一种验证东亚小花蝽胁迫影响大豆蚜体内内源物质活力的方法
1.本发明涉及一种验证东亚小花蝽胁迫影响大豆蚜体内内源物质活力的方法,属于农业科学技术领域。
背景技术:
2.东亚小花蝽(orius sauteri)隶属于半翅目花蝽科,是我国北方的重要捕食性天敌昆虫之一,具有发生早、时间长、分布广、数量多、食性杂、活动能力强等特点(杨淑斐等,2009)。它主要取食蓟马、蚜虫、叶螨、粉虱等小型昆虫、植物花粉,以及夜蛾类昆虫的卵和小幼虫(nagayi k,1997),在控制果园、农田、温室花卉和林木等虫害的发生发挥着重要作用,已经成为昆虫天敌开发利用的主要品种。国内先后对东亚小花蝽的生物学特性、发生、消长、活动规律和人工饲养等进行了相关研究(xu x n and enkegaard a,2009;武予清等,2010;霍捷等,2011;刘文静等,2011;黄增玉等,2011;郅军锐等,2011;蒋兴川等,2012;张昌容等,2012;马广民等,2012;王洪亮等,2013),但对其释放和控制效果方面的研究报道较少(蒋月丽等,2011;孙猛等,2012)。大豆蚜作为田间大豆的主要害虫之一,亦可引发由其作为媒介的多种病毒病等,若对其不加以及时控制,将带来无法估计的损失。充分发挥天敌资源的作用,是建立大豆蚜防控体系的重要环节。在室内研究中东亚小花蝽已经表现出对大豆蚜很强的捕食能力和控制潜力。捕食者-猎物相互作用是节肢动物害虫生物防治的核心,在农业害虫管理和自然资源保护等领域具有重要的应用价值(tscharntke t.et al.,2007;straub c.s.et al.,2008;zehnder g.et al.,2007;de clercq p.et al.,2011;lu y.et al.,2012;attia s.et al.,2013)。减少劳动力、农药和设备的成本,以及可能返回到入侵前的生态服务,构成了在未来几年加强保护生物控制的有说服力的论据(liu yq.et al.,2014;pickett c h.and bugg r.l.,1998;landis d a.et al.,2000)。波寇诺-甘塔评估芒果粉蚧生物控制经济效益时,发现收益-成本比为145:1(bokonon-ganta.et al.,2002;de groote h.et al.,2003)。这还没有衡量有些未成功的生物控制计划的成本(cortesero a.et al.,2000;altieri m a.and nicholls c i.,2004;messing r h.and wrightmg.,2006;jonsson m.et al.,2008)。然而,应注意与非目标生物体相互作用的可能性(howarth f g.,1991;louda s m.et al.,2013)。为了减少或避免这种相互作用,已经开发了新的工具,包括尽可能在目标栖息地进行的宿主特异性试验(desneux n.et al.,2009;desneux n.,2012)。今天,大量捕食的节肢动物被收集,并且大规模饲养,然后在增强生物控制计划的目标农业生态系统中释放(pickett ch.and buggrl.,1998;symondson w.,2002)。这些计划受益于可用于吸引,收集和监测自然栖息地和目标作物的捕食性节肢动物的工具。自然界中存在的大豆蚜天敌种类繁多,戴长春和王坤等通过对大豆蚜田间天敌昆虫的群落结构分析发现,在大豆蚜发生初期,少量的小花蝽、龟纹瓢虫、叶色草蛉和捕食性蜘蛛即在田间活动,在一定程度上延缓了大豆蚜暴发期的临近,减少其初期发生数量;异色瓢虫、中华草蛉、黑食蚜盲蝽、窄姬猎蝽、黑带食蚜蝇等大多数天敌优势种群在大豆蚜发生数量较多时
出现,在大豆蚜发生高峰期起到一定的控害作用。而蚜小蜂、食蚜瘿蚊在大豆蚜的发生后期大量发生,减少下一年迁飞回豆田的虫口基数,压低虫源(戴长春等,2009)。对于农业生态系统,明确害虫与天敌资源,掌握群落的结构和特点,了解生态系统中昆虫群落的结构及其动态,以农田生态系统为调控单位,从群落的角度出发,充分发挥自然控制力,即充分利用生物自身的相互促进,相互制约的关系及非生物环境来调控有害生物的种群数量,以达到防止其造成经济损害的目的,是维护自然平衡和治理有害生物的基础(wratten,1987;丁岩钦,1993;戈峰,1998)。利用捕食性天敌昆虫在自然界中的调控作用,以虫治虫,控制虫灾,是保证农业增产、保护农业清洁环境的有效途径,也有利于实现农业生态平衡、保护生物多样性和实现人与各种生物和谐相处的崇高目标。东亚小花蝽作为一种具有很好利用价值的天敌资源,在棉花、玉米、高粱、黄豆和多种蔬菜等作物上,对蚜虫、蓟马和粉虱等一些小型害虫以及夜蛾类昆虫的卵和小幼虫都有良好的控制与调节作用(董天宇等,2015)。作为大豆蚜的天敌,其存在必定对大豆蚜会产生一定的胁迫作用,大豆蚜在经过视觉冲击、气味感受刺激与直接接触后会产生一定的体内物质含量的波动,这种波动的产生必定与体内参与呼吸作用、神经传递、解毒能力和产生自我保护功能的各种物质产生与变化有关,而一种物质的变化,为维持体内的恒定,必将导致与其有关的物质也一定会产生变化。如生物受到刺激后乙酰胆碱的提升,为维护体内的安全,其体内分解乙酰胆碱的酯酶含量也会随之增加,使得生物体内在一定程度下维持一个相对的平衡,保护大豆蚜不会因一种物质过度的变化而使机体受损,这种机制我们常称为自我保护机制,存在于各种生物体中。本发明中,通过测定分析受东亚小花蝽胁迫后不同龄期大豆蚜体内各生理指标的变化差异显著,初步探讨了不同龄期大豆蚜体内不同生化物质含量及保护酶活性变化与东亚小花蝽胁迫对大豆蚜防御机制之间的关系,为研究东亚小花蝽的胁迫机制及综合治理提供了理论依据。海藻糖是昆虫体内主要的血糖,海藻糖有不同于其他碳水化合物的生物学特性,能在恶劣环境下保护生物体细胞蛋白质、脂肪、糖类、核酸等组分不受损害。在恶劣环境条件下在细胞表面能形成独特的保护膜,有效地保护蛋自质分子不变性失活,从而维持生命体的生命过程和生物特征(crowe et al.,1990;williams,1990;程媛,2016)。本发明中,利用东亚小花蝽对不同龄期大豆蚜进行多种方式胁迫后,各个龄期的大豆蚜体内海藻糖含量都显著高于对照组,这表明受胁迫后,大豆蚜的自身保护机制启动,导致大豆蚜体内的保护性物质海藻糖的合成都显著提升,理论上证明东亚小花蝽的胁迫作用可以有效刺激大豆蚜启动防御机制,从而导致大豆蚜提前处于应激状态,一段时间后体质下降,致使大豆蚜的防御机制下降,这与(王颖颖等,2015)浅黄恩蚜小蜂寄生对烟粉虱海藻糖含量的影响研究结果相类似。从总体来看,直接胁迫对大豆蚜体内海藻糖含量的影响更为显著,而视觉胁迫随龄期的降低其显著性有所下降。这表明高龄时大豆蚜的海藻糖含量受东亚小花蝽视觉胁迫影响更为显著,这可能由于成蚜的发育比较完善,所以对视觉胁迫的感受能力更强。但也可能是因为24h的胁迫处理后,低龄大豆蚜较高龄大豆蚜体内的海藻糖含量先受海藻糖酶影响恢复为一个较为稳定的状态,而造成影响合成或解离相关的蛋白及其基因还应做进一步的研究。还原糖主要包括葡萄糖、果糖和麦芽糖等,其中葡萄糖和果糖是呼吸作用的主要底物。所以在东亚小花蝽胁迫后,大豆蚜体内的呼吸作用加强,需要进行糖酵解的底物需要大量增加,以供呼吸作用消耗,所以受胁迫后,大豆蚜体内的还原糖含量大幅度增加,但是成蚜阶段的还原糖含量虽然略有提升,但在0.05水平并无显著性差异,原因可能是成蚜受胁迫后对其
呼吸作用并无太大影响,而低龄期的大豆蚜受到的影响在0.01水平有显著性差异,这说明东亚小花蝽胁迫对低龄期的大豆蚜影响十分显著。总体上,东亚小花蝽接触胁迫和气味胁迫对大豆蚜的影响更为显著,这说明低龄期的大豆蚜受气味胁迫的影响较比视觉胁迫更为显著。昆虫中的蛋白质有很多种,这些蛋白质不仅与组织形成的物质代谢有关,而且与虫体生长发育的激素调控以及产生抗药性和免疫机制等都有密切的关系(孙昊雨等,2014)。本发明中分别利用视觉冲击、气味感受刺激与直接接触对各龄期大豆蚜进行处理。结果表明,总体上受东亚小花蝽胁迫后的各龄期大豆蚜体内的蛋白含量均显著低于对照组,这可能是由于大豆蚜受胁迫影响后,其体内的各种酶类等物质的合成与呼吸等系统的消耗。而从龄期来看的话,视觉冲击造成的胁迫对高龄期大豆蚜体内蛋白含量的影响较为显著,而在低龄期时,东亚小花蝽与大豆蚜直接接触产生的胁迫对大豆蚜体内的蛋白含量影响比较显著,这可能是因为大豆蚜在成蚜阶段其视觉系统发育的更加完善,而且因东亚小花蝽的捕食性,其更喜好捕食低龄期的大豆蚜,所以接触可能对高龄期的大豆蚜影响不如低龄期显著。而就胁迫方式之间的差异来看,【直接胁迫影响的更为彻底(变化大),而气味与视觉对其影响较为稳定】。造成蛋白如此变化必定会导致酶类物质的变化,而造成影响合成或解离相关的蛋白及其基因还应做进一步的研究。昆虫体内的保护酶相互协调作用,保护昆虫体内的平衡,防御不良条件和物质对生物体的损害。寄主保护酶活性变化可作为测定外界刺激物的强弱和昆虫耐药及抗逆能力的重要指标(任晓亚,2014;武海斌,2018)。大豆蚜在受东亚小花蝽胁迫后产生多种应激防御反应机制,而酶恰好是这些反应的基础,其中,ache,care和gsts是重要的解毒和保护酶。乙酰胆碱酯酶属于丝氨酸水解酶,广泛存在于各种动物组织和血清中。乙酰胆碱酯酶催化乙酰胆碱水解,在神经传导中起调节作用。在本发明中,各个龄期大豆蚜受到东亚小花蝽胁迫后,高龄期大豆蚜乙酰胆碱酯酶比活力与对照组虽有显著性差异,但较低龄期大豆蚜的乙酰胆碱酯酶比活力与对照组相比,变化差异幅度相差很多。这说明东亚小花蝽的胁迫对低龄期的大豆蚜更为显著。而且气味和直接接触造成的胁迫是对低龄期大豆蚜产生刺激更为显著的因素。羧酸酯酶参与脂质的运输和代谢,并且与多种药物、毒物的解毒和代谢有关。对其产生影响与否是对大豆蚜进行生物防治和化学防治等综合防治的重要影响因子。因此如何验证东亚小花蝽胁迫影响大豆蚜体内内源物质的活力成为急需解决的一大难题,采用东亚小花蝽胁迫大豆蚜,经过实验,可以看出,东亚小花蝽胁迫对大豆蚜的羧酸酯酶比活力影响显著。并且发现3龄、4龄和成蚜组成的高龄期大豆蚜其羧酸酯酶比活力受东亚小花蝽视觉胁迫的影响更为显著,而1龄和2龄组成的低龄期大豆蚜其羧酸酯酶比活力受东亚小花蝽气味胁迫的影响更为显著。其原因可能是由于高龄期大豆蚜视觉发育完善,而低龄期大豆蚜视觉发育不完善,所以更依靠触角对气味的识别,从而被东亚小花蝽气味胁迫造成的影响更为明显。谷胱甘肽-s-转移酶是体内解毒酶系统的重要组成部分,主要催化各种化学物质及其代谢产物与gsh的巯基共价结合,使亲电化合物变为亲水物质,达到将体内各种潜在或具备毒性的物质降解并排出体外的目的。所以受东亚小花蝽胁迫后的大豆蚜体内的谷胱甘肽-s-转移酶比活力与羧酸酯酶呈现出大致相同的情况,受胁迫的成蚜和4龄若蚜组成的高龄期大豆蚜,其受东亚小花蝽视觉胁迫的影响更为显著,而3龄、2龄和1龄若蚜组成的低龄期大豆蚜,其受东亚小花蝽气味胁迫和直接接触造成的胁迫影响更为显著,发明一种验证东亚小花蝽胁迫影响大豆蚜体内内源物质活力的方法是必要的。
技术实现要素:
3.为了克服如何验证东亚小花蝽胁迫影响大豆蚜体内内源物质的活力的难题,本发明提供了一种验证东亚小花蝽胁迫影响大豆蚜体内内源物质活力的方法,该种一种验证东亚小花蝽胁迫影响大豆蚜体内内源物质活力的方法采用东亚小花蝽胁迫大豆蚜,经过实验,可以看出,东亚小花蝽胁迫对大豆蚜的羧酸酯酶比活力影响显著。并且发现3龄、4龄和成蚜组成的高龄期大豆蚜其羧酸酯酶比活力受东亚小花蝽视觉胁迫的影响更为显著,而1龄和2龄组成的低龄期大豆蚜其羧酸酯酶比活力受东亚小花蝽气味胁迫的影响更为显著。其原因可能是由于高龄期大豆蚜视觉发育完善,而低龄期大豆蚜视觉发育不完善,所以更依靠触角对气味的识别,从而被东亚小花蝽气味胁迫造成的影响更为明显。谷胱甘肽-s-转移酶是体内解毒酶系统的重要组成部分,主要催化各种化学物质及其代谢产物与gsh的巯基共价结合,使亲电化合物变为亲水物质,达到将体内各种潜在或具备毒性的物质降解并排出体外的目的。所以受东亚小花蝽胁迫后的大豆蚜体内的谷胱甘肽-s-转移酶比活力与羧酸酯酶呈现出大致相同的情况,受胁迫的成蚜和4龄若蚜组成的高龄期大豆蚜,其受东亚小花蝽视觉胁迫的影响更为显著,而3龄、2龄和1龄若蚜组成的低龄期大豆蚜,其受东亚小花蝽气味胁迫和直接接触造成的胁迫影响更为显著。
4.本发明解决其技术问题所采用的技术方案是:
5.本发明一种验证东亚小花蝽胁迫影响大豆蚜体内内源物质活力的方法,具体实验步骤如下:
6.1材料与方法
7.1.1供试材料及仪器-8.1.1.1供试昆虫
9.东亚小花蝽orius sauteri采自黑龙江省向阳试验农场大豆田间,饲养在恒温培养箱(温度t=25
±
1℃、相对湿度rh=70%
±
5%、光照l∶d=16∶8)内,用大豆蚜饲喂,用黄豆芽作产卵植物繁殖东亚小花蝽。
10.大豆蚜aphis glycines为实验室长期保留的哈尔滨本地敏感种群,人工盆栽大豆苗饲养,置于温度25
±
1℃,相对湿度65%-70%,光周期的14l:10d的人工气候箱内,供试大豆品种为东农52。
11.1.1.2实验试剂
12.考马斯亮蓝g-250和牛血清蛋白(bsa)购自北京宝日医生物技术(takara)有限公司。磷酸缓冲液(pb)购自上海生工生物工程股份有限公司。乙酰胆碱酯酶(ache)、羧酸酯酶(care)、谷胱甘肽-s-转移酶(gsts)、海藻糖、还原糖均购自北京索莱宝(solarbio)科技有限公司。
13.1.1.3仪器设备
14.高速低温冷冻离心机(thermosorvallst16r)、人工气候箱(bic,上海博讯实业有限公司医疗设备厂)、多功能酶标仪(infinite-200pro,瑞士tecan公司)、烘箱(dph781,湖北省黄石市医疗器械厂)、冰箱(美菱)、电热恒温水浴锅(dzkw-s-4,北京永光明医疗仪器厂)、石英砂(分析纯,天津市巴斯夫化工有限公司)、无水乙醇(天津市天力化学试剂有限公司)
15.1.2东亚小花蝽对大豆蚜的胁迫处理
16.用东亚小花蝽成虫对各龄期的大豆蚜进行胁迫处理,处理方式分为三种:接触胁迫、气味胁迫和视觉胁迫,对照组为无东亚小花蝽胁迫。每个自制培养皿上层放入一定数量的大豆蚜,下层放入东亚小花蝽成虫,培养皿中间用透明可见的塑料板和不可见的带有针孔的透气滤隔离。处理后置入温度24
±
1℃、相对湿度70%
±
5%、光照为16l:8d的培养箱内。24h后,取受胁迫的大豆蚜分别进行各种生化物质测定。
17.1.3酶液制备
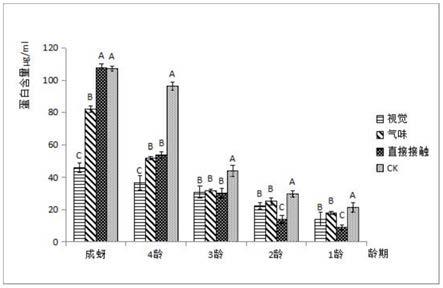
18.参照高欢欢等(2016)的酶液提取法,24h后,分别于各个处理中挑取5个龄期的大豆蚜,每个龄期的大豆蚜收集5只存活的虫体置于1.5ml的eppendorf离心管中,加入400μl0.02mol/l的磷酸缓冲液(nacl 136.89mm;kcl 2.67mm;na2hpo4 8.1mm;kh2po4 1.76mm)和少量石英砂,冰浴充分研磨,并将研磨后的匀浆置于高速冷冻离心机8000g、4℃离心10min。取上清液分装到新的1.5ml的eppendorf离心管置于冰上待测。各个处理的5个虫龄均设置3次重复。
19.1.4蛋白质、还原糖和海藻糖含量的测定
20.1.4.1蛋白质含量的测定
21.参照bradford(1976)的考马斯亮蓝g-250染色法测定。分别配制考马斯亮蓝g-250溶液和牛血清蛋白标准溶液,制作标准曲线。每个样品取4μl酶液于酶标板的加样孔中,对照孔中加入4μl的磷酸缓冲液,再加入200μl复温的考马斯亮蓝g-250试剂,充分混合,室温下放置5min后,在酶标仪波长设定在595nm波长处进行测定,记录od值,重复3次,根据蛋白标准曲线,计算出各个待测样中的蛋白浓度。
22.1.4.2还原糖、海藻糖含量的测定
23.采用蒽酮硫酸法测定海藻糖的含量,分别配制蒽酮溶液和10mg/ml的海藻糖标准溶液,制作标准曲线。每个样品取60μl样本液和240μl配置好的蒽酮试剂于1.5ml的eppendorf离心管中混匀,95℃水浴10min,(盖紧,防止水分散失)自然冷却至室温,取200μl至酶标板中,在620nm波长下进行测定,记录od值,重复3次,根据海藻糖标准曲线,计算出各个待测样中的海藻糖含量。
24.采用dns法测定还原糖的含量,分别配置3,5-二硝基水杨酸(dns)碱性溶液和10mg/ml的葡萄糖标准溶液,制作标准曲线。测定管中,每个样品取175μl样本液和125μl配置好的dns试剂于1.5ml的eppendorf离心管中混均匀,沸水浴中加热5min(盖紧,防止水分散失),取出后立即冷却至室温,混匀。对照管中,每个样品取175μl样本液和125μl蒸馏水于1.5ml的eppendorf离心管中混均匀,沸水浴中加热5min(盖紧,防止水分散失),取出后立即冷却至室温,混匀。取200μl至96孔酶标板中,在540nm波长下进行测定,记录od值,重复3次。计算测定管减去对照管的吸光值差值,根据葡萄糖标准曲线,计算出各个待测样中的还原糖含量(仵均祥等,2004;丁慧梅,2011;程媛等,2016)。
25.1.5酶活性的测定
26.ache、care和gsts酶比活力的测定均按照北京索莱宝科技有限公司试剂盒中的说明进行检测。酶活性值根据试剂盒说明书中的公式进行计算。ache、care和gsts酶比活力测定波长分别为412、450和340nm。ache活性单位定义为每毫克组织蛋白每分钟催化产生1nmol摩尔消光系数(tnb)为一个酶活单位;care活性单位定义为37℃下每毫升反应体系每毫克组织蛋白每分钟催化吸光值增加1定义为一个酶活力单位;gst活性单位定义为在25℃
或者37℃中,每毫克蛋白每分钟催化1μmol1-氯-2,4-二硝基苯(cdnb)与谷胱甘肽(gsh)结合为一个酶活性单位。
27.1.6数据分析
28.所有试验数据均采用microsoft excel表格处理软件和spss base ver.19.0统计软件进行数据分析和作图,通过duncan's新复极差法检验分析处理间差异性。对受东亚小花蝽胁迫后大豆蚜体内蛋白质、海藻糖和还原糖含量以及ache、care和gsts活性分别采用单因素方差分析。
29.2结果
30.2.1东亚小花蝽胁迫对大豆蚜蛋白含量的影响
31.东亚小花蝽胁迫大豆蚜24h后,随着龄期的增长其酶液蛋白含量呈一直上升的趋势。由图1显示,和对照组相比,在成蚜阶段时,气味胁迫与视觉胁迫处理组体内酶液蛋白含量分别降低了23.42%和57.27%,且视觉胁迫处理组与其他组间差异极显著,而直接接触的胁迫与对照组间并无显著差异;在4龄时和对照组相比,气味胁迫和视觉胁迫处理组的蛋白含量分别降低了46.36%和61.85%,直接接触的胁迫则降低了44.33%;在3龄时,气味胁迫和视觉胁迫处理组分别降低了27.99%和30.15%,直接接触的胁迫则降低了31.54%,且三个处理组间无显著性差异;在2龄时,气味胁迫和视觉胁迫处理组的蛋白含量分别降低了15.82%和24.64%,直接接触的胁迫则降低了52.73%;在1龄时,气味胁迫、视觉胁迫和直接接触胁迫处理组分别降低了15.13%,33.32%和57.01%,且直接接触胁迫处理组中的蛋白含量达到最低值(9.12μg/ml),极显著低于对照组、气味胁迫和视觉胁迫处理组(f=167.694,df=3,p《0.001)。
32.2.2东亚小花蝽胁迫对大豆蚜海藻糖含量的影响
33.东亚小花蝽对大豆蚜胁迫24h后,随着龄期的增长其海藻糖含量表现为一直上升的趋势。由图2显示,和对照组相比,在成蚜阶段时,气味胁迫与视觉胁迫处理组体内海藻糖含量分别升高了97.77%和43.49%,且直接接触胁迫处理组达到最高值为(3.4607mg/mg prot),而三种胁迫方式与对照组间均有极显著差异;在4龄时,气味胁迫和视觉胁迫处理组的海藻糖含量分别升高了67.08%和32.56%,直接接触的胁迫则升高了104.12%;在3龄时,气味胁迫和视觉胁迫处理组分别升高了52.29%和22.22%,直接接触的胁迫则升高了115.33%;在2龄时,气味胁迫和视觉胁迫处理组的海藻糖含量分别升高了72.21%和30.22%,直接接触的胁迫则升高了145.48%;在1龄时,气味胁迫、视觉胁迫和直接接触胁迫处理组分别升高了57.24%,37.96%和160.72%,而气味胁迫和视觉胁迫处理组间并无极显著差异。(f=167.694,df=3,p《0.001)。
34.2.3东亚小花蝽胁迫对大豆蚜还原糖含量的影响
35.东亚小花蝽对大豆蚜胁迫24h后,随着龄期的增长其还原糖含量表现为一直降低的趋势。由图3显示,和对照组相比,在成蚜阶段时,气味胁迫、视觉胁迫和直接接触胁迫处理组与对照组的还原糖含量均无显著性差异;在4龄时,气味胁迫和视觉胁迫处理组的还原糖含量分别升高了27.29%和11.36%,直接接触的胁迫则升高了46.10%,且三个处理组的还原糖含量均显著高于对照组,而直接接触胁迫处理组极显著高于气味胁迫与视觉胁迫处理组;在3龄时,气味胁迫和视觉胁迫处理组分别升高了69.72%和55.27%,直接接触的胁迫则升高了120%,且气味胁迫与视觉胁迫处理组间并无显著性差异;在2龄时,气味胁迫和
视觉胁迫处理组的还原糖含量分别升高了122.67%和114.88%,直接接触的胁迫则升高了168.73%;在1龄时,气味胁迫、视觉胁迫和直接接触胁迫处理组分别升高了308.95%,193.53%和371.54%,且直接接触胁迫处理组中的还原糖含量达到最高值(2.89μg/mg prot),极显著高于对照组、气味胁迫和视觉胁迫处理组(f=167.694,df=3,p《0.001)。
36.2.4东亚小花蝽胁迫对大豆蚜乙酰胆碱酯酶比活力的影响
37.东亚小花蝽对大豆蚜胁迫24h后,随着龄期的增长其乙酰胆碱酯酶比活力表现为一直降低的趋势。由图4显示,和对照组相比,在成蚜阶段时,气味胁迫与视觉胁迫处理组体内乙酰胆碱酯酶比活力分别降低了68.34%和70.57%,且气味胁迫和视觉胁迫处理组间并无显著性差异极,而直接接触的胁迫与对照组间并无显著差异;在4龄时,气味胁迫和直接接触胁迫处理组的乙酰胆碱酯酶比活力分别升高了56.91%和104.57%,且气味胁迫和直接接触胁迫处理组均极显著高于对照组,而视觉胁迫处理组与对照组间并无显著性差异;在3龄时,气味胁迫和视觉胁迫处理组分别升高了115.19%和28.70%,直接接触的胁迫则升高了174.31%,且三个处理组间均有显著性差异并都极显著高于对照组;在2龄时,气味胁迫和视觉胁迫处理组的乙酰胆碱酯酶比活力分别升高了368.71%和136.04%,直接接触的胁迫则升高了918.05%;在1龄时,气味胁迫、视觉胁迫和直接接触胁迫处理组分别升高了4228.37%,1628.48%和8506.98%,且直接接触胁迫处理组中的乙酰胆碱酯酶比活力达到最高值(198.79u/mg prot),极显著高于对照组、气味胁迫和视觉胁迫处理组(f=167.694,df=3,p《0.001)。
38.2.5东亚小花蝽胁迫对大豆蚜羧酸酯酶比活力的影响
39.东亚小花蝽对大豆蚜胁迫24h后,随着龄期的增长其羧酸酯酶比活力表现为下降明显的趋势。由图5显示,和对照组相比,在成蚜阶段时,气味胁迫与视觉胁迫处理组体内羧酸酯酶比活力分别升高了174.30%和887.64%,且视觉胁迫与气味胁迫处理组均极显著高于对照组,而直接接触的胁迫与对照组间并无显著差异;在4龄时,气味胁迫和视觉胁迫处理组羧酸酯酶比活力分别升高了126.16%和586.81%,直接接触的胁迫则升高了124.52%,且三种胁迫方式均极显著高于对照组,而气味胁迫与直接接触胁迫处理组间并无显著性差异;在3龄时,气味胁迫和视觉胁迫处理组分别升高了55.48%和1508.95%,直接接触的胁迫则升高了173.87%,且三个处理组间均有极显著差异;在2龄时,气味胁迫和视觉胁迫处理组的羧酸酯酶比活力分别升高了1135.72%和875.35%,直接接触的胁迫则升高了581.85%;在1龄时,气味胁迫、视觉胁迫和直接接触胁迫处理组分别升高了1889.46%,357.15%和1713.34%,且气味胁迫处理组中的羧酸酯酶比活力达到最高值(0.0038u/mg prot),显著高于对照组、气味胁迫和视觉胁迫处理组(f=167.694,df=3,p《0.001)。
40.2.6东亚小花蝽胁迫对大豆蚜谷胱甘肽-s-转移酶比活力的影响
41.东亚小花蝽对大豆蚜胁迫24h后,随着龄期的增长其谷胱甘肽-s-转移酶比活力表现为下降明显的趋势。由图6显示,和对照组相比,在成蚜阶段时,气味胁迫与视觉胁迫处理组体内谷胱甘肽-s-转移酶比活力分别升高了97.62%和229.91%,直接接触的胁迫则降低了58.5%,且三个处理组与对照组间差异极显著,而直接接触胁迫处理组的谷胱甘肽-s-转移酶比活力极显著低于对照组;在4龄时,气味胁迫和视觉胁迫处理组的谷胱甘肽-s-转移酶比活力分别升高了81.78%和219.26%,直接接触的胁迫则降低了12.19%,且直接接触
胁迫处理组与对照组间无显著性差异;在3龄时,气味胁迫和视觉胁迫处理组的分别升高了20.55%和10%,直接接触的胁迫则降低了58.45%;在2龄时,气味胁迫和视觉胁迫处理组的谷胱甘肽-s-转移酶比活力分别升高了31.17%和7.39%,直接接触的胁迫则升高了156.59%,且直接接触胁迫和气味胁迫处理组均显著性高于对照组,直接接触胁迫处理组中的谷胱甘肽-s-转移酶比活力达到最高值(0.06nmol/min/mg prot),而视觉胁迫处理组与对照组间并无显著性差异;在1龄时,气味胁迫、视觉胁迫和直接接触胁迫处理组分别升高了10.23%,2.54%和104.42%,直接胁迫极显著高于对照组、气味胁迫和视觉胁迫处理组(f=167.694,df=3,p《0.001)。
42.本发明的有益效果为,一种验证东亚小花蝽胁迫影响大豆蚜体内内源物质活力的方法,采用东亚小花蝽胁迫大豆蚜,经过实验,可以看出,东亚小花蝽胁迫对大豆蚜的羧酸酯酶比活力影响显著。并且发现3龄、4龄和成蚜组成的高龄期大豆蚜其羧酸酯酶比活力受东亚小花蝽视觉胁迫的影响更为显著,而1龄和2龄组成的低龄期大豆蚜其羧酸酯酶比活力受东亚小花蝽气味胁迫的影响更为显著。其原因可能是由于高龄期大豆蚜视觉发育完善,而低龄期大豆蚜视觉发育不完善,所以更依靠触角对气味的识别,从而被东亚小花蝽气味胁迫造成的影响更为明显。谷胱甘肽-s-转移酶是体内解毒酶系统的重要组成部分,主要催化各种化学物质及其代谢产物与gsh的巯基共价结合,使亲电化合物变为亲水物质,达到将体内各种潜在或具备毒性的物质降解并排出体外的目的。所以受东亚小花蝽胁迫后的大豆蚜体内的谷胱甘肽-s-转移酶比活力与羧酸酯酶呈现出大致相同的情况,受胁迫的成蚜和4龄若蚜组成的高龄期大豆蚜,其受东亚小花蝽视觉胁迫的影响更为显著,而3龄、2龄和1龄若蚜组成的低龄期大豆蚜,其受东亚小花蝽气味胁迫和直接接触造成的胁迫影响更为显著。本发明操作简单,使用方便,效果明显,验证了东亚小花蝽胁迫影响大豆蚜体内内源物质活力,是一种有效的验证方法。
附图说明
43.下面结合附图对本发明进一步说明。
44.图1为本发明一种验证东亚小花蝽胁迫影响大豆蚜体内内源物质活力的方法的受东亚小花蝽胁迫后大豆蚜不同龄期蛋白含量值图。
45.图2为本发明一种验证东亚小花蝽胁迫影响大豆蚜体内内源物质活力的方法的受东亚小花蝽胁迫后大豆蚜不同龄期海藻糖含量变化图。
46.图3为本发明一种验证东亚小花蝽胁迫影响大豆蚜体内内源物质活力的方法的受东亚小花蝽胁迫后大豆蚜不同龄期还原糖含量变化图。
47.图4为本发明一种验证东亚小花蝽胁迫影响大豆蚜体内内源物质活力的方法的受胁迫后大豆蚜不同龄期乙酰胆碱酯酶比活力变化图。
48.图5为本发明一种验证东亚小花蝽胁迫影响大豆蚜体内内源物质活力的方法的受胁迫后大豆蚜不同龄期羧酸酯酶比活力变化图。
49.图6为本发明一种验证东亚小花蝽胁迫影响大豆蚜体内内源物质活力的方法的受胁迫后大豆蚜不同龄期谷胱甘肽-s-转移酶比活力变化图。
具体实施方式
50.实施例一:
51.本发明一种验证东亚小花蝽胁迫影响大豆蚜体内内源物质活力的方法,目的:大豆蚜aphis glycines是危害我国大豆产量的重要刺吸式害虫,东亚小花蝽oriussauteri是我国东北地区大豆蚜的主控天敌之一。明确东亚小花蝽与大豆蚜之间的胁迫影响,有助于我们了解害虫对天敌的应答关系,探索东亚小花蝽作为生物农药实践应用价值,指导精准用药,为害虫综合治理提供科学依据。方法:在实验室不同胁迫条件下,测定了不同龄期大豆蚜体内总蛋白含量及乙酰胆碱酯酶、羧酸酯酶和谷胱甘肽-s-转移酶的比活力。结果:经过东亚小花蝽胁迫的1-4龄大豆蚜与没有受到胁迫的对照组相同龄期大豆蚜相比,体内乙酰胆碱酯酶和羧酸酯酶的比活力显著提升,且一龄时羧酸酯酶比活力达到最大值为0.0035u/mg prot。而受到胁迫的末龄大豆蚜与对照组相比,乙酰胆碱酯酶和羧酸酯酶的比活力并没有显著提升。受到胁迫的3-5龄大豆蚜,其谷胱甘肽-s-转移酶的比活力低于处理组,而1-2龄受胁迫的大豆蚜其体内谷胱甘肽-s-转移酶的比活力显著高于对照组,且二龄时谷胱甘肽-s-转移酶比活力达到最大值为0.1622nmol/min/mg prot。受到胁迫的1-4龄大豆蚜的总蛋白的含量显著低于对照组,而受到胁迫的末龄大豆蚜的总蛋白含量与对照组并无显著差异。结论:东亚小花蝽胁迫对低龄期大豆蚜体内各种生化物质含量影响明显,而对末龄大豆蚜的体内生化物质含量影响不大。
52.2材料与方法
53.1.1供试材料及仪器-54.1.1.1供试昆虫
55.东亚小花蝽orius sauteri采自黑龙江省向阳试验农场大豆田间,饲养在恒温培养箱(温度t=25
±
1℃、相对湿度rh=70%
±
5%、光照l∶d=16∶8)内,用大豆蚜饲喂,用黄豆芽作产卵植物繁殖东亚小花蝽。
56.大豆蚜aphis glycines为实验室长期保留的哈尔滨本地敏感种群,人工盆栽大豆苗饲养,置于温度25
±
1℃,相对湿度65%-70%,光周期的14l:10d的人工气候箱内,供试大豆品种为东农52。
57.1.1.2实验试剂
58.考马斯亮蓝g-250和牛血清蛋白(bsa)购自北京宝日医生物技术(takara)有限公司。磷酸缓冲液(pb)购自上海生工生物工程股份有限公司。乙酰胆碱酯酶(ache)、羧酸酯酶(care)、谷胱甘肽-s-转移酶(gsts)、海藻糖、还原糖均购自北京索莱宝(solarbio)科技有限公司。
59.1.1.3仪器设备
60.高速低温冷冻离心机(thermosorvallst16r)、人工气候箱(bic,上海博讯实业有限公司医疗设备厂)、多功能酶标仪(infinite-200pro,瑞士tecan公司)、烘箱(dph781,湖北省黄石市医疗器械厂)、冰箱(美菱)、电热恒温水浴锅(dzkw-s-4,北京永光明医疗仪器厂)、石英砂(分析纯,天津市巴斯夫化工有限公司)、无水乙醇(天津市天力化学试剂有限公司)
61.1.2东亚小花蝽对大豆蚜的胁迫处理
62.用东亚小花蝽成虫对各龄期的大豆蚜进行胁迫处理,处理方式分为三种:接触胁
迫、气味胁迫和视觉胁迫,对照组为无东亚小花蝽胁迫。每个自制培养皿上层放入一定数量的大豆蚜,下层放入东亚小花蝽成虫,培养皿中间用透明可见的塑料板和不可见的带有针孔的透气滤隔离。处理后置入温度24
±
1℃、相对湿度70%
±
5%、光照为16l:8d的培养箱内。24h后,取受胁迫的大豆蚜分别进行各种生化物质测定。
63.1.3酶液制备
64.参照高欢欢等(2016)的酶液提取法,24h后,分别于各个处理中挑取5个龄期的大豆蚜,每个龄期的大豆蚜收集5只存活的虫体置于1.5ml的eppendorf离心管中,加入400μl0.02mol/l的磷酸缓冲液(nacl 136.89mm;kcl 2.67mm;na2hpo4 8.1mm;kh2po4 1.76mm)和少量石英砂,冰浴充分研磨,并将研磨后的匀浆置于高速冷冻离心机8000g、4℃离心10min。取上清液分装到新的1.5ml的eppendorf离心管置于冰上待测。各个处理的5个虫龄均设置3次重复。
65.1.4蛋白质、还原糖和海藻糖含量的测定
66.1.4.1蛋白质含量的测定
67.参照bradford(1976)的考马斯亮蓝g-250染色法测定。分别配制考马斯亮蓝g-250溶液和牛血清蛋白标准溶液,制作标准曲线。每个样品取4μl酶液于酶标板的加样孔中,对照孔中加入4μl的磷酸缓冲液,再加入200μl复温的考马斯亮蓝g-250试剂,充分混合,室温下放置5min后,在酶标仪波长设定在595nm波长处进行测定,记录od值,重复3次,根据蛋白标准曲线,计算出各个待测样中的蛋白浓度。
68.1.4.2还原糖、海藻糖含量的测定
69.采用蒽酮硫酸法测定海藻糖的含量,分别配制蒽酮溶液和10mg/ml的海藻糖标准溶液,制作标准曲线。每个样品取60μl样本液和240μl配置好的蒽酮试剂于1.5ml的eppendorf离心管中混匀,95℃水浴10min,(盖紧,防止水分散失)自然冷却至室温,取200μl至酶标板中,在620nm波长下进行测定,记录od值,重复3次,根据海藻糖标准曲线,计算出各个待测样中的海藻糖含量。
70.采用dns法测定还原糖的含量,分别配置3,5-二硝基水杨酸(dns)碱性溶液和10mg/ml的葡萄糖标准溶液,制作标准曲线。测定管中,每个样品取175μl样本液和125μl配置好的dns试剂于1.5ml的eppendorf离心管中混均匀,沸水浴中加热5min(盖紧,防止水分散失),取出后立即冷却至室温,混匀。对照管中,每个样品取175μl样本液和125μl蒸馏水于1.5ml的eppendorf离心管中混均匀,沸水浴中加热5min(盖紧,防止水分散失),取出后立即冷却至室温,混匀。取200μl至96孔酶标板中,在540nm波长下进行测定,记录od值,重复3次。计算测定管减去对照管的吸光值差值,根据葡萄糖标准曲线,计算出各个待测样中的还原糖含量(仵均祥等,2004;丁慧梅,2011;程媛等,2016)。
71.1.5酶活性的测定
72.ache、care和gsts酶比活力的测定均按照北京索莱宝科技有限公司试剂盒中的说明进行检测。酶活性值根据试剂盒说明书中的公式进行计算。ache、care和gsts酶比活力测定波长分别为412、450和340nm。ache活性单位定义为每毫克组织蛋白每分钟催化产生1nmol摩尔消光系数(tnb)为一个酶活单位;care活性单位定义为37℃下每毫升反应体系每毫克组织蛋白每分钟催化吸光值增加1定义为一个酶活力单位;gst活性单位定义为在25℃或者37℃中,每毫克蛋白每分钟催化1μmol1-氯-2,4-二硝基苯(cdnb)与谷胱甘肽(gsh)结
合为一个酶活性单位。
73.1.6数据分析
74.所有试验数据均采用microsoft excel表格处理软件和spss base ver.19.0统计软件进行数据分析和作图,通过duncan's新复极差法检验分析处理间差异性。对受东亚小花蝽胁迫后大豆蚜体内蛋白质、海藻糖和还原糖含量以及ache、care和gsts活性分别采用单因素方差分析。
75.2结果
76.2.1东亚小花蝽胁迫对大豆蚜蛋白含量的影响
77.东亚小花蝽胁迫大豆蚜24h后,随着龄期的增长其酶液蛋白含量呈一直上升的趋势。由图1显示,和对照组相比,在成蚜阶段时,气味胁迫与视觉胁迫处理组体内酶液蛋白含量分别降低了23.42%和57.27%,且视觉胁迫处理组与其他组间差异极显著,而直接接触的胁迫与对照组间并无显著差异;在4龄时和对照组相比,气味胁迫和视觉胁迫处理组的蛋白含量分别降低了46.36%和61.85%,直接接触的胁迫则降低了44.33%;在3龄时,气味胁迫和视觉胁迫处理组分别降低了27.99%和30.15%,直接接触的胁迫则降低了31.54%,且三个处理组间无显著性差异;在2龄时,气味胁迫和视觉胁迫处理组的蛋白含量分别降低了15.82%和24.64%,直接接触的胁迫则降低了52.73%;在1龄时,气味胁迫、视觉胁迫和直接接触胁迫处理组分别降低了15.13%,33.32%和57.01%,且直接接触胁迫处理组中的蛋白含量达到最低值(9.12μg/ml),极显著低于对照组、气味胁迫和视觉胁迫处理组(f=167.694,df=3,p《0.001)。
78.2.2东亚小花蝽胁迫对大豆蚜海藻糖含量的影响
79.东亚小花蝽对大豆蚜胁迫24h后,随着龄期的增长其海藻糖含量表现为一直上升的趋势。由图2显示,和对照组相比,在成蚜阶段时,气味胁迫与视觉胁迫处理组体内海藻糖含量分别升高了97.77%和43.49%,且直接接触胁迫处理组达到最高值为(3.4607mg/mg prot),而三种胁迫方式与对照组间均有极显著差异;在4龄时,气味胁迫和视觉胁迫处理组的海藻糖含量分别升高了67.08%和32.56%,直接接触的胁迫则升高了104.12%;在3龄时,气味胁迫和视觉胁迫处理组分别升高了52.29%和22.22%,直接接触的胁迫则升高了115.33%;在2龄时,气味胁迫和视觉胁迫处理组的海藻糖含量分别升高了72.21%和30.22%,直接接触的胁迫则升高了145.48%;在1龄时,气味胁迫、视觉胁迫和直接接触胁迫处理组分别升高了57.24%,37.96%和160.72%,而气味胁迫和视觉胁迫处理组间并无极显著差异。(f=167.694,df=3,p《0.001)。
80.2.3东亚小花蝽胁迫对大豆蚜还原糖含量的影响
81.东亚小花蝽对大豆蚜胁迫24h后,随着龄期的增长其还原糖含量表现为一直降低的趋势。由图3显示,和对照组相比,在成蚜阶段时,气味胁迫、视觉胁迫和直接接触胁迫处理组与对照组的还原糖含量均无显著性差异;在4龄时,气味胁迫和视觉胁迫处理组的还原糖含量分别升高了27.29%和11.36%,直接接触的胁迫则升高了46.10%,且三个处理组的还原糖含量均显著高于对照组,而直接接触胁迫处理组极显著高于气味胁迫与视觉胁迫处理组;在3龄时,气味胁迫和视觉胁迫处理组分别升高了69.72%和55.27%,直接接触的胁迫则升高了120%,且气味胁迫与视觉胁迫处理组间并无显著性差异;在2龄时,气味胁迫和视觉胁迫处理组的还原糖含量分别升高了122.67%和114.88%,直接接触的胁迫则升高了
168.73%;在1龄时,气味胁迫、视觉胁迫和直接接触胁迫处理组分别升高了308.95%,193.53%和371.54%,且直接接触胁迫处理组中的还原糖含量达到最高值(2.89μg/mg prot),极显著高于对照组、气味胁迫和视觉胁迫处理组(f=167.694,df=3,p《0.001)。
82.2.4东亚小花蝽胁迫对大豆蚜乙酰胆碱酯酶比活力的影响
83.东亚小花蝽对大豆蚜胁迫24h后,随着龄期的增长其乙酰胆碱酯酶比活力表现为一直降低的趋势。由图4显示,和对照组相比,在成蚜阶段时,气味胁迫与视觉胁迫处理组体内乙酰胆碱酯酶比活力分别降低了68.34%和70.57%,且气味胁迫和视觉胁迫处理组间并无显著性差异极,而直接接触的胁迫与对照组间并无显著差异;在4龄时,气味胁迫和直接接触胁迫处理组的乙酰胆碱酯酶比活力分别升高了56.91%和104.57%,且气味胁迫和直接接触胁迫处理组均极显著高于对照组,而视觉胁迫处理组与对照组间并无显著性差异;在3龄时,气味胁迫和视觉胁迫处理组分别升高了115.19%和28.70%,直接接触的胁迫则升高了174.31%,且三个处理组间均有显著性差异并都极显著高于对照组;在2龄时,气味胁迫和视觉胁迫处理组的乙酰胆碱酯酶比活力分别升高了368.71%和136.04%,直接接触的胁迫则升高了918.05%;在1龄时,气味胁迫、视觉胁迫和直接接触胁迫处理组分别升高了4228.37%,1628.48%和8506.98%,且直接接触胁迫处理组中的乙酰胆碱酯酶比活力达到最高值(198.79u/mg prot),极显著高于对照组、气味胁迫和视觉胁迫处理组(f=167.694,df=3,p《0.001)。
84.2.5东亚小花蝽胁迫对大豆蚜羧酸酯酶比活力的影响
85.东亚小花蝽对大豆蚜胁迫24h后,随着龄期的增长其羧酸酯酶比活力表现为下降明显的趋势。由图5显示,和对照组相比,在成蚜阶段时,气味胁迫与视觉胁迫处理组体内羧酸酯酶比活力分别升高了174.30%和887.64%,且视觉胁迫与气味胁迫处理组均极显著高于对照组,而直接接触的胁迫与对照组间并无显著差异;在4龄时,气味胁迫和视觉胁迫处理组羧酸酯酶比活力分别升高了126.16%和586.81%,直接接触的胁迫则升高了124.52%,且三种胁迫方式均极显著高于对照组,而气味胁迫与直接接触胁迫处理组间并无显著性差异;在3龄时,气味胁迫和视觉胁迫处理组分别升高了55.48%和1508.95%,直接接触的胁迫则升高了173.87%,且三个处理组间均有极显著差异;在2龄时,气味胁迫和视觉胁迫处理组的羧酸酯酶比活力分别升高了1135.72%和875.35%,直接接触的胁迫则升高了581.85%;在1龄时,气味胁迫、视觉胁迫和直接接触胁迫处理组分别升高了1889.46%,357.15%和1713.34%,且气味胁迫处理组中的羧酸酯酶比活力达到最高值(0.0038u/mg prot),显著高于对照组、气味胁迫和视觉胁迫处理组(f=167.694,df=3,p《0.001)。
86.2.6东亚小花蝽胁迫对大豆蚜谷胱甘肽-s-转移酶比活力的影响
87.东亚小花蝽对大豆蚜胁迫24h后,随着龄期的增长其谷胱甘肽-s-转移酶比活力表现为下降明显的趋势。由图6显示,和对照组相比,在成蚜阶段时,气味胁迫与视觉胁迫处理组体内谷胱甘肽-s-转移酶比活力分别升高了97.62%和229.91%,直接接触的胁迫则降低了58.5%,且三个处理组与对照组间差异极显著,而直接接触胁迫处理组的谷胱甘肽-s-转移酶比活力极显著低于对照组;在4龄时,气味胁迫和视觉胁迫处理组的谷胱甘肽-s-转移酶比活力分别升高了81.78%和219.26%,直接接触的胁迫则降低了12.19%,且直接接触胁迫处理组与对照组间无显著性差异;在3龄时,气味胁迫和视觉胁迫处理组的分别升高了
20.55%和10%,直接接触的胁迫则降低了58.45%;在2龄时,气味胁迫和视觉胁迫处理组的谷胱甘肽-s-转移酶比活力分别升高了31.17%和7.39%,直接接触的胁迫则升高了156.59%,且直接接触胁迫和气味胁迫处理组均显著性高于对照组,直接接触胁迫处理组中的谷胱甘肽-s-转移酶比活力达到最高值(0.06nmol/min/mg prot),而视觉胁迫处理组与对照组间并无显著性差异;在1龄时,气味胁迫、视觉胁迫和直接接触胁迫处理组分别升高了10.23%,2.54%和104.42%,直接胁迫极显著高于对照组、气味胁迫和视觉胁迫处理组(f=167.694,df=3,p《0.001)。
88.以上显示和描述了本发明的基本原理和主要特征和本发明的优点。本行业的技术人员应了解,本发明不受上述实施例的限制,上述实施例和说明书中描述的只是说明本发明的原理,在不脱离本发明精神和范围的前提下,本发明还会有各种变化和改进,这些变化和改进都落入要求保护的本发明范围内,本发明要求保护范围由所附的权利要求书其等效物界定。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1