不包含microrna-155。
在其他实施例中,例如,在非特异性抑制和/或干扰精氨酸酶表达和/或精氨酸酶mrna的中和的情况下,所述抑制和/或干扰核酸分子可以包括和/或基本上由microrna-155组成。
在优选的实施例中,使用在细胞中表达和/或转录抑制性和/或定向干扰rna的载体。此类载体允许长期基因特异性沉默。此类载体,例如,用于使精氨酸酶1或精氨酸酶2沉默的载体,可购自例如santacruzbiotechnology,inc.,usa,该公司提供了完整的rnai基因沉默产品线,包括sirna、shrna质粒和shrna慢病毒产品。因此,可以施用编码干扰rna的质粒或载体,或者可以例如使用合适的递送载体将干扰rna定向引入细胞中。根据另一实例,病毒颗粒可以用于施用shrna编码质粒。对人精氨酸酶2mrna具有特异性的示例性sirna分子的序列可以源自精氨酸酶2的rna或dna序列,例如,genbank中以登录号nm_001172所公开。sirna分子还如settyba,etal.hypoxicproliferationofosteosarcomacellsdependsonarginaseii,cell.physiol.biochem.39(2),802-813(2016)所公开。
根据本发明,精氨酸酶活性和/或表达的降低可以是组成性的或诱导性的。在一些实施例中,精氨酸酶活性的降低是诱导性的,即,可以由可以控制的外部因素引发,例如,由进行过继细胞转移的细胞修饰的工作人员进行控制。精氨酸酶表达的降低可以通过合适启动子的存在而被诱导,例如,在编码干扰rna的载体上。由于启动子,基因沉默仅在启动子活化后发生。例如,可以选择启动子以使其可以被特定的小分子活化,所述小分子可以单独施用于细胞或接受细胞的患者。
另一方面,通过使用具有组成性活性的启动子,沉默rna可以组成性表达,导致精氨酸酶表达的组成性受损。
此外,精氨酸酶活性的降低可以是瞬时的或稳定的。精氨酸酶表达的瞬时或稳定损伤可以通过选择干扰rna和细胞处理方式确定。例如,通过用适当的sirna直接转染细胞,精氨酸酶的表达可以瞬时沉默。通过在启动子的控制下用编码干扰rna的载体转染,精氨酸酶的表达可以在细胞中稳定沉默(以诱导性或组成性的方式)。
在一些实施例中,免疫细胞进一步包括用于治疗癌症的其他有利特征和/或功能。优选地,将细胞进一步修饰以具有改善的癌症治疗功能。免疫细胞的进一步修饰可以与上述精氨酸酶活性受损同时进行,或者可以使用例如相同的免疫细胞在单独的、先前的或后续的步骤中进行。本发明还涵盖独立地基因改造其他免疫细胞以具有改善的癌症治疗功能,其中在根据本发明的治疗中,将这些其他细胞与本发明的免疫细胞同时或顺序施用。
在实施例中,所述免疫细胞是肿瘤浸润淋巴细胞(til)和/或所述方法包括施用til。优选地,所述til是肿瘤浸润t细胞。优选地,所述til表现出组成性或诱导性精氨酸酶活性受损。优选地,向有需要的受试者(特别是患有癌症的受试者)施用所述til。优选地,所述til预先从所述受试者中分离和/或纯化。优选地,所述方法包括损害所述til中的精氨酸酶活性,优选地离体进行,例如,在分离和/或纯化所述til之后和/或在施用所述til之前。
在实施例中,免疫细胞包含和/或表达嵌合抗原受体(car)和/或转基因t细胞受体(tcr)。在实施例中,所述car包括经接头与t细胞信号传导结构域融合的抗原结合结构域,特别是cd3ζ(ζ)信号传导结构域。抗原结合结构域优选包括抗体的一个或多个可变结构域。优选地,所述接头包括跨膜结构域。当在car细胞的表面表达时,抗原结合结构域在细胞的细胞外侧,而信号传导结构域在细胞的细胞内侧。
根据car的设计和功能,可以选择合适的细胞内结构域。如果选择细胞外结构域以识别(结合)癌细胞上的靶标,则优选地选择细胞内结构域以在结合细胞外结构域时活化免疫细胞。通常,在cart细胞中,例如,car的结合通常导致t细胞增殖和细胞因子的产生。cart细胞与靶位点(诸如cd19)的结合可以定向导致癌细胞被杀死,例如,通过诱导癌细胞凋亡。因此,通过选择适当的细胞内结构域来控制从细胞外结构域的结合获得的特异性作用。通常,car免疫细胞包括信号传导结构域(包括cd3ζ)和任选的其他信号传导结构域。然而,本发明不限于免疫细胞可以表达的car的胞内或胞外结构域。本领域技术人员可以根据待靶向的特定癌细胞以及希望通过结合引发的免疫应答来选择合适的结构域。
例如,抗原结合结构域可以对b细胞抗原cd19具有特异性。当前针对血液癌症(诸如白血病和淋巴瘤)的细胞转移疗法基于cart细胞,其中car的抗体可变结构域对cd19具有特异性。
根据待治疗的癌症,car的抗体可变结构域优选对癌细胞表面表达的靶标具有特异性。在本领域中,针对通过car疗法治疗相应的癌症,已经检测了以下抗原结合结构域特异性。对以下项具有特异性的抗原结合结构域:碳酸酐酶,特别是用于抗肾癌的car细胞疗法;表皮生长因子受体,特别是用于治疗胶质母细胞瘤的变体egfrviiicar;抗前列腺癌的car治疗中的前列腺特异性抗原(psma)和/或前列腺干细胞抗原(psca);治疗选自卵巢癌、输卵管癌和原发性腹膜癌中任一项的卵巢肿瘤抗原粘蛋白16(muc16)。
根据本发明的实施例,免疫细胞优选经工程改造以表达一种或多种car,诸如具有选自上述抗原的组的特异性的car。
在实施例中,免疫细胞表达car和/或所述方法包括施用表达car的免疫细胞。优选地,所述表达car的细胞表现出组成性或诱导性精氨酸酶活性受损。优选地,向有需要的受试者(特别是患有癌症的受试者)施用表达car的细胞。优选地,预先从所述受试者分离和/或纯化的所述免疫细胞和所述方法包括处理所述细胞的步骤,以表达所述car和/或提供预先从患者或其他供体分离的细胞,并且所述细胞预先经处理以表达所述car。在实施例中,所述方法包括优选损害所述car表达细胞中的精氨酸酶活性,优选离体进行。
根据本发明所涵盖的免疫细胞的另一种修饰是内部信号传导结构域的转换。
在实施例中,免疫细胞包括和/或表达il-4受体,其经修饰或失活以减少、消除和/或逆转由肿瘤微环境引发的抑制性il-4信号传导。例如,可以将il-4受体的信号传导结构域转换为il-7受体的信号传导结构域,以逆转抑制性il-4信号传导。
在实施例中,免疫细胞表达两种car。第一car优选免疫细胞靶向特定癌细胞,具有定向于癌细胞表达的抗原(诸如psca)的抗原结合结构域。第二car可以例如与合成或外源化合物(例如,小分子)结合。所述化合物可以单独和/或与car细胞一起向患者施用。当施用时,小分子与第二car结合。当免疫细胞与靶细胞接触时,与化合物的共同刺激导致免疫细胞的有效和/或强活化。共表达两种car的t细胞,与rimiducid(具有同型二聚活性的脂质渗透性他克莫司类似物)的第二结合,目前在进行i期试验。
在实施例中,免疫细胞经工程改造以表达失活的检查点蛋白或缺乏抑制性检查点蛋白。示例性免疫检查点调节剂是pd-l1/pd1、ctla4、b7-h3(cd276)、b7-h4(b7x/b7s1/vtcn1)、hhla2(b7h7/b7-h5)、vista(pd1h、dd1alpha、c10orf54、gi24、dies1、sisp1)、vsig、lag-3、tigit、cd96、cd39、cd73、腺苷a2受体、cd47、嗜乳脂蛋白(btn)和/或tim-3(t细胞-免疫球蛋白-粘蛋白结构域)。
在免疫细胞的实施例中,检查点蛋白的免疫应答抑制信号传导结构域被失活或与共刺激结构域交换(取代)。例如,可以使pd1的抑制性cd28结构域(程序性细胞死亡蛋白1)突变,使其失活或可以用共刺激结构域(例如,cd3ζ信号传导结构域或cd137结构域)进行转变。
通过使免疫细胞的免疫检查点蛋白失活或通过将细胞内结构域转换为刺激性结构域(例如,通过取代细胞内结构域),可以避免肿瘤细胞和/或肿瘤微环境的免疫抑制活性。
在实施例中,免疫细胞重组表达当被外部因素引发时能够诱导细胞凋亡的蛋白。这可以看作是一种安全性措施,例如,在向患者施用免疫细胞后,如果发现非预期的副作用,则可以靶向方式破坏本发明的免疫细胞。例如,可以以与细胞外结构域结合导致细胞凋亡的方式提供信号传导结构域。如本说明书其他地方所述,细胞外结构域可以结合人工化合物,诸如小分子。
通过用合适的载体转染细胞,或在基因组中插入用于表达受体的基因和/或启动子,例如,使用诸如crispr/cas9技术的基因编辑,可以对免疫细胞进行工程改造以表达car和/或转基因t细胞受体。
为了治疗癌症,优选地向个体施用免疫细胞,例如,向需要癌症治疗的个体施用。所述个体优选是患有癌症的患者。可以以任何合适的方式,优选地肠胃外施用免疫细胞。在优选的实施例中,静脉内施用免疫细胞。
在优选的实施例中,除了另一种癌症治疗之外,还施用免疫细胞。优选地,免疫细胞与另一种癌症治疗联合使用。该另一种癌症治疗可以同时和/或单独施用。此外,该另一种癌症治疗可以以单独的组合物的形式单独施用,或者可以以单一组合物联合施用。
在优选的实施例中,使用免疫细胞和/或与靶向和/或特异性结合负性免疫检查点调节剂的癌症治疗联合施用。免疫检查点调节剂已在本说明书的其他地方公开。在一个实施例中,免疫细胞可以表达与可以由癌细胞表达的免疫检查点调节剂结合的细胞表面蛋白,例如,受体。例如,如上所述,免疫细胞可以表达与免疫检查点调节蛋白结合的car。根据该实施例,阻断免疫检查点调节剂的实体可以以免疫细胞上的细胞表面蛋白的形式表达,而不是未结合/游离的抗体。
在另一实施例中,所述(其他或另外的)癌症治疗包括抗癌剂和/或分子,例如,免疫检查点调节剂抑制剂。优选地,所述癌症治疗包括抗体,优选地与免疫检查点调节剂特异性结合的抗体。在优选的实施例中,所述癌症治疗包括与一种或多种选自以下项的组特异性结合的抗体:pd1、pd-l1、ctla4、b7-h3、b7-h4、hhla2、vista、vsig、lag-3、tigit、cd96、cd39、cd73、腺苷a2受体、cd47、嗜乳脂蛋白(btn)和/或tim-3。
特异性结合免疫检查点调节剂的抗体,诸如纳武利尤单抗和帕博利珠单抗,二者均特异性结合pd-1,可商购获得,并用于治疗多种不同的癌症。
此类抗体通常可称为免疫检查点调节剂。本发明人已观察到,当本发明的精氨酸酶受损的免疫细胞与包括免疫检查点调节剂抑制剂的治疗联合使用时,产生协同的抗癌活性。在实施例中,抗体特异性结合由癌细胞表达并在癌细胞表面上提供的免疫检查点调节蛋白。在另一实施例中,抗体特异性结合蛋白,例如,本发明的免疫细胞表面表达的受体,其中,所述抗体阻止免疫细胞的蛋白与癌细胞上表达的相应蛋白质接触。在优选的实施例中,免疫检查点调节剂抑制剂是单克隆抗体。
在优选的实施例中,免疫细胞用于治疗和/或预防癌症的方法。在实施例中,选择用于治疗的癌症选自包括白血病、淋巴瘤和/或实体瘤的组。本发明人的一个惊人的发现是免疫细胞不仅在减少血液癌症和/或血液学癌症方面有效,而且在减小实体瘤的大小和/或体积方面特别有效。这是令人惊讶的,因为当前使用的基于cart细胞的过继细胞转移疗法被用于治疗血液癌症,通常是由于难以获得针对实体瘤的足够疗效。在当前的cart细胞疗法中,实体瘤的治疗有时伴随着重大的副作用和毒性问题。本发明人公开了在不存在毒性副作用的情况下通过过继细胞转移治疗实体瘤。
本发明涉及几种方法,包括治疗和/或预防癌症的方法、免疫治疗方法、细胞治疗方法、改善现有免疫治疗的方法、产生可用于治疗和预防癌症的免疫细胞的方法、用于准备和/或产生癌症治疗的方法,和/或改善免疫细胞的抗癌活性和/或疗效的方法。如本说明书中详述,优选地将免疫细胞处理为具有改善的抗癌活性。更具体地,细胞优选在由癌细胞(包括实体瘤细胞)产生的免疫抑制环境中具有增加的活性和/或存活率。优选地,免疫细胞通过在某种程度上对癌细胞的至少一些免疫抑制活性保持惰性来保持其抗癌活性和/或功能。
优选地,对免疫细胞进行处理和/或工程改造以表现出改善的抗癌活性。如本说明书中详述,所述处理优选导致精氨酸酶活性降低。细胞的处理优选离体地和/或在体外进行,即,优选在人或动物体外。在实施例中,本发明的方法包括从个体提取和/或收集尚未处理或野生型的免疫细胞。在实施例中,本发明的方法包括在施用所述免疫细胞之前,离体损害所述免疫细胞中的精氨酸酶活性和/或表达。在实施例中,所述方法包括向患者施用免疫细胞。
如本说明书其他地方所公开的,免疫细胞可以源自待治疗的患者、供体,或者可以源自培养物,例如,获自干细胞。
免疫细胞优选以药物组合物的形式提供。优选地,所述组合物包括合适的辅料和/或载体,例如,选自其中悬浮细胞的近似生理和/或等渗溶液的溶液,例如,生理盐水。在一些实施例中,药物组合物可以包括血清,例如,从待治疗的患者或从供体获得。
本发明还提供了用于治疗癌症方法的试剂盒。所述试剂盒优选包括适合于损害免疫细胞中的精氨酸酶活性和/或表达的制剂。在一些实施例中,所述试剂盒包括当施用于免疫细胞时能够诱导rna干扰的试剂,其中所述rnai导致精氨酸酶表达受损。例如,所述试剂盒包括一种或多种如本说明书其他地方公开的试剂(sirna载体等)。在一些实施例中,所述试剂盒包括免疫细胞,例如,精氨酸酶活性受损的现成免疫细胞,例如,arg2-/-免疫细胞。
尽管以上已经描述并具体示例性说明了本发明的某些优选实施例,但并不表示本发明限于这些实施例。如所附权利要求书所述,在不背离本发明的范围和精神的情况下可以对其进行各种修改。在下文中,公开了本发明的实例。这些实例仅用于说明,而并不旨在限制本发明的范围。
实例
实例1:在arg2-/-小鼠中肿瘤生长受损和抗肿瘤免疫力增强
对于本实例,使用从charlesriverlaboratories,inc.获得的arg2缺陷型(双基因敲除)小鼠。作为第一种方法,比较了wt和arg2-/-小鼠的肿瘤生长。使用两种可移植的肿瘤模型:b16黑色素瘤模型和mc38结肠癌模型。在这两个模型中,均使用表达卵清蛋白(ova)作为替代肿瘤抗原的肿瘤变体。
在第一个实验中,皮下注射0.5x106个b16-ova或mc38-ova细胞至wt或arg2-/-小鼠的背部,并监测肿瘤生长2周。n=9,数据来自两个独立实验的合并。
在第二个实验中,按照第一个实验植入肿瘤细胞。肿瘤注射后11(b16-ova)或13(mc38-ova)天,i.v.注射高剂量的细胞紫色示踪剂(ctvhi)ova脉冲和低剂量(ctvlo)非脉冲的wt脾细胞进行荧光标记。二十四小时后,切除肿瘤并通过流式细胞术分析来自tdln和ndln的细胞悬浮液。特定的体内杀伤计算如下:[1-(%ctvhitdln/%ctvlotdln)/(%ctvhindln/%ctvlotdln)]x100。
结果显示在图1a、1b,2a和2b中(*,p<0.05;**,p<0.01;****,p<0.0001)。如图1a和1b所示,在arg2-/-小鼠中,b16-ova和mc38-ova肿瘤的生长均显著受损。肿瘤生长受损与肿瘤引流淋巴结中体内ova特异性肿瘤细胞杀伤的增加有关(图2a和2b)。对于b16-ova,显示了两个实验的代表。对于mc38-ova,数据来自两个独立的实验。
为了进一步研究,我们主要关注mc38-ova模型。
实例2:cd8+t细胞对控制肿瘤生长和动物存活的贡献
为了确定arg2-/-小鼠中mc38-ova生长受损是否可能是由于cd8+t细胞的增强控制,我们进行了cd8+t细胞消耗实验。
s.c.注射0.5x106个mc38-ova细胞至wt或arg2-/-小鼠的背部,并监测肿瘤生长和动物存活4周。cd8+t细胞消耗通过数次i.p.注射抗cd8a+消耗抗体(αcd8a)或igg2a同种型对照抗体(αigg2a)。
如图3a和3b所示,在arg2-/-小鼠中,肿瘤生长降低和动物存活率增加通过cd8+t细胞的消耗而显著恢复,表明cd8+t细胞介导的免疫控制具有重要作用。但是,抗肿瘤cd8+t细胞应答并不是唯一起作用的机制,因为cd8+t细胞缺失的arg2-/-小鼠的肿瘤生长并未完全恢复到cd8+t细胞缺失的wt小鼠中观察到的结果。
实例3:抗pd1治疗和arg2缺陷对肿瘤生长抑制和动物存活的协同作用
mc38肿瘤对使用可阻断t细胞抑制性pdl1-pd1检查点轴的抗体的免疫疗法敏感。为了确定由pdl1-pd1阻断诱导的对mc38肿瘤控制增强是否可以与由arg2缺陷导致的对mc38肿瘤控制增强的机制协同,mc38-ova荷瘤的wt和arg2-/-小鼠接受抗pd1抗体治疗。
s.c.注射0.5x106个mc38-ova细胞至wt或arg2-/-小鼠的背部。肿瘤注射后第9、11和14天,向小鼠注射抗pd1(αpd-1)ab或igg2a同型对照ab(αigg2a)(绿色箭头)。
从图4a中可以看出(数据来自2个实验的合并,****,p<0.0001),接受抗pd1抗体治疗的wt小鼠中的肿瘤生长降低到与未接受治疗的arg2-/-小鼠中的肿瘤生长降低相同的程度。重要的是,接受抗pd1抗体治疗arg2-/-小鼠导致肿瘤生长几乎完全消除。实际上,许多小鼠中的肿瘤已经清除,如图4b所示。图4c示出了在接受抗pd1抗体治疗的arg2-/-小鼠中动物存活大大增加。因此,抗pd1治疗和arg2缺陷对肿瘤生长表现出很强的协同作用。
实例4:bm来源的细胞中的arg2缺陷可改善对肿瘤生长的控制
产生相互的骨髓(bm)嵌合小鼠,以确定是否受损的mc38-ova生长是bm来源细胞或非造血来源细胞中arg2缺陷的结果。wt和arg2-/-小鼠受亚致死性照射以破坏宿主bm。然后以所有四种配对组合,通过移植来自wt或arg2-/-小鼠的bm细胞重建造血。
s.c.注射0.5x106个mc38-ova细胞至bm嵌合小鼠的背部,并监测肿瘤生长4周。n=11只小鼠。
从图5可以看出(数据抽取自3个独立实验的合并,****,p<0.0001),与接受来自wt小鼠的bm细胞相比(wt>wt和wt>arg2-/-),接受来自arg2-/-小鼠(arg2-/->wt和arg2-/->arg2-/-)的bm细胞的嵌合体表现出强降低肿瘤生长。这些结果表明,肿瘤生长的降低主要归因于bm来源的细胞中的arg2缺陷。
实例5:arg2-/-oti细胞在体外表现出活化和增殖增强
为了确定ccd8+t细胞中arg2缺陷是否可能导致对肿瘤生长的控制改善,我们将arg2突变进入oti小鼠中以获得双纯合的arg2-/-oti小鼠。
oti小鼠仅具有ova特异性cd8+t细胞,因为它们表达编码mhci类限制性ova特异性tcr的转基因。这使我们能够比较aarg2+/+和arg2-/-ova特异性t细胞的功能特性。
从杂交小鼠中分离出wtoti或arg2-/-otit细胞,并在体外用抗cd3和抗cd28ab活化,在96孔板的rpmi中培养,并在60℃下评估其活化后的第1、2和3天的活化(cd69染色)。在活化后第4天测定t细胞增殖(羧荧光素琥珀酰亚胺酯(cfse)稀释)。
结果显示在图6a和6b中(数据代表2次独立实验,**,p<0.01;***,p<0.001)。
图6a和6b所示的体外t细胞活化试验表明,与arg2+/+oti细胞相比,arg2-/-oti细胞表现出活化和增殖增强。
实例6:mc38-ova肿瘤的otit细胞疗法
开发了一种体内系统,以比较wt背景下arg2+/+和arg2-/-oti细胞对mc38-ova肿瘤生长和动物存活的控制。
体内系统的产生在图7a中示出。首先,产生混合的bm嵌合体,其中用rag2-/-小鼠来源的bm和arg2+/+或arg2-/-oti小鼠来源的bm细胞的9:1混合物重建受辐照的wt小鼠。在这些混合的bm嵌合体中,arg2-/-和arg2+/+otit细胞在表现出正常arg2表达的环境中产生,因此其功能特性的任何差异均可归因于arg2表达的细胞内在差异。应当注意,源自arg2-/-和arg2+/+otibm的b细胞在arg2状态方面当然也会有所不同,但这不太可能产生任何影响,因为检查immgen联合数据表明b细胞不表达arg2mrna。
混合的bm嵌合体用作arg2+/+和arg2-/-otit细胞的供体,这些细胞被过继转移至mc38-ova荷瘤的wt小鼠中。更具体地,将106个脾和淋巴结arg2+/+或arg2-/-otit细胞从混合的bm嵌合体转移到5天前已注射0.5x106个mc38-ova肿瘤细胞的wt受体中。
然后在otit细胞转移后一天,用cpg-b+ova1肽免疫荷瘤受体,并监测肿瘤的生长。
结果显示在图7b和7c中。与未接受oti细胞或arg2+/+oti细胞的小鼠相比,接受arg2-/-otit细胞的小鼠的肿瘤生长明显降低(图7b),动物存活率提高(图7c)。该实验设置正式证明arg2-/-otit细胞比arg2+/+oti细胞在wt环境中控制mc38-ova肿瘤的生长方面更好。
实例7:arg2-/-oti细胞在mc38-ova荷瘤动物中表现出ifn产生增加,消耗较少且持续时间较长
为了进一步研究arg2缺陷对体内cd8+t细胞的效应子功能的影响,将相等数量的原始oti和arg2-/-oticd8+t细胞转移至mc38-ova荷瘤的wt小鼠中。oti转移后一天,然后用cpg-b+ova-1肽免疫荷瘤受体,免疫后7天,引流ln(dln)中的oti细胞和流式细胞仪分析肿瘤。在dln和肿瘤中,arg2-/-oti细胞中ifn+细胞的频率均较高(图8a)。在肿瘤中的arg2-/-oti细胞上,pd-1表达水平显著降低(图8b)。
为了研究arg2缺陷对肿瘤特异性t细胞应答的时间和空间动态的影响,将相等数量的原始oti和arg2-/-oticd8+t细胞转移到mc38-ova荷瘤小鼠中,在ova257–264免疫后的不同时间,通过流式细胞术评估dln和肿瘤中它们在宿主中的分布。cd45.1标志物用于区分oti(cd45.1+/+)和arg2-/-oti(cd45.1+/-)细胞。至第15天,在dln和肿瘤中,arg2-/-oti细胞的频率明显高于otiwt细胞(图8c),表明它们具有更持久的抗肿瘤应答。
实例8:t细胞固有的arg2缺陷与pd-1阻断协同
我们探索了在wtmc38-ova荷瘤小鼠中联合使用过继arg2-/-oti转移和pd1阻断的益处。wt小鼠受到mc38-ova肿瘤的攻击,五天后,当肿瘤可被触知时,小鼠接受如图7a所示的过继转移细胞,除了供体不是嵌合体小鼠外,其于次日进行免疫。在t细胞转移后的第8、11和14天,小鼠经i.p.注射接受200μg相关抗体。在这种情况下,我们再次观察到t细胞固有的arg2缺乏与pd1阻断之间的协同作用。与仅接受arg2-/-ot-i细胞或抗pd1抗体的小鼠相比,接受联合治疗的小鼠表现出更强的肿瘤生长降低(图9a),延长的存活时间(图9b),和增加的肿瘤清除率(图9c)。
实例9:抑制arg增强体外人t细胞活化
接下来,我们确定了人cd4+和cd8+t细胞中arg的抑制作用是否可能影响它们在体外的活化。从pbmc纯化的人t细胞未经处理或在体外用抗cd3和抗cd28ab活化,在存在或不存在arg抑制剂的情况下在96孔板的rpmi中培养,并评估其活化24h后的活化情况(cd69染色)。抑制arg酶功能增加cd4+(图10a)或cd8+细胞(图10b)中cd69+细胞的频率,表明arg抑制在体外增加人t细胞的活化。
- 还没有人留言评论。精彩留言会获得点赞!