一种利用植物雄性不育材料进行回交育种的方法及应用与流程
本发明属于植物育种技术领域,涉及一种利用植物雄性不育材料进行回交育种的方法及其在多系品种培育中的应用。
背景技术:
随着大量植物有利性状控制基因(或遗传位点)的鉴定及克隆,对这些基因(或遗传位点)进行育种应用已经成为当前育种的主要任务。对已知基因(或遗传位点)进行育种利用主要通过分子标记辅助选择方法进行,目前的主流做法是将少数几个有利基因通过分子标记辅助选择聚合于某个品种中,而随着有利基因的数量增加,将之聚合于一个品种变得十分困难,如果欲将位于相同染色体上的不同基因聚集于一个品种更是难上加难。而培育多系品种则可绕开这些困难,又可达到类似聚合多个有利基因的效果。
多系品种是将不同品系以一定比例混合而成的品种。组成多系品种的不同品系可以是近等基因系,也可以是肉眼可见农艺性状无明显差异但相互之间遗传差异比近等基因系之间要大一些的品系。多系品种培育的核心步骤在于其组成的不同品系的培育,这些遗传背景和农艺性状几乎一致但各自具有不同特异优良性状的多个品系一般通过回交方法选育而来。多系品种不同品系的培育因涉及多个基因的分子标记辅助回交育种,对于不易去雄的作物,进行杂交时去雄工作需耗费大量劳力,需收获较多杂交种时这种情况更加严重,使得多个基因的回交导入工作变得十分困难,育种周期较长,这限制了多系品种的培育。因此,实践中需要一种提高工作效率的方法,以加快多系品种的选育进程。
植物雄性不育在生产中往往用于避免自交,例如应用于杂种优势利用和轮回选择。植物中自然产生或通过诱变发生的雄性不育一般分为细胞质雄性不育和核雄性不育两种类型,与细胞质雄性不育相比,核雄性不育更为普遍,而核雄性不育中,又以隐性雄性不育最为常见,迄今为止在不同作物中鉴定了很多隐性雄性不育基因。由于十分易于创制,遗传育种领域研究者在积极挖掘隐性雄性不育基因的用途,不过与细胞质雄性不育已广泛应用于杂种生产相比,核隐性不育真正应用于生产的较少。目前为止,核雄性不育仅被应用于轮回选择和作为新一代杂种生产技术的spt(seedproduction)技术中,且未大范围推广应用。
技术实现要素:
本发明旨在针对现有植物多系品种培育工作量大、效率低下的缺点,提出一种利用植物雄性不育材料进行回交育种的方法及应用,在多系品种不同品系的培育过程中引入一种雄性不育系,将其应用于回交选育过程,可大幅度减少去雄工作的劳力耗费,并且易于增加杂交种数量从而增加目标单株的出现概率进而缩短育种进程,提高效率。
为了实现上述技术目的,本发明是通过如下技术方案实现的:
第一方面,本发明提供了一种利用植物雄性不育材料进行回交育种的方法,包含如下步骤:
1)将轮回亲本改造成不育性状受单个核隐性雄性不育基因座位控制的雄性不育株系;
2)以步骤1)改造得到的轮回亲本不育株系为母本,以含有某种已知有利目的基因的供体亲本为父本进行杂交,获得杂交种f1;
3)繁殖杂交种f1,以其为父本、轮回亲本不育株系为母本进行杂交,获得bc1f1种;
4)繁殖bc1f1群体,选择肉眼可见农艺性状倾向于轮回亲本且含有供体亲本有利基因的可育或不育单株继续回交:
当选择的单株为可育单株时,以其为父本、轮回亲本不育株系为母本进行杂交获得bc2f1种;
当选择的单株为不育单株时,以其为母本、以原始轮回亲本或轮回亲本不育基因杂合可育株系为父本进行杂交获得bc2f1种;
5)繁殖bc2f1群体,依照步骤4)回交的方法继续回交1次以上,获得bcnf1群体;
6)在步骤5)最后一次回交获得的bcnf1群体中选择含有目的基因且肉眼可见农艺性状与轮回亲本无明显差异的可育株自交,收获其种子繁殖获得bcnf2分离群体,从中选择供体亲本有利基因纯合且不含不育基因的可育株即完成回交育种。
优选地,所述植物为二倍体植物。
通常,选定一个优良亲本作为背景亲本,将其定为回交改良的轮回亲本;优选地,步骤1)中,用于改造的方法包括:基因编辑技术、(传统)诱变育种技术和回交育种技术,更优选地,选择基因编辑技术。
优选地,步骤1)中,控制雄性不育的核隐性雄性不育基因位点具有如下特点:其控制产生的不育株与正常可育株在田间易于识别;不会引起除雄性不育之外的农艺性状变化。
优选地,步骤1)中所述的核隐性雄性不育基因与步骤2)中所述的有利目的基因处于不同的染色体上。更优选地,当向轮回亲本中导入的有利目的基因数量较多,应用一个雄性不育基因不能满足这一要求时,当引入与第一个雄性不育基因位于不同染色体的第二个雄性不育基因。
优选地,轮回亲本不育株系通过收获不育基因为杂合的单株的种子进行繁殖或者通过利用纯合不育株的营养器官进行无性繁殖。例如,水稻可利用纯合不育株的稻桩进行无性繁殖。
优选地,所述植物为水稻时,将tdr基因(msu水稻基因组数据库基因编号为loc_os02g02820;rap水稻基因组数据库编号为os02g0120500)应用于轮回亲本的雄性不育改造。
更优选地,通过crispr\cas9系统介导的基因敲除技术敲除轮回亲本中具有正常功能的tdr基因从而获得雄性不育性状,方法参考中国发明专利202010401496.9。
第二方面,本发明提供了前述利用植物雄性不育材料进行回交育种的方法的应用,即应用于多系品种的培育。
优选地,上述多系品种指由两个以上近等基因系或两个以上肉眼可见农艺性状无明显差异的品系混合而成的植物品种。
更优选地,上述多系品种的部分或全部品系通过前述利用植物雄性不育材料进行的回交育种方法选育而来。
有益效果
在生产呼唤多系品种而其培育进程缓慢的形势下,本发明提供了一种利用雄性不育材料进行回交育种的方法,并将这一方法应用于多系品种不同品系的培育,可大幅度减少了回交过程中人工杂交的工作量,使得本发明的实施者一方面可以同时进行更多有利基因的回交导入工作,另一方面可以扩大杂交规模以增加杂交种产量,从而增加后代分离群体的规模,提高农艺性状与轮回亲本接近的单株出现的概率,缩短回交育种进程,进而提高整个多系品种培育的效率,最终加速多系品种向市场推广的进程,这是本发明的第一个有益效果。
本发明的第二个有益效果是,在应用本发明提供的方法培育得到多系品种的所有品系后,可以十分便利地将回交选育过程中使用的雄性不育株系与多系品种的不同品系杂交以合成轮回选择育种的初始群体,然后进一步进行轮回选择育种的操作,选育出满足某种需求的多系品种群体或从中分离纯系。
附图说明
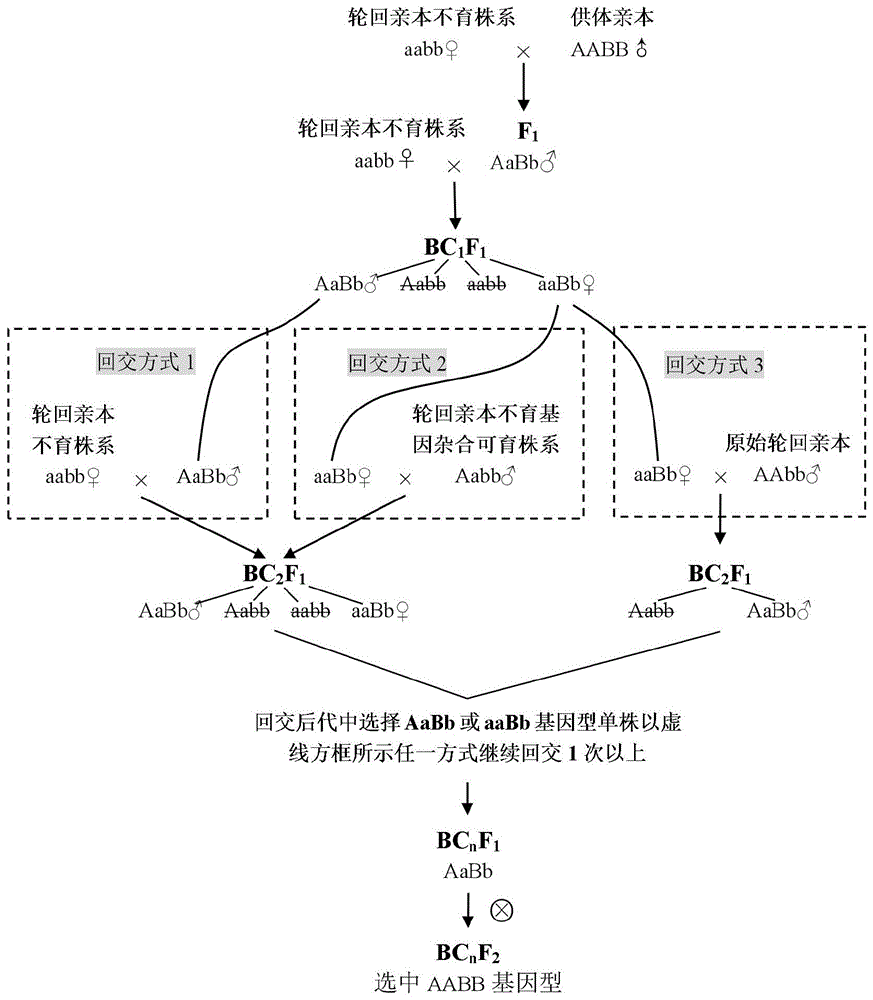
图1:本发明中利用植物隐性雄性不育材料进行回交育种的过程示意图。图中a/a表示控制雄性不育的等位基因,a为野生型,a为突变型,突变基因纯合时植株表现雄性不育;b/b表示控制某一农艺性状的等位基因,b为有利等位基因(大写表示并不一定意味着为显性),b为无对应有利功能的等位基因。基因符号全部大写或小写时表示该基因座位的两个等位基因是纯合的,而大写和小写并存时则表示杂合。图中虚线框内的内容表示3种回交方式。♀和♂分别表示杂交时作母本和父本,基因型符号中删除线表示该种基因型植株应被舍弃,×和
图2:本发明回交育种利用的水稻雄性不育基因与低镉基因序列与常规品种中野生型序列的对比。上图示雄性不育基因tdr,下图示低镉基因nramp5;图中数字表示该碱基在日本晴参考基因组tdr基因序列开放阅读框中的位置;wt,野生型;mut,突变体;下划线字母示新插入碱基;短横线示缺失碱基;tdr突变体与野生型序列相比缺失9个碱基同时插入一个新碱基a;nramp5突变体与野生型相比缺失28个碱基。
图3:回交育种所用低镉性状供体亲本osnramp5基因突变体与野生型的高分辨率溶解曲线分型效果。
具体实施方式
下面结合具体实施例对本发明进行详细说明。以下实施例将有助于本领域的技术人员进一步理解本发明,但不以任何形式限制本发明。应当指出的是,对本领域的普通技术人员来说在不脱离本发明构思的前提下,还可以做出若干变形和改进。这些都属于本发明的保护范围。
本发明提供的一种利用植物雄性不育材料进行回交育种的方法,回交过程如图1所示,主要包含如下步骤:
1)将轮回亲本改造成不育性状受单个核隐性雄性不育基因座位(a/a)控制的雄性不育株系(aabb);
2)以步骤1)改造得到的轮回亲本不育株系为母本(aabb♀),以含有某种已知有利目的基因(b)的供体亲本为父本(aabb♂)进行杂交,获得杂交种f1(aabb);
3)繁殖杂交种f1,以其为父本(aabb♂)、轮回亲本不育株系为母本(aabb♀)进行杂交,获得bc1f1种(aabb,aabb,aabb,aabb);
4)繁殖bc1f1群体,选择肉眼可见农艺性状倾向于轮回亲本且含有供体亲本有利基因的可育或不育单株(aabb或aabb)继续回交:
当选择的单株为可育单株时,以其为父本(aabb♂)、轮回亲本不育株系为母本(aabb♀)进行杂交获得bc2f1种(如图1回交方式1所示);
当选择的单株为不育单株时,以其为母本(aabb♀)、以原始轮回亲本或轮回亲本不育基因杂合可育株系为父本(aabb♂或aabb♂)进行杂交获得bc2f1种(如图1回交方式2或3所示);
5)繁殖bc2f1群体,依照步骤4)回交的方法继续回交1次以上,获得bcnf1群体;
6)在步骤5)最后一次回交获得的bcnf1群体中选择含有目的基因且肉眼可见农艺性状与轮回亲本无明显差异的可育株(aabb)自交,收获其种子繁殖获得bcnf2分离群体,从中选择供体亲本有利基因纯合且不含不育基因的可育株(aabb♂)即完成回交育种。
下列实施例中未注明具体条件的实验方法,通常按照常规条件,例如书籍《分子克隆实验指南(第3版)》(j.萨姆布鲁克,d.w.拉塞尔著;黄培堂等译;科学出版社,2008)中所述的条件,或按照制造试剂或设备厂商所建议的条件。
农艺性状(agronomictraits)指农作物的生育期、株高、叶面积、果实重量等可以代表作物品种特点的相关性状。不同农作物“肉眼可见农艺性状无明显差异”依据本领域现有的判定标准,在此不做限定。
实施例1、在水稻中利用tdr基因功能缺失型隐性雄性不育材料进行回交育种
水稻是我国最为重要的农作物,也是一种典型的二倍体植物,在本发明的具体实施方式中,我们以水稻为实施对象对本发明的技术方案进行展示。具体地,利用tdr基因功能缺失引起的雄性不育进行回交育种,向背景亲本品种台南11(一种热带粳稻品种)中导入osnramp5突变基因从而创制低镉台南11品系。具体步骤如下:
1、将背景亲本台南11改造成雄性不育材料
通过crispr/cas9系统介导的基因敲除技术创制tdr基因突变的台南11雄性不育材料(试验过程及雄性不育材料的繁殖参见发明专利:利用crispr\cas9系统对水稻tdr基因进行定点突变及检测的方法,申请号202010401496.9实施例1所述),本发明实施例选用的台南11雄性不育株系为发明专利202010401496.9实施例中所述的对靶位点2进行定点敲除获得的编号为##07的无标记纯合突变株系,其tdr基因突变序列与野生型序列的差别如图2上图所示。tdr突变基因与野生型的基因分型通过高分辨率溶解曲线分型法进行(方法参考发明专利202010401496.9)。雄性不育株系通过将杂合型可育株与纯合突变不育株系进行人工杂交的方式繁殖,后代群体维持杂合可育株与纯合突变不育株1:1比例不变。
2、将品种五丰b的osnramp5基因突变株系作为低镉供体亲本
品种五丰b的无标记低镉osnramp5基因突变株系通过crispr/cas9系统介导的基因敲除技术创制(参见文献:龙起樟,黄永兰,唐秀英,王会民,芦明,袁林峰,万建林.利用crispr/cas9敲除osnramp5基因创制低镉籼稻,中国水稻科学,2019,33(5):407-420)。突变株系中osnramp5基因的序列与野生型的差别如图2下图所示。突变体与野生型之间进行基因分型采用高分辨率溶解曲线方法进行(参见本段前述文献或发明专利:一种osnramp5基因特定位点突变的高分辨率溶解曲线检测方法,201910394240.7),分型效果如图3所示。
3、配制f1杂种
在南昌于6月中旬播种,将台南11雄性不育株系与其杂合体群体进行种植,而五丰b低镉株系因其生育期较短,延迟至7月中旬播种。在9月上旬在田间通过观察花药是否含有花粉选出台南11群体中的雄性不育株系,将其作母本与五丰b低镉株系进行杂交,收获杂种约200粒。
4、bc1f1种子的获得
冬季在海南繁殖f1杂种,同期繁殖台南11雄性不育株系与杂合可育株混合的群体,将f1植株作父本与台南11群体中的雄性不育株系进行人工杂交获得bc1f1种子,收获杂交种子约1000粒。
5、bc2f1种子的获得
正季在南昌繁殖bc1f1群体,分单株种植,同期繁殖台南11原始亲本,最终存活bc1f1植株654株,抽穗扬花期观察发现,bc1f1群体存在可育株和不育株的分离,大约各占一半,从不育株中选出株叶形态类同台南11亲本者20株,利用高分辨率熔解曲线基因分型技术对其nramp5基因型进行检测,以选出杂合突变型单株,结果发现8株为杂合突变型(另外12株是野生型)。以nramp5杂合突变型不育株作母本,台南11原始亲本作父本进行人工杂交获得bc2f1种子,收获800粒左右杂交种。
6、bc3f1种子的获得
冬季在海南繁殖bc2f1群体,同期种植台南11雄性不育株系及其杂合体群体,最终存活bc2f1植株486株,观察发现植株全部为可育,筛选20株株叶形态与野生型高度类似者,通过高分辨率熔解曲线基因分型法对这些植株的nramp5基因型进行鉴定,发现9株为杂合突变型(余为野生型),将nramp5杂合突变型可育株作为父本与台南11群体中的雄性不育株系杂交,获得bc3f1种,收获种子约500粒。
7、bc3f2种子的获得
正季在南昌繁殖bc3f1群体,获得344个单株,观察发现不育株与可育株约各占一半,从可育株中选择与台南11株叶形态几乎相同的单株10株,通过高分辨率熔解曲线基因分型法对这些植株的nramp5基因型进行鉴定,发现4株为杂合突变型(余为野生型),使此4个单株自交繁殖,获得bc3f2种子。
8、完成回交选育过程
在海南繁殖bc3f2群体约300株,选择92株与野生型株叶形态一致的可育株通过高分辨率熔解曲线分型技术进行基因型鉴定,筛选出tdr基因为野生型同时nramp5基因为纯合突变型的单株,共获得4个目标单株,让这些单株自交。回交过程结束,完成低镉基因导入。
在本实施例中,通过2次回交即获得了株叶形态与野生型高度类似的单株,而回交3次则与野生型无异,且基本稳定,较常规方法回交次数至少减少2次以上,能够减少回交次数是因为试验中回交群体较常规做法大得多,这增加了野生型类似者出现的概率。仅导入少数基因时这种做法优势不明显,但导入较多基因时则可极大提高工作效率,大幅度缩短育种年限。
- 还没有人留言评论。精彩留言会获得点赞!