用于测定抗体或配体结合和功能的基于细胞的测定的制作方法
本发明涉及用于测定抗体或配体结合和功能的新的基于细胞的测定。
背景技术:
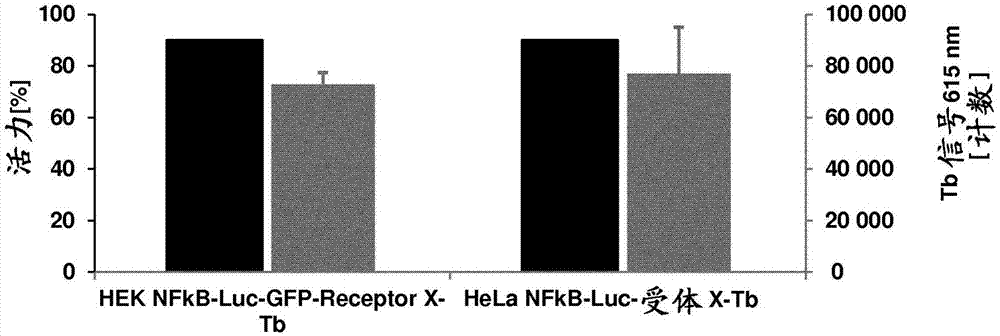
:化学疗法直到现在仍是最常用的癌症治疗之一。但是,由于化疗剂主动靶向分裂细胞(癌细胞的一个特征),健康的分裂细胞如血细胞;肠、口腔和毛发中的细胞也受影响,产生很强的副作用。科学家一直致力于改善化疗剂的施用和组合,以最小化那些副作用。此外,基于抗体的治疗在过去15年中演变,现在在恶性血液肿瘤和实体瘤的治疗中是化学疗法方法的有价值的组合或替代。与化学疗法不一样,抗体治疗靶向癌细胞上的特异性抗原,从而允许更定向的治疗,从而减少对健康组织的副作用。在开发基于抗体的治疗剂的过程中,需要多种测定来鉴定最好的候选治疗剂进入临床试验,最终上市。在第一个早期临床前阶段,必须产生抗体,并分析它们的靶标特异性,以及它们对靶标的亲和力和功能性。结合特性可以用多种蛋白质-蛋白质相互作用测定分析,如基于fret的方法、表面等离振子共振(spr)、萦光激活细胞分选(facs)或alphascreentm。通常在设计为尽可能接近生理环境的多种基于细胞的测定中测试功能性,以鉴定最佳候选者来在进入临床试验之前在动物模型中测试。这些功能性测定通常用原代细胞、肿瘤细胞系或设计为在激活特异性途径时表达报道分子的报道细胞进行。在抗体治疗分子开发过程的早期阶段设计允许评估结合和功能性的组合测定具有重要意义。本发明的发明人开发了将抗体和抗体样构建体(例如配体)的结合和功能性评估组合在同一个小管(vial)中的新测定。此新测定用于例如筛选或表征目的。此新测定是不仅针对结合还针对功能性进行早期筛选的有价值的工具,其将允许在开发候选药物的早期阶段鉴定最佳结合剂。发明概述在一个实施方案中,提供用于测定特异性结合靶抗原的抗体或配体的结合和功能性的体外测定,其包括以下步骤:i)提供细胞,该细胞a)在其表面表达靶抗原;b)用能量供体化合物共价或非共价标记;和c)包含靶抗原反应元件控制下的报道基因;ii)加入待测试的抗体或配体;iii)通过测定能量转移来测量与靶抗原的结合,其中能量受体化合物共价或非共价缀合于待测试的抗体或结合一抗的二抗;和iv)通过将报道基因表达水平与靶抗原激活或抑制水平相关来测定抗体或配体的功能性。在一个实施方案中,该能量转移供体和受体化合物是萦光共振能量转移(fret)能量供体和受体化合物,在步骤iii)中测定的能量转移是萦光共振能量转移(fret)。在一个实施方案中,该fret是时间分辨fret。在一个实施方案中,该fret能量供体化合物是铽穴合物(cryptate)和/或该fret能量受体化合物是d2。在一个实施方案中,该能量供体和受体化合物是生物发光能量转移(bret)能量供体和受体化合物,在步骤iii)中测定的能量转移是生物发光能量转移(bret)。在一个实施方案中,该能量供体和受体化合物是alphascreen受体和供体小球,在步骤iii)中测定的能量转移是受体小球内从单线态氧至二甲基噻吩衍生物的能量转移。在一个实施方案中,在同一个小管中测量抗体与靶抗原的结合和功能性。在一个实施方案中,用能量供体化合物共价或非共价标记靶抗原。在一个实施方案中,该能量供体化合物与麦胚凝集素(wga)共价或非共价连接。在一个实施方案中,该报道基因选自编码萦光蛋白质的基因或编码可以检测其催化活性的酶的基因。在一个实施方案中,该报道基因编码绿色萦光蛋白(gfp)或萦光素酶。在一个实施方案中,该靶抗原是细胞表面受体。在一个实施方案中,步骤iii)和iv)顺次进行或同时进行。在一个实施方案中,该靶抗原和反应元件是nf-κb途径的部分。在一个实施方案中,该反应元件包含具有dna序列seqidno:1、2、3、4或5的至少一个dna重复。在一个实施方案中,该反应元件包含dna序列seqidno6、7、8或9。在一个实施方案中,该测定包括用含有靶抗原反应元件控制下的编码报道基因的dna序列的表达载体转染细胞的预备步骤。附图简述图1.snap-受体x转染和供体萦光染料铽标记后细胞的转染效率测定及活力测量。在615nm波长测量每孔10000个细胞的铽信号。比较了转染试剂lipofectamine2000、lipofectamineltx和xtremegenehp。图2.snap-受体x转染和供体萦光染料铽标记后heknfκb-luc-gfp和helanfκb-luc细胞的lipofectamineltx转染效率测定及活力测量。在615nm波长测量每孔10000个细胞的铽信号。图3.抗体-i和抗体-ii与表达受体x的heknfκb-luc-gfp和helanfκb-luc细胞的间接结合测定。抗体按2倍稀释系列从25nm稀释至0.05nm。使用浓度为150nm的d2标记二抗。用graphpadprism6.0通过非线性回归拟合结合曲线。图4.抗体-i与表达受体x的heknfκb-luc-gfp细胞结合的间接结合测定。抗体-i按2倍稀释系列从12.5nm稀释至0.025nm。使用浓度为150nm或75nm的d2标记二抗。用graphpadprism6.0通过非线性回归拟合结合曲线。图5.抗体-i、抗体-iii和抗体-iv与表达受体x的heknfκb-luc-gfp细胞的间接结合测定。所有抗体按2倍稀释系列从20nm稀释至0.04nm。使用浓度为75nm的d2标记二抗。用graphpadprism6.0通过非线性回归拟合结合曲线。图6.抗体-iii与表达受体x的heknfκb-luc-gfp细胞的间接结合测定。抗体以2倍稀释系列按从20nm至0.04nm的浓度稀释。使用浓度为75nm的d2标记二抗。比较室温孵育和37℃孵育。用graphpadprism6.0通过非线性回归拟合结合曲线。图7.抗体-iii与表达受体x的heknfκb-luc-gfp细胞结合的间接结合测定。抗体以2倍稀释系列按从20nm至0.04nm的浓度稀释。使用浓度为75nm的d2标记二抗。比较用于稀释的不同缓冲液和培养基。用graphpadprism6.0通过非线性回归拟合结合曲线。图8.抗体-iii与表达受体x的heknfκb-luc-gfp细胞结合的间接结合测定。在解冻细胞后直接及在随后3天的每一天进行实验。抗体以2倍稀释系列从20nm稀释至0.04nm。使用浓度为75nm的d2标记二抗。用graphpadprism6.0通过非线性回归拟合结合曲线。图9.标记的表达受体x的heknfκb-luc-gfp细胞的铽信号和活力的测定。解冻后直接及在随后3天的每一天测量铽信号。每天接种每孔10000个细胞,并在615nm波长测量。图10.孵育6小时后抗体-i和tnfα与表达受体x的heknfκb-luc-gfp细胞的萦光素酶1000测定系统。tnfα稀释系列(2倍)从25至0.8ng/ml,抗体-i稀释系列(4倍)从30至0.03nm。用抗体-i和二抗单独作为对照。用graphpadprism6.0通过非线性回归拟合结合曲线。图11.tnf-a激活孵育48小时后one-glotm萦光素酶测定系统与萦光素酶1000测定系统比较。通过tnfα(50ng/ml)激活稳定转染的heknfκb-luc-gfp-受体x细胞和瞬时转染的heknfκb-luc-gfp-受体x-tb细胞的nfκb途径。图12.孵育6小时和48小时后的one-glotm萦光素酶测定系统。通过抗体-iii(10nm)作为一抗和抗人(anti-hu)iggfcγ-特异性山羊iggf(ab)2作为二抗(40nm)来激活稳定转染的heknfκb-luc-gfp-受体x细胞和瞬时转染的heknfκb-luc-gfp-受体x-tb细胞的nfκb途径。图13.孵育24小时后在96孔板中及在384孔板中进行的one-glotm萦光素酶测定系统。通过抗体-iii(10nm)作为一抗和抗人iggfcγ-特异性山羊iggf(ab)2作为二抗(40nm)来激活稳定转染的heknfκb-luc-gfp-受体x细胞的nfκb途径。图14.孵育6小时后在384孔板中进行的one-glotm测定系统。通过抗体-iii(10nm)作为一抗和抗人iggfcγ-特异性山羊iggf(ab)2作为二抗(40nm)来激活稳定转染的heknfκb-luc-gfp-受体x细胞的nfκb途径。图15.抗体-iii与表达受体x的heknfκb-luc-gfp细胞的wga-htrf间接结合测定。抗体-v作为阴性对照。两种抗体都以2倍稀释系列从1.56nm稀释至0.01nm。使用浓度为75nm的d2标记二抗。用graphpadprism6.0通过非线性回归拟合结合曲线。图16.抗体-iii与瞬时超转染和瞬时转染的表达受体x的heknfκb-luc-gfp细胞的间接结合测定。所有抗体都以2倍稀释系列从12.5nm稀释至0.02nm。使用浓度为75nm的d2标记二抗。用graphpadprism6.0通过非线性回归拟合结合曲线。图17.孵育24小时后在one-glotm萦光素酶测定系统中测试稳定转染、瞬时转染和瞬时超转染的表达受体x的heknfκb-luc-gfp细胞。通过从40至2.5nm2倍系列稀释的抗体-iii激活nfκb途径。作为二抗的抗人iggfcγ-特异性山羊iggf(ab)2按与一抗相比为4倍的摩尔量(extent)加入。图18.孵育24小时后在one-glotm萦光素酶测定系统中测试瞬时超转染的表达受体x的heknfκb-luc-gfp细胞。通过浓度为40nm的抗体-iii激活nfκb途径。按1:1至1:5之间(一抗:二抗)的比值测试作为二抗的抗人iggfcγ-特异性山羊iggf(ab)2和抗人igg-d2。图19.抗体-iii与瞬时超转染的表达受体x的heknfκb-luc-gfp细胞的间接结合测定。抗体-iii以2倍稀释系列从40nm稀释至0.02nm。使用浓度为120nm的d2标记二抗。用graphpadprism6.0通过非线性回归拟合结合曲线。图20.孵育24小时后在one-glotm萦光素酶测定系统中测试瞬时转染的表达受体x的heknfκb-luc-gfp细胞。通过从40至0.02nm2倍系列稀释的抗体-iii激活nfκb途径。作为二抗的抗人igg-d2按与一抗相比为3倍的摩尔量加入。图21.drozitumab和对照抗体aby与表达hudr5的hekebna细胞的alphascreen结合测定。所有抗体按在孔中终浓度从0.63nm至0.05nm2倍系列稀释。(a)通过非线性回归拟合的drozitumab和aby的s型结合曲线。(b)通过非线性回归测定drozitumab和aby的kd值。图22.abz和对照抗体aby与表达受体z的hekebna细胞的alphascreen结合测定。所有抗体按在孔中终浓度从2.5nm至0.05nm2倍系列稀释。(a)通过非线性回归拟合的abz和aby的s型结合曲线。(b)通过非线性回归测定abz和aby的kd值。图23.pbs/1%fcs(a)或生长培养基(b)中抗cd3与表达cd3的细胞的wga-fret结合测定。此外,在两个测定中都包括非结合对照igg作为阴性对照。用graphpadprism单位点特异性结合函数拟合曲线。图24.pbs/1%fcs(a)或生长培养基(b)中抗cd3结合时的nfkb途径激活。此外,在两个测定中都用非结合对照igg作为阴性对照。用one-glotm萦光素酶测定系统测量萦光素酶活性。用graphpadprisms型剂量反应拟合测定ec50值。发明详述i.定义本文所用的―报道基因”指可测定其表达的基因。在优选实施方案中,“报道基因”是编码蛋白质的基因,用该蛋白质的产生和检测作为替代来间接检测待测试抗体或配体的活性。报道蛋白是由报道基因编码的蛋白质。优选地,报道基因编码可通过简单的测定方法检测其催化活性的酶,或编码下述蛋白质,其具有诸如内萦光的特性,使得可以在需要最少样品制备的简单和快速的测定中检测报道基因的表达。可以检测其催化活性的酶的非限制性实例是萦光素酶、β半乳糖苷酶、碱性磷酸酶。萦光素酶是分子量(mw)为61kda的单体酶。它作为催化剂发挥作用,能够在腺苷三磷酸(atp)和mg2+存在下将d-萦光素转化为萦光素腺苷酸。此外,作为副产物产生焦磷酸(ppi)和腺苷单磷酸(amp)。然后中间体萦光素腺苷酸氧化为氧化萦光素、二氧化碳(co2)和光。氧化萤光素是生物发光产物,其可以在发光计中通过从反应释放的光来定量测量。萦光素酶报告测定(luciferasereporterassay)可购得,且为本领域已知,例如luciferase1000assaysystem和one-glotmluciferaseassaysystem。术语“具有内萦光的蛋白质”指能够通过蛋白质内的内部氨基酸的环化和氧化或通过酶促加入萦光辅因子而形成高萦光内在生色团的蛋白质。术语“具有内萦光的蛋白质”包括野生型萦光蛋白质和显示改变的光谱或物理特性的突变体。该术语不包括仅由于蛋白质内未修饰的酪氨酸、色氨酸、组氨酸和苯丙氨酸集团的萦光贡献而显示弱萦光的蛋白质。具有内萦光的蛋白质为本领域已知,例如绿色萦光蛋白(gfp)、红色萦光蛋白(rfp)、蓝色萦光蛋白(bfp,heim等1994,1996)、称为cfp的青色萦光变体(heim等1996;tsien1998)、称为yfp的黄色萦光变体(ormo等1996;wachter等1998)、称为蓝宝石(sapphire)的紫外激发绿色萦光变体(tsien1998;zapata-hommer等2003)、称为增强型绿色萦光蛋白或egfp(yang等1996)且可通过例如活细胞成像(例如incucyte)或萦光分光光度法测量的青色激发绿色萦光变体。本文所用的术语“抗体或配体的功能性”指抗体或配体的生物学活性,例如抗体或配体引出细胞反应的能力。例如,通过结合靶抗原,抗体激活或抑制细胞信号传导途径,即激活或抑制靶抗原的功能。例如,待测试抗体结合激活nf-κb途径的受体,并通过此结合激活细胞核中的反应元件。在此反应元件与报道基因连接时,可以在本发明的测定中容易地监测该激活。术语―功能性”还包括抗体的效应子功能,例如,c1q结合和依赖补体的细胞毒性(cdc);fc受体结合;依赖抗体的细胞毒性(adcc);依赖抗体的吞噬作用(adcp)、细胞因子分泌、免疫复合物介导的抗原呈递细胞的抗原摄取;细胞表面受体(例如b细胞受体)的下调;及b细胞激活。本文所用的“靶抗原”指抗体或其片段可以靶向的任何细胞表面抗原。它还指配体可以靶向的受体。“靶抗原反应元件”指特异性转录因子结合元件,或通过结合某些转录因子激活或沉默的顺式作用元件。在一个实施方案中,该反应元件是位于最小启动子(例如tata框启动子)上游的顺式作用增强子元件,该最小启动子通过转录因子结合驱动报道基因的表达。本文所用的“nf-κb”指“激活b细胞核因子κ轻链增强子”,其参与调节许多编码凋亡、病毒复制、肿瘤发生、多种自身免疫病和炎症反应的介质的基因。nf-κb存在于几乎所有真核细胞中。通常,如果与抑制性κb(iκb)蛋白质形成复合物,则它以无活性状态定位在细胞质中。通过配体与完整膜受体(也称为“nf-κb途径受体”)的结合,iκb激酶(ikk)激活。ikk是由两种激酶和调节亚基组成的酶复合物。此复合物磷酸化iκb蛋白质,导致泛素化,从而通过蛋白酶体降解那些蛋白质。最后,游离nfκb处于活性状态,转运(translocate)至细胞核,结合κbdna元件,诱导靶基因的转录。本文所用的“κbdna途径”指导致nf-κb活性的调节的刺激。例如,通过配体或抗体的结合激活toll样受体信号发放、tnf受体信号发放、t细胞受体和b细胞受体信号发放导致nf-κb的激活。随后,磷酸化的nf-κb二聚体结合κbdna元件,诱导靶基因的转录。本文所用的示例性κbdna元件称为“nf-κb途径反应元件”。因此,“nf-κb途径受体”指可以触发nf-κb活性调节的受体。“nf-κb途径受体的实例”是toll样受体、tnf受体、t细胞受体和b细胞受体。与其靶标结合时导致nf-κb活性调节的抗体的非限制性实例是抗cd40抗体、抗dr5抗体、抗dr4抗体、抗41bb抗体、抗ox40抗体和抗gitr抗体。与其靶标结合时导致nf-κb活性调节的配体的非限制性实例是ox40配体、4-1bb配体或cd40配体。本文所用的“高通量筛选”应理解为指可用本文所述的新测定针对结合和功能性分析相对大数量的不同候选抗体或配体。典型的这种高通量筛选在多孔微量滴定板中,例如在96孔板或384孔板或具有1536或3456个孔的板中进行。术语“能量供体化合物”指萦光共振能量转移(fret)能量供体化合物、生物发光能量转移(bret)能量供体化合物和alphascreen供体小球。术语“能量受体化合物”指萦光共振能量转移(fret)能量受体化合物、生物发光能量转移(bret)能量受体化合物和alphascreen受体小球。术语“fret”指发生在两个生色团之间的萦光共振能量转移过程。本文所用的生色团包括例如萦光、发光和其他非萦光成分。“fret”、“萦光共振能量转移”、共振能量转移”和“共振能量转移”在本文中可互换使用。本文所用的术语“时间分辨fret”指基于受体萦光体的发射的时间分辨检测的发生在两个生色团之间的能量转移过程(morrison,l.e.,1988.anal.biochem.,174(1)101)。本文中所用的“fret能量供体化合物”也称为“fret供体”或“fret能量供体”,指用于fret且为本领域已知的供体萦光团。能量供体化合物的非限制性实例在表b中列出。优选地,fret能量供体化合物是稀土元素,例如铽穴合物(“lumi4-tb”)和铕穴合物(eu3+穴合物)。适合用于该方法的镧系螯合物包括描述于例如美国专利号5,622,821、5,639,615、5,656,433和4,822,733中的那些。本文所用的“fret能量受体化合物”也称为“fret受体”或“fret能量受体”,指用于fret且为本领域已知的受体萦光团,例如alexa萦光染料(lifetechnologies)或cy5、yfp、fitc、化学修饰的别藻蓝蛋白(allophycocyanine)(xl665)、d2。能量受体化合物的其他非限制性实例在表b中列出。优选地,该fret能量受体化合物是d2。术语“bret”指化学发光供体部分(即bret能量供体化合物)和萦光受体部分(即bret能量受体化合物)之间的共振能量转移(ret)。术语“alphascreen”指放大发光亲和均相测定(amplifiedluminescentproximityhomogeneousassay),已描述于例如―luminescentoxygenchannelingimmunoassay:measurementofparticlebindingkineticsbychemiluminescence.”ullman,ef等proc.natl.acad.sci.usa,91卷,5426–5430页,1994年6月中。“亲和力”指分子(例如抗体或配体)的单个结合部位与其结合配偶体(例如抗原或受体)之间非共价相互作用的总和的强度。除非另有说明,本文所用的“结合亲和力”指内在的结合亲和力,其反映结合对(例如抗体和抗原)的成员之间的1:1相互作用。分子x对其配偶体y的亲和力一般可通过解离常数(kd)来表示。亲和力可以通过本领域公知的方法来测量,包括本文中所述的那些。用于测量结合亲和力的具体的说明性和示例性实施方案在下文中描述。“亲和力成熟的”抗体指与不具有这类改变的亲本抗体相比在一个或多个高变区(hvr)中具有一个或多个改变的抗体,这类改变导致抗体对抗原的亲和力的改善。本文所用的术语“配体”指能够结合另一分子的任何分子。配体分子的实例包括但不限于肽、蛋白质、糖类、脂质或核酸。待用本文所述测定分析的优选配体是能够结合靶抗原的肽或蛋白质。通常,这种靶抗原是细胞表面受体。术语“抗体”在本文中以最广泛的含义使用并涵盖多种抗体结构,包括但不限于单克隆抗体、多克隆抗体、多特异性抗体(例如双特异性抗体)和抗体片段,只要它们显示希望的抗原结合活性。“抗体片段”指除完整抗体以外的包含完整抗体的部分的分子,其结合该完整抗体所结合的抗原。抗体片段的实例包括但不限于fv、fab、fab'、fab’-sh、f(ab')2;双抗体;交叉fab片段;线性抗体;单链抗体分子(例如scfv);及从抗体片段形成的多特异性抗体。scfv抗体例如描述于houston,j.s.,methodsinenzymol.203(1991)46-96)中。此外,抗体片段包括具有vh结构域的特征(即能够与vl结构域一起组装为功能性抗原结合部位,从而提供全长抗体的抗原结合特性)或vl结构域的特征(即能够与vh结构域一起组装为功能性抗原结合部位,从而提供全长抗体的抗原结合特性)的单链多肽。本文所用的“fab片段”指包含含有轻链的vl结构域和恒定结构域(cl)的轻链片段及重链的vh结构域和第一恒定结构域(ch1)的抗体片段。术语“免疫球蛋白分子”指具有天然存在的抗体的结构的蛋白质。例如,igg种类的免疫球蛋白是由二硫键键合的两条相同的轻链和两条相同的重链组成的、约150,000道尔顿的异源四聚体糖蛋白。从n端至c端,每条重链具有可变区(vh),也称为可变重链结构域或重链可变结构域,随后是三个恒定结构域(ch1、ch2和ch3),也称为重链恒定区。类似地,从n端至c端,每条轻链具有可变区(vl),也称为可变轻链结构域或轻链可变结构域,随后是恒定轻链(cl)结构域,也称为轻链恒定区。可将免疫球蛋白的重链分配至称为α(iga)、δ(igd)、ε(ige),γ(igg)或μ(igm)的五种类型之一,其中一些可进一步分为亚型,例如γ1(igg1)、γ2(igg2)、γ3(igg3)、γ4(igg4)、α1(iga1)和α2(iga2)。根据其恒定结构域的氨基酸序列,可以将免疫球蛋白的轻链分配至称为κ和λ的两种类型之一。免疫球蛋白基本上由通过免疫球蛋白铰链区连接的两个fab分子和fc结构域组成。术语“抗原结合结构域”指抗原结合分子的部分,其包含特异性结合抗原的部分或全部并与之互补的区域。在抗原较大时,抗原结合分子可以仅结合抗原的特定部分,该部分称为表位。抗原结合结构域可以由例如一个或多个抗体可变结构域(也称为抗体可变区)提供。优选地,抗原结合结构域包含抗体轻链可变区(vl)和抗体重链可变区(vh)。术语“嵌合”抗体指这样的抗体,其中部分重链和/或轻链源自特定来源或物种,而重链和/或轻链的其余部分源自不同的来源或物种,通常通过重组dna技术制备。优选包含兔可变区和人恒定区的嵌合抗体。本发明所涵盖的“嵌合抗体”的其他优选形式是这样的形式,其中恒定区已从原抗体的恒定区修饰或改变,以产生本发明的尤其是就c1q结合和/或fc受体(fcr)结合而言的特性。这类嵌合抗体也称为“种类转换抗体”。嵌合抗体是所表达的包含编码免疫球蛋白可变区的dna区段和编码免疫球蛋白恒定区的dna区段的免疫球蛋白基因的产物。用于产生嵌合抗体的方法涉及本领域公知的常规重组dna和基因转染技术。参见例如morrison,s.l.等,proc.natl.acad.sci.usa81(1984)6851-6855;美国专利号5,202,238和5,204,244。“效应子功能”指可归因于抗体fc区且随抗体同种型而变的那些生物活性。抗体效应子功能的实例包括:c1q结合和依赖补体的细胞毒性(cdc);fc受体结合;依赖抗体的细胞毒性(adcc);依赖抗体的吞噬作用(adcp);细胞因子分泌;免疫复合物介导的抗原呈递细胞的抗原摄取;细胞表面受体(例如b细胞受体)的下调;及b细胞激活。认为本文所用的术语“改造”包括对天然存在的多肽或重组多肽或其片段的肽骨架或翻译后修饰的任意操作。改造包括氨基酸序列、糖基化模式或单个氨基酸的侧链基团的修饰,以及这些途径的组合。本文所用的术语“氨基酸突变”意在涵盖氨基酸取代、缺失、插入和修饰。可以产生取代、缺失、插入和修饰的任意组合来达到最终的构建体,只要最终构建体具有希望得到的特征,例如与fc受体的结合减少或与另一肽的结合增加。氨基酸序列缺失和插入包括氨基端和/或羧基端的氨基酸缺失和插入。为了改变例如fc区的结合特征的目的,尤其优选非保守氨基酸取代,即用另一具有不同结构和/或化学特性的氨基酸取代氨基酸。氨基酸取代包括用非天然存在的氨基酸或用二十种标准氨基酸的天然存在的氨基酸衍生物(例如4-羟脯氨酸、3-甲基组氨酸、鸟氨酸、高丝氨酸、5-羟赖氨酸)取代。氨基酸突变可以用本领域公知的遗传或化学方法产生。遗传方法可以包括定位诱变、pcr、基因合成等。考虑可以使用通过遗传改造之外的方法(如化学修饰)改变氨基酸的侧链基团的方法。例如,fc结构域329位脯氨酸至甘氨酸的取代可以表示为329g、g329、g329、p329g或pro329gly。活性剂(例如药物制剂)的“有效量”指对在必要的剂量和时期下达到希望得到的治疗或预防结果有效的量。本文的术语“fc结构域”或“fc区”用于定义免疫球蛋白重链的c端区域,其包含至少部分恒定区。该术语包括天然序列fc区和变体fc区。虽然igg重链fc区的边界可以略有变动,但人igg重链fc区通常定义为从cys226或从pro230延伸至重链的羧基端。但是,fc区的c端赖氨酸(lys447)可以存在或不存在。除非本文另有说明,fc区或恒定区中氨基酸残基的编号按照kabat等,sequencesofproteinsofimmunologicalinterest,第5版publichealthservice,nationalinstitutesofhealth,bethesda,md,1991中所述的eu编号系统,也称为eu指数。本文所用的fc结构域的“亚基”指形成二聚体fc结构域的两条多肽之一,即包含免疫球蛋白重链的c端恒定区、能够稳定地自我结合的多肽。例如,iggfc结构域的亚基包含iggch2和iggch3恒定结构域。“促进fc结构域的第一和第二亚基的结合的修饰”是fc结构域亚基的肽骨架或翻译后修饰的操作,其减少或阻止包含fc结构域亚基的多肽与相同多肽结合形成同二聚体。本文所用的促进结合的修饰尤其包括对希望结合的两个fc结构域亚基(即fc结构域的第一和第二亚基)中的每一个进行的分开的修饰,其中修饰彼此互补,以促进两个fc结构域亚基结合。例如,促进结合的修饰可以改变fc结构域亚基之一或二者的结构或电荷,以分别使它们的结合在空间上或静电上有利。因此,在包含第一fc结构域亚基的多肽和包含第二fc结构域亚基的多肽之间发生(异)二聚化,在与各亚基融合的其他成分(例如抗原结合部分)不同的意义上,两条多肽可以是不同的。在一些实施方案中,促进结合的修饰包括fc结构域中的氨基酸突变,尤其是氨基酸取代。在具体实施方案中,促进结合的修饰包括fc结构域两个亚基的每一个中分开的氨基酸突变,尤其是氨基酸取代。术语“全长抗体”、“完整抗体”和“全抗体”在本文中可互换使用,指具有基本类似于天然抗体结构的结构或具有包含本文所定义的fc区的重链的抗体。术语“宿主细胞”、“宿主细胞系”和“宿主细胞培养物”可互换使用,指已将外源核酸引入其中的细胞,包括这类细胞的后代。宿主细胞包括“转化体”和“转化细胞”,其包括最初转化的细胞及从其衍生的后代,而不考虑传代数。后代的核酸含量可以并非与亲本细胞完全相同,而是可以包含突变。本文包括具有与在最初转化的细胞中筛选或选择的功能或生物学活性相同的功能和生物学活性的突变体后代。“人抗体”是具有这样的氨基酸序列的抗体,该氨基酸序列对应于由人或人细胞产生或源自利用人抗体库或其他人抗体编码序列的非人来源的抗体的氨基酸序列。人抗体的此定义明确排除了包含非人抗原结合残基的人源化抗体。如针对本发明的嵌合和人源化抗体所提到,本文所用的术语“人抗体”还包括例如通过“种类转换”(即fc部分的改变或突变(例如igg1至igg4和/或igg1/igg4突变))在恒定区中修饰以产生本发明的尤其是就c1q结合和/或fcr结合而言的特性的这类抗体。本文所用的术语“重组人抗体”旨在包括通过重组手段制备、表达、产生或分离的所有人抗体,如从诸如ns0或cho细胞的宿主细胞或从人免疫球蛋白基因的转基因动物(例如小鼠)分离的抗体,或用转染入宿主细胞的重组表达载体表达的抗体。这类重组人抗体具有重排形式的可变区和恒定区。本发明的重组人抗体已经历了体内体细胞高变。因此,重组抗体的vh和vl区的氨基酸序列是这样的序列,其虽然源自人种系vh和vl序列并与人种系vh和vl序列相关,但可以并非在体内天然存在于人抗体种系库内。用于本文目的的“受体人构架”是包含源自下文定义的人免疫球蛋白构架或人共有构架的轻链可变结构域(vl)构架或重链可变结构域(vh)构架的氨基酸序列的构架。“源自”人免疫球蛋白构架或人共有构架的受体人构架可以包含其相同氨基酸序列,或者它可以包含氨基酸序列改变。在一些实施方案中,氨基酸改变的数目为10或更少、9或更少、8或更少、7或更少、6或更少、5或更少、4或更少、3或更少、或2或更少。在一些实施方案中,vl受体人构架在序列上与vl人免疫球蛋白构架序列或人共有构架序列相同。“构架”或“fr”指高变区(hvr)残基之外的可变结构域残基。可变结构域的fr一般由四个fr结构域组成:fr1、fr2、fr3和fr4。因此,hvr和fr序列一般按以下顺序出现在vh(或vl)中:fr1-h1(l1)-fr2-h2(l2)-fr3-h3(l3)-fr4。“人共有构架”是代表一系列人免疫球蛋白vl或vh构架序列中最常出现的氨基酸残基的构架。通常,该系列人免疫球蛋白vl或vh序列来自可变结构域序列亚组。通常,该序列亚组是如kabat等,sequencesofproteinsofimmunologicalinterest,第5版,nihpublication91-3242,bethesdamd(1991),1-3卷中的亚组。在一个实施方案中,对于vl,该亚组是如kabat等,上文中的κi亚组。在一个实施方案中,对于vh,该亚组是kabat等,上文中的iii亚组。“人源化”抗体指包含来自非人hvr的氨基酸残基和来自人fr的氨基酸残基的嵌合抗体。在某些实施方案中,人源化抗体将包含至少一个且通常为两个可变结构域的基本上全部,其中全部或基本上全部hvr(例如cdr)对应于非人抗体的那些,而全部或基本上全部fr对应于人抗体的那些。人源化抗体可以可选地包含源自人抗体的抗体恒定区的至少一部分。抗体(例如非人抗体)的“人源化形式”指已进行了人源化的抗体。本发明所涵盖的“人源化抗体”的其他形式是这样的形式,其中恒定区已从原抗体的恒定区额外地修饰或改变,以产生本发明的尤其是就c1q结合和/或fc受体(fcr)结合而言的特性。本文所用的术语“高变区”或“hvr”指抗体可变结构域在序列上高变和/或形成结构确定的环(“高变环”)的区域中的每一个。通常,天然四链抗体包含六个hvr;三个在vh中(h1、h2、h3),三个在vl中(l1、l2、l3)。hvr一般包含来自高变环和/或来自“互补决定区”(cdr)的氨基酸残基,后者具有最高序列变异性和/或涉及抗原识别。示例性高变环存在于氨基酸残基26-32(l1)、50-52(l2)、91-96(l3)、26-32(h1)、53-55(h2)和96-101(h3)(chothia和lesk,j.mol.biol.196:901-917(1987))。示例性cdr(cdr-l1、cdr-l2、cdr-l3、cdr-h1、cdr-h2和cdr-h3)存在于l1的氨基酸残基24-34、l2的氨基酸残基50-56、l3的氨基酸残基89-97、h1的氨基酸残基31-35b、h2的氨基酸残基50-65和h3的氨基酸残基95-102(kabat等,sequencesofproteinsofimmunologicalinterest,第5版publichealthservice,nationalinstitutesofhealth,bethesda,md(1991))。高变区(hvr)也称为互补决定区(cdr),在提及形成抗原结合区的可变结构域的部分时,这些术语在本文中可互换使用。此特定区域已由kabat等,u.s.dept.ofhealthandhumanservices,"sequencesofproteinsofimmunologicalinterest"(1983)和chothia等,j.mol.biol.196:901-917(1987)描述,其中定义包括相互比较时的重叠或氨基酸残基亚组。然后,用任一种定义来提到抗体的cdr或其变体旨在处于本文所定义和使用的术语的范围之内。下文表a中显示涵盖上文引用的参考文献中的每一篇所定义的cdr的适当氨基酸残基。涵盖特定cdr的确切残基编号将取决于cdr的序列和大小而变。鉴于抗体的可变区氨基酸序列,本领域技术人员可以常规确定哪些残基包含特定cdr。表a.cdr定义1cdrkabatchothiaabm2vhcdr131-3526-3226-35vhcdr250-6552-5850-58vhcdr395-10295-10295-102vlcdr124-3426-3224-34vlcdr250-5650-5250-56vlcdr389-9791-9689-971表a中所有cdr定义的编号按照kabat等所述的编号惯例(参见下文)。2表a中所用的带有小写字母“b”的“abm”指oxfordmolecular's"abm"抗体建模软件所定义的cdr。kabat等还定义了适用于任何抗体的可变区序列编号系统。本领域普通技术人员可以将此“kabat编号”系统明确分配至任何可变区序列,而不依赖于序列本身之外的任何实验数据。“kabat编号”指kabat等,u.s.dept.ofhealthandhumanservices,"sequenceofproteinsofimmunologicalinterest"(1983)所述的编号系统。除非另有说明,对抗体可变区中具体氨基酸残基位置编号的引用是按照kabat编号系统。除vh中的cdr1外,cdr通常包含形成高变环的氨基酸残基。cdr还包含“特异性决定残基”或“sdr”,其是接触抗原的残基。sdr包含在cdr的称为缩短的cdr或a-cdr的区域内。示例性a-cdr(a-cdr-l1、a-cdr-l2、a-cdr-l3、a-cdr-h1、a-cdr-h2和a-cdr-h3)存在于l1的氨基酸残基31-34、l2的氨基酸残基50-55、l3的氨基酸残基89-96、h1的氨基酸残基31-35b、h2的氨基酸残基50-58和h3的氨基酸残基95-102处。(参见almagro和fransson,front.biosci.13:1619-1633(2008))。除非另有说明,本文按照kabat等,上文编号可变结构域中的hvr残基和其他残基(例如fr残基)。“免疫缀合物”是与一个或多个异源分子缀合的抗体,该异源分子包括但不限于细胞毒剂。“个体(individual)”或“对象(subject)”是哺乳动物。哺乳动物包括但不限于饲养动物(例如牛、绵羊、猫、狗和马)、灵长类(例如人类和非人灵长类,如猴)、兔和啮齿类(例如小鼠和大鼠)。在某些实施方案中,该个体或对象是人。“分离的”抗体是已从其天然环境的成分分开的抗体。在一些实施方案中,将抗体纯化至通过例如电泳(例如sds-page、等电聚焦(ief)、毛细管电泳)或层析(例如离子交换或反相hplc)测定的纯度大于95%或99%。用于评估抗体纯度的方法综述参见例如flatman等,j.chromatogr.b848:79-87(2007)。“分离的”核酸指已从其天然环境的成分分开的核酸分子。分离的核酸包括包含在通常含有该核酸分子的细胞中的核酸分子,但该核酸分子存在于染色体外或不同于其天然染色体位置的染色体位置上。本文所用的术语“单克隆抗体”指获自基本同质的抗体群体的抗体,即除了例如包含天然存在的突变或在产生单克隆抗体制剂期间出现的可能的变体抗体(这类变体通常以较小的量存在)外,包含该群体的单种抗体相同和/或结合相同的表位。与通常包含抗不同决定簇(表位)的不同抗体的多克隆抗体制剂不同,单克隆抗体制剂的每种单克隆抗体抗抗原上的单个决定簇。因此,修饰词“单克隆”指抗体获自基本同质的抗体群体的特征,而不解释为需要通过任意具体方法来产生该抗体。例如,将要按照本发明使用的单克隆抗体可以通过多种技术制备,其包括但不限于杂交瘤法、重组dna法、噬菌体展示法及利用含有全部或部分人免疫球蛋白基因座的转基因动物的方法,本文描述用于制备单克隆抗体的这类方法和其他示例性方法。“裸抗体”指未与异源部分(例如细胞毒性部分)或放射性标记缀合的抗体。裸抗体可以存在于药物制剂中。“天然抗体”指天然存在的具有不同结构的免疫球蛋白分子。例如,天然igg抗体是由二硫键键合的由两条相同的轻链和两条相同的重链组成的、约150,000道尔顿的异源四聚体糖蛋白。从n端至c端,每条重链具有可变区(vh),也称为可变重链结构域或重链可变结构域,随后是三个恒定结构域(ch1、ch2和ch3)。类似地,从n端至c端,每条轻链具有可变区(vl),也称为可变轻链结构域或轻链可变结构域,随后是恒定轻链(cl)结构域。根据其恒定结构域的氨基酸序列,可以将抗体轻链分配至称为κ和λ的两个类型之一。术语“包装说明书”用来指通常包含在治疗产品的商业包装中的说明书,其包含关于适应症、用法、剂量、施用、联合治疗、禁忌症的信息和/或有关这类治疗性产品的用途的警告。“无实质性交叉反应”是指,尤其是在与靶抗原相比时,分子(例如抗体)不识别或特异性结合不同于该分子的实际靶抗原的抗原(例如与该靶抗原密切相关的抗原)。例如,抗体可以结合不到约10%至不到约5%的不同于实际靶抗原的抗原,或可以按不到约10%、9%、8%7%、6%、5%、4%、3%、2%、1%、0.5%、0.2%或0.1%、优选不到约2%、1%或0.5%、最优选不到约0.2%或0.1%的不同于实际靶抗原的抗原。就参考多肽序列而言的“百分比(%)氨基酸序列同一性”定义为,在比对序列并在必要时引入缺口来达到最大百分比序列同一性,而不将任意保守取代视为序列同一性的部分之后,候选序列中与参考多肽序列中的氨基酸残基相同的氨基酸残基的百分比。可以以本领域技术之内的多种方式来达到以测定百分比氨基酸序列同一性为目的的比对,例如,使用公开可得的计算机软件,如blast、blast-2、align或megalign(dnastar)软件。本领域技术人员可以确定用于比对序列的适当参数,包括在所比较的序列全长内达到最大比对所需的任意算法。但是,为了本文的目的,用序列比较计算机程序align-2来产生%氨基酸序列同一性值。align-2序列比较计算机程序由genentech,inc.编写,源代码已随用户文件提交美国版权办公室,washingtond.c.,20559,它在美国版权办公室注册在美国版权注册号txu510087之下。align-2程序从genentech,inc.,southsanfrancisco,california公开可得,或者可以从源代码编译。align-2程序应编译用在unix操作系统上,包括数字unixv4.0d。所有序列比较参数由align-2程序设定且不变动。在利用align-2来进行氨基酸序列比较的情况下,按以下计算给定氨基酸序列a与给定氨基酸序列b的%氨基酸序列同一性(备选地,其可以叙述为具有或包含与给定氨基酸序列b的某%氨基酸序列同一性的给定氨基酸序列a):100乘以分数x/y,其中x是在该程序的a和b比对中通过序列比对程序align-2评分为相同的匹配氨基酸残基的数目,且其中y是b中氨基酸残基总数。应理解,在氨基酸序列a的长度不等于氨基酸序列b的长度时,a与b的%氨基酸序列同一性将不等于b与a的%氨基酸序列同一性。除非另有明确说明,本文中使用的所有%氨基酸序列同一性值都是用align-2计算机程序按前一段落中所述获得。术语“药物制剂”指这样的制备物,其处于这样的形式,使得包含在其中的活性成分的生物活性有效,且不包含对将要施用该制剂的个体具有不可接受的毒性的附加成分。“可药用载体”指药物制剂中除活性成分外的成分,其对个体无毒性。可药用载体包括但不限于缓冲剂、赋形剂、稳定剂或防腐剂。本文所用的“处理”(及其语法变形)指改变所处理的个体的自然过程的尝试中的临床干预,且可以为了预防而进行或在临床病理的过程中进行。希望得到的处理作用包括但不限于防止疾病的发生或复发、减轻症状、减少疾病的任何直接或间接的病理结果、防止转移、降低疾病进展的速率、改善或缓和疾病状态及消退或改善的预后。术语“可变区”或“可变结构域”指抗体重链或轻链的涉及该抗体与抗原结合的结构域。天然抗体的重链和轻链的可变结构域(分别为vh和vl)通常具有相似的结构,每个结构域包含四个保守的构架区(fr)和三个高变区(hvr)(参见例如kindt等kubyimmunology,第6版,w.h.freemanandco.,第91页(2007))。单个vh或vl结构域可以足以赋予抗原结合特异性。此外,可以分别用来自结合该抗原的抗体的vh或vl结构域筛选互补vl或vh结构域的文库来分离结合特定抗原的抗体。参见例如portolano等,j.immunol.150:880-887(1993);clarkson等,nature352:624-628(1991)。在本文中使用时,术语“抗体的抗原结合部位”指抗体的负责抗原结合的氨基酸残基。抗体的抗原结合部分包含来自“互补决定区”或“cdr”的氨基酸残基。“构架”或“fr”区是除本文定义的高变区残基外的那些可变结构域残基。因此,抗体的轻链和重链可变结构域从n端至c端包含fr1、cdr1、fr2、cdr2、fr3、cdr3和fr4。尤其是,重链的cdr3是对抗原结合贡献最多并定义抗体特性的区域。cdr和fr区按照kabat等,sequencesofproteinsofimmunologicalinterest,第5版,publichealthservice,nationalinstitutesofhealth,bethesda,md(1991)的标准定义确定和/或是来自“高变环”的那些残基。抗体特异性指抗体对抗原的特定表位的选择性识别。例如天然抗体是单特异性的。本文所用的术语“单特异性”抗体指具有一个或多个结合部位的抗体,其中每个结合部位结合同一抗原的同一表位。本文所用的术语“双特异性”抗体指具有至少两个结合部位的抗体,每个结合部位结合同一抗原或不同抗原的不同表位。用于制备多特异性抗体的技术包括但不限于重组共表达两个具有不同特异性的免疫球蛋白重链-轻链对(参见milstein和cuello,nature305:537(1983);wo93/08829;及traunecker等,emboj.10:3655(1991))和“杵入臼”改造(参见例如美国专利号5,731,168)。还可以通过以下来制备多特异性抗体:用于制备抗体fc-异二聚体分子的工程静电引导作用(wo2009/089004);交联两种或多种抗体或片段(参见例如美国专利号4,676,980和brennan等,science,229:81(1985));用亮氨酸拉链来产生双特异性抗体(参见例如kostelny等,j.immunol.,148(5):1547-1553(1992));使用用于制备双特异性抗体片段的“双抗体”技术(参见例如hollinger等,proc.natl.acad.sci.usa,90:6444-6448(1993));使用单链fv(sfv)二聚体(参见例如gruber等,j.immunol.,152:5368(1994));按例如tutt等j.immunol.147:60(1991)中所述制备三特异性抗体。本文还包括具有三个或多个功能性抗原结合部位的改造抗体,包括“章鱼抗体”(参见例如us2006/0025576a1)。本文的抗体或片段还可以包括“双重作用fab”或“daf”(参见例如us2008/0069820)。本申请内所用的术语“价”指抗体分子中存在指定数目的结合部位。因此,术语“二价”、“四价”和“六价”分别指抗体分子中存在两个结合部位、四个结合部位和六个结合部位。本发明的双特异性抗体至少为“二价”,且可以是“三价”或“多价”(例如“四价”或“六价”)。本发明的抗体具有两个或多个结合部位且是双特异性的。也就是说,即使在有两个以上结合部位(即该抗体为三价或多价)的情况下,该抗体也可以是双特异性的。本发明的双特异性抗体包括例如多价单链抗体、双抗体和三抗体,以及具有可通过一个或多个肽接头连接其他抗原结合部位(例如单链fv、vh结构域和/或vl结构域、fab或(fab)2)的全长抗体恒定结构域结构的抗体。抗体可以是来自单个物种的全长抗体,或嵌合或人源化抗体。本文所用的术语“载体”指能够扩增与它连接的另一核酸的核酸分子。该术语包括作为自主复制核酸结构的载体,以及掺入它所引入的宿主细胞基因组中的载体。某些载体能够指导与它们有效连接的核酸的表达。这类载体在本文中称为“表达载体”。本申请内所用的术语“氨基酸”指天然存在的羧基α-氨基酸,包括丙氨酸(三字母代码:ala,单字母代码:a)、精氨酸(arg,r)、天冬酰胺(asn,n)、天冬氨酸(asp,d)、半胱氨酸(cys,c)、谷氨酰胺(gln,q)、谷氨酸(glu,e)、甘氨酸(gly,g)、组氨酸(his,h)、异亮氨酸(ile,i)、亮氨酸(leu,l)、赖氨酸(lys,k)、甲硫氨酸(met,m)、苯丙氨酸(phe,f)、脯氨酸(pro,p)、丝氨酸(ser,s)、苏氨酸(thr,t)、色氨酸(trp,w)、酪氨酸(tyr,y)和缬氨酸(val,v)。本文所用的表述“细胞”、“细胞系”和“细胞培养物”可互换使用,且所有这类命名包括后代。因此,词语“转染体”和“转染细胞”包括最初的主题细胞和从其衍生的培养物,而不考虑转移的数目。还应理解,由于有意或无意的突变,所有后代可以在dna含量上并非完全相同。本文包括具有与在最初转化的细胞中筛选的功能或生物学活性相同的功能或生物学活性的变体后代。“亲和力”指分子(例如抗体或配体)的单个结合部位与其结合配偶体(例如抗原)之间非共价相互作用的总和的强度。除非另有说明,本文所用的“结合亲和力”指内在的结合亲和力,其反映结合对(例如抗体和抗原/配体和受体)的成员之间的1:1相互作用。分子x对其配偶体y的亲和力一般可通过解离常数(kd)来表示。亲和力可以通过本领域公知的方法来测量,包括本文中所述的那些。用于测量结合亲和力的具体的说明性和示例性实施方案在下文中描述。本文所用的术语“结合”或“特异性结合”指优选通过萦光共振能量转移(fret)进行的体外测定中,抗体与抗原表位的结合或配体与受体的结合。术语“表位”包括能够与抗体特异性结合的任意多肽决定簇。在某些实施方案中,表位决定簇包括诸如氨基酸、糖侧链、磷酰基或磺酰基的分子的具有化学活性的表面分组,且在某些实施方案中可以具有特异的三维结构特征,或特异的电荷特征。表位是抗体结合的抗原区域。本文所用的尤其是加上前缀―糖”的术语―改造”以及术语―糖基化改造”视为包括天然存在的多肽或重组多肽或其片段的糖基化模式的任何操作。糖基化改造包括细胞糖基化机器的代谢性改造,包括寡糖合成途径的遗传操作,以达到改变细胞中表达的糖蛋白的糖基化。此外,糖基化改造包括突变和细胞环境对糖基化的作用。在一个实施方案中,糖基化改造是糖基转移酶活性的改变。在具体实施方案中,改造导致葡糖氨基转移酶活性和/或岩藻糖基转移酶活性的改变。ii.新测定发明人开发了适合用于高通量型式(384孔)的测定,该测定使得能够在一个孔或小管中组合分析抗体或配体结合和功能性。通过使用在激活反应元件时具有报道基因表达的转染报告细胞系来评价抗体或配体的功能性(例如,抗体或配体的生物学活性,例如,抗体或配体引出细胞反应,例如激活或抑制靶抗原的能力)。在一个实施方案中,该报道基因选自编码萦光蛋白(例如绿色萦光蛋白,gfp)的基因和/或编码可检测其催化活性的酶(例如萦光素酶)的基因。为了研究结合,应用基于细胞的fret方法、bret或alphascreen。在一个实施方案中,提供用于测定特异性结合靶抗原的抗体或配体的结合和功能性的体外测定,其包括以下步骤:i)提供细胞,该细胞a)在其表面表达靶抗原;b)用能量供体化合物共价或非共价标记;和c)包含靶抗原反应元件控制下的报道基因;ii)加入待测试的抗体或配体;iii)通过测定能量转移来测量与靶抗原的结合,其中能量受体化合物共价或非共价缀合于待测试的抗体或结合一抗的二抗;和iv)通过将报道基因表达水平与靶抗原激活或抑制水平相关,来测定抗体或配体的功能性。在一个实施方案中,该能量转移供体和受体化合物是萦光共振能量转移(fret)能量供体和受体化合物,在步骤iii)中测定的能量转移是萦光共振能量转移(fret)。在一个实施方案中,该fret是时间分辨fret。时间分辨fret是以微量滴定板型式研究蛋白质-蛋白质相互作用的技术。时间分辨萦光共振能量转移(tr-fret)应用基于供体和受体分子之间的能量转移。fret是从萦光供体分子至适当受体分子的非辐射性能量转移。在通过光源激发供体分子时,它产生萦光。如果受体分子很靠近,且供体分子的发射光谱与受体分子的激发光谱重叠,则供体分子可以将其激发能转移至受体分子而不是发射萦光。出现受体发射波长的萦光表示供体和受体分子相互很靠近,因为供体和受体分子需要相隔不到约20nm(例如相隔5-10nm)才能发生能量转移。通常,通过生物亲和相互作用(例如蛋白质-蛋白质结合、抗原-抗体结合、配体-受体结合、dna杂交和dna-蛋白质结合)达到供体和受体分子之间的靠近。存在多种不同的供体和受体分子。通常,用于fret测定的供体和受体分子是具有短半衰期的萦光团。来自样品成分如缓冲液、蛋白质、化学化合物和细胞裂解物的背景萦光可以降低传统fret化学的性能。必须针对此自发萦光矫正所检测到的萦光强度,这降低了测定灵敏度,并可使结果解释变得复杂。这类背景萦光是瞬时的(半衰期在纳秒范围内),因此可以用时间分辨方法消除。tr-fret利用镧系离子如铕(eu)、铽(tb)、钐(sm)和镝(dy)离子的独特特性。由于它们特异的光物理和光谱特性,稀土离子的络合物对生物学中的萦光应用具有重要意义。具体而言,与更传统的萦光团相比,它们具有大的stoke位移(stoke'sshifts)和长发射半衰期(从微秒至毫秒)。fret能量受体可在perkinelmer(例如产品)、invitrogen(例如产品)和cisbiobioassays(例如产品)处购得。在一些实施方案中,可以根据下表中所列的一种或多种萦光团来选择fret能量供体和fret能量受体。表b:fret能量供体和能量受体分子的实例在选择fret能量供体和fret能量受体组合时,还可考虑以下信息。美国专利号5,998,146(在此引入作为参考)描述了镧系螯合物络合物尤其是铕和铽络合物与萦光团或淬灭剂组合的用途。它还描述了长寿命镧系螯合物络合物的特性。基于作为能量供体的金属络合物和作为能量受体的来自藻胆蛋白类的染料的fret系统为本领域已知(ep76695;hemmilae,chemicalanalysis117,johnwiley&sons,inc.,(1991)135-139)。已建立的市售系统(例如来自wallac、oy或cisbiopackard)使用由作为金属络合物的镧系螯合物和藻胆蛋白组成的fret对。镧系螯合物络合物尤其是铕或铽络合物的特性是已知的,可与淬灭剂组合以及与萦光团组合使用。钌络合物本身用作萦光团或发光团,尤其是对于电化学发光。钌螯合物络合物例如从ep178450和ep772616已知,其中还描述了用于将这些络合物与生物分子偶联的方法。其中未讨论它们在fret系统中作为能量供体的用途。别藻蓝蛋白具有诸如异常高的消光系数(约700000l/mcm)以及极高的发射系数的特性。这些对于它们在fret系统中作为萦光团受体的用途而言是有用的前提。此外,已知这些染料易溶于水且稳定。术语低分子萦光团指分子量在300和3000da之间的萦光染料。就重要特征而言,与apc相比,这类低分子萦光团(如呫吨、花青苷、罗丹明和噁嗪)具有显著优势。因此,例如它们的消光系数实质性更低,且在ca.100000l/mcm的范围内。在一些实施方案中,本发明的方法和测定利用同源tr-fret测定技术。tr-fret是时间分辨(trf)和fret的组合。trf通过使读取萦光信号的时间延迟例如10纳秒来降低背景萦光。此延迟(即门控期)之后,测量样品中持续时间更长的萦光。因此,使用tr-fret,例如可由样品中的干扰物质引起的干扰背景萦光不再共检测到,而是仅测量由能量转移产生或抑制的萦光。借助适当的测量装置测定tr-fret系统所产生的萦光。这类时间分辨检测系统使用例如脉冲激光二极管、发光二极管(led)或脉冲染料激光器作为发射光源。测量发生在适当的时间延迟之后,即干扰背景萦光衰减之后。本领域描述了用于测定时间分辨fret信号的装置和方法。此技术要求目的信号必须对应于具有长萦光寿命的化合物。这类长寿命萦光化合物是稀土稀土镧系元素。例如,eu3+具有毫秒级萦光寿命。tr-fret需要上文所述的fret能量供体和fret能量受体。与fret一样,tr-fret能量供体和受体对可以基于以下一项或多项(包括全部)来选择:(1)fret能量供体的发射光谱应与fret能量受体的激发光谱重叠;(2)fret配偶体(即fret能量供体和fret能量受体)的发射光谱应显示非重叠萦光;(3)fret量子产率(即从fret供体转移至fret受体的能量)应尽可能高(例如,fret在1-20nm(例如5-10nm)的测量距离内应具有月1-100%,例如30%、40%、50%、60%、70%、80%、85%、90%、95%、98%和99%效率);(4)fret信号(即萦光)必须可与样品产生的萦光(例如自发萦光)区分;和(5)fret供体和fret受体应具有允许检测fret信号的半衰期(例如,fret可以明亮,且可在从10-9秒至10-4秒范围内的时间尺度发生)。在一些实施方案中,tr-fret供体和tr-fret受体可以基于上表中所列萦光团中的一种或多种来选择。在一些实施方案中,tr-fret供体可以是镧系离子,例如结合于螯合剂的镧系离子。在一些实施方案中,该镧系离子可以是铕、铽、钐或镝离子。本文所用的eu包括eu和所有eu离子,例如eu3+。在一些实施方案中,tr-fret供体可以是dsred。在一些实施方案中,tr-fret供体可以是ri2。应理解,适当tr-fret供体的选择需要考虑上文所列的标准和所选择的具体tr-fret受体。在一些实施方案中,tr-fret受体可以选自萦光素、cy5、别藻蓝蛋白(apc—例如xl665、d2(cisbio)和bg-647)和萦光蛋白质(例如gfp、cfp、yfp、bfp、rfp和其他gfp变体)。在一些实施方案中,tr-fret供体可以是铽,tr-fret受体可以是萦光素。在一些实施方案中,tr-fret供体可以是eu,tr-fret受体可以是cy5或apc(例如xl665、d2(cisbio)、bg-647和cy5相关tr-fret受体)。在一个实施方案中,fret能量供体化合物是铽穴合物和/或fret能量受体化合物是d2。在一些实施方案中,tr-fret供体和tr-fret受体可以与增强tr-fret供体和/或tr-fret受体功能的第二化合物组合。例如,tr-fret供体和tr-fret受体可以与穴合物包裹组合来延长萦光团的半衰期。备选地,或此外,tr-fret供体和tr-fret受体可以与例如增强系统组合。在一些实施方案中,tr-fret供体和tr-fret受体可以与例如增强或延长tr-fret信号的寿命或检测的缓冲剂、盐、增强剂、螯合剂和稳定剂(例如光稳定剂)组合。可以用上文及美国专利号4,925,804、美国专利号5,637,509、美国专利号4,761,481、美国专利号4,920,195、美国专利号5,032,677、美国专利号5,202,423、美国专利号5,324,825、美国专利号5,457,186、美国专利号5,571,897、美国专利号7,250,517、us2005/0202565(在此引入作为参考)中所述的适宜的fret或tr-fret供体和受体直接或间接标记分子(例如蛋白质)。通过在供体激发和受体发射测量之间引入约50-150μs的时间延迟来以时间分辨的方式测量时间分辨fret。因此,测量不到由介质、生物制剂或受体直接激发产生的短寿命背景萦光,测定灵敏度提高。常用的受体分子是称为d2的有机红色萦光染料,其比xl665小100倍,显示与xl665非常相似的光物理特性。(amoravainm,lyotards,jagad,lebretonml,serventf,bomeru.introductionofanewhtrfacceptor,d2:developmentofacompletegpcrplatformforags,giandgqscreening.sbs;11thannualconference;geneva.2005.)。可以用多种发光仪和光检测仪中的任一种来起始fret(例如激发fret供体或激发能够激发fret供体的试剂)和/或检测从该fret产生的发射。可以目测、通过照相、通过感光测定、通过分光光度测定或通过任何其他方便的手段检测或测量由于上述方法而由fret对的第一和第二成员产生的光发射来测定其量,该量与介质中每种成分的量相关。可以定性(即通过fret信号的存在或缺乏)测定抗体或配体与靶抗原的结合;无任何fret信号指示无结合。通常,“无fret信号”定义为某个阈值,即扣除任何背景信号。背景信号通常通过用除待测试的抗体或配体外的所有试剂进行fret测定来测定。可以定量测定抗体或配体与靶抗原的结合,即可以用fret法测定结合的水平或强度。为此,以不同浓度测试待测试的抗体或配体,并测定半数最大有效浓度(ec50)。ec50指抗体或配体的浓度,在该浓度下,抗体或配体在指定的暴露时间后的结合达到基线和最大值之间的中点。剂量反应曲线的ec50因此代表观察到其最大结合的50%的抗体或配体浓度。kd(解离常数)可以通过本领域已知的方法从剂量反应曲线计算。在一个实施方案中,用生物发光共振能量转移(bret)测定抗体或配体与靶抗原的结合。因此,在一个实施方案中,能量供体和受体化合物是生物发光能量转移(bret)能量供体和受体化合物,步骤iii)中测定的能量转移是生物发光能量转移(bret)。bret测定技术基于生物发光供体部分和萦光受体部分之间有效的共振能量转移(ret)。bret是天然存在的现象,与fret的不同在于它用萦光素酶作为供体。在一个实施方案中,用分离自海肾(renillareniformis)的萦光素酶(rluc)和称为deepbluec(dbc)的腔肠萦光素(coelenterazine)作为供体。在氧存在下,rluc催化dbc转化为coelenteramide,伴随最大395nm(蓝光)的光发射。在紧靠适宜的受体时,通过ret捕获蓝光能量。在一个实施方案中,bret中的受体是改造为最大程度吸收rluc/dbc反应所发射的能量的gfp变体(gfp2)。ret激发gfp2产生510nm绿光发射。通过用受体发射强度除以供体发射强度来比例测定rluc/dbc和gfp2之间的能量转移效率。此比例测量称为bret信号,反应了rluc与gfp的靠近程度。在另一实施方案中,用rluc作为供体,腔肠萦光素的衍生物作为其底物,黄色萦光蛋白(yfp)作为受体。在一个实施方案中,用alphascreen(放大发光亲和均相测定)测定抗体或配体与靶抗原的结合。因此,在一个实施方案中,能量供体和受体化合物是alphascreen受体和供体小球,步骤iii)中测定的能量转移是受体小球内从单线态氧至二甲基噻吩衍生物的能量转移。alphascreen是使用―受体”和―供体”小球的非辐射、均相靠近测定。alphascreen是用于以微孔板型式研究生物分子相互作用的基于小球的检测系统。捕获在小球上的分子的结合导致从一个小球至另一个小球的能量转移,最终产生发光产物/萦光信号。每个alphascreen测定包含两种小球类型,供体小球和受体小球。两种小球类型都包被有最小化非特异性结合和自我聚集的水凝胶,提供反应性醛基用于将生物分子缀合至小球表面。每个alphascreen测定包含两种小球类型:供体小球和受体小球。每种小球类型包含不同化学药品组合,其是alphascreen技术的关键要素。供体小球包含光敏剂酞菁,其在680nm激光照射时将环境氧转化为单线态氧的激发形式。与其他激发分子一样,单线态氧在回到基态之前寿命有限。在4秒的半衰期内,单线态氧可以在溶液中扩散约200nm。如果受体小球在该距离内,则发生从单线态氧至受体小球内的二甲基噻吩衍生物的能量转移,然后以产生520-620nm光告终。在缺乏受体小球的情况下,单线态氧回到基态,不产生信号。在一个实施方案中,用于测定的细胞用结合于链霉抗生物素蛋白包被的供体小球的生物素化wga标记。在一个实施方案中,受体小球缀合于蛋白a。待测试的抗体可以容易地用这些受体小球标记,因为蛋白a结合抗体的fc结构域。标记用于bret、fret或alphascreen的细胞表面和待测试的抗体或配体或二抗的方法为本领域已知。例如,可以例如按yang等,analyticalbiochemistry351:158-160,2006(在此引入作为参考)所述直接或间接(例如通过标签,如非肽标签,如生物素、洋地黄毒苷、萦光素、二硝基苯酚,以及肽标签,如flag、histag、cmyc、ha、v5、streptag、acp/mcp标签,或使用抗生物素蛋白-链霉抗生物素蛋白相互作用)标记结合配偶体。acp标签和mcp标签是基于酰基载体蛋白的小蛋白质标签。acp标签或mcp标签与其底物(其是辅酶a的衍生物)间共价连接的形成需要所加入的合酶的存在。在标记反应中,重组合酶将缀合于coa的基团共价附着至acp标签或mcp标签。可以用用于acp标签标记的acp合酶(neb#p9301)或用于双重acp和mcp标签标记的sfp合酶(neb#p9302)将标记共价附着至标签。在一个实施方案中,用能量供体化合物共价或非共价标记靶抗原。例如,可以通过将它连接至20kdadna修复酶人o6-烷基鸟嘌呤-dna-烷基转移酶(agt)来标记待分析的蛋白质(例如本文中要分析的抗体或配体的靶抗原)。此酶可以对o6-苄基鸟嘌呤(bg)衍生物建立共价键,该衍生物本身与供体或受体萦光染料偶联。为了产生表达标记目的靶抗原的细胞,用编码与目的蛋白融合的人o6-烷基鸟嘌呤-dna-烷基转移酶(agt)的质粒转染细胞。转染后,用bg萦光团标记表达在细胞表面的融合蛋白质,然后可以用于基于fret的相互作用研究。优选地,用fret供体分子标记靶抗原。可以用能量供体或受体化合物(例如萦光染料或alphascreen小球)共价或非共价标记待分析的抗体或配体(其结合表达在细胞表面的标记靶抗原)。还可能标记结合待分析的抗体或配体的二抗(例如抗人igg)。也可以通过fret供体或受体分子标记的麦胚凝集素(wga)达到标记。因此,在一个实施方案中,将能量供体化合物共价或非共价连接至麦胚凝集素(wga)。因此,可以用供体分子标记细胞本身,而不是按上文所述转染细胞并标记融合蛋白质。wga以高亲和力结合诸如n-乙酰葡糖胺(glcnac)和n-乙酰神经氨酸(也称为唾液酸)的糖类。由于这类糖型在细胞表面普遍存在,缀合供体或受体萦光团的wga的结合允许随机标记细胞表面。然后可以通过从wga-铽至直接偶联待评价的抗体或配体或第二检测抗体的受体标记的能量转移,来检测结合包埋在细胞膜中的目的抗原的抗体或配体的相互作用。在一个实施方案中,报道基因选自编码萦光蛋白质的基因或编码可检测其催化活性的酶的基因。在一个实施方案中,萦光蛋白质选自绿色萦光蛋白(gfp)、黄色萦光蛋白(yfp)、红色萦光蛋白(rfp)、蓝色萦光蛋白(bfp,heim等1994,1996)、称为cfp的青色萦光变体(heim等1996;tsien1998)、称为yfp的黄色萦光变体(ormo等1996;wachter等1998)、称为蓝宝石的紫外激发绿色萦光变体(tsien1998;zapata-hommer等2003)、称为增强型绿色萦光蛋白或egfp(yang等1996)且可通过例如活细胞成像(例如incucyte)或萦光分光光度法测量的青色激发绿色萦光变体。在一个实施方案中,可检测其催化活性的酶选自萦光素酶、β半乳糖苷酶、碱性磷酸酶。在一个优选实施方案中,报道基因编码gfp。在一个实施方案中,报道基因编码萦光素酶。萦光素酶的活性可以通过市售测定,例如通过luciferase1000assaysystem或one-glotmluciferaseassaysystem(均来自promega)来检测。luciferase1000assaysystem除作为底物的萦光素外还包含辅酶a(coa),产生持续至少一分钟的强烈光强度。为了测定胞内萦光素酶,有必要在检测之前裂解细胞。因此,独立于luciferase1000assaysystem提供细胞裂解缓冲液。相比之下,one-glotmluciferaseassaysystem将萦光素酶底物与细胞裂解试剂组合,还显示更稳定的信号。通过发光计从整个可见光谱收集作为反应副产物产生的光。在本文所示的实例中,信号与所产生的萦光素酶的量成比例,因此与nfκb启动子的激活强度成比例。在另一实施方案中,使用萦光素酶测定,其中萦光素酶从细胞分泌。因此,可在不裂解细胞的情况下进行测定。可将报道基因的表达与待测试的配体或抗体的功能性直接相关。例如,在用编码萦光蛋白质的基因或编码萦光素酶的基因作为报道基因时,从细胞检测到的光的量与待测试的抗体或配体的靶抗原激活或抑制直接相关。在一个实施方案中,以不同浓度测试抗体或配体,并测定报道基因激活或抑制的半数最大有效浓度(ec50)。ec50指抗体或配体的浓度,在该浓度下,抗体或配体在指定的暴露时间后对报道基因的激活或抑制达到基线和最大值之间的中点。剂量反应曲线的ec50因此代表观察到其对靶抗原的最大激活或抑制作用的50%的抗体或配体浓度。在一个实施方案中,细胞天然表达靶抗原。在一个实施方案中,靶抗原是细胞表面受体。在另一实施方案中,细胞在用编码该靶抗原的基因拷贝转染后表达该靶抗原。在另一实施方案中,用编码该靶抗原的附加基因拷贝转染天然表达该靶抗原的细胞。在一个实施方案中,用与编码人o6-烷基鸟嘌呤-dna-烷基转移酶(agt)的基因拷贝融合的编码靶抗原的基因拷贝转染细胞。在一个实施方案中,该细胞是真核细胞,优选人或灵长类细胞。在一个实施方案中,将结合测量为稀释曲线用于测定上文所述的kd和ec50值。抗体或配体与靶抗原的结合直接或通过细胞信号发放级联引出导致反应元件活性调节的细胞反应。反应元件是可通过转录因子等沉默或激活的dna元件。反应元件为本领域已知,且可购得(例如在报告载体中)。通常,反应元件包含dna重复元件,是位于通过转录因子结合驱动报道基因表达的最小启动子上游的顺式作用增强子元件。下表中提到本文中所使用的反应元件及其转录因子的实例:在一个实施方案中,该靶抗原的反应元件是细胞的细胞核中的核反应元件。在另一实施方案中,该反应元件定位在细胞中的质粒上。在一个实施方案中,该测定包括用包含靶抗原反应元件控制下的编码报道基因的dna序列的表达载体转染细胞的预备步骤。在一个实施方案中,靶抗原是细胞表面受体。在一个实施方案中,在同一小管或孔中测量抗体或配体与靶抗原的结合和功能性。在一个实施方案中,在同一测试介质中测量抗体或配体与靶抗原的结合和功能性。本文所述新测定的优势是无需洗涤步骤。优选地,测试介质是为细胞提供存活至多48小时的条件的介质。适宜的介质是例如实施例中所列的fluorobright或dmem。在一个实施方案中,在微量滴定板中进行测定。在一个实施方案中,微量滴定板适合用于高通量筛选。本发明的测定可以以允许快速配制、处理和分析多个反应的任何型式进行。这可以是例如在多孔测定板(例如96孔或384孔)中。多种试剂的母液可以手动或自动配制,所有后续吸取、稀释、混合、分散、洗涤、孵育、样品读取、数据采集和分析可以用市售分析软件、机器人和能够检测fret、bret或alphascreen信号的检测仪器自动进行。在一个实施方案中,步骤iii)和iv)顺次或同时进行。在一个实施方案中,该fret是时间分辨fret。在一个实施方案中,该fret能量供体化合物是铽穴合物和/或该fret能量受体化合物是d2。在一个实施方案中,该靶抗原和反应元件是nf-κb途径的部分。在一个实施方案中,该靶抗原选自toll样受体、tnf受体、t细胞受体和b细胞受体上的靶抗原。结合其靶标时导致nf-κb活性调节的抗体的非限制性实例是抗cd40抗体、抗dr5抗体、抗dr4抗体、抗41bb抗体、抗ox40抗体和抗gitr抗体。在一个实施方案中,该反应元件是nf-κb反应元件。在一个实施方案中,该反应元件包含一个或多个以下dna重复:gggaatttcc(seqidno:1)、ggggactttcc(seqidno:2)、gggactttcc(seqidno:3)、gggacttcc(seqidno:4)、attgtagcgta(seqidno:5)。在一个实施方案中,该反应元件包含3至6个、3个或6个上文提到的dna重复。在一个实施方案中,该反应元件包含3至6个、3个或6个上文提到的dna重复,及1、2、3或4个附加核苷酸。在一个实施方案中,该反应元件包含以下dna序列:gggaatttccggggactttccccggaatttccggggactttccgggaatttcc(seqidno:6),gggaatttccgggaatttccgggaatttccgggaatttccgggaatttccgggaatttcc(seqidno:7),gggacttccgggactttccgggactttccgggactttccgggactttccgggactttcc(seqidno:8),gggactttccattgtagcgtagggactttccattgtagcgtagggctttccattgtagcgtagggctttcc(seqidno:9),在一个实施方案中,将抗体或配体的结合和/或功能性与参照抗体或配体的结合和/或功能性(都用本文所述的测定测量)相比较。参照抗体或配体是现有技术抗体或配体,其与待测试的抗体或配体结合同一抗原,且具有已知的特性(例如结合和/或功能性)。在一个实施方案中,该测定包括用包含靶抗原反应元件控制下的编码报道基因的dna序列的表达载体转染细胞的预备步骤。在一个实施方案中,在步骤i)中提供每孔1000至40000个细胞。在优选实施方案中,在步骤i)中提供每孔5000至30000个细胞或5000至10000个细胞。在一个实施方案中,在步骤ii)中提供抗体或配体以达到每孔120-0.02nm抗体或配体的浓度。iii.实施例以下是本发明的方法和组合物的实例。应理解,由于上文提供的一般描述,可以实施多种其他实施方案。虽然已为了理解的清晰性的目的,以说明和实例的方式较为详细地描述了前述发明,但该描述和实例不应解释为限制本发明的范围。本文引用的所有专利和科学文献的公开内容明确以其整体引入作为参考。为了组合功能性和结合评估,首先优化了两种测定,然后组合:提供关于抗体功能性的信息的萦光素酶测定和允许研究蛋白质-蛋白质相互作用的测定如fret测定。对于两种测定的组合,需要用与snap标记融合的命名为“受体x”的靶抗原转染并用供体萦光团标记的报告细胞系。转染之前,通过进行萦光素酶测定来针对它们的nfκb激活评价报告细胞系helanfκb-luc和heknfκb-luc-gfp。用tnfα而不是抗受体x抗体来测试nfκb途径的激活,其导致萦光素酶表达。表1:缩略词表表2:材料实施例1:用snap-受体x融合瞬时转染和标记细胞helanfκb-luc细胞瞬时转染的优化helanfκb-luc细胞必须用携带编码receptorx融合的基因转染。为了找出最佳转染方法,评价了每孔接种的细胞数,以及三种不同的转染试剂2000、ltx和x-tremegenehpdna转染试剂)。细胞数的评价在6孔板中接种六个不同的细胞数(每孔300000、400000直至800000个活细胞),以找到用于在24小时孵育时间后达到70%汇合的最佳数目。转染试剂的评价为了评价转染试剂,在6孔板中接种每孔500000个活helanfκb-luc细胞,37℃、5%co2孵育24小时。用1mlpbs洗涤细胞,加入2ml预热的培养基,然后用不同转染试剂处理各孔。通过在无菌1.5mleppendorf反应管中混合150μl低血清培养基和10μl2000dna转染试剂来配制第一转染混合物。在第二个管中,将150μl低血清培养基与3.5μgsnap-受体x质粒混合。室温下至少5分钟孵育时间后,将两个管的内容物混合,并在室温孵育25分钟。对于第二转染混合物,将1.25μg质粒稀释在500μl低血清培养基中。随后,加入5μlltx,轻轻混匀,室温孵育25分钟。通过将2μg质粒稀释在200μl低血清培养基中来配制第三转染混合物。然后,加入6μlx-tremegenehpdna转染试剂,室温孵育30分钟。以逐滴方式将各转染混合物加至细胞。轻轻摇动6孔板以保证在整个孔内均匀分布。然后在调湿培养箱中37℃、5%co2孵育细胞24小时。孵育后,小心去除含转染混合物的培养基,用3mlpbs洗涤转染细胞一次。为了用snap-lumi4-tb标记细胞,向转染细胞加入1.5ml稀释标记试剂(100nm于缓冲液中),在调湿培养箱中孵育1小时。然后,去除标记试剂,通过使用500μl细胞解离缓冲液来使细胞脱附。将脱附的细胞重悬在2.5mlpbs,转移至15mlfalcon管,通过300g、5分钟的离心步骤用5ml磷酸缓冲盐溶液(pbs)洗涤三次。将细胞沉淀重悬在140μl反应缓冲液中。用100μl细胞悬液进行1:5稀释,以通过来自beckmanncoulter的vi-celltmxr测定细胞活率和活细胞浓度。为了评价转染后的蛋白质表达,将剩余40μl细胞悬液分入白色平底384孔板的2个孔中,以在酶标仪(platereader,victor3tm)中在615nm波长测量铽信号。为了在不同转染方法之间比较转染水平,将铽信号对每孔10000个活细胞的细胞数归一化。结果为了找出最佳转染方法,在6孔板中测试了三种不同的转染试剂:lipofectamine2000、lipofectamineltx和xtremegenehp。对于每种试剂,每孔接种500000个活细胞。然后用供体萦光团铽标记snap标签融合蛋白来进行前瞻性实验。此外,在615nm波长测量萦光时,铽标记允许测定转染效率。此外,测定转染和标记后的细胞活率(图1)。图1显示,对于转染hela细胞,lipofectamineltx是最好的试剂。除最好的转染和标记后细胞活率外,还可以达到最高的铽信号。相反,xtremegenehp以及lipofectamine2000杀死更多细胞,且转染水平更低。从6孔板至t150细胞培养瓶的转染规模放大确定最佳转染方法后,必须为进行中的项目转染和标记大量细胞用于所计划的实验。除helanfκb-luc细胞外,还必须转染heknfκb-luc-gfp细胞。因此,在转染前一天,在t150细胞培养瓶中在25mldmem+10%fbs中接种15x106(15mio)个活helanfκb-luc细胞和11x106个活heknfκb-luc-gfp细胞。通过将18.75μgsnap-受体x质粒加至7.5ml低血清培养基来配制用于hela细胞的转染复合物,而对于hek细胞,将37.5μgdna稀释在相同体积的转染培养基中。轻轻混匀稀释液后,加入75μlltxdna转染试剂。洗涤t150细胞培养瓶中的细胞后,逐滴加入转染复合物,并在调湿培养箱中孵育24小时。然后,再次洗涤转染的细胞,并通过在调湿培养箱中37℃孵育1小时来用10ml100nm标记试剂稀释液标记。pbs中洗涤细胞三次,用beckmanncoultervi-celltmxr计数。然后300g离心细胞5分钟,重选在反应缓冲液中,以测定蛋白质表达。因此,将每孔10000个活细胞用于体积20μl的反应缓冲液。在615nm波长一式两份测量铽信号。再次离心细胞悬液,弃上清,沉淀重选在适当体积的冻存培养基(含90%热灭活fbs和10%dmso)中,以达到1x106细胞/ml。用含异丙醇的冻存管冷冻盒将0.5和1ml的分装样品于冻存管中冻存在-80℃。以与上文所述相同的方式,但使用每个t75瓶4x106个细胞、10μgdna稀释在4mlopti低血清培养基中和37.5μlltx,进行具有受体x-snap的稳定受体x表达细胞的瞬时转染。其中这些细胞称为“超转染”。结果对于进行中的项目,需要更大量的细胞。因此,用lipofectamineltx在t150细胞培养瓶中转染heknfκb-luc-gfp和helanfκb-luc细胞,并测定铽信号(图2)。对于两个细胞系,铽信号以及活率显示相似的结果。因此,推测snap-受体x融合蛋白在细胞表面的表达相似,因此,用这些细胞系进行的前瞻性测定的结果可比。实施例2:间接结合测定一般流程配制称为抗体-i的抗人受体x抗体在反应缓冲液中的从100nm至0.2nm的两倍稀释系列。在相同缓冲液中稀释抗人igg-d2检测抗体至每孔150nm的终浓度。每个测定中使用用snap-受体x质粒瞬时转染并用铽标记的heknfκb-luc-gfp细胞。解冻细胞系,通过350g离心5分钟来用10mlpbs洗涤。弃上清,沉淀重悬在反应缓冲液中,以获得1x106细胞/ml。在测定中,细胞不按某个细胞数目使用,而是将它们调整至某个tb信号。为了找出测定中每个孔是用多少个细胞,在384孔板中一式三份地将10μl重悬细胞(每孔10000个细胞)与10μl反应缓冲液混匀,在victor3tm酶标仪中测定615nm铽信号。对于测定,然后调整细胞数,以获得通过victor3tm酶标仪测定的每孔20000至30000个计数之间的铽信号。用10μl稀释细胞、5μl抗人igg-d2和5μl抗体稀释系列进行测定。作为空白,吸取10μl细胞、5μl二抗和5μl缓冲液。所有测量均一式三份地进行。室温孵育0小时、2小时和4小时后用tecanm1000pro酶标仪测量板。所有后续实验(包括不同抗受体xigg和构建体的比较,多种检测抗体浓度、孵育温度的评价,及孵育培养基的影响)均用此流程进行。偏离一般流程的例外或变化在结果章节中陈述。结果转染和标记后,用靶向受体x的两种不同igg(抗体-i和-ii)在间接结合测定中测试了两种细胞系(图3)。图3显示,抗体-i与两种细胞系(表达受体x的heknfκb-luc-gfp和helanfκb-luc细胞)的结合都比抗体-ii好。抗体-i对两种细胞系的结合信号具有相同的强度。对于抗体-ii,对helanfκb-luc-受体x-tb细胞仅可检测到弱信号,在heknfκb-luc-gfp-受体x-tb细胞上无可见信号。因此,将抗体-i用于进行中的实验。用作二抗的d2标记抗人igg与一抗相比应至少三倍摩尔过量。测试了每孔75nm终浓度并比较了结果(图4),而不是如之前测定中使用150nm的浓度。图4显示,150或75nm浓度二抗之间无显著差异。对于150nm的结合曲线,测定kd值为0.19nm±0.05nm(r2=0.93),而对于75nm浓度,发现kd为0.16nm±0.03nm(r2=0.95)。因此,如果使用至多20nm浓度的一抗,则在所有进行中的实验中使用75nm的二抗。除使用igg抗体-i外,还针对其结合测试了两种不同的受体x抗体构建体(即抗体-iii和-iv)(图5)。所有抗体和抗体样构建体都显示相似的结合和kd值,抗体-i为0.11nm±0.03nm(r2=0.85),抗体-iii为0.18nm±0.02nm(r2=0.97),抗体-iv为0.28nm±0.04nm(r2=0.98)。对于萦光素酶测定与结合测定的组合,可能有必要在37℃而不是常用于结合测定的室温(rt)下孵育细胞。因此,必须评估37℃下的结合(图6)。rt孵育测定的kd值为0.18nm±0.02nm(r2=0.97),仅略低于37℃孵育(kd为0.43nm±0.18nm(r2=0.85))。但是,二者的结合都在低纳摩尔范围,因此相对的强。除用缓冲液或pbs进行结合测定外,还测试了可能更适于萦光素酶测定的其他介质(图7)。图7显示,pbs和dmem减少抗体构建体与受体的结合。dmem/10%fbs或fluorobritedmem/10%fbs与一般使用的缓冲液相比显示相似的结果,因此对组合测定而言是适宜的替代。解冻后的瞬时转染细胞上的受体x的时间依赖稳定性用snap-受体x质粒瞬时转染heknfκb-luc-gfp细胞后,冷冻细胞并用于日常实验。为了确定对解冻的瞬时转染细胞进行培养后受体的稳定性,每天进行间接结合测定,进行三天。因此,将解冻的细胞分入6孔板的4个孔中,培养在4mlfluorobritetmdmem+10%fbs中。每天,脱附6孔板的一个孔的细胞,350g离心5分钟,重悬在fluorobritetmdmem+10%fbs中至终浓度为1x106细胞/ml。然后将每孔10000个细胞与抗体-iii在fluorobritetmdmem+10%fbs培养基中的从80nm至0.16nm的两倍稀释系列混合来测试结合。将抗人igg-d2检测抗体在相同培养基中稀释至每孔75nm的终浓度。每日监测铽信号。结果监测受体在瞬时转染细胞的细胞表面上的存在随时间的变化。直接在解冻细胞后(第0天)以及在随后3天,通过使用每孔10000个解冻细胞或培养细胞来进行间接结合测定(图8)以及细胞铽信号测量(图9)。图8显示,结合信号从第0天至第1天显著减少,在第2和3天彻底没有信号。时间点第0天的665nm与620nm的比值*10000约为10000,而1天后,bmax约为3000,其代表原始信号的70%。时间点第0天和第1天的kd值都在纳摩尔范围内,分别为0.49nm±0.08nm(r2=0.98)和0.12nm±0.03nm(r2=0.87)。图9与图8中所示的结果一致,显示铽信号随时间推移减少。相反,活率与日递增。深度评价测定参数后,萦光素酶测定的条件也必须优化。htrf原始数据的评价对于测定的分析,首先通过microsoftexcel编辑原始数据。在htrf测定中,结果的变异可由于细胞、培养基添加剂的吸取步骤而发生在孔与孔之间,及发生自每孔裂解的细胞数。为了最小化这些变异,通过计算665nm与620nm比值将受体的发射对各孔中供体信号的发射归一化:―比值=”―665nm”/―620nm”―*10000”。通过称为graphpadprism6.0的软件评价计算的比值。用非线性回归拟合结合曲线。用以下方程用“单部位特异性结合”测定bmax和kd:―y=―(―b”_―max”―*x”)/(―(――k”_―d””+x)”)。实施例3:萦光素酶测定两种不同细胞系的萦光素酶激活的评价针对其在用tnfα刺激时的萦光素酶活性,比较了用于瞬时转染的helanfκb-luc细胞以及已经稳定转染的nfκb-luc-受体x细胞。所用方法为luciferase1000assaysystem(promega)。因此,激活前一天,将两种细胞系按每孔20000个细胞的细胞数接种在白色96孔板中的100μl生长培养基(dmem+10%fbs+1%glutamax-i+200μg/ml潮霉素b)中。此外,还将相同的细胞接种在透明96孔板的3各孔中来在激活前通过显微镜测定汇合、附着和污染状态。37℃、5%co2孵育24小时后,配制tnfα的从25ng/ml至0.8ng/ml的两倍稀释系列。将100μl各稀释液一式三份加至细胞,然后进行37℃、5%co2孵育6小时的步骤。然后,通过350g、5分钟的离心步骤用每孔200μlpbs洗涤细胞三次。为了检测所产生的萦光素酶,必须裂解细胞。因此,向各孔加入40μl裂解缓冲液并在-80℃孵育2小时以确保裂解。调节细胞至室温后,避光向各孔加入100μlluciferase1000测定试剂,立即以500ms积分时间通过spectramaxm5/m5e酶标仪测量整个可见光谱所有波长的光发射。作为空白,扣除含萦光素酶试剂的裂解细胞的信号。通过发光计从整个可见光谱采集作为反应副产物产生的光。还用相同的流程进行了helanfκb-luc细胞与heknfκb-luc-gfp细胞系的比较。不同激活方法比较一式三份地进行与上文所述相同的测定,只是将瞬时转染的heknfκb-luc-gfp-snap-受体x细胞按每孔80000个细胞接种在dmem+10%fbs中。除通过tnfα受体激活外,用抗体-i来结合在寡聚化时也应能够激活nfκb途径的受体x。因此,配制抗体-i的从120至0.03nm的四倍稀释系列,将50μl各浓度加至细胞,然后进行37℃、5%co2孵育15分钟的步骤。对于激活nfκb途径所需的受体的超交联,使用50μl二抗(抗人iggfcγ特异性山羊iggf(ab)2)。此二抗的浓度保持480nm恒定,其转化为与一抗相比至少四倍摩尔过量。luciferase1000和one-glotmluciferaseassaysystem的比较除luciferase1000测定系统外,还测试了one-glotmluciferase测定系统,并对结果进行比较。因此,与作为阳性对照的稳定转染heknfκb-luc-gfp-受体x细胞相比较,测试了瞬时转染的heknfκb-luc-gfp-snap-受体x细胞。此外,分析了tnfα激活及受体x激活。激活前一天,将两种细胞系按每孔30000个细胞的细胞数接种在两个白色96孔板(每种测定系统一块板)中的150μldmem+10%fbs中。此外,还将相同的细胞接种在透明96孔板的3个孔中,以在激活前通过显微镜确定汇合、附着和污染状态。37℃、5%co2孵育24小时后,向测试受体x激活的孔中加入10nm一抗抗体-iii。再孵育15分钟后,加入40nm二抗或50ng/mltnfα。37℃、5%co2孵育平板48小时。一块板按上文针对luciferase1000测定系统所述处理。简言之,洗涤、裂解、冷冻、解冻细胞,并在加入萦光素酶试剂后测量。作为空白,扣除未激活的裂解细胞的信号。另一块板用于one-glotmluciferase测定系统。因此,每孔去除适当体积的上清,使终体积为每孔100μl。每孔加入100μlone-glotmluciferase试剂。避光孵育5分钟后,以500ms积分时间通过spectramaxm5/m5e酶标仪测量光发射。作为空白,扣除含萦光素酶试剂的裂解的非激活细胞的信号。一式三份进行测量。结果首先在萦光素酶测定中将helanfκb-luc细胞与helanfκbluc-受体x细胞相比较。已知后者通过tnfα受体激活nfκb途径的能力,因此用它们作为阳性对照(数据未显示)。helanfκb-luc细胞达到了和阳性对照几乎相同的发光信号。因此,tnfα可以很好地激活nfκb途径,意味着该细胞可以用于用含有编码snap-受体x的基因的质粒转染。测试heknfκb-luc-gfp细胞,并与helanfκb-luc细胞相比较(数据未显示错误!未找到引用源。)。由于其可作为用于检测基因表达的附加工具的第二报道基因,此细胞系将优选用于开发组合测定。heknfκb-luc-gfp细胞显示比helanfκb-luc细胞高的信号。由于两种细胞系都可以用于萦光素酶测定,用snap-受体x质粒转染它们中的每一种。用受体x-snap转染和标记heknfκb-luc-gfp细胞后,用抗体-i进行初步萦光素酶测定,并与tnfα激活比较。单独使用作为一抗的抗体-i以及二抗作为阴性对照(图10)。图10显示tnfα对nfκb途径的强烈激活。抗体-i和二抗单独不能激活该途径。两种抗体组合仅可检测到非常弱的发光信号。总之,确定抗体-i是最佳结合剂,但在此背景中未显示良好的功能性。此外,评价了两种不同的萦光素酶测定系统one-glotm和luciferase1000系统(图11)。因此,用tnfα激活稳定和瞬时转染的表达受体x的heknfκb-luc-gfp细胞的nfκb途径。使用48小时来给细胞更多时间表达萦光素酶,而不是激活后孵育6小时。图11显示one-glotmluciferase测定系统对萦光素酶催化的从萦光素至氧化萦光素的反应所释放的光更敏感。由于更高的发光信号以及更快和更容易进行测定流程,将one-glotmluciferase测定系统用于进行中的实验。在以下实验中,通过使用与前一实验中相同的细胞比较了6小时和48小时的孵育时间。作为一抗的抗体-iii按10nm浓度使用,二抗(抗人iggfcγ特异性山羊iggf(ab)2)按40nm浓度使用(图12)。稳定转染的heknfκb-luc-gfp-受体x细胞清楚地显示,孵育48小时后,细胞产生了更多的萦光素酶,产生更强的生物发光信号。抗体-iii单独如预期地以较低程度激活该途径,因为它已经可以三聚化受体。瞬时转染细胞的信号显著更低。尤其是对于6小时孵育,两种抗体组合和各抗体自身之间生物发光没有大的差异,而48小时的值略有差异。从96孔板至384孔板的“规模缩小”对于功能性和结合的组合测定,有必要将通常在96孔板中进行的萦光素酶测定转移至384孔板。one-glotmluciferase测定用96孔板进行,但不同于使用每孔30000个细胞,仅使用5000个细胞。一抗和二抗分别保持在10nm和40nm的恒定浓度,而tnfα按50ng/ml的浓度使用。37℃、5%co2孵育24小时后,去除适当体积的上清,使各孔中剩余15μl体积。每孔加入15μlone-glotmluciferase试剂。用稳定heknfκb-luc-gfp-受体x细胞一式三份进行测量。此外,用与上文所述相同的流程评价了两种不同的细胞数(每孔5000和10000个细胞)。结果考虑将组合功能性和结合评估用于高通量筛选(hts)。由于此原因及由于成本和使萦光素酶测定与实验一致的原因,测试了从96孔板至384孔板的规模缩小(图13)。整体上,图13显示96孔板的信号比384孔板略强。然而,即使384孔板中的信号低于96孔板,关于激活或功能性的结果保持不变。因此,在384孔板中进行了进行中的萦光素酶测定。对于萦光素酶测定的规模缩小,计算每孔5000个活细胞的细胞数,但对于结合实验,有时需要每孔10000个细胞,以具有足够高的供体萦光团萦光信号用于fret信号。使用稳定转染的heknfκb-luc-gfp-受体x细胞,用one-glotmluciferase测定系统测试了使用5000或10000个细胞的可行性(图14)。图14显示,在使用每孔10000个活细胞时,生物发光信号甚至更高。尽管保持其他参数(如所加入的抗体量)不变,细胞可产生更多的萦光素酶,导致生物发光信号更强。现在,比对了用于结合和萦光素酶测定的组合的所有参数。不幸的是,没有一个细胞系适合用于组合测定。有稳定转染的heknfκb-luc-gfp-受体x细胞可用,其在功能性测定中显示良好的结果,但由于缺乏供体萦光团而不能用于正常测定,有瞬时转染的heknfκb-luc-gfp-受体x-tb细胞可用,其在结合测定中显示良好的结果,但不能非常好地激活。因此,测试了两种方法。一方面是稳定转染细胞上的基于wga的fret,另一方面是用受体x-snap构建体超转染稳定转染的细胞。一抗浓度优化所有萦光素酶测定都用10nm一抗和40nm二抗进行。此测定的目的是滴定五个稀释度的一抗并保持二抗为1:4的比例进行检测。用瞬时转染的heknfκb-luc-gfpsnap-受体x细胞、稳定转染的heknfκb-luc-gfp-受体x细胞以及瞬时超转染细胞进行one-glotmluciferase测定。对于全部三个细胞系,将每孔5000个细胞接种在384孔板中的15μldmem+10%fbs中,并在激活前37℃、5%co2孵育至少12小时。配制一抗抗体-iii的两倍稀释系列,按每孔40nm至2.5nm加至孔中。孵育15分钟后,按每孔160nm至10nm的浓度将二抗(抗人iggfcγ特异性山羊iggf(ab)2)加至孔中,保持二抗与一抗相比处于四倍摩尔过量。37℃、5%co2孵育平板。孵育24小时后,去除适当体积的上清,使每孔剩余15μl终体积。每孔加入15μlone-glotmluciferase试剂。避光孵育5分钟后,按500ms积分时间通过spectramaxm5/m5e酶标仪测量光发射。作为空白,扣除含萦光素酶试剂的裂解的非激活细胞的信号。一式三份进行测量。交联抗体的比值优化对于功能性测定,用二抗(抗人iggfcγ特异性山羊iggf(ab)2)进行交联。为了将萦光素酶测定与结合测定组合,用受体萦光团d2标记的抗体是必要的。或者必须标记抗人iggfcγ特异性山羊iggf(ab)2,或者可以使用来自cisbio的抗人igg-d2。一抗保持在之前的测定中优化的40nm恒定浓度,而一抗与二抗的比值从1:1变化至1:5。实施例4:间接wga-htrf将抗人igg-d2检测抗体在反应缓冲液中稀释至每孔75nm终浓度。配制抗体-iii在反应缓冲液中的从1.56nm至0.01nm的两倍稀释系列。解冻稳定heknfκb-luc-gfp受体x细胞,通过350g离心5分钟用10mlpbs洗涤。弃上清,将沉淀重悬在反应缓冲液中至1x106细胞/ml。向细胞悬液中加入0.05ng/μlwga-铽,并在室温孵育30分钟。然后,再次洗涤细胞,重悬在适当体积的反应缓冲液中至终浓度为1x106细胞/ml。在384孔板中将10μl重选的wga-铽标记细胞与10μl反应缓冲液混匀。用victor3tm酶标仪在615nm波长一式三份测定铽信号。用10μl具有某种铽信号的细胞、5μl抗人igg-d2和用于滴定的5μl抗体稀释液进行测定。作为空白,吸取10μl细胞、5μl二抗和5μl缓冲液。在384孔板中一式三份进行测定,室温孵育0小时、2小时和4小时后用tecanm1000pro酶标仪测量。按上文所述进行原始数据的评价。结果用铽缀合的凝集素wga孵育稳定转染细胞,通过用抗人igg-d2作为受体标记二抗来建立间接htrf测定(图15)。在图15中,与阴性对照相比,检测到了抗体-iii的结合信号。测量值同样地符合实验,使得可以测定和比较kd值。此实验中的kd值为0.15nm±0.03nm(r2=0.97)。实施例5:组合测定对于组合测定,配制抗体-iii在fluorobritedmem+10%fbs中的从200nm至0.1nm的两倍稀释系列。这导致每孔最终浓度从40nm到0.02nm变化。抗人igg-d2检测抗体稀释在相同的缓冲液中至每孔120nm终浓度。使用snap-受体x质粒瞬时超转染和铽标记的heknfκb-luc-gfp细胞。解冻细胞系,通过350g离心5分钟用10mlpbs洗涤。弃上清,将沉淀重悬在fluorobritedmem+10%fbs中。用每孔15μl培养基中的10000个细胞、5μl抗人igg-d2和5μl抗体稀释系列在384孔板中进行测定。作为空白,吸取15μl细胞、5μl二抗和5μl缓冲液。所有测量都一式三份进行。37℃、5%co2孵育1小时、2小时和4小时后用tecanm1000pro酶标仪测量平板。孵育24小时后,去除10μl上清,使每孔剩余15μl终体积。每孔加入15μlone-glotmluciferase试剂。避光孵育5分钟后,按500ms积分时间通过spectramaxm5/m5e酶标仪测量光发射。作为空白,扣除含萦光素酶试剂的裂解的非激活细胞的信号。结果适合用于组合功能性和结合评估的细胞系的评价新方法是“瞬时超转染”稳定转染的heknfκb-luc-gfp-受体x细胞。这是指用编码snap-受体x的质粒再次转染heknfκb-luc-gfp-受体x细胞,使受体x与融合。然后,用铽标记细胞,该细胞可用于正常的间接结合实验(图16)。用瞬时转染细胞作为对照,因为他们在初步实验中显示与抗体-iii的良好结合。图16显示,与瞬时转染细胞相比,瞬时超转染细胞的比值更低。但是,信号高至足以获得漂亮的曲线拟合和测定kd值。两种细胞类型的kd相同,heknfκb-luc-gfp受体x-tb超转染和转染细胞分别为0.09nm±0.02nm(r2=0.92)和0.09nm±0.02nm(r2=0.96)。因此,那些超转染细胞具有在组合功能性和结合评估的测定中用作模型的潜能。在将测定与萦光素酶测定组合之前最后的实验中,在萦光素酶测定中找到一抗和二抗的最佳浓度。首先,以2.5至40nm之间的五个浓度滴定作为一抗的抗体-iii,而一抗与二抗的比值保持1:4恒定比值。除稳定和瞬时转染的表达受体x的heknfκb-luc-gfp细胞外,还测试了瞬时超转染细胞(图17)。与另外两个细胞系相比,瞬时超转染细胞显示非常高的生物发光信号。此外,在所有细胞系中,可以清楚地看到,浓度40nm的一抗显示最佳结果。因此,在后续测定中保持抗体-iii的此浓度恒定,而与二抗的比值从1:1比值变化至1:5比值。此外,将通常在萦光素酶测定中用于交联的二抗(抗人iggfcγ特异性山羊iggf(ab)2)与常用于实验的d2标记二抗(抗人igg-d2)相比较(图18)。图18显示,在用d2标记抗人igg交联时生物发光信号更强。因此,120nm浓度(1:3比值)二抗已经足够。对于抗人iggfcγ特异性山羊iggf(ab)2,200nm浓度(1:5比值)显示最好的结果。最终条件的组合测定评价和确定了将结合测定与功能性测定组合的所有必要参数。用瞬时超转染的表达受体x的heknfκb-luc-gfp细胞进行组合测定。因此,在384孔板中将每孔10000个细胞与抗体-iii和二抗混匀。在4小时时间内进行间接结合测定(图19)。激活受体x后24小时,用one-glotm萦光素酶试剂裂解细胞。由于此试剂还包含萦光素酶底物,通过发光计测量从萦光素氧化至氧化萦光素所释放的光(图20)。对于heknfκb-luc-gfp-受体x-tb超转染细胞,图28中测定的kd为0.10nm±0.01nm(r2=0.98)。图20显示,随着一抗浓度逐渐提高至10nm,发光信号也提高。因此,一抗和二抗的组合导致nfκb途径激活。总之,抗体-iii能够结合受体x,它还显示良好的功能性。虽然显示和描述了目前优选的本发明的实施方案,但应清楚地理解,本发明不限于该实施方案,而是可以在以下权利要求的范围内包含和实施多种其他实施方案。讨论细胞系的评价转染snap-受体x质粒之前,在萦光素酶测定中测试了heknfκb-luc-gfp和helanfκb-luc。heknfκb-luc-gfp细胞的信号显著高于用helanfκb-luc细胞细胞检测到的信号,但这两种细胞系不应进行比较,因为它们的生长速率完全不同。激活前一天接种相同量的细胞。由于heknfκb-luc-gfp细胞分裂得更快,次日可以激活更多的细胞。对于进一步的实验,helanfκb-luc细胞必须以高于heknfκb-luc-gfp细胞的细胞数接种。但是,由于两种细胞系都可以很好地激活,用它们进行与融合的靶受体的转染。蛋白质-蛋白质相互作用研究在间接结合测定中,将抗体-i与抗体-ii相比较。这些抗体之间的差异是在抗体-i轻链的互补决定区(cdr)中交换了一个氨基酸。这肯定是抗体-i不像抗体-ii一样结合其靶标的原因。除使用反应缓冲液外,还可以使用fluorobritedmem/10%fbs和dmem/10%fbs。相反,pbs或dmem不支持抗体与靶受体的结合。dmem和dmem/10%fbs的比较是一个好的实例,fbs增加结合。fbs作为培养基添加剂为细胞提供营养物,且包含生长因子。使用包含fbs的培养基时结合的增加可能仅仅是由于更低的细胞死亡或更好的细胞生长。pbs减少结合的解释可能也是如此,在pbs中孵育细胞4小时时,它们可能开始结合。反应缓冲液的组成未知,但它似乎支持两种蛋白质的结合,且保持细胞是活的。在heknfκb-luc-gfp细胞上瞬时转染snap-受体x质粒后,通过在解冻细胞后每天进行测定来测试受体稳定性。观察到从第0天(解冻后直接进行)至第1天的强烈减少。但这并非必然意味着细胞表面受体丢失。细胞刚进行了瞬时转染,因此,在增殖后,子细胞不具有表达受体x的基因。由于通过每天使用相同量的细胞进行测定,在第1天用于测定的许多细胞可以是不具有此受体的那些子细胞。第2和3天表达受体x的细胞完全生长超过非表达受体x的子细胞。此推测也符合每日测量的铽信号和活率。在第0天,测定到高铽信号,但活率低。因此,在解冻后即刻,细胞没有那么健康,但大多数细胞表达受体x。日复一日,细胞增殖产生更健康的子细胞,但在其细胞表面不具有标记的靶受体。另一种推测是受体的内化和降解。对于结合和功能性的组合测定,此结果没有任何担忧,因为细胞在解冻后直接激活。然后,在4小时的时期内测量结合,在几个小时后测量功能性,受体在细胞表面的存在不发挥任何作用。激活细胞和测量功能性之后的孵育时期对获得高萦光素酶表达很必要。萦光素酶测定用在测定中显示良好结合的抗体-i进行瞬时转染和标记后的初步萦光素酶测定。此外,还尝试通过tnfα激活nfκb途径。tnfα激活表现得非常好。这证明细胞的转染和标记未改变细胞,nfκb途径仍在工作。还有细胞用于测定而不进行激活,作为从信号扣除的空白。这确认细胞不组成性表达任何萦光素酶。在用抗体-i和二抗激活细胞时仅观察到弱发光信号。这是显示进行组合测定的必要性的好的实例,因为抗体-i测定为最佳结合剂之一。在另一萦光素酶测定中,用稳定和瞬时转染的表达受体x的heknfκb-luc-gfp细胞比较了两种不同孵育时间(6小时和48小时)。可以清楚地看到,48小时后稳定转染细胞中表达更多的萦光素酶,其产生更强的发光信号。不幸的是,瞬时转染细胞仅显示非常弱的发光信号。瞬时转染细胞表达仅与融合的受体x。由于nfκb途径的激活仅在受体x寡聚化时发生,的大小(20kda)可能导致空间位阻和寡聚化抑制。间接wga-htrfwga通过避免直接标记表达在细胞表面的受体而使得能够以新方式进行基于细胞的蛋白质-蛋白质相互作用测量。避免和允许用稳定转染细胞进行组合测定的方法是通过使用wga来用铽标记它们。因此,可以进行间接htrf测定。kd值为0.15nm±0.03nm,与在正常实验中在瞬时转染的表达受体x的heknfκb-luc-gfp细胞上测定的值处于相同范围。因此,wga作为标记工具也可以用此设置来进行组合测定,但由于时间限制未进行进一步评价。适合用于组合功能性和结合评估的细胞系的评价第二种方法是瞬时超转染稳定转染的heknfκb-luc-gfp-受体x细胞,其使得能够进行正常的间接结合实验。除在结合测定中测试它们外,还用此细胞系评价了萦光素酶测定。因此,用这些细胞来找出最佳一抗浓度,同时将它们与瞬时和稳定转染的表达受体x的heknfκb-luc-gfp细胞相比较。对于每个抗体-iii浓度,超转染细胞清楚地显示最强的发光信号。超转染细胞与稳定细胞相比显示更高信号的一个原因是细胞表面存在更多的靶受体。因此,更多抗体可以结合,更多的交联可以发生,导致nfκb途径的激活更强。稳定转染细胞的信号在此测定中一般极低。原因可能是传代细胞的错误处理。如果将它们分开得非常晚,细胞具有高汇合,可观察到细胞不能那么好地激活。或者一些细胞丢失其细胞表面的受体,或者它们以某种方式下调信号发放事件。还推测,由于空间位阻,瞬时超转染细胞的信号高于瞬时转染细胞。超转染细胞在其细胞表面过量表达受体x。它们中的一些与融合,因为用snap-受体x质粒对它们进行了瞬时转染,但它们中的一些不具有因为用稳定转染的表达受体x的heknfκb-luc-gfp细胞系进行snap-受体x的瞬时转染。很可能,这些细胞的激活由于在携带的受体与不含的受体发生寡聚化时的空间位阻更小的原因而更好。此外,在瞬时超转染的表达受体x的heknfκb-luc-gfp细胞上比较了两种二抗。与常用于实验的d2标记二抗(抗人igg-d2)相比,常用于萦光素酶测定的二抗(抗人iggfcγ-特异性山羊iggf(ab)2片段)显示更低的信号。可能的原因是抗人igg-d2是多克隆抗体。多克隆抗体理想地适合作为二抗,因为那些抗体不限于其靶标上的一个表位,因此可以同时交联多种构建体。原先使用的二抗是仅结合一个特异性表位的单克隆(fab)2片段,其对达到良好的交联并不理想。最后,用瞬时超转染的表达受体x的heknfκb-luc-gfp细胞作为用于组合测定的模型。结论可能比对萦光素酶和测定这两种测定的参数来开发二者组合在一个孔中的新测定。例如,测定可以在37℃进行而不是在室温孵育。此外,可以使用培养基而不是反应缓冲液,以及每孔接种的细胞量可以在很大程度上变动而不改变结果。此外,one-glotmluciferase测定系统的规模缩小表现得很好。因此,组合测定可以在384孔板中进行,其过去也用于实验。此规模的优势不仅是需要更低量的试剂,导致总体成本的下降,还有其对进行hts的适合性。最终的组合测定用瞬时超转染细胞进行,因为没有除靶受体外还包含snap标签的稳定细胞系可用。对于以后使用组合测定的实验,也可能产生具有该标签的细胞系,或超转染它们。此外,通过进行wga-htrf,可以使用无的细胞。总之,组合测定的开发不仅可用于nfκb信号发放途径,也有将该测定扩展至其他功能性读出(例如细胞因子释放或其他信号发放途径)的潜能。实施例6:alphascreen对于不同测定的评价,使用了hudr5特异性抗体drozitumab。hudr5通过hekebna细胞表达。对于基于alphascreen的方法,将生物素化wga结合于链霉抗生物素蛋白供体小球(perkinelmer),用结合抗体fc部分的prota受体小球(perkinelmer)检测drozitumab与dr5的结合。值得注意的是,由于小球对光敏感,所有吸取步骤和测量均避光进行。确定alphascreen的测定窗口为了确定测定窗口,用1.4nmwga-生物素和10000个细胞进行alphascreen测定。perkinelmer建议每种小球(受体和供体)的浓度为10μg/ml。首先,将10μg/ml链霉抗生物素蛋白供体小球与1.4nmwga-生物素混匀,并孵育30分钟。同时,按1:4稀释步骤配制drozitumab的从1200nm浓度开始至0.0011nm的稀释系列。解冻表达hudr5snaptag的hekebna细胞,用10ml1xpbs洗涤,350g离心8分钟。弃上清。将沉淀重悬在适当体积中以获得1x106细胞/ml。将wga-生物素标记的链霉抗生物素蛋白小球加至重悬的细胞,室温孵育30分钟。孵育细胞的同时,将5μl稀释抗体一式两份转移至384孔板,加入5μl蛋白a受体小球(10μg/ml)。孵育384孔板30分钟。用1x反应缓冲液将细胞补足至10ml,350g离心8分钟。弃上清,用1x反应缓冲液重悬沉淀在适当体积中以获得1.0x106细胞/ml。将10μlwga链霉抗生物素蛋白小球标记的细胞转移至384孔板的各孔中。用10μlwga-链霉抗生物素蛋白标记的细胞、5μl蛋白a受体小球和5μl1x反应缓冲液配制空白。所有测量均一式两份进行。1小时、2小时、3小时和4小时后用tecanm1000pro酶标仪用alphascreen模板测量384孔板。在excel中通过扣除空白评价原始数据,并进一步在graphpadprism中分析。每孔细胞数和wga-生物素的评价首先,将10μg/ml链霉抗生物素蛋白供体小球与1.4nm或2.8nmwga-生物素混匀,并孵育30分钟。同时,按1:2稀释步骤配制drozitumab的从6.25nm浓度开始至0.020nm的稀释系列。解冻表达hudr5snaptag的hekebna细胞,分入两个15mlfalcon管,用10ml1xpbs洗涤,350g离心8分钟。弃上清。将两个管的沉淀重悬在适当体积中以获得1x106细胞/ml或2.0x106细胞/ml。将wga-生物素标记的链霉抗生物素蛋白小球转移至重悬的细胞,室温孵育30分钟。孵育细胞的同时,将5μl稀释抗体一式两份转移至384孔板,加入5μl蛋白a受体小球(10μg/ml)。孵育384孔板30分钟。用1x反应缓冲液将细胞补足至10ml,350g离心8分钟。弃上清,用1x反应缓冲液重悬沉淀在适当体积中以获得1.0x106细胞/ml和2.0x106细胞/ml。将10μlwga链霉抗生物素蛋白小球标记的细胞转移至384孔板。用10μlwga-链霉抗生物素蛋白标记的细胞、5μl蛋白a受体小球和5μl1x反应缓冲液配制空白。所有测量均一式两份进行。1小时、2小时、3小时和4小时后用tecanm1000pro酶标仪用alphascreen模板测量384孔板。在excel中通过扣除空白评价原始数据,并进一步在graphpadprism中分析。不同受体/供体对的评价为了确定哪个受体/供体对表现最好,用2.8nmwga-生物素和20000个细胞进行alphascreen测定。首先,将10μg/ml链霉抗生物素蛋白供体小球或10μg/ml链霉抗生物素蛋白受体小球与2.8nmwga-生物素混匀,并孵育30分钟。同时,按1:2稀释度配制drozitumab和aby的从25nm浓度开始至0.024nm的稀释系列。解冻表达hudr5snaptag的hekebna细胞,分入两个15mlfalcon管,每个管用于各链霉抗生物素蛋白小球(供体或受体)。用10ml1xpbs洗涤细胞,350g离心8分钟。弃上清。将两个管的沉淀重悬在适当体积中以获得2.0x106细胞/ml。将wga-生物素标记的链霉抗生物素蛋白小球转移至重悬的细胞,室温孵育30分钟。孵育细胞的同时,将5μl稀释抗体一式两份转移至384孔板,加入5μl蛋白a供体小球(10μg/ml)或蛋白a受体小球(10μg/ml)。孵育384孔板30分钟。用1x反应缓冲液将细胞补足至10ml,350g离心8分钟。弃上清,用1x反应缓冲液重悬两个管的沉淀在适当体积中以获得2.0x106细胞/ml。将10μlwga链霉抗生物素蛋白小球标记的细胞转移至384孔板。用10μlwga-链霉抗生物素蛋白标记的细胞、5μl蛋白a小球和5μl1x反应缓冲液配制空白。所有测量均一式两份进行。1小时、2小时、3小时和4小时后用tecanm1000pro酶标仪用alphascreen模板测量384孔板。在excel中通过扣除空白评价原始数据,并进一步在graphpadprism中分析。最终条件的alphascreen首先,将10μg/ml链霉抗生物素蛋白受体小球与2.8nmwga-生物素混匀,并孵育30分钟。同时,配制drozitumab的从2.5nm浓度开始至0.020nm的稀释系列。稀释按1:2稀释步骤进行。解冻表达hudr5snaptag的hekebna细胞,用10ml1xpbs洗涤,350g离心8分钟。弃上清。将沉淀重悬在适当体积中以获得2.0x106细胞/ml。将wga-生物素标记的链霉抗生物素蛋白小球转移至重悬的细胞,室温孵育30分钟。孵育细胞的同时,将5μl稀释抗体一式两份转移至384孔板,加入5μl蛋白a供体小球(10μg/ml)。孵育384孔板30分钟。用1x反应缓冲液将细胞补足至10ml,350g离心8分钟。弃上清,用1x反应缓冲液重悬沉淀在适当体积中以获得2.0x106细胞/ml。将10μlwga链霉抗生物素蛋白小球标记的细胞转移至384孔板。用10μlwga-链霉抗生物素蛋白标记的细胞、5μl蛋白a供体小球和5μl1x反应缓冲液配制空白。所有测量均一式三份进行。1小时、2小时、3小时和4小时后用tecanm1000pro酶标仪用alphascreen模板测量384孔板。在excel中通过扣除空白评价原始数据。通过非线性回归计算kd值。alphascreen原始数据的评价将数据对背景归一化。通过graphpadprism6.0软件通过非线性回归评价归一化的数据来确定结合曲线和kd值。用s型剂量-反应(可变斜率)按照以下方程拟合结合曲线:用单部位结合模型按照以下方程确定kd:结果确定alphascreen的测定窗口在第一测定中,每孔中表达hudr5的hekebna细胞数、每孔中生物素化wga的量和wga-生物素与细胞的孵育时间采纳自wga-htrf。每孔使用10000个细胞和1.4nmwga-生物素。两种抗体都按1:4稀释度按在孔中的终浓度从300nm开始至3.0*104nm稀释。alphascreen测定的第一次测试显示,测定设置的想法应可行。但是,drozitumab的曲线显示从浓度0.018nm至19nm的相对小的测定窗口。每孔细胞数和wga-生物素的评价为了改善测定,在下一步中,将细胞数设为每孔10000和20000个细胞,wga-生物素的量设为每孔1.4nm或2.8nm。为了得到更好的测定窗口,稀释步骤降低至从孔中终浓度50nm开始1:2而不是1:5稀释。两种抗体都按1:2稀释度按孔中终浓度从50nm开始至0.049nm稀释。使用每孔1.4nmwga-生物素的两个测定设置仅显示微弱结合信号或无结合信号。每孔10000个细胞和每孔2.8nmwga-生物素的测定显示弱结合曲线。用每孔20000个细胞和2.8nmwga-生物素获得了最佳结果。不同受体/供体对的评价为了进行alphascreen测定,可能使用链霉抗生物素蛋白供体小球和prota受体小球或链霉抗生物素蛋白受体小球和prota供体小球。为了找出最佳供体/受体对,将它们相互进行比较。两种抗体都按1:2稀释度按孔中终浓度从1.6nm开始至0.006nm稀释。两个不同的供体/受体对显示几乎相同的结合曲线。甚至其r2值的差异也几乎相同。用链霉抗生物素蛋白供体小球和prota受体小球测量的测定具有0.99的r2值,用链霉抗生物素蛋白受体小球与prota受体小球配对测量的测定具有0.99的r2值。最终条件的alphascreen评价alphascreen的结果是按以下设置(表2)进行alphascreen。表2:最终评价的进行alphascreen测定的参数用针对alphascreen测定评价的最终条件,测定了drozitumab与hudr5结合的kd值(图21)。drozitumab的s型拟合显示s型结合曲线。通过非线性回归测定的kd为0.084nm+/-0.0053nm。由于aby显示与hudr5无结合,未计算kd值。与wga-htrf测定中一样,测定评价的结果必须用另一种抗体和抗原确认,测定了abz与其靶受体z的kd(图22)。如之前的测定中,阴性对照抗体aby显示无结合。通过非线性回归测定kd值。abz-受体z结合的kd为0.26nm+/-0.068nm,r2值为0.96。由于aby是受体z的非结合剂,未计算kd值。实施例7:用jurkatnfat报告细胞系在一个孔中测量cd3结合和功能性jurkatnfat萦光素酶报告细胞系设计用于监测导致活化t细胞核因子(nuclearfactorofactivatortcells,nfat)活性调节的t细胞激活。转录因子nfat在t细胞发育和功能中发挥重要作用。在非活化t细胞中,nfat主要定位在细胞质中,处于高度磷酸化形式。经tcr/cd3受体复合物进行抗原性t细胞刺激后,ca2+依赖性磷酸酶神经钙蛋白去磷酸化nfat上的多个磷酸丝氨酸,导致其核转运和t细胞激活基因的下游基因表达。nfat报告细胞系是通过用nfat-萦光素酶报告载体稳定转染的jurkat细胞的克隆选择而产生,其包含多个重复的nfat反应元件和萤火虫萦光素酶编码区上游的最小启动子。已确认用pma/离子霉素诱导该报告细胞后nfat激活。方法用wga-tb标记jurkatnfat报告细胞:离心收集jurkatnfat细胞,重悬在pbs/1%fcs或生长培养基中,以获得4x106细胞/ml。随后,通过加入0.5ng/ulwga-铽并在rt孵育30分钟来标记1x106个细胞。孵育后,通过280g离心8分钟,用pbs洗涤细胞两次。最后,将细胞重悬在pbs/1%fcs或生长培养基中,以获得4x106细胞/ml的最终细胞数。结合将d2标记的结合cd3的抗体以及d2标记的非结合对照igg重悬在pbs/1%fcs或生长培养基中,按1:2稀释步骤产生终浓度50-0.1nm的系列稀释。将40000(10ul)个标记细胞在384孔板中与5ul标记抗体和5ulpbs/1%fcs或生长培养基混匀。37℃孵育结合测定5小时。通过测定665nm吸光度以及在60秒延迟时间后测定615nm吸光度来用m1000proreader测量tr-fret信号。40000个wga-tb标记的jurkat-nfat细胞作为空白。测定各孔的665nm/615nm比值来将特异性fret信号对各孔中的供体信号归一化。此外,在graphpadprism6.0中测定kd和ec50值之前从样品的比值扣除空白的比值。用非线性回归拟合结合曲线。使用方程y=―(―b”_―max”―*x”)/(―(――k”_―d””+x)”),用“单部位特异性结合”模型测定bmax和kd。评价htrf原始数据的评价。对于测定的分析,首先通过microsoftexcel编辑原始数据。在htrf测定中,结果的变异可由于细胞、培养基添加剂的吸取步骤而发生在孔与孔之间,及发生自每孔裂解的细胞数。为了最小化这些变异,通过计算665nm与620nm比值将受体的发射对各孔中供体信号的发射归一化:―比值=――665nm”/―620nm”―*10000”。通过称为graphpadprism6.0的软件评价计算的比值。用非线性回归拟合结合曲线。用以下方程用“单部位特异性结合”模型测定bmax和kd:―y=―(―b”_―max”―*x”)/(―(――k”_―d””+x)”)。功能测定-萦光素酶测定通过在37℃孵育6小时后裂解细胞来从与结合测定相同的孔测量cd3信号发放。因此,每孔去除5μl测定混合物,然后加入15μone-gloluciferase试剂。rt避光孵育5-10分钟后,以1000ms积分时间用m1000proreader测量发光信号。作为空白,扣除含萦光素酶试剂的裂解的非激活细胞的信号。测量一式三份进行。结果结合测定:可以在生长培养基(图23a)以及pbs/1%fcs(图23b)中检测到cd3结合。非结合对照在此测定系统中未显示任何结合。抗cd3在生长培养基中与cd3的结合kd值比在pbs/1%中高约7倍,分别为7nm和0.98nm。功能测定结果显示,虽然在两种条件下观察到相似的结合,但cd3结合时的信号发放仅在生长培养基中存在(图24a),而在pbs/1%fcs中不存在(图24b)。萦光素酶活性的ec50位1.4nm。结论jurkatnfat报告细胞系中的cd3结合和功能性评估实验是其中可在一个孔中以高通量形式测量结合和功能性的测定系统的另一实例。当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1