敲减SphK2的小干扰RNA在制备逆转肝癌对瑞戈非尼耐药药物中的应用的制作方法
本发明涉及肿瘤治疗尤其是肿瘤耐药的逆转治疗领域,具体来说是指敲减sphk2的小干扰rna在制备逆转肝癌对瑞戈非尼耐药药物中的应用。
背景技术:
肝细胞肝癌(hepatocellularcarcinoma,hcc)是威胁我国人民生命和健康的重大疾病,瑞戈非尼(regorafenib)是治疗中晚期hcc的标准二线药物,能够在一线药物索拉非尼(sorafenib)治疗失败后继续生效,体内外研究中均展现出良好的抗hcc效果。然而获得性耐药的发生不仅限制了hcc患者从瑞戈非尼治疗中长久获益,还使疾病恶化,病程加速;且更为严重的是,患者对瑞戈非尼耐药后尚无其他可选药物。因此寻找可能的逆转hcc瑞戈非尼获得性耐药的潜在药物具有非常重要的临床意义。目前针对hcc瑞戈非尼获得性耐药问题的研究非常少,至今还未有成熟的解决方案。公开报道的在体内外具有确切的逆转hcc瑞戈非尼获得性耐药的潜在药物也几乎没有。
近年来肿瘤中的异常鞘磷脂代谢受到广泛关注,鞘氨醇激酶2(sphingosinekinase,sphk2)通过催化鞘氨醇-1-磷酸(sphingosine-1-phosphate,s1p)的生成在鞘磷脂代谢中发挥关键作用。研究显示sphk2由于存在bh3类似结构域,能够诱导多种细胞凋亡并抑制细胞增殖;后来的部分研究认为sphk2通过其具有促癌作用的催化产物s1p促进肿瘤的发生发展。然而,目前为止sphk2在肝癌的发生发展和耐药等生物学过程中的作用和机制尚不清楚。
由于肿瘤的获得性耐药是一个特殊的肿瘤生物学过程,对肿瘤具有抑制作用的药物不一定具有逆转肿瘤获得性耐药的功能。而且,药物之间具有联合之后的协调效应,也不意味着其中的一个药物可以逆转肿瘤对另外一个药物的耐药。更进一步的sphk2在肿瘤发生发展中的作用尚存在争议,且肿瘤耐药与肿瘤的其他生物学行为不同、有其特殊之处,因此其在耐药中的作用和机制尚不清楚。肝癌对瑞戈非尼的获得性耐药也是一个刚刚出现研究命题,其过程和机制尚不清楚,因此,敲减sphk2能否对肝癌的瑞戈非尼获得性耐药产生逆转作用尚存争议。
因此,目前敲减sphk2能否在hcc上展示抑癌效果不得而知,更进一步其能否逆转hcc对瑞戈非尼的耐药也处于迷茫之中。本发明将通过体外hcc瑞戈非尼获得性耐药模型评估敲减sphk2的小干扰rna对获得性耐药的逆转作用。
技术实现要素:
本发明的目的是通过体外研究揭示sphk2在肝癌细胞对瑞戈非尼获得性耐药中的作用,并且用小干扰rna敲减sphk2的方法来逆转了肝癌细胞对瑞戈非尼的耐药。因此,本发明的主要内容概括为:采用小干扰rna敲减sphk2的方法降低sphk2表达量能够逆转肝癌对瑞戈非尼的耐药,敲减sphk2的小干扰rna能够用于制备逆转肝癌对瑞戈非尼耐药的药物。
本发明内容详细情况如下:
1.我们首次揭示瑞戈非尼对耐药hcc细胞抑制增殖和促进凋亡的作用显著下降,耐药细胞对瑞戈非尼的敏感性降低;
2.我们首次揭示sphk2在耐药细胞中的表达比在非耐药细胞中高,而sphk1在两种细胞中的表达没有明显差别,且sphk2的表达量与hcc细胞对瑞戈非尼的敏感度具有负相关关系;
3.我们首次揭示sphk2的催化产物s1p刺激非耐药细胞也可以促进细胞对瑞戈非尼产生抵抗。
4.我们首次揭示过表达sphk2可以降低非耐药细胞对瑞戈非尼的敏感性,促进耐药发生;
5.我们首次使用敲减sphk2的小干扰rna降低sphk2蛋白表达水平后,耐药细胞对瑞戈非尼的敏感性升高,逆转耐药。联合应用敲减sphk2的小干扰rna瑞戈非尼具有显著体外逆转对瑞戈非尼耐药的逆转效果。由此可见,敲减sphk2的小干扰rna可以用于肿瘤对瑞戈非尼获得性耐药的逆转。
附图说明
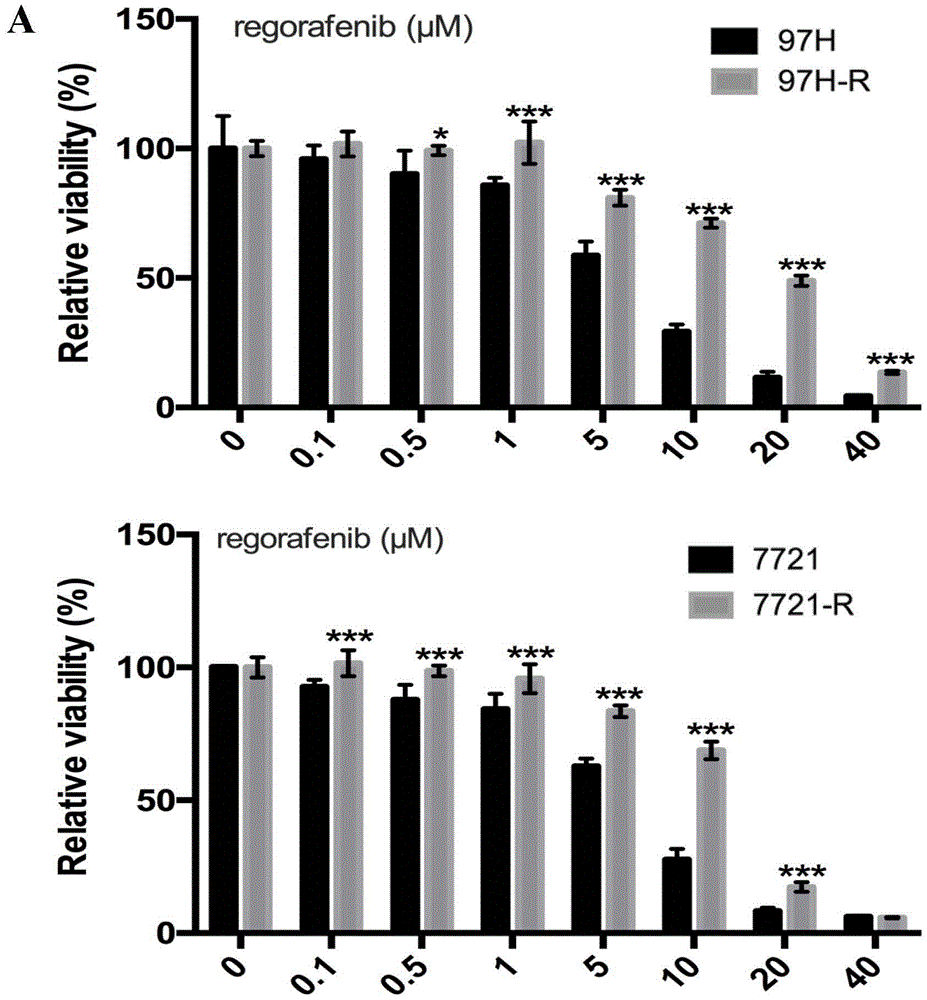
图1.瑞戈非尼获得性耐药细胞的构建:(a)瑞戈非尼耐药细胞和非耐药母系细胞用不同浓度瑞戈非尼(0、0.5、1、5、10、20、40(μmol/l))处理48h后,cck-8检测细胞活力,并计算相应的ic50值。(b)10μmol/l瑞戈非尼处理瑞戈非尼获得性耐药细胞97h-r、7721-r和非耐药母系细胞48h后,分别检测两种细胞的凋亡比例。(c)10μmol/l瑞戈非尼处理耐药细胞和母系细胞48h后,分别检测凋亡细胞的比例。(d)在克隆形成实验中,用5μmol/l瑞戈非尼抑制耐药细胞和母系细胞的增殖,7721和7721-r连续培养14天,97h和97h-r连续培养10天,检测瑞戈非尼对细胞增殖的影响。(e)10μmol/l瑞戈非尼处理耐药细胞和母系细胞48h后,westernblot检测不同组细胞中bcl2、cleaved-parp、cdk2、ckd4、cyclind1的蛋白表达水平,根据实验结果分析各条带灰度值。图中所示结果为三次独立实验中具有代表性的实验结果,结果用平均值±标准差(mean±sd)表示;*p<0.05,**p<0.01,***p<0.001。
图2.sphk2而非sphk1在与hcc瑞戈非尼耐药相关:(a)westernblot检测瑞戈非尼获得性耐药细胞97h-r、7721-r与对应的非耐药母系细胞mhcc97h、smmc-7721中sphk1蛋白表达量的差异。(b)瑞戈非尼获得性耐药细胞与非耐药母系细胞sphk2蛋白表达量的差异。(c)westernblot检测bel-7402、huh-7、plc/prf/5、smmc-7721、mhcc97h五株hcc细胞sphk2的蛋白表达水平,并分析各株细胞sphk2条带灰度值与瑞戈非尼ic50值的关系。图中所示结果为三次独立实验中具有代表性的实验结果,结果用平均值±标准差(mean±sd)表示;*p<0.05,**p<0.01,***p<0.001,ns:没有显著统计学差异。
图3.降低耐药细胞sphk2蛋白表达水平后,细胞对瑞戈非尼敏感性升高:(a)sphk2-sirna或阴性对照sirna干扰瑞戈非尼获得性耐药细胞97h-r、7721-r后,westernblot检测细胞sphk2的蛋白表达量确认是否敲除成功,根据结果用imagej分析各条带灰度值。(b)成功降低耐药细胞sphk2蛋白表达水平后,用不同浓度的瑞戈非尼(0、0.5、1、5、10、20、40(μmol/l))处理细胞48h,cck-8实验检测细胞活力并计算相应的瑞戈非尼ic50值。(c)用10μmol/l瑞戈非尼分别处理sphk2干扰成功的耐药细胞和对照细胞48h后,annexinv-fitc/pi检测凋亡细胞的比例。(d)10μmol/l瑞戈非尼分别处理sphk2干扰成功的耐药细胞和对照细胞48h后,流式细胞周期实验检测细胞周期的分布。(e)在克隆形成实验中瑞戈非尼的浓度为5μmol/l,处理时间分别为:7721-r14天,97h-r10天,检测敲除sphk2后瑞戈非尼对耐药细胞增殖能力的影响。(f)10μmol/l瑞戈非尼处理sphk2干扰成功的耐药细胞和对照细胞48h后,westernblot检测不同组细胞中bcl2、cleaved-parp、cdk2、ckd4、cyclind1的蛋白表达水平,根据实验结果分析各条带灰度值。图中所示结果为三次独立实验中具有代表性的实验结果,结果用平均值±标准差(mean±sd)表示;*p<0.05,**p<0.01,***p<0.001。
图4.升高非耐药hcc细胞sphk2蛋白表达水平后,细胞对瑞戈非尼的敏感性降低:(a)westernblot检测过表达sphk2细胞和对照细胞中sphk2蛋白表达水平,根据结果分析各条带灰度值。(b)成功升高hcc细胞sphk2表达后,用不同浓度的瑞戈非尼(0、0.5、1、5、10、20、40(μmol/l))处理细胞48h,cck-8实验检测细胞活力并计算相应的瑞戈非尼ic50值。(c)用10μmol/l处理sphk2慢病毒过表达成功的hcc细胞和对照细胞48h后,annexinv-fitc/pi检测凋亡细胞的比例。(d)10μmol/l处理sphk2慢病毒过表达成功的hcc细胞和对照细胞48h后,流式细胞周期实验检测细胞周期的分布。(e)在克隆形成实验中瑞戈非尼的浓度为5μmol/l,处理时间分别为:7721-r14天,97h-r10天,检测过表达sphk2后瑞戈非尼对细胞增殖的影响。(f)10μmol/l瑞戈非尼处理sphk2慢病毒过表达成功的hcc细胞和对照细胞48h后,westernblot检测不同组细胞中bcl2、cleaved-parp、cdk2、ckd4、cyclind1的蛋白表达水平,根据结果分析各条带灰度值。图中所示结果为三次独立实验中具有代表性的实验结果,结果用平均值±标准差(mean±sd)表示;*p<0.05,**p<0.01,***p<0.001,ns:没有显著统计学差异。
图5.s1p刺激促进hcc细胞对瑞戈非尼的耐受:(a)用不同浓度s1p刺激非hcc细胞48h,cck-8检测细胞活力。(b)1μmol/ls1p刺激hcc细胞,同时用不同浓度的瑞戈非尼(0、0.5、1、5、10、20、40(μmol/l))处理细胞48h,cck-8实验检测细胞活力并计算相应的瑞戈非尼ic50值。(c)用10μmol/l瑞戈非尼分别处理s1p或dmso刺激的hcc细胞和对照细胞48h后,annexinv-fitc/pi检测各实验组中凋亡细胞的比例。(d)10μmol/l瑞戈非尼分别处理s1p或dmso刺激的hcc细胞和对照细胞48h后,流式细胞周期实验检测各实验组中细胞周期的分布。(e)在克隆形成实验中瑞戈非尼的浓度为5μmol/l,处理时间分别为:7721-r14天,97h-r10天,检测s1p或dmso刺激后瑞戈非尼对细胞增殖的影响。(f)10μmol/l瑞戈非尼处理s1p或dmso刺激的hcc细胞48h后,westernblot检测不同实验组细胞中bcl2、cleaved-parp、cdk2、ckd4、cyclind1的蛋白表达水平,并根据检测结果分析各条带灰度值。图中所示结果为三次独立实验中具有代表性的实验结果,结果用平均值±标准差(mean±sd)表示;*p<0.05,**p<0.01,***p<0.001。
具体实施方式
实施例部分所涉到的所有实验材料和方法
1.细胞培养
人hcc细胞株bel-7402、huh-7、plc/prf/5、smmc-7721均购于上海中国科学院细胞库。细胞株mhcc97h由复旦大学附属中山医院贾凡教授赠送。这五种hcc细胞的培养基均为添加了10%胎牛血清、100u/ml盘尼西林和100μg/ml链霉素的dmem完全培养基。细胞在温度为37℃、co2浓度为5%的恒温培养箱中持续培养。
2.细胞活力检测(cck-8法)
用含10%胎牛血清的完全培养基制备细胞混悬液,以每孔5000个细胞/100μl的浓度接种于96孔板内,接种好的细胞培养24h使之贴壁后用不同药物处理细胞相应时间后,每孔加入cck-8试剂10μl,37℃恒温培养箱内继续孵育1~2h,酶标仪检测波长为450nm处各孔吸光度值并记录,并做相关分析,每次实验重复三次。
3.瑞戈非尼获得性耐药细胞建立
我们使用浓度梯度递增法建立瑞戈非尼获得性耐药hcc细胞株。首先利用cck-8实验检测5个hcc细胞株对瑞戈非尼的敏感性,根据cck-8检测结果计算各株细胞对应的瑞戈非尼ic50值,选择对瑞戈非尼相对敏感(瑞戈非尼ic50值较低)的两株hcc细胞进行耐药细胞的建立。初始瑞戈非尼的处理浓度略低于细胞瑞戈非尼ic50(smmc-7721、mhcc-97h初始瑞戈非尼处理浓度均为5μmol/l),每隔两天去除陈旧培养基,pbs清洗细胞后更换为新鲜培养基。之后每周缓慢增加瑞戈非尼的浓度0.5μmol/l,并用pbs洗涤去除死亡细胞,剩余存活hcc细胞继续培养传代。在用瑞戈非尼连续刺激hcc细胞6个月后,再用不同浓度的瑞戈非尼处理细胞48h(瑞戈非尼浓度设置为0、0.5、1、5、10、20、30(μmol/l)),利用cck-8实验检测细胞活力,根据cck-8结果的计算这些细胞对瑞戈拉非尼ic50值,并与母系非耐药细胞的瑞戈非尼ic50值比较,计算耐药指数(resistanceindex,ri)(ri=耐药细胞瑞戈非尼ic50值/母系细胞瑞戈非尼ic50值),确定瑞戈非尼获得性耐药hcc细胞株是否建立成功。构建成功的瑞戈非尼耐药细胞继续用含4μmol/l瑞戈非尼的完全培养基持续培养以维持其耐药性。
4.细胞凋亡流式检测(annexinv-fitc/pi双染法)
hcc细胞在经过不同处理后,胰酶消化收集约1×106个细胞。1000rpm离心5min,去除上清;加入5μlannexinv-fitc染料和5μl碘化丙啶(propidiumiodide,pi)染料,轻轻混匀,室温避光孵育染色30min,孵育过程中轻轻重悬细胞2~3次使得染色更均匀充分。染色结束后用筛网过滤细胞制备成单细胞悬液,使用bdfacscalibur流式仪对凋亡细胞进行检测,flowjo软件分析结果。所获取的结果图中,q1(左上象限)、q2(右上象限)、q3(右下象限)和q4(左下象限)分别代表死亡细胞、晚期凋亡细胞、早期凋亡细胞和存活细胞。每次实验独立重复三次。
5.细胞周期流式检测
hcc细胞在经过不同处理后,胰酶消化收集约1×106个细胞。1000rpm离心5min,去除上清;4℃、70%的酒精2ml固定细胞30min,1000rpm离心5min,去除上清;rnasea的pbs溶液处理细胞30min,加入5μlpi染料,对细胞进行室温避光孵育染色30min,孵育过程中轻轻重悬细胞2~3次使得染色更均匀充分。染色结束后筛网过滤细胞制备成单细胞悬液,bdfacscalibur流式仪进行检测,flowjo软件分析结果。每次实验独立重复三次。
6.细胞克隆形成实验
取对数生长期的hcc细胞,胰酶消化,完全培养基终止并用pbs制备细胞悬液。以每孔1×103个细胞的密度接种于六孔板中,恒温培养箱孵育过夜使细胞贴壁,之后将培养基更换为含5μmol/l瑞戈非尼或dmso对照的完全培养基。每天在显微镜下观察细胞生长情况,每三天更换一次新鲜细胞培养基。连续培养10~14天后(smcc-7721和7721-r培养14天,mhcc-97h和97h-r培养10天),去除细胞培养基,pbs清洗细胞;用4%多聚甲醛固定细胞20min,pbs清洗;0.4%结晶紫溶液对细胞染色30min,pbs洗涤去除染色液并干燥,拍照计数。在显微镜(低倍镜)下人工计数≥30个细胞的克隆数。每次实验独立重复三次。
7.小干扰rna(smallinterferingrna,sirna)转染
我们通过sirna干扰技术降低sphk1和sphk2的表达,按照说明书使用lipofectaminetm2000转染试剂将sirna转染入hcc细胞。实验涉及到的sirna序列详见附表。具体步骤如下:将适量hcc细胞种植在六孔板中,恒温培养箱孵育过夜使细胞贴壁,保证转染时细胞汇合度在30~50%左右为宜。4℃,12000rpm离心sirna粉末,无酶水溶解。每孔转染方式如下:a管:将60pmsirna溶于150μl的optiprotmsfm无血清培养基中;b管:将4μllipofectaminetm2000溶于150μl的optiprotmsfm无血清培养基中;c管:将a管和b管混合至c管中,室温静置20min。在静置c管期间,将六孔板培养基更换为无血清培养基,每孔800μl,将静置20min后的c管混合液加入到六孔板中,每孔300μl,4~6h后将培养基更换为含胎牛血清的dmem完全培养基。sirna转染细胞48h后,westernblot检测sphk1和sphk2的蛋白表达水平,确认是否敲除成功。
8.慢病毒转染
纯化的高滴度慢病毒悬液购自上海吉玛公司。慢病毒载体构建、慢病毒的扩增和纯化以及滴度测定均由该公司完成,所使用的慢病毒载体为gv492,元件顺序为ubi-mcs-3flag-cbh-gcgfp-ires-puromycin,将目的片段插入mcs区后调控sphk2表达。
将适量hcc细胞种植在六孔板中,每孔培养基体积为2ml,恒温培养箱孵育过夜使细胞贴壁,保证转染时细胞汇合度在30~50%左右为宜。根据细胞的感染复数(multiplicityofinfection,moi)在培养基中加入相应比例1×108tu/ml的病毒悬液,继续培养细胞72~96h后荧光显微镜下观察细胞荧光表达情况确认慢病毒是否转染成功,同时在细胞培养基中加入适量的嘌呤霉素筛选细胞,以获得稳定过表达sphk2的hcc细胞株。之后westernblot检测sphk2的蛋白表达水平,确认是否过表达成功。
9.数据分析
所获的实验数据用spss19.0统计软件进行分析,具有代表性的独立实验的结果用平均值±标准差表征。两组之间的比较采用独立t检验方法,p值低于0.05(p<0.05)认为具有统计学上的显著差异。
实施例1肝癌瑞戈非尼获得性耐药细胞的成功构建
我们通过cck-8实验检测了bel-7402、huh-7、plc/prf/5、smmc-7721、mhcc-97h五株hcc细胞对瑞戈非尼的敏感性,根据检测结果计算得到各细胞对应的瑞戈非尼ic50值。可以发现,smmc-7721、mhcc-97h的瑞戈非尼的ic50在五株细胞中处于较低水平(表1),证明这两株细胞对瑞戈非尼相对敏感。由此我们选取这两株细胞利用浓度梯度递增法构建瑞戈非尼获得性耐药hcc细胞株。瑞戈非尼持续刺激smmc-7721、mhcc-97h六个月后,我们再次检测了这两株细胞对瑞戈非尼的敏感性,发现这两株细胞瑞戈非尼的ic50值显著升高,耐药指数ri分别为3.133(97h-r/97h)、2.26(7721-r/7721)(图1.a,表2),说明对瑞戈非尼的敏感性降低,确认瑞戈非尼获得性耐药细胞构建成功,由此获得的耐药hcc细胞分别命名为7721-r和97h-r。此外,我们还通过细胞周期实验以及克隆形成实验检测瑞戈非尼对耐药细胞的生长抑制作用。结果显示10μmol/l瑞戈非尼处理细胞48h后,阻滞在g1期的耐药细胞比阻滞在g1期的非耐药母系细胞少;5μmol/l瑞戈非尼在耐药细胞中抑制克隆形成的效力也要比在非耐药母系细胞中差(图1.c-d),说明瑞戈非尼抑制耐药细胞增殖的效力显著下降。同时10μmol/l瑞戈非尼处理hcc细胞48h后,耐药细胞中凋亡细胞的比例比非耐药细胞中低(图1.b),说明瑞戈非尼诱导耐药细胞凋亡的能力也明显下降。并且westernblot结果显示与细胞周期相关的蛋白:g1/s特异性周期蛋白-d1(cyclind1)和周期蛋白依赖性激酶2(cyclin-dependentkinases2,cdk2)、cdk4的表达水平以及细胞凋亡的标志性蛋白:剪切形式的腺苷二磷酸核糖聚合酶(cleaved-polyadp-ribosepolymerase,cleaved-parp)和抑制细胞凋亡的蛋白b细胞淋巴瘤2(b-celllymphoma-2,bcl2)的蛋白水平变化与现象相符(图1.f)。综合以上各项实验结果,我们确认hcc瑞戈非尼获得性耐药细胞构建成功,瑞戈非尼抑制耐药细胞增殖、促进耐药细胞凋亡的能力均显著下降。
表1.五株hcc细胞的瑞戈非尼ic50值(μmol/l)
表2.非耐药细胞和耐药细胞的瑞戈非尼ic50值(μmol/l)
实施例2sphk2表达水平与瑞戈非尼获得性耐药相关
为了验证sphks是否参与hcc瑞戈非尼获得性耐药的调控,我们首先比较了耐药细胞与非耐药母系细胞之间sphk1、sphk2蛋白表达水平差异。westernblot实验结果显示,mhcc97h和97h-r、smmc-7721和7721-r的sphk1蛋白表达没有显著差异,而sphk2在耐药细胞97h-r和7721-r中表达水平显著升高(图2.a-b),说明sphk2而非sphk1可能介导了hcc瑞戈非尼获得性耐药。此外我们还发现:在五株hcc细胞bel-7402、huh-7、plc/prf/5、smmc-7721、mhcc97h中,细胞sphk2的westernblot条带灰度值越高,对应的瑞戈非尼ic50值也越高。进一步数据分析显示:细胞的sphk2表达水平与瑞戈非尼ic50值的皮尔逊相关系数r2=0.8889(图2.c),存在显著的正相关关系,说明hcc细胞sphk2的表达量越高,细胞对瑞戈非尼敏感性越低。综合以上结果,我们发现sphk2在hcc的瑞戈非尼耐药过程中扮演着重要角色。
实施例3敲减sphk2升高hcc耐药细胞对瑞戈非尼的敏感性
我们利用sirna干扰技术,降低了97h-r和7721-r这两株瑞戈非尼获得性耐药细胞中sphk2的蛋白表达量,并通过westernblot方法进行了验证,确认sphk2敲除成功(图3.a)。cck-8检测结果显示:成功敲除sphk2的耐药细胞在经过不同浓度瑞戈非尼刺激48h后细胞活力比阴性对照低,且其瑞戈非尼ic50值也明显降低(图3.b,表3),说明降低sphk2蛋白表达水平后耐药细胞对瑞戈非尼的敏感性增加。此外,将成功敲除sphk2的耐药细胞和对照细胞分别置于含有相同浓度瑞戈非尼的完全培养基中孵育相同时间后,成功敲除sphk2的耐药细胞所形成的克隆数要比对照细胞少(图3.e),阻滞在g1期的细胞也明显增多(图3.d)。同时cyclind1和cdk2、cdk4的表达水平在经过sphk2敲除和瑞戈非尼刺激共同处理的耐药细胞中的水平最低(图3.f)。这些结果说明瑞戈非尼能够将成功敲除sphk2的耐药细胞阻滞在g1期,抑制其生长、增殖。annexinv-fitc/pi凋亡检测发现成功敲除sphk2的耐药细胞在经过10μmol/l瑞戈非尼刺激后凋亡细胞的比例比对照组高(图3.c),cleaved-parp表达升高,bcl2表达降低(图3.f),说明瑞戈非尼能够诱导成功敲除sphk2的耐药细胞凋亡。综合以上各项实验结果,我们证明了降低sphk2蛋白表达水平可以部分恢复耐药细胞对瑞戈非尼的敏感性。
表3.敲减sphk2的耐药细胞与阴性对照细胞瑞戈非尼ic50值
实施例4过表达sphk2降低hcc细胞对瑞戈非尼的敏感性
鉴于降低sphk2蛋白表达水平可以升高耐药细胞对瑞戈非尼的敏感性,我们进一步检测升高sphk2蛋白表达水平是否能够降低非耐药细胞对瑞戈非尼的敏感,正向验证sphk2在hcc瑞戈非尼获得性耐药中的重要作用。
我们利用慢病毒转染技术升高非耐药细胞mhcc97h、smmc-7721中sphk2的蛋白表达水平,并通过westernblot实验进行了验证,结果显示非耐药hcc细胞sphk2过表达成功(图4.a)。用不同浓度瑞戈非尼刺激成功过表达sphk2的hcc细胞和对照hcc细胞48h后,cck-8检测发现过表达sphk2的hcc细胞活力要比对照细胞高(图4.b),根据细胞活力计算得到的瑞戈非尼ic50值也更高(表4),说明过表达sphk2后hcc细胞对瑞戈非尼的敏感性降低。此外,将成功过表达sphk2的耐药细胞和对照细胞分别置于含有相同浓度瑞戈非尼的完全培养基中培养相同时间后,过表达sphk2的hcc细胞所形成的克隆数要比对照细胞多(图4.e),阻滞在g1期的细胞也相对较少(图4.d)。同时过表达sphk2的hcc细胞中细胞周期相关蛋白cyclind1、cdk2、cdk4的表达水平在经过瑞戈非尼刺激后没有明显降低,而对照细胞中这些蛋白表达水平在瑞戈非尼刺激后明显降低(图4.f)。说明,在升高hcc细胞sphk2的蛋白水平后,瑞戈非尼无法有效将细胞阻滞在g1/s期也无法抑制细胞生长、增殖。annexinv-fitc/pi凋亡检测发现成功过表达sphk2的hcc细胞在经过10μmol/l瑞戈非尼刺激后凋亡细胞的比例比对照组低(图4.c),westernblot结果显示对照细胞的细胞凋亡标志性蛋白cleaved-parp表达水平更高,抑制细胞凋亡的蛋白bcl2则在过表达sphk2的hcc细胞中水平更高(图4.f),说明瑞戈非尼诱导凋亡的作用在sphk2过表达的hcc中减弱。综合以上各项实验结果,我们证明了升高sphk2蛋白表达水平可以降低hcc细胞对瑞戈非尼的敏感性,促进hcc细胞瑞戈非尼耐药的产生。
表4.过表达sphk2的非耐药细胞在与对照细胞瑞戈非尼的ic50值
实施例5外源性s1p刺激降低hcc细胞对瑞戈非尼的敏感性
我们通过敲除和过表达sphk2正反向证明了sphk2在hcc瑞戈非尼获得性耐药中的重要作用,鉴于sphk2的主要功能是催化s1p的生成,我们推测sphk2促进hcc瑞戈非尼获得性耐药是通过s1p介导的。首先我们检测了不同浓度的s1p对hcc细胞活力的影响,cck-8结果显示1μmol/l的s1p对mhcc97h和smmc-7721两株hcc细胞均可产生一定程度的生长促进作用。且由于人外周血中检测到的s1p浓度约为0.1~1μmol/l,研究鞘脂代谢的文章中多用1μmol/l的s1p刺激细胞,于是我们选择1μmol/l的s1p对hcc细胞进行刺激。
当用不同浓度的瑞戈非尼处理1μmol/l的外源性s1p或dmso刺激的hcc细胞48h后,cck-8检测发现s1p刺激的hcc细胞活力要比dmso刺激的细胞高(图5.a),其对应的瑞戈非尼ic50值也更高(表5),说明hcc细胞在1μmol/ls1p作用后对瑞戈非尼的敏感性降低。此外,将hcc细胞分别置于含有1μmol/ls1p或dmso的完全培养基中,再加入同等浓度的瑞戈非尼分别培养相同时间后,s1p刺激的hcc细胞所形成的克隆数要比dmso处理的细胞多(图5.d),细胞周期结果显示阻滞在g1期的细胞也相对较少(图5.c)。同时s1p刺激的hcc细胞中细胞周期相关蛋白cyclind1、cdk2、cdk4的表达水平在经过瑞戈非尼处理后没有明显降低,而dmso刺激的hcc细胞中这些蛋白表达水平在瑞戈非尼处理后明显降低(图5.f)。说明hcc在经过适量s1p刺激后,瑞戈非尼无法有效将细胞阻滞在g1/s期以及无法抑制细胞生长、增殖。此外,annexinv-fitc/pi凋亡检测发现s1p刺激的hcc细胞在经过10μmol/l瑞戈非尼作用后凋亡细胞的比例比对照组低(图5.b),细胞凋亡的标志性蛋白cleaved-parp表达水平更低,抑制细胞凋亡的蛋白bcl2表达水平更高(图5.f),说明瑞戈非尼诱导凋亡的作用在s1p刺激的hcc细胞中减弱。综合以上各项实验结果,我们证明了适量s1p刺激同样也可以降低hcc细胞对瑞戈非尼的敏感性,促进hcc细胞瑞戈非尼耐药的产生,sphk2的粗瑞戈非尼耐药作用可能由s1p介导。
表5.非耐药细胞在s1p刺激后与对照细胞瑞戈非尼的ic50值
发明小结
综合以上研究结果,我们首次揭示了鞘磷脂代谢的限速酶sphk2在hcc瑞戈非尼获得性耐药中的关键作用,由于本领域之前对sphk2在hcc进展以及hcc的耐药中的作用和相应的机制都不清楚,我们的发现对于本领域具有重要意义。更加重要的是我们首次利用小干扰rna干扰降低sphk2的蛋白表达水平,产生了明确的逆转hcc的瑞戈非尼获得性耐药的作用。综上所述,本发明所提供的敲减sphk2的特异性小干扰rna能够用于制备逆转肿瘤的瑞戈非尼获得性耐药,尤其是逆转hcc的瑞戈非尼获得性耐药的药物。
序列表
<110>南京大学
<120>敲减sphk2的小干扰rna在制备逆转肝癌对瑞戈非尼耐药药物中的应用
<130>20200608
<160>2
<170>siposequencelisting1.0
<210>1
<211>21
<212>dna
<213>人工序列(.)
<400>1
gggcaaggccuugcagcuctt21
<210>2
<211>21
<212>dna
<213>人工序列(.)
<400>2
uucuccgaacgugucacgutt21
- 还没有人留言评论。精彩留言会获得点赞!