生物标志物导向的多靶点免疫治疗的制作方法
本发明提供了用于评估疾病中生物标志物表达并提供针对所述疾病的多靶点基于李斯特菌的免疫疗法的方法和组合物。
背景技术:
生物标志物是一种可测量的特征,其反映某种疾病状态的严重性或存在性或者与该疾病状态相关,并可用作生物体的特定疾病状态或一些其他生理学状态的指征。生物标志物可为特异的细胞、分子、基因、基因产物、酶、受体、任何这些细胞元件的突变形式或者激素,它们可用于鉴定和/或衡量疾病状态的存在或进展,例如特定的癌症或肿瘤。此外,熟知的是,肿瘤和癌症可表达成套的肿瘤生物标志物,可将它们用于鉴定肿瘤或癌症的存在或者衡量它们的进展或治疗效果。
尽管将生物标志物大量用于诊断疾病和监测疾病进展,但是仍亟需开发利用这种信息的治疗手段来特异靶向由该疾病表达的靶生物标志物,这些生物标志物与增生或疾病状态的存在以及随后受试者整体健康的恶化直接相关。
单核细胞增多性李斯特菌(Listeria monocytogenes)(Lm)为细胞内病原体,它们主要感染抗原呈递细胞,已适应于生活在这些细胞的细胞质中。单核细胞增多性李斯特菌及其产生的称为李斯特菌溶血素O(LLO)的蛋白具有强佐剂特性,与大多数用于基于细胞的免疫治疗的佐剂不同,其可在提供抗原特异性处理后给药,或者,当目的抗原与该李斯特菌表达的佐剂蛋白如LLO或ActA蛋白融合时,其自身可用于提供抗原特异性处理。
本发明通过提供组合的、多靶点免疫疗法来满足该需求,其中将每个包含表达不同的疾病相关抗原的重组李斯特菌菌株的各个组合物(而不是存在于分开组合物中的对等李斯特菌)分开地向具有疾病的受试者给药,或者将这些组合物作为单次剂量给药而联合用药。本发明还通过采用表达至少一种融合蛋白的李斯特菌来提供预定数量的疾病相关抗原或其片段来满足该需求,该融合蛋白包含融合至免疫原性李斯特菌肽如N末端LLO、截短的LLO、ActA蛋白片段或PEST肽的该抗原。这些组合物的使用将使得具有表达不止一种生物标志物的患病细胞亚群的疾病(包括肿瘤、癌症或其他疾病)被成功治疗。
技术实现要素:
在一个方面,本发明涉及在具有疾病的受试者中诱导针对所述疾病的免疫应答的方法,该方法包括以下步骤:a.从所述受试者获得生物样品;b.评估所述生物样品中预定数量的生物标志物的表达;c.以包含重组李斯特菌菌株的组合物向所述受试者给药,所述菌株包含编码至少一种融合蛋白的核酸序列,其中所述融合蛋白包含在所述生物样品中鉴定的生物标志物,其中所述生物标志物与所述疾病相关,其中所述生物标志物融合至含PEST的多肽,并且其中所述李斯特菌内的每种融合蛋白包含不同的生物标志物,因而在所述受试者中诱导多靶点抗疾病免疫应答。
在一个方面,本发明涉及在具有疾病的受试者中诱导针对所述疾病的免疫应答的方法,该方法包括以下步骤:a.从所述受试者获得生物样品;b.评估所述生物样品中预定数量的生物标志物的表达;c.以每个包含重组李斯特菌菌株的组合物的混合物向所述受试者给药,所述菌株包含编码融合蛋白的核酸序列,其中所述融合蛋白包含在所述生物样品中鉴定的生物标志物,其中所述生物标志物与所述疾病相关,其中所述生物标志物融合至含PEST的多肽,并且其中每个组合物内的每种李斯特菌菌株在所述融合蛋白中包含不同的生物标志物,因而在所述受试者中诱导多靶点抗疾病免疫应答。
在一相关方面,本发明涉及治疗受试者中疾病的方法,该方法包括以下步骤:a.从所述受试者获得生物样品;b.评估所述生物样品中预定数量的生物标志物的表达;c.以包含重组李斯特菌菌株的组合物向所述受试者给药,所述菌株包含编码至少一种融合蛋白的核酸序列,其中所述融合蛋白包含在所述生物样品中鉴定的生物标志物,其中所述生物标志物与所述疾病相关,其中所述生物标志物融合至含PEST的多肽,并且其中所述李斯特菌内的每种融合蛋白包含不同的生物标志物,因而治疗所述受试者中的所述疾病。
在一相关方面,本发明涉及治疗受试者中疾病的方法,该方法包括以下步骤:a.从所述受试者获得生物样品;b.评估所述生物样品中预定数量的生物标志物的表达;c.以每个包含重组李斯特菌菌株的组合物的混合物向所述受试者给药,所述菌株包含编码融合蛋白的核酸序列,其中所述融合蛋白包含在所述生物样品中鉴定的生物标志物,其中所述生物标志物与所述疾病相关,其中所述生物标志物融合至含PEST的多肽,并且其中每个组合物内的每种李斯特菌菌株包含不同的生物标志物,因而治疗所述受试者中的所述疾病。
在另一方面,本发明涉及预防受试者中疾病复发的方法,该方法包括以下步骤:a.从所述受试者获得生物样品;b.评估所述生物样品中预定数量的生物标志物的表达;c.以包含重组李斯特菌菌株的组合物向所述受试者给药,所述菌株包含编码至少一种融合蛋白的核酸序列,其中所述融合蛋白包含在所述生物样品中鉴定的生物标志物,其中所述生物标志物与所述疾病相关,其中所述生物标志物融合至含PEST的多肽,并且其中所述李斯特菌内的每种融合蛋白包含不同的生物标志物,因而在所述受试者中预防所述疾病的复发。
在另一方面,本发明涉及防止受试者中疾病复发的方法,该方法包括以下步骤:a.从所述受试者获得生物样品;b.评估所述生物样品中预定数量的生物标志物的表达;c.以每个包含重组李斯特菌菌株的组合物的混合物向所述受试者给药,所述菌株包含编码融合蛋白的核酸序列,其中所述融合蛋白包含在所述生物样品中鉴定的生物标志物,其中所述生物标志物与所述疾病相关,其中所述生物标志物融合至含PEST的多肽,并且其中每个组合物内的每种李斯特菌菌株包含不同的生物标志物,因而在所述受试者中防止所述疾病的复发。
本发明的其他特征和优点将由于以下详细描述的实例和附图而变得显而易见。但是,应当理解,该详细描述和具体实例尽管指出了本发明的优选实施例,但是仅仅通过例证给出,因为本发明的精神和范围内的各种变化和修改对阅读了该详细描述的本领域技术人员而言将是显而易见的。
附图说明
以下附图形成了本说明书的一部分,将它们包括进来以进一步说明本公开的某些方面,通过参考一个或多个这些附图并结合本文提供的对具体实施例的描述可更好地理解本发明。本专利或申请文件包含至少一个用彩色表现的附图。根据请求并支付必要的费用后,专利局将提供带有彩色附图的本专利或专利申请公开的副本。
图1显示Lm-E7和Lm-LLO-E7使用不同的表达系统来表达和分泌E7。通过将基因盒引入单核细胞增多性李斯特菌基因组的orfZ结构域中来产生Lm-E7(A)。hly启动子驱动hly信号序列和LLO的前五个氨基酸(AA)以及随后的HPV-16E7的表达。B)通过以质粒pGG-55转化prfA-菌株XFL-7来产生Lm-LLO-E7。pGG-55具有驱动LLO-E7非溶血性融合体的表达的hly启动子。pGG-55也含有prfA基因,以选择XFL-7在体内对质粒的保留。
图2显示Lm-E7和Lm-LLO-E7分泌E7。使Lm-Gag(泳道1)、Lm-E7(泳道2)、Lm-LLO-NP(泳道3)、Lm-LLO-E7(泳道4)、XFL-7(泳道5)和10403S(泳道6)在Luria-Bertoni液体培养基中于37℃生长过夜。沉淀相等数目的细菌(通过600nm处吸光度OD确定),并将18ml的每种上清液进行TCA沉淀。通过Western印迹分析E7表达。用抗E7mAb、随后用HRP-偶联的抗小鼠(Amersham)探测该印迹,接着使用ECL检测试剂显影。
图3显示LLO-E7融合体的肿瘤免疫治疗功效。显示了肿瘤接种后第7、第14、第21、第28和第56天小鼠中以毫米计的肿瘤大小。未暴露的小鼠:空心圆圈;Lm-LLO-E7:实心圆圈;Lm-E7:正方形;Lm-Gag:空心菱形;和Lm-LLO-NP:实心三角形。
图4显示来自Lm-LLO-E7-免疫的小鼠的脾细胞在暴露于TC-1细胞时增殖。将C57BL/6小鼠用Lm-LLO-E7、Lm-E7或对照rLm菌株免疫和强化。在强化后6天收获脾细胞,并以显示的比率与经照射的TC-1细胞一起铺板。将细胞用3H胸苷脉冲处理并收获。将Cpm定义为(实验cpm)-(无TC-1对照)。
图5A显示,(A)Western印迹证实Lm-ActA-E7分泌E7。泳道1:Lm-LLO-E7;泳道2:Lm-ActA-E7.001;泳道3;Lm-ActA-E7-2.5.3;泳道4:Lm-ActA-E7-2.5.4。
图5B显示Lm-ActA-E7(矩形)、Lm-E7(椭圆形)、Lm-LLO-E7(X)和未暴露的小鼠(未接种疫苗;实心三角形)给药的小鼠中的肿瘤大小。
图6A显示用于制备4种LM疫苗的质粒插入片段的示意图。Lm-LLO-E7插入片段含有所有使用的李斯特菌基因。它含有hly启动子、hly基因(其编码蛋白LLO)的前1.3kb和HPV-16E7基因。hly的该前1.3kb包括信号序列(ss)和PEST区域。Lm-PEST-E7包括hly启动子、信号序列以及PEST和E7序列,但是不包括截短的LLO基因的剩余部分。Lm-ΔPEST-E7不包括PEST区域,但是含有hly启动子、信号序列、E7和截短的LLO的剩余部分。Lm-E7epi仅具有hly启动子、信号序列和E7。
图6B上部插图:含有PEST区域的李斯特菌构建体诱导肿瘤消退。下部插图:在2个单独实验中肿瘤挑战后第28天时的平均肿瘤大小。
图6C显示含有PEST区域的李斯特菌构建体在脾中诱导更高百分比的E7特异性淋巴细胞。显示了来自3个实验的数据的平均值和SE。
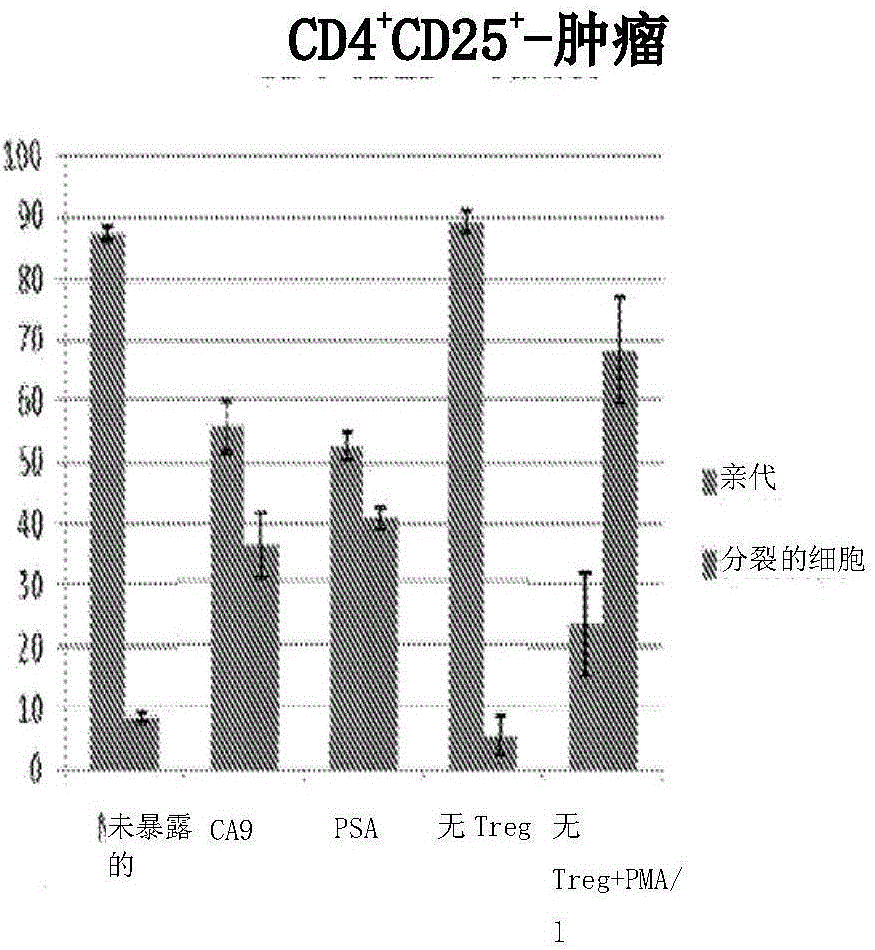
图7A显示,在施用TC-1肿瘤细胞并随后以Lm-E7、Lm-LLO-E7、Lm-ActA-E7给药的或无疫苗(未暴露的)的小鼠中,脾中E7特异性的分泌IFN-γ的CD8+T细胞的诱导和浸润肿瘤的数目。
图7B显示,针对(A)描述的小鼠的脾和肿瘤中E7特异性的CD8+细胞的诱导和浸润。
图8显示,含有PEST区域的李斯特菌构建体在肿瘤中诱导更高百分比的E7特异性淋巴细胞。(A)来自1个实验的代表性数据。(B)来自所有3个实验的数据的平均值和SE。
图9A显示了大肠杆菌(E.coli)-李斯特菌穿梭质粒pGG55的示意图谱。CAT(-):大肠杆菌氯霉素转移酶;CAT(+):李斯特菌氯霉素转移酶;Ori Lm:李斯特菌的复制起点;Ori Ec:大肠杆菌的p15复制起点;prfA:李斯特菌致病性调节因子A;LLO:C端截短的李斯特菌溶血素O,包括其启动子;E7:HPV E7。还描绘了所选的限制位点。
图9B显示了大肠杆菌-李斯特菌穿梭质粒pTV3的示意图谱(下图)。CAT(-):大肠杆菌氯霉素转移酶;CAT(+):李斯特菌氯霉素转移酶;Ori Lm:李斯特菌的复制起点;Ori Ec:大肠杆菌的p15复制起点;prfA:李斯特菌致病性调节因子A;LLO:C端截短的李斯特菌溶血素O,包括其启动子;E7:HPV E7;p60-dal:p60启动子和李斯特菌dal基因的表达盒。还描绘了所选的限制位点。
图10显示了存在于李斯特菌菌株EGD的基因组上的inlC区域的上游和下游的DNA序列(SEQ ID NO:81)。DNA-上游(红色)、inlC基因(蓝色)和DNA-下游(黑色)。
图11显示了克隆进温度敏感的质粒pKSV7中以产生inl C缺失突变体的DNA序列(SEQ ID NO:82)。用于克隆这些区域的限制性酶位点用大写字母表示且标有下划线。GAATTC-EcoRI、GGATCC-BamHI和CTGCAg-PstI。EcoRI-PstI插入片段克隆进该载体pKSV7中。
图12显示了Lm-dd和Lm-ddD actA菌株的示意图。凝胶显示了使用寡聚物的1/2和寡聚物的3/4将菌株Lm-dd和Lm-ddΔactA的e染色体DNA作为模板得到的PCR产物的大小。
图13显示了存在于李斯特菌染色体中的actA基因的上游和下游的DNA序列(SEQ ID NO:60)。以斜体字显示的区域含有存在于LmddΔactA菌株中的残余actA序列元件。标有下划线的序列gtcgac代表XhoI的限制位点,其为actA的N-T和C-T区域之间的连接处。
图14显示了响应于LM疫苗菌株(A)的给药而引起的肿瘤消退。圆圈代表未暴露的小鼠,倒三角形代表Lmdd-TV3给药的小鼠,十字叉代表Lm-LLOE7给药的小鼠。
图15显示了(A)pAdv164的质粒图谱,其具有枯草芽孢杆菌(bacillus subtilis)dal基因,该基因受组成型李斯特菌p60启动子的控制,以用于补充LmddA菌株中的染色体dal-dat缺失。它还含有截短的LLO(1-441)与嵌合人Her2/neu基因的融合体,该融合体通过直接融合3个片段Her2/neu:EC1(氨基酸40-170)、EC2(氨基酸359-518)和ICI(氨基酸679-808)而构建。(B)通过对用抗LLO抗体印迹的TCA沉淀的细胞培养物上清液进行Western印迹分析,在Lm-LLO-ChHer2(Lm-LLO-138)和LmddA-LLO-ChHer2(ADXS31-164)中检测tLLO-ChHer2的表达和分泌。~104KD的差异带对应tLLO-ChHer2。内源LLO作为58KD的带被检测到。李斯特菌对照缺乏ChHer2表达。
图16(A)将NT-2细胞用作刺激物,将3T3/neu细胞用作靶标,测试Her2/neu基于李斯特菌的疫苗在来自经免疫小鼠的脾细胞中引起的细胞毒性T细胞应答。Lm-对照是基于LmddA背景,除了表达不相关抗原(HPV16-E7)外,在所有方面都相同。(B)在体外用丝裂霉素C处理的NT-2细胞刺激24小时以后,通过ELISA测定来自经免疫的FVB/N小鼠的脾细胞分泌进细胞培养基中的IFN-γ。(C)响应于与来自蛋白的不同区域的肽一起在体外温育,来自用嵌合疫苗免疫的HLA-A2转基因小鼠的脾细胞的IFN-γ分泌。如图例中列出,将重组ChHer2蛋白用作阳性对照,不相关肽或没有肽组构成阴性对照。使用在72小时共温育之后收获的细胞培养物上清液,通过ELISA测定检测IFN-γ的分泌。每个数据点是一式三份数据的平均值±标准误差。*P值<0.001。
图17代表了来自Her2/neu转基因小鼠的结果,该Her2/neu转基因小鼠经每种重组李斯特菌-ChHer2或对照李斯特菌疫苗注射六次。在6周龄开始免疫,并且继续每三周一次直到第21周。每周监测肿瘤的出现,并且将其表示为无肿瘤小鼠的百分比。*p<0.05,每组N=9。
图18显示了以1×106个NT-2细胞皮下接种FVB/N小鼠,并且用每种疫苗以一周的间隔免疫三次。在第二次免疫接种之后7天收获脾。在分离免疫细胞之后,将它们染色用于通过抗CD3、CD4、CD25和FoxP3抗体检测Treg。来自代表性实验的Treg的点图显示了CD25+/FoxP3+T细胞的频率,将其表示为跨不同处理组的总CD3+或CD3+CD4+T细胞的百分比。
图19显示了以1×106个NT-2细胞皮下接种FVB/N小鼠,并且用每种疫苗以一周的间隔免疫三次。在第二次免疫接种之后7天收获肿瘤。在分离免疫细胞之后,将它们染色用于通过抗CD3、CD4、CD25和FoxP3抗体检测Treg。(A).来自代表性实验的Treg的点图。(B).CD25+/FoxP3+T细胞的频率,表示为跨不同处理组的总CD3+或CD3+CD4+T细胞(左插图)的百分比和肿瘤内CD8/Treg比率(右插图)。数据显示为得自2个独立实验的均值±SEM。
图20显示了pAdv134质粒和双重质粒的示意图。指出了将用于克隆抗原1(Xho I和SpeI)和抗原2(XbaI和SacI或BglII)基因的限制位点。黑箭头代表转录的方向。p15ori和RepR是指李斯特菌和大肠杆菌复制起点。tLLO是截短的李斯特菌溶血素O蛋白(1-441氨基酸),tActA是截短的ActA(1-233氨基酸)蛋白。芽孢杆菌-dal基因编码D-丙氨酸消旋酶,该酶补充LmΔdal dat菌株中的D-丙氨酸的合成。
图21显示了肿瘤中MDSC和Treg的减少。在Lm接种(LmddAPSA和LmddAE7)后右侧插图的MDSC数量(B)和左侧插图的Treg数量(A)。
图22A-22D显示,抑制测定数据证实来自TPSA23肿瘤(表达PSA的肿瘤)的单核细胞MDSC在李斯特菌接种后抑制性较差。MDSC的抑制能力的这种变化不是抗原特异性,因为用PSA-抗原特异性T细胞以及用非特异性刺激的T细胞观察到相同的抑制降低。在图22A和22B中,佛波醇-肉豆蔻酸酯-乙酸酯和离子霉素(PMA/I)代表非特异性刺激。在图22C和22D中,术语“肽”代表特异性抗原刺激。百分比(%)CD3+CD22+代表%效应(应答)T细胞。无MDSC组显示当它们不受刺激时缺乏应答T细胞的分裂,最后一组(添加了PMA/I或肽)显示MDSC不存在时受刺激的细胞的分裂。图22A和22C显示了每个组的单个细胞分裂周期。图22B和22D显示了合并的分裂周期。
图23A-23D显示了抑制测定数据,证实李斯特菌对脾单核细胞MDSC无影响,且它们仅以抗原特异性方式产生抑制。在图23A和23B中,PMA/I代表非特异性刺激。在图23C和23D中,术语“肽”代表特异性抗原刺激。百分比(%)CD3+CD8+代表%效应(应答)T细胞。无MDSC组显示当它们不受刺激时缺乏应答T细胞的分裂,最后一组(添加了PMA/I或肽)显示MDSC不存在时受刺激的细胞的分裂。图23A和23C显示了每个组的单个细胞分裂周期。图23B和23D显示了合并的分裂周期。
图24A-24D显示了抑制测定数据,证实来自肿瘤的粒细胞MDSC在李斯特菌接种后减弱了抑制T细胞的能力。MDSC的抑制能力的这种变化不是抗原特异性,因为用PSA-抗原特异性T细胞以及用非特异性刺激的T细胞观察到相同的抑制降低。在图24A和24B中,PMA/I代表非特异性刺激。在图24C和24D中,术语“肽”代表特异性抗原刺激。百分比(%)CD3+CD8+代表%效应(应答)T细胞。无MDSC组显示当它们不受刺激时缺乏应答T细胞的分裂,最后一组(添加了PMA/I或肽)显示MDSC不存在时受刺激的细胞的分裂。图24A和24C显示了每个组的单个细胞分裂周期。图24B和24D显示了合并的分裂百分比。
图25A-25D显示了抑制测定数据,证实李斯特菌对脾粒细胞MDSC无影响,且它们仅以抗原特异性方式产生抑制。在图25A和25B中,PMA/I代表非特异性刺激。在图25C和25D中,术语“肽”代表特异性抗原刺激。百分比(%)CD3+CD8+代表%效应(应答)T细胞。无MDSC组显示当它们不受刺激时缺乏应答T细胞的分裂,最后一组(添加了PMA/I或肽)显示MDSC不存在时受刺激的细胞的分裂。图25A和25C显示了每个组的单个细胞分裂周期。图25B和25D显示了合并的分裂百分比。
图26A-26D显示了抑制测定数据,证实来自肿瘤的Treg仍是抑制性的。在该肿瘤模型中,Treg的抑制能力以非抗原特异性方式轻微下降。在图26A和26B中,PMA/I代表非特异性刺激。在图26C和26D中,术语“肽”代表特异性抗原刺激。百分比(%)CD3+CD8+代表%效应(应答)T细胞。无Treg组显示当它们不受刺激时缺乏应答T细胞的分裂,最后一组(添加了PMA/I或肽)显示Treg不存在时受刺激的细胞的分裂。图26A和26C显示了每个组的单个细胞分裂周期。图26B和26D显示了合并的分裂百分比。
图27A-27D显示了抑制测定数据,证实脾Treg仍是抑制性的。在图27A和27B中,PMA/I代表非特异性刺激。在图27C和27D中,术语“肽”代表特异性抗原刺激。百分比(%)CD3+CD8+代表%效应(应答)T细胞。无Treg组显示当它们不受刺激时缺乏应答T细胞的分裂,最后一组(添加了PMA/I或肽)显示Treg不存在时受刺激的细胞的分裂。图27A和27C显示了每个组的单个细胞分裂周期。图27B和27D显示了合并的分裂百分比。
图28A-28D显示了抑制测定数据,证实常规的CD4+T细胞对细胞分裂无影响,无论它们是在小鼠的肿瘤还是脾中发现的。在图28A和28B中,PMA/I代表非特异性刺激。在图28C和28D中,术语“肽”代表特异性抗原刺激。百分比(%)CD3+CD8+代表%效应(应答)T细胞。无Treg组显示当它们不受刺激时缺乏应答T细胞的分裂,最后一组(添加了PMA/I或肽)显示Treg不存在时受刺激的细胞的分裂。图28C-28D显示来自合并的百分比分裂的数据。
图29A-29D显示了抑制测定数据,证实来自4T1肿瘤(表达Her2的肿瘤)的单核细胞MDSC在李斯特菌接种后具有减弱的抑制能力。MDSC的抑制能力的这种变化不是抗原特异性,因为用Her2/neu抗原特异性T细胞以及用非特异性刺激的T细胞观察到相同的抑制降低。在图29A和29B中,PMA/I代表非特异性刺激。在图29C和29D中,术语“肽”代表特异性抗原刺激。百分比(%)CD8+代表%效应(应答)T细胞。无MDSC组显示当它们不受刺激时缺乏应答T细胞的分裂,最后一组(添加了PMA/I或肽)显示MDSC不存在时受刺激的细胞的分裂。图29A和29C显示了每个组的单个细胞分裂周期。图29B和29D显示了合并的分裂百分比。
图30A-30D显示了抑制测定数据,证实对脾单核细胞MDSC无李斯特菌特异性影响。在图30A和30B中,PMA/I代表非特异性刺激。在图30C和30D中,术语“肽”代表特异性抗原刺激。百分比(%)CD8+代表%效应(应答)T细胞。无MDSC组显示当它们不受刺激时缺乏应答T细胞的分裂,最后一组(添加了PMA/I或肽)显示MDSC不存在时受刺激的细胞的分裂。图30A和30C显示了每个组的单个细胞分裂周期。图30B和30D显示了合并的分裂百分比。
图31A-31D显示了抑制测定数据,证实来自4T1肿瘤(表达Her2的肿瘤)的粒细胞MDSC在李斯特菌接种后具有减弱的抑制能力。MDSC的抑制能力的这种变化不是抗原特异性,因为用Her2/neu抗原特异性T细胞以及用非特异性刺激的T细胞观察到相同的抑制降低。在图31A和31B中,PMA/I代表非特异性刺激。在图31C和31D中,术语“肽”代表特异性抗原刺激。百分比(%)CD8+代表%效应(应答)T细胞。无MDSC组显示当它们不受刺激时缺乏应答T细胞的分裂,最后一组(添加了PMA/I或肽)显示MDSC不存在时受刺激的细胞的分裂。图31A和31C显示了每个组的单个细胞分裂周期。图31B和31D显示了合并的分裂百分比。
图32A-32D显示了抑制测定数据,证实对脾粒细胞MDSC无李斯特菌特异性影响。在图32A和32B中,PMA/I代表非特异性刺激。在图32C和32D中,术语“肽”代表特异性抗原刺激。百分比(%)CD8+代表%效应(应答)T细胞。无MDSC组显示当它们不受刺激时缺乏应答T细胞的分裂,最后一组(添加了PMA/I或肽)显示MDSC不存在时受刺激的细胞的分裂。图32A和32C显示了每个组的单个细胞分裂周期。图32B和32D显示了合并的分裂百分比。
图33A-33D显示了抑制测定数据,证实来自4T1肿瘤(表达Her2的肿瘤)的Treg在李斯特菌接种后抑制能力减弱。在图33A和33B中,PMA/I代表非特异性刺激。在图33C和33D中,术语“肽”代表特异性抗原刺激。百分比(%)CD8+代表%效应(应答)T细胞。这种减弱为非抗原特异性的,因为用Her2/neu特异性和非特异性应答T细胞都看到Treg抑制能力的变化。图33A和33C显示了每个组的单个细胞分裂周期。图33B和33D显示了合并的分裂百分比。
图34A-34D显示了抑制测定数据,证实对脾Treg无李斯特菌特异性影响。应答T细胞都能够分裂,无论它们是否是抗原特异性的。在图34A和34B中,PMA/I代表非特异性刺激。在图34C和34D中,术语“肽”代表特异性抗原刺激。百分比(%)CD8+代表%效应(应答)T细胞。图34A和34C显示了每个组的单个细胞分裂周期。图34B和34D显示了合并的分裂百分比。
图35A-35D显示了抑制测定数据,证实粒细胞MDSC的抑制能力是由于tLLO的过表达,且独立于伴侣融合抗原。左侧插图(图35A和35C)显示了每个组的单个细胞分裂周期。右侧插图(图35B和35D)显示了合并的分裂百分比。
图36A-36D显示了抑制测定数据,也证实单核细胞MDSC的抑制能力是由于tLLO的过表达,且独立于伴侣融合抗原。左侧插图(图36A和36C)显示了每个组的单个细胞分裂周期。右侧插图(图36B和36D)显示了合并的分裂百分比。
图37A-37D显示了抑制测定数据,证实在Lm接种后,从脾纯化的粒细胞MDSC保留了它们抑制抗原特异性应答T细胞分裂的能力(图37A和37B)。但是,在非特异性刺激以后,(以PMA/离子霉素)活化的T细胞仍然能够分裂(图37C和37D)。左侧插图显示了每个组的单个细胞分裂周期。右侧插图显示了合并的分裂百分比。
图38A-38D显示了抑制测定数据,证实在Lm接种后,从脾纯化的单核细胞MDSC保留了它们抑制抗原特异性应答T细胞分裂的能力(图38A和38B)。但是,在非特异性活化(以PMA/离子霉素刺激)以后,T细胞仍然能够分裂(图38C和38D)。左侧插图显示了每个组的单个细胞分裂周期。右侧插图显示了合并的分裂百分比。
图39A-39D显示了抑制测定数据,证实不论应答细胞是抗原特异性(图39A和39B)还是非特异性(图39C和39D)活化的,从任意Lm处理组的肿瘤纯化的Treg具有稍微减弱的抑制应答T细胞分裂的能力。左侧插图显示了每个组的单个细胞分裂周期。右侧插图显示了合并的分裂百分比。
图40A-40D显示了抑制测定数据,证实从脾纯化的Treg仍能够抑制抗原特异性(图40A-40B)和非特异性(图40C和40D)活化的应答T细胞的分裂。
图41A-41D显示了抑制测定数据,证实肿瘤Tcon细胞不能抑制T细胞的分裂,无论该应答细胞是抗原特异性(图41A和41B)还是非特异性活化的(图41C和41D)。
图42A-42D显示了抑制测定数据,证实脾Tcon细胞不能抑制T细胞的分裂,无论该应答细胞是抗原特异性(图42B和42B)还是非特异性活化的(图42C和42D)。
图43.ISG15在小鼠乳腺肿瘤中升高的表达。(A)从来自FVB/N HER2/neu转基因小鼠的原部位(autochthonous)小鼠乳腺肿瘤(n=9)和来自FVB/N小鼠的正常乳腺组织(n=4)提取mRNA.在cDNA转化后,进行qPCR分析来确定相对ISG15mRNA表达。(B)用抗ISG15抗体对来自正常乳腺组织和HER2/neu乳腺肿瘤组织的组织裂解物进行Western印迹分析(上部插图),用抗GAPDH抗体证实相等的蛋白上样(下部插图)。(C)就SG15mRNA的表达,将来自乳腺肿瘤细胞系NT2和4T1-Luc的cDNA的qPCR与正常乳腺组织和非转化细胞系NIH-3T3进行比较(n=3)。(D)ISG15在一组正常组织(n=3)中的表达的qPCR分析与来自HER2/neu转基因小鼠的原部位乳腺肿瘤(n=7)比较。
图44.针对ISG15的基于李斯特菌的CTL疫苗的构建。(A)图解描述李斯特菌表达载体pGG34-LLO-ISG15,其被电穿孔进prfA-XFL7李斯特菌菌株,以构建减毒李斯特菌疫苗Lm-LLO-ISG15。(B)来自Lm-LLO-ISG15培养基和对照Lm疫苗Lm-LLO-OVA培养物的TCA沉淀蛋白的Western印迹分析。用针对小鼠ISG15(上部插图)、鸡卵清蛋白(中间插图)以及李斯特菌溶血素O(下部插图)的抗体使沉淀的蛋白接受SDS-PAGE和Western印迹分析。(C)来自8周龄IBalb/c小鼠的脾细胞的SG15特异性IFNγ应答的ELISpot分析,这些小鼠经Lm-LLO-ISG15或对照Lm腹膜内接种两次。结果显示为分泌IFNγ的SFC/2×106个脾细胞。(D)经对照Lm疫苗(2×108个CFU)或Lm-LLO-ISG15(2×108个CFU)接种的雌性小鼠每窝的幼崽数量。(E)来自出生后第一天每个雌性接种组的同窝平均幼崽重量,以克描述。
图45.Lm-LLO-ISG15接种后对小鼠乳腺肿瘤的治疗影响。(A)肿瘤负荷研究以确定Lm-LLO-ISG15对植入的NT2乳腺肿瘤的有效性。将NT2肿瘤细胞皮下植入FVB/N小鼠的后胁腹中,随后以Lm-LLO-ISG15或对照Lm接种。用卡尺监测肿瘤大小直至实验结束并计算肿瘤体积。(B)肿瘤负荷研究以确定Lm-LLO-ISG15接种控制植入的原发4T1-Luc乳腺肿瘤的生长的能力。将4T1-Luc肿瘤细胞植入Balb/c小鼠的乳腺组织中,随后以Lm-LLO-ISG15或对照Lm接种小鼠。C)转移性肿瘤研究以确定Lm-LLO-ISG15接种控制4T1-Luc在植入乳腺后转移性扩散的能力。简而言之,将4T1-Luc细胞植入Balb/c小鼠的乳腺组织中,随后以Lm-LLO-ISG15或对照Lm接种小鼠。在植入32天后,从接种的荷瘤小鼠取出肺并用PBS灌注。然后用光学显微镜对肺表面转移性结节进行计数。
图46.通过Lm-LLO-ISG15延迟了HER2/neu+原部位乳腺肿瘤的进展和表位扩展。(A)将FVB/N Her2/neu转基因小鼠模型用于确定,与对照Lm接种相比,Lm-LLO-ISG15接种是否可延迟原部位乳腺肿瘤进展。以Lm-LLO-ISG15(2×108个CFU)或对照Lm疫苗Lm-LLO-OVA(2×108个CFU)注射FVB/N HER2/neu转基因小鼠六次,其中从6周龄开始,每3周继续,直至第21周。每周一次监测肿瘤发生率。(B)来自未暴露的小鼠的自发乳腺肿瘤中ISG15特异性IFN-γ应答的ELISpot分析。在让肿瘤形成后,用对照Lm和Lm-LLO-ISG15接种荷瘤小鼠两次(第0和第7天),随后在第14天取出肿瘤,进行ELISpot分析。(C)ELISpot分析证实,在实验结束时,经Lm-LLO-ISG15接种的带有NT2肿瘤的FVB/N HER2/neu转基因小鼠脾细胞中表位扩展至HER2/neu。(D)TIL四聚体分析证实,与对照Lm接种小鼠相比,经Lm-LLO-ISG15接种的带有4T1-Luc肿瘤的小鼠的肿瘤中增加的HER2/neu特异性CD8+62L-百分比。
图47.ISG15接种的治疗影响是CD8依赖性的。(A)带有4T1-Luc肿瘤的小鼠的CD8耗尽实验。简而言之,将4T1-Luc肿瘤细胞植入Balb/c小鼠中,除了接种Lm-LLO-ISG15或对照Lm外,还耗尽CD8+细胞或不进行耗尽。(B)进行Winn测定以测量Lm-LLO-ISG15富含CD8的脾细胞的直接细胞裂解活性。将来自Lm-LLO-ISG15或对照Lm接种的小鼠的耗尽CD4的脾细胞与4T1-Luc细胞混合,并植入未暴露的Balb/c小鼠中。(C)曲线图描绘来自图47B中所示的实验的Balb/c小鼠的无肿瘤存活百分比。
图48.ISG15特异性CTL克隆的体内扩增引起抗肿瘤应答。在雌性Balb/c小鼠的乳腺组织中植入4T1-Luc肿瘤细胞后,接着以PBS或CpG连同对照或ISG15表位肽接种小鼠。(A)在整个实验过程中测量每个组的肿瘤体积。(B)在实验结束时,取出原发肿瘤并计算每个接种组的平均肿瘤质量。(C)另外,在实验结束时也取出每个接种组小鼠的肺,用于检查表面转移。为每个接种组计算平均肺表面转移的数量。(D)ELISpot分析来自PBS和pISG15d1/CPG接种小鼠的肿瘤浸润淋巴细胞(TIL)的ISG15d1特异性IFN-γ应答。(E)ELISpot分析来自PBS和pISG15d2/CPG接种小鼠的TIL的ISG15d2特异性IFN-γ应答。
图49A显示表达Flk-1/VEGFR2的基于Lm的构建体的设计。将每个基因片段克隆进表达载体pGG34,该载体与LLO融合并置于hly启动子控制下。
图49B显示表达Flk-1/VEGFR2的基于Lm的构建体的设计。来自培养上清液的Western印迹显示从所列的构建体表达每种融合蛋白。将多克隆兔抗PEST抗体用于融合蛋白检测(下部),将鼠抗LLO抗体用于验证(上部)。注意,所有的泳道都取自相同的Western印迹。
图49C显示了表达Flk-1/VEGFR2的基于Lm的构建体的设计。IFN-g ELISpot显示了在以每种构建体免疫后的离体CD8+T细胞限制性应答。未暴露的组只注射PBS;所有的组都含有对照Lm组。应答是针对每种Flk片段的对应的作图表位。每组N=5。图表显示均值±SEM;*p<0.05,Mann-Whitney统计检验,实验重复一次。
图50A显示了表达Flk-1/VEGFR2的基于Lm的构建体的设计。对于每一建成的构建体用方框标出了克隆区域,突出显示的/粗体氨基酸显示针对H2d/q MHC I单体型的作图的CTL表位。
图50B显示了flk基因的图谱,显示了用在本发明中的片段的一个实施例,
图50C显示了草图,显示用在本发明一个实施例中的flk片段如何与该flk基因的各个结构域相关联。
图50D显示了按方法中的描述进行巨噬细胞感染测定。将J774A.1细胞与李斯特菌构建体一起温育,洗涤,然后与庆大霉素一起温育,能够感染巨噬细胞并逃逸至细胞质中的细菌以Alexa-488(绿色)显示,PE CD11b+环(红色)标出了细胞的形状和大小。
图51A显示了Lm-LLO-Flk-1疫苗可在体内诱导建立的Her-2/neu+肿瘤的消退。来自每种构建体处理的小鼠的NT-2肿瘤体积(mm3)。曲线图显示均值±SEM;*p<0.05,Mann-Whitney统计检验,每组N=8只小鼠,实验重复两次。
图51B显示了IFN-g ELISpots,显示表位扩展至各种Her-2/neu区域。来自64天时间点的脾细胞以Her-2/neu肽表位离体再刺激。图表显示均值±SEM;*p<0.05,Mann-Whitney统计检验,每组N=5只小鼠,实验重复一次。
图51C显示在最初建立NT-2肿瘤后,在三周时间期间免疫小鼠三次。在这张图中,我们显示了对泛内皮细胞标志物CD31-PE的染色和采用DAPI的细胞核染色。在顺序切片上使用了同种型对照,如右侧所显示的。通过Image Pro软件进行血管密度的定量。图表显示均值±SEM;*p<0.05,Mann-Whitney检验,ns=无显著性。
图51D显示了对泛内皮细胞标志物CD31-PE染色,采用DAPI的细胞核染色和对核缺氧标志物缺氧诱导因子-1α(HIF-1α)的染色。
图52A显示具有完全消退的肿瘤的小鼠,显示出对肿瘤再次挑战的长期记忆。在第100天用NT-2在对侧胁腹内再次挑战具有完全消退肿瘤的小鼠。将盐水处理组用作我们的肿瘤生长阴性对照。
图52B显示了第100天再次挑战无肿瘤小鼠后生长肿瘤的小鼠的肿瘤体积。两图指的是单个实验。对于Flk-E1核Flk-I1组,无肿瘤小鼠的数量为2/8,盐水组具有5只小鼠。
图53A显示了抗血管生成疫苗在耐受HER-2/neu的小鼠中是无效的。A.向FVB/N野生型(WT)或FVB/N转基因(Tg)小鼠皮下注射1×106个NT-2细胞,在处理开始前让肿瘤生长直至可触知。一共免疫小鼠三次,这里显示平均肿瘤大小,直至肿瘤接种后69天。曲线图显示均值±SEM;*p<0.05,Mann-Whitney检验,实验重复两次。
图53B显示对脾进行处理用于IFN-g ELISpots,以各种Her-2/neu肽或者作为阴性对照的第三方肽(pGag)离体刺激。图表显示均值±SEM;*p<0.05,Mann-Whitney检验,实验重复一次。
图53C显示合并来自每个组的肿瘤,并进行消化用于TIL;这里我们显示针对CD8α和EC1或IC1特异性四聚体的Her-2/neu特异性T细胞染色。在野生型(WT)而不是转基因(Tg)小鼠中发现显著更多的Her-2/neu特异性T细胞;对照Lm组显示低的背景。实验重复一次,给出类似的结果。
图54A显示,当用4T1实验性转移瘤挑战时,经抗Flk-1Lm-疫苗保护的小鼠显示出降低的原发肿瘤生长,肿瘤负荷,以及减小的发病率和死亡率。A.在经Lm-LLO-Flk-1保护的动物中原发皮下4T1肿瘤生长较慢。用每种疫苗免疫小鼠三次,然后用50,000个4T1细胞皮下和静脉内注射。曲线图显示肿瘤体积的均值±SEM。
图54B显示了肿瘤负荷,显示为4T1细胞皮下挑战后无肿瘤小鼠的百分比。曲线图显示每个处理组8只小鼠的均值。
图54C展示了曲线图,该图显示基于可视检测和观察无疾病/健康小鼠的百分比。每组N=8只小鼠。
图55A显示,在乳腺癌的更有侵袭性的肿瘤模型中,Flk-1疫苗可保护小鼠免于实验性转移,并诱导弱的Her-2/neu表位扩展。A.以每种疫苗免疫小鼠两次,然后静脉内注射50,000个4T1细胞,让肿瘤生长25天,然后处死小鼠。在肺组织上进行H+E染色切片,手工对肿瘤结节计数。图表显示了每只动物每个肺叶的肺转移数量,均值±SEM;*p<0.05,Mann-Whitney检验,实验重复一次,显示了N=5只小鼠。
图55B显示了对来自这些动物的脾进行处理,并在IFN-g ELISpot测定中离体再次挑战,以用于Her-2/neu表位扩展。4T1细胞系确实表达低水平的小鼠Her-2/neu。仅在Flk-1-E1免疫小鼠中看到扩展。图显示与对照Lm组相对比的每孔斑点数的均值±SEM;*p<0.05,Mann-Whitney检验,实验重复一次,每组N=5。
图55C.显示了这样的实验,其中通过在一连串的三周以每种疫苗免疫来保护小鼠,然后静脉内注射50,000个4T1-Luc细胞,在四周时间期间对小鼠纵向成像,寻找肺种植的发生率和转移速率。
图55D显示了对于ROI内设门的表面积(sr),以每cm2每秒捕获的光子(p)计的平均发光度。曲线图显示均值±SEM;*p<0.05,Mann-Whitney检验。小鼠显著性如下:第18天,仅Flk-E1显著的;第25天,Flk-E1和Flk-I1与对照Lm比较均显著不同。
图56A显示采用抗血管发生Flk-1疫苗的安全性研究。如所有之前实验中进行的那样,将小鼠免疫三次,然后让小鼠交配或者进入伤口愈合研究。让小鼠(n=5/组)与同基因FVB/N雄性交配,在性交后观察到阴道栓时确认妊娠。这认为是dpc 0.5天。测量总的妊娠时长、到期幼崽质量(pup mass at term)以及总窝产崽数,图表显示均值±SEM;*p<0.05。
图56B.在每只接种小鼠的后背上产生一对无菌皮肤活检样本(n=5/组)。每天观察愈合。在第14天,所有的测试组都完成了愈合,对所有组都观察到几乎相同的愈合。图表显示直至伤口闭合的天数,均值±SEM;*p<0.05,Mann-Whitney检验。
图57.Flk-1疫苗诱导的表位扩展可能不是由于Flk-1和Her-2/neu共有结构域之间的交叉反应性。用对照Lm或Flk-I1疫苗免疫小鼠三次。处理脾细胞并离体再刺激,以响应于肽挑战而分泌IFN-g。所包括的肽为之前作图的pFlk-I1表位(PGGPLMVIV;SEQ ID NO:102),用于Her-2/neu的推定pIC1表位(GSGAFGTVYK;SEQ ID NO:99)或者所讨论的表位:推定的Her-2/neu和Flk-1激酶结构域之间共有的表位(GRGAFGQVI;SEQ ID NO:103),以及用作阴性对照的第三方表位(pGag)。图表显示均值±SEM,N=3/组。
图58A.在Her-2/neu乳腺癌的自发、常位模型中,Flk-1疫苗可显著延迟肿瘤生长。用每种Flk疫苗或单独的对照Lm免疫转基因FVB-rHer-2/neu小鼠三次。对来自每只小鼠的肿瘤检测突变的Her-2/neu信息。收集信使RNA,合成cDNA并测序。将得到的序列与野生型序列并排比对以确定突变的残基。只有出现4次或更多次的突变才考虑为真正的突变。在左侧有全部突变的汇总,这显示每组的N至少为3但不超过5只小鼠。合并所有的突变数据并叠加在大鼠Her-2/neu野生型序列上。粗体氨基酸残基为当疫苗针对Her-2/neu结构域时产生的突变。红色突出显示的氨基酸残基为当使用Flk-1疫苗时产生的突变。蓝色突出显示的区域显示Her-2/neu激酶结构域。绿色突出显示的结构域显示ATP结合结构域。
图58B.肿瘤生长是由于负责保持肿瘤被抑制的关键CTL表位出现了突变。更近地观察“热点”或成串的突变残基,我们发现几个突变的残基可在之前作图的CTL表位内找到。一个这样的表位显示关键氨基酸中导致将该表位锚定至H2Dq MHC I分子的突变。针对新的CTL表位,正在对其他的“热点”进行研究。
图59A.抗Her-2/neu人嵌合疫苗可延迟转移性乳腺癌细胞系在受保护小鼠的脑中的生长。以每种疫苗、抗人Her-2/neu或对照接种NYESO1,免疫Balb/c小鼠三次。使EMT6-Luc细胞在体外生长,然后以5,000个细胞/小鼠注射进麻醉小鼠的脑中。EMT6-Luc细胞表达低水平的小鼠Her-2/neu(数据未示出)。让细胞生长,然后在指定的天数成像。EMT6-Luc细胞产生荧光素酶,当它们在体内代谢D-荧光素时,副产物为光子,采用Xenogen X-100相机离体捕获这些光子并采用热图来展示。将像素密度作为每cm^2表面积每秒的光子数(以平均发光度表示)来作图。
图59B.抗HMWMAA人疫苗可延迟转移性黑素瘤细胞系在受保护小鼠的脑中的生长。以每种疫苗、抗人HMWMAA-C或对照接种NYESO1,免疫C57Bl/6小鼠三次。使B16F10-Luc细胞在体外生长,然后以5,000个细胞/小鼠注射进麻醉小鼠的脑中。B16F10亲代品系不表达HMWMAA,因此HMWMAA的唯一来源是周细胞和胶质细胞。荧光素酶机制和图像捕获与图59A中相同。
图60.内皮因子(CD105)的序列。克隆进Lm-LLO-CD5的原始片段为粗体并标有下划线,该原始片段基于Reisfeld小组克隆的序列。注意,Rankpep和其他MHC表位预测程序已显示,对于b、d和k H-2单倍体型,有几个备选的位于该区域之外的推定CTL表位(红色突出显示的)。
图61.新的表达CD105A和CD105B的李斯特菌构建体的设计。A.每一构建体的克隆区域为粗体,两个推定的表位标有下划线;Lm-LLO-CD105A和Lm-LLOCD105B合起来几乎覆盖了整个内皮因子基因,并涵盖更多的潜在CTL表位。B.将每个标下划线的片段克隆进表达载体pGG34,该表达载体与佐剂LLO融合。
图62.通过抗LLO抗体和Western印迹检测,Lm-LLO-CD105A表达并分泌大概大小为~80kD的蛋白。采用电穿孔用CD105A质粒转化XFL7菌株。将所转化的XFL7细胞在37μg/mL和250μg/μL的氯霉素和链霉素上铺板。让在两天的温育期中形成的集落生长在LB培养基中,快速离心(spum down),对上清液和细胞裂解物进行Western印迹以检测融合蛋白,或者是作为上清液中的分泌蛋白,或者是作为受限在细菌细胞内的内源蛋白。
图63.通过抗LLO抗体和Western印迹检测,Lm-LLO-CD105B表达并分泌大概大小为~74kD的蛋白。采用电穿孔用CD105A质粒转化XFL7菌株。将所转化的XFL7细胞在37μg/mL和250μg/μL的氯霉素和链霉素上铺板。让在两天的温育期中形成的集落生长在LB培养基中,快速离心(spum down),对上清液和细胞裂解物进行Western印迹以检测融合蛋白,或者是作为上清液中的分泌蛋白,或者是作为受限在细菌细胞内的内源蛋白。
图64.4T1肿瘤(2×105个细胞植入乳腺脂肪垫中)的生长,经Lm-LLO-CD105A和B免疫的Balb/c小鼠与对照疫苗Lm-LLO-NY-ESO-1比较。在指出的天数用2×108个cfu的每种疫苗接种小鼠。
图65.在第32天处死来自显示在图53B中的实验的小鼠,取出肺并以PBS灌注。在解剖显微镜下对可见的表面转移计数。与未暴露的(p<0.01)或Lm-LLO-NY-ESO1(p<0.05)相比,仅在Lm-LLO-CD105B观察到显著减少。
图66.用Lm-LLO-CD105A和B免疫诱导表位扩展至内源抗原HER-2/neu和gp70,并在脾中诱导抗原特异性T细胞。在显示在图53B中的实验中的肿瘤植入后的第22天,从3只小鼠取出脾,合并,在所显示的肽刺激后通过ELISpot分析单细胞悬液。注意,Kd和Dd为来自内皮因子序列的经预测会结合这些MHC I类分子的两种肽。它们位于CD105A:AGPRTVTVM(Dd)(SEQ ID NO:120)和CD105B AYSSCGMKV(Kd)(SEQ ID NO:121)中。
图67.用Lm-LLO-CD105A和B免疫诱导表位扩展至内源抗原HER-2/neu和gp70,并诱导浸润肿瘤的抗原特异性T细胞。在显示在图53B中的实验中的肿瘤植入后的第22天,从3只小鼠取出肿瘤,合并,并处理用于FACS分析,以EC1、EC2、IC1和AH1四聚体、抗CD8和CD62L、CD11B染色。将CD11B-群体设门CD8+,CD62L低,并采用所显示的四聚体分析抗原特异性。
图68.皮下植入FVB小鼠中的NT-2(1×106个细胞)肿瘤的生长,这些小鼠随后在第4、11和18天以Lm-LLO-CD105A和B或对照疫苗Lm-LLO-NY-ESO-1免疫,每种疫苗2×108个cfu。
图69A-B.SOE诱变策略。通过突变LLO的第4结构域实现LLO毒力的降低/下降。该结构域含有胆固醇结合位点,使得其能结合细胞膜,在这里其寡聚化以形成孔。
图70。图表显示各个小鼠和以下肿瘤测定日的肿瘤大小:在各种基于李斯特菌病的构建体给药后的第11、18和21天。
图71.A.工程化以表达和分泌作为融合蛋白的两种抗原的李斯特菌菌株LmddA244G的构建。抗原Her2嵌合体在基因上融合至基因组李斯特菌溶血素O,第二抗原HMW-MAA-C(HMC)融合至质粒中的截短的李斯特菌溶血素O。B.分别采用抗LLO和抗FLAG抗体通过Western印迹检测融合蛋白LLO-ChHer2和tLLO-HMC的分泌。
图72.采用山羊红血细胞定量LmddA244G-168的溶血活性。当与10403S相比时,观察到二价免疫治疗剂LmddA244G-168的溶血活性降低1.5倍。B.二价和单价免疫治疗剂在J774细胞系中的细胞内生长。LmddA244G-168的细胞内生长类似于单价治疗剂LmddA164和LmddA168。
图73.A.植入建立的NT2肿瘤,以单价治疗和二价治疗剂在第6、13和20天处理。未暴露的组为未处理的小鼠。B.不同的处理和未处理未暴露的组中的无肿瘤小鼠百分比。C.在LmddA244G-168处理后建立的NT2肿瘤的体积。
图74.在表达嵌合体Her2的单价(LmddA164)以及二价免疫治疗剂(LmddA244G-168)给药后,小鼠中Her2特异性免疫应答的产生。在采用FvB IC1肽表位-RLLQETELV(Seavey等人2009,Clin Cancer Res.2009年2月1日;15(3):924-32)的基于ELIspot的测定法中评估Her2特异性免疫应答。B.在表达HMW-MAA-C的单价(LmddA168)以及二价免疫治疗剂(LmddA244G-168)给药后,小鼠中HMW-MAA-C特异性免疫应答的产生。采用亲和纯化的HMA-MAA-C片段在基于ELISA的测定中评估Her2特异性免疫应答。
图75.以单价治疗剂和二价治疗剂在第1、8和15天处理建立的4T1肿瘤。未暴露的组为未处理的小鼠。
图76.以单价治疗剂、二价治疗剂或顺序单价治疗剂处理建立的NT2肿瘤。未暴露的组为未处理的小鼠。
应当理解,为了阐述的简洁和清楚,显示在图中的要素未必是按比率绘制的。例如,为了清楚,一些要素的尺寸可以是相对其他要素而放大的。另外,在认为适合时,附图标记可以在附图之间重复,以指出对应的或类似的要素。
具体实施方式
在一个实施例中,本文提供的发明旨在评估与受试者中疾病相关的生物标志物的表达或存在性,让它们在得自该受试者的生物样品中表达,以便鉴定并靶向与该疾病相关的生物标志物,由此治疗该受试者中的该疾病。在另一个实施例中,治疗该受试者的方法包括以靶向受试者中疾病表达的两种或更多种生物标志物的组合物或组合物的混合物给药。
在一个实施例中,本文提供的组合物包含重组李斯特菌菌株,所述菌株包含编码融合蛋白的核酸序列,其中所述融合蛋白包含从得自具有疾病的受试者的生物样品中鉴定的生物标志物,其中所述生物标志物与所述疾病相关,并且其中所述生物标志物融合至含PEST的多肽。在另一个实施例中,该重组李斯特菌包含编码重组多肽的核酸,该重组多肽包含融合蛋白。在另一个实施例中,该重组多肽为融合蛋白。
在一个实施例中,当以单一组合物(与组合物的混合物相反,即与作为混合物方案的一部分给药无关)向具有疾病的受试者给药时,该组合物包含含有编码至少一种融合蛋白的核酸序列的重组李斯特菌,该融合蛋白包含在得自该受试者的生物样品中鉴定的生物标志物,其中所述生物标志物与所述疾病相关,其中所述生物标志物融合至含PEST的多肽。在另一个实施例中,该李斯特菌包含编码1至3个、1至4个或1至6个融合蛋白的核酸序列,每个融合蛋白包含与疾病相关的不同生物标志物。在另一个实施例中,该李斯特菌包含含有1至3个、1至4个或1至6个开放阅读框(ORF)的核酸序列,每个开放阅读框编码包含与疾病相关的生物标志物的融合蛋白。在另一个实施例中,所述至少一个融合蛋白每个中的每个生物标志物与存在于从该核酸序列内的不同开放阅读框表达的另一融合蛋白中的另一生物标志物不同。
在另一个实施例中,当组合物作为混合物的一部分向受试者给药时,该混合物中的每个组合物包含重组李斯特菌菌株,所述菌株包含编码融合蛋白的核酸序列,其中所述融合蛋白包含在得自该受试者的生物样品中鉴定的生物标志物,其中所述生物标志物与所述疾病相关,其中所述生物标志物融合至含PEST的多肽,并且其中每个组合物中的每个李斯特菌菌株包含与其余生物标志物不同的生物标志物。在一个实施例中,混合物为疫苗混合物。在另一个实施例中,混合物包含预订数量的组合物,每个组合物包含重组李斯特菌,该重组李斯特菌表达在所述疾病中表达的生物标志物和含PEST多肽的融合蛋白。在另一个实施例中,组合物的混合物为组合物的组合。
技术人员应当理解,组合物的混合物中的每个组合物都可以在单次剂量中同时给药或者随时间分开给药,并且可靶向疾病表达的多于一个生物标志物。在另一个实施例中,组合物的混合物中的每个组合物都可以在单次剂量中同时给药,并将在同一时间靶向疾病表达的多于一个生物标志物。在另一个实施例中,包含表达至少一种融合蛋白的重组李斯特菌的组合物在同一时间靶向疾病表达的多于一个生物标志物。在一个实施例中,包含表达至少一种融合蛋白的重组李斯特菌的组合物的混合物中的预定数量的混合物同时或者随时间分开向受试者给药。
在一个实施例中,当以组合物的混合物向具有疾病的受试者给药时,该混合物中的每个组合物可以间隔1至2天、间隔1至3天、间隔1至5天、间隔1至10天或间隔1至14天给药。
在一个实施例中,当以组合物的混合物向具有疾病的受试者给药时,每个组合物可以以预定的剂量给药,该预定的剂量之前已确定为接受该给药的受试者的最佳剂量。这种最佳剂量可以由临床医生或技术人员在以该混合物给药之前通过实验确定。在另一个实施例中,混合物包含预订数量的组合物,每个组合物包含重组李斯特菌,该重组李斯特菌表达生物标志物和含PEST多肽的融合蛋白。在另一个实施例中,组合物的混合物为组合物的组合,每个组合物包含重组李斯特菌菌株,该重组李斯特菌菌株表达融合至含PEST的多肽的生物标志物的单一融合蛋白。
在一个方面,本发明涉及在具有疾病的受试者中诱导针对所述疾病的免疫应答的方法,该方法包括以下步骤:a.从所述受试者获得生物样品;b.评估所述生物样品中预定数量的生物标志物的表达;c.以包含重组李斯特菌菌株的组合物向所述受试者给药,所述菌株包含编码至少一种融合蛋白的核酸序列,其中所述融合蛋白包含在得自所述受试者的所述生物样品中鉴定的生物标志物,其中所述生物标志物与所述疾病相关,其中所述生物标志物融合至含PEST的多肽,并且其中所述李斯特菌内的每种融合蛋白包含不同的生物标志物,因而在所述受试者中诱导多靶点抗疾病免疫应答。
在一个实施例中,包含编码至少一种融合蛋白的核酸序列的李斯特菌菌株编码1至2种融合蛋白。在另一个实施例中,该核酸序列编码1至3种融合蛋白、1至4种融合蛋白、1至5种融合蛋白、1至10种融合蛋白、2至3种融合蛋白、2至4种融合蛋白、2至5种融合蛋白或2至10种融合蛋白。
在一个方面,本发明涉及在具有疾病的受试者中诱导针对所述疾病的免疫应答的方法,该方法包括以下步骤:a.从所述受试者获得生物样品;b.评估所述生物样品中预定数量的生物标志物的表达;c.以预定数量的每个包含重组李斯特菌菌株的组合物向所述受试者给药,所述菌株包含编码融合蛋白的核酸序列,其中所述融合蛋白包含在得自所述受试者的所述生物样品中鉴定的生物标志物,其中所述生物标志物与所述疾病相关,其中所述生物标志物融合至含PEST的多肽,并且其中每个组合物内的每种李斯特菌菌株在所述融合蛋白中包含不同的生物标志物,因而在所述受试者中诱导多靶点抗疾病免疫应答。
在一个实施例中,本发明涉及治疗受试者中疾病的方法,该方法包括以下步骤:a.从所述受试者获得生物样品;b.评估所述生物样品中预定数量的生物标志物的表达;c.以包含重组李斯特菌菌株的组合物向所述受试者给药,所述菌株包含编码至少一种融合蛋白的核酸序列,其中所述融合蛋白包含在得自所述受试者的所述生物样品中鉴定的生物标志物,其中所述生物标志物与所述疾病相关,其中所述生物标志物融合至含PEST的多肽,并且其中所述李斯特菌内的每种融合蛋白包含不同的生物标志物,因而治疗所述受试者中的所述疾病。
在另一个实施例中,本发明涉及治疗受试者中疾病的方法,该方法包括以下步骤:a.从所述受试者获得生物样品;b.评估所述生物样品中预定数量的生物标志物的表达;c.以预定数量的每个包含重组李斯特菌菌株的组合物向所述受试者给药,所述菌株包含编码融合蛋白的核酸序列,其中所述融合蛋白包含在所述生物样品中鉴定的生物标志物,其中所述生物标志物与所述疾病相关,其中所述生物标志物融合至含PEST的多肽,并且其中每个组合物内的每种李斯特菌菌株包含不同的生物标志物,因而治疗所述受试者中的所述疾病。
在另一个实施例中,本发明涉及预防受试者中疾病复发的方法,该方法包括以下步骤:a.从所述受试者获得生物样品;b.评估所述生物样品中预定数量的生物标志物的表达;c.以包含重组李斯特菌菌株的组合物向所述受试者给药,所述菌株包含编码至少一种融合蛋白的核酸序列,其中所述融合蛋白包含在所述生物样品中鉴定的生物标志物,其中所述生物标志物与所述疾病相关,其中所述生物标志物融合至含PEST的多肽,并且其中所述李斯特菌内的每种融合蛋白包含不同的生物标志物,因而预防所述受试者中所述疾病的复发。
在一个实施例中,本文提供的疾病为癌症或肿瘤生长。在另一个实施例中,本文提供的疾病为感染性疾病、呼吸道疾病、炎症性疾病或受试者具有Th2持续情况的疾病。在另一个实施例中,该疾病为局部疾病,即限于具体的疾病部位,或者为全身性疾病。
在一个实施例中,所述融合蛋白为转录的融合蛋白。在另一个实施例中,本文提供的李斯特菌表达该融合蛋白,该融合蛋白包含融合至本文提供的含PEST的多肽的生物标志物。在另一个实施例中,该融合蛋白表达本文提供的生物标志物。
在一个实施例中,本发明的方法和组合物的李斯特菌为单核细胞增多性李斯特菌。在另一个实施例中,该李斯特菌为伊氏李斯特菌(Listeria ivanovii)。在另一个实施例中,该李斯特菌为威氏利斯特菌(Listeria welshimeri)。在另一个实施例中,该李斯特菌为西尔李斯特菌(Listeria seeligeri)。每种类型的李斯特菌代表了本发明的独立实施例。
在一个实施例中,本文提供了组合物的混合物,其中每个组合物包含表达生物标志物的重组李斯特菌菌株,该生物标志物靶向针对疾病中存在的各个生物标志物的免疫应答。在另一个实施例中,该生物标志物存在于与所述疾病相关的组织的表面上。在另一个实施例中,本文提供的疾病为肿瘤或癌症。在另一个实施例中,生物标志物为表面生物标志物。在另一个实施例中,生物标志物为细胞内生物标志物。在另一个实施例中,当疾病为肿瘤或癌症时,该生物标志物为与肿瘤或癌症脉管系统相关的血管生成生物标志物。
在另一个实施例中,预定数量的每个包含李斯特菌菌株的组合物为至少1至约10个组合物。在另一个实施例中,预定数量的组合物为至少1至约20个组合物。在另一个实施例中,预定数量的组合物为2至5个组合物、3至6个组合物、4至7个组合物、5至10个组合物、6至11个组合物或7至12个组合物。
在一个实施例中,预定数量的生物标志物为至少1至约10个生物标志物。在另一个实施例中,预定数量的生物标志物为至少1至约20个生物标志物、至少1至约30个生物标志物、至少1至约40个生物标志物、至少1至约50个生物标志物、至少51至约60个生物标志物、至少71至约80个生物标志物、至少81至约90个生物标志物或至少1至约100个生物标志物。
本领域技术人员应该理解,术语“生物标志物”可以涵盖抗原,包括异源抗原、肿瘤抗原、血管生成抗原等。该术语还可以涵盖与疾病相关的或由疾病表达的蛋白、DNA、RNA、肽,该疾病包括但不限于癌症、肿瘤、感染性疾病、自身免疫疾病、先天性疾病等。还应当理解,与健康宿主中生物标志物的正常表达水平相比,这些生物标志物可以在具有疾病的受试者中过表达。在另一个实施例中,受试者耐受生物标志物,以致表达该生物标志物的疾病可容易地进展而没有引起恰当的针对它的免疫应答。在另一个实施例中,术语“生物标志物”指由疾病表达的抗原。在另一个实施例中,该生物标志物为异源抗原或其片段。在一个实施例中,该生物标志物为肿瘤抗原或其片段。在另一个实施例中,本文提供的生物标志物为在宿主内引起变态反应或炎症性反应的变应原。
在一个实施例中,术语“肿瘤标志物”和“肿瘤抗原”在本文中可互换使用,并指由肿瘤表达的抗原。在另一个实施例中,该肿瘤标志物为异源肿瘤抗原或其片段。在一个实施例中,肿瘤标志物与所述肿瘤的形成或增生相关。在另一个实施例中,肿瘤标志物由所述肿瘤或由所述肿瘤的脉管系统表达。在另一个实施例中,肿瘤标志物由肿瘤分泌。在另一个实施例中,本文提供的肿瘤标志物与局部组织环境相关,该局部组织环境进一步与癌症的发展或转移相关。在另一个实施例中,肿瘤标志物与肿瘤逃避或抵抗癌症相关,或者为血管生成抗原。在另一个实施例中,肿瘤标志物由肿瘤在肿瘤表面上表达。在另一个实施例中,肿瘤标志物存在于肿瘤内,并在肿瘤细胞溶解时释放到细胞外环境。
在一个实施例中,对生物标志物情况或生物标志物表达水平的评估包括从具有疾病的受试者获得生物样品并检测所述样品中生物标志物的表达水平。在一个实施例中,本文提供的生物标志物情况为肿瘤标志物情况。在另一个实施例中,对肿瘤标志物情况或肿瘤标志物表达水平的评估包括从具有肿瘤的受试者获得生物样品并获得所述样品中肿瘤标志物的表达水平。
技术人员应当理解,生物标志物表达情况可以通过使用本领域已知的可用于测定生物标志物表达水平的任何测定法来测定。这些测定法包括但不限于免疫测定(例如ELISA)、FACS、免疫组化测定、基于荧光的测定、PCR、单独或与质谱联用的定量HPLC或者本领域已知的任何其他测定法。然后,可将所测定的表达情况与诸如来自健康受试者的对照情况进行比较,以有效地诊断受试者中的疾病(例如肿瘤或癌症)。在另一个实施例中,在以包含重组李斯特菌菌株的组合物给药之前,可在得自具有疾病的受试者的生物样品中检测出生物标志物。
在另一个实施例中,本文提供了监测受试者中疾病进展以便确定以本发明的组合物或组合物的混合物给药的最佳时间的方法,该方法包括从该受试者获得生物样品并测定该生物样品中生物标志物的表达情况的步骤,其中在该受试者中测定生物标志物表达水平高于在对照样品中观察到的水平使得能够监测受试者中疾病的进展,并且其中本发明的组合物或组合物的混合物在将实现最大疗效的预定时间给药。本发明涵盖的疾病包括但不限于癌症、肿瘤生长、感染性疾病或受试者具有Th2偏向情况的疾病。这些疾病根据受试者中疾病进展进一步表征或分期。这种信息可用于确定以本发明的一种或多种组合物给药的最佳时期。
技术人员应当理解,所述生物样品可以包括但不限于组织、血液、血清、DNA、RNA、尿、精液、滑液、痰或脑脊液(CSF)。
在一个实施例中,“Th2偏向的受试者”也称为具有Th2表型的受试者,其中Th1免疫应答作为受试者中感染性疾病(包括但不限于寄生虫感染)或癌症的结果是有缺陷的、缺乏的或者被压制的。在另一个实施例中,Th2偏向的受试者指这样的受试者,其中Th2应答并非排他性地存在于受试者中,而是在该受试者中相对Th1应答占优势。在另一个实施例中,Th2偏向的受试者指这样的受试者,其中Th2应答排他性地存在于受试者中,而Th1应答的指示物(即细胞因子、趋化因子或其他已知的标志物)的水平极低或微不足道。
在一个实施例中,组合物的混合物每个包含1至5个组合物,每个组合物包含表达单一融合蛋白的重组李斯特菌菌株。在另一个实施例中,该混合物包含1至10个组合物、1至15个组合物、5至10个组合物、5至15个组合物、5至20个组合物,每个包含表达含PEST的多肽与生物标志物的单一融合蛋白的重组李斯特菌菌株。在另一个实施例中,混合物中的每个组合物包含表达不同生物标志物的李斯特菌。
在另一个实施例中,本文提供的组合物或组合物的混合物与不同于最初的组合物或组合物的混合物的替代形式的疫苗或组合物的给药同时给药。在另一个实施例中,替代形式的疫苗与本文提供的组合物或组合物的混合物的给药分开给药。技术人员应答理解,替代形式的疫苗或组合物可以包括但不限于:编码包含生物标志物和含PEST的肽或其片段的融合蛋白的DNA疫苗,包含所述融合蛋白的病毒载体,包含本文提供的生物标志物的病毒载体,包含所述融合蛋白的病毒样颗粒,包含本文提供的生物标志物的病毒样颗粒,包含所述融合蛋白的重组肽或重组多肽,表达本文提供的生物标志物的基于细胞的疫苗,或者包含单独的本文提供的或本文进一步提供的融合蛋白形式的生物标志物的活的重组非李斯特菌细菌载体。技术人员还应该理解,这些替代形式不仅可以与本文提供的组合物或组合物的混合物联合使用,还可以在它们给药之前或之后给药。
在一个实施例中,本文提供的重组多肽包含本文提供的融合蛋白。在另一个实施例中,本文提供的重组多肽为本文提供的融合蛋白。
在另一个实施例中,用在上文描述的任何方法中的组合物或组合物的组合具有本发明的疫苗和组合物的任意特征。
术语“免疫原性组合物”、“组合物”和“药物组合物”可以互换使用。例如,在一个实施例中,本发明的组合物可以包含本文描述的重组李斯特菌和佐剂。在另一个实施例中,免疫原性组合物包含本文提供的重组李斯特菌。在另一个实施例中,免疫原性组合物包含本领域已知的或本文提供的佐剂。还应理解的是,这些组合物的给药增强免疫应答或增加T效应细胞与调节性T细胞比率或者引起抗肿瘤免疫应答,如本文进一步提供的。
术语“药物组合物”可以涵盖治疗有效量的包括含李斯特菌菌株的组合物在内的一种或多种活性成分,连同药学上可接受的载体或稀释剂。
技术人员应理解,术语“给药”可以涵盖让受试者与本发明的组合物接触。在一个实施例中,给药可在体外即在试管中完成,或者在体内即活生物体(如人)的细胞或组织内完成。在一个实施例中,本发明包括以本发明的李斯特菌菌株及其组合物向受试者给药。
在一个实施例中,细菌载体为细胞内病原体。在另一个实施例中,载体源自细胞质病原体。在另一个实施例中,载体源自细胞内病原体。在另一个实施例中,细胞内病原体诱导主要是细胞介导的免疫应答。在另一个实施例中,该载体为沙门氏菌(Salmonella)菌株。在另一个实施例中,该载体为BCG菌株。在另一个实施例中,该载体为细菌载体。在另一个实施例中,可以以经本发明的载体转导的树突状细胞向受试者给药,以上调该受试者的免疫应答,在一个实施例中其通过上调CTL活性来完成。
在另一个实施例中,重组疫苗载体诱导主要是Th1型免疫应答。
在另一个实施例中,载体选自沙门氏菌(Salmonella sp.)、志贺氏菌(Shigella sp.)、BCG、单核细胞增多性李斯特菌、大肠杆菌以及戈氏链球菌(S.gordonii)。在另一个实施例中,由经修饰以逃避吞噬溶酶体融合并生存在细胞的细胞质中的重组细菌载体来递送融合蛋白。在另一个实施例中,载体为病毒载体。在其他实施例中,载体选自牛痘病毒、禽痘病毒、腺病毒、AAV、牛痘病毒NYVAC、经修饰的安卡拉株牛痘病毒(MVA)、塞姆利基森林病毒、委内瑞拉马脑炎病毒、疱疹病毒以及逆转录病毒。在另一个实施例中,载体为裸DNA载体。在另一个实施例中,载体为本领域已知的任何其他载体。每种可能性代表了本发明的独立实施例。
在一个实施例中,病毒载体为腺病毒载体、逆转录病毒载体、慢病毒载体、痘病毒载体、杆状病毒载体、单纯疱疹病毒载体、腺相关病毒载体、纳米工程化病毒样物质或本领域已知用在疫苗中的任何病毒载体。在另一个实施例中,该病毒载体为牛痘病毒载体。
技术人员应当理解,基于细胞的疫苗可以包括活细胞或死细胞,并且还可以包括自体或异源来源的肿瘤细胞。
在一个实施例中,在以本文所提供的任何疫苗给药之前获得生物标志物表达情况。
在一个实施例中,基于肽的疫苗包含融合至本文提供的肿瘤标志物或抗原的去毒LLO,该LLO具有突变的或缺失的胆固醇结合结构域(参见实例38-39)。在另一个实施例中,基于肽的疫苗与本文提供的包含重组李斯特菌菌株的组合物联合,以用于提供对包括癌症在内的疾病的多靶点免疫治疗。
在另一个实施例中,本发明的方法还包括向接受了本文提供的免疫治疗的受试者以强化剂量给药的步骤。在另一个实施例中,该强化剂量在本文提供的组合物或组合物的混合物的初次给药后给药。在另一个实施例中,该方法还包括在本文提供的组合物或组合物的混合物的首次给药后从所述受试者获得生物标志物情况并以强化剂量给药的步骤。在另一个实施例中,所给药的强化剂量包含本文提供的组合物以及本文进一步提供的替代形式的疫苗。在另一个实施例中,该方法还包括在以本文提供的组合物或组合物的混合物在先给药后获得第二、第三、第四、第五次等等的生物标志物情况,并在获得所述第三、第四、第五次等等的生物标志物情况后以强化剂量给药或者与替代形式的疫苗或组合物联合。在另一个实施例中,该生物标志物为肿瘤标志物,该生物标志物情况为肿瘤标志物情况。在另一个实施例中,强化剂量与本文提供的组合物或者组合物的混合物中的任一组合物的初次剂量相同或不同。
在一个实施例中,强化接种在单次初免接种或给药之后。在另一个实施例中,单次强化接种在初免接种后给药。在另一个实施例中,两次强化接种在初免接种后给药。在另一个实施例中,三次强化接种在初免接种后给药。在一个实施例中,初免和强化疫苗之间的间隔时间由技术人员通过实验确定。在另一个实施例中,初免和强化疫苗之间的间隔时间为1周,在另一个实施例中为2周,在另一个实施例中为3周,在另一个实施例中为4周,在另一个实施例中为5周,在另一个实施例中为6-8周,在又另一个实施例中强化疫苗在初免疫苗后8-10周给药。
异源的“初免强化”策略对于增强免疫应答和防御众多病原体已是有效的。Schneider等人.,Immunol.Rev.170:29-38(1999);Robinson,H.L.,Nat.Rev.Immunol.2:239-50(2002);Gonzalo,R.M.等人,Vaccine 20:1226-31(2002);Tanghe,A.,Infect.Immun.69:3041-7(2001)。将抗原以不同形式提供在初免和强化注射中似乎使针对该抗原的免疫应答最大化。以DNA疫苗初免,接着以佐剂中的蛋白或者通过病毒载体递送编码抗原的DNA来强化似乎是改善抗原特异性抗体以及各自的CD4+T细胞应答或CD8+T细胞应答的最有效方式。Shiver J.W.等人,Nature 415:331-5(2002);Gilbert,S.C.等人,Vaccine 20:1039-45(2002);Billaut-Mulot,O.等人,Vaccine 19:95-102(2000);Sin,J.I.等人,DNA Cell Biol.18:771-9(1999)。最近的来自猴接种研究的数据提示,当以HIV gag DNA对猴进行初免接种后接着以表达HIV gag的腺病毒载体(Ad5-gag)强化时,向编码HIV gag抗原的DNA添加CRL1005泊洛沙姆(12kDa,5%POE)增强了T细胞应答。针对DNA/泊洛沙姆初免和随后的Ad5-gag强化的细胞免疫应答强于以DNA(无泊洛沙姆)初免和随后进行Ad5-gag强化所诱导的应答或者针对单独的Ad5-gag的应答。Shiver,J.W.等人Nature 415:331-5(2002)。美国专利申请公开US 2002/0165172 A1描述了以编码抗原的免疫原性部分的载体构建体和包含抗原的该免疫原性部分的蛋白同时给药,由此产生免疫应答。该文献限于乙型肝炎抗原和HIV抗原。此外,美国专利No.6,500,432涉及通过以多核苷酸和目的多肽同时给药来增强核酸接种的免疫应答的方法。根据该专利,同时给药指在相同的免疫应答期间以多核苷酸和多肽给药,优选在彼此的0-10天或3-7天内。该专利所考虑的抗原包括:肝炎(所有形式)、HSV、HIV、CMV、EBV、RSV、VZV、HPV、脊髓灰质炎、流感、寄生虫(例如,来自疟原虫(Plasmodium)属)和病原菌(包括但不限于结核分枝杆菌(M.tuberculosis)、麻风杆菌(M.leprae)、衣原体(Chlamydia)、志贺氏菌(Shigella)、伯氏疏螺旋体(B.burgdorferi)、肠毒素大肠杆菌(enterotoxigenic E.coli)、伤寒沙门氏菌(S.typhosa)、幽门螺杆菌(H.pylori)、霍乱弧菌(V.cholerae)、百日咳杆菌(B.pertussis)等)抗原等。通过引用将以上所有文献全文并入本文。
在一个实施例中,本文提供的组合物可以称作疫苗,本文提供的组合物的混合物或者组合物的组合可以称作疫苗组合。在其他实施例中,疫苗组合可以在本文提供的组合物或组合物的组合之外包括替代形式的疫苗。
技术人员应当理解,可以基于在耐药肿瘤或复发肿瘤中检测到的变化或者在初次治疗或给药后的时间点收集的数据对疫苗组合或给药进行调整(以靶向另外的或新的肿瘤标志物)。例如,如果肿瘤表达标志物A、B、C和D,则所给予的免疫疗法的总剂量将会由靶向针对每一单独标志物的免疫应答的包含本文提供的重组李斯特菌菌株的组合物或组合物的混合物构成。在一个实施例中,当以组合物的混合物给药时,以单次剂量同时给药,组合物的混合物中的至少一种组合物在不同时间给药,也就是说,其中包含特异李斯特菌菌株的来自该混合物的一个组合物靶向生物标志物A和B,而在不同的时间,包含另一种重组李斯特菌菌株的来自该混合物的另一个组合物靶向生物标志物C和D。
在另一个实施例中,10个组合物的混合物中的2至4个组合物在该混合物中的其余组合物之前给药。在另一个实施例中,10个组合物的混合物中的2至6个组合物在该混合物中的其余组合物之前给药。在另一个实施例中,10个组合物的混合物中的5至8个组合物在该混合物中的其余组合物之前给药。在另一个实施例中,10个组合物的混合物中的全部组合物都在不同时间点给药。在另一个实施例中,10个组合物的混合物中的全部组合物都同时给药。在另一个实施例中,该混合物包含5至10个组合物、11至15个组合物或16至20个组合物。
在一个实施例中,本文提供的方法增加具有疾病的受试者内或该受试者的疾病部位内浸润性T淋巴细胞/抑制细胞的比率。在另一个实施例中,本文提供的方法增加具有疾病的受试者内或该受试者的疾病部位内CD8+T细胞/抑制细胞的比率。在另一个实施例中,本文提供的增加浸润性T淋巴细胞/抑制细胞或CD8+T细胞/抑制细胞比率的方法包括以包含所述疫苗的组合物或本文提供的组合物向该受试者给药的步骤。
在一个实施例中,本文提供的方法增加具有疾病的受试者内或该受试者的疾病部位内浸润性T淋巴细胞/髓源性抑制细胞(MDSC)的比率。在另一个实施例中,本文提供的方法增加具有疾病的受试者内或该受试者的疾病部位内CD8+T细胞/髓源性抑制细胞(MDSC)的比率。在另一个实施例中,本文提供的增加浸润性T淋巴细胞/髓源性抑制细胞(MDSC)或CD8+T细胞/髓源性抑制细胞比率的方法包括以包含所述疫苗的组合物或本文提供的组合物向该受试者给药的步骤。
在一个实施例中,该浸润性T淋巴细胞为肿瘤浸润性T淋巴细胞。在一个实施例中,本文提供的抑制细胞为调节性T细胞(Treg)。在一个实施例中,该抑制细胞为髓源性抑制细胞(MDSC)。
在一个实施例中,本文提供的方法降低抑制针对疾病的免疫应答的细胞的量。在另一个实施例中,该抑制免疫应答的细胞为抑制性细胞。在另一个实施例中,该抑制性细胞为髓源性抑制细胞(MDSC)。在另一个实施例中,该抑制性细胞为调节性T细胞(Treg)。
在一个实施例中,肿瘤MDSC可出乎意料地抑制抗原特异性功能和非特异性T细胞功能,而脾MDSC仅能抑制抗原特异性T细胞的功能。如下文实例(参见实例17-20)中所显示的,与疾病部位(例如肿瘤部位)处TIL群相比,本文提供的活的减毒李斯特菌减少疾病中抑制细胞的百分比。
在一个实施例中,本文提供的包含在基于单核细胞增多性李斯特菌(Lm)的疫苗中的重组李斯特菌菌株降低疾病部位Treg和MDSC的百分比,在疾病部位具有对应的效应细胞与抑制细胞比率的偏移。在另一个实施例中,本文提供的基于Lm的疫苗可用于通过降低受试者中特定疾病部位Treg和MDSC的百分比以及MDSC的绝对数量来改善免疫应答。这种部位可为变态反应、创伤、感染、疾病导致的炎症部位,或者该部位可为肿瘤部位。
在另一个实施例中,从经李斯特菌处理的小鼠的肿瘤纯化的单核细胞和粒细胞MDSC,与从未处理小鼠的肿瘤纯化的MDSC相比,抑制CD8+T细胞分裂的能力较弱,然而从这些相同荷瘤小鼠的脾纯化的单核细胞和粒细胞MDSC未显示出以李斯特菌接种后它们的功能变化(参见本文实例17-20)。在一个实施例中,观察到这种效应是因为脾MDSC的抑制性为抗原特异性方式。因而,以李斯特菌处理具有的明显优势在于使得能肿瘤特异性抑制肿瘤抑制性细胞,例如Treg和MDSC。本文提供的方法和组合物的活的减毒李斯特菌菌株提供的另一出乎意料的优势为,在肿瘤中有较低量的Treg,并且仍存在的也丧失了抑制T细胞复制的能力。
在另一个实施例中,从经表达截短的LLO的李斯特菌处理的小鼠的肿瘤纯化的单核细胞和粒细胞MDSC,与从未处理小鼠的肿瘤纯化的MDSC相比,抑制CD8+T细胞分裂的能力较弱,然而从这些相同荷瘤小鼠的脾纯化的单核细胞和粒细胞MDSC未显示出在以表达截短的LLO的李斯特菌免疫接种后它们的功能变化(参见本文实例21)。在一个实施例中,观察到这种效应是因为脾MDSC的抑制性仅为抗原特异性方式。因而,以表达截短的LLO的李斯特菌处理具有的明显优势在于使得能肿瘤特异性抑制肿瘤抑制性细胞,例如Treg和MDSC。本文提供的方法和组合物的表达截短的LLO的活的减毒李斯特菌菌株提供的另一出乎意料的优势为,在肿瘤中有较低量的Treg和MDSC,并且仍存在的Treg和MDSC丧失了抑制T细胞复制的能力,这种效应甚至在不存在LLO融合伴侣(例如异源抗原)的情况下也观察到。
在另一个实施例中,表达截短的LLO的活的减毒李斯特菌菌株的给药通过抑制Treg和MDSC介导的T细胞抑制来增强抗肿瘤T细胞应答(参见本文实例21)。
在一个实施例中,本文提供了降低受试者中疾病部位内抑制细胞百分比的方法,该方法包括以本文提供的包含活的减毒李斯特菌菌株的组合物或者包含活的减毒李斯特菌疫苗菌株的组合物的混合物向该受试者给药的步骤。
在另一个实施例中,本文提供了降低受试者中疾病部位内抑制细胞抑制T细胞复制的能力的方法,该方法包括以本文提供的包含活的减毒李斯特菌菌株的组合物或者包含活的减毒李斯特菌疫苗菌株的组合物的混合物向该受试者给药的步骤。
在一个实施例中,降低疾病部位内抑制细胞的数量有效地治疗了疾病。在另一个实施例中,降低疾病部位抑制细胞的数量增强了具有疾病的受试者中在该疾病部位的抗疾病免疫应答。在另一个实施例中,免疫应答是细胞介导的免疫应答。在另一个实施例中,免疫应答是肿瘤浸润性T淋巴细胞(TIL)免疫应答。
在一个实施例中,本文提供的方法降低了受试者中疾病内抑制细胞的百分比并增强了受试者中抗疾病的治疗应答。
在另一个实施例中,本文提供的方法降低了受试者中疾病内抑制细胞抑制T细胞复制的能力并增强了受试者中抗疾病的治疗应答。
在一个实施例中,术语“降低...的百分比”为Treg或MDSC抑制细胞的量的表示,它们在疾病部位的存在相对于T浸润性细胞的存在减少了或降低了,如在测定法或免疫应答中测量的。
在另一个实施例中,术语“降低...的数量”指,作为本文提供的包含在基于李斯特菌的疫苗中的活的减毒李斯特菌菌株或者也在本文其他地方描述的获得类似效果的这种疫苗的替代形式给药的结果,Treg或MDSC这些抑制细胞的绝对数量降低了或减少了。
在一个实施例中,本文提供了含PEST的多肽,所述生物标志物或者肿瘤标志物与其融合。在另一个实施例中,该含PEST的多肽为N末端李斯特菌溶血素O(LLO)或截短的LLO、PEST氨基酸序列或PEST序列或N末端ActA序列或截短的ActA。
在另一个实施例中,本发明包括与包含启动子/调控序列的核酸可操作地连接的编码截短的ActA、截短的LLO或PEST氨基酸序列的分离的核酸以及编码肿瘤标志物或其免疫原性片段的分离的核酸,使得该核酸优选地能够指导由该核酸编码的蛋白的表达。本发明还包括包含本发明的分离的核酸的载体。因此,本发明涵盖表达载体和将外源DNA引入细胞并在该细胞中同时表达该外源DNA的方法,例如在Sambrook等人(1989,Molecular Cloning:A Laboratory Manual,Cold Spring Harbor Laboratory,New York)以及在Ausubel等人(1997,Current Protocols in Molecular Biology,John Wiley&Sons,New York)中描述的那些。
在另一个实施例中,术语“核酸”或“核苷酸”指一串至少两个碱基-糖-磷酸的组合。该术语在一个实施例中包括DNA和RNA。“核苷酸”在一个实施例中指核酸聚合物的单体单元。RNA在一个实施例中为tRNA(转运RNA)、snRNA(小核RNA)、rRNA(核糖体RNA)、mRNA(信使RNA)、反义RNA、小抑制性RNA(siRNA)、微RNA(miRNA)和核酶的形式。siRNA和miRNA的用途已有描述(Caudy AA等人,Genes&Devel 16:2491-96和其中引用的参考文献)。DNA在另一个实施例中可为质粒DNA、病毒DNA、线性DNA或染色体DNA或这些组的衍生物的形式。此外,这些形式的DNA和RNA可以是单链、双链、三链或四链的。在另一个实施例中,该术语还包括人工核酸,其含有其他类型的主链,但含有相同的碱基。在一个实施例中,该人工核酸是PNA(肽核酸)。PNA含有肽骨架和核苷酸碱基,且在一个实施例中,能够结合DNA和RNA分子。在另一个实施例中,该核苷酸被氧杂环丁烷修饰。在另一个实施例中,该核苷酸通过用硫代磷脂键置换一个或多个磷酸二酯键而被修饰。在另一个实施例中,所述人工核酸含有本领域已知的天然核酸的磷酸酯骨架的任何其他变体。硫代磷酸酯核酸和PNA的用途是本领域技术人员已知的,例如描述在Neilsen PE,Curr Opin Struct Biol9:353-57和Raz NK等人Biochem Biophys Res Commun.297:1075-84中。核酸的制备和用途是本领域技术人员已知的,例如描述在Molecular Cloning,(2001),Sambrook和Russell编以及Methods in Enzymology:Methods for molecular cloning in eukaryotic cells(2003)Purchio和G.C.Fareed.中。每种核酸衍生物代表了本发明的独立实施例。
技术人员应该理解,术语“分离的核酸”可以涵盖已从天然存在的状态中在侧面与其连接的序列中分离出来的核酸区段或片段,例如已从通常与该片段邻接的序列中移出的DNA片段,该序列例如在其天然存在的基因组中与所述片段邻接。该术语还适用于已从天然陪伴该核酸的其他组分(例如在细胞中与其陪伴的RNA或DNA或蛋白)中基本上纯化的核酸。该术语因此包括例如并入到载体中的、并入自主复制的质粒或病毒中的或并入原核细胞或真核细胞的基因组DNA中的重组DNA,或者其作为独立于其他序列的单独分子(例如,作为cDNA或者通过PCR或限制酶消化产生的基因组或cDNA片段)存在。其还包括作为编码另外的多肽序列的杂种基因的一部分的重组DNA。
在一个实施例中,编码本文提供的重组多肽或融合蛋白的核酸还包含信号肽或序列。在一个实施例中,异源抗原可以通过使用信号序列来表达,例如李斯特菌信号序列,例如溶血素信号序列或actA信号序列。备选地,例如,外来基因可从单核细胞增多性李斯特菌启动子下游表达,无需产生融合蛋白。在另一个实施例中,该信号肽为细菌的(李斯特菌或非李斯特菌的)。在一个实施例中,该信号肽为该细菌自有的。在另一个实施例中,该信号肽对该细菌为外来的。在另一个实施例中,该信号肽为来自单核细胞增多性李斯特菌的信号肽,例如secA1信号肽。在另一个实施例中,该信号肽为来自乳酸乳球菌(Lactococcus lactis)的Usp45信号肽,或者来自炭疽芽孢杆菌(Bacillus anthracis)的保护性抗原信号肽。在另一个实施例中,该信号肽为来自单核细胞增多性李斯特菌的secA2信号肽,例如p60信号肽。此外,该重组核酸分子任选地包含编码p60或其片段的第三多核苷酸。在另一个实施例中,该信号肽为Tat信号肽,例如枯草芽孢杆菌(B.subtilis)Tat信号肽(例如,PhoD)。在一个实施例中,该信号肽处于编码重组多肽的相同翻译阅读框中。
在另一个实施例中,本发明提供了编码本发明的信号肽或重组多肽的分离的核酸。在一个实施例中,该分离的核酸包含与编码本发明的信号肽或重组多肽的核酸共有至少65%同源性的序列。在另一个实施例中,该分离的核酸包含与编码本发明的信号肽或重组多肽的核酸共有至少75%同源性的序列。在另一个实施例中,该分离的核酸包含与编码本发明的信号肽或重组多肽的核酸共有至少85%同源性的序列。在另一个实施例中,该分离的核酸包含与编码本发明的信号肽或重组多肽的核酸共有至少90%同源性的序列。在另一个实施例中,该分离的核酸包含与编码本发明的信号肽或重组多肽的核酸共有至少95%同源性的序列。在另一个实施例中,该分离的核酸包含与编码本发明的信号肽或重组多肽的核酸共有至少97%同源性的序列。在另一个实施例中,该分离的核酸包含与编码本发明的信号肽或重组多肽的核酸共有至少99%同源性的序列。
在一个实施例中,本发明提供了包含编码本发明的多肽的寡核苷酸的载体。在一个实施例中,术语“寡核苷酸”指短的核酸聚合物,通常具有二十个或更少的碱基。在一个实施例中,本发明提供了包含编码本发明的多肽的多核苷酸的载体。在一个实施例中,术语“多核苷酸”指多个核苷酸的链,其在一个实施例中多于5个,在另一个实施例中多于10个,在另一个实施例中多于20个,在另一个实施例中多于50个核苷酸。在一个实施例中,寡核苷酸或多核苷酸或核酸可以指原核序列、真核mRNA、来自真核mRNA的cDNA、来自真核(例如哺乳动物的)DNA的基因组DNA序列或合成的DNA序列。该术语还指包括任何已知的DNA和RNA碱基类似物的序列。
在一个实施例中,本发明提供了李斯特菌,其在一个实施例中为包含本发明的分离的核酸或载体的李斯特菌疫苗菌株。
在一个实施例中,本文提供的重组多肽或融合蛋白由本文提供的李斯特菌菌株表达。在另一个实施例中,重组多肽或融合蛋白从存在于所述李斯特菌菌株内的质粒表达。在另一个实施例中,该重组多肽从所述李斯特菌的染色体表达。在一个实施例中,该重组多肽包含本文提供的融合蛋白。在另一个实施例中,该重组多肽为本文提供的融合蛋白。
在另一个实施例中,包含在本文提供的组合物中的活的减毒李斯特菌菌株包含重组核酸序列,该重组核酸序列包含第一和第二开放阅读框,每个编码第一和第二多肽,其中该第一和第二多肽每个包含融合至含PEST的多肽的异源抗原或其片段。
在一个实施例中,本文提供了包含游离型重组核酸分子的重组李斯特菌菌株,该核酸分子包含第一和第二开放阅读框,每个编码第一和第二多肽,其中该第一和第二多肽每个包含融合至含PEST的多肽的异源抗原或其片段,并且其中该核酸还包含编码代谢酶的开放阅读框。在一个实施例中,术语“游离型的”或“游离体”指存在于宿主细胞(例如李斯特菌)中的质粒。
在另一个实施例中,该李斯特菌菌株包含含有1至5个开放阅读框的重组核苷酸,每个开放阅读框编码融合至含PEST的多肽的异源抗原或其片段。在一个实施例中,本文提供的异源抗原或其片段和含PEST的多肽在单个开放阅读框中翻译。在另一个实施例中,本文提供的每个异源抗原性多肽和含PEST的多肽在分开翻译后融合。
在一个实施例中,包含重组李斯特菌菌株的组合物包含1至5个重组核酸分子,每个编码融合至含PEST的多肽的异源抗原或其片段。
在另一个实施例中,含PEST的多肽为N末端截短的LLO多肽、N末端ActA多肽或PEST肽或它们的片段。在另一个实施例中,该片段为功能性片段。在另一个实施例中,该片段为免疫原性片段。
在一个实施例中,本文提供的核酸分子包含编码异源抗原的第一开放阅读框。在另一个实施例中,本文提供的核酸分子还包含编码代谢酶的第二开放阅读框。在另一个实施例中,该代谢酶补充重组李斯特菌菌株的染色体中缺乏的内源基因。在另一个实施例中,由该第二开放阅读框编码的代谢酶为丙氨酸消旋酶(dal)。在另一个实施例中,由该第二开放阅读框编码的代谢酶为D-氨基酸转移酶(dat)。在一个实施例中,该李斯特菌还包含编码另外代谢酶的第三开放阅读框。在另一个实施例中,由该第三开放阅读框编码的代谢酶为D-氨基酸转移酶。在另一个实施例中,所述核酸分子包含编码异源抗原或其片段的第四开放阅读框。在另一个实施例中,本文提供的重组李斯特菌在基因组dal/dat基因中包含突变或缺失。在另一个实施例中,重组李斯特菌菌株缺乏dal/dat基因。在另一个实施例中,该dal/dat基因在本文提供的重组李斯特菌中失活。在一个实施例中,当指基因组毒力基因时,术语“缺乏”指该毒力基因或者是突变的,或者说是未从染色体有功能地表达。该术语还可以涵盖该毒力基因在染色体中的部分缺失或全部基因缺失。
在另一个实施例中,本发明的方法和组合物的核酸分子可操作地连接至启动子/调控序列。在另一个实施例中,本发明的方法和组合物的第一开放阅读框可操作地连接至启动子/调控序列。在另一个实施例中,本发明的方法和组合物的第二开放阅读框可操作地连接至启动子/调控序列。在另一个实施例中,每一开放阅读框可操作地连接至启动子/调控序列。每种可能性代表了本发明的独立实施例。
“代谢酶”在另一个实施例中指参与宿主细菌所需的营养物的合成的酶。在另一个实施例中,该术语指宿主细菌所需的营养物的合成所需的酶。在另一个实施例中,该术语指参与宿主细菌所利用的营养物的合成的酶。在另一个实施例中,该术语指参与宿主细菌持续生长所需的营养物的合成的酶。在另一个实施例中,该酶为营养物的合成所需。每种可能性代表了本发明的独立实施例。
在另一个实施例中,所述重组李斯特菌为减毒的营养缺陷型菌株。
在一个实施例中,减毒的李斯特菌菌株为Lm dal(-)dat(-)(Lmdd)。在另一个实施例中,该减毒的菌株为Lm dal(-)dat(-)ΔactA(LmddA)。LmddA基于李斯特菌疫苗载体,该载体因缺失毒力基因actA而为减毒的并保留了质粒,以通过补充dal基因用于期望的异源抗原或截短的LLO在体内和体外表达。在另一个实施例中,该减毒菌株为LmΔactA。在另一个实施例中,该减毒菌株为LmΔPrfA。在另一个实施例中,该减毒菌株为LmΔPlcB。在另一个实施例中,该减毒菌株为LmΔPlcA。在另一个实施例中,该菌株为任何上文提及的菌株的双突变体或三突变体。在另一个实施例中,这种菌株发挥强的佐剂效应,这是基于李斯特菌的疫苗的固有特性。在另一个实施例中,这种菌株从EGD李斯特菌骨架构建。在另一个实施例中,用在本发明中的菌株为表达非溶血性LLO的李斯特菌菌株。
在另一个实施例中,本文提供的李斯特菌菌株为营养缺陷型突变体。在另一个实施例中,该李斯特菌菌株在编码维生素合成基因的基因中是有缺陷的。在另一个实施例中,该李斯特菌菌株在编码泛酸合酶的基因中是有缺陷的。
在另一个实施例中,本文提供的李斯特菌菌株在氨基酸代谢酶中是有缺陷的。在另一个实施例中,该李斯特菌菌株在D-谷氨酸合酶基因中是有缺陷的。在另一个实施例中,该李斯特菌菌株在dat基因中是有缺陷的。在另一个实施例中,该李斯特菌菌株在dal基因中是有缺陷的。在另一个实施例中,该李斯特菌菌株在dga基因中是有缺陷的。在另一个实施例中,该李斯特菌菌株在参与合成二氨基庚二酸的基因中是有缺陷的。CysK.在另一个实施例中,该基因为与维生素-B12无关的甲硫氨酸合酶。在另一个实施例中,该基因为trpA。在另一个实施例中,该基因为trpB。在另一个实施例中,该基因为trpE。在另一个实施例中,该基因为asnB。在另一个实施例中,该基因为gltD。在另一个实施例中,该基因为gltB。在另一个实施例中,该基因为leuA。在另一个实施例中,该基因为argG。在另一个实施例中,该基因为thrC。在另一个实施例中,该李斯特菌菌株在一个或多个上文描述的基因中是有缺陷的。
在另一个实施例中,本文提供的李斯特菌菌株在合酶基因中是有缺陷的。在另一个实施例中,该基因为氨基酸合成基因。在另一个实施例中,该基因为folP。在另一个实施例中,该基因为二氢尿苷合酶家族蛋白。在另一个实施例中,该基因为ispD。在另一个实施例中,该基因为ispF。在另一个实施例中,该基因为磷酸烯醇丙酮酸合酶。在另一个实施例中,该基因为hisF。在另一个实施例中,该基因为hisH。在另一个实施例中,该基因为fliI。在另一个实施例中,该基因为核糖体大亚基假尿苷合酶。在另一个实施例中,该基因为ispD。在另一个实施例中,该基因为双功能GMP合酶/谷氨酰胺氨基转移酶蛋白。在另一个实施例中,该基因为cobS。在另一个实施例中,该基因为cobB。在另一个实施例中,该基因为cbiD。在另一个实施例中,该基因为尿卟啉-III C-甲基转移酶/尿卟啉-III合酶。在另一个实施例中,该基因为cobQ。在另一个实施例中,该基因为uppS。在另一个实施例中,该基因为truB。在另一个实施例中,该基因为dxs。在另一个实施例中,该基因为mvaS。在另一个实施例中,该基因为dapA。在另一个实施例中,该基因为ispG。在另一个实施例中,该基因为folC。在另一个实施例中,该基因为柠檬酸合酶。在另一个实施例中,该基因为argJ。在另一个实施例中,该基因为3-脱氧-D-阿拉伯-庚酮糖-7-磷酸合酶。在另一个实施例中,该基因为吲哚-3-甘油-磷酸合酶。在另一个实施例中,该基因为邻氨基苯甲酸合酶/谷氨酰胺氨基转移酶组分。在另一个实施例中,该基因为menB。在另一个实施例中,该基因为甲基萘醌特异的异分支酸合酶。在另一个实施例中,该基因为磷酸核糖甲酰甘氨脒合酶I或II。在另一个实施例中,该基因为磷酸核糖氨基咪唑-琥珀酸甲酰胺合酶。在另一个实施例中,该基因为carB。在另一个实施例中,该基因为carA。在另一个实施例中,该基因为thyA。在另一个实施例中,该基因为mgsA。在另一个实施例中,该基因为aroB。在另一个实施例中,该基因为hepB。在另一个实施例中,该基因为rluB。在另一个实施例中,该基因为ilvB。在另一个实施例中,该基因为ilvN。在另一个实施例中,该基因为alsS。在另一个实施例中,该基因为fabF。在另一个实施例中,该基因为fabH。在另一个实施例中,该基因为假尿苷合酶。在另一个实施例中,该基因为pyrG。在另一个实施例中,该基因为truA。在另一个实施例中,该基因为pabB。在另一个实施例中,该基因为atp合酶基因(例如,atpC、atpD-2、aptG、atpA-2等)。
在另一个实施例中,该基因为phoP。在另一个实施例中,该基因为aroA。在另一个实施例中,该基因为aroC。在另一个实施例中,该基因为aroD。在另一个实施例中,该基因为plcB。
在另一个实施例中,所述李斯特菌菌株在肽转运蛋白中是有缺陷的。在另一个实施例中,该基因为ABC转运蛋白/ATP结合/通透酶蛋白。在另一个实施例中,该基因为寡肽ABC转运蛋白/寡肽结合蛋白。在另一个实施例中,该基因为寡肽ABC转运蛋白/通透酶蛋白。在另一个实施例中,该基因为锌ABC转运蛋白/锌结合蛋白。在另一个实施例中,该基因为糖ABC转运蛋白。在另一个实施例中,该基因为磷酸转运蛋白。在另一个实施例中,该基因为ZIP锌转运蛋白。在另一个实施例中,该基因为EmrB/QacA家族的耐药转运蛋白。在另一个实施例中,该基因为硫酸转运蛋白。在另一个实施例中,该基因为质子依赖性寡肽转运蛋白。在另一个实施例中,该基因为镁转运蛋白。在另一个实施例中,该基因为甲酸/硝酸转运蛋白。在另一个实施例中,该基因为亚精胺/腐胺ABC转运蛋白。在另一个实施例中,该基因为Na/Pi-协同转运蛋白。在另一个实施例中,该基因为磷酸糖转运蛋白。在另一个实施例中,该基因为谷氨酰胺ABC转运蛋白。在另一个实施例中,该基因为主要协助家族转运蛋白。在另一个实施例中,该基因为甘氨酸甜菜碱/L-脯氨酸ABC转运蛋白。在另一个实施例中,该基因为钼ABC转运蛋白。在另一个实施例中,该基因为磷壁酸ABC转运蛋白。在另一个实施例中,该基因为钴ABC转运蛋白。在另一个实施例中,该基因为铵转运蛋白。在另一个实施例中,该基因为氨基酸ABC转运蛋白。在另一个实施例中,该基因为细胞分裂ABC转运蛋白。在另一个实施例中,该基因为锰ABC转运蛋白。在另一个实施例中,该基因为铁化合物ABC转运蛋白。在另一个实施例中,该基因为麦芽糖/麦芽糖糊精ABC转运蛋白。在另一个实施例中,该基因为Bcr/CflA家族的耐药转运蛋白。在另一个实施例中,该基因为来自上述蛋白之一的亚基。
在一个实施例中,本文提供了核酸分子,该核酸分子用于转化李斯特菌以便获得重组李斯特菌。在另一个实施例中,本文提供的用于转化李斯特菌的核酸缺乏毒力基因。在另一个实施例中,该核酸分子整合进李斯特菌基因组并携带非功能性毒力基因。在另一个实施例中,该毒力基因在该重组李斯特菌中是突变的。在又另一个实施例中,该核酸分子用于让李斯特菌基因组中存在的内源基因失活。在又另一个实施例中,该毒力基因为actA基因、inlA基因、inlB基因、inlC基因、inlJ基因、plbC基因、bsh基因或prfA基因。技术人员应当理解,该毒力基因可为本领域已知的任何与重组李斯特菌中的毒力相关的基因。
在又另一个实施例中,本文提供的李斯特菌菌株为inlA突变体、inlB突变体、inlC突变体、inlJ突变体、prfA突变体、ActA突变体、prfA突变体、plcB缺失突变体或缺乏plcA和plcB的双突变体。在另一个实施例中,该李斯特菌包含这些基因的缺失或突变,无论是单独的,还是组合的。在另一个实施例中,本文提供的李斯特菌缺乏基因的每一个。在另一个实施例中,本文提供的李斯特菌缺乏至少1个且最多10个本文提供的任何基因,包括actA、prfA以及dal/dat基因。在一个实施例中,所述活的减毒李斯特菌为重组李斯特菌。在另一个实施例中,该重组李斯特菌包含基因组内化素B(inlB)基因的突变或缺失。在另一个实施例中,该重组李斯特菌包含基因组actA基因和基因组内化素B基因的突变或缺失。在另一个实施例中,该重组李斯特菌包含基因组内化素C(inlC)基因的突变或缺失。在一个实施例中,李斯特菌向邻近细胞的迁移通过参与该过程的actA基因和/或inlC基因的缺失而被抑制,由此产生出乎意料的高水平减毒,具有增加的免疫原性,并用作疫苗的骨架。
每种可能性代表了本发明的独立实施例。
在一个实施例中,所述代谢基因、毒力基因等在李斯特菌菌株的染色体中是缺乏的。在另一个实施例中,所述代谢基因、毒力基因等在李斯特菌菌株的染色体以及任何游离型遗传元件中是缺乏的。在另一个实施例中,所述代谢基因、毒力基因等在毒力菌株的染色体中是缺乏的。在另一个实施例中,该毒力基因在染色体中是突变的。在另一个实施例中,该毒力基因从染色体缺失。每种可能性代表了本发明的独立实施例。
在一个实施例中,为了选择包含编码代谢酶或补充本文提供的基因的质粒的营养缺陷型细菌,让转化的营养缺陷型细菌生长在将选择表达氨基酸代谢基因或补充基因的培养基上。在另一个实施例中,以包含用于D-谷氨酸合成的基因的质粒转化D-谷氨酸合成缺陷型细菌,该营养缺陷型细菌将在不存在D-谷氨酸情况下生长,而尚未被该质粒转化或者不表达编码用于D-谷氨酸合成的蛋白的质粒的营养缺陷型细菌将不会生长。在另一个实施例中,如果本发明的质粒包含编码用于D-丙氨酸合成的氨基酸代谢酶的分离的核酸,D-丙氨酸合成营养缺陷型的细菌当被转化且表达所述质粒时,将在不存在D-丙氨酸的情况下生长。这样的用于制备适当的培养基(其包含或缺少必需生长因子、补充物、氨基酸、维生素、抗生素等)的方法是本领域众所公知的,且可商购获得(Becton-Dickinson,Franklin Lakes,NJ)。每种方法代表了本发明的独立实施例。
在另一个实施例中,在适当的培养基上选择包含本发明的质粒的营养缺陷型细菌以后,让细菌在选择压力存在下繁殖。这样的繁殖包括在没有营养缺陷因子存在的培养基中使该细菌生长。表达氨基酸代谢酶的质粒在营养缺陷型细菌中的存在会确保,该质粒将与该细菌一起复制,从而连续地选择携带该质粒的细菌。技术人员在获知本文的公开内容和方法后将能够容易地通过调整培养基(包含质粒的营养缺陷型细菌在其中生长)的体积来放大李斯特菌疫苗载体的生产。
技术人员应当理解,在另一个实施例中,其他营养缺陷型菌株和互补系统适合与本发明一起使用。
在另一个实施例中,本文提供的构建体或核酸分子采用同源重组整合进李斯特菌染色体。用于同源重组的技术在本领域是公知的,并例如在Baloglu S,Boyle SM等人(Immune responses of mice to vaccinia virus recombinants expressing either Listeriamonocytogenes partial listeriolysin or Brucella abortus ribosomal L7/L12protein.Vet Microbiol 2005,109(1-2):11-7)和Jiang LL,Song HH等人(Characterization of a mutant Listeriamonocytogenes strain expressing green fluorescent protein.Acta Biochim Biophys Sin(Shanghai)2005,37(1):19-24)中有描述。在另一个实施例中,按美国专利No.6,855,320中的描述进行同源重组。在这种情况下,通过hly启动子控制下的E7基因的染色体整合,制备表达E7的重组Lm菌株,并且包含hly信号序列以确保基因产物的分泌,从而产生称作Lm-AZ/E7的重组体。在另一个实施例中,将温度敏感的质粒用来选择重组体。每种技术代表了本发明的独立实施例。
在另一个实施例中,所述构建体或核酸分子采用转座子插入整合进李斯特菌染色体。用于转座子插入的技术在本领域是公知的,尤其是Sun等人(Infection and Immunity 1990,58:3770-3778)在构建DP-L967中所描述的。在另一个实施例中,转座子诱变具有可形成稳定的基因组插入突变体的优点,但缺点是外源基因在基因组中的插入位置是未知的。
在另一个实施例中,使用噬菌体整合位点(Lauer P,Chow MY等人,Construction,characterization,and use of two Listeriamonocytogenes site-specific phage integration vectors.J Bacteriol 2002;184(15):4177-86)将构建体或核酸分子整合进李斯特菌染色体。在该方法的某些实施例中,使用噬菌体(例如U153或PSA李斯特噬菌体)的整合酶基因和附着位点将异源基因插入对应的附着位点中,该附着位点可以是基因组中的任何适当的位点(例如comK或arg tRNA基因的3’末端)。在另一个实施例中,内源原噬菌体在构建体或异源基因整合之前从所利用的附着位点解离。在另一个实施例中,这种方法产生单拷贝的整合体。在另一个实施例中,本发明还包括用于临床应用的基于噬菌体的染色体整合系统,其中可使用必需酶(包括但不限于d-丙氨酸消旋酶)营养缺陷的宿主菌株,例如Lmdal(-)dat(-)。在另一个实施例中,为了避免“噬菌体解离步骤”,使用基于PSA的噬菌体整合系统。这在另一个实施例中需要通过抗生素持续选择以维持整合的基因。因此,在另一个实施例中,本发明使得能建立基于噬菌体的染色体整合系统,该系统不需要以抗生素选择。作为替代,可补充营养缺陷型宿主菌株。每种可能性代表了本发明的独立实施例。
“噬菌体表达载体”或“噬粒”指用于在体外或体内在任何细胞(包括原核、酵母、真菌、植物、昆虫或哺乳动物细胞)中组成型或诱导型表达本文提供的方法和组合物的核酸序列的目的的任何基于噬菌体的重组表达系统。噬菌体表达载体通常既可在细菌细胞中繁殖,又可在适合条件下产生噬菌体颗粒。该术语还包括线状或环状表达系统,并涵盖保持游离或者整合进宿主细胞基因组的两种基于噬菌体的表达载体。
在另一个实施例中,所述构建体或核酸分子从游离型或质粒载体表达,具有编码LLO、PEST或ActA序列或其片段的内源核酸序列。在另一个实施例中,该质粒在不存在抗生素选择情况下稳定保持在重组李斯特菌疫苗菌株中。在另一个实施例中,该质粒不会赋予重组李斯特菌抗生素抗性。在另一个实施例中,该片段为功能性片段。在另一个实施例中,该片段为免疫原性片段。在另一个实施例中,所述构建体或核酸分子包含第一和至少第二开放阅读框,每个编码第一和至少第二多肽,其中该第一和至少第二多肽每个包含融合至含PEST的多肽的异源抗原或其功能性片段。
在一个实施例中,“开放阅读框”或“ORF”为生物体基因组的一部分,其含有可潜在地编码蛋白的碱基序列。在另一个实施例中,该ORF的开始和结束端不等同于mRNA的末端,但它们通常包含在该mRNA内。在一个实施例中,ORF位于基因的开始密码子序列(起始密码子)和结束密码子序列(终止密码子)之间。因此,在一个实施例中,作为带有内源多肽的开放阅读框可操作地整合进基因组的核酸分子为已整合进基因组中与内源多肽处于相同开放阅读框中的核酸分子。
在另一个实施例中,“稳定保持”指核酸分子或质粒在没有选择(例如抗生素选择)的情况下保持10代,而没有可检测的损失。在另一个实施例中,该周期为15代。在另一个实施例中,该周期为20代。在另一个实施例中,该周期为25代。在另一个实施例中,该周期为30代。在另一个实施例中,该周期为40代。在另一个实施例中,该周期为50代。在另一个实施例中,该周期为60代。在另一个实施例中,该周期为80代。在另一个实施例中,该周期为100代。在另一个实施例中,该周期为150代。在另一个实施例中,该周期为200代。在另一个实施例中,该周期为300代。在另一个实施例中,该周期为500代。在另一个实施例中,该周期为更多代。在另一个实施例中,该核酸分子或质粒在体外(例如培养中)稳定的保持。在另一个实施例中,该核酸分子或质粒在体内稳定的保持。在另一个实施例中,该核酸分子或质粒在体外和体内都稳定的保持。每种可能性代表了本发明的独立实施例。
在另一个实施例中,“功能性片段”为当向受试者单独给药或者在本文提供的疫苗或组合物中向受试者给药时能够引起免疫应答的免疫原性片段。在另一个实施例中,功能性片段具有生物学活性,如技术人员所了解的以及如本文所进一步提供的。
在其他实施例中,本文提供的抗原与以下疾病之一相关:霍乱、白喉、嗜血杆菌、甲型肝炎、乙型肝炎、流感、麻疹、脑膜炎、腮腺炎、百日咳、天花、肺炎球菌肺炎、脑灰质炎、狂犬病、风疹、破伤风、肺结核、伤寒、水痘-带状疱疹、百日咳、黄热病、来自爱迪生氏病的免疫原和抗原、过敏症、过敏反应、布鲁顿综合征、癌症、包括实体和血源性肿瘤、湿疹、桥本氏甲状腺炎、多肌炎、皮肌炎、1型糖尿病、获得性免疫缺陷综合征、移植排斥、例如肾、心脏、胰腺、肺、骨骼和肝脏移植物、格雷夫斯病、多内分泌自身免疫病、肝炎、显微镜下多动脉炎、结节性多动脉炎、天疱疮、原发性胆汁性肝硬化、恶性贫血、乳糜泻、抗体介导的肾炎、血管球性肾炎、风湿病、全身性红斑狼疮、类风湿性关节炎、血清阴性脊椎关节病、鼻炎、Sjogren综合征、全身性硬化症、硬化性胆管炎、韦格纳氏肉芽肿、疱疹样皮炎、牛皮癣、白癜风、多发性硬化症、脑脊髓炎、吉兰-巴雷综合征、重症肌无力、兰伯特-伊顿综合征、巩膜、巩膜外层、色素层炎、慢性粘膜皮肤念珠菌病、风疹、婴儿短暂性低丙种球蛋白血症、骨髓瘤、X-连锁高IgM综合征、斯科特-奥尔德里奇综合征、共济失调性毛细血管扩张症、自身免疫性溶血性贫血、自身免疫性血小板减少症、自身免疫性中性粒细胞减少症、瓦尔登斯特伦巨球蛋白血、淀粉样变性、慢性淋巴细胞白血病、非霍奇金淋巴瘤、疟疾circumsporozite蛋白、微生物抗原、病毒抗原、自身抗原以及李斯特菌病。
在一个实施例中,本文提供的疾病为感染性疾病。在一个实施例中,感染性疾病为由以下病原体的任一种但并不限于它们所引起的感染性疾病:BCG/结核病、疟疾、恶性疟原虫、三日疟原虫、间日疟原虫、轮状病毒、霍乱、白喉-破伤风、百日咳、流感嗜血杆菌、乙型肝炎、人乳头瘤病毒、季节性流感)、A型流感(H1N1)流行病、麻疹和风疹、腮腺炎、脑膜炎球菌A+C、口服脊髓灰质炎疫苗(单价、二价和三价)、肺炎球菌、狂犬病、破伤风类毒素、黄热病、炭疽芽孢杆菌(炭疽)、肉毒棱状芽孢杆菌毒素(肉毒中毒)、鼠疫耶尔森菌(瘟疫)、重型天花(天花)和其他有关的痘病毒、土拉热弗朗西丝菌(土拉菌病)、病毒性出血热、沙粒病毒(LCM、胡宁病毒、马丘波病毒、瓜纳瑞托病毒、拉沙热)、布尼亚病毒(汉坦病毒属、裂谷热)、黄病毒属(登革热)、线状病毒(埃博拉病毒、马尔堡病毒)、假鼻疽伯克霍尔德菌、伯内特考克斯体(Q热)、布鲁杆菌属种(布鲁杆菌病)、鼻疽伯克霍尔德菌(鼻疽)、鹦鹉衣原体(鹦鹉热)、蓖麻蛋白毒素(来自蓖麻)、产气荚膜梭菌的ε毒素、葡萄球菌肠毒素B、斑疹伤寒热(普氏立克次体)、其他立克次体、食品和水传播的病原体、细菌(致腹泻性的大肠杆菌、致病性的弧菌属、志贺氏菌属种、沙门氏菌属BCG/、空肠弯曲杆菌、小肠结肠炎耶尔森菌)、病毒(杯状病毒、甲型肝炎、西尼罗病毒、LaCrosse、加利福尼亚脑炎、VEE、EEE、WEE、日本脑炎病毒、科萨努尔森林病毒、尼帕病毒、汉坦病毒属、蜱传出血热病毒、切昆贡亚病毒、克里米亚-刚果出血热病毒、蜱传脑炎病毒、乙型肝炎病毒、丙型肝炎病毒、单纯疱疹病毒(HSV)、人免疫缺陷病毒(HIV)、人乳头状瘤病毒(HPV)、原生动物(小隐孢子虫、卡晏环孢子虫、兰伯贾第虫、溶组织内阿米巴、弓形虫属)、真菌(微孢子目)、黄热病、结核病(包括耐药的TB)、狂犬病、朊病毒、严重急性呼吸综合征相关的冠状病毒(SARS-CoV)、Coccidioides posadasii、粗球孢子菌、细菌性阴道病、沙眼衣原体、巨细胞病毒、腹股沟肉芽肿、杜克雷嗜血杆菌、淋病奈瑟球菌、苍白密螺旋体、阴道毛滴虫或本领域已知的在本文中没有列出的任何其他感染性疾病。
在另一个实施例中,该感染性疾病是家畜感染性疾病。在另一个实施例中,家畜疾病可传播给人,并被称作“人畜共患病”。在另一个实施例中,这些疾病包括但不限于口蹄疫、西尼罗病毒、狂犬病、犬细小病毒、猫白血病病毒、马流感病毒、传染性牛鼻气管炎(IBR)、伪狂犬病、古典猪瘟(CSF)、由牛1型牛疱疹病毒(BHV-1)感染造成的IBR以及猪伪狂犬病(奥尔斯基病)、弓形虫病、炭疽、水疱性口炎病毒、马红球菌、土拉菌病、瘟疫(鼠疫耶尔森菌)、毛滴虫。
在另一个实施例中,本文提供的疾病为呼吸道疾病或炎症性疾病。在另一个实施例中,该呼吸道疾病或炎症性疾病为慢性阻塞性肺病(COPD)。在另一个实施例中,该疾病为哮喘。
在一个实施例中,活的减毒李斯特菌菌株能够缓解哮喘症状,无需其他治疗剂的共给药,例如抗炎剂或支气管扩张剂。在一个实施例中,本文提供的方法还包括以活的减毒李斯特菌菌株和一种或多种治疗剂向受试者共给药的步骤。在另一个实施例中,该治疗剂为平喘药。在另一个实施例中,该治疗剂为抗炎剂、非甾体抗炎剂、抗生素、抗胆碱能药、支气管扩张药、皮质类固醇、短效β受体激动剂、长效β受体激动剂、组合吸入剂、抗组织胺药或它们的组合。
在一个实施例中,本文提供的疾病为癌症或肿瘤。在一个实施例中,由本发明的方法处理的癌症为乳腺癌。在另一个实施例中,该癌症为子宫颈癌。在另一个实施例中,该癌症为含HER2的癌症。在另一个实施例中,该癌症为黑素瘤。在另一个实施例中,该癌症为胰腺癌。在另一个实施例中,该癌症为卵巢癌。在另一个实施例中,该癌症为胃癌。在另一个实施例中,该癌症为胰腺的癌性病变。在另一个实施例中,该癌症为肺腺癌。在另一个实施例中,其为多形性胶质母细胞瘤。在另一个实施例中,其为间皮瘤。在另一个实施例中,该癌症为结直肠腺癌。在另一个实施例中,该癌症为肺鳞癌。在另一个实施例中,该癌症为胃腺癌。在另一个实施例中,该癌症为卵巢表面上皮肿瘤(例如其良性的、增生性的或恶性的种类)。在另一个实施例中,该癌症为口腔鳞状细胞癌。在另一个实施例中,该癌症为非小细胞肺癌。在另一个实施例中,该癌症为子宫内膜癌。在另一个实施例中,该癌症为膀胱癌。在另一个实施例中,该癌症为头颈癌。在另一个实施例中,该癌症为前列腺癌。在另一个实施例中,该癌症为口咽癌。在另一个实施例中,该癌症为肺癌。在另一个实施例中,该癌症为肛门癌。在另一个实施例中,该癌症为结肠直肠癌。在另一个实施例中,该癌症为食道癌。每种可能性代表了本发明的独立实施例。
在一个实施例中,本文提供的肿瘤标志物为异源肿瘤抗原,其在本文中也称为“肿瘤抗原、“抗原性多肽”或“抗原”。在另一个实施例中,该抗原为人类乳头瘤病毒E7(HPV-E7)抗原,其在一个实施例中来自HPV16(在一个实施例中,GenBank登录号AAD33253),在另一个实施例中来自HPV18(在一个实施例中,GenBank登录号P06788)。在另一个实施例中,该抗原性多肽为HPV-E6,其在一个实施例中来自HPV16(在一个实施例中,GenBank登录号AAD33252、AAM51854、AAM51853或AAB67615),在另一个实施例中来自HPV18(在一个实施例中,GenBank登录号P06463)。在另一个实施例中,该抗原性多肽为Her/2-neu抗原。在另一个实施例中,该抗原性多肽为前列腺特异性抗原(PSA)(在一个实施例中GenBank登录号CAD30844、CAD54617、AAA58802或NP_001639)。在另一个实施例中,该抗原性多肽为角质层胰凝乳蛋白酶(SCCE)抗原(在一个实施例中,GenBank登录号AAK69652、AAK69624、AAG33360、AAF01139或AAC37551)。在另一个实施例中,该抗原性多肽为Wilms肿瘤抗原1,其在另一个实施例中为WT-1端粒酶(GenBank登录号P49952、P22561、NP_659032、CAC39220.2或EAW68222.1)。在另一个实施例中,该抗原性多肽为hTERT或端粒酶(GenBank登录号NM003219(变体1)、NM198255(变体2)、NM 198253(变体3)或NM 198254(变体4)。在另一个实施例中,该抗原性多肽为蛋白酶3(在一个实施例中,GenBank登录号M29142、M75154、M96839、X55668、NM 00277、M96628或X56606)。在另一个实施例中,该抗原性多肽为酪氨酸酶相关蛋白2(TRP2)(在一个实施例中,GenBank登录号NP_001913、ABI73976、AAP33051或Q95119)。在另一个实施例中,该抗原性多肽为高分子量黑素瘤相关抗原(HMW-MAA)(在一个实施例中,GenBank登录号NPNP_001888、AAI28111或AAQ62842)。在另一个实施例中,该抗原性多肽为Testisin(在一个实施例中,GenBank登录号AAF79020、AAF79019、AAG02255、AAK29360、AAD41588或NP_659206)。在另一个实施例中,该抗原性多肽为NY-ESO-1抗原(在一个实施例中,GenBank登录号CAA05908、P78358、AAB49693或NP_640343)。在另一个实施例中,该抗原性多肽为PSCA(在一个实施例中,GenBank登录号AAH65183、NP_005663、NP_082492、O43653或CAB97347)。在另一个实施例中,该抗原性多肽为白细胞介素(IL)13受体α(在一个实施例中,GenBank登录号NP_000631、NP_001551、NP_032382、NP_598751、NP_001003075或NP_999506)。在另一个实施例中,该抗原性多肽为碳酸酐酶IX(CAIX)(在一个实施例中,GenBank登录号CAI13455、CAI10985、EAW58359、NP_001207、NP_647466或NP_001101426)。在另一个实施例中,该抗原性多肽为癌胚抗原(CEA)(在一个实施例中,GenBank登录号AAA66186、CAA79884、CAA66955、AAA51966、AAD15250或AAA51970)。在另一个实施例中,该抗原性多肽为MAGE-A(在一个实施例中,GenBank登录号NP_786885、NP_786884、NP_005352、NP_004979、NP_005358或NP_005353)。在另一个实施例中,该抗原性多肽为存活素(在一个实施例中,GenBank登录号AAC51660、AAY15202、ABF60110、NP_001003019或NP_001082350)。在另一个实施例中,该抗原性多肽为GP100(在一个实施例中,GenBank登录号AAC60634、YP_655861或AAB31176)。在另一个实施例中,该抗原性多肽为本领域已知的任何其他抗原性多肽。在另一个实施例中,本发明的组合物和方法的抗原性肽包含所述抗原性多肽的免疫原性部分。每种可能性代表了本发明的独立实施例。
在另一个实施例中,本文提供的抗原为HPV-E6。在另一个实施例中,该抗原为端粒酶(TERT)。在另一个实施例中,该抗原为LMP-1。在另一个实施例中,该抗原为p53。在另一个实施例中,该抗原为间皮素。在另一个实施例中,该抗原为EGFRVIII。在另一个实施例中,该抗原为碳酸酐酶IX(CAIX)。在另一个实施例中,该抗原为PSMA。在另一个实施例中,该抗原为HMW-MAA。在另一个实施例中,该抗原为HIV-1Gag。在另一个实施例中,该抗原为酪氨酸酶相关蛋白2。在另一个实施例中,该抗原选自HPV-E7、HPV-E6、Her-2、HIV-1Gag、LMP-1、p53、PSMA、癌胚抗原(CEA)、LMP-1、激肽释放酶相关肽酶3(KLK3)、KLK9、Muc、酪氨酸酶相关蛋白2、Muc1、FAP、IL-13Rα2、PSA(前列腺特异性抗原)、MAGE-1、MAGE-3、gp-100、热休克蛋白70(HSP-70)、β-HCG、EGFR-III、VEGFR2、粒细胞集落刺激因子(G-CSF)、血管生成素、促血管生成素-1、Del-1、成纤维细胞生长因子:酸性(aFGF)或碱性(bFGF)、卵泡抑素、粒细胞集落刺激因子(G-CSF)、肝细胞生长因子(HGF)/分散因子(SF)、白细胞介素-8(IL-8)、痩素、中期因子、胎盘生长因子、血小板衍生内皮细胞生长因子(PD-ECGF)、血小板衍生生长因子-BB(PDGF-BB)、多效生长因子(PTN)、颗粒蛋白前体、增殖素、转化生长因子-α(TGF-α)、转化生长因子-β(TGF-β)、肿瘤坏死因子-α(TNF-α)、血管内皮生长因子(VEGF)/血管渗透因子(VPF)、VEGFR、VEGFR2(KDR/FLK-1)或其片段、FLK-1或其表位、FLK-E1、FLK-E2、FLK-I1、内皮因子或其片段、神经菌毛素1(NRP-1)、促血管生成素-1(Ang1)、Tie2、血小板衍生生长因子(PDGF)、血小板衍生生长因子受体(PDGFR)、转化生长因子-β(TGF-β)、内皮因子、TGF-β受体、单核细胞趋化蛋白-1(MCP-1)、VE-钙粘蛋白、CD31、ephrin、ICAM-1、V-CAM-1、VAP-1、E-选择素、纤溶酶原激活物、纤溶酶原激活物抑制剂-1、一氧化氮合酶(NOS)、COX-2、AC133或Id1/Id3、促血管生成素3、促血管生成素4、促血管生成素6、CD105、EDG、HHT1、ORW、ORW1或TGFβ共受体或它们的组合。对本文提供的抗原的片段的使用也涵盖在本发明中。
在另一个实施例中,本文提供的异源抗原为肿瘤相关抗原,其在一个实施例中为以下肿瘤抗原之一:MAGE(黑素瘤相关抗原E)蛋白,例如MAGE 1、MAGE 2、MAGE 3、MAGE 4、酪氨酸酶;突变体ras蛋白;突变体p53蛋白;p97黑素瘤抗原、与晚期癌症相关的ras肽或p53肽;与子宫颈癌相关的HPV 16/18抗原、与乳腺癌相关的KLH抗原、与结肠直肠癌相关的CEA(癌胚抗原)、与黑素瘤相关的MART1抗原或与前列腺癌相关的PSA抗原。在另一个实施例中,用于本文提供的组合物和方法的抗原为黑素瘤相关抗原,其在一个实施例中为TRP-2、MAGE-1、MAGE-3、gp-100、酪氨酸酶、HSP-70、β-HCG或它们的组合。在一个实施例中,所述抗原为美国专利申请公开US2011/0142791中描述的嵌合Her2抗原,通过引用将该专利全文并入本文。
应当理解,技术人员能够将本文未提到但本领域已知的任何异源抗原用在本文提供的方法和组合物中。
在一个实施例中,抗原源自真菌病原体、细菌、寄生虫、蠕虫或病毒。在另一个实施例中,所述抗原选自破伤风类毒素、来自流感病毒的血凝素分子、白喉类毒素、HIV gp120、HIV gag蛋白、IgA蛋白酶、胰岛素肽B、马铃薯粉痂病菌(Spongospora subterranea)抗原、弧菌抗原、沙门氏菌抗原、肺炎球菌抗原、呼吸道合胞体病毒抗原、流感嗜血杆菌(Haemophilus influenza)外膜蛋白、幽门螺杆菌(Helicobacter pylori)脲酶、脑膜炎奈瑟球菌(Neisseria meningitidis)菌毛蛋白、淋病奈瑟菌(N.gonorrhoeae)菌毛蛋白、黑素瘤相关抗原(TRP-2、MAGE-1、MAGE-3、gp-100、酪氨酸酶、MART-1、HSP-70、β-HCG)、来自HPV-16、-18、-31、-33、-35或-45型人类乳头瘤病毒的人类乳头瘤病毒抗原E1和E2、肿瘤抗原CEA、突变的或其他形式的ras蛋白、突变的或其他形式的p53蛋白、Muc1、间皮素、EGFRVIII或pSA。
在一个实施例中,用在本发明的组合物和方法中的血管生成因子为VEGFR2。
在一个实施例中,血管内皮生长因子(VEGF)为重要的参与血管发生(胚胎循环系统的形成)和血管生成(从现存脉管系统生长血管)的信号蛋白。在一个实施例中,VEGF活性主要限于血管内皮的细胞,尽管其对有限数量的其他细胞类型有作用(例如刺激单核细胞/巨噬细胞迁移)。VEGF在体外已显示出刺激内皮细胞有丝分裂和细胞迁移。VEGF还增强微血管渗透性,并且有时也被称为血管通透性因子。
在一个实施例中,VEGF家族的所有成员都刺激细胞应答,这通过结合细胞表面上的酪氨酸激酶受体(VEGFR),导致它们二聚化并经由转磷酸作用而被活化来实现。该VEGF受体具有由7个免疫球蛋白样结构域组成的胞外部分、单个跨膜区和含有分开的酪氨酸激酶结构域的胞内部分。
在一个实施例中,VEGF-A为VEGFR-2(KDR/Flk-1)配体以及VEGFR-1(Flt-1)配体。在一个实施例中,VEGFR介导几乎全部已知的对VEGF的细胞应答。VEGFR-1的功能不太明确,尽管在一个实施例中认为其通过隔离VEGF与VEGFR-2结合而调节VEGFR-2信号转导,在一个实施例中,这在胚胎的血管发生期间是尤其重要的。在一个实施例中,VEGF-C和VEGF-D为VEGFR-3受体的配体,该VEGFR-3受体在一个实施例中介导淋巴管生成。
在一个实施例中,本发明的重组李斯特菌表达VEGF受体或其片段,其在一个实施例中为VEGFR-2,在另一个实施例中为VEGFR-1,以及在另一个实施例中为VEGFR-3。
在一个实施例中,血管内皮生长因子受体2(VEGFR2)在活化的内皮细胞(EC)上高表达并参与新血管的形成。在一个实施例中,VEGFR2结合VEGF的全部5个同种型。在一个实施例中,VEGF经由VEGFR2在EC上的信号转导诱导增殖、迁移和最终的分化。在一个实施例中,VEGFR2的小鼠同源物为胎儿肝激酶基因-1(Flk-1),其为强治疗靶标,并在肿瘤生长、入侵和转移中起重要作用。在一个实施例中,VEGFR2也称为激酶插入区受体(III型受体酪氨酸激酶)(KDR)、分化群309(CD309)、FLK1、Ly73、Krd-1、VEGFR、VEGFR-2或6130401C07。
在另一个实施例中,用在本发明的组合物中的VEGFR2蛋白具有以下序列:
(GenBank登录号NP_034742.2、AAH20530.1或EDL37891.1;SEQ ID NO:137;GenBank登录号NM_010612.2或BC020530.1列出了核酸序列)。在一个实施例中,氨基酸68-277对应本文描述的E1,氨基酸545-730对应本文描述的E2,以及氨基酸792-1081对应本文描述的I1。在另一个实施例中,将以上序列用作并入本发明的疫苗中的VEGFR2片段的来源。在另一个实施例中,本发明的方法和组合物的VEGFR2氨基酸序列为SEQ ID NO:137的同源物。在另一个实施例中,该VEGFR2氨基酸序列为SEQ ID NO:137的变体。在另一个实施例中,该VEGFR2氨基酸序列为SEQ ID NO:137的片段。在另一个实施例中,该VEGFR2氨基酸序列为SEQ ID NO:137的同种型。每种可能性代表了本发明的独立实施例。
在另一个实施例中,该VEGFR2具有以下GenBank条目之一中列出的氨基酸序列:EDL37891.1、CAA61917.1、BAC27532.1、BAE24892.1、AAH20530.1、AAB25043.1、CAA42040.1或CAA50192.1。在另一个实施例中,该VEGFR2具有以下GenBank条目之一中列出的氨基酸序列:EAX05462.1、EAX05463.1、EAX05464.1、CAA61916.1、BAD93138.1、AAB88005.1、AAC16450.1、BAG57114.1、AAI31823.1、ACF47599.1、AAA59459.1或CAA43837.1。在另一个实施例中,该VEGFR2具有以下GenBank条目之一中列出的氨基酸序列:EDL89914.1、EDL89915.1、EDL89916.1、AAH87029.1、AAB97508.1或AAB97509.1。在另一个实施例中,该VEGFR2具有以下GenBank条目之一中列出的氨基酸序列:CAQ13438.1、AAF03237.1、AAN47136.1、AAL16381.1、AAI29159.1、CAM73177.1、AAB18415.1、AAB41042.1或AAB62405.1。在另一个实施例中,该VEGFR2具有本领域已知的任何VEGFR2氨基酸序列。在另一个实施例中,该VEGFR2为来自上述GenBank条目之一的序列的同源物。在另一个实施例中,该VEGFR2为来自上述GenBank条目之一的序列的变体。在另一个实施例中,该VEGFR2为来自上述GenBank条目之一的序列的同种型。在另一个实施例中,该VEGFR2为来自上述GenBank条目之一的序列的片段。每种可能性代表了本发明的独立实施例。
在另一个实施例中,该VEGFR2具有以下GenBank条目之一中列出的核酸序列:AC124615.11、AC134903.4、AC160723.2、AF061804.1、AF153058.1、CH466524.1、X89777.1、AK031739.1、AK054510.1、AK141938.1、BC020530.1、S53103.1、X59397.1或X70842.1。在另一个实施例中,该VEGFR2具有以下GenBank条目之一中列出的核酸序列:AC021220.7、AC111194.4、CH471057.1、EAX05463.1、EAX05464.1、X89776.1、AB209901.1、AF035121.1、AF063658.1、AK293668.1、BC131822.1、BP280621.1、CR606055.1、EU826563.1、L04947.1或X61656.1。在另一个实施例中,该VEGFR2具有以下GenBank条目之一中列出的核酸序列:CH473981.1、BC087029.1、U93306.1或U93307.1。在另一个实施例中,该VEGFR2具有以下GenBank条目之一中列出的核酸序列:AL935131.7、BX247946.6、CR759732.9、AF180354.1、AF487829.1、AY056466.1、BC129158.1、CU458916.1、U75995.1、U82383.1、U89515.1。在另一个实施例中,该VEGFR2具有任何本领域已知的VEGFR2核酸序列。在另一个实施例中,该VEGFR2为来自上述GenBank条目之一的序列的同源物。在另一个实施例中,该VEGFR2为来自上述GenBank条目之一的序列的变体。在另一个实施例中,该VEGFR2为来自上述GenBank条目之一的序列的同种型。在另一个实施例中,该VEGFR2为来自上述GenBank条目之一的序列的片段。每种可能性代表了本发明的独立实施例。
在另一个实施例中,将VEGFR2多肽片段用在本发明的组合物和方法中。在另一个实施例中,该VEGFR2片段包含VEGFR2蛋白的氨基酸68-277,其在一个实施例中称为Flk1-E1。在另一个实施例中,该VEGFR2多肽片段具有以下序列:
RDSEERVLVTECGGGDSIFCKTLTIPRVVGNDTGAYKCSYRDVDIASTVYVYVRDYRSPFIASVSDQHGIVYITENKNKTVVIPCRGSISNLNVSLCARYPEKRFVPDGNRISWDSEIGFTLPSYMISYAGMVFCEAKINDETYQSIMYIVVVVGYRIYDVILSPPHEIELSAGEKLVLNCTARTELNVGLDFTWHSPPSKSHHKKIVNR(SEQ ID NO:138)。在另一个实施例中,本发明的方法和组合物的VEGFR2氨基酸序列包含SEQ ID NO:138中列出的序列。在另一个实施例中,该VEGFR2氨基酸序列为SEQ ID NO:138的同源物。在另一个实施例中,该VEGFR2氨基酸序列为SEQ ID NO:138的变体。在另一个实施例中,该VEGFR2氨基酸序列为SEQ ID NO:138的片段。在另一个实施例中,该VEGFR2氨基酸序列为SEQ ID NO:138中列出的同种型。每种可能性代表了本发明的独立实施例。
在另一个实施例中,该VEGFR2片段包含VEGFR2蛋白的氨基酸545-730,其在一个实施例中称为Flk1-E2。在另一个实施例中,该VEGFR2多肽片段具有以下序列:
VIRGPEITVQPAAQPTEQESVSLLCTADRNTFENLTWYKLGSQATSVHMGESLTPVCKNLDALWKLNGTMFSNSTNDILIVAFQNASLQDQGDYVCSAQDKKTKKRHCLVKQLIILERMAPMITGNLENQTTTIGETIEVTCPASGNPTPHITWFKDNETLVEDSGIVLRDGNRNLTIRRVRKEDG(SEQ ID NO:139)。在另一个实施例中,本发明的方法和组合物的VEGFR2氨基酸序列包含SEQ ID NO:139中列出的序列。在另一个实施例中,该VEGFR2氨基酸序列为SEQ ID NO:139的同源物。在另一个实施例中,该VEGFR2氨基酸序列为SEQ ID NO:139的变体。在另一个实施例中,该VEGFR2氨基酸序列为SEQ ID NO:139的片段。在另一个实施例中,该VEGFR2氨基酸序列为SEQ ID NO:139的同种型。每种可能性代表了本发明的独立实施例。
在另一个实施例中,该VEGFR2片段包含VEGFR2蛋白的氨基酸792-1081,其在一个实施例中称为Flk1-I1。在另一个实施例中,该VEGFR2多肽片段具有以下序列:
EGELKTGYLSIVMDPDELPLDERCERLPYDASKWEFPRDRLKLGKPLGRGAFGQVIEADAFGIDKTATCKTVAVKMLKEGATHSEHRALMSELKILIHIGHHLNVVNLLGACTKPGGPLMVIVEFCKFGNLSTYLRGKRNEFVPYKSKGARFRQGKDYVGELSVDLKRRLDSITSSQSSASSGFVEEKSLSDVEEEEASEELYKDFLTLEHLICYSFQVAKGMEFLASRKCIHRDLAARNILLSEKNVVKICDFGLARDIYKDPDYVRKGDARLPLKWMAPETIFDRVYT(SEQ ID NO:140)。在另一个实施例中,本发明的方法和组合物的VEGFR2氨基酸序列包含SEQ ID NO:140中列出的序列。在另一个实施例中,该VEGFR2氨基酸序列为SEQ ID NO:140的同源物。在另一个实施例中,该VEGFR2氨基酸序列为SEQ ID NO:140的变体。在另一个实施例中,该VEGFR2氨基酸序列为SEQ ID NO:140的片段。在另一个实施例中,该VEGFR2氨基酸序列为SEQ ID NO:140的同种型。每种可能性代表了本发明的独立实施例。
在另一个实施例中,该VEGFR2片段包含VEGFR2蛋白的氨基酸1082-1237,其在一个实施例中称为Flk1-I2。在另一个实施例中,该VEGFR2多肽片段具有以下序列:
IQSDVWSFGVLLWEIFSLGASPYPGVKIDEEFCRRLKEGTRMRAPDYTTPEMYQTMLDCWHEDPNQRPSFSELVEHLGNLLQANAQQDGKDYIVLPMSETLSMEEDSGLSLPTSPVSCMEEEEVCDPKFHYDNTAGISHYLQNSKRKSRPVSVKTF(SEQ ID NO:141)。在另一个实施例中,本发明的方法和组合物的VEGFR2氨基酸序列包含SEQ ID NO:141中列出的序列。在另一个实施例中,该VEGFR2氨基酸序列为SEQ ID NO:141的同源物。在另一个实施例中,该VEGFR2氨基酸序列为SEQ ID NO:141的变体。在另一个实施例中,该VEGFR2氨基酸序列为SEQ ID NO:141的片段。在另一个实施例中,该VEGFR2氨基酸序列为SEQ ID NO:141的同种型。每种可能性代表了本发明的独立实施例。
在另一个实施例中,用在本发明的组合物和方法中的VEGFR2片段基于分析VEGFR2氨基酸序列含有T细胞表位的区域,在一个实施例中,这通过让VEGFR2序列运行穿过表位预测程序(其中几个在本领域是已知的)来确定,在另一个实施例中,这通过预测性表位作图来确定。在另一个实施例中,通过采用与其他物种(在一个实施例中,小鼠或大鼠)中已知包含T细胞表位的VEGFR2序列同源的人序列来使用VEGFR2片段。在另一个实施例中,用在本发明的组合物和方法中的VEGFR2片段基于本领域中对含有T细胞表位的VEGFR2区域的了解。
在一个实施例中,用在本发明的组合物和方法中的人HLA-A0201片段包含VEGFR2蛋白的氨基酸766-774。在另一个实施例中,该VEGFR2多肽片段包含序列IILVGTAVI(SEQ ID NO:142)。在另一个实施例中,用在本发明的组合物和方法中的人HLA-A0201片段包含VEGFR2蛋白的氨基酸781-789。在另一个实施例中,该VEGFR2多肽片段包含序列LLVIILRTV(SEQ ID NO:143)。在另一个实施例中,用在本发明的组合物和方法中的人HLA-A0201片段包含VEGFR2蛋白的氨基酸1034-1042。在另一个实施例中,该VEGFR2多肽片段包含序列ILLSEKNVV(SEQ ID NO:144)。在另一个实施例中,用在本发明的组合物和方法中的人HLA-A0201片段包含VEGFR2蛋白的氨基酸1076-1084。在另一个实施例中,该VEGFR2多肽片段包含序列TIFDRVYTI(SEQ ID NO:145)。在另一个实施例中,用在本发明的组合物和方法中的人HLA-A0201片段包含VEGFR2蛋白的氨基酸1093-1101。在另一个实施例中,该VEGFR2多肽片段包含序列VLLWEIFSL(SEQ ID NO:146)。
在一个实施例中,内皮因子蛋白在以下序列中列出:
MDRGVLPLPITLLLFEIYSFEPTTGLAERVGCDLQPVDPTRGEVTFTTSQVSEGCVAQAANAVREVHVLFLDFPGMLSHLELTLQASKQNGTETREVFLVLVSNKNVFVKFQAPEIPLHLAYDSSLVIFQGQPRVNITVLPSLTSRKQILDWAATKGAITSIAALDDPQSIVLQLGQDPKAPFLCLPEAHKDMGATLEWQPRAQTPVQSCRLEGVSGHKEAYILRILPGSEAGPRTVTVMMELSCTSGDAILILHGPPYVSWFIDINHSMQILTTGEYSVKIFPGSKVKGVELPDTPQGLIAEARKLNASIVTSFVELPLVSNVSLRASSCGGVFQTTPAPVVTTPPKDTCSPVLLMSLIQPKCGNQVMTLALNKKHVQTLQCTITGLTFWDSSCQAEDTDDHLVLSSAYSSCGMKVTAHVVSNEVIISFPSGSPPLRKKVQCIDMDSLSFQLGLYLSPHFLQASNTIELGQQAFVQVSVSPLTSEVTVQLDSCHLDLGPEGDMVELIQSRTAKGSCVTLLSPSPEGDPRFSFLLRVYMVPTPTAGTLSCNLALRPSTLSQEVYKTVSMRLNVVSPDLSGKGLVLPSVLGITFGAFLIGALLTAALWYIYSHTRGPSKREPVVAVAAPASSESSSTNHSIGSTQSTPCSTSSMA(SEQ ID NO:147;图60)。在一个实施例中,该内皮因子为本领域中可得到的任何内皮因子,其包括但不限于以下登录号:CAA54917.1、NP_001010968.1、NP_001074356.1、AAC63386.1、CAA50891。在另一个实施例中,氨基酸17-319对应构建体CD105A。在另一个实施例中,氨基酸359-599对应构建体CD105B。
基于李斯特菌的疫苗可以含有活的减毒李斯特菌菌株的混合物和共给药的治疗剂二者。该活的减毒李斯特菌菌株和共给药的治疗剂也可以处于不同的药物组合物中。
在一个实施例中,该治疗剂包括吸入性皮质类固醇,其包括氟地松(Flovent Diskus、Flovent HFA)、布地奈德(Pulmicort Flexhaler)、莫米松(Asmanex)、氟尼缩松(Aerobid)、倍氯米松(Qvar)等。它们为处方中最常见类型的长期哮喘药。与口服皮质类固醇不同,这些皮质类固醇药物具有相对较低的副作用,通常对长期使用是安全的。
该治疗剂可为白三烯调节剂。这些口服药物包括孟鲁斯特(Singulair)、扎鲁司特(Accolate)和齐留通(Zyflo,Zyflo CR)。它们帮助预防哮喘症状长达24小时。
此外,该治疗剂可为长效β激动剂(LABA)。这些吸入性药物包括沙美特罗(Serevent Diskus)和福莫特罗(Foradil Aerolizer)。LABA开放气道并减轻炎症。但是,它们与严重哮喘发作相关联。LABA应该仅与吸入性皮质类固醇联合使用。
在一个实施例中,本文提供的组合物或组合物的混合物包含佐剂。在另一个实施例中,该佐剂为粒细胞/巨噬细胞集落刺激因子(GM-CSF)蛋白。在另一个实施例中,该佐剂包含GM-CSF蛋白。在另一个实施例中,该佐剂为编码GM-CSF的核苷酸分子。在另一个实施例中,该佐剂为包含编码GM-CSF的核苷酸分子。在另一个实施例中,该佐剂为皂苷QS21。在另一个实施例中,该佐剂包含皂苷QS21。在另一个实施例中,该佐剂为单磷酰脂质A。在另一个实施例中,该佐剂包含单磷酰脂质A。在另一个实施例中,该佐剂为SBAS2。在另一个实施例中,该佐剂包含SBAS2。在另一个实施例中,该佐剂为含有未甲基化的CpG的寡核苷酸。在另一个实施例中,该佐剂包含含有未甲基化的CpG的寡核苷酸。在另一个实施例中,该佐剂为免疫刺激性细胞因子。在另一个实施例中,该佐剂包含免疫刺激性细胞因子。在另一个实施例中,该佐剂为编码免疫刺激性细胞因子的核苷酸分子。在另一个实施例中,该佐剂包含编码免疫刺激性细胞因子的核苷酸分子。在另一个实施例中,该佐剂为或包含刺糖苷(quill glycoside)。在另一个实施例中,该佐剂为或包含细菌有丝分裂原。在另一个实施例中,该佐剂为或包含细菌毒素。在另一个实施例中,该佐剂为或包含本领域已知的任何其他佐剂。每种可能性代表了本发明的独立实施例。
在一个实施例中,本文提供的组合物或组合物的混合物包含另外的活性剂。在一个实施例中,所述另外的活性剂包含溶瘤病毒。在另一个实施例中,该另外的活性剂包含T细胞受体工程化T细胞(受体工程化T细胞)。在另一个实施例中,该另外的活性剂包含嵌合抗原受体工程化细胞(CAR T细胞)。在另一个实施例中,该另外的活性剂包含治疗性或免疫调节性单克隆抗体。在另一个实施例中,该另外的活性剂包含靶向性胸苷激酶抑制剂(TKI)。在另一个实施例中,该另外的活性剂包含掺有工程化T细胞受体的过继转移细胞。在另一个实施例中,本发明的另外的活性剂包含减毒溶瘤病毒、T细胞受体工程化T细胞(受体工程化T细胞)、嵌合抗原受体工程化T细胞(CAR T细胞)、治疗性或免疫调节性单克隆抗体、靶向性胸苷激酶抑制剂(TKI)或掺有工程化T细胞受体的过继转移细胞或它们的任何组合。
在另一个实施例中,本文提供的组合物或组合物的混合物包含另外的活性剂。在另一个实施例中,该活性剂为免疫检查点抑制剂。
在一个实施例中,该免疫检查点蛋白抑制剂为程序性死亡1(PD-1)信号通路抑制剂。在另一个实施例中,该PD-1信号通路抑制剂为阻断PD-1受体与PD-1配体1(PD-L1)和PD-1配体2(PD-L2)相互作用的分子。在另一个实施例中,PD-L1也称为CD274或B7-H1。在另一个实施例中,PD-L2也称为CD273或B7-DC。在另一个实施例中,该阻断PD-1受体与PD-1配体1(PD-L1)和PD-1配体2(PD-L2)相互作用的分子为与PD-1、PD-L1或PD-L2相互作用的分子。在另一个实施例中,该阻断PD-1受体与PD-1配体1(PD-L1)或PD-1配体2(PD-L2)相互作用的分子为与PD-1、PD-L1或PD-L2相互作用的分子。术语“相互作用”或其语法上的等同物可以涵盖结合或接触另一分子。在另一个实施例中,该分子结合PD-1。在另一个实施例中,该PD-1信号通路抑制剂为抗PD1抗体。在另一个实施例中,与PD-L2相互作用的分子为抗PD-L1抗体,或者结合PD-L1的小分子。在另一个实施例中,与PD-L2相互作用的分子为抗PD-L2抗体,或者结合PD-L2的小分子。
在一个实施例中,与PD-1相互作用的该分子为截短的PD-L1蛋白。在另一个实施例中,该截短的PD-L1蛋白包含PD-L1蛋白的细胞质结构域。在另一个实施例中,与PD-1相互作用的该分子为截短的PD-L2蛋白。在另一个实施例中,该截短的PD-L2蛋白包含PD-L2蛋白的细胞质结构域。在另一个实施例中,该阻断PD-1受体与PD-1配体1(PD-L1)和PD-1配体2(PD-L2)相互作用的分子为与PD-L1和PD-L2相互作用的分子。在另一个实施例中,与PD-L1或PD-L2相互作用的该分子为截短的PD-1蛋白、PD-1模拟物或结合PD-L1或PD-L2的小分子。在另一个实施例中,该截短的PD-1蛋白包含PD-L1蛋白的细胞质结构域。
在一个实施例中,该免疫检查点抑制剂为CD80/86信号通路抑制剂。在另一个实施例中,CD80也称为B7.1。在另一个实施例中,CD86也称为B7.2。在另一个实施例中,该CD80信号通路抑制剂为与CD80相互作用的小分子。在另一个实施例中,该CD80抑制剂为抗CD80抗体。在另一个实施例中,该CD86信号通路抑制剂为与CD86相互作用的小分子。在另一个实施例中,该CD86抑制剂为抗CD86抗体。
在一个实施例中,该免疫检查点抑制剂为CTLA-4信号通路抑制剂。在另一个实施例中,CTLA-4也称为CD152。在另一个实施例中,该CTLA-4信号通路抑制剂为与CTLA-4相互作用的小分子。在另一个实施例中,该CTLA-4抑制剂为抗CTLA-4抗体。在另一个实施例中,该免疫检查点抑制剂为CD40信号通路抑制剂。在另一个实施例中,该免疫检查点抑制剂为本领域已知的任何其他抗原呈递细胞:T细胞信号通路抑制剂。
技术人员应当理解,本领域任何已知的免疫检查点蛋白都可被免疫检查点抑制剂靶向。免疫检查点蛋白可以选自但不限于以下:程序性细胞死亡蛋白1(PD1)、T细胞膜蛋白3(TIM3)、腺苷A2a受体(A2aR)和淋巴细胞活化基因3(LAG3)、杀伤细胞免疫球蛋白受体(KIR)或细胞毒性T淋巴细胞抗原4(CTLA-4)。在另一个实施例中,该检查点抑制剂蛋白属于B7/CD28受体超家族的成员。在一个实施例中,该T细胞刺激剂为抗原呈递细胞(APC)/T细胞激动剂。在另一个实施例中,该T细胞刺激剂为CD134或其配体或其片段、CD-137或其配体或其片段或诱导型T细胞共刺激剂(ICOS)或其配体或其片段。
在一个实施例中,本文提供的方法还包括将本文提供的免疫原性组合物与吲哚胺2,3-双加氧酶(IDO)通路抑制剂共给药的步骤。用于本发明的IDO通路抑制剂包括任何本领域已知的IDO通路抑制剂,其包括但不限于1-甲基色氨酸(1MT)、1-甲基色氨酸(1MT)、Necrostatin-1、吡哆醛异烟腙、依布硒啉、5-甲基吲哚M-3-甲醛、CAY10581、抗IDO抗体或小分子IDO抑制剂。在另一个实施例中,本文提供的组合物和方法还与在前的或随后的化疗或放疗方案联合使用。
在一个实施例中,本文提供的方法还包括将本文提供的免疫原性组合物与在所述受试者中增强抗肿瘤免疫应答的肿瘤激酶抑制剂共给药的步骤。肿瘤激酶抑制剂(TKI)起干扰特定细胞信号通路的作用,由此使得能对所选的恶性肿瘤进行靶特异性治疗。TKI为公知的,技术人员应理解其包括在以下表1中列出的那些以及任何其他的已知增强抗肿瘤应答的TKI。
表1.
在另一个实施例中,存在于本文提供的向受试者给药的免疫原性组合物中的免疫检查点抑制剂(例如,PD-1信号通路抑制剂)的剂量为每2周5-10mg/kg、每3周5-10mg/kg或每3周1-2mg/kg。在另一个实施例中,该剂量为每周1-10mg/kg的范围。在另一个实施例中,该剂量为每2周1-10mg/kg的范围。在另一个实施例中,该剂量为每3周1-10mg/kg的范围。在另一个实施例中,该剂量为每4周1-10mg/kg的范围。这些剂量为示例性的,且无意进行限制。
在一个实施例中,本发明的组合物或组合物的混合物包含特异结合GITR或其部分的抗体或其功能性片段。在另一个实施例中,本发明的组合物或组合物的混合物包含特异结合OX40或其部分的抗体或其功能性片段。在另一个实施例中,本发明的组合物或组合物的混合物包含特异结合GITR或其部分的抗体和特异结合OX40的抗体。在另一个实施例中,本发明的组合物或组合物的混合物包含Lm菌株和特异结合GITR的抗体或其功能性片段。在另一个实施例中,本发明的组合物或组合物的混合物包含Lm菌株和特异结合OX40的抗体或其功能性片段。在另一个实施例中,本发明的组合物或组合物的混合物包含Lm菌株以及特异结合GITR或其部分的抗体和特异结合OX40或其部分的抗体。
存在于相同或不同组合物中的不同抗体不必具有相同的形式,例如一种抗体可以是单克隆抗体,另一种可以是Fab片段。每种可能性代表了本发明的不同实施例。
术语“抗体功能性片段”指完整抗体的能够特异结合抗原的一部分。抗体片段的例子包括但不限于Fab、Fab'、F(ab')2和Fv片段、线性抗体、scFv抗体以及从抗体片段形成的多特异性抗体。
技术人员应理解,就抗体而言的术语“结合”或“特异结合”涵盖抗体或其功能性片段,其识别特异抗原,而基本上不识别或结合样品中的其他分子。例如,特异结合来自一个物种的抗原的抗体可能也结合来自一个或多个物种的该抗原,但是,这种跨物种反应性本身不改变将抗体分类为特异性的。在另一例子中,特异结合抗原的抗体可能也结合该抗原的不同等位基因形式。但是,这种交叉反应性不改变将抗体分类为特异性的。在一些情况下,可在提及抗体、蛋白或肽与第二化学物相互作用中使用术语“特异性结合”或“特异地结合”,指该相互作用依赖于该化学物上特定结构(例如抗原决定簇或表位)的存在;例如,抗体识别并结合特异蛋白结构而不是特异氨基酸序列。
在一个实施例中,本发明的组合物包含重组的单核细胞增多性李斯特菌(Lm)菌株。在另一个实施例中,本发明的组合物包含抗体或其功能性片段,如本文所描述的。
在一个实施例中,本文提供的组合物,无论是作为单一组合物给药的一部分还是作为组合物的混合物的一部分,都包含本文提供的抗体或其功能性片段,以及本文提供的重组减毒李斯特菌。在另一个实施例中,本文提供的组合物的每个组分在本文提供的组合物的另一组分之前、同时或之后给药。在一个实施例中,即使同时给药时,基于李斯特菌的组合物和抗体或其功能性片段也可以作为两个分开的组合物给药。替代性地,在另一个实施例中,基于李斯特菌的组合物可以包含抗体或其功能性片段。
在一个实施例中,本发明的任意组合物都诱导强的干扰素-γ固有刺激,该干扰素-γ在一个实施例中具有抗血管生成性质。在一个实施例中,本发明的李斯特菌诱导强的干扰素-γ固有刺激,该干扰素-γ在一个实施例中具有抗血管生成性质(Dominiecki等人,Cancer Immunol Immunother.2005年5月;54(5):477-88.2004年10月6日电子版,通过引用将其全文并入本文;Beatty和Paterson,J Immunol.2001年2月15日;166(4):2276-82,通过引用将其全文并入本文)。在一个实施例中,李斯特菌的抗血管生成性质通过CD4+T细胞(Beatty和Paterson,2001)介导。在另一个实施例中,李斯特菌的抗血管生成性质通过CD8+T细胞介导。在另一个实施例中,由李斯特菌接种而产生的IFN-γ分泌由NK细胞、NKT细胞、Th1CD4+T细胞、TC1CD8+T细胞或它们的组合介导。
在另一个实施例中,本发明的任何组合物都诱导一种或多种抗血管生成蛋白或因子的产生。在一个实施例中,该抗血管生成蛋白为IFN-γ。在另一个实施例中,该抗血管生成蛋白为色素上皮衍生因子(PEDF)、血管生成抑制素、内皮抑素、fms样酪氨酸激酶sFlt)-1或可溶性内皮因子(sEng)。在一个实施例中,本发明的李斯特菌参与抗血管生成因子的释放,由此,在一个实施例中,除了作为载体向受试者引入抗原外还具有治疗作用。每个李斯特菌菌株及其类型代表了本发明的独立实施例。
在另一个实施例中,本发明提供了包含游离型重组核酸分子的重组李斯特菌菌株,该核酸分子包含第一和至少第二开放阅读框,每个编码第一和至少第二多肽,其中第一和至少第二多肽每个包含融合至含PEST的多肽的异源抗原或其功能性片段,其中该核酸还包含质粒复制控制区。在另一个实施例中,该质粒控制区调节从所述第一和至少第二开放阅读框的表达。在另一个实施例中,该质粒控制区包含编码转录阻遏物的开放阅读框,该转录阻遏物阻遏从所述第一或至少第二开放阅读框的异源抗原表达。在另一个实施例中,该质粒控制区包含编码转录诱导物的开放阅读框,该转录诱导物诱导从所述第一和至少第二开放阅读框的异源抗原表达。在另一个实施例中,该核酸分子包含1-4个开放阅读框,每个编码1-4个重组多肽,其中所述重组多肽每个包含融合至含PEST的多肽的异源抗原或其功能性片段。在另一个实施例中,质粒控制区阻遏从所述第一至第四开放阅读框的异源抗原表达。在另一个实施例中,该质粒控制区包含编码转录诱导物的开放阅读框,该转录诱导物诱导从所述第一至第四开放阅读框的异源抗原表达。
在一个实施例中,本领域已知有不同类型的转录调节机制,这些机制包括但不限于“负控制”和“正控制”。在负控制中,调节蛋白或阻遏蛋白结合操纵子并防止RNA聚合酶正确结合至启动子序列。备选地,阻遏蛋白可以以不能阻断RNA聚合酶结合启动子的无活性形式合成;然后该阻遏蛋白通过结合辅阻遏物被活化,来防止RNA聚合酶结合至启动子。这种类型的控制在合成代谢途径(例如,精氨酸生物合成)中最常见,其中辅阻遏物通常是合成代谢途径的终产物。备选地,阻遏蛋白以活性形式合成,结合至操纵子并防止RNA聚合酶结合至启动子。当诱导物结合阻遏物时,阻遏物变成无活性的,由此现在RNA聚合酶自由地启动转录。这种类型的控制在分解代谢途径(例如,乳糖分解代谢)中最常见。该诱导物通常是待降解的底物形式。在正控制中,称为激活蛋白的调节蛋白结合操纵子,并且该激活物分子稳定RNA聚合酶结合启动子区域。其例子包括阿拉伯糖分解代谢。调节蛋白(对于正和负调节二者而言)由调节基因编码,并且可以以低水平持续合成。它们可以是自身调节的,因而高浓度的调节蛋白(与高质粒产量相关)结合其自身的操纵子并阻遏RNA聚合酶结合启动子区域。这让转录停止,直至其水平下降。这些类型的调节的几个例子包括乳糖操纵子、精氨酸操纵子、白喉毒素基因调节系统等。转录阻遏物及其使用方法为本领域熟知的,并考虑用在本发明中。
在一个实施例中,本文提供的方法包括在将本文提供的表达多个融合蛋白的重组李斯特菌用于临床环境之前测定重组李斯特菌中的代谢负荷的步骤。这种情况下,技术人员可容易地确定哪种最佳条件用于表达本文提供的包含异源抗原的融合蛋白。在另一个实施例中,测量代谢负荷通过发明时本领域已知的任何方式完成,其包括但不限于:测量疫苗菌株的生长速率、最佳密度读数、集落形成单位(CFU)铺板等。在另一个实施例中,细菌细胞的代谢负荷通过测量细菌细胞的活力来确定。测量细菌活力的方法在本领域是已知的和可得到的,其中一些包括但不限于:用于活力计数的细菌铺板、测量ATP以及流式细胞术。在ATP染色中,检测基于采用荧光素酶反应来测量来自活细胞的ATP的量,其中细胞中ATP的量与细胞活力相关。至于流式细胞术,该方法可以多种方式使用,在本领域中也是已知的,例如在使用活力染料后,该染料被活细菌细胞排出,被死亡细菌细胞吸收或吸附。技术人员可容易地理解,这些以及任何其他本领域中已知的测定细菌活力的方法均可用在本发明中。应当理解,技术人员将能够实践发明时本领域可获得的知识来测量李斯特菌的生长速率或李斯特菌菌株对标志物基因的表达,这使得能确定表达多个异源抗原或其功能性片段的李斯特菌菌株的代谢负荷。
在一个实施例中,术语“至少第二核酸分子”指两个或多个核酸分子,或者其指三个、四个、五个核酸分子,以此类推。在另一个实施例中,该术语指多至10个核酸分子,或多至20或多至30个核酸分子。
在一个实施例中,本文提供的重组李斯特菌菌株包含递送两种或更多种抗原的多价质粒。在另一个实施例中,该质粒为本文描述的双重质粒(dual plasmid)(参见图20和实例40)。在另一个实施例中,本文提供了编码该多价质粒的游离型重组核酸分子。在另一个实施例中,该多价质粒递送两种至五种抗原。在另一个实施例中,该多价质粒递送两种至十种抗原。在另一个实施例中,该多价质粒中的抗原融合至含PEST的氨基酸序列。
在一个实施例中,本文提供的质粒保持在染色体外或者为游离的,即在转染进细菌之后不整合进宿主细菌的染色体中。在另一个实施例中,本文提供的质粒为整合型质粒,其在转染进细菌后整合进宿主细菌的染色体序列中(特异地或随机地)。
在另一个实施例中,游离型重组核酸骨架由包含SEQ ID NO:1的序列编码。在另一个实施例中,本文提供的游离型重组核酸由SEQ ID NO:1组成的序列编码。在另一个实施例中,本文提供的游离型重组核酸由SEQ ID NO:1中列出的序列编码。
ggagtgtatactggcttactatgttggcactgatgagggtgtcagtgaagtgcttcatgtggcaggagaaaaaaggctgcaccggtgcgtcagcagaatatgtgatacaggatatattccgcttcctcgctcactgactcgctacgctcggtcgttcgactgcggcgagcggaaatggcttacgaacggggcggagatttcctggaagatgccaggaagatacttaacagggaagtgagagggccgcggcaaagccgtttttccataggctccgcccccctgacaagcatcacgaaatctgacgctcaaatcagtggtggcgaaacccgacaggactataaagataccaggcgtttccccctggcggctccctcgtgcgctctcctgttcctgcctttcggtttaccggtgtcattccgctgttatggccgcgtttgtctcattccacgcctgacactcagttccgggtaggcagttcgctccaagctggactgtatgcacgaaccccccgttcagtccgaccgctgcgccttatccggtaactatcgtcttgagtccaacccggaaagacatgcaaaagcaccactggcagcagccactggtaattgatttagaggagttagtcttgaagtcatgcgccggttaaggctaaactgaaaggacaagttttggtgactgcgctcctccaagccagttacctcggttcaaagagttggtagctcagagaaccttcgaaaaaccgccctgcaaggcggttttttcgttttcagagcaagagattacgcgcagaccaaaacgatctcaagaagatcatcttattaatcagataaaatatttctagccctcctttgattagtatattcctatcttaaagttacttttatgtggaggcattaacatttgttaatgacgtcaaaaggatagcaagactagaataaagctataaagcaagcatataatattgcgtttcatctttagaagcgaatttcgccaatattataattatcaaaagagaggggtggcaaacggtatttggcattattaggttaaaaaatgtagaaggagagtgaaacccatgaaaaaaataatgctagtttttattacacttatattagttagtctaccaattgcgcaacaaactgaagcaaaggatgcatctgcattcaataaagaaaattcaatttcatccatggcaccaccagcatctccgcctgcaagtcctaagacgccaatcgaaaagaaacacgcggatgaaatcgataagtatatacaaggattggattacaataaaaacaatgtattagtataccacggagatgcagtgacaaatgtgccgccaagaaaaggttacaaagatggaaatgaatatattgttgtggagaaaaagaagaaatccatcaatcaaaataatgcagacattcaagttgtgaatgcaatttcgagcctaacctatccaggtgctctcgtaaaagcgaattcggaattagtagaaaatcaaccagatgttctccctgtaaaacgtgattcattaacactcagcattgatttgccaggtatgactaatcaagacaataaaatagttgtaaaaaatgccactaaatcaaacgttaacaacgcagtaaatacattagtggaaagatggaatgaaaaatatgctcaagcttatccaaatgtaagtgcaaaaattgattatgatgacgaaatggcttacagtgaatcacaattaattgcgaaatttggtacagcatttaaagctgtaaataatagcttgaatgtaaacttcggcgcaatcagtgaagggaaaatgcaagaagaagtcattagttttaaacaaatttactataacgtgaatgttaatgaacctacaagaccttccagatttttcggcaaagctgttactaaagagcagttgcaagcgcttggagtgaatgcagaaaatcctcctgcatatatctcaagtgtggcgtatggccgtcaagtttatttgaaattatcaactaattcccatagtactaaagtaaaagctgcttttgatgctgccgtaagcggaaaatctgtctcaggtgatgtagaactaacaaatatcatcaaaaattcttccttcaaagccgtaatttacggaggttccgcaaaagatgaagttcaaatcatcgacggcaacctcggagacttacgcgatattttgaaaaaaggcgctacttttaatcgagaaacaccaggagttcccattgcttatacaacaaacttcctaaaagacaatgaattagctgttattaaaaacaactcagaatatattgaaacaacttcaaaagcttatacagatggaaaaattaacatcgatcactctggaggatacgttgctcaattcaacatttcttgggatgaagtaaattatgatctcgagactagttctagatttatcacgtacccatttccccgcatcttttatttttttaaatactttagggaaaaatggtttttgatttgcttttaaaggttgtggtgtagactcgtctgctgactgcatgctagaatctaagtcactttcagaagcatccacaactgactctttcgccacttttctcttatttgcttttgttggtttatctggataagtaaggctttcaagctcactatccgacgacgctatggcttttcttctttttttaatttccgctgcgctatccgatgacagacctggatgacgacgctccacttgcagagttggtcggtcgactcctgaagcctcttcatttatagccacatttcctgtttgctcaccgttgttattattgttattcggacctttctctgcttttgctttcaacattgctattaggtctgctttgttcgtatttttcactttattcgatttttctagttcctcaatatcacgtgaacttacttcacgtgcagtttcgtatcttggtcccgtatttacctcgcttggctgctcttctgttttttcttcttcccattcatctgtgtttagactggaatcttcgctatctgtcgctgcaaatattatgtcggggttaatcgtaatgcagttggcagtaatgaaaactaccatcatcgcacgcataaatctgtttaatcccacttatactccctcctcgtgatacgctaatacaacctttttagaacaaggaaaattcggccttcattttcactaatttgttccgttaaaaattggattagcagttagttatcttcttaattagctaatataagaaaaaatattcatgaattattttaagaatatcacttggagaattaatttttctctaacatttgttaatcagttaaccccaactgcttcccaagcttcacccgggccactaactcaacgctagtagtggatttaatcccaaatgagccaacagaaccagaaccagaaacagaacaagtaacattggagttagaaatggaagaagaaaaaagcaatgatttcgtgtgaataatgcacgaaatcattgcttatttttttaaaaagcgatatactagatataacgaaacaacgaactgaataaagaatacaaaaaaagagccacgaccagttaaagcctgagaaactttaactgcgagccttaattgattaccaccaatcaattaaagaagtcgagacccaaaatttggtaaagtatttaattactttattaatcagatacttaaatatctgtaaacccattatatcgggtttttgaggggatttcaagtctttaagaagataccaggcaatcaattaagaaaaacttagttgattgccttttttgttgtgattcaactttgatcgtagcttctaactaattaattttcgtaagaaaggagaacagctgaatgaatatcccttttgttgtagaaactgtgcttcatgacggcttgttaaagtacaaatttaaaaatagtaaaattcgctcaatcactaccaagccaggtaaaagtaaaggggctatttttgcgtatcgctcaaaaaaaagcatgattggcggacgtggcgttgttctgacttccgaagaagcgattcacgaaaatcaagatacatttacgcattggacaccaaacgtttatcgttatggtacgtatgcagacgaaaaccgttcatacactaaaggacattctgaaaacaatttaagacaaatcaataccttctttattgattttgatattcacacggaaaaagaaactatttcagcaagcgatattttaacaacagctattgatttaggttttatgcctacgttaattatcaaatctgataaaggttatcaagcatattttgttttagaaacgccagtctatgtgacttcaaaatcagaatttaaatctgtcaaagcagccaaaataatctcgcaaaatatccgagaatattttggaaagtctttgccagttgatctaacgtgcaatcattttgggattgctcgtataccaagaacggacaatgtagaattttttgatcccaattaccgttattctttcaaagaatggcaagattggtctttcaaacaaacagataataagggctttactcgttcaagtctaacggttttaagcggtacagaaggcaaaaaacaagtagatgaaccctggtttaatctcttattgcacgaaacgaaattttcaggagaaaagggtttagtagggcgcaatagcgttatgtttaccctctctttagcctactttagttcaggctattcaatcgaaacgtgcgaatataatatgtttgagtttaataatcgattagatcaacccttagaagaaaaagaagtaatcaaaattgttagaagtgcctattcagaaaactatcaaggggctaatagggaatacattaccattctttgcaaagcttgggtatcaagtgatttaaccagtaaagatttatttgtccgtcaagggtggtttaaattcaagaaaaaaagaagcgaacgtcaacgtgttcatttgtcagaatggaaagaagatttaatggcttatattagcgaaaaaagcgatgtatacaagccttatttagcgacgaccaaaaaagagattagagaagtgctaggcattcctgaacggacattagataaattgctgaaggtactgaaggcgaatcaggaaattttctttaagattaaaccaggaagaaatggtggcattcaacttgctagtgttaaatcattgttgctatcgatcattaaattaaaaaaagaagaacgagaaagctatataaaggcgctgacagcttcgtttaatttagaacgtacatttattcaagaaactctaaacaaattggcagaacgccccaaaacggacccacaactcgatttgtttagctacgatacaggctgaaaataaaacccgcactatgccattacatttatatctatgatacgtgtttgtttttctttgctggctagcttaattgcttatatttacctgcaataaaggatttcttacttccattatactcccattttccaaaaacatacggggaacacgggaacttattgtacaggccacctcatagttaatggtttcgagccttcctgcaatctcatccatggaaatatattcatccccctgccggcctattaatgtgacttttgtgcccggcggatattcctgatccagctccaccataaattggtccatgcaaattcggccggcaattttcaggcgttttcccttcacaaggatgtcggtccctttcaattttcggagccagccgtccgcatagcctacaggcaccgtcccgatccatgtgtctttttccgctgtgtactcggctccgtagctgacgctctcgccttttctgatcagtttgacatgtgacagtgtcgaatgcagggtaaatgccggacgcagctgaaacggtatctcgtccgacatgtcagcagacgggcgaaggccatacatgccgatgccgaatctgactgcattaaaaaagccttttttcagccggagtccagcggcgctgttcgcgcagtggaccattagattctttaacggcagcggagcaatcagctctttaaagcgctcaaactgcattaagaaatagcctctttctttttcatccgctgtcgcaaaatgggtaaatacccctttgcactttaaacgagggttgcggtcaagaattgccatcacgttctgaacttcttcctctgtttttacaccaagtctgttcatccccgtatcgaccttcagatgaaaatgaagagaaccttttttcgtgtggcgggctgcctcctgaagccattcaacagaataacctgttaaggtcacgtcatactcagcagcgattgccacatactccgggggaaccgcgccaagcaccaatataggcgccttcaatccctttttgcgcagtgaaatcgcttcatccaaaatggccacggccaagcatgaagcacctgcgtcaagagcagcctttgctgtttctgcatcaccatgcccgtaggcgtttgctttcacaactgccatcaagtggacatgttcaccgatatgttttttcatattgctgacattttcctttatcacggacaagtcaatttccgcccacgtatctctgtaaaaaggttttgtgctcatggaaaactcctctcttttttcagaaaatcccagtacgtaattaagtatttgagaattaattttatattgattaatactaagtttacccagttttcacctaaaaaacaaatgatgagataatagctccaaaggctaaagaggactataccaactatttgttaat(SEQ ID NO:1)。
在一个实施例中,多价质粒骨架包含编码至少两个抗原的至少两个核酸序列。在另一个实施例中,该重组游离型核酸编码质粒骨架序列和至少两个抗原。在另一个实施例中,该抗原为携带该质粒的细菌宿主的异源抗原。在另一个实施例中,该抗原为携带该质粒的李斯特菌宿主的异源抗原。在另一个实施例中,编码质粒骨架和至少两个异源抗原的该重组游离型核酸序列包含SEQ ID NO:2。在另一个实施例中,编码质粒骨架和至少两个异源抗原的该重组游离型核酸序列由SEQ ID NO:2组成。
在另一个实施例中,由SEQ ID NO:2内的序列编码的该抗原之一为E7(SEQ ID NO:2中以粗体表示)。在另一个实施例中,该E7序列在SEQ ID NO:3
Ctcgagcatggagatacacctacattgcatgaatatatgttagatttgcaaccagagacaactgatctctactgttatgagcaattaaatgacagctcagaggaggaggatgaaatagatggtccagctggacaagcagaaccggacagagcccattacaatattgtaaccttttgttgcaagtgtgactctacgcttcggttgtgcgtacaaagcacacacgtagacattcgtactttggaagacctgttaatgggcacactaggaattgtgtgccccatctgttctcagaaaccataaactagt(SEQ ID NO:3)中列出。
在一个实施例中,由SEQ ID NO:2内的序列编码的该抗原之一为嵌合Her2-neu抗原(SEQ ID NO:2中以斜体表示)。在另一个实施例中,该嵌合Her2-neu序列在SEQ ID NO:4中列出。
ctagtggtgatggtgatgatggagctcagatctgtctaagaggcagccatagggcataagctgtgtcaccagctgcaccgtggatgtcaggcagatgcccagaaggcgggagacatatggggagcccacaccagccatcacgtatgcttcgtctaagatttctttgttggctttgggggatgtgttttccctcaacactttgatggccactggaattttcacattctccccatcagggatccagatgcccttgtagactgtgccaaaagcgccagatccaagcaccttcaccttcctcagctccgtctctttcaggatccgcatctgcgcctggttgggcatcgctccgctaggtgtcagcggctccaccagctccgtttcctgcagcagtctccgcatcgtgtacttccggatcttctgctgccctcgggcgcacagctggtggcaggccaggccctcgcccacacactcgtcctctggccggttggcagtgtggagcagagcttggtgcgggttccgaaagagctggtcccagggcaccgtgtgcacgaagcagaggtgggtgttatggtggatgagggccagtccactgcccagttccctcagtgagcgcagccccagccagctgatgcccagcccttgcagggtcagcgagtaggcgccattgtgcagaattcgtccccggattacttgcaggttctggaagacgctgaggtcaggcaggctgtccggccatgctgagatgtataggtaacctgtgatctcttccagagtctcaaacacttggagctgctctggctggagcggggcagtgttggaggctgggtccccatcaaagctctccggcagaaatgccaggctcccaaagatcttcttgcagccagcaaactcctggatattcttccacaaaatcgtgtcctggtagcagagctgggggttccgctggatcaagacccctcctttcaagatctctgtgaggcttcgaagctgcagctcccgcaggcctcctggggaggcccctgtgacaggggtggtattgttcagcgggtctccattgtctagcacggccagggcatagttgtcctcaaagagctgggtgcctcgcacaatccgcagcctctgcagtgggacctgcctcacttggttgtgagcgatgagcacgtagccctgcacctcctggatatcctgcaggaaggacaggctggcattggtgggcaggtaggtgagttccaggtttccctgcaccacctggcagccctggtagaggtggcggagcatgtccaggtgggttctagat(SEQ ID NO:4)。
在另一个实施例中,本文提供的编码代谢酶的基因在李斯特菌p60启动子控制下表达。在另一个实施例中,使用inlA(编码内化素)启动子。在另一个实施例中,使用hyl启动子。在另一个实施例中,使用actA启动子。当具备了本公开和本文提供的方法,技术人员将容易理解,不同的转录启动子、终止子、携带载体或特异基因序列(例如可购得的克隆载体中的序列)可成功地用在本发明的方法和组合物中。如本发明所预料的,这些功能在例如可购得的称为pUC系列的载体中有提供。在另一个实施例中,除去非必需DNA系列(例如抗生素抗性基因)。每种可能性代表了本发明的独立实施例。
在另一个实施例中,整合酶基因在任何其他革兰氏阳性启动子控制下表达。在另一个实施例中,编码代谢酶的基因在李斯特菌中有功能的任何其他启动子控制下表达。技术人员应理解,其他的启动子或多顺反子表达盒可以用于驱动该基因的表达。每种可能性代表了本发明的独立实施例。
在一个实施例中,“组成型”启动子为这样的核苷酸序列,当可操作地连接编码或规定某基因产物的多核苷酸时,其导致在活的人细胞中在该细胞的大多数或全部生理条件下都生成该基因产物。
在一个实施例中,“诱导型”启动子为这样的核苷酸序列,当可操作地连接编码或规定某基因产物的多核苷酸时,其导致在活细胞中基本上仅在对应于该启动子的诱导物存在于该细胞中时才生成该基因产物。
在一个实施例中,“组织特异性”启动子为这样的核苷酸序列,当可操作地连接编码或规定某基因产物的多核苷酸时,其导致在活的人细胞中基本上仅在该细胞为对应于该启动子的组织类型的细胞时才生成该基因产物。
在一个实施例中,本文提供的重组李斯特菌菌株已经在动物宿主中传代。在另一个实施例中,该动物宿主为非人动物宿主。在另一个实施例中,该传代让该菌株作为疫苗载体的效能最佳化。在另一个实施例中,该传代稳定了李斯特菌菌株的免疫原性。在另一个实施例中,该传代稳定了李斯特菌菌株的毒力。在另一个实施例中,该传代增强了李斯特菌菌株的免疫原性。在另一个实施例中,该传代增强了李斯特菌菌株的毒力。在另一个实施例中,该传代除去李斯特菌菌株的不稳定亚株。在另一个实施例中,该传代减少了李斯特菌菌株的不稳定亚株的普遍性。在另一个实施例中,该传代让菌株减毒,或者在另一个实施例中让菌株毒力减小。让重组李斯特菌菌株在动物宿主中传代的方法为本领域所公知的,例如在美国专利申请No.10/541,614中有描述。在一个实施例中,李斯特菌传代的动物为哺乳动物,其在一个实施例中为小鼠。本发明考虑采用哺乳动物传代,例如小鼠、兔、豚鼠、仓鼠、沙鼠、大鼠等。这些哺乳动物在本领域是公知的,技术人员可通过各种各样的批发商、经销商和诸如Jackson Laboratories(Bar Harbor,Me.)的实验室获得。让重组李斯特菌菌株在动物宿主中传代的方法为本领域所已知的,例如在美国专利申请No.10/541,614中有描述。每种可能性代表了本发明的独立实施例。
在一个实施例中,本文提供了在人受试者中预防疾病、治疗疾病和接种的方法和组合物。在另一个实施例中,本发明提供了在动物受试者中预防疾病、治疗疾病和接种的方法和组合物。
在另一个实施例中,本发明涉及增强人或动物的抗肿瘤免疫应答。在另一个实施例中,通过以本文提供的组合物或本文提供的组合物的混合物给药来增强受试者中抗肿瘤应答的方法可与其他已知的抗肿瘤或抗癌疗法联合。在另一个实施例中,本发明的组合物可单独使用,或者与任何这样的疗法联合:其中佐剂是适合的或者不通常使用佐剂但佐剂可能在其中有功用的情形,例如化疗或放疗。
在另一个实施例中,本文提供的方法还包括克服或“打破”对作为自身抗原的异源抗原的耐受。这些抗原可以由各种肿瘤异常表达,这些肿瘤屈从于本发明范围下采用本文提供的方法和组合物的治疗或预防。
在一个实施例中,由本文提供的方法和组合物诱导的免疫应答为治疗性免疫应答。在另一个实施例中,其为预防性免疫应答。在另一个实施例中,其为增强的免疫应答,超出本领域中可得到的用于在受本文提供的病症折磨的受试者中诱导免疫应答的方法。在另一个实施例中,该免疫应答导致折磨该受试者的本文提供的肿瘤的清除。
在一个实施例中,肿瘤为缺氧实体肿瘤。在另一个实施例中,该肿瘤为实体肿瘤。在另一个实施例中,该肿瘤为与本文提供的任何癌相关的以及本领域已知的任何肿瘤。
在一个实施例中,表达截短的李斯特菌溶血素O的重组减毒李斯特菌与其他的治疗模式联合,用于增强免疫应答以及用于预防和治疗包括癌或实体肿瘤在内的疾病。在一个实施例中,表达截短的ActA的重组减毒李斯特菌与其他的治疗模式联合,用于增强免疫应答以及用于预防和治疗包括癌或实体肿瘤在内的疾病。在一个实施例中,表达PEST氨基酸序列的重组减毒李斯特菌与其他的治疗模式联合,用于增强免疫应答以及用于预防和治疗包括癌或实体肿瘤在内的疾病。
在另一个实施例中,本文提供了改善治疗性疫苗的免疫原性的方法,该方法包括将该疫苗和包含表达多种融合蛋白的单一重组李斯特菌的组合物或每种包含表达本发明的融合蛋白的重组李斯特菌的组合物的混合物向受试者共给药,其中每种组合物或组合物的混合物增强该疫苗的免疫原性并引起抗原特异性免疫应答,由此改善该疫苗的免疫原性。在一个实施例中,该方法使得能治疗该疫苗特异针对的肿瘤。在另一个实施例中,该疫苗为药物疫苗、化疗剂、肽疫苗或本领域已知的任何其他类型的疫苗。
在另一个实施例中,用在本文提供的方法和组合物中的LLO为李斯特菌LLO。在一个实施例中,该LLO所源于的李斯特菌为单核细胞增多性李斯特菌(Lm)。在另一个实施例中,该李斯特菌为伊氏李斯特菌(Listeria ivanovii)。在另一个实施例中,该李斯特菌为威氏利斯特菌(Listeria welshimeri)。在另一个实施例中,该李斯特菌为西尔李斯特菌(Listeria seeligeri)。
在一个实施例中,该LLO蛋白由以下(SEQ ID NO:5)中列出的核酸序列编码。
atgaaaaaaataatgctagtttttattacacttatattagttagtctaccaattgcgcaacaaactgaagcaaaggatgcatctgcattcaataaagaaaattcaatttcatccatggcaccaccagcatctccgcctgcaagtcctaagacgccaatcgaaaagaaacacgcggatgaaatcgataagtatatacaaggattggattacaataaaaacaatgtattagtataccacggagatgcagtgacaaatgtgccgccaagaaaaggttacaaagatggaaatgaatatattgttgtggagaaaaagaagaaatccatcaatcaaaataatgcagacattcaagttgtgaatgcaatttcgagcctaacctatccaggtgctctcgtaaaagcgaattcggaattagtagaaaatcaaccagatgttctccctgtaaaacgtgattcattaacactcagcattgatttgccaggtatgactaatcaagacaataaaatagttgtaaaaaatgccactaaatcaaacgttaacaacgcagtaaatacattagtggaaagatggaatgaaaaatatgctcaagcttatccaaatgtaagtgcaaaaattgattatgatgacgaaatggcttacagtgaatcacaattaattgcgaaatttggtacagcatttaaagctgtaaataatagcttgaatgtaaacttcggcgcaatcagtgaagggaaaatgcaagaagaagtcattagttttaaacaaatttactataacgtgaatgttaatgaacctacaagaccttccagatttttcggcaaagctgttactaaagagcagttgcaagcgcttggagtgaatgcagaaaatcctcctgcatatatctcaagtgtggcgtatggccgtcaagtttatttgaaattatcaactaattcccatagtactaaagtaaaagctgcttttgatgctgccgtaagcggaaaatctgtctcaggtgatgtagaactaacaaatatcatcaaaaattcttccttcaaagccgtaatttacggaggttccgcaaaagatgaagttcaaatcatcgacggcaacctcggagacttacgcgatattttgaaaaaaggcgctacttttaatcgagaaacaccaggagttcccattgcttatacaacaaacttcctaaaagacaatgaattagctgttattaaaaacaactcagaatatattgaaacaacttcaaaagcttatacagatggaaaaattaacatcgatcactctggaggatacgttgctcaattcaacatttcttgggatgaagtaaattatgatctcgag(SEQ ID NO:5)。
在另一个实施例中,该LLO蛋白具有序列SEQ ID NO:6。在另一个实施例中,该LLO蛋白包含SEQ ID NO:6中列出的序列。
M K K I M L V F I T L I L V S L P I A Q Q T E A K D A S A F N K E N S I S S M A P P A S P P A S P K T P I E K K H A D E I D K Y I Q G L D Y N K N N V L V Y H G D A V T N V P P R K G Y K D G N E Y I V V E K K K K S I N Q N N A D I Q V V N A I S S L T Y P G A L V K A N S E L V E N Q P D V L P V K R D S L T L S I D L P G M T N Q D N K I V V K N A T K S N V N N A V N T L V E R W N E K Y A Q A Y P N V S A K I D Y D D E M A Y S E S Q L I A K F G T A F K A V N N S L N V N F G A I S E G K M Q E E V I S F K Q I Y Y N V N V N E P T R P S R F F G K A V T K E Q L Q A L G V N A E N P P A Y I S S V A Y G R Q V Y L K L S T N S H S T K V K A A F D A A V S G K S V S G D V E L T N I I K N S S F K A V I Y G G S A K D E V Q I I D G N L G D L R D I L K K G A T F N R E T P G V P I A Y T T N F L K D N E L A V I K N N S E Y I E T T S K A Y T D G K I N I D H S G G Y V A Q F N I S W D E V N Y D L(SEQ ID NO:6)
对应于该序列的前蛋白的前25个氨基酸为信号序列,并在由细菌分泌时从LLO切除。因此,在本实施例中,全长的活性LLO蛋白为504个残基长。在另一个实施例中,该LLO蛋白具有GenBank登录号DQ054588、DQ054589、AY878649、U25452或U25452中列出的序列。在另一个实施例中,该LLO蛋白为LLO蛋白的变体。在另一个实施例中,该LLO蛋白为LLO蛋白的同源物。每种可能性代表了本发明的独立实施例。
在另一个实施例中,用于构建本发明的(任何形式的)组合物的该LLO蛋白(在另一个实施例中,用作并入本文提供的组合物中的LLO片段的来源)在另一个实施例中具有以下序列:
MKKIMLVFITLILVSLPIAQQTEAKDASAFNKENSISSMAPPASPPASPKTPIEKKHADEIDKYIQGLDYNKNNVLVYHGDAVTNVPPRKGYKDGNEYIVVEKKKKSINQNNADIQVVNAISSLTYPGALVKANSELVENQPDVLPVKRDSLTLSIDLPGMTNQDNKIVVKNATKSNVNNAVNTLVERWNEKYAQAYPNVSAKIDYDDEMAYSESQLIAKFGTAFKAVNNSLNVNFGAISEGKMQEEVISFKQIYYNVNVNEPTRPSRFFGKAVTKEQLQALGVNAENPPAYISSVAYGRQVYLKLSTNSHSTKVKAAFDAAVSGKSVSGDVELTNIIKNSSFKAVIYGGSAKDEVQIIDGNLGDLRDILKKGATFNRETPGVPIAYTTNFLKDNELAVIKNNSEYIETTSKAYTDGKINIDHSGGYVAQFNISWDEVNYDPEGNEIVQHKNWSENNKSKLAHFTSSIYLPGNARNINVYAKECTGLAWEWWRTVIDDRNLPLVKNRNISIWGTTLYPKYSNKVDNPIE(GenBank登录号P13128;SEQ ID NO:123;核酸序列在GenBank登录号X15127中列出)。对应于该序列的前蛋白的前25个AA为信号序列,并在由细菌分泌时从LLO切除。因此,在本实施例中,全长的活性LLO蛋白为504个残基长。在另一个实施例中,该LLO蛋白为SEQ ID NO:123的同源物。在另一个实施例中,该LLO蛋白为SEQ ID NO:123的变体。在另一个实施例中,该LLO蛋白为SEQ ID NO:123的异构体。在另一个实施例中,该LLO蛋白为SEQ ID NO:123的片段。在另一个实施例中,该LLO蛋白为SEQ ID NO:123的同源物的片段。在另一个实施例中,该LLO蛋白为SEQ ID NO:123的变体的片段。在另一个实施例中,该LLO蛋白为SEQ ID NO:123的异构体的片段。每种可能性代表了本发明的独立实施例。
在另一个实施例中,“截短的LLO”或“tLLO”指包含PEST氨基酸序列结构域的LLO片段。在另一个实施例中,该术语指不包含氨基末端的活化结构域且不包含胱氨酸484的LLO片段。在另一个实施例中,该LLO蛋白为ctLLO。在另一个实施例中,ctLLO为全长LLO,其中胆固醇结合结构域(CBD)已被抗原肽或其表位所替换。在另一个实施例中,“替换”可指经由置换或缺失突变。在另一个实施例中,该LLO蛋白为mutLLO。在另一个实施例中,mutLLO为其中CBD已突变的mutLLO。在另一个实施例中,该mutLLO为其中CBD中的氨基酸已突变的mutLLO。在另一个实施例中,该突变为点突变、缺失、倒位、置换或它们的组合。在另一个实施例中,该突变为本领域已知的任何突变。在另一个实施例中,该突变的LLO蛋白包含CBD中缺失、置换或点突变的任何组合和/或LLO的信号序列的缺失。在另一个实施例中,让CBD突变减小了LLO的溶血活性。在另一个实施例中,该CBD被已知的HLAI类限制性表位替换,以便用作疫苗。在另一个实施例中,该突变的LLO从大肠杆菌表达系统中表达和纯化。
在另一个实施例中,该LLO片段由PEST序列构成。在另一个实施例中,该LLO片段包含PEST序列。在另一个实施例中,该LLO片段由529个氨基酸的全长LLO蛋白的约前400至441个氨基酸构成。在另一个实施例中,该LLO片段为LLO蛋白的非溶血形式。
在另一个实施例中,本发明提供了重组多肽,其包含(a)突变的LLO蛋白,其中该突变的LLO蛋白含有内部缺失,该内部缺失包含该突变LLO蛋白的胆固醇结合结构域;和(b)目的异源肽。在另一个实施例中,该LLO胆固醇结合结构域的序列在本领域是公知的,并在美国专利No.8,771,702中有描述,通过引用将其并入本文。在另一个实施例中,该内部缺失为11-50个氨基酸的内部缺失。在另一个实施例中,该内部缺失就该重组蛋白或多肽的溶血活性而言是去活性的。在另一个实施例中,该重组蛋白或多肽相对于野生型LLO表现出溶血活性的减小。在另一个实施例中,本文提供了包含本文提供的重组蛋白或重组多肽的李斯特菌。每种可能性代表了本发明的另一个实施例。
在另一个实施例中,本发明提供了重组蛋白或多肽,其包含(a)突变的LLO蛋白,其中该突变的LLO蛋白含有内部缺失,该内部缺失包含该突变LLO蛋白的胆固醇结合结构域的片段;和(b)目的异源肽。在另一个实施例中,该内部缺失为1-11个氨基酸的内部缺失。在另一个实施例中,该胆固醇结合结构域的序列在SEQ ID NO:130中列出。在另一个实施例中,该内部缺失就该重组蛋白或多肽的溶血活性而言是去活性的。在另一个实施例中,该重组蛋白或多肽相对于野生型LLO表现出溶血活性的减小。每种可能性代表了本发明的另一个实施例。
本发明的方法和组合物的突变区域在另一个实施例中包含SEQ ID NO:123的残基C484。在另一个实施例中,该突变区域包含同源LLO蛋白的相应半胱氨酸残基。在另一个实施例中,该突变区域包含SEQ ID NO:123的残基W491。在另一个实施例中,该突变区域包含同源LLO蛋白的相应色氨酸残基。在另一个实施例中,该突变区域包含SEQ ID NO:123的残基W492。在另一个实施例中,该突变区域包含SEQ ID NO:123的残基C484、W491和W492。在另一个实施例中,该突变区域包含同源LLO蛋白的相应色氨酸残基。鉴定同源蛋白的相应残基的方法在本领域是公知的,并且例如包括序列比对。每种可能性代表了本发明的独立实施例。
在另一个实施例中,该突变区域包含残基C484和W491。在另一个实施例中,该突变区域包含残基C484和W492。在另一个实施例中,该突变区域包含残基C491和W492。在另一个实施例中,该突变区域包含残基C484、C491和W492。每种可能性代表了本发明的独立实施例。
在另一个实施例中,本发明的方法和组合物中提供的LLO蛋白的突变区域包含该突变的LLO蛋白的胆固醇结合结构域或其片段。例如,由SEQ ID NO:37的残基470-500、470-510或480-500构成的突变区域包含其CBD(残基483-493)。在另一个实施例中,该突变区域为该突变LLO蛋白或其片段的CBD的片段。例如,如本文所提供的,残基C484、W491和W492(每个都为CBD的片段)突变为丙氨酸残基(实例38)。另外,如本文提供的,CBD的片段(残基484-492)被来自NY-ESO-1的异源序列所替换(实例39)。在另一个实施例中,该突变区域与该突变LLO蛋白或其片段的CBD重叠。例如,由SEQ ID NO:123的残基470-490、480-488、490-500或486-510构成的突变区域包含其CBD。在另一个实施例中,单一的肽可以具有信号序列中的缺失和CBD中的突变或置换。每种可能性代表了本发明的独立实施例。
在另一个实施例中,本发明的方法和组合物的LLO蛋白中的内部缺失包含LLO蛋白的CBD或其片段。例如,由SEQ ID NO:37的残基470-500、470-510或480-500构成的内部缺失包含其CBD(残基483-493)。在另一个实施例中,该内部缺失为该突变LLO蛋白或其片段的CBD的片段。例如,残基484-492、485-490和486-488都为SEQ ID NO:123的CDB的片段。在另一个实施例中,该内部缺失与该突变LLO蛋白或其片段的CBD重叠。例如,由SEQ ID NO:123的残基470-490、480-488、490-500或486-510构成的内部缺失包含其CBD。每种可能性代表了本发明的独立实施例。
在一个实施例中,该LLO片段大概由残基1-25构成。在另一个实施例中,该LLO片段大概由残基1-50构成。在另一个实施例中,该LLO片段大概由残基1-75构成。在另一个实施例中,该LLO片段大概由残基1-100构成。在另一个实施例中,该LLO片段大概由残基1-125构成。在另一个实施例中,该LLO片段大概由残基1-150构成。在另一个实施例中,该LLO片段大概由残基1-175构成。在另一个实施例中,该LLO片段大概由残基1-200构成。在另一个实施例中,该LLO片段大概由残基1-225构成。在另一个实施例中,该LLO片段大概由残基1-250构成。在另一个实施例中,该LLO片段大概由残基1-275构成。在另一个实施例中,该LLO片段大概由残基1-300构成。在另一个实施例中,该LLO片段大概由残基1-325构成。在另一个实施例中,该LLO片段大概由残基1-350构成。在另一个实施例中,该LLO片段大概由残基1-375构成。在另一个实施例中,该LLO片段大概由残基1-400构成。在另一个实施例中,该LLO片段大概由残基1-425构成。在另一个实施例中,该LLO片段大概由残基1-441构成。每种可能性代表了本发明的独立实施例。
在另一个实施例中,该LLO片段含有对应于以上AA范围之一的同源LLO蛋白的残基。残基编号在另一个实施例中不必精确对应以上列举的残基编号;例如,如果该同源LLO蛋白相对于本文所用的LLO蛋白具有插入或缺失。
在另一个实施例中,本发明中可以使用来自其他物种的LLO的同源物,包括已知的溶解素,例如链球菌溶血素O、气荚膜梭菌溶血素O、肺炎球菌溶血素等或它们的片段。
在一个实施例中,本文提供的活的、减毒李斯特菌或重组李斯特菌表达ActA蛋白或其片段。在本发明的方法和组合物的另一个实施例中,ActA蛋白的片段融合至本文也提供的异源抗原或其片段。在另一个实施例中,该ActA蛋白的片段具有以下序列:
MRAMMVVFITANCITINPDIIFAATDSEDSSLNTDEWEEEKTEEQPSEVNTGPRYETAREVSSRDIKELEKSNKVRNTNKADLIAMLKEKAEKGPNINNNNSEQTENAAINEEASGADRPAIQVERRHPGLPSDSAAEIKKRRKAIASSDSELESLTYPDKPTKVNKKKVAKESVADASESDLDSSMQSADESSPQPLKANQQPFFPKVFKKIKDAGKWVRDKIDENPEVKKAIVDKSAGLIDQLLTKKKSEEVNASDFPPPPTDEELRLALPETPMLLGFNAPATSEPSSFEFPPPPTDEELRLALPETPMLLGFNAPATSEPSSFEFPPPPTEDELEIIRETASSLDSSFTRGDLASLRNAINRHSQNFSDFPPIPTEEELNGRGGRP(SEQ ID NO:7)。在另一个实施例中,本发明的方法和组合物的ActA AA序列包含SEQ ID NO:7中列出的序列。在另一个实施例中,该ActAAA序列为SEQ ID NO:7的同源物。在另一个实施例中,该ActA AA序列为SEQ ID NO:7的变体。在另一个实施例中,该ActA AA序列为SEQ ID NO:7的片段。在另一个实施例中,该ActA AA序列为SEQ ID NO:5的同种型。每种可能性代表了本发明的独立实施例。
在另一个实施例中,该ActA片段由包含以下序列的重组核苷酸编码:
ATGCGTGCGATGATGGTGGTTTTCATTACTGCCAATTGCATTACGATTAACCCCGACATAATATTTGCAGCGACAGATAGCGAAGATTCTAGTCTAAACACAGATGAATGGGAAGAAGAAAAAACAGAAGAGCAACCAAGCGAGGTAAATACGGGACCAAGATACGAAACTGCACGTGAAGTAAGTTCACGTGATATTAAAGAACTAGAAAAATCGAATAAAGTGAGAAATACGAACAAAGCAGACCTAATAGCAATGTTGAAAGAAAAAGCAGAAAAAGGTCCAAATATCAATAATAACAACAGTGAACAAACTGAGAATGCGGCTATAAATGAAGAGGCTTCAGGAGCCGACCGACCAGCTATACAAGTGGAGCGTCGTCATCCAGGATTGCCATCGGATAGCGCAGCGGAAATTAAAAAAAGAAGGAAAGCCATAGCATCATCGGATAGTGAGCTTGAAAGCCTTACTTATCCGGATAAACCAACAAAAGTAAATAAGAAAAAAGTGGCGAAAGAGTCAGTTGCGGATGCTTCTGAAAGTGACTTAGATTCTAGCATGCAGTCAGCAGATGAGTCTTCACCACAACCTTTAAAAGCAAACCAACAACCATTTTTCCCTAAAGTATTTAAAAAAATAAAAGATGCGGGGAAATGGGTACGTGATAAAATCGACGAAAATCCTGAAGTAAAGAAAGCGATTGTTGATAAAAGTGCAGGGTTAATTGACCAATTATTAACCAAAAAGAAAAGTGAAGAGGTAAATGCTTCGGACTTCCCGCCACCACCTACGGATGAAGAGTTAAGACTTGCTTTGCCAGAGACACCAATGCTTCTTGGTTTTAATGCTCCTGCTACATCAGAACCGAGCTCATTCGAATTTCCACCACCACCTACGGATGAAGAGTTAAGACTTGCTTTGCCAGAGACGCCAATGCTTCTTGGTTTTAATGCTCCTGCTACATCGGAACCGAGCTCGTTCGAATTTCCACCGCCTCCAACAGAAGATGAACTAGAAATCATCCGGGAAACAGCATCCTCGCTAGATTCTAGTTTTACAAGAGGGGATTTAGCTAGTTTGAGAAATGCTATTAATCGCCATAGTCAAAATTTCTCTGATTTCCCACCAATCCCAACAGAAGAAGAGTTGAACGGGAGAGGCGGTAGACCA(SEQ ID NO:8)。在另一个实施例中,该重组核苷酸具有SEQ ID NO:8中列出的序列。在另一个实施例中,本发明的方法和组合物的ActA编码核苷酸包含SEQ ID NO:8中列出的序列。在另一个实施例中,该ActA编码核苷酸为SEQ ID NO:8的同源物。在另一个实施例中,该ActA编码核苷酸为SEQ ID NO:8的变体。在另一个实施例中,该ActA编码核苷酸为SEQ ID NO:8的片段。在另一个实施例中,该ActA编码核苷酸为SEQ ID NO:8的同种型。每种可能性代表了本发明的独立实施例。
在另一个实施例中,该ActA片段由包含以下序列的重组核苷酸编码:
Tttatcacgtacccatttccccgcatcttttatttttttaaatactttagggaaaaatggtttttgatttgcttttaaaggttgtggtgtagactcgtctgctgactgcatgctagaatctaagtcactttcagaagcatccacaactgactctttcgccacttttctcttatttgcttttgttggtttatctggataagtaaggctttcaagctcactatccgacgacgctatggcttttcttctttttttaatttccgctgcgctatccgatgacagacctggatgacgacgctccacttgcagagttggtcggtcgactcctgaagcctcttcatttatagccacatttcctgtttgctcaccgttgttattattgttattcggacctttctctgcttttgctttcaacattgctattaggtctgctttgttcgtatttttcactttattcgatttttctagttcctcaatatcacgtgaacttacttcacgtgcagtttcgtatcttggtcccgtatttacctcgcttggctgctcttctgttttttcttcttcccattcatctgtgtttagactggaatcttcgctatctgtcgctgcaaatattatgtcggggttaatcgtaatgcagttggcagtaatgaaaactaccatcatcgcacgcat(SEQ ID NO:9)。在另一个实施例中,该重组核苷酸具有SEQ ID NO:9中列出的序列。在另一个实施例中,本发明的方法和组合物的ActA编码核苷酸包含SEQ ID NO:9中列出的序列。在另一个实施例中,该ActA编码核苷酸为SEQ ID NO:9的同源物。在另一个实施例中,该ActA编码核苷酸为SEQ ID NO:9的变体。在另一个实施例中,该ActA编码核苷酸为SEQ ID NO:9的片段。在另一个实施例中,该ActA编码核苷酸为SEQ ID NO:9的同种型。在另一个实施例中,SEQ ID NO:9被用来获得也在本文提供的构建体SEQ ID NO:2。每种可能性代表了本发明的独立实施例。
在本发明的方法和组合物的另一个实施例中,ActA蛋白的片段融合至异源抗原或其片段。在另一个实施例中,ActA蛋白的片段具有Genbank登录号AAF04762中列出的序列。在另一个实施例中,本发明的方法和组合物的ActA AA序列包含Genbank登录号AAF04762中列出的序列。在另一个实施例中,该ActA AA序列为Genbank登录号AAF04762的同源物。在另一个实施例中,该ActA AA序列为Genbank登录号AAF04762的变体。在另一个实施例中,该ActA AA序列为Genbank登录号AAF04762的片段。在另一个实施例中,该ActA AA序列为Genbank登录号AAF04762的同种型。每种可能性代表了本发明的独立实施例。
本发明的方法和组合物中使用的ActA蛋白的N末端片段在另一个实施例中具有SEQ ID NO:10:
MRAMMVVFITANCITINPDIIFAATDSEDSSLNTDEWEEEKTEEQPSEVNTGPRYETAREVSSRDIKELEKSNKVRNTNKADLIAMLKEKAEKGPNINNNNSEQTENAAINEEASGADRPAIQVERRHPGLPSDSAAEIKKRRKAIASSDSELESLTYPDKPTKVNKKKVAKESVADASESDLDSSMQSADESSPQPLKANQQPFFPKVFKKIKDAGKWVRDKIDENPEVKKAIVDKSAGLIDQLLTKKKSEEVNASDFPPPPTDEELRLALPETPMLLGFNAPATSEPSSFEFPPPPTDEELRLALPETPMLLGFNAPATSEPSSFEFPPPPTEDELEIIRETASSLDSSFTRGDLASLRNAINRHSQNFSDFPPIPTEEELNGRGGRP中列出的序列。在另一个实施例中,该ActA片段包含SEQ ID NO:10中列出的序列。在另一个实施例中,该ActA片段为本领域已知的任何其他ActA片段。每种可能性代表了本发明的独立实施例。
在另一个实施例中,编码ActA蛋白的片段的重组核苷酸包含SEQ ID NO:11:
Atgcgtgcgatgatggtggttttcattactgccaattgcattacgattaaccccgacataatatttgcagcgacagatagcgaagattctagtctaaacacagatgaatgggaagaagaaaaaacagaagagcaaccaagcgaggtaaatacgggaccaagatacgaaactgcacgtgaagtaagttcacgtgatattaaagaactagaaaaatcgaataaagtgagaaatacgaacaaagcagacctaatagcaatgttgaaagaaaaagcagaaaaaggtccaaatatcaataataacaacagtgaacaaactgagaatgcggctataaatgaagaggcttcaggagccgaccgaccagctatacaagtggagcgtcgtcatccaggattgccatcggatagcgcagcggaaattaaaaaaagaaggaaagccatagcatcatcggatagtgagcttgaaagccttacttatccggataaaccaacaaaagtaaataagaaaaaagtggcgaaagagtcagttgcggatgcttctgaaagtgacttagattctagcatgcagtcagcagatgagtcttcaccacaacctttaaaagcaaaccaacaaccatttttccctaaagtatttaaaaaaataaaagatgcggggaaatgggtacgtgataaaatcgacgaaaatcctgaagtaaagaaagcgattgttgataaaagtgcagggttaattgaccaattattaaccaaaaagaaaagtgaagaggtaaatgcttcggacttcccgccaccacctacggatgaagagttaagacttgctttgccagagacaccaatgcttcttggttttaatgctcctgctacatcagaaccgagctcattcgaatttccaccaccacctacggatgaagagttaagacttgctttgccagagacgccaatgcttcttggttttaatgctcctgctacatcggaaccgagctcgttcgaatttccaccgcctccaacagaagatgaactagaaatcatccgggaaacagcatcctcgctagattctagttttacaagaggggatttagctagtttgagaaatgctattaatcgccatagtcaaaatttctctgatttcccaccaatcccaacagaagaagagttgaacgggagaggcggtagacca中列出的序列。在另一个实施例中,该重组核苷酸具有SEQ ID NO:11中列出的序列。在另一个实施例中,该重组核苷酸包含任何其他的编码ActA蛋白的片段的序列。每种可能性代表了本发明的独立实施例。
在另一个实施例中,该ActA片段由包含Genbank登录号AF103807中列出的序列的重组核苷酸编码。另一个实施例中,该重组核苷酸具有Genbank登录号AF103807中列出的序列。在另一个实施例中,本发明的方法和组合物的ActA编码序列包含Genbank登录号AF103807中列出的序列。在另一个实施例中,该ActA编码序列为Genbank登录号AF103807的同源物。在另一个实施例中,该ActA编码序列为Genbank登录号AF103807的变体。在另一个实施例中,该ActA编码序列为Genbank登录号AF103807的片段。在另一个实施例中,该ActA编码序列为Genbank登录号AF103807的同种型。每种可能性代表了本发明的独立实施例。在另一个实施例中,截短的ActA为ActA-N100或其修饰形式(称为ActA-N100*),其中PEST基序被删除并含有非保守QDNKR置换,如美国专利公开No.2014/0186387中所描述的。
在另一个实施例中,该ActA片段为本领域已知的任何其他ActA片段。在另一个实施例中,本发明的重组核苷酸包含任何其他的编码ActA蛋白的片段的序列。在另一个实施例中,该重组核苷酸包含任何其他的编码完整ActA蛋白的序列。每种可能性代表了本发明的独立实施例。
在一个实施例中,本文提供的活的减毒李斯特菌或重组李斯特菌表达PEST序列肽。在本发明的方法和组合物的另一个实施例中,PEST AA序列融合至异源抗原或其片段。在另一个实施例中,PEST AA序列为KENSISSMAPPASPPASPKTPIEKKHADEIDK(SEQ ID NO:12)。在另一个实施例中,PEST AA序列为KENSISSMAPPASPPASPK(SEQ ID NO:13)。在另一个实施例中,抗原与包括本文列举的PEST AA序列之一的任何LLO序列的融合可增强细胞介导的针对抗原的免疫。
在另一个实施例中,PEST AA序列为来自李斯特菌ActA蛋白的PEST序列。在另一个实施例中,该PEST序列为KTEEQPSEVNTGPR(SEQ ID NO:14)、KASVTDTSEGDLDSSMQSADESTPQPLK(SEQ ID NO:15)、KNEEVNASDFPPPPTDEELR(SEQ ID NO:16)或RGGIPTSEEFSSLNSGDFTDDENSETTEEEIDR(SEQ ID NO:17)。在另一个实施例中,该PEST序列为上文描述的PEST序列的变体,其在一个实施例中为KESVVDASESDLDSSMQSADESTPQPLK(SEQ ID NO:18)、KSEEVNASDFPPPPTDEELR(SEQ ID NO:19)或RGGRPTSEEFSSLNSGDFTDDENSETTEEEIDR(SEQ ID NO:20),如技术人员将会理解的那样。在另一个实施例中,该PEST序列来自西尔李斯特菌细胞溶素,由lso基因编码。在另一个实施例中,该PEST序列为RSEVTISPAETPESPPATP(SEQ ID NO:21)。在另一个实施例中,该PEST序列来自链球菌属菌种(Streptococcus sp)的链球菌溶血素O蛋白。在另一个实施例中,该PEST序列来自酿脓链球菌(Streptococcus pyogenes)的链球菌溶血素O,例如AA 35-51处的KQNTASTETTTTNEQPK(SEQ ID NO:22)。在另一个实施例中,该PEST序列来自类马链球菌(Streptococcus equisimilis)的链球菌溶血素O,例如AA 38-54处的KQNTASTETTTTNEQPK(SEQ ID NO:23)。在另一个实施例中,该PEST具有选自SEQ ID NO:14-23的序列。在另一个实施例中,该PEST序列为源自原核生物体的另一PEST AA序列。
对PEST氨基酸序列或“PEST序列”的鉴定在本领域是公知的,例如在Rogers S等人(Amino acid sequences common to rapidly degraded proteins:the PEST hypothesis.Science 1986;234(4774):364-8)和Rechsteiner等人(PEST sequences and regulation by proteolysis.Trends Biochem Sci 1996;21(7):267-71)中有描述。“PEST序列”在另一个实施例中指富含脯氨酸(P)、谷氨酸(E)、丝氨酸(S)和苏氨酸(T)残基的区域。在另一个实施例中,该PEST序列的侧翼为一个多个含有几个带正电荷氨基酸的簇。在另一个实施例中,该PEST序列介导含有它的蛋白的快速细胞内降解。在另一个实施例中,该PEST序列适合Rogers等人公开的算法。在另一个实施例中,该PEST序列适合Rechsteiner等人公开的算法。在另一个实施例中,该PEST序列含有一个或多个内部磷酸化位点,这些位点处的磷酸化先于蛋白降解。
在一个实施例中,原核生物体的PEST序列按照例如Rechsteiner和Rogers(1996,Trends Biochem.Sci.21:267-271)针对LM描述的方法和Rogers S等人(Science 1986;234(4774):364-8)描述的方法来鉴定。备选地,来自其他原核生物体的PEST AA序列也可基于这种方法来鉴定。可预期其中PEST AA序列的其他原核生物体包括但不限于其他的李斯特菌菌种。在一个实施例中,该PEST序列适合Rogers等人公开的算法。在另一个实施例中,该PEST序列适合Rechsteiner等人公开的算法。在另一个实施例中,该PEST序列采用PEST-发现程序(PEST-find program)来鉴定。
在另一个实施例中,通过在指定的蛋白序列中初步扫描带正电荷的AA R、H和K来实现对PEST基序的鉴定。对带正电荷侧翼之间的所有AA都计数,且仅进一步考虑这样的基序:其含有的AA数量等于或高于窗口尺寸参数。在另一个实施例中,PEST样序列必须含有至少1个P、1个D或E,以及至少1个S或T。
在另一个实施例中,PEST基序的质量通过基于关键AA的局部富集的得分参数以及该基序的疏水性来优化。D、E、P、S和T的富集以质量百分比(w/w)表示,并相对1当量的D或E、1当量的P和1当量的S或T进行校正。在另一个实施例中,对疏水性的计算按J.Kyte和R.F.Doolittle(Kyte,J和Dootlittle,RF.J.Mol.Biol.157,105(1982))的方法中的原则进行。
在另一个实施例中,潜在的PEST基序的疏水性计算为,每个AA种类的摩尔百分比和疏水性指数的乘积的总和。得到期望的PEST得分,其作为由以下等式表示的局部富集项和疏水性项的组合:
PEST得分=0.55*DEPST-0.5*疏水性指数。
应该理解,术语“PEST氨基酸序列”、“PEST序列”、“PEST样序列”或“PEST样序列肽”可涵盖采用以上算法得分至少为+5的肽。在另一个实施例中,该术语指得分至少为6的肽。在另一个实施例中,该肽具有至少为7的得分。在另一个实施例中,该得分至少为8。在另一个实施例中,该得分至少为9。在另一个实施例中,该得分至少为10。在另一个实施例中,该得分至少为11。在另一个实施例中,该得分至少为12。在另一个实施例中,该得分至少为13。在另一个实施例中,该得分至少为14。在另一个实施例中,该得分至少为15。在另一个实施例中,该得分至少为16。在另一个实施例中,该得分至少为17。在另一个实施例中,该得分至少为18。在另一个实施例中,该得分至少为19。在另一个实施例中,该得分至少为20。在另一个实施例中,该得分至少为21。在另一个实施例中,该得分至少为22。在另一个实施例中,该得分至少为22。在另一个实施例中,该得分至少为24。在另一个实施例中,该得分至少为24。在另一个实施例中,该得分至少为25。在另一个实施例中,该得分至少为26。在另一个实施例中,该得分至少为27。在另一个实施例中,该得分至少为28。在另一个实施例中,该得分至少为29。在另一个实施例中,该得分至少为30。在另一个实施例中,该得分至少为32。在另一个实施例中,该得分至少为35。在另一个实施例中,该得分至少为38。在另一个实施例中,该得分至少为40。在另一个实施例中,该得分至少为45。每种可能性代表了本发明的独立实施例。
在另一个实施例中,使用本领域已知的任何其他方法或算法,例如CaSPredictor(Garay-Malpartida HM,Occhiucci JM,Alves J,Belizario JE.Bioinformatics.2005年6月;21增刊1:i169-76),来鉴定PEST序列。在另一个实施例中,采用以下方法:
通过将1的值分配给AA Ser、Thr、Pro、Glu、Asp、Asn或Gln,计算每个适当长度段(例如30-35个氨基酸的段)的PEST指数。每个PEST残基的系数值(CV)为1,每个其他氨基酸(非-PEST)的系数值为0。
每种用于鉴别PEST样序列的方法代表本发明的单独实施例。
在另一个实施例中,该PEST序列为本领域已知的任何其他PEST序列。每个PEST序列及其类型代表了本发明的独立实施例。
应理解,术语“融合至PEST序列”可以涵盖融合至包含PEST序列的蛋白片段。在另一个实施例中,该术语包括这样的情况:其中该蛋白片段包含除了PEST序列以外的周围序列。在另一个实施例中,该蛋白片段由PEST序列构成。还应当理解,术语“融合”涵盖融合至两个肽或蛋白片段,无论是在它们各自的末端连接在一起还是一个嵌入另一个中。
在另一个实施例中,本文提供了包含本发明的重组形式的李斯特菌的疫苗。
在另一个实施例中,本文提供了本发明的重组形式的李斯特菌的培养物。
另一个实施例为诸如pCR2.1(Invitrogen,La Jolla,CA)的质粒,其为原核表达载体,带有原核复制起点和启动子/调控元件,以促进在原核生物体中表达。在另一个实施例中,除去外来的核苷酸序列以降低质粒大小并增加可置于其中的表达盒的大小。
这样的方法在本领域是公知的,例如在Sambrook等人(1989,Molecular Cloning:A Laboratory Manual,Cold Spring Harbor Laboratory Press,New York)和Ausubei等人(1997,Current Protocols in Molecular Biology,Green&Wiley,New York)中有描述。
抗生素抗性基因用于在分子生物学和疫苗制备中常用的常规选择和克隆过程。在本发明中考虑的抗生素抗性基因包括但不限于这样的基因产物:其赋予对氨苄西林、青霉素、甲氧西林、链霉素、红霉素、卡那霉素、四环素、氯霉素(CAT)、新霉素、潮霉素、庆大霉素和本领域公知的其他抗生素的抗性。每种基因代表了本发明的独立实施例。
用于转化细菌的方法是本领域公知的,并包括基于氯化钙感受态细胞的方法、电穿孔方法、噬菌体介导的转导、化学和物理转化技术(de Boer等人,1989,Cell 56:641-649;Miller等人,1995,FASEB J.,9:190-199;Sambrook等人1989,Molecular Cloning:A Laboratory Manual,Cold Spring Harbor Laboratory,New York;Ausubel等人,1997,Current Protocols in Molecular Biology,John Wiley&Sons,New York;Gerhardt等人编,1994,Methods for General and Molecular Bacteriology,American Society for Microbiology,Washington,DC;Miller,1992,A Short Course in Bacterial Genetics,Cold Spring Harbor Laboratory Press,Cold Spring Harbor,N.Y.)。在另一个实施例中,本发明的李斯特菌疫苗通过电穿孔转化。每种方法代表了本发明的独立实施例。
在另一个实施例中,使用接合将遗传物质和/或质粒引入细菌中。接合的方法是本领域公知的,并例如在Nikodinovic J等人(A second generation snp-derived Escherichia coli-Streptomyces shuttle expression vector that is generally transferable by conjugation.Plasmid.2006年11月;56(3):223-7)和Auchtung JM等人(Regulation of a Bacillus subtilis mobile genetic element by intercellular signaling and the global DNA damage response.Proc Natl Acad Sci U S A.2005年8月30日;102(35):12554-9)中有描述。每种方法代表了本发明的独立实施例。
应该理解,术语“转化”可与术语“转染”等同地使用,并指工程化细菌细胞以吸收质粒或其他异源DNA分子。还应理解,术语“转化”可指工程化细菌细胞以表达质粒或其他异源DNA分子的基因。
在一个实施例中,将可购得的质粒用在本发明中。这些质粒可从各种各样的来源获得,例如Invitrogen(La Jolla,CA)、Stratagene(La Jolla,CA)、Clontech(Palo Alto,CA),或者可采用本领域公知的方法构建。可用在本发明中的质粒和其他表达载体在本文其他地方有描述,并可包括诸如启动子/调控序列、革兰氏阴性和革兰氏阳性细菌的复制起点、编码融合蛋白的分离的核酸以及编码氨基酸代谢基因的分离的核酸等特征。此外,编码融合蛋白和氨基酸代谢基因的分离核酸将具有适合驱动这样的分离核酸的表达的启动子。可用于在细菌系统中驱动表达的启动子是本领域公知的,并包括噬菌体λ、pBR322的β-内酰胺酶基因的bla启动子和pBR325的氯霉素乙酰基转移酶基因的CAT启动子。原核启动子的另外例子包括:噬菌体λ的主要右和左启动子(PL和PR),大肠杆菌的trp、recA、lacZ、lad和gal启动子,枯草芽孢杆菌的α-淀粉酶(Ulmanen等人,1985.J.Bacteriol.162:176-182)和S28特异性启动子(Gilman等人,1984Gene 32:11-20),芽孢杆菌属的噬菌体的启动子(Gryczan,1982,In:The Molecular Biology of the Bacilli,Academic Press,Inc.,New York)以及链霉菌属启动子(Ward等人,1986,Mol.Gen.Genet.203:468-478)。在本发明中考虑的另外的原核启动子例如在Glick(1987,J.Ind.Microbiol.1:277-282)、Cenatiempo(1986,Biochimie,68:505-516)和Gottesman(1984,Ann.Rev.Genet.18:415-442)中有综述。在本发明中考虑的启动子/调控元件的另外例子包括但不限于:李斯特菌prfA启动子、李斯特菌hly启动子、李斯特菌p60启动子和李斯特菌ActA启动子(GenBank登录。号NC_003210)或其片段。
本发明的重组蛋白在另一个实施例中采用重组DNA方法学合成。这在一个实施例中涉及:生成DNA序列,将该DNA置于特定启动子/调控元件控制下的表达盒(例如本发明的质粒)中,以及表达该蛋白。在另一个实施例中,通过任何适当的方法制备编码本发明的重组蛋白(例如非溶血性LLO)的DNA,所述方法例如包括克隆和限制性酶切适当的序列,或者通过以下方法直接化学合成:例如Narang等人(1979,Meth.Enzymol.68:90-99)的磷酸三酯法;Brown等人(1979,Meth.Enzymol 68:109-151)的磷酸二酯法;Beaucage等人(1981,Tetra.Lett.,22:15 1859-1862)的二乙基亚磷酰胺法;以及美国专利No.4,458,066的固相载体法。
在另一个实施例中,将化学合成用于产生单链寡核苷酸。在不同的实施例中,通过与互补序列杂交,或通过将该单链作为模板以DNA聚合酶进行聚合,将该单链的寡核苷酸转化成双链DNA。本领域技术人员会认识到,尽管DNA的化学合成限于约100个碱基的序列,但是通过连接较短的序列可以得到较长的序列。在另一个实施例中,克隆子序列,并使用适当的限制性酶切割适当的子序列。然后连接片段以产生期望的DNA序列。
在另一个实施例中,使用诸如聚合酶链式反应(PCR)的DNA扩增方法克隆编码本发明的重组蛋白的DNA。因此,采用包含合适的限制位点的正义引物和包含另一限制位点(例如用于促进克隆的不同限制位点)的反义引物,PCR扩增非溶血性LLO的基因。
在另一个实施例中,将编码融合蛋白的重组基因可操作地连接至每种宿主的适当的表达控制序列。在本文别处详细描述了启动子/调控序列。在另一个实施例中,所述质粒还包含另外的启动子调控元件,以及核糖体结合位点和转录终止信号。对于真核细胞,该控制序列将包括源自例如免疫球蛋白基因、SV40、巨细胞病毒等的启动子和增强子,以及聚腺苷酸化序列。在另一个实施例中,该序列包括剪接供体和受体序列。
在一个实施例中,术语“可操作地连接”指并列连接,其中如此描述的组件的关系允许它们以其预期方式起作用。与编码序列“可操作地连接”的控制序列是以这样的方式连接,使得在与该控制序列相容的条件下实现该编码序列的表达。
在一个实施例中,本文提供了以本发明的组合物或组合物的混合物给药的方法。在另一个实施例中,本文提供了以本发明的疫苗给药的方法。在另一个实施例中,本文提供了以本发明的免疫治疗组合物给药的方法。在另一个实施例中,本文提供了以本发明的减毒的重组形式李斯特菌给药的方法。
在一个实施例中,由本发明的方法和组合物引起的免疫应答包括CD8+T细胞介导的应答。在另一个实施例中,该免疫应答主要由CD8+T细胞介导的应答构成。在另一个实施例中,该免疫应答的唯一可检测组成部分为CD8+T细胞介导的应答。
在另一个实施例中,由本发明的方法和组合物引起的免疫应答包括CD4+T细胞介导的应答。在另一个实施例中,该免疫应答主要由CD4+T细胞介导的应答构成。在另一个实施例中,该免疫应答的唯一可检测组成部分为CD4+T细胞介导的应答。
在另一个实施例中,由本发明的方法和组合物引起的免疫应答包括固有免疫应答。在另一个实施例中,该免疫应答主要由固有免疫应答构成。在另一个实施例中,该免疫应答的唯一可检测组成部分为固有免疫应答。技术人员应理解,固有免疫应答的激活可以涉及巨噬细胞(例如M1巨噬细胞)、天然杀伤细胞以及树突状细胞(DC)的激活。
在另一个实施例中,本发明提供了降低癌症或传染性疾病或过敏的发生率的方法,其包括以本发明的组合物给药。在另一个实施例中,本发明提供了缓解癌症或传染性疾病或过敏的方法,其包括以本发明的组合物给药。每种可能性代表了本发明的独立实施例。
在一个实施例中,用在本发明中的重组单核细胞增多性李斯特菌分泌异源肽。在另一个实施例中,用在本发明中的重组单核细胞增多性李斯特菌表达异源肽。
在另一个实施例中,用在本发明中的重组单核细胞增多性李斯特菌表达并分泌含PEST的多肽(例如非溶血性LLO),如本文所描述的。
在一个实施例中,本发明的处理方案为治疗性的。在另一个实施例中,该方案为预防性的。在另一个实施例中,将本发明的疫苗用于保护人们免于诸如乳腺癌的癌症或其他类型的肿瘤的风险,因为家族性遗传或其他情况使得它们易于患有这些类型的疾病,如技术人员将理解的那样。在另一个实施例中,将本发明的疫苗用于治疗具有诸如乳腺癌的癌症或其他类型的肿瘤的人,因为家族性遗传或其他情况使得它们易于患有这些类型的疾病,如技术人员将理解的那样。在另一个实施例中,将本发明的疫苗在具有诸如乳腺癌的癌症或其他类型的肿瘤的人中的备选治疗之前或之后使用,因为家族性遗传或其他情况使得它们易于患有这些类型的疾病,如技术人员将理解的那样。在另一个实施例中,这些治疗包括化疗、手术、放射等。在这些治疗后,以本发明的疫苗给药,以致对疫苗的肿瘤抗原的CTL应答破坏剩余的转移并延长该癌症的缓解。在另一个实施例中,将本发明的疫苗用于影响之前建立的肿瘤的生长并杀死现有的肿瘤细胞。每种可能性代表了本发明的独立实施例。
本发明考虑剂量范围的各种实施例。在一个实施例中,对于疫苗载体的情况,该剂量在0.4LD50/剂的范围内。在另一个实施例中,该剂量为约0.4-4.9LD50/剂。在另一个实施例中,该剂量为约0.5-0.59LD50/剂。在另一个实施例中,该剂量为约0.6-0.69LD50/剂。在另一个实施例中,该剂量为约0.7-0.79LD50/剂。在另一个实施例中,该剂量为约0.8LD50/剂。在另一个实施例中,该剂量为约0.4LD50/剂至0.8LD50/剂。
在另一个实施例中,该剂量为107个细菌/剂。在另一个实施例中,该剂量为1.5×107个细菌/剂。在另一个实施例中,该剂量为2×107个细菌/剂。在另一个实施例中,该剂量为3×107个细菌/剂。在另一个实施例中,该剂量为4×107个细菌/剂。在另一个实施例中,该剂量为6×107个细菌/剂。在另一个实施例中,该剂量为8×107个细菌/剂。在另一个实施例中,该剂量为1×108个细菌/剂。在另一个实施例中,该剂量为1.5×108个细菌/剂。在另一个实施例中,该剂量为2×108个细菌/剂。在另一个实施例中,该剂量为3×108个细菌/剂。在另一个实施例中,该剂量为4×108个细菌/剂。在另一个实施例中,该剂量为6×108个细菌/剂。在另一个实施例中,该剂量为8×108个细菌/剂。在另一个实施例中,该剂量为1×109个细菌/剂。在另一个实施例中,该剂量为1.5×109个细菌/剂。在另一个实施例中,该剂量为2×109个细菌/剂。在另一个实施例中,该剂量为3×109个细菌/剂。在另一个实施例中,该剂量为5×109个细菌/剂。在另一个实施例中,该剂量为6×109个细菌/剂。在另一个实施例中,该剂量为8×109个细菌/剂。在另一个实施例中,该剂量为1×1010个细菌/剂。在另一个实施例中,该剂量为1.5×1010个细菌/剂。在另一个实施例中,该剂量为2×1010个细菌/剂。在另一个实施例中,该剂量为3×1010个细菌/剂。在另一个实施例中,该剂量为5×1010个细菌/剂。在另一个实施例中,该剂量为6×1010个细菌/剂。在另一个实施例中,该剂量为8×1010个细菌/剂。在另一个实施例中,该剂量为8×109个细菌/剂。在另一个实施例中,该剂量为1×1011个细菌/剂。在另一个实施例中,该剂量为1.5×1011个细菌/剂。在另一个实施例中,该剂量为2×1011个细菌/剂。在另一个实施例中,该剂量为3×1011个细菌/剂。在另一个实施例中,该剂量为5×1011个细菌/剂。在另一个实施例中,该剂量为6×1011个细菌/剂。在另一个实施例中,该剂量为8×1011个细菌/剂。每种可能性代表了本发明的独立实施例。
当提及本文提供的任何蛋白或肽时,术语“同源性”、“同源的”等在一个实施例中指,在将序列对齐并在必要时引入缺口以达到最大同源性百分比后,且在不将任何保守置换视为序列一致性的一部分的情况下,候选序列中与相应天然多肽的残基相同的氨基酸残基的百分比。用于比对的方法和计算机程序是本领域所公知的。在另一个实施例中,本发明的方法和组合物使用本发明的异源抗原或LLO序列的同源物。
在另一个实施例中,当提及任何核酸序列时,术语“同源性”类似地指候选序列中与相应天然核酸序列的核苷酸相同的核苷酸的百分比。
在另一个实施例中,术语“同源性”指分离的编码本发明的信号肽或重组多肽的核酸与编码本发明的信号肽或重组多肽的核酸共有至少65%同源性。在另一个实施例中,该分离的核酸包含与编码本发明的信号肽或重组多肽的核酸共有至少75%同源性的序列。在另一个实施例中,该分离的核酸包含与编码本发明的信号肽或重组多肽的核酸共有至少85%同源性的序列。在另一个实施例中,该分离的核酸包含与编码本发明的信号肽或重组多肽的核酸共有至少90%同源性的序列。在另一个实施例中,该分离的核酸包含与编码本发明的信号肽或重组多肽的核酸共有至少95%同源性的序列。在另一个实施例中,该分离的核酸包含与编码本发明的信号肽或重组多肽的核酸共有至少97%同源性的序列。在另一个实施例中,该分离的核酸包含与编码本发明的信号肽或重组多肽的核酸共有至少99%同源性的序列。在另一个实施例中,以上同源性范围适用于信号肽或重组多肽的氨基酸序列与本文提供的信号肽或重组多肽的氨基酸序列之间的共有同源性。在另一个实施例中,本文提供的含PEST的多肽为重组多肽。
在一个实施例中,通过序列比对的计算机算法、通过本领域中充分描述的方法来确定同源性。例如,核酸序列同源性的计算机算法分析可以包括使用任何数目的可用软件包,例如,BLAST、DOMAIN、BEAUTY(BLAST Enhanced Alignment Utility)、GENPEPT和TREMBL软件包。
在另一个实施例中,“同源性”指与选自本文提供的序列的序列有大于60%的一致性。在另一个实施例中,“同源性”指与选自本文提供的序列的序列有大于70%的一致性。在另一个实施例中,该一致性大于75%。在另一个实施例中,该一致性大于78%。在另一个实施例中,该一致性大于80%。在另一个实施例中,该一致性大于82%。在另一个实施例中,该一致性大于83%。在另一个实施例中,该一致性大于85%。在另一个实施例中,该一致性大于87%。在另一个实施例中,该一致性大于88%。在另一个实施例中,该一致性大于90%。在另一个实施例中,该一致性大于92%。在另一个实施例中,该一致性大于93%。在另一个实施例中,该一致性大于95%。在另一个实施例中,该一致性大于96%。在另一个实施例中,该一致性大于97%。在另一个实施例中,该一致性大于98%。在另一个实施例中,该一致性大于99%。在另一个实施例中,该一致性为100%。每种可能性代表了本发明的独立实施例。
在另一个实施例中,通过确定候选序列杂交而确定同源性,其方法在现有技术中有充分描述(参见,例如“Nucleic Acid Hybridization"Hames,B.D.和Higgins S.J.编(1985);Sambrook等人,2001,Molecular Cloning,A Laboratory Manual,Cold Spring Harbor Press,N.Y.;以及Ausubel等人,1989,Current Protocols in Molecular Biology,Green Publishing Associates and Wiley Interscience,N.Y)。例如,可在中等至严格条件下进行与编码天然caspase肽的DNA的互补序列杂交的方法。杂交条件例如为在包含下述物质的溶液中在42℃温育过夜:10-20%甲酰胺、5X SSC(150mM NaCl、15mM柠檬酸三钠)、50mM磷酸钠(pH7.6)、5X登哈特溶液、10%硫酸葡聚糖和20μg/ml变性的经剪切的鲑鱼精DNA。
在一个实施例中,通过本领域充分描述的方法确定蛋白和/或肽对本文列出的任何氨基酸序列的同源性,所述方法包括免疫印迹分析,或通过建立的方法采用可获得的多种软件包的任一种通过氨基酸序列的计算机算法分析进行。这些软件包中的一些例如可以包括FASTA、BLAST、MPsrch或Scanps软件包,并且例如可以将Smith和Waterman算法,和/或整体/局部或BLOCKS比对用于分析。每种确定同源性的方法代表了本发明的独立实施例。
在另一个实施例中,本发明提供了包括用于进行本发明的方法的试剂的试剂盒。在另一个实施例中,本发明提供了包括本发明的组合物、工具或仪器的试剂盒。
应该理解,术语“接触”或“给药”可涵盖以本发明的组合物直接接触癌细胞、受试者、肿瘤或疾病部位。在另一个实施例中,该术语指以本发明的组合物间接接触癌细胞、肿瘤或疾病部位。在另一个实施例中,本发明的方法包括这样的方法:其中让受试者与本发明的组合物接触,然后让该组合物通过扩散或本领域已知的任何其他主动运输或被动运输过程(由此使化合物在体内循环)与癌细胞、肿瘤或疾病部位接触。每种可能性代表了本发明的独立实施例。
在另一个实施例中,术语“基因”和“重组基因”指包含编码本发明多肽的开放阅读框的核酸分子。这些天然等位基因变异通常可导致给定基因中核苷酸序列的1-5%差异。可通过在数个不同的个体或生物体中对目的基因测序来鉴定备选等位基因。这可使用杂交探针来鉴定多种个体或生物体中的相同基因座而容易地进行。由天然等位基因变异所造成且不改变功能活性的任何和所有这样的核苷酸变异和产生的氨基酸多态性或变异都旨在落入本发明的范围内。
在另一个实施例中,本发明的组合物通过本领域技术人员已知的任何方法向受试者给药,例如胃肠外、癌旁、透粘膜、透皮、肌肉内、静脉内、真皮内、皮下、腹膜内、心室内、颅内、阴道内或肿瘤内给药。
在本文提供的方法和组合物的另一个实施例中,该疫苗或组合物口服给药,并因此将其被配制为适合口服给药的形式,即,作为固体或液体制备物。适合的固体口服制剂包括片剂、胶囊剂、丸剂、颗粒剂、药丸等。适合的液体口服制剂包括溶液、混悬液、分散剂、乳剂、油剂等。在本发明的另一个实施例中,将活性成分配制在胶囊中。根据该实施例,本发明的组合物除了包含活性化合物和惰性载体或稀释剂以外,还包含明胶胶囊。
在另一个实施例中,通过静脉内、动脉内或肌肉内注射液体制备物来以疫苗或组合物给药。适合的液体制剂包括溶液、混悬液、分散剂、乳剂、油剂等。在一个实施例中,将药物组合物静脉内给药,因此将其配制为适合静脉内给药的形式。在另一个实施例中,将药物组合物动脉内给药,因此将其配制为适合动脉内给药的形式。在另一个实施例中,将药物组合物肌肉内给药,因此将其配制为适合肌肉内给药的形式。
在一些实施例中,当抗体或其功能性片段与包含重组Lm菌株的组合物分开给药时,该抗体可以静脉内注射、皮下注射或直接注射进肿瘤或瘤床中。在一个实施例中,将包含抗体的组合物注射进肿瘤手术摘除后留下的空间内,例如在摘除前列腺肿瘤后的前列腺腺体内的空间。
在一个实施例中,术语“免疫原性组合物”可以涵盖本文提供的重组李斯特菌、佐剂、和抗体或其功能性片段或者它们的组合。在另一个实施例中,免疫原性组合物包含本文提供的重组李斯特菌。在另一个实施例中,免疫原性组合物包含本领域已知的或本文提供的佐剂。还应理解的是,这些组合物的给药增强免疫应答或增加T效应细胞与调节性T细胞比率或者引起抗肿瘤免疫应答,如本文进一步提供的。
遍及本申请,本发明的各个实施例可以以范围的形式表示。应理解,以范围形式描述仅是出于方便和简洁,而不应认为是对发明范围的呆板限制。因此,对范围的描述应认为是已具体公开了所有可能的子范围以及该范围内的各个数值。例如,对诸如1至6的范围的描述应理解为已具体公开了诸如1至3、1至4、1至5、2至4、2至6、3至6等的子范围,以及该范围内的各个数值,例如1、2、3、4、5和6。这种适用性与范围的广度无关。
每当在本文中指出数值范围时,其旨在包括该指出的范围内的任何引用的数字(分数或整数)。短语第一指定数和第二指定数“之间的范围”和“从”第一指定数“至”第二指定数的“范围”在本文中可互换使用,旨在包括第一和第二指定数以及它们之间的所有分数和整数。
技术人员应理解,术语“治疗”可以涵盖治愈疾病、预防疾病、降低疾病的发生率、缓解疾病的症状、诱导疾病的减轻、减缓疾病的进展。术语“减小”、“遏制”和“抑制”在另一个实施例中指减弱或降低。
技术人员应理解,术语“治疗有效剂量”或“治疗有效量”可以涵盖产生期望的作用(为此而给药)的剂量。确切剂量可由本领域技术人员使用已知的技术确定。
技术人员应理解,术语”约”可以涵盖定量项加或减5%,或在另一个实施例中加或减10%,或在另一个实施例中加或减15%,或在另一个实施例中表示加或减20%。
技术人员应理解,术语“受试者”可以涵盖需要治疗病症或其后遗症或者易感于病症或其后遗症的包括人在内的哺乳动物,并且还可以包括狗、猫、猪、牛、绵羊、山羊、马、大鼠和小鼠和人类。应当理解,该术语可以涵盖家畜。术语”受试者”不排除在所有方面都正常的个体。
为了更充分地举例说明本发明的优选实施例,提供了以下实例。但是,它们绝不应解释为限制本发明的广泛范围。
实例
材料和实验方法(实例1-2)
实例1:LLO-抗原融合蛋白诱导抗肿瘤免疫
细胞系
C57BL/6同基因TC-1肿瘤经HPV-16E6和E7永生化并以c-Ha-ras癌基因转化。T.C.Wu(Johns Hopkins University School of Medicine,Baltimore,MD)提供的TC-1为高度致瘤的肺上皮细胞,该细胞表达低水平的HPV-16E6和E7,并被c-Ha-ras癌基因转化。让TC-1于37℃及10%CO2下生长在RPMI 1640、10%FCS、2mM L-谷氨酰胺、100U/ml青霉素、100μg/ml链霉素、100μM非必需氨基酸、1mM丙酮酸钠、50微摩尔(mcM)2-ME、400微克(mcg)/ml G418以及10%国家典型菌种保藏中心-109培养基(National Collection Type Culture-109medium)中。C3为来自C57BL/6小鼠的小鼠胚胎细胞,该细胞经完整HPV 16基因组永生化并经pEJ-ras转化。EL-4/E7为经E7逆转录病毒转导的胸腺瘤EL-4。
单核细胞增多性李斯特菌菌株和繁殖
所用的李斯特菌菌株为Lm-LLO-E7(在游离型表达系统中的hly-E7融合基因;图1A)、Lm-E7(整合进李斯特菌基因组中的单拷贝E7基因盒)、Lm-LLO-NP(“DP-L2028”;在游离型表达系统中的hly-NP融合基因)和Lm-Gag(“ZY-18”;整合进染色体中的单拷贝HIV-1Gag基因盒)。将E7用引物5'-GGCTCGAGCATGGAGATACACC-3'(SEQ ID NO:24;XhoI位点标有下划线)和5'-GGGGACTAGTTTATGGTTTCTGAGAACA-3'(SEQ ID NO:25;SpeI位点标有下划线)通过PCR扩增并连接进pCR2.1(Invitrogen,San Diego,CA)。通过XhoI/SpeI消化从pCR2.1切离E7,并将其连接进pGG-55。将hly-E7融合基因和多能转录因子prfA克隆进pAM401,即一种多拷贝穿梭质粒(Wirth R等人,J Bacteriol,165:831,1986),从而产生pGG-55。hly启动子驱动hly基因产物的前441个氨基酸(缺乏溶血性C-端,在下文中被称作“ΔLLO”)的表达,其通过XhoI位点连接至E7基因,从而产生hly-E7融合基因,该基因被转录和分泌成LLO-E7。用为了在体内保留质粒而选择的pGG-55转化prfA阴性李斯特菌菌株XFL-7(由Hao Shen博士提供,University of Pennsylvania)(图1A-B)。采用引物5'-GGGGGCTAGCCCTCCTTTGATTAGTATATTC-3'(SEQ ID NO:26;NheI位点标有下划线)和5'-CTCCCTCGAGATCATAATTTACTTCATC-3'(SEQ ID NO:27;XhoI位点标有下划线)生成hly启动子和基因片段。采用引物5'-GACTACAAGGACGATGACCGACAAGTGATAACCCGGGATCTAAATAAATCCGTTT-3'(SEQ ID NO:28;XbaI位点标有下划线)和5'-CCCGTCGACCAGCTCTTCTTGGTGAAG-3'(SEQ ID NO:29;SalI位点标有下划线)PCR扩增prfA基因。通过将含有驱动E7的表达和分泌的hly启动子和信号序列的表达盒引入LM基因组的orfZ结构域中,产生Lm-E7。将E7用引物5'-GCGGATCCCATGGAGATACACCTAC-3'(SEQ ID NO:30;BamHI位点标有下划线)和5'-GCTCTAGATTATGGTTTCTGAG-3'(SEQ ID NO:31;XbaI位点标有下划线)通过PCR扩增。然后将E7连接进pZY-21穿梭载体。用得到的质粒pZY-21-E7转化LM菌株10403S,该质粒包括插入在与LM基因组的orfX、Y、Z结构域对应的1.6-kb序列的中央的表达盒。该同源结构域允许E7基因盒通过同源重组插入orfZ结构域中。针对E7基因盒在orfZ结构域中的整合,筛选克隆。在含有(Lm-LLO-E7和Lm-LLO-NP)或不含(Lm-E7和ZY-18)氯霉素(20μg/ml)的脑心浸液培养基中培养细菌。在-80℃以等分试样冷冻细菌。通过Western印迹法验证表达(图2)。
Western印迹法
使李斯特菌菌株在Luria-Bertoni培养基中于37℃生长,并在600nm处测得的相同光密度下收获。将上清液TCA沉淀,并重悬于补充了0.1N NaOH的1x样品缓冲液中。将相同量的每种细胞沉淀物或每种经TCA沉淀的上清液上样至4-20%Tris-甘氨酸SDS-PAGE凝胶(NOVEX,San Diego,CA)。将凝胶转移至聚偏二氟乙烯,并用抗-E7单克隆抗体(mAb)(Zymed Laboratories,South San Francisco,CA)探测,然后与HRP-偶联的抗小鼠二抗(Amersham Pharmacia Biotech,Little Chalfont,U.K.)一起温育,用Amersham ECL检测试剂显影,并暴露于Hyperfilm(Amersham Pharmacia Biotech)。
肿瘤生长的测量
每隔一天用卡尺跨最短和最长表面直径测量肿瘤。将这两个测量结果的平均值作为平均肿瘤直径(以毫米计)相对于不同时间点作图。当肿瘤直径达到20mm时,处死小鼠。仅显示存活小鼠在每个时间点的肿瘤测量结果。
李斯特菌重组体对建立的肿瘤生长的影响
6-8周龄C57BL/6小鼠(Charles River)在左胁腹皮下接受2×105个TC-1细胞。肿瘤接种后一周,肿瘤已经达到直径为4-5mm的可触知大小。然后在第7和14天用0.1LD50腹膜内Lm-LLO-E7(107个CFU)、Lm-E7(106个CFU)、Lm-LLO-NP(107个CFU)或Lm-Gag(5×105个CFU)处理8只小鼠的组。
51Cr释放测定
以0.1LD50Lm-LLO-E7、Lm-E7、Lm-LLO-NP或Lm-Gag腹膜内免疫6-8周龄C57BL/6小鼠。免疫接种后十天,收获脾。将经照射的TC-1细胞作为饲养细胞,在培养物中建立脾细胞(100:1,脾细胞:TC-1);在体外刺激5天,然后用在标准51Cr释放测定中,其中使用下述靶标:EL-4、EL-4/E7或经E7H-2b肽(RAHYNIVTF)(SEQ ID NO:32)脉冲处理的EL-4。一式三份地进行的E:T细胞比率为:80:1、40:1、20:1、10:1、5:1和2.5:1。在37℃温育4h以后,将细胞沉淀,并从每个孔取出50μl上清液。以Wallac 1450闪烁计数器(Gaithersburg,MD)测定样品。将特异性裂解百分比确定为[(每分钟的实验计数(cpm)-自发cpm)/(总cpm-自发cpm)]×100。
TC-1特异性增殖
将C57BL/6小鼠用0.1LD50免疫,并在20天后通过腹膜内注射1LD50Lm-LLO-E7、Lm-E7、Lm-LLO-NP或Lm-Gag进行强化。强化后六天,从经免疫的小鼠和未暴露的小鼠收获脾。以2.5×104、1.25×104、6×103或3×103个经照射的TC-1细胞/孔作为E7Ag的来源,或不用TC-1细胞或用10μg/ml Con A,在平底96-孔平板中以5×105孔在培养物中建立脾细胞。45h后以0.5μCi[3H]胸苷/孔脉冲处理细胞。18h以后,采用Tomtec采集器96(Orange,CT)收获平板,并用Wallac 1450闪烁计数器评估增殖。将以cpm计的变化计算为实验cpm–无Ag cpm。
流式细胞术分析
将C57BL/6小鼠用0.1LD50Lm-LLO-E7或Lm-E7静脉内(i.v.)免疫,并在30天后强化。采用带软件(Becton Dickinson,Mountain View,CA)的流式细胞计对CD8(53-6.7,PE偶联的)、CD62配体(CD62L;MEL-14,APC偶联的)和E7H-2Db四聚体进行三色流式细胞术。将强化后第5天收获的脾细胞在室温(rt)用负载了E7肽(RAHYNIVTF)(SEQ ID NO:32)或对照(HIV-Gag)肽的H-2Db四聚体染色。以1/200稀释度使用四聚体,其由Larry R.Pease博士(Mayo Clinic,Rochester,MN)以及由NIAID Tetramer Core Facility和NIH AIDS Research and Reference Reagent Program提供。分析四聚体+、CD8+、CD62L低细胞。
B16F0-Ova实验
给24只C57BL/6小鼠接种5×105个B16F0-Ova细胞。在第3、10和17天,将8只小鼠的组用0.1LD50Lm-OVA(106个cfu)、Lm-LLO-OVA(108个cfu)免疫,另八只动物保持不处理。
统计
为了对比肿瘤直径,确定每个组的肿瘤大小的平均值和SD,并通过学生t检验确定统计显著性。p≤0.05视作显著的。
结果
对比了Lm-E7和Lm-LLO-E7影响TC-1生长的能力。在C57BL/6小鼠的左胁腹上建立皮下肿瘤。七天后,肿瘤已经达到可触知的大小(4-5mm)。在第7和14天,给小鼠接种0.1LD50Lm-E7、Lm-LLO-E7或作为对照的Lm-Gag和Lm-LLO-NP。Lm-LLO-E7诱导75%的建立的TC-1肿瘤的完全消退,而在该组中的其他2只小鼠中控制了肿瘤生长(图3)。相反,使用Lm-E7和Lm-Gag的免疫接种没有诱导肿瘤消退。该实验重复多次,总是得到非常类似的结果。另外,在不同的免疫方案下,Lm-LLO-E7也得到了类似的结果。在另一实验中,单次免疫能够治愈小鼠建立的5mm TC-1肿瘤。
在其他实验中,用另外2个表达E7的肿瘤细胞系得到了类似的结果:C3和EL-4/E7。为了确认以Lm-LLO-E7接种的效力,分别在第60天或第40天用TC-1或EL-4/E7肿瘤细胞重新挑战已经消除了它们的肿瘤的动物。经Lm-LLO-E7免疫的动物保持无肿瘤直到实验终止(就TC-1而言,第124天;就EL-4/E7而言,第54天)。
因此,抗原作为与ΔLLO的融合蛋白的表达会增强所述抗原的免疫原性。
实例2:LM-LLO-E7处理引起TC-1特异性脾细胞增殖
为了用Lm-LLO-E7测量Lm-E7对T细胞的诱导,在经免疫的小鼠中测量了E7特异性增殖性应答(抗原特异性免疫活性的度量)。来自经Lm-LLO-E7免疫的小鼠的脾细胞当以20:1、40:1、80:1和160:1的脾细胞:TC-1比率暴露于经照射的TC-1细胞(作为E7的来源)时发生增殖(图4)。反之,来自经Lm-E7和rLm对照免疫的小鼠的脾细胞仅表现出背景水平的增殖。
实例3:ActA-E7和PEST-E7融合体赋予抗肿瘤免疫
材料和实验方法
Lm-ActA-E7的构建
Lm-ActA-E7是LM的重组菌株,其包含表达与截短形式的actA蛋白融合的E7蛋白的质粒。通过将质粒载体pDD-1(通过修饰pDP-2028而构建)引入李斯特菌中,产生Lm-actA-E7。pDD-1包含这样的表达盒,其表达:一个拷贝的310bp hly启动子和hly信号序列(ss),其驱动ActA-E7的表达和分泌;1170bp的actA基因,其包含四个PEST序列(SEQ ID NO:11)(该截短的ActA多肽由该分子的前390个氨基酸组成,SEQ ID NO:10);300bp HPV E7基因;1019bp prfA基因(控制毒力基因的表达);以及CAT基因(氯霉素抗性基因),用于选择转化的细菌克隆(Sewell等人(2004),Arch.Otolaryngol.Head Neck Surg.,130:92-97)。
采用引物5'-GGGGTCTAGACCTCCTTTGATTAGTATATTC-3'(Xba I位点标有下划线;SEQ ID NO:33)和引物5'-ATCTTCGCTATCTGTCGCCGCGGCGCGTGCTTCAGTTTGTTGCGC-'3(Not I位点标有下划线,前18个核苷酸是ActA基因重叠;SEQ ID NO:34)从pGG55(实例1)PCR扩增hly启动子(pHly)和基因片段。采用引物5'-GCGCAACAAACTGAAGCAGCGGCCGCGGCGACAGATAGCGAAGAT-3'(NotI位点标有下划线;SEQ ID NO:35)和引物5'-TGTAGGTGTATCTCCATGCTCGAGAGCTAGGCGATCAATTTC-3'(XhoI位点标有下划线;SEQ ID NO:36)从LM 10403s野生型基因组PCR扩增actA基因。采用引物5'-GGAATTGATCGCCTAGCTCTCGAGCATGGAGATACACCTACA-3'(XhoI位点标有下划线;SEQ ID NO:37)和引物5'-AAACGGATTTATTTAGATCCCGGGTTATGGTTTCTGAGAACA-3'(XmaI位点标有下划线;SEQ ID NO:38)从pGG55(pLLO-E7)PCR扩增E7基因。采用引物5'-TGTTCTCAGAAACCATAACCCGGGATCTAAATAAATCCGTTT-3'(XmaI位点标有下划线;SEQ ID NO:39)和引物5'-GGGGGTCGACCAGCTCTTCTTGGTGAAG-3'(SalI位点标有下划线;SEQ ID NO:40)从LM 10403s野生型基因组PCR扩增prfA基因。通过PCR生成hly启动子-actA基因融合体(pHly-actA),并使用上游pHly引物(SEQ ID NO:33)和下游actA引物(SEQ ID NO:36)从纯化的pHly DNA和纯化的actA DNA扩增。
通过PCR产生与prfA基因融合的E7基因(E7-prfA),并使用上游E7引物(SEQ ID NO:37)和下游prfA基因引物(SEQ ID NO:40)从纯化的E7DNA和纯化的prfA DNA扩增。
通过PCR产生与E7-prfA融合产物融合的pHly-actA融合产物,并使用上游pHly引物(SEQ ID NO:33)和下游prfA基因引物(SEQ ID NO:40)从纯化的融合的pHly-actA DNA产物和纯化的融合的E7-prfA DNA产物扩增,并连接进pCRII(Invitrogen,La Jolla,Calif.)。以pCRII-ActAE7转化感受态大肠杆菌(TOP10'F,Invitrogen,La Jolla,Calif.)。裂解和分离以后,使用BamHI(预期的片段大小770bp和6400bp(或当将插入片段逆向插入载体时:2500bp和4100bp))和BstXI(预期的片段大小2800bp和3900bp)通过限制性分析来筛选质粒,并且还使用上游pHly引物(SEQ ID NO:33)和下游prfA基因引物(SEQ ID NO:40)通过PCR分析进行筛选。
将pHly-actA-E7-prfA DNA插入片段通过Xba I和Sal I双重消化从pCRII切离,并连接进也用Xba I和Sal I消化的pDP-2028中。在用表达系统pActAE7转化TOP10'F感受态大肠杆菌(Invitrogen,La Jolla,Calif.)后,使用上游pHly引物(SEQ ID NO:33)和下游PrfA基因引物(SEQ ID NO:40)通过PCR分析来筛选氯霉素抗性克隆。使包含pActAE7的克隆在脑心浸液培养基(含有氯霉素(20mcg(微克)/ml(毫升),Difco,Detroit,Mich.)中生长,并使用midiprep DNA纯化系统试剂盒(Promega,Madison,Wis.)从细菌细胞分离pActAE7。如在Ikonomidis等人(1994,J.Exp.Med.180:2209-2218)中所描述的,用表达系统pActAE7转化青霉素处理的李斯特菌的prfA阴性菌株(菌株XFL-7),并针对体内质粒保留来选择克隆。于37℃在含有氯霉素(20mcg/ml)的脑心浸液中培养克隆。在-80℃在等分试样中冷冻细菌。
抗原表达的免疫印迹验证
为了验证Lm-ActA-E7分泌ActA-E7(约64kD),使李斯特菌菌株在Luria-Bertoni(LB)培养基中在37℃培养。用三氯乙酸(TCA)从培养物上清液中沉淀蛋白,重悬于含有0.1N氢氧化钠的1x样品缓冲液中。将相等量的每种经TCA沉淀的上清液加样至4%至20%Tris-甘氨酸十二烷基硫酸钠-聚丙烯酰胺凝胶(NOVEX,San Diego,Calif)。将凝胶转移至聚偏二氟乙烯膜,并用1:2500抗-E7单克隆抗体(Zymed Laboratories,South San Francisco,Calif)探测,然后用1:5000辣根过氧化物酶偶联的抗小鼠IgG(Amersham Pharmacia Biotech,Little Chalfont,England)探测。将印迹用Amersham增强的化学发光检测试剂显影,并暴露于放射自显影胶片(Amersham)(图5A)。
Lm-PEST-E7、Lm-ΔPEST-E7和Lm-E7epi的构建(图6A)
Lm-PEST-E7与Lm-LLO-E7相同,但是它仅含有hly基因的启动子和PEST序列,具体而言,LLO的前50个氨基酸。为了构建Lm-PEST-E7,使用SOE(通过重叠延伸实现的基因剪接)PCR技术,让hly启动子和PEST区域与全长E7基因融合。从质粒pGG-55(其含有LLO的前441个氨基酸)扩增E7基因和hly-PEST基因片段,并通过常规PCR技术剪接到一起。为了建立最终的质粒pVS16.5,将hly-PEST-E7片段和prfA基因亚克隆进质粒pAM401(其包括用于体外选择的氯霉素抗性基因)中,并使用得到的质粒转化XFL-7。
Lm-ΔPEST-E7是与Lm-LLO-E7相同的重组李斯特菌菌株,但是它缺少PEST序列。基本上如针对Lm-PEST-E7所描述的那样进行制备,但是使用设计成从hly-E7融合基因除去含有PEST的区域(bp 333-387)的引物来构建游离型表达系统。Lm-E7epi是分泌不含PEST区域或LLO的E7的重组菌株。用于转化该菌株的质粒含有与E7基因融合的hly启动子和信号序列的基因片段。该构建体不同于原始的Lm-E7,该Lm-E7表达整合进染色体中的单个拷贝的E7基因。除了表达的E7抗原的形式以外,Lm-E7epi是与Lm-LLO-E7、Lm-PEST-E7和Lm-ΔPEST-E7完全同基因的。
结果
为了对比Lm-ActA-E7相对于Lm-LLO-E7诱导的抗肿瘤免疫,将2×105个TC-1肿瘤细胞皮下植入小鼠中,并让其生长至可触知的大小(大约5毫米[mm])。在第7和14天,用1LD50的Lm-ActA-E7(5×108个CFU)(十字叉)、Lm-LLO-E7(108个CFU)(正方形)或Lm-E7(106个CFU)(圆形)腹膜内免疫小鼠。到第26天,在Lm-LLO-E7和Lm-ActA-E7中的所有动物都没有肿瘤且保持如此,而所有未暴露的动物(三角形)和用Lm-E7免疫的动物生长出大肿瘤(图5B)。因而,使用ActA-E7融合体的接种造成肿瘤消退。
另外,针对它们引起表达E7的肿瘤的消退的能力,对比了Lm-LLO-E7、Lm-PEST-E7、Lm-ΔPEST-E7和Lm-E7epi。在40只C57BL/6小鼠的左胁腹上建立皮下TC-1肿瘤。在肿瘤已经达到4-5mm后,将小鼠分成5组,每组8只。每组用4种重组LM疫苗之一处理,1个组保持不处理。Lm-LLO-E7和Lm-PEST-E7分别在5/8和3/8的病例中诱导了建立的肿瘤的消退。在任何时间点在用Lm-PEST-E7或Lm-LLO-E7处理的小鼠的平均肿瘤大小之间没有统计差异。但是,表达不含PEST序列的E7、Lm-ΔPEST-E7和Lm-E7epi的疫苗在除了一只以外的所有小鼠中没有造成肿瘤消退(图6B,上方插图)。这代表2个实验,其中在用Lm-LLO-E7或Lm-PEST-E7处理的肿瘤与用Lm-E7epi或Lm-ΔPEST-E7处理的肿瘤之间,观察到在第28天时平均肿瘤大小的统计上显著的差异;P<0.001,学生t检验;图6B,下方插图)。另外,在经含有PEST的疫苗接种的小鼠的脾中,在3个实验中可再现地观察到四聚体阳性脾细胞的增加的百分比(图6C)。因而,使用PEST-E7融合体的接种造成肿瘤消退。
实例4:E7与LLO、ActA或PEST样序列的融合增强E7特异性免疫并产生浸润肿瘤的E7特异性CD8+细胞
材料和实验方法
在12只C57BL/6小鼠(n=3)的左胁腹皮下植入500mcl(微升)其包含100微升的在磷酸盐缓冲盐水(PBS)中的2×105个TC-1肿瘤细胞和400微升的(BD Biosciences,Franklin Lakes,N.J.)。在第7、14和21天腹膜内免疫小鼠,并在第28天收获脾和肿瘤。将肿瘤MATRIGEL从小鼠取出,并在含有2毫升(ml)RP 10培养基的试管中在冰上4℃温育过夜。将肿瘤用镊子切碎,切成2mm块,并与3ml酶混合物(PBS中0.2mg/ml胶原酶P、1mg/ml DNAse-1)一起在37℃温育1小时。将组织悬液穿过尼龙网过滤,并用5%胎牛血清+0.05%的NaN3在PBS中的溶液洗涤,以用于四聚体和IFN-γ染色。
在有布雷菲德菌素A存在下以107个细胞/ml将脾细胞和肿瘤细胞与1微摩尔(mcm)E7肽一起温育5小时。将细胞洗涤两次,并在50微升的抗小鼠Fc受体上清液(2.4G2)中4℃温育1小时或过夜。针对表面分子CD8和CD62L将细胞染色,渗透化,使用渗透化试剂盒或(Pharmingen,San Diego,Calif.)固定,并针对IFN-γ染色。使用双激光流式细胞计FACSCalibur获取500,000个事件,并使用Cellquest软件(Becton Dickinson,Franklin Lakes,NJ)分析。计算活化的(CD62L低)CD8+T细胞内的IFN-γ分泌细胞的百分比。
对于四聚体染色,让H-2Db四聚体负载藻红蛋白(PE)偶联的E7肽(RAHYNIVTF,SEQ ID NO:32),在室温染色1小时,并用抗别藻蓝蛋白(APC)偶联的MEL-14(CD62L)和FITC偶联的CD8β在4℃染色30min。通过对比脾中和肿瘤中的四聚体+CD8+CD62L低细胞来分析细胞。
结果
为了分析Lm-ActA-E7增强抗原特异性免疫的能力,给小鼠植入TC-1肿瘤细胞,并用Lm-LLO-E7(1×107个CFU)、Lm-E7(1×106个CFU)或Lm-ActA-E7(2×108个CFU)免疫,或者不处理(未暴露的)。来自Lm-LLO-E7和Lm-ActA-E7组的小鼠肿瘤含有比在Lm-E7或未暴露的小鼠中更高百分比的分泌IFN-γ的CD8+细胞(图7A)和四聚体特异性CD8+细胞(图7B)。
在另一个实验中,给荷瘤小鼠施用Lm-LLO-E7、Lm-PEST-E7、Lm-ΔPEST-E7或Lm-E7epi,并测量肿瘤内的E7特异性淋巴细胞的水平。在第7和14天用0.1LD50的4种疫苗处理小鼠。在第21天收获肿瘤,并用针对CD62L、CD8的抗体和用E7/Db四聚体染色。在接种Lm-LLO-E7和Lm-PEST-E7的小鼠中观察到四聚体阳性的淋巴细胞在肿瘤内增加的百分比(图8A)。该结果在三个实验中是可再现的(图8B)。
因而,Lm-LLO-E7、Lm-ActA-E7和Lm-PEST-E7各自有效地诱导浸润肿瘤的CD8+T细胞和肿瘤消退。
材料和实验方法(实例5-10)
细菌菌株、转化和筛选
采用标准方案,将大肠杆菌菌株MB2159用于转化。通过以H2O洗涤,将细菌细胞制备为用于电穿孔。
大肠杆菌菌株MB2159(Strych U等人,FEMS Microbiol Lett.2001年3月15日;196(2):93-8)为alr(-)/dadX(-)缺陷突变体,其不能合成D-丙氨酸消旋酶。类似地,李斯特菌菌株Lm dal(-)/dat(-)(Lmdd)由于dal和dat基因的部分缺失,也不能合成D-丙氨酸消旋酶。
质粒构建
使用公开的plcA基因序列(Mengaud等人,Infect.Immun.198957,3695-3701),采用PCR来扩增来自染色体DNA的基因。然后使用SalI和XbaI产生的DNA末端将扩增产物连接进pAM401中,以产生pDP1462。
如下构建含有单独prfA的质粒pDP1500:在用XbaI和PstI限制酶切后,从pDP1462删除plcA基因,即碱基429-1349(Mengaud等人,出处同上),用T4DNA聚合酶处理DNA末端以使它们成为平末端,并进行分子内连接。
如下构建含有plcA启动子和plcA的3'末端的一部分的质粒pDP1499:在用PstI和NsiI限制酶切后,从pDP1339删除plcA内部片段,即碱基428-882(Mengaud等人,Infect.Immun.198957,3695-3701),并进行分子内连接。
通过单次连接下述三部分而构建pDP1526(pKSV7::ΔplcA):用BAMHI和XbaI限制酶切的pKSV7;XbaI和NsiI从pAM401::plcA产生的468bp片段,其含有plcA的5'末端(碱基882-1351;Mengaud等人,出处同上);以及PstI-和BamHI从pAM401::plcA prfA产生的501bp片段,其含有plcA的3'末端(碱基77至429;Mengaud等人,出处同上)。
通过EcoRI和PstI双重消化pDP1462,分离出prfA启动子,即碱基1-429(Mengaud等人,出处同上),随后将该片段连接进EcoRI和PstI限制酶切的pKSV7中,以产生pDP1498。将两个随机的HindIII产生的10403S染色体DNA片段(长度为大约3kb)连接进HindIII限制酶切的pKSV7中,以产生随机整合对照质粒pDP1519和pDP1521。
单核细胞增多性李斯特菌突变菌株的构建
从SLCC 5764的Tn917-LTV3库分离出作为具有减少的卵磷脂酶(PC-PLC)的突变体的单核细胞增多性李斯特菌菌株DP-L1387,所述库按之前所描述的进行构建(Camilli等人,J.Bacteriol.1990,172,3738-3744)。如之前所描述的(Sun等人,Infect.Immun.199058,3770-3778),通过对一个转座子-染色体DNA连接处测序来确定Tn917-LTV3插入位点。如之前所描述的(Camilli等人,出处同上),用质粒DNA转化单核细胞增多性李斯特菌。在每ml培养基有10μg氯霉素存在下,施加用于在单核细胞增多性李斯特菌中维持pAM401、pKSV7和它们的衍生物的选择压力。另外,pKSV7衍生物的维持需要在30℃(革兰氏阳性细菌中质粒复制的许可温度)生长。
pKSV7衍生物向单核细胞增多性李斯特菌染色体中的整合,通过质粒上的单核细胞增多性李斯特菌DNA序列和它们的对应染色体等位基因之间的同源重组而发生。通过在每ml培养基含有10μg氯霉素的脑心浸液(BHI)液体培养基中于40℃(pKSV7复制的非许可温度)生长大约30代,富集整合突变体。随后在每ml培养基含有10μg氯霉素的BHI琼脂上集落纯化每个整合菌株,并在40℃温育。从每个整合菌株分离出的染色体DNA的Southern印迹分析证实了整合的质粒的存在。
如上所述,通过pKSV7衍生物pDP1526的整合,实现DP-L1552的构建,以产生部分二倍体中间体。整合的质粒通过分子内同源重组实现的自发切除以低频率发生。通过在BHI液体培养基中于30℃生长大约50代,富集质粒已从染色体切离的细菌。在该步骤中的选择压力的性质是未知的,但是可能是由于含有整合的温度敏感质粒的菌株的轻微生长缺陷。大约50%的切除事件,即,由缺失的3'侧的序列之间的同源重组产生的那些事件,导致ΔplcA对染色体上的野生型等位基因的等位基因交换。
通过使所述细菌在BHI中于40℃生长大约30代,消除切离的质粒。通过以下方面鉴定在染色体上保留ΔplcA等位基因的消除了质粒的细菌:在含有5ml BHI琼脂/2.5%蛋黄/2.5%磷酸盐缓冲盐水(PBS)的覆盖层(BHI/蛋黄琼脂)的BHI琼脂平板上培养以后,它们不能在集落周围产生浑浊带。浑浊带源自蛋黄中PI的PI-PLC水解,从而产生不溶性的二酰甘油沉淀物。通过使用PCR扩增缺失的等位基因和跨缺失测序,证实在单核细胞增多性李斯特菌染色体上的正确plcA缺失。
因而,根据本发明,可以使用PI-PLC阴性突变体(plcA缺失突变体)来产生减毒的单核细胞增多性李斯特菌疫苗。使用相同方法制备其他突变体,即actA缺失突变体、plcB缺失突变体和缺乏plcA和plcB二者的双重突变体,它们也都可以根据本公开内容用于产生减毒的单核细胞增多性李斯特菌疫苗。鉴于本公开内容,本领域技术人员将能够制备除了上述那些以外的其他减毒突变体。
Lmdd的构建
首先借助于染色体基因和温度敏感的穿梭质粒pKSV7之间的双等位基因交换(Smith K等人,Biochimie.1992年7-8月;74(7-8):705-11),将dal基因灭活,该穿梭质粒pKSV7携带在来自原始850-bp dal基因PCR产物的5'末端的450bp片段和来自dal基因PCR产物的3'末端的450bp片段之间的红霉素抗性基因。随后,通过与pKSV7的类似交换反应来构建覆盖82%基因的dal缺失突变体,该pKSV7携带来自期望的缺失周围的完整基因(包括所述基因的上游和下游序列)的5'和3'末端的同源性区域。使用PCR分析确认该染色体缺失的结构。
通过类似的等位基因交换反应,让染色体dat基因失活。将pKSV7修饰成携带450bp片段,该片段通过PCR衍生自完整dat基因(包括该基因的上游和下游序列)的5'和3'末端。通过适当PCR连接这两个片段。该构建体向染色体中的交换导致dat基因的30%中央碱基的缺失,这通过PCR分析进行证实。
李斯特菌的细菌培养物和体内传代
按照标准方法,培养大肠杆菌。使李斯特菌在含有50μg/ml链霉素的LB培养基(Difco,Detroit,MI)中于37℃、250rpm摇动下生长,并在指数生长期中收获。对于Lm-LLOE7,将37μg/ml氯霉素加入培养基中。对于生长动力学确定,使细菌在10ml LB+抗生素中生长16小时。测量OD600nm,并将培养物密度在菌株之间归一化。将培养物1:50稀释进LB+合适的抗生素和D-丙氨酸(如果适用的话)中。
LM在小鼠中的传代
将1×108个CFU腹膜内(i.p.)注射进C57BL/6小鼠。在第三天,分离脾,并在PBS中匀化。将脾悬液的等分试样铺板在含有抗生素(如果适用的话)的LB平板上。扩增几个集落,并混合以建立注射储备液。
不含抗生素抗性因子的质粒pTV3的构建
p60-dal盒的构建。构建不含抗生素抗性基因的载体的第一步是构建截短的p60启动子与dal基因的融合体。通过从LM菌株10403S的基因组PCR扩增,分离出LM丙氨酸消旋酶(dal)基因(正向引物:5'-CCA TGG TGA CAG GCT GGC ATC-3';SEQ ID NO:41)(反向引物:5'-GCT AGC CTA ATG GAT GTA TTT TCT AGG-3';SEQ ID NO:42)和最小p60启动子序列(正向引物:5'-TTA ATT AAC AAA TAG TTG GTA TAG TCC-3';SEQ ID NO:43)(反向引物:5'-GAC GAT GCC AGC CTG TCA CCA TGG AAA ACT CCT CTC-3';SEQ ID NO:44)。这些引物会引入p60序列上游的PacI位点、dal序列下游的NheI位点(限制位点以粗体显示)和p60启动子下游的重叠dal序列(前18bp),以用于随后通过剪接重叠延伸(SOE)PCR来融合p60和dal。截短的p60启动子的序列为:CAAATAGTTGGTATAGTCCTCTTTAGCCTTTGGAGTATTATCTCATCATTTGTTTTTTAGGTGAAAACTGGGTAAACTTAGTATTATCAATATAAAATTAATTCTCAAATACTTAATTACGTACTGGGATTTTCTGAAAAAAGAGAGGAGTTTTCC(SEQ ID NO:45)(Kohler等人,J Bacteriol 173:4668-74,1991)。采用SOE-PCR,将p60和dal的PCR产物融合并克隆进载体pCR2.1(Invitrogen,La Jolla,CA)中。
从pGG55除去抗生素抗性基因。从pGG55除去氯霉素乙酰基转移酶(CAT)基因和引入p60-dal盒的后续克隆策略也会间断地导致革兰氏阳性复制区域的移除(oriRep;Brantl等人,Nucleic Acid Res18:4783-4790,1990)。为了重新引入该革兰氏阳性oriRep,使用在该序列上游添加NarI/EheI位点的5'-引物(GGCGCCACTAACTCAACGCTAGTAG,SEQ ID NO:46)和在该序列下游添加NheI位点的3'-引物(GCTAGCCAGCAAAGAAAAACAAACACG,SEQ ID NO:47)从pGG55PCR扩增oriRep。将PCR产物克隆进克隆载体pCR2.1中,并验证序列。
为了将p60-dal序列并入pGG55载体中,通过PacI/NheI双重消化,从pCR-p60dal切离p60-dal表达盒。通过PCR(引物1,5'-GTC GAC GGT CAC CGG CGC CAC TAA CTC AAC GCT AGT AG-3';SEQ ID NO:48)、(引物2,5'-TTA ATT AAG CTA GCC AGC AAA GAA AAA CAA ACA CG-3';SEQ ID NO:49),从pCR-oriRep扩增pGG55中用于革兰氏阳性细菌的复制区域,以引入另外的EheI和NheI限制位点。将PCR产物连接进pCR2.1-TOPO(Invitrogen,Carlsbad,Calif.)中,并验证序列。通过EheI/NheI消化切离复制区域,并用EheI和NheI双重消化载体pGG55,从而从质粒同时除去两个CAT基因。将两个插入片段即p60-dal和oriRep与pGG55片段连接在一起,产生pTV3(图9)。pTV3也含有prfA(致病性调节因子A)基因。该基因不是pTV3的功能所必需的,但是可以用于这样的情形:其中需要或期望另外的选择标志物。
用于实时PCR的DNA的制备
使用Total DNA试剂盒(Epicentre,Madison,WI),制备全李斯特菌DNA。将李斯特菌在25ml Luria-Bertoni液体培养基(LB)中在37℃和250rpm摇动下培养24小时。通过离心让细菌细胞沉淀,重悬于补充了5mg/ml溶菌酶的PBS中,并在37℃温育20分钟,此后分离DNA。
为了得到用于实时PCR的标准靶DNA,从pGG55PCR扩增LLO-E7基因(5'-ATGAAAAAAATAATGCTAGTTTTTATTAC-3'(SEQ ID NO:50);5'-GCGGCCGCTTAATGATGATGATGATGATGTGGTTTCTG AGAACAGATG-3'(SEQ ID NO:51)),并克隆进载体pETblue1(Novagen,San Diego,CA)。类似地,将plcA扩增子克隆进pCR2.1。分别用pET-LLOE7和pCR-plcA转化大肠杆菌,并制备纯化的质粒DNA用于实时PCR。
实时PCR
采用下列引物:5'-GCAAGTGTGACTCTACGCTTCG-3'(SEQ ID NO:52)、5'-TGCCCATTAACAGGTCTTCCA-3'(SEQ ID NO:53)、5'-FAM-TGCGTA CAAAGCACACACGTAGACATTCGTAC-TAMRA-3'(SEQ ID NO:54)和单拷贝基因plcA(TGACATCGTTTGTGTTTGAGCTAG-3'(SEQ ID NO:55)、5'-GCAGCGCTCTCTATACCAGGTAC-3'(SEQ ID NO:56)、5'-TET-TTAATGTCCATGTTA TGTCTCCGTTATAGCTCATCGTA-TAMRA-3'、SEQ ID NO:57)作为李斯特菌基因组靶标,采用ABI PrimerExpress软件(Applied Biosystems),以E7作为质粒靶标,设计Taqman引物-探针组(Applied Biosystems,Foster City,CA)。
按照生产商的推荐,将0.4μM引物和0.05mM探针与PuRE Taq RTG PCR珠(Amersham,Piscataway,NJ)混合。使用纯化的质粒DNA、pET-LLOE7和pCR-plcA(内标),制备每种靶标的标准曲线,并用于计算未知样品中的基因拷贝数。基于标准曲线计算E7拷贝/plcA拷贝的平均比率,并通过将针对Lmdd-TV3和Lm-LLOE7的结果除以来自Lm-E7(含有整合进基因组中的单个拷贝的E7基因的李斯特菌属菌株)的结果来进行校准。在每个重复三次的qPCR测定中,一式三份地运行所有样品。使用KyPlot软件,通过双因素方差分析(Two-Way ANOVA),分析样品之间的差异。如果p<0.05,则将结果视作统计上显著的。
生长测定
在Luria Bertani(LB)培养基+/-100微克(μg)/ml D-丙氨酸和/或37μg/ml氯霉素中,使细菌在37℃、250rpm摇动下生长。基于OD600nm测量结果,调节起始接种物至所有菌株为相同值。
溶血裂解测定
解冻4×109个CFU的李斯特菌,通过离心(1分钟,14000rpm)进行沉淀,并重悬于100μl含有1M半胱氨酸的PBS(pH5.5)中。将细菌进行1:2系列稀释,并在37℃温育45分钟,以便活化分泌的LLO。将去纤维蛋白的全绵羊血(Cedarlane,Hornby,Ontario,Canada)用5倍体积的PBS洗涤两次,并用6倍体积的PBS-半胱氨酸洗涤三至四次,直到上清液保持澄清,在洗涤步骤之间将细胞以3000x g沉淀8分钟,然后在PBS-半胱氨酸中重悬至10%(v/v)的终浓度。将100μl的10%经洗涤血细胞与100μl李斯特菌悬液混合,并在37℃再温育45分钟。然后通过离心(10分钟,1000x g),沉淀未裂解的血细胞。将100μl上清液转移进新平板,并测定OD530nm,相对于样品稀释度绘图。
Lmdd-Tv3的治疗效果
将105个TC-1(ATCC,Manassas,VA)皮下植入C57BL/6小鼠(n=8)中,并让其生长约7天,此后肿瘤是可触知的。TC-1是经HPV E6和E7永生化并用活化的ras转化的C57BL/6上皮细胞系,其在皮下植入后形成肿瘤。在肿瘤细胞植入后第7和14天,用0.1LD50的适当李斯特菌属菌株免疫小鼠。还包括未免疫的对照组(未暴露的)。用电子卡尺测量肿瘤生长。
ActA缺失突变体的制备
通过毒力因子ActA的不可逆缺失,使菌株Lm dal dat(Lmdd)减毒。构建actA在Lmdaldat(Lmdd)背景下的框内缺失,以避免对下游基因的表达的任何极化效应。Lm dal datΔactA含有在N-端处的前19个氨基酸和C-端的28个氨基酸残基,缺失了ActA的591个氨基酸。使用在actA缺失区域的外部退火的引物,验证该基因在染色体位置中的缺失。这些为如图12中所示的引物3(Adv 305-tgggatggccaagaaattc)(SEQ ID NO:58)和4(Adv304-ctaccatgtcttccgttgcttg)(SEQ ID NO:59)。在分离自Lmdd和Lm-ddΔactA的染色体DNA上进行PCR分析。预期在Lm-dd染色体DNA中用两组不同的引物对1、2和3、4扩增以后的DNA片段的大小为3.0Kb和3.4Kb。但是,对于Lm-ddΔactA,使用引物对1、2和3、4的PCR的预期大小为1.2Kb和1.6Kb。因而,在图12中的PCR分析证实了actA的1.8kb区域在菌株Lm-ddΔactA中的缺失。还对PCR产物进行了DNA测序,以证实含有actA的区域在菌株Lm-ddΔactA中的缺失(图13,SEQ ID NO:60)。
炎性细胞因子的产生:
以诸如Lm prfA-(pGG55)、Lm dal dat、Lm dal dat actA、Lm dal dat actAΔinlC和Lm dal datΔinlC的不同李斯特菌骨架感染巨噬细胞如RAW264.7,并在不同的时间点收获上清液,以使用不同的基于ELISA的试剂盒定量不同细胞因子的水平。所定量的细胞因子包括IFN-γ、TNF-α和IL-6。
体内细胞因子产生:
为了测量体内细胞因子的产生和嗜中性粒细胞的募集,向C57BL/6小鼠腹膜内注射不同的108个CFU的Lm prfA-(pGG55)、Lm dal dat、Lm dal dat actA、Lm dal dat actAΔinlC和Lm dal datΔinlC、李斯特菌或等体积的盐水。12h后,杀死小鼠,用2mL PBS洗涤腹膜腔。在生长培养基上铺板以后,检查腹膜洗液的细菌载量,并分析促炎细胞因子,如MIP-1α、KC、MCP等。采用流式细胞术,在诸如Gr-1、CD11b和F4/80的标志物染色后确定中性粒细胞和巨噬细胞的数量,并采用CellQuest软件进一步定量这些细胞群体。
Transwell迁移测定:
进行该测定,以确定在用inlC缺失菌株感染骨髓源巨噬细胞或树突细胞以后,是否存在嗜中性粒细胞迁移的增加。从诸如C57BL/6的小鼠分离骨髓源巨噬细胞或树突状细胞,并以inlC缺失突变体或对照李斯特菌感染。使用感染的细胞,采用corning costar Transwell平板,建立transwell测定。最初使用3、5或8微米孔transwell平板将测定标准化。为了试验嗜中性粒细胞迁移,将经感染的APC铺板在平板的底部,并将嗜中性粒细胞铺板在室内孔的顶部。在不同的时间点对细胞计数,以确定已迁移至底部的嗜中性粒细胞的数目。
Lm dal dat actAΔinlC突变体的治疗效果:
为了确定inlC突变体的治疗效果,使用人前列腺特异性抗原(PSA)作为肿瘤抗原,用作概念性验证。以含有人PSA表达盒的质粒pAdv142转化骨架Lm dal dat actA inlC,得到LmddAinlC142。对菌株LmddAinlC142表征融合蛋白tLLO-PSA的表达和分泌。此外,将菌株LmddAinlC142在小鼠体内传代两次,并针对融合蛋白tLLO-PSA的表达和分泌,检查在两次体内传代以后得到的集落。从第二次体内传代后得到的集落制备疫苗工作储备液,并将其用于评估治疗效果和免疫原性。-
对肿瘤微环境的影响:
确定LmddA、LmddAΔactA、LmddAΔPlcA、LmddAΔPlcB、LmddAΔprfA、LmddAinlC142、LmddA142以及其他对照菌株引起免疫细胞在肿瘤微环境中浸润的能力。在该研究中,在第0天向小鼠接种1×106个TPSA23肿瘤细胞,并在第7、14和21天接种108个CFU的LmddAinlC142、LmddA142和其他对照菌株。在第28天收获肿瘤,并进行处理以用于以诸如Gr-1、CD11b、CD3、CD4、CD8、CD25、Foxp3、NK1.1和CD62L的不同细胞表面标志物进一步染色。采用这些标志物,所检查的细胞群包括巨噬细胞(CD11b+)、NK细胞(NK1.1+)、中性粒细胞(Gr-1+CD11b+)、髓源性抑制细胞(MDSC)(Gr-1+CD11b+)、调节性T细胞(CD4+CD25+Foxp3+)以及效应T细胞(CD8+CD3+CD62L低)。此外,表征了效应T细胞产生诸如IFN-γ、TNF-α和IL-2的效应细胞因子的功能性能力。测试了瘤内调节性T细胞和MDSC造成T细胞增殖受到抑制的能力。
结果
实例5:含有氨基酸代谢酶而不是抗生素抗性基因的质粒在体外和在体内都被保留在大肠杆菌和LM中
使用基于D-丙氨酸消旋酶的营养缺陷型互补系统来介导质粒在LM中的保留,而不使用抗生素抗性基因。大肠杆菌菌株MB2159是不能合成D-丙氨酸消旋酶的alr(-)/dadX(-)缺陷型突变体。类似地,李斯特菌菌株Lm dal(-)/dat(-)(Lmdd)由于dal和dat基因的部分缺失,也不能合成D-丙氨酸消旋酶。通过除去两个CAT基因,并用在李斯特菌p60启动子控制下的p60-dal表达盒替换它们,来修饰基于大肠杆菌-李斯特菌穿梭载体pAM401的质粒pGG55,以产生pTV3(图9)。从几个集落纯化DNA。
实例6:含有代谢酶的质粒不增加细菌的毒力
因为毒力与LLO功能有关,所以对比了Lmdd-TV3和Lm-LLOE7之间的溶血裂解活性。该测定通过红血细胞的裂解来测试LLO功能,且可用培养物上清液、纯化的LLO或细菌细胞进行。Lmdd-TV3展示出比Lm-LLOE7更高的溶血裂解活性。
还通过确定LD50值来测量体内毒力,这是一种更直接的因此更准确的测量毒力的方式。Lmdd-TV3(0.75×109)的LD50非常接近于Lm-LLOE7(1×109)的,表明含有代谢酶的质粒不增加细菌毒力。
实例7:通过含有代谢酶的质粒诱导抗肿瘤免疫
在肿瘤消退模型中确定了含有代谢酶的质粒作为癌症疫苗的效力。使用TC-1细胞系模型,其针对HPV疫苗开发进行了充分表征,且允许受控地对比在Lmdd-TV3或Lm-LLOE7免疫以后建立的类似大小的肿瘤的消退。在两个分开的实验中,Lmdd-TV3和Lm-LLOE7对小鼠的免疫导致类似的肿瘤消退(图14),在接种疫苗的组之间没有统计上显著的差异(p<0.05)。所有经免疫的小鼠在63天后仍然存活,而未经免疫的小鼠在它们的肿瘤达到20mm直径时必须处死。直至实验结束,治愈的小鼠保持无肿瘤。
因而,含有代谢酶的质粒作为治疗性癌症疫苗是有效的。因为治疗性癌症疫苗需要的免疫应答强于预防性癌症疫苗需要的免疫应答,所以这些结果证实了作为预防性癌症疫苗的同样的实用性。
实例8:inlC缺失突变体产生显著高水平的趋化因子和细胞因子。
inlC缺失突变体产生显著高水平的趋化因子,如MIP-1α、KC(IL-8的小鼠同系物)、MCP,从而导致嗜中性粒细胞和白细胞向感染部位的浸润。因此,当腹膜内施用不同的李斯特菌菌株时,inlC突变体表现出这些细胞因子和趋化因子的产生的增加,它们将嗜中性粒细胞和巨噬细胞吸引到注射后12h得到的腹膜液中。此外,当与对照菌株相比,inlC缺失突变体产生显著高水平的炎性细胞因子。
实例9:inlC缺失突变体诱导嗜中性粒细胞迁移
当与其他对照菌株比较,被inlC缺失突变体感染的巨噬细胞在不同的时间点表现出嗜中性粒细胞迁移的显著增加。该实验的结果有力地支持了该菌株在感染过程中吸引诸如嗜中性粒细胞的免疫细胞的能力。
实例10:inlC缺失突变体实现治疗性抗肿瘤应答
使用LmddA142和LmddAinlC142的抗肿瘤研究的结果彼此非常相当,且观察到肿瘤的治疗性消退。此外,两剂LmddAinlC142与三剂菌株LmddA142相当,因为它能够产生高水平的先天性应答并增加促炎细胞因子分泌。
材料与方法(实例11-16)
寡核苷酸由Invitrogen(Carlsbad,CA)合成,并且由Genewiz Inc,South Plainfield,NJ进行DNA测序。流式细胞术试剂购自Becton Dickinson Biosciences(BD,San Diego,CA)。除非指出,否则细胞培养基、补充物和所有其他试剂都得自Sigma(St.Louise,MO)。由EZbiolabs(Westfield,IN)合成Her2/neu HLA-A2肽。完全RPMI 1640(C-RPMI)培养基含有2mM谷氨酰胺、0.1mM非必需氨基酸和1mM丙酮酸钠、10%胎牛血清、青霉素/链霉素、Hepes(25mM)。之前描述了多克隆抗LLO抗体,抗Her2/neu抗体购自Sigma。
小鼠和细胞系
根据IACUC核准的方案在University of Pennsylvania或Rutgers University进行所有动物实验。FVB/N小鼠购自Jackson laboratories(Bar Harbor,ME)。在University of Pennsylvania的动物中心机构安置和饲养FVB/N Her2/neu转基因小鼠,其过表达大鼠Her2/neu肿瘤蛋白。NT-2肿瘤细胞系表达高水平的大鼠Her2/neu蛋白,源自这些小鼠中的自发性乳腺肿瘤,并按之前的描述生长。DHFR-G8(3T3/neu)细胞得自ATCC,并根据ATCC建议生长。EMT6-Luc细胞系是得自John Ohlfest博士(University of Minnesota,MN)的慷慨礼物,在完全C-RPMI培养基中生长。在University of Pennsylvania(Philadelphia,PA)的小动物成像机构(Small Animal Imaging Facility(SAIF))的指导下进行生物发光工作。
李斯特菌构建体和抗原表达
Her2/neu-pGEM7Z由University of Pennsylvania的Mark Greene博士友好提供,并含有克隆进pGEM7Z质粒(Promega,Madison WI)中的全长人Her2/neu(hHer2)基因。将该质粒用作模板,以使用表2所示的pfx DNA聚合酶(Invitrogen)和寡聚物,通过PCR扩增hHer-2/neu的三个区段,即EC1、EC2和IC1。
表2:用于克隆人her-2嵌合体的引物
通过SOEing PCR方法,并将每个单独的hHer-2/neu区段作为模板,通过直接融合产生Her-2/neu嵌合构建体。引物显示在表3中。
用于扩增不同区段人Her2区域的引物的序列。
用于扩增不同区段人Her2区域的引物的序列。
使用XhoI和SpeI限制性酶从pAdv138切离ChHer2基因,并将其与LLO的截短的、非溶血性片段同框克隆进Lmdd穿梭载体pAdv134中。通过DNA测序分析,确认插入片段、LLO和hly启动子的序列。将该质粒电穿孔进电感受态的actA、dal、dat突变体单核细胞增多性李斯特菌菌株中,在含有链霉素(250μg/ml)的脑心浸液(BHI)琼脂平板上选择LmddA和阳性克隆。在一些实验中,表达hHer2/neu(Lm-hHer2)片段的类似李斯特菌菌株用于对比目的。这些在之前有描述。在所有研究中,都包括不相关的李斯特菌构建体(Lm对照),以解释李斯特菌对免疫系统的抗原非依赖性作用。Lm对照基于与ADXS31-164相同的李斯特菌平台,但表达不同的抗原,如HPV16-E7或NY-ESO-1。测试了来自李斯特菌的融合蛋白的表达和分泌。每种构建体都在体内传代两次。
细胞毒性测定
将3-5只FVB/N小鼠的组以一周间隔用1×108个集落形成单位(CFU)的Lm-LLO-ChHer2、ADXS31-164、Lm-hHer2ICI或Lm对照(表达不相关抗原)免疫三次或保持未暴露的。使NT-2细胞在体外生长,用胰蛋白酶分离,并用丝裂霉素C(250μg/ml,在无血清C-RPMI培养基中)在37℃处理45分钟。洗涤5次之后,将它们与从免疫动物或未暴露的动物收获的脾细胞以1:5的(刺激物:应答者)比率一起在37℃和5%CO2下共温育5天。使用铕标记的3T3/neu(DHFR-G8)细胞作为靶标,根据之前描述的方法,进行标准细胞毒性测定。在4小时温育之后,使用分光光度计(Perkin Elmer,Victor2)在590nm处测量从杀死的靶细胞释放的铕。将特异性裂解百分比定义为(实验组的裂解-自发裂解)/(最大裂解-自发裂解)。
来自免疫小鼠的脾细胞的干扰素γ分泌
将3-5只FVB/N或HLA-A2转基因小鼠的组以一周间隔用1×108个CFU的ADXS31-164(即阴性李斯特菌对照(表达不相关抗原))免疫三次或保持未暴露的。最后一次免疫接种之后一周,从FVB/N小鼠分离脾细胞,并且以5×106个细胞/孔在24孔板中C-RPMI培养基内在经丝裂霉素C处理的NT-2细胞存在下共培养。在1μM HLA-A2特异性肽或1μg/ml重组His-标记的ChHer2蛋白(大肠杆菌中产生并且通过基于镍的亲和色谱系统纯化)存在下,温育来自HLA-A2转基因小鼠的脾细胞。24或72小时之后,从上清液获得样品,并使用小鼠IFN-γ酶联免疫吸附测定(ELISA)试剂盒根据制造商的建议测试干扰素-γ(IFN-γ)的存在。
Her2转基因动物中的肿瘤研究
用5×108个CFU的Lm-LLO-ChHer2、ADXS31-164或Lm对照免疫六周龄FVB/N大鼠Her2/neu转基因小鼠(9-14只/组)6次。一周两次观察它们的自发性乳腺肿瘤的出现,使用电子卡尺测量,直至52周。当逃逸的肿瘤达到平均直径1cm2大小时将其切除,并且在-20℃保存于RNAlater中。为了确定Her2/neu蛋白中的突变对这些肿瘤逃逸的影响,使用基因组DNA分离试剂盒提取基因组DNA并且测序。
ADXS31-164对脾和肿瘤中的调节性T细胞的影响
向小鼠皮下(s.c.)植入1×106个NT-2细胞。在第7、14和21天,用1×108个CFU的ADXS31-164、LmddA对照免疫它们或保持未暴露的。在第28天提取肿瘤和脾,并且通过FACS分析来测试CD3+/CD4+/FoxP3+Treg的存在。简而言之,通过在C-RPMI培养基中在两个载玻片之间匀化脾,分离脾细胞。使用无菌剃刀刀片切碎肿瘤,并用在PBS中含有DNA酶(12U/ml)和胶原酶(2mg/ml)的缓冲液消化。在室温下搅拌温育60min之后,通过强力吹打而分离细胞。用RBC裂解缓冲液裂解红血细胞,随后用含有10%FBS的完全RPMI-1640培养基洗涤数次。穿过尼龙网过滤之后,将肿瘤细胞和脾细胞重悬于FACS缓冲液(2%FBS/PBS)中,并用抗CD3-PerCP-Cy5.5、CD4-FITC、CD25-APC抗体染色,随后进行透化,并用抗Foxp3-PE染色。使用4色FACS calibur(BD)进行流式细胞术分析,并使用cell quest软件(BD)分析数据。
统计分析
将对数秩卡方检验用于存活数据,并将学生t检验用于CTL和ELISA测定,所述测定一式三份地进行。在这些分析中,小于0.05的p值(标记为*)被认为是统计上显著的。所有统计分析均用Prism软件V.4.0a(2006)或SPSS软件V.15.0(2006)进行。除非另有说明,否则对于所有的FVB/N大鼠Her2/neu转基因研究,我们均使用8-14只小鼠/组,对于所有野生型FVB/N研究,我们均使用至少8只小鼠/组。除了在Her2/neu转基因小鼠模型中的长期肿瘤研究以外,所有研究重复至少一次。
结果
实例11:分泌与Her-2片段融合的LLO片段的单核细胞增多性李斯特菌菌株的制备:ADXS31-164的构建。
之前描述了嵌合的Her2/neu基因(ChHer2)的构建。简而言之,通过SOEing PCR方法,通过直接融合Her2/neu蛋白的两个胞外片段(氨基酸40-170和氨基酸359-433)和一个胞内片段(氨基酸678-808),制备ChHer2基因。该嵌合蛋白具有该蛋白的已知人MHC I类表位的大部分。从质粒pAdv138(其用于构建Lm-LLO-ChHer2)切离ChHer2基因,并克隆进LmddA穿梭质粒,从而产生质粒pAdv164(图15A)。在这两个质粒骨架之间存在两大差异。1)尽管pAdv138使用氯霉素抗性标记(cat)用于重组细菌的体外选择,但是pAdv164具有来自枯草芽孢杆菌(bacillus subtilis)的D-丙氨酸消旋酶基因(dal),其使用代谢互补路径进行体外选择并在缺乏dal-dat基因的LmddA菌株中进行体内质粒保持。设计和开发该疫苗平台,以解决FDA关于工程化李斯特菌疫苗菌株的抗生素抗性的担忧。2)不同于pAdv138,pAdv164在质粒中没有prfA基因拷贝(参见下面的序列和图15A),因为这对于Lmdd菌株的体内互补而言不是必需的。LmddA疫苗菌株也缺乏actA基因(负责李斯特菌的胞内运动和细胞至细胞扩散),所以源自该骨架的重组疫苗菌株的毒力比源自其亲代菌株Lmdd的毒力小100倍。基于LmddA的疫苗也远比来自免疫小鼠脾的基于Lmdd的疫苗更快地清除(短于48小时)。体外生长8小时之后经TCA沉淀的细胞培养物上清液中,来自该菌株的融合蛋白tLLO-ChHer2的表达和分泌与Lm-LLO-ChHer2的表达和分泌相当(图15B),因为使用Western印迹分析通过抗LLO抗体检测到~104KD的带。将仅表达tLLO的李斯特菌骨架菌株用作阴性对照。
pAdv164序列(7075个碱基对)(参加图15):
cggagtgtatactggcttactatgttggcactgatgagggtgtcagtgaagtgcttcatgtggcaggagaaaaaaggctgcaccggtgcgtcagcagaatatgtgatacaggatatattccgcttcctcgctcactgactcgctacgctcggtcgttcgactgcggcgagcggaaatggcttacgaacggggcggagatttcctggaagatgccaggaagatacttaacagggaagtgagagggccgcggcaaagccgtttttccataggctccgcccccctgacaagcatcacgaaatctgacgctcaaatcagtggtggcgaaacccgacaggactataaagataccaggcgtttccccctggcggctccctcgtgcgctctcctgttcctgcctttcggtttaccggtgtcattccgctgttatggccgcgtttgtctcattccacgcctgacactcagttccgggtaggcagttcgctccaagctggactgtatgcacgaaccccccgttcagtccgaccgctgcgccttatccggtaactatcgtcttgagtccaacccggaaagacatgcaaaagcaccactggcagcagccactggtaattgatttagaggagttagtcttgaagtcatgcgccggttaaggctaaactgaaaggacaagttttggtgactgcgctcctccaagccagttacctcggttcaaagagttggtagctcagagaaccttcgaaaaaccgccctgcaaggcggttttttcgttttcagagcaagagattacgcgcagaccaaaacgatctcaagaagatcatcttattaatcagataaaatatttctagccctcctttgattagtatattcctatcttaaagttacttttatgtggaggcattaacatttgttaatgacgtcaaaaggatagcaagactagaataaagctataaagcaagcatataatattgcgtttcatctttagaagcgaatttcgccaatattataattatcaaaagagaggggtggcaaacggtatttggcattattaggttaaaaaatgtagaaggagagtgaaacccatgaaaaaaataatgctagtttttattacacttatattagttagtctaccaattgcgcaacaaactgaagcaaaggatgcatctgcattcaataaagaaaattcaatttcatccatggcaccaccagcatctccgcctgcaagtcctaagacgccaatcgaaaagaaacacgcggatgaaatcgataagtatatacaaggattggattacaataaaaacaatgtattagtataccacggagatgcagtgacaaatgtgccgccaagaaaaggttacaaagatggaaatgaatatattgttgtggagaaaaagaagaaatccatcaatcaaaataatgcagacattcaagttgtgaatgcaatttcgagcctaacctatccaggtgctctcgtaaaagcgaattcggaattagtagaaaatcaaccagatgttctccctgtaaaacgtgattcattaacactcagcattgatttgccaggtatgactaatcaagacaataaaatagttgtaaaaaatgccactaaatcaaacgttaacaacgcagtaaatacattagtggaaagatggaatgaaaaatatgctcaagcttatccaaatgtaagtgcaaaaattgattatgatgacgaaatggcttacagtgaatcacaattaattgcgaaatttggtacagcatttaaagctgtaaataatagcttgaatgtaaacttcggcgcaatcagtgaagggaaaatgcaagaagaagtcattagttttaaacaaatttactataacgtgaatgttaatgaacctacaagaccttccagatttttcggcaaagctgttactaaagagcagttgcaagcgcttggagtgaatgcagaaaatcctcctgcatatatctcaagtgtggcgtatggccgtcaagtttatttgaaattatcaactaattcccatagtactaaagtaaaagctgcttttgatgctgccgtaagcggaaaatctgtctcaggtgatgtagaactaacaaatatcatcaaaaattcttccttcaaagccgtaatttacggaggttccgcaaaagatgaagttcaaatcatcgacggcaacctcggagacttacgcgatattttgaaaaaaggcgctacttttaatcgagaaacaccaggagttcccattgcttatacaacaaacttcctaaaagacaatgaattagctgttattaaaaacaactcagaatatattgaaacaacttcaaaagcttatacagatggaaaaattaacatcgatcactctggaggatacgttgctcaattcaacatttcttgggatgaagtaaattatgatctcgagacccacctggacatgctccgccacctctaccagggctgccaggtggtgcagggaaacctggaactcacctacctgcccaccaatgccagcctgtccttcctgcaggatatccaggaggtgcagggctacgtgctcatcgctcacaaccaagtgaggcaggtcccactgcagaggctgcggattgtgcgaggcacccagctctttgaggacaactatgccctggccgtgctagacaatggagacccgctgaacaataccacccctgtcacaggggcctccccaggaggcctgcgggagctgcagcttcgaagcctcacagagatcttgaaaggaggggtcttgatccagcggaacccccagctctgctaccaggacacgattttgtggaagaatatccaggagtttgctggctgcaagaagatctttgggagcctggcatttctgccggagagctttgatggggacccagcctccaacactgccccgctccagccagagcagctccaagtgtttgagactctggaagagatcacaggttacctatacatctcagcatggccggacagcctgcctgacctcagcgtcttccagaacctgcaagtaatccggggacgaattctgcacaatggcgcctactcgctgaccctgcaagggctgggcatcagctggctggggctgcgctcactgagggaactgggcagtggactggccctcatccaccataacacccacctctgcttcgtgcacacggtgccctgggaccagctctttcggaacccgcaccaagctctgctccacactgccaaccggccagaggacgagtgtgtgggcgagggcctggcctgccaccagctgtgcgcccgagggcagcagaagatccggaagtacacgatgcggagactgctgcaggaaacggagctggtggagccgctgacacctagcggagcgatgcccaaccaggcgcagatgcggatcctgaaagagacggagctgaggaaggtgaaggtgcttggatctggcgcttttggcacagtctacaagggcatctggatccctgatggggagaatgtgaaaattccagtggccatcaaagtgttgagggaaaacacatcccccaaagccaacaaagaaatcttagacgaagcatacgtgatggctggtgtgggctccccatatgtctcccgccttctgggcatctgcctgacatccacggtgcagctggtgacacagcttatgccctatggctgcctcttagactaatctagacccgggccactaactcaacgctagtagtggatttaatcccaaatgagccaacagaaccagaaccagaaacagaacaagtaacattggagttagaaatggaagaagaaaaaagcaatgatttcgtgtgaataatgcacgaaatcattgcttatttttttaaaaagcgatatactagatataacgaaacaacgaactgaataaagaatacaaaaaaagagccacgaccagttaaagcctgagaaactttaactgcgagccttaattgattaccaccaatcaattaaagaagtcgagacccaaaatttggtaaagtatttaattactttattaatcagatacttaaatatctgtaaacccattatatcgggtttttgaggggatttcaagtctttaagaagataccaggcaatcaattaagaaaaacttagttgattgccttttttgttgtgattcaactttgatcgtagcttctaactaattaattttcgtaagaaaggagaacagctgaatgaatatcccttttgttgtagaaactgtgcttcatgacggcttgttaaagtacaaatttaaaaatagtaaaattcgctcaatcactaccaagccaggtaaaagtaaaggggctatttttgcgtatcgctcaaaaaaaagcatgattggcggacgtggcgttgttctgacttccgaagaagcgattcacgaaaatcaagatacatttacgcattggacaccaaacgtttatcgttatggtacgtatgcagacgaaaaccgttcatacactaaaggacattctgaaaacaatttaagacaaatcaataccttctttattgattttgatattcacacggaaaaagaaactatttcagcaagcgatattttaacaacagctattgatttaggttttatgcctacgttaattatcaaatctgataaaggttatcaagcatattttgttttagaaacgccagtctatgtgacttcaaaatcagaatttaaatctgtcaaagcagccaaaataatctcgcaaaatatccgagaatattttggaaagtctttgccagttgatctaacgtgcaatcattttgggattgctcgtataccaagaacggacaatgtagaattttttgatcccaattaccgttattctttcaaagaatggcaagattggtctttcaaacaaacagataataagggctttactcgttcaagtctaacggttttaagcggtacagaaggcaaaaaacaagtagatgaaccctggtttaatctcttattgcacgaaacgaaattttcaggagaaaagggtttagtagggcgcaatagcgttatgtttaccctctctttagcctactttagttcaggctattcaatcgaaacgtgcgaatataatatgtttgagtttaataatcgattagatcaacccttagaagaaaaagaagtaatcaaaattgttagaagtgcctattcagaaaactatcaaggggctaatagggaatacattaccattctttgcaaagcttgggtatcaagtgatttaaccagtaaagatttatttgtccgtcaagggtggtttaaattcaagaaaaaaagaagcgaacgtcaacgtgttcatttgtcagaatggaaagaagatttaatggcttatattagcgaaaaaagcgatgtatacaagccttatttagcgacgaccaaaaaagagattagagaagtgctaggcattcctgaacggacattagataaattgctgaaggtactgaaggcgaatcaggaaattttctttaagattaaaccaggaagaaatggtggcattcaacttgctagtgttaaatcattgttgctatcgatcattaaattaaaaaaagaagaacgagaaagctatataaaggcgctgacagcttcgtttaatttagaacgtacatttattcaagaaactctaaacaaattggcagaacgccccaaaacggacccacaactcgatttgtttagctacgatacaggctgaaaataaaacccgcactatgccattacatttatatctatgatacgtgtttgtttttctttgctggctagcttaattgcttatatttacctgcaataaaggatttcttacttccattatactcccattttccaaaaacatacggggaacacgggaacttattgtacaggccacctcatagttaatggtttcgagccttcctgcaatctcatccatggaaatatattcatccccctgccggcctattaatgtgacttttgtgcccggcggatattcctgatccagctccaccataaattggtccatgcaaattcggccggcaattttcaggcgttttcccttcacaaggatgtcggtccctttcaattttcggagccagccgtccgcatagcctacaggcaccgtcccgatccatgtgtctttttccgctgtgtactcggctccgtagctgacgctctcgccttttctgatcagtttgacatgtgacagtgtcgaatgcagggtaaatgccggacgcagctgaaacggtatctcgtccgacatgtcagcagacgggcgaaggccatacatgccgatgccgaatctgactgcattaaaaaagccttttttcagccggagtccagcggcgctgttcgcgcagtggaccattagattctttaacggcagcggagcaatcagctctttaaagcgctcaaactgcattaagaaatagcctctttctttttcatccgctgtcgcaaaatgggtaaatacccctttgcactttaaacgagggttgcggtcaagaattgccatcacgttctgaacttcttcctctgtttttacaccaagtctgttcatccccgtatcgaccttcagatgaaaatgaagagaaccttttttcgtgtggcgggctgcctcctgaagccattcaacagaataacctgttaaggtcacgtcatactcagcagcgattgccacatactccgggggaaccgcgccaagcaccaatataggcgccttcaatccctttttgcgcagtgaaatcgcttcatccaaaatggccacggccaagcatgaagcacctgcgtcaagagcagcctttgctgtttctgcatcaccatgcccgtaggcgtttgctttcacaactgccatcaagtggacatgttcaccgatatgttttttcatattgctgacattttcctttatcgcggacaagtcaatttccgcccacgtatctctgtaaaaaggttttgtgctcatggaaaactcctctcttttttcagaaaatcccagtacgtaattaagtatttgagaattaattttatattgattaatactaagtttacccagttttcacctaaaaaacaaatgatgagataatagctccaaaggctaaagaggactataccaactatttgttaattaa(SEQ ID NO:73)
实例12:ADXS31-164的免疫原性如同LM-LLO-ChHER2。
在标准CTL测定中,将ADXS31-164在产生抗Her2/neu特异性细胞毒性T细胞方面的免疫原性性质与Lm-LLO-ChHer2疫苗的进行比较。两种疫苗引起对由3T3/neu靶细胞表达的Her2/neu抗原强烈的但是相当的细胞毒性T细胞应答。因此,用仅表达与LLO融合的Her2的胞内片段的李斯特菌免疫的小鼠显示比含有更多MHC I类表位的嵌合体更低的裂解活性。在未暴露的动物或注射不相关李斯特菌疫苗的小鼠中没有检测到CTL活性(图16A)。ADXS31-164也能够刺激得自野生型FVB/N小鼠的脾细胞分泌IFN-γ(图16B)。这在与经丝裂霉素C处理的表达高水平的Her2/neu抗原的NT-2细胞共培养的这些细胞的培养物上清液中检测到(图19C)。
用ADXS31-164免疫之后,在HLA-A2小鼠中测试了人MHC I类表位的适当加工和呈递。将来自经免疫的HLA-A2转基因动物的脾细胞与对应于作图的HLA-A2限制性表位的肽一起共温育72小时,所述HLA-A2限制性表位位于Her2/neu分子的细胞外结构域(HLYQGCQVV SEQ ID NO:74或KIFGSLAFL SEQ ID NO:75)或细胞内结构域(RLLQETELV SEQ ID NO:76)(图16C)。用重组ChHer2蛋白作为阳性对照,不相关肽或没有肽作为阴性对照。来自该实验的数据表明:ADXS31-164能够引起针对位于靶抗原的不同结构域处的人表位的抗Her2/neu特异性免疫应答。
实例13:ADXS31-164在预防自发性乳腺肿瘤的发作中比LM-LLO-ChHER2更有效。
对比了ADXS31-164与Lm-LLO-ChHer2在Her2/neu转基因动物中的抗肿瘤效果,该转基因动物在20-25周龄时产生缓慢生长的、自发性乳腺肿瘤。所有经不相关李斯特菌对照疫苗免疫的动物在第21-25周内产生乳腺肿瘤,在第33周之前处死。相比而言,李斯特菌-Her2/neu重组疫苗导致乳腺肿瘤形成的明显延迟。在第45周,超过50%的ADXS31-164接种小鼠(9只中的5只)仍然无肿瘤,与之相比,25%的经Lm-LLO-ChHer2免疫的小鼠无肿瘤。在第52周,8只经ADXS31-164免疫的小鼠中的2只仍保持无肿瘤,而来自其他实验组的所有小鼠均已死于它们的疾病(图17)。这些结果指出:尽管ADXS31-164更加减毒,但是其在Her2/neu转基因动物中预防自发性乳腺肿瘤的发作方面比Lm-LLO-ChHer2更有效。
实例14:经ADXS31-164免疫后HER2/NEU基因的突变
Her2/neu的MHC I类表位中的突变已被认为是造成以小片段疫苗或曲妥珠单抗(赫赛汀,一种靶向Her2/neu的胞外结构域中的表位的单克隆抗体)免疫后的肿瘤逃逸的原因。为了对此进行评估,从转基因动物的逃逸肿瘤中提取基因组材料,并对经嵌合疫苗或对照疫苗免疫的肿瘤中neu基因的对应片段进行测序。在任何接种疫苗的肿瘤样品的Her-2/neu基因中均未观察到突变,这提示替代性逃逸机制(数据未显示)。
实例15:ADXS31-164引起肿瘤内调节性T细胞的明显下降。
为了阐明ADXS31-164对脾和肿瘤中的调节性T细胞的频率的影响,向小鼠植入NT-2肿瘤细胞。三次免疫之后,分离脾细胞和肿瘤内淋巴细胞,并且针对Treg染色,Treg被定义为CD3+/CD4+/CD25+/FoxP3+细胞,尽管当单独分析时,用FoxP3或CD25标志物获得了相当的结果。结果指出:与不相关李斯特菌疫苗或未暴露的动物相比,ADXS31-164免疫对脾中Treg的频率没有影响(参见图18)。相反地,以李斯特菌疫苗进行免疫对肿瘤中Treg的存在造成了明显的影响(图19A)。然而在未处理肿瘤中所有CD3+T细胞中平均19.0%是Treg,该频率对于不相关疫苗而言下降至4.2%,并且对于ADXS31-164而言下降至3.4%,肿瘤内Treg的频率下降了5倍(图19B)。经任一种LmddA疫苗处理的小鼠中肿瘤内Treg频率的下降不能归因于肿瘤尺寸的差异。在代表性实验中,来自经ADXS31-164免疫的小鼠的肿瘤[平均直径(mm)±SD,6.71±0.43,n=5]明显小于来自未处理小鼠(8.69±0.98,n=5,p<0.01)或用不相关疫苗处理的小鼠(8.41±1.47,n=5,p=0.04)的肿瘤,而这后两组的对比未显示出肿瘤大小的统计上显著的差异(p=0.73)。经LmddA疫苗处理的肿瘤中更低频率的Treg导致肿瘤内CD8/Treg比率的增加,这提示:在经LmddA疫苗免疫之后可获得更有利的肿瘤微环境。但是,仅有表达靶抗原HER2/neu的疫苗(ADXS31-164)能够减少肿瘤生长,表明Treg的下降仅仅在肿瘤中存在抗原特异性应答的情况下有作用。
实例16:同时递送两种异种抗原的双重质粒的构建
采用以下引物ActA-F-5’-atcccgggtgaagcttgggaagcagttggg-3’(XmaI)(SEQ ID NO:77)和ActA-R-attctagatttatcacgtacccatttccccgc(XbaI)(SEQ ID NO:78)通过聚合酶链式反应(PCR)从李斯特菌基因组DNA扩增对应于actA启动子区域和ActA的N末端1-233氨基酸的DNA。用于克隆的限制位点标有下划线。将XmaI/XbaI区段克隆进质粒pNEB193中,以产生pNEB193-ActA。使用引物Ch-Her2-F–5’-attctagaacccacctggacatgctccgccac-3’(XbaI)(SEQ ID NO:79)和Ch-Her2-R-5’-gtcgacactagtctagtggtgatggtgatgatggagctcagatctgtctaagaggcagccatagggc-3’(RE位点-SalI-SpeI-SacI-BglII)(SEQ ID NO:80)PCR扩增作为嵌合体Her2的另外抗原2。将Ch-Her2的XbaI和SalI片段克隆进质粒pNEB193-ActA中,以产生pNEB193-ActA-Ch-Her2质粒。在Ch-Her2反向引物序列中SacI和SpeI限制位点之间包含His标签DNA序列。将与来自质粒pNEB193-ActA-Ch-Her2的tActA-Ch-Her2-His对应的XmaI/SpeI片段切离,用于克隆进XmaI/SpeI限制的pAdv134中,以建立双重质粒。
然后生成基于李斯特菌的质粒,其同时递送两种重组抗原作为融合蛋白。该质粒表达的两种融合蛋白包括tLLO-抗原1和tActA-抗原2。抗原1的表达和分泌是在hly启动子和LLO信号序列的控制下,且它被表达为与李斯特菌溶血素O的非溶血性片段(截短的LLO或tLLO)的融合体。抗原2的表达和分泌是在actA启动子和ActA信号序列的控制下,且它被表达为与ActA的1-233氨基酸(截短的ActA或tActA)的融合体。之前已经描述了不含抗生素标志物的质粒pAdv134的构建,它含有用于表达tLLO-抗原1融合蛋白的基因盒。存在于pAdv134中tLLO-抗原1下游的SpeI和Xma I限制位点被用于克隆actA启动子-tActA-抗原2DNA区段,见图20。将限制位点XbaI、SacI和BglII加入表达盒中,以方便抗原2插入片段在XbaI/SacI或XbaI/BglII处的克隆。将编码His标签的DNA序列添加在SacI位点之后,以有利于tActA-抗原2-his融合蛋白的检测。该双重质粒能够同时表达和分泌两种不同抗原作为融合蛋白。
材料与方法(实例17-21)
MDSC和Treg功能
取决于肿瘤模型,将肿瘤植入小鼠的胁腹或生理学部位。7天后,向小鼠接种疫苗,接种首日取决于使用的肿瘤模型。然后在给予疫苗后一周,用强化疫苗对小鼠给药。
然后在强化后1周处死小鼠,并收获肿瘤和脾,或者在侵袭性肿瘤型的情况下,在强化后3-4天收获肿瘤和脾。在收获肿瘤之前五天,给非荷瘤小鼠接种疫苗,以用于应答T细胞。采用标准方法学制备脾细胞。
简而言之,制备肿瘤和脾的单细胞悬液。手工地破碎脾,并裂解红血细胞。切碎肿瘤,并与胶原酶/DNA酶一起温育。备选地,将GENTLEMACSTM解离器与肿瘤解离试剂盒一起使用。
采用Miltenyi试剂盒和柱或autoMACs分离器,从肿瘤和脾纯化MDSC或Treg。然后对细胞计数。
制备单细胞悬液,并裂解红血细胞。然后以CFSE标记应答T细胞。
以1×105个T细胞/孔的密度,将应答T细胞(来自所有分裂周期阶段)与MDSC或Treg以2:1比率一起铺板在96孔板中。然后用适当的肽(PSA或CA9)刺激应答T细胞,或者用PMA/离子霉素非特异性地刺激。将细胞在5%CO2下在37℃在暗处温育2天。两天后,将细胞染色进行FACS,并在FACS机器上分析。
T细胞应答分析
对于通过ELISA进行的细胞因子分析,收获脾细胞,并在有培养基、SEA或conA(作为阳性对照)存在下,以150万个细胞/孔铺板在48孔平板中。温育72小时后,收获上清液,通过ELISA(BD)分析细胞因子水平。对于抗原特异性IFN-γELISpot,收获脾细胞,并在有培养基、特定CTL肽、不相关肽、特定辅助肽或conA(作为阳性对照)存在下,以30万和15万个细胞/孔铺板在IFN-γELISpot平板中。温育20小时后,进行ELISpots(BD),并通过Immunospot分析仪(C.T.L.)对斑点计数。将每一百万个脾细胞的斑点数绘图。
使用Coulter Counter,Z1对脾细胞计数。使用标准的基于IFN-γ的ELISPOT测定,确定在以gag-CTL、gag-辅助、培养基、不相关抗原和con A(阳性对照)再刺激以后,产生IFN-γ的CD8+T细胞的频率。
简而言之,以5mg/ml使用mAb R46-A2和以最佳稀释度使用多克隆兔抗IFN-γ(由University of Pennsylvania,Philadelphia,PA的Phillip Scott博士友情提供)检测IFN-γ。通过与使用鼠rI++FN-γ(Life Technologies,Gaithersburg,MD)绘制的标准曲线进行对比,计算IFN-γ的水平。使用过氧化物酶偶联的山羊抗兔IgG Ab(IFN-γ)让平板显影。然后在405nm处读取平板。测定法的检测下限为30pg/ml。
结果
实例17:在李斯特菌疫苗处理以后的抑制性细胞功能
在第0天,将肿瘤植入小鼠中。在第7天,给小鼠接种Lmdda-E7或LmddA-PSA。在第14天,收获肿瘤,并测量接种组和未暴露的组的浸润性MDSC和Treg的数目和百分比。据发现,在李斯特菌处理过的小鼠的肿瘤中MDSC和Treg的百分比以及MDSC的绝对数目的减小,而在脾或引流淋巴结(TLDN)中没有观察到该效应(图21)。
将从上述实验中的荷瘤小鼠提取的分离的脾细胞和肿瘤浸润淋巴细胞(TIL)合并,并针对CD3和CD8染色,以阐明Lm-LLO-E7、Lm-LLO-PSA和Lm-LLO-CA9、Lm-LLO-Her2免疫对MDSC和Treg(脾和肿瘤的MDSC和Treg)在肿瘤中的存在的影响(图22-34)。每个列代表处于特定细胞分裂阶段的T细胞群体的百分比,并且在特定处理组(未暴露的、肽CA9或PSA处理、无MDSC/Treg和无MDSC+PMA/离子霉素)下分成小组(参见图22-34)。
针对存在的Treg和MDSC的百分比,分析来自荷瘤小鼠的血液。在Lm接种后,在小鼠血液中存在MDSC和Treg二者的下降。
实例18:在李斯特菌接种后,来自TPSA23肿瘤而不是脾的MDSC具有更低的抑制性。
用非特异性地活化的未暴露的鼠细胞和特异性地活化的细胞(PSA、CA9、PMA/离子霉素),使用分离自TPSA23肿瘤的单核细胞和粒细胞MDSC,进行抑制测定。结果证实:与来自未暴露的小鼠的肿瘤的MDSC相比,分离自Lm接种组的肿瘤的MDSC具有减小的抑制活化的T细胞分裂的能力。(参见图22和24中的Lm-LLO-PSA和Lm-LLO处理组,在图中右侧的插图代表来自左侧插图的合并的细胞分裂数据)。另外,来自未处理小鼠(其中不存在MDSC,并且其中细胞未被刺激/活化)的T应答细胞保持处于它们的亲代(休眠)状态(图22和24),而观察到用PMA或离子霉素刺激的T细胞发生复制(图22和24)。另外,据观察,Gr+Ly6G+和GrdimLy6G-MDSC在李斯特菌疫苗处理后抑制性较低。这适用于它们降低的抑制活化的PSA特异性T细胞和非特异性(PMA/离子霉素刺激的)T细胞分裂的能力。
此外,采用分离自TPSA23肿瘤的MDSC以及非特异性地活化的未暴露的鼠细胞进行的抑制测定证实:与来自未暴露的小鼠的肿瘤的MDSC相比,分离自Lm接种组的肿瘤的MDSC具有减小的抑制活化的T细胞分裂的能力(参见图22和24)。
另外,当使用脾MDSC时,没有观察到上文刚刚关于图22和24讨论的观察结果。在后者中,来自未暴露的组、李斯特菌处理组(PSA、CA9)和PMA/离子霉素刺激组(阳性对照)的脾细胞/T细胞都表现出相同的复制水平(图23和25)。因此,这些结果表明:李斯特菌介导的对肿瘤中抑制性细胞的抑制以抗原特异性和非特异性方式起作用,而李斯特菌对脾粒细胞MDSC没有影响,因为它们仅以抗原特异性方式产生抑制。
实例19:肿瘤调节性T细胞的减少的抑制
使用在李斯特菌处理后分离自TPSA23肿瘤的Treg,进行抑制测定。据观察,在李斯特菌处理后,来自肿瘤的Treg的抑制能力下降(图26),但是,发现脾Treg仍然是抑制性的(图27)。
作为对照,使用常规CD4+T细胞替代MDSC或Treg,并发现对细胞分裂没有影响(图28)。
实例20:在李斯特菌接种后,来自4T1肿瘤而不是脾的MDSC和TREG具有较低的抑制性。
如上所述的,采用4T1肿瘤进行了相同实验,并且得到了相同的观察结果,即,在李斯特菌接种后MDSC具有较低的抑制性(图29和31),李斯特菌对脾单核细胞MDSC没有特异性作用(图30和32),在李斯特菌接种后来自4T1肿瘤的Treg的抑制能力下降(图33),以及李斯特菌对脾Treg的抑制能力没有影响(图34)。
最后,据观察,李斯特菌对脾Treg的抑制能力没有影响。
实例21:粒细胞和单核细胞MDSC的抑制能力的变化是由于tLLO的过表达。
LLO质粒显示出与含有TAA或不相关抗原的李斯特菌疫苗类似的结果(图35)。这意味着,粒细胞MDSC的抑制能力的变化是由于tLLO的过表达,且独立于伴侣融合抗原。单独的空质粒构建体也会导致MDSC的抑制能力的变化,尽管不是与在质粒上含有截短LLO的任意疫苗刚好相同的水平。3个独立实验的平均值表明:空质粒和其他含有tLLO(含有和不含肿瘤抗原)的质粒之间的抑制差异是显著的。不论是使用抗原特异性还是非特异性刺激的应答T细胞,MDSC抑制能力的下降是相同的。
类似于粒细胞MDSC,3个独立实验的平均值表明:当与其他疫苗构建体相比,在用Lm空质粒接种以后,从肿瘤纯化的单核细胞MDSC观察到的抑制能力的差异是显著的(图36)。
类似于以上观察结果,在Lm接种以后,从脾纯化的粒细胞MDSC保留了它们抑制抗原特异性应答T细胞分裂的能力(图37)。但是,在非特异性刺激以后,(以PMA/离子霉素)活化的T细胞仍然能够分裂。这些结果都没有随单独LLO或空质粒疫苗的使用而改变,表明基于Lm的疫苗不会影响脾粒细胞MDSC(图37)。
类似地,在Lm接种以后,从脾纯化的单核细胞MDSC保留它们的抑制抗原特异性应答T细胞分裂的能力。但是,在非特异性活化(以PMA/离子霉素刺激)以后,T细胞仍然能够分裂。这些结果都没有随单独LLO或空质粒疫苗的使用而改变,表明Lm疫苗不会影响脾单核细胞MDSC(图38)。
不论应答细胞是抗原特异性还是非特异性活化的,从任意Lm处理组的肿瘤纯化的Treg都具有略微降低的抑制应答T细胞分裂的能力。特别是对于非特异性活化的应答T细胞而言,看起来好像含有空质粒的疫苗显示出与在质粒上含有LLO的所有疫苗相同的结果。该实验与其他实验的平均值表明,该差异不是显著的(图39)。
从脾纯化的Treg仍然能够抑制抗原特异性和非特异性活化的应答T细胞的分裂。Lm处理对脾Treg的抑制能力没有影响(图40)。
不论应答细胞是抗原特异性还是非特异性活化的,Tcon细胞都不能抑制T细胞的分裂,这与这些细胞不具有抑制性的事实一致。Lm对这些细胞没有影响,如果从小鼠的肿瘤或脾纯化这些细胞,则不存在差异(图41-42)。
材料与方法(实例22-28)
小鼠
将来自Charles River Laboratories的Balb/c雌性小鼠(6-8周龄)用于所有涉及4T1肿瘤系的实验。将来自Jackson Laboratories的FVB/NJ雌性小鼠(6-8周龄)用于所有涉及NT2肿瘤系的实验。将FVB/NJ背景中的大鼠Her2/neu转基因小鼠品系用于涉及自发肿瘤形成的研究以及用于原部位乳腺肿瘤形成的预防研究,安置和饲养在University of Pennsylvania的动物中心机构。所有的小鼠实验根据宾夕法尼亚大学实验动物管理与使用委员会(Institutional Animal Care and Use Committee of the University of Pennsylvania)的规章进行。
李斯特菌菌株
为了构建基于减毒李斯特菌的ISG15疫苗,首先采用以下引物:Lm-LLO-ISG15.FOR 5’-TAAT-CTCGAG-ATGGCCTGGGACCTAAAG-3’(SEQ ID NO:83)和Lm-LLO-ISG15.REV 5’-ATTA-ACTAGT-TTAGGCACACTGGTCCCC-3’(SEQ ID NO:84)从包含来自Balb/c小鼠的鼠ISG15cDNA的构建体扩增编码鼠ISG15的基因。将正向引物中标下划线的XhoI序列和反向引物中标下划线的SpeI序列用于连接。将每个片段扩增子都用限制酶消化并连接进李斯特菌表达质粒pGG34。每一序列都在基因上融合至编码截短的李斯特菌溶血素O(tLLO)的序列的下游,置于hly启动子控制下。随后,让pGG34-LLO-ISG15电穿孔进减毒单核细胞增多性李斯特菌(Lm)菌株XFL7,并在BHI氯霉素平板上进行含质粒集落的抗性筛选。为了验证Lm-LLO-ISG15的正确构建,让基于减毒李斯特菌的疫苗在BHI-氯霉素选择培养基中生长,所分泌的蛋白以三氯乙酸沉淀。在SDS样品缓冲液中煮沸之后,对分泌的蛋白进行SDS-PAGE分析并转移至PVDF膜。用抗小鼠ISG15抗体(Santa Cruz Biotech,Santa Cruz,CA)在膜上进行Western分析以验证tLLO-ISG15融合蛋白的分泌,对抗鸡卵清蛋白使用3A11.2单克隆抗体,以及对野生型LLO使用B3-19单克隆抗体。类似地构建由tLLO在基因上融合至鸡卵清蛋白而构成的对照疫苗Lm-LLO-OVA。所有的基于李斯特菌的疫苗都以200μl PBS中2×108或5×108个CFU腹膜内(i.p.)施用。类似地构建对照疫苗Lm-LLO-OVA和Lm-LLO-NYESO-1。
细胞系
将转移性乳腺癌肿瘤细胞系4T1用在Balb/c小鼠中的肿瘤植入研究中。将过表达大鼠Her2/neu的NT2乳腺癌细胞系用于FVB小鼠中的肿瘤植入研究中。将4T1-Luc维持在补加了10%胎牛血清、2mML-谷氨酰胺、1mM丙酮酸钠、50U/mL青霉素以及50μg/mL链霉素的DMEM中。将NT2细胞维持在补加了10%胎牛血清、20μg/mL胰岛素、2mML-谷氨酰胺、1mM丙酮酸钠、50U/mL青霉素以及50μg/mL链霉素的RPMI 1640培养基中。未转化的NIH-3T3成纤维细胞系从ATCC获得。将NIH-3T3细胞维持在补加了10%胎牛血清、2mML-谷氨酰胺、1mM丙酮酸钠、50U/mL青霉素以及50μg/mL链霉素的DMEM中。
ISG15在正常和肿瘤鼠组织中的表达
采用来自Qiagen的RNeasy RNA提取试剂盒从组织或细胞提取RNA,并将其转变为cDNA。然后以特异于ISG15的引物qISG15.FOR5’-ATGGCCTGGGACCTAAAG-3’(SEQ ID NO:85)和qISG15.REV5’-TTAGGCACACTGGTCCCC-3’(SEQ ID NO:86),特异于18S rRNA的引物18SRNA.FOR 5’-CGGCTACCACATCCAAGGAA-3’(SEQ ID NO:87)和18SRNA.REV 5’-GCTGGAATTACCGCGGCT-3’(SEQ ID NO:88),以及特异于β-肌动蛋白的引物ACTIN.FOR 5’-GTGGGCCGCTCTAGGCACCAA-3’(SEQ ID NO:89)和ACTIN.REV 5’-CTCTTTGATGTCACGCACGATTTC-3’(SEQ ID NO:90)使cDNA接受PCR分析。将ISG15表达归一化至18S rRNA(图43C和D)或β-肌动蛋白(图43A)。
乳腺组织裂解物的Western印迹分析
将来自FVB/N小鼠(n=4)的正常乳腺组织和来自FVB/N背景(n=9)中的HER2/neu转基因小鼠的原部位乳腺肿瘤组织切下并加工成裂解物。简而言之,将组织样品快速冷冻在液体N2中,粉碎并溶解在补加了蛋白酶抑制剂混合物的裂解缓冲液(PBS,含2%Triton X-100和0.02%皂苷)中。将裂解物与4X LDS上样缓冲液混合,进行SDS-PAGE。在将分离的蛋白转移至PVDF膜后,以抗小鼠ISG15抗体进行Western印迹分析。将相同的裂解物单独进行SDA-PAGE,并以考马斯蓝对凝胶染色,以便观察总蛋白,作为对蛋白上样的度量。
用ISG15肽的免疫治疗。
将4T1-Luc肿瘤细胞(105个)植入Balb/c小鼠的乳腺组织,并随后在第5、12和19天以100μl的PBS或在100μL的PBS中与对照、HIV-gag H-2Kd CTL表位肽(AMQMLKETI)(SEQ ID NO:91)或ISG15特异性肽(100μg)、pISG15d1(RGHSNIYEV)(SEQ ID NO:92)和pISG15d2(LGPSSTVML)(SEQ ID NO:93)混合的50μg CpG寡脱氧核苷酸(ODN)在颈部淋巴结的近侧对小鼠皮下接种。在整个实验过程中通过垂直卡尺测量来监测肿瘤体积。肿瘤体积计算为(肿瘤直径)3/2。
ISG15肽肿瘤负荷研究
将4T1-Luc肿瘤细胞(105个)植入乳腺组织,随后在第4、11和18天或者以100μl的PBS中单独的50μg CpG或者以100μl的PBS中连同对照或ISG15特异性肽(100μg)的CpG(50μg)在颈部淋巴结的近侧对小鼠皮下接种。在第32天实验结束时,测量每个接种组的肿瘤质量,按“ELISpot分析”中所描述的那样对肿瘤分析ISG15特异性IFN-γ应答,并按“转移性肿瘤研究”中所描述的那样测量肺转移。
转移性肿瘤研究
将4T1-Luc肿瘤细胞(105个)植入乳腺组织,随后在第4、11和18天以肽或基于李斯特菌的疫苗对小鼠接种。然后在第32天处死小鼠,分离肺并以PBS灌注。接着以附连到Fostec 8375发光器和Ringlight的Nikon SMZ1B Zoom立体显微镜对每个肺的肺表面转移性结节进行计数。
ELISpot分析
以100μl的PBS中15μg/ml大鼠抗小鼠IFN-γ抗体包被96孔过滤板(Millipore,Bedford,MA)。在4℃温育过夜后,洗涤孔,并以补加了10%胎牛血清的DMEM封闭。对于图2C,将来自每个实验组的脾细胞连同HIV-gag H-2Kd CTL表位肽(AMQMLKETI)(SEQ ID NO:91)或预测的ISG15特异性H-2Kd CTL表位肽ISG15-d1(RGHSNIYEV)(SEQ ID NO:92)和ISG15-d2(LGPSSTVML)(SEQ ID NO:93)(5μg/ml)+IL-2(5U/ml)添加至孔中。ISG15特异性H-2Kd CTL表位是采用http://bio.dfci.harvard.edu/Tools/rankpep.html上的RANKPEP预测软件从Balb/c小鼠中的ISG15蛋白序列预测的。对于图47B,将来自每个实验组的脾细胞连同HIV-gag H-2Kd CTL表位肽(AMQMLKETI)(SEQ ID NO:91)或Her2/neu-特异性H-2Kd表位肽Her2-EC1(PYNYLSTEV)(SEQ ID NO:94)、Her2-EC2(LFRNPHQALL)(SEQ ID NO:95)和Her2-IC1(PYVSRLLGI)(SEQ ID NO:96)加到孔中。让细胞在37℃温育24h。洗涤平板,然后与100μl PBS中的1μg/ml生物素化IFN-γ抗体(克隆R4-6A2,MABTECH,Mariemont,OH)在4℃温育过夜。洗涤后,添加100μl PBS中的1:100链霉亲和素-辣根过氧化物酶,并在室温温育1h。洗涤后通过添加100μl底物来显示斑点,并在室温温育15min。通过在dH2O中充分洗涤终止显色,以ELISpot读取仪对斑点形成细胞(SFC)计数。
耗尽实验
在肿瘤植入后的第6、7、8、10、12和14天通过向小鼠注射0.5mgα-CD8抗体(单克隆抗体克隆2.43)让4T1-Luc荷瘤小鼠中的CD8+细胞耗尽。对照组小鼠也在相同条件下处理,只是采用同种型匹配的、特异于β-半乳糖苷酶的对照抗体。同时进行的肿瘤负荷研究遵循本文方法章节中“用Lm-LLO-ISG15的免疫治疗”中的描述。
用于效应细胞的体内测定的Winn测定法。
按以前的描述但有一些改动进行Winn测定法。简而言之,让4T1-Luc肿瘤细胞(2×105个)与来自两次对照Lm接种的或两次Lm-LLO-ISG15接种的Balb/c小鼠的CD4耗尽的脾细胞(以CD4+Dynabeads耗尽并通过FACS分析确认)(2×107个)以1个肿瘤细胞对100个CD4耗尽的脾细胞的比率混合,并植入乳腺组织。然后按本文方法章节中“用Lm-LLO-ISG15的免疫治疗”中的描述测量肿瘤生长。
HER2/neu特异性肿瘤浸润淋巴细胞(TIL)的检测
将4T1-Luc肿瘤细胞植入Balb/c小鼠,以对照Lm或Lm-LLO-ISG15腹膜内免疫,并在7天后强化。在强化后9天收获肿瘤,手工解离为单细胞悬液。然后将肿瘤细胞悬液Ficoll纯化以通过在离心后排除低密度组分来除去死亡细胞和细胞碎片。然后对剩下的肿瘤细胞针对CD8(53–6.7,FITC偶联的)、CD62配体(CD62L;MEL-14,APC偶联的)和HER2/neu-EC2H-2Dq四聚体-PE偶联的(特异于PDSLRDLSVF,SEQ ID NO:97)采用带有Cell Quest软件的FACSCalibur流式细胞仪进行三色流式细胞术。四聚体由National Institute of Allergy and Infectious Diseases Tetramer Core Facility提供,并以1/200的稀释度使用。按上文描述的对结果进行分析,以比较Lm-LLO-ISG15和对照Lm接种诱导四聚体+、CD8+、CD62L-、Her2/neu特异性TIL的能力。
统计分析
对所有的最终肿瘤体积进行单尾学生t检验,带有Welch’s校正的转移性负荷和免疫应答研究应适用于以原部位HER2/neu乳腺肿瘤进行的基因表达研究。对原部位HER2/neu乳腺肿瘤发生率研究进行对数秩检验。采用GraphPad Prism version 4.0a for Macintosh(www.graphpad.com)进行统计学分析。对于所有的比较,显著性p值如下描绘在图中:*=p值<0.05,**=p值<0.01。以及***=p值<0.001。
结果
实例22:鼠乳腺肿瘤中ISG15的升高的表达
ISG15在人恶性肿瘤中升高的表达在许多肿瘤模型中有充分的表征。但是,在鼠肿瘤模型中缺乏ISG15水平类似升高的证据。为了确定ISG15表达是否在乳腺癌的小鼠模型中升高,在来自HER2/neu转基因小鼠的原部位小鼠乳腺肿瘤、小鼠乳腺肿瘤细胞系和一组正常和未转化的乳腺组织和细胞系中测定了ISG15的表达。如同在人乳腺癌中观察到的,与正常小鼠乳腺组织相比,ISG15mRNA的表达在原部位小鼠乳腺肿瘤中显著升高(图43A)。为了证实升高的ISG15mRNA表达导致升高的蛋白生成,对正常和HER2/neu肿瘤小鼠乳腺组织的裂解物以抗ISG15抗体进行了Western印迹分析。与正常小鼠乳腺组织(图43B,上部插图,泳道1)相比,偶联形式的ISG15蛋白(高于20KD标记物的带)在HER2/neu乳腺肿瘤组织中升高(图43B,上部插图,泳道2-5)。与正常乳腺组织(图43B,上部插图,泳道1)相比,非偶联形式的ISG15蛋白的升高的表达在小鼠乳腺肿瘤组织(图43B,上部插图,泳道2和4)中也是明显的。通过以相同裂解物探测管家蛋白GAPDH的表达,表明蛋白上样量是等量的(图43B,下部插图,泳道1-5)。与正常小鼠乳腺组织和未转化的小鼠细胞系NIH-3T3相比,ISG15mRNA表达在小鼠乳腺肿瘤细胞系4T1-Luc和NT2中类似地升高(图43C)。为了缓解对非恶性肿瘤组织中升高的ISG15表达的担忧,与HER2/neu乳腺小鼠肿瘤组织相比较,在一组正常小鼠组织中分析了ISG15mRNA表达。与每个正常组织类型相比,类似地观察到乳腺肿瘤组织中ISG15mRNA显著升高的表达(图43D)。这种表达分析证实ISG15表达在乳腺癌小鼠模型中显著升高。结合ISG15mRNA在一组正常组织中名义上表达的发现,这提示ISG15可能是有希望的新肿瘤相关抗原(TAA)。
实例23:构建ISG15特异性CTL疫苗
为了评估ISG15作为新TAA的可能性,开发了基于李斯特菌的CTL疫苗以靶向具有升高的ISG15表达的肿瘤。通过将来自Balb/c小鼠的小鼠ISG15基因遗传上融合至已存在于单核细胞增多性李斯特菌(Lm)表达载体pGG34中的编码截短形式的李斯特菌溶血素O(LLO)的基因下游来完成疫苗Lm-LLO-ISG15的构建,所述表达载体含有信号序列以使得能正确分泌融合蛋白。随后将pGG34-LLO-ISG15构建体电穿孔进减毒的感受态Lm菌株XFL7(图44A)。通过以抗小鼠ISG15抗体对来自Lm-LLO-ISG15生长培养物的培养基的TCA沉淀蛋白进行Western印迹分析,证实tLLO-ISG15融合蛋白的正确分泌(图44B,下部插图)。当以抗卵清蛋白抗体探测时,从我们的对照Lm观察到tLLO融合至鸡卵清蛋白的融合蛋白的类似产生和分泌(图44B,中部插图)。来自Lm-LLO-ISG15和对照Lm的分泌的蛋白也用野生型LLO抗体探测,以证实相同量的分泌蛋白上样量(图44B,下部插图)。从第6周开始,每周一次,通过以Lm-LLO-ISG15和对照Lm疫苗向雌性Balb/c小鼠给药,来测定ISG15特异性CTL应答的产生。在第三次接种后一周,对来自每个接种组的脾细胞进行ELISpot分析,以考察针对对照表位和两个通过RANKPEP预测的ISG15特异性H2-Kd限制性CD8+T细胞表位的IFN-γ应答。与对照肽刺激相比较,只在以每种预测的ISG15特异性CTL表位刺激后从来自经Lm-LLO-ISG15接种的小鼠的脾细胞中观察到IFN-γ分泌性SFC的显著增加(图44C)。这些结果提示:ISG15特异性的适应性应答可通过针对ISG15的基于减毒Lm的CTL疫苗产生。
尽管在正常条件下,SG15表达在正常组织中处于低的或者检测不到的水平,但是有证据表明妊娠期间在胎盘植入位点有升高的ISG15表达。为了确定ISG15特异性免疫应答是否可能严重影响Lm-LLO-ISG15接种雌性小鼠中的繁殖力,进行了妊娠研究。与对照Lm接种的雌性小鼠相比,通过窝产崽数和幼崽体重度量(分别为图44D和E),Lm-LLO-ISG15接种的雌性小鼠的繁殖力没有受到明显损害。产生ISG15特异性的适应性免疫应答而没有明显的不良反应,鼓舞了研究其在乳腺癌小鼠模型中的有效性。
实例24:LM-LLO-ISG15接种后对鼠乳腺肿瘤的治疗性影响
最初在乳腺癌的植入原发和转移小鼠模型上考察了通过Lm-LLO-ISG15产生的针对乳腺癌的ISG15特异的适应性免疫应答的治疗潜力。将NT2肿瘤细胞皮下植入FVB/N小鼠的后胁腹并随后以Lm-LLO-ISG15接种,导致与对照接种相比显著减小的肿瘤体积(图45A)。类似地,Lm-LLO-ISG15治疗性接种显著地抑制了乳腺组织植入的4T1-Luc原发肿瘤的生长(图45B)。在植入乳腺中之后4T1-Luc肿瘤自然转移的能力使得能进一步考察ISG15特异性CTL应答对乳腺癌的更强侵袭性模型的有效性。与对照Lm相比,在Lm-LLO-ISG15给药后观察到显著减少的4T1-Luc转移性肺病变的出现(图45C)。
实例25:通过Lm-LLO-ISG15产生的HER2/NEU+原部位乳腺肿瘤延迟的进展和表位扩展
为了确定Lm-LLO-ISG15是否也能在人乳腺肿瘤发展的更加临床相关的模型中提供治疗效果,我们利用了FVB/N HER2/neu转基因小鼠模型,该小鼠模型在缺乏治疗干预的情况下超过4月龄就生长原部位乳腺肿瘤。从出生后第6至第21周,每三周以Lm-LLO-ISG15或对照Lm接种转基因雌性小鼠,随后监测乳腺肿瘤发生率。与对照Lm接种组相比,Lm-LLO-ISG15给药的小鼠展示了肿瘤进展的显著延迟(p<0.0001)(图46A)。实际上,大于80%的Lm-LLO-ISG15接种小鼠在出生后49周仍然是无肿瘤的,而所有的对照Lm接种小鼠都生长出乳腺肿瘤,进展中位时间为31周。为了确定Lm-LLO-ISG15接种后ISG15特异性CTL浸润到原部位肿瘤中是否为这种延迟的进展的可能机制,在Lm-LLO-ISG15接种后对这些肿瘤的TIL进行了IFNγELISpot分析。在允许原部位肿瘤形成后,在第0和第7天以对照Lm疫苗或Lm-LLO-ISG15接种荷瘤小鼠两次。在最后一次接种后一周,切除肿瘤,纯化TIL,并进行处理用于ELISpot分析。如所预料的,Lm-LLO-ISG15接种的肿瘤,与对照Lm接种小鼠的肿瘤相比,含有显著较多数量的特异于ISG15的TIL,如ISG15表位肽刺激后测量它们分泌IFNγ的能力所显示的(图46B)。这些结果提示:Lm-LLO-ISG15引起的原部位乳腺肿瘤的延迟的进展部分地是由ISG15特异性CTL的浸润所介导的。
最近的研究表明,癌症疫苗的临床效果与它们刺激交叉致敏和表位扩展至另外TAA的能力显著相关。之前采用基于Lm的癌症疫苗观察到类似的结果,其中表位扩展至另外TAA的发生与疫苗效果相关。为了评估表位扩展是否是在Lm-LLO-ISG15接种后发生,对来自对照Lm或Lm-LLO-ISG15给药后的NT2荷瘤小鼠的脾细胞进行了检测HER2/neu特异性应答的ELISpot。与对照Lm接种小鼠相比,Lm-LLO-ISG15接种小鼠的脾细胞含有显著较多数量的特异于HER2/neu内已知CTL表位的SFC(图46C)。该结果提示:Lm-LLO-ISG15接种导致表位扩展至另外的TAA。实际上,表位扩展的证据还在针对4T1-Luc肿瘤的Lm-LLO-ISG15接种后观察到,该4T1-Luc肿瘤细胞系表达Her2/neu非常弱。来自Lm-LLO-ISG15接种小鼠的4T1-Luc肿瘤含有比来自对照Lm接种小鼠的4T1-Luc显著较高百分比的Her2/neu特异性CD8+62L-TIL(图46D)。尽管表位扩展至HER2/neu可提供某些治疗效果,但不清楚这种二次应答是否足够强烈,以致有必要担忧心脏毒性安全。总体而言,这些肿瘤负荷研究证实:针对ISG15的接种可抑制原发植入小鼠乳腺肿瘤的生长,抑制转移性扩散,延迟原部位乳腺肿瘤的进展和产生表位扩展至另外TAA。
实例26:ISG15接种的治疗性影响是CD-8依赖性的
虽然产生强烈的IFN-γ应答和显著的治疗性肿瘤影响提示强的CTL应答,但是研究了Lm-LLO-ISG15效果中对ISG15特异性CD8+T细胞功能的依赖性。与对照抗体的模拟耗尽相比,在4T1-Luc荷瘤小鼠中耗尽CD8+细胞完全消除了Lm-LLO-ISG15的抗肿瘤效果(图47A)。作为ISG15特异性CTL肿瘤细胞裂解的体内测量,我们进行了Winn测定来评估富集了CD8+T细胞的来自Lm-LLO-ISG15接种小鼠的脾细胞是否可直接抑制4T1-Luc肿瘤形成。将来自以Lm-LLO-ISG15或对照Lm两次接种的小鼠的脾细胞耗尽CD4+细胞并与4T1-Luc肿瘤细胞短暂温育。然后将肿瘤细胞和脾细胞混合物植入Balb/c小鼠的乳腺组织并监测肿瘤进展。与来自对照Lm接种小鼠的脾细胞相比,来自Lm-LLO-ISG15接种小鼠的富集了CD8+T细胞的脾细胞显著抑制了肿瘤生长(图47B)。另外,所有的对照Lm脾细胞接收小鼠在植入21天后出现肿瘤,而接收ISG15特异性脾细胞的小鼠中的40%在第43天时仍然无肿瘤(图47C)。该结果提示:Lm-LLO-ISG15诱导CD8依赖性的适应性免疫应答,这导致肿瘤细胞的直接裂解,并很可能由CD8+T细胞介导。
实例27:ISG15特异性CTL克隆的体内扩增引起抗肿瘤应答
为了评估单一ISG15特异性CD8+T细胞克隆的扩增是否可引起抗肿瘤效果,将4T1-Luc肿瘤细胞植入小鼠并以单独的PBS或与每种ISG15H2Kd表位肽或对照肽混合的佐剂CpG ODN接种。在以CpG ODN和ISG15H2Kd肽接种的小鼠中,4T1-Luc肿瘤体积和重量质量与单独的PBS和对照肽接种相比都显著减小(分别为图48A和B)。与单独的PBS或对照肽接种相比,以每种ISG15肽接种后4T1-Luc肿瘤肺转移也显著减少(图48C)。另外,响应于每种ISG15H2Kd表位肽刺激的IFNγ分泌仅在来自以它们各自的ISG15H2Kd表位肽接种的小鼠的TIL中观察到,提示存在成功的到达靶肿瘤的ISG15特异性CTL扩增(图48D和E)。这些数据强烈提示:ISG15特异性CD8+T细胞的扩增可直接抑制具有升高的ISG15表达的肿瘤的生长。
材料与方法(实例28-37)
小鼠。
雌性FVB/N小鼠购自Charles River Laboratories。在University of Pennsylvania的动物中心机构安置和饲养FVB/N Her-2/neu转基因小鼠。在实验开始时小鼠为六至八周龄,这些实验根据宾夕法尼亚大学的实验动物管理与使用委员会的规章进行。
肽和抗体。
抗小鼠CD31、抗小鼠CD8α-PE、大鼠IgG2a-PE同种型对照购自BD Biosciences(San Jose,CA)。兔抗李斯特菌抗血清多克隆抗体,血清型1,4购自Difco BD Biosciences。兔抗HIF-1α购自Novus Biologicals(Littleton,CO)。山羊抗兔-Alexa-488二抗购自Invitrogen。DAPI购自Sigma(St.Louis,MO)。大鼠抗小鼠IFN-g(克隆AN18)购自MABTECH(Mariemont,OH)。大鼠抗小鼠IFN-g(克隆XMG1.2)购自eBioscience(San Deigo,CA)。用在用于融合蛋白表达的Western印迹中的抗体是针对LLO蛋白的前三十个残基(PEST)产生的多克隆兔血清(Sewell等人,2004,Cancer research.64:8821-8825),或者是产生自杂交瘤上清液的特异于全长LLO的抗LLO小鼠抗体,克隆#B5-19(Edelson等人,2001,Immunity.14:503-512)。所有的肽都购自EZBiolabs(Westfield,IN)。四聚体由National Institutes of Health AIDS Research and Reference Reagent Program的Amy Stout博士提供。所用的四聚体都是PE偶联的H-2Dq,并含有用于Her-2/neu区域EC1(ASPETHLDML;SEQ ID NO:98)或EC2(PDSLRDLSVF;SEQ ID NO:97)或IC1(GSGAFGTVYK;SEQ ID NO:99)的表位。用在这些研究中的肽如下;Flk-E1210-219(TYQSIMYIV;SEQ ID NO:100)、Flk-E2613-622(MFSNSTNDI;SEQ ID NO:101)、Flk-I1906-915(PGGPLMVIV;SEQ ID NO:102)、Flk-I1839-848(GRGAFGQVI;SEQ ID NO:103);(Her2-pEC1302-310(PYNYLSTEV;SEQ ID NO:94)、Her2-pEC2420-429(PDSLRDLSVF;SEQ ID NO:97)、Her2-pIC1732-741(GSGAFGTVYK;SEQ ID NO:99);HIV-pGag(AMQMLKETI;SEQ ID NO:91)。
ELISpots
小鼠脾细胞响应于肽刺激而分泌IFN-g通过酶联免疫斑点(ELISpot)测定法来测试。我们优选使用ELISpots而不是其他测定法,是因为对低频率、抗原特异性细胞可能获得的灵敏度水平,还因为我们可直接离体测试抗Her-2/neu和抗Flk-1特异性T细胞,而无需体外操作。简而言之,在10μg/ml肽和5U/ml的IL-2存在下,将分离的脾细胞以1×106个细胞/孔铺板或者滴定在包被了7μg/ml大鼠抗小鼠IFN-g抗体(克隆AN18,MABTECH,Mariemont,OH)的96孔板上。将生物素化的抗IFN-g二抗(克隆XMG1.2,eBioscience)以终浓度2μg/ml添加至每孔。37℃温育过夜后,通过链霉亲和素-辣根过氧化物酶(1:1000稀释度)以及随后的底物TMB(Vector laboratories,ABC试剂盒)将平板在室温下显色1小时。采用Immunospot C.T.L.扫描仪和计数软件(CTL,Cleveland,OH)对斑点计数。
细胞系。
细胞培养基和补充物购自Gibco(Invitrogen)。NT-2和J774A.1细胞按之前的描述维持。所有的细胞培养物都保持在37℃和5%CO2下。4T1和稳定表达萤火虫荧光素酶基因的4T1细胞(4T1-Luc)为Ellen Pure博士(Wistar Institute)的友好礼物,并维持在细胞培养基中。
Lm-LLO-Flk-1疫苗的构建。
Flk-1基因的来源为Ralph Reisfeld博士(The Scripps Research Institute,La Jolla,CA)慷慨提供的DNA疫苗质粒。对应于残基68至1081的片段采用以下引物通过PCR扩增:Flk-E1(F):5’-GGGCTCGAGCGTGATTCTGAGGAAAGGGTATT-3’(SEQ ID NO:104),Flk-E1(R):5’GGGACTAGTTTACCCGGTTTACAATCTTCTTAT-3’(SEQ ID NO:105),(氨基酸68-277);Flk-E2(F):5’-GGGCTCGAGGTGATCAGGGGTCCTGAAATTA-3’(SEQ ID NO:106),Flk-E2(R):5’-GGGACTAGTTTAGCCTCCATCCTCCTTCCT-3’(SEQ ID NO:107),(氨基酸545-730);Flk-I1(F):5’-GGGCTCGAGGAAGGGGAACTGAAGACAGCC-3’(SEQ ID NO:108),Flk-I1(R):5’-GGGACTAGTTTATGTGTATACTCTGTCAAAAATGGTTTC-3’(SEQ ID NO:109),(氨基酸792-1081)。对正向(F)引物下划线标出XhoI序列,对反向(R)引物下划线标出SpeI序列,黑体标出终止密码子。将PCR产物连接进pCR2.1-TOPO质粒(Invitrogen),通过测序确认,随后通过以XhoI和SpeI(New England Biolabs)双重消化而切离。将片段连接进基于pGG34的质粒下游,并融合至编码LLO蛋白的前441个残基的基因,该蛋白的表达由hly启动子驱动。pGG34质粒的构建在本文其他地方有详细描述。将所得的质粒电穿孔进PrfA缺陷型Lm菌株XFL-7,该菌株源自Lm菌株10403S。在补加了34μg/ml氯霉素和250μg/ml链霉素的脑心浸液(BHI,Difco)平板上选择阳性克隆。所得的菌株命名为Lm-LLO-Flk-E1、Lm-LLO-Flk-E2和Lm-LLO-Flk-I1。
Lm疫苗剂量的生长和制备
疫苗储备液于-80℃在1X PBS中的10%甘油中保存。将每种储备液在氯霉素/链霉素平板上划线接种并生长过夜。将单个集落用于在抗生素选择下的5ml BHI培养基的过夜培养物中生长。让该培养物在摇床中于37℃进一步扩增4h,生长直至微生物密度达到0.4-0.8OD600,这时洗涤微生物并在10%甘油中无菌冷冻,保持在-80℃直至使用。对产生的每批次储备液都测定效价。将单个批次用于一个连续的实验,不同批次用于每个重复,未观察到批次间差异。对每个批次通过Western印迹以抗PEST和抗LLO抗体检查融合蛋白的表达。对于每个剂量,选择一个小瓶,解冻,在稀释和使用前在1X PBS中洗涤两次;弃去未使用的微生物。
Lm-LLO-Flk-1疫苗对肿瘤生长的影响
将200μl PBS中的1×106个NT-2肿瘤细胞皮下注射进FVB/N小鼠胁腹。在肿瘤接种后第4天,用5×108个CFU的Lm-LLO-Flk-E1、Lm-LLO-Flk-E2或Lm-LLO-Flk-I1腹膜内免疫小鼠。该剂量确定为观察到对小鼠有不良反应的最低剂量的十分之一,并用在全部实验中。每周重复免疫,对全部实验都是一共3剂疫苗。在对照组中,小鼠接受了对照Lm疫苗–Lm-LLO-NY-ESO-1101-156。Lm-LLO-NY-ESO-1101-156用作不相关或第三方Lm疫苗作为针对LLO或李斯特菌感染的免疫应答的对照,我们通常将这种疫苗以可比浓度用作测试疫苗的对照。每3天用卡尺测量肿瘤,对每个肿瘤记录最短(宽度)和最长表面直径。采用如下等式进行肿瘤体积的计算:[(宽度)2×长度×0.52]。如果小鼠出现开放伤口或肿瘤达到直径20mm,就将它们处死。对经NT-2挑战而无肿瘤的存活小鼠在第一次接种后至少10周以相同的细胞系在对侧胁腹再次挑战。
肿瘤免疫荧光
在肿瘤接种后第64天,处死小鼠,手术切下NT-2肿瘤,在OCT冷冻介质中冷冻保存,并进行冷冻切片以提供8-10mm厚的切片。对于免疫荧光,将样品解冻,采用4%福尔马林固定。封闭(2.4G2条件化培养基/10%FBS/5%正常大鼠和小鼠血清)后,在加湿室中于37℃用封闭溶液中的一抗对切片染色1小时。按用于一抗染色的相同操作,以二抗对样品染色。根据制造商的说明进行DAPI(Invitrogen)染色。对HIF-1α的细胞内染色在PBS/0.1%Tween/1%BSA溶液中进行。采用含有抗褪色剂的封片溶液(Biomeda)对载玻片加盖玻片,静置24小时,并保持在4℃,直至采用Spot Image软件(2006版)和BX51系列Olympus荧光显微镜成像。采用Spot Image软件合并图像,采用Image Pr软件(2006版)在设门ROI后进行定量。所有的图像都是相同曝光时间捕获的三个不同通道的合并系列。对于采用抗CD31的微血管密度定量,我们的分析基于之前发表的工作,其中采用与测量小鼠肿瘤模型中MVD相似的策略。
转移研究和生物发光成像
对小鼠总共接种三次,接着在最后一次接种后第7天,在静脉内注射50,000个表达整合的荧光素酶报告基因的4T1细胞(4T1-Luc)。在成像前,将对应的底物D-荧光素以200μl PBS中5-10mg/小鼠腹膜内注射。在腹膜内注射氯胺酮(80mg/kg)/甲苯噻嗪(12mg/kg)(Sigma,St.Louis,MO)后在麻醉下,将小鼠置于Xenogen IVIS成像系统(X-100(Xenogen Corporation,Alameda,CA)的暗室中。照相和荧光图像用CCD相机捕获,荧光密度采用来自Xenogen的Living Image软件(2.11版)按制造商的说明进行定量。每周进行纵向成像,直至肿瘤接种后至少4周。所有的小鼠都在相同曝光和时间长度下成像。图像显示归一化的图形。对于病理学研究,进行相同的实验,只是在对肿瘤计数(手工)前将肺组织灌注、提取、蜡包埋和以H+E染色。
妊娠和伤口愈合安全性研究。
用对照Lm疫苗或Lm-LLO-Flk-1疫苗免疫六至八周龄FVB/N雌性小鼠,每周一次,连续三次。在第四周进行安全性研究。对于妊娠和繁殖力,让每组5只小鼠与单独安置的雄性交配。监测交配并通过阴道栓的存在来确认。测定妊娠前时间、出生时幼崽重量和总窝产崽数。用于本研究的伤口愈合测定按之前描述的方法进行。简而言之,将小鼠麻醉,去毛并用无菌布进清洁皮肤。采用无菌的皮肤活检工具(Acuderm)从皮肤打孔两个直径3mm的圆形伤口。不处理伤口,未观察到感染。监测伤口闭合的平均时间,当形成伤痕而没留下任何可见的痂时认为结束。
统计分析和定量方法。
采用非参数Mann-Whitney检验来分析数据。将对数秩卡方检验用于所有的存活数据。所有的统计学分析都采用Prism软件vs.4.0a(2006)完成。统计显著性基于p值<0.05。在所有的非转基因研究中,我们在每组包括至少8只小鼠。所有的研究都重复至少一次。
实例28
构建LLO-FLK-1构建体
一共测试了三个构建体,每个含有Flk-1的不同区域:E1(AA 68-277)、E2(AA 545-730)和I1(792-1081)(图49A)。区域基于预测的表位而选择。因为我们对在基于FVB/N的乳腺癌模型中测试这些疫苗感兴趣,所以我们决定克隆可能最适合于用于测试的模型单体型(即,FVB/N,H2q)的片段。所选的E1、E2和I1结构域含有几个可能的用于H-2q小鼠MHC I单体型的表位(图50A)。
每个片段被克隆成与截短的LLO蛋白的融合蛋白(图49A)。为了测试Lm-LLO-Flk-1构建体是否产生并分泌LLO-Flk-1融合蛋白,我们采用多克隆抗PEST抗体(图49B,下部)或抗LLO抗体(图49B,上部)通过Western印迹(图49B)分析了来自培养上清液的蛋白。检测到针对每个融合构建体的带:LLO-Flk-E1(~81kDa)、LLO-Flk-E2(~78kDa)和LLO-Flk-I1(~89kDa)。大约60-70kDa的带为内源LLO;发现截短的融合蛋白LLO为约60-50kDa。将抗LLO印迹用作对照,以显示我们的融合蛋白为LLO-Flk相关的。所有三个构建体都能够感染、生长和逃脱吞噬溶酶体,如通过在J774A.1巨噬细胞中的复制所证实的(图50D)。每种疫苗还能够针对克隆的Flk-1区域免疫小鼠,如通过离体IFN-g脾细胞应答所显示的(图49C)。在这些FVB/N ELISpot实验中用于再次挑战的肽起初作为免疫显性Flk-1表位作图在H2d Balb/c小鼠中。我们按常规在H2q模型中使用H2d作图的表位,因为H2d鉴定的表位也可用作H2q表位,这可能是由于H2d和H2q分子的高度同源性。
实例29
LM-LLO-FLK-1疫苗在表达HER-2/NEU的肿瘤模型中的治疗效果
为了测试我们的疫苗诱导Her-2/neu+乳腺肿瘤消退的能力,我们采用了NT-2肿瘤模型,该模型将大鼠Her-2/neu作为转基因过表达,并且最初源自FVB/N Her-2/neu转基因小鼠中的自发乳腺肿瘤。该NT-2细胞系不表达Flk-1分子,因此我们的目的抗原仅位于宿主脉管系统上。细胞在体外生长,并皮下移植进FVB/N小鼠的胁腹。在第4天,当形成了可触知的肿瘤(直径~4-5mm)时,接种小鼠,然后每周强化,一共接种三次。疫苗Flk-E1和Flk-I1能够诱导消退,在一些小鼠中到种植后64天完全消除(Flk-E1:2/8;Flk-I1:2/8)移植的肿瘤(图51A)。但是,Flk-E2不能控制肿瘤生长,其类似于经对照Lm处理的组。在肿瘤接种后第100天对肿瘤完全消退的小鼠以NT-2在相对侧再次挑战,未观察到再生长新的肿瘤,提示了长寿命的抗肿瘤免疫(图52A和B)。
通过以泛内皮细胞标志物CD31染色评估了64天肿瘤的微血管密度(MVD)并以核标志物DAPI复染。如所预料的,来自Flk-E2处理组的肿瘤中MVD与来自对照处理小鼠的肿瘤类似。但是,在Flk-I1处理小鼠中看到CD31+血管密度的降低,并在采用Flk-E1接种时观察到进一步的降低(图51C)。这种CD31+血管的降低与Flk-E1和Flk-I1处理组中核缺氧标志物缺氧诱导因子-1α(HIF-1α)的染色的增加相关,而不是对照组(图51D)。可以假设这些Her-2/neu+肿瘤的消退,除了肿瘤MVD的减少之外,还由于抗VEGFR2细胞毒性T细胞杀伤参与肿瘤血管生成的内皮细胞,从而可能引起肿瘤破坏或生长限制,导致观察到的消退。之后,所吞噬的肿瘤碎片可由引流淋巴结中的局部树突状细胞交叉呈递,并呈递至抗Her-2/neu CTL,而它们的表位之前已在FVB/N小鼠中作图。如果发生了这种分子间的表位扩展,我们可以预期,表现出最明显消退的小鼠可能也具有高频率的抗Her-2/neu CD8+T细胞。为了测试这种假设,我们从64天小鼠收获脾细胞,并进行IFN-g ELISpot,以来自Her-2/neu的三个不同区域的三个已知表位再次挑战。我们决定采用ELISpot测定法来测量抗Her-2/neu应答,这是因为我们之前已针对Her-2/neu分子的不同区域对CTL表位作图,并且ELISpot测定法对于检测低频率的特异性T细胞是足够灵敏的,不像几个细胞毒性测定法需要体外刺激和扩增。我们发现,Flk-E1和Flk-I1显示出最大的表位扩展,而Flk-E2显示出最弱的表位扩展(图51B,*p<0.05),与体内发现的肿瘤消退程度强相关(图51A)。
实例30
抗血管生成诱导的肿瘤消退依赖于表位扩展至内源肿瘤抗原
Her-2/neu表位扩展的存在提示:肿瘤消退可能不仅仅依赖于抗血管事件,而且还依赖于对肿瘤抗原HER-2/neu的免疫应答。为了测试这种假设,我们使用两种最有效的疫苗Flk-E1和Flk-I1重复了相同的实验,但是,除了野生型FVB/N小鼠的接种之外,我们还将NT-2细胞皮下注射进其起源品系FVB/N Her-2/neu转基因小鼠(其表现出对大鼠Her-2/neu分子的高度耐受性)。Flk-E1和Flk-I1又一次减慢了NT-2肿瘤在野生型FVB/N小鼠中的生长,如之前所证实的(图53A,左侧插图)。但是,在抗HER-2/neu应答被耐受所限制的转基因宿主中,我们观察到所有肿瘤的生长(图53A,右侧插图)。这二者的结果都反映了所观察到的在脾中(图53B)和如Flk-E1接种所示的在肿瘤部位处(图53C)所证实的向内源Her-2/neu蛋白的表位扩展。这提示:抗血管事件对于肿瘤消退是不够的,而是需要对肿瘤脉管系统和直接对肿瘤细胞的组合作用才能实现肿瘤死亡和最终的消退。
实例31
以LM-LLO-FLK-1疫苗片段接种可防止实验性转移的生长
抗血管生成疫苗的重要用途可为用于治疗或预防乳腺癌转移。转移的肿瘤细胞高度依赖于新血管的生长,尽管较小的肿瘤不完全依靠新的脉管系统。但是,据假设,一旦它们生长超过某个尺寸,肿瘤就变得高度依赖于新血管的形成,并因此成为抗VEGFR2CTL的可能的靶标。为了测试我们的疫苗是否可防止乳腺肿瘤散播,我们使用了涉及将体外培养的肿瘤细胞直接接种进小鼠尾静脉的实验性转移系统,使得能在几个下游器官特别是肺快速定植。由于在尾静脉接种后,NT-2模型不能很好地定植肺(数据未示出),所以我们采用了4T1,其为来自Balb/c小鼠的侵袭性小鼠乳腺癌细胞系。在三周的期间以Lm-LLO-Flk-E1或Lm-LLO-Flk-I1或对照Lm疫苗免疫Balb/c小鼠三次。然后向小鼠静脉内注射50,000个4T1细胞,也向该相同动物皮下注射。进行该皮下部位注射,以使得我们可测量原发肿瘤生长,而静脉内注射则模拟转移。与对照小鼠相比,经Flk-1疫苗处理的小鼠具有延期的肿瘤生长,减慢的原发性皮下肿瘤大小,增加的存活率,以及降低的发病率(图54)。与看到的针对原发性4T1肿瘤的较差应答不同,与对照小鼠相比,在经处理动物的每只动物中发现的种植速率和总的转移显著较低(图55A)。观察到低水平的向Her-2/neu的表位扩展(图55B),这可能是因为4T1弱表达小鼠Her-2/neu。
为了更严谨地测试该假设,即针对Flk-1的免疫可防止实验性转移在肺组织的种植,我们采用了生物发光模型,其中各个肿瘤细胞和肿瘤块可采用非侵入性成像来观察。在几轮Lm-Flk-E1和–I1接种后,向小鼠静脉内注射50,000个表达萤火虫荧光素酶基因的4T1细胞(4T1-Luc)。每周一次,将小鼠麻醉,注射荧光素酶底物(D-荧光素)并成像。肺种植到第11天是可见的,并且对照处理的小鼠到第25天变得快速被4T1-Luc细胞定植,而直至至少第32天,Lm-LLO-Flk-E1和Lm-LLO-Flk-I1处理的小鼠均未显示出任何肺种植的迹象,这时对照处理小鼠已发病并被处死(图55C)。在第32天,只有25%的Flk-1接种小鼠显示出任何肺肿瘤。有可能肿瘤块在这个时间点不能被该生物发光方法检测到,因为对于Lm-Flk-E1处理组在第25天观察到肿瘤细胞信号,而在第32天未观察到。第25天该非常小的信号低于1000个细胞阈值,并且可能在接下来的一周内失去一些细胞团块,以致低于该系统的检测限。经对照Lm免疫的小鼠因肺肿瘤而快速患病,但是Flk-E1和Flk-I1Lm接种延迟了肿瘤负荷、进展时间(对照处理的为第25天相对Flk-1处理的第32天)以及最终的患病(降低的发病率,如图54所示)。
实例32
以FLK-1免疫对小鼠中伤口愈合、妊娠或繁殖力无影响
为了评估Lm-LLO-Flk-1疫苗是否引起与血管生成抑制相关的毒性,我们在经免疫的小鼠中研究了伤口愈合、妊娠和繁殖力。小鼠在交配或者制造无菌伤口穿孔之前,以Lm-LLO-Flk-E1、Lm-LLO-Flk-E2、Lm-LLO-Flk-I1、对照Lm或单独的盐水免疫三次。我们观察交配小鼠从性交到妊娠的时长、平均到期幼崽质量和总窝产崽数。伤口穿孔是无菌的,但是小鼠关在一起。伤口愈合技术按之前描述的方法进行。给来自每个免疫组的五只小鼠剃毛,制造无菌伤口穿孔,每只动物两个,然后让它们随时间愈合。测量伤口闭合时间。当伤口闭合时没有留下的痂,认为完全伤口愈合完成。以Lm-LLO-Flk-E1、Lm-LLO-Flk-E2或Lm-LLO-Flk-I1免疫对繁殖力、妊娠时长或出生时幼崽质量没有影响(图56A)。类似地,免疫对伤口闭合所需的时间也没有显著影响(图56B)。
为了评估在Flk-I1免疫后观察到的对Her-2/neu的免疫应答是否是由于Flk-1和Her-2/neu之间的共有表位之间的交叉反应性,对经Flk-I1疫苗免疫的FVB/N小鼠评估对FLK-I1839-848的免疫,该FLK-I1839-848与大鼠Her-2/neu表位GSGAFGTVYK(SEQ ID NO:99)具有交叉反应性。以Lm-LLO-Flk-I1接种小鼠导致针对之前作图的Flk-I1表位PGGPLMVIV(SEQ ID NO:102)的极好应答。但是,未观察到对小鼠Flk-I1839-848表位或同源的大鼠Her-2/neu IC1732-741表位的明显应答(图57)。因此,在Flk-I1免疫后观察到的对Her-2/neu的免疫应答最可能是由于表位扩展,而不是由于共有表位之间的交叉反应性。
合起来,Lm-LLO-Flk-1疫苗能够清除一些建立的乳腺肿瘤,在余下肿瘤中减小微血管密度,防止肿瘤再次挑战和实验性转移以及诱导表位扩展至肿瘤相关抗原Her-2/neu的各个区域。发现肿瘤清除依赖于表位扩展至HER-2/neu,而不单单是由于减少肿瘤脉管系统。但是,疫苗效果既不影响正常的伤口愈合,也对妊娠没有有毒副作用。因此,抗血管生成疫苗可克服对宿主脉管系统的耐受,从而驱动表位扩展至内源肿瘤蛋白并驱动有效的肿瘤消退。因此,本文提供了采用单一疫苗来靶向肿瘤脉管系统和内源肿瘤抗原(Her-2/neu)的新方法。
实例33
逃逸突变体中出现突变
小鼠
在University of Pennsylvania的动物中心机构安置和饲养FVB/N Her-2/neu转基因小鼠。在实验开始时使用的小鼠为六至八周龄,这些实验根据宾夕法尼亚大学的实验动物管理与使用委员会的规章进行。
李斯特菌疫苗菌株。
所用的菌株为Lm-LLO-Flk-E1和Lm-LLO-Flk-I1。将菌株Lm-LLO-NYESO1用作用于抗原特异性第三方对照疫苗。将细菌在补加了34μg/ml氯霉素和250μg/ml链霉素的脑心浸液(BHI,Difco)平板上选择,然后在液体培养物中生长,以1ml的等分试样在-80℃冷冻。对于注射,在给药前以无菌PBS洗涤疫苗两次。
原部位肿瘤保护。
为了测试抗Flk-1李斯特菌疫苗影响自发出现的肿瘤的能力,我们采用了FVB/N大鼠Her-2/neu转基因雌性小鼠,其过表达大鼠Her-2/neu分子并自发产生乳腺肿瘤。对于这些长期保护研究,我们从6周龄开始每三周腹膜内免疫一次直至21周龄,一共免疫雌性小鼠(N=15)六次。疫苗Lm-LLO-Flk-E1、Lm-LLO-Flk-I1或Lm-LLO-NYESO-1作为PBS中的悬浮液以0.1LD50注射。每周一次跟踪肿瘤负荷。一旦肿瘤超过10mm尺寸,处死动物并取出肿瘤用于分析。采用GraphPad Prism软件,使用Kaplan-Meier对数秩检验进行原部位肿瘤生长差异的统计学分析,比较每一疫苗组和对照组之间肿瘤生长开始的时间。
突变的分析和作图。
新切下肿瘤,将其置于RNAlater溶液中,4℃下保存少于2周。我们采用Qiagen mRNA试剂盒(Invitrogen)从保存的肿瘤提取mRNA,然后通过PCR产生cDNA。将各个PCR样品进一步细分,使得能对Her-2/neu的500-800bp长度的每个单独片段测序(EC1、EC2、EC3、IC1、IC2),如在别出描述的(Singh,2007)。由Children’s Hospital of Philadelphia(CHOP)Sequencing Facility测序,然后采用4Peaks软件1.7.2分析。将没有在四个或更多个的单个PCR和测序反应中出现的突变作为PCR诱导的突变弃去。采用MacPyMol建立分子模型。
PCR引物序列:
EC1FP:AGGGCTGTCAGGTAGTGC(SEQ ID NO:110)
EC1RP:TGACCTCTTGGTTATTCG(SEQ ID NO:111)
EC2FP:ACCTGCCCCTACAACTAC(SEQ ID NO:112)
EC2RP:GACGCCCTCTACAGTTGC(SEQ ID NO:113)
EC3FP:GTGGATTGGCTCTGATTC(SEQ ID NO:114)
EC3RP:TGAGTTACAGACCAAGCC(SEQ ID NO:115)
IC1FP:CAAACGAAGGAGACAGAAG(SEQ ID NO:116)
IC1RP:CACCATCAAACACATCGG(SEQ ID NO:117)
IC2FP:CACTGCTGGAAGATGATG(SEQ ID NO:118)
IC2RP:TTTGTGGCGATGGAGACC(SEQ ID NO:119)
从6周龄开始,每3周以Flk-E1、Flk-I1或对照Lm接种表达大鼠Her-2/neu的转基因FVB/N小鼠,在最后一次接种之后每周测量肿瘤。与对照Lm接种比较,Flk-E1和Flk-I1接种增加了无肿瘤小鼠的百分比。在第35周和第40周之间,在Flk-E1和Flk-I1接种小鼠中有一些小鼠出现肿瘤。对来自每只小鼠的肿瘤检测突变的Her-2/neu信息。收集信使RNA,合成cDNA并测序。将得到的序列与野生型序列并排比对以确定突变的残基。只有出现4次或更多次的突变才考虑为真正突变(图58A)。几个“热点”内的突变残基或成串的突变残基处于之前作图的CTL表位内。一个这样的表位显示关键氨基酸中的突变导致了将该表位锚定至H2Dq MHC I分子(图58B)。
实例34
靶向乳腺和黑素瘤脑转移
采用上文描述的方法进行实验。
以每种疫苗、抗人Her-2/neu或对照接种NYESO1,免疫Balb/c小鼠三次。使稳定表达萤火虫荧光素酶基因的鼠乳腺癌细胞(来自University of Minnesota的John Ohlfest实验室的EMT6-Luc细胞)在体外生长,然后以5,000个细胞/小鼠注射进麻醉小鼠的脑中。EMT6-Luc细胞表达低水平的小鼠Her-2/neu(数据未示出)。让细胞生长,然后在指定的天数成像。尽管在NYESO1接种的小鼠中清楚地看到了脑转移,但是抗人Her-2/neu接种在实验性诱导转移后的第3、8和11天控制了脑肿瘤(图59A)。
以每种疫苗、抗人HMWMAA-C或对照接种NYESO1,免疫C57Bl/6小鼠三次。使B16F10-Luc小鼠黑素瘤细胞(来自UCSF的Jeff Miller实验室)在体外生长,然后以5,000个细胞/小鼠注射进麻醉小鼠的脑中。B16F10亲代品系不表达HMWMAA(个人交流),因此HMWMAA的唯一来源是周细胞和胶质细胞。用抗人HMW-MAA-C接种小鼠在实验性诱导转移后的第11和第15天减少了脑肿瘤(图59B)。因此,以HMW-MAAC或Her-2/neu接种防止了脑转移,甚至当肿瘤细胞不表达HMW-MAA时。
实例35
新的基于李斯特菌的抗CD105/内皮因子的疫苗的构建-治疗性的
表达内皮因子(图60)的相当大片段的Lm菌株的构建体当与LLO融合时不分泌该片段,因此,将其重设计为两个新的Lm构建体:Lm-LLO-CD105A(aa17-319)和Lm-LLO-CD105B(359-588),它们几乎覆盖了整个内皮因子基因(图61A),并包括位于之前被靶向的内皮因子(图60)区域外部的推定(采用RANKpep确定的)CTL表位。通过在这些新构建体中潜在地包括更多的免疫显性表位,将CTL表位池的扩充用来增强疫苗效果。通过进一步让融合蛋白变小并从构建体除去高度疏水性的区域,这些融合蛋白由Lm更好地分泌。将编码这些片段的基因克隆进CD105pGG-34(图61B)。Lm-LLO-CD105A(图62)和Lm-LLO-CD105B(图63)都表达并分泌适当大小的片段。
实例36
LM-LLO-CD105A和B影响BALB/C小鼠中乳腺肿瘤4T1的原发性和转移性生长
BALB/c小鼠4T1乳腺肿瘤为我们的乳腺肿瘤模型中更恶性的,因为当它植入乳腺中时快速转移,将其选择为实例8中所示的疫苗的首个测试。将2×105个4T1细胞植入Balb/c小鼠的乳腺脂肪垫中。以2×108个cfu的每种疫苗在第1、8和15天或在第4、11和18天接种小鼠。与未暴露的或对照接种小鼠相比,两种疫苗方案都显示了肿瘤生长的显著减慢(图64)。第32天时,处死小鼠,取出它们的肺,并检查转移性扩散。有趣的是,仅Lm-LLO-CD105B显示了表面肺转移统计学上显著的减少(图65)。
接下来考查了这些小鼠中的CTL应答。作为确定可能被CD8+T细胞识别的内皮因子分子的免疫原性区域的最初尝试,让两个片段接受RANKpep(http://bio.dfci.harvard.edu/RANKPEP/)和SYFPEITHI(http://www.syfpeithi.de/)分析。由此选择了两个对CD105A:AGPRTVTVM(SEQ ID NO:120)(Dd结合者)和对CD105B:AYSSCGMKV(SEQ ID NO:121)(Kd结合者)最有希望的肽。它们在内皮因子中的位置在图61A中标有下划线。
将这两种肽用在ELISpot分析中以刺激在最后一次接种后四天从显示在图16B中的小鼠(这些小鼠在第4、11和18天被接种)取得的脾细胞。但是,与来自HIV Gag的对照H-2d限制性肽相比,它们不刺激T细胞分泌干扰素-γ,这提示它们不是CTL表位(图55)。还分析了向由4T1低水平表达的两种内源肿瘤抗原的表位扩展。第一种为来自内源亲嗜性鼠白血病病毒的包膜糖蛋白gp70。已针对BALB/c小鼠,对具有Ld限制性的来自gp70的命名为AH1的表位SPSYVYHQF(SEQ ID NO:122)作图。有趣的是,发现Lm-LLO-CD105A和B都诱导表位扩展至该抗原。还研究了向HER-2/neu的表位扩展。使用HER-2/neu的细胞外结构域中两个已知的表位EC1和EC2以及来自细胞内结构域的一个。尽管与对照疫苗Lm-LLO-NY-ESO-1相比,对于任一种内皮因子疫苗,在针对IC1的IFN-γELISpot中没有观察到显著增加,但是采用Lm-LLO-CD105A疫苗有扩展至EC1和EC2的证据(图65)。
对来自小鼠的肿瘤检查抗原特异性浸润T细胞,采用FACS和四聚体分析从中收获脾细胞用于HER-2/neu和gp70特异性T细胞。观察到肿瘤中EC1、EC2和AH1特异性T细胞的显著增加,还观察到与Lm-LLO-NY-ESO-1接种的那些相比来自Lm-LLO-CD105接种小鼠的IC1特异性T细胞温和增加(图66)。
实例37
研究LM-LLO-CD105A和B的使用对源自FVB HER-2/NEU转基因小鼠的HER-2/NEU阳性乳腺肿瘤NT2生长的影响
采用可移植的HER-2/neu肿瘤NT2在FVB小鼠中的其他乳腺肿瘤模型中测试了该内皮因子疫苗。进一步地,将1×106个肿瘤细胞皮下植入FVB小鼠,将它们在第4、11和18天以Lm-LLO-CD105A和B免疫,每种疫苗2×108个cfu。将Lm-LLO-NY-ESO-1用作对照疫苗。两种疫苗都显著影响了肿瘤生长(图67),在第60天,50%的经Lm-LLO-CD105A免疫的小鼠无肿瘤,25%的经Lm-LLO-CD105B接种的小鼠无肿瘤,作为比较,未接种组或经Lm-LLO-NYESO1接种的组没有小鼠无肿瘤。
实例38:LLO胆固醇结合结构域的定点突变
采用以下策略对LLO进行定点突变,以便在CBD中引入失活点突变。所得的蛋白命名为“mutLLO”:
将LLO亚克隆进pET29b
野生型LLO的氨基酸序列为:
信号肽和胆固醇结合结构域(CBD)标有下划线,CBD中的3个关键性残基(C484、W491和W492)为加粗斜体。
在LLO的C末端添加6xHis标签(HHHHHH)。加了His标签的LLO的氨基酸序列为:
将编码加His标签的LLO蛋白的基因以NdeI/BamHI消化,并将NdeI/BamHI亚克隆进表达载体pET29b的NdeI和BamHI位点之间。编码LLO蛋白的基因的序列为:
从序列起点开始,标下划线的序列为NdeI位点、NheI位点、CBG编码区、6x His标签和BamHI位点。将在下一步中突变的CBD残基为加粗斜体。
重叠延伸拼接(SOE)PCR
步骤1:PCR反应#1和#2在pET29b-LLO模板上进行。PCR反应#1利用引物#1和#2扩增NheI位点和CBD(含)之间的片段,向CBD中引入突变。PCR反应#2利用引物#3和#4扩增CBD和BamHI(含)之间的片段,向CBD中引入突变(图69A)。
PCR反应#1循环:A)94℃2min30s,B)94℃30s,C)55℃30s,D)72℃1min,重复步骤B至D 29次(一共30个循环),E)72℃10min。
PCR反应#2循环:A)94℃2min30s,B)94℃30s,C)60℃30s,D)72℃1min,重复步骤B至D 29次(一共30个循环),E)72℃10min。
步骤2:将PCR反应#1和#2的产物混合,让它们退火(在突变的CBD编码区),并用引物#1和#4再进行PCR 25个循环(图8B)。PCR反应循环:A)94℃2min30s,B)94℃30s,C)72℃1min,重复步骤B至C 9次(一共10个循环),添加引物#1和#4,D)94℃30s,E)55℃30s,F)72℃1min,重复步骤D至F 24次(一共25个循环),G)72℃10min。
引物序列:
引物1:GCTAGCTCATTTCACATCGT(SEQ ID NO:126;NheI序列标有下划线)。
引物2:
(SEQ ID NO:127;CBD编码序列标有下划线;突变的密码子为加粗斜体)。
引物3:
(SEQ ID NO:128;CBD编码序列标有下划线;突变的密码子为加粗斜体)。
引物4:GGATCCTTATTAGTGGTGGTGGTGGTGGTGTTCGATTGG(SEQ ID NO:129;BamHI序列标有下划线)。
野生型CBD序列为ECTGLAWEWWR(SEQ ID NO:130)。
突变的CBD序列为EATGLAWEAAR(SEQ ID NO:131)。
突变的NheI-BamHI片段的序列为
实例39:以CTL表位替换LLO CBD的一部分
对LLO进行定点突变,以将CBD的9个氨基酸(AA)用来自抗原NY-ESO-1的CTL表位替换。CBD的序列(SEQ ID NO:130)被序列ESLLMWITQCR(SEQ ID NO:133;突变的残基标有下划线)替换,该序列含有来自NY-ESO-1的157-165HLA-A2限制性表位,命名为“ctLLO”。
所采用的亚克隆策略与之前的实例类似。
所用的引物如下:
引物1:GCTAGCTCATTTCACATCGT(SEQ ID NO:126;NheI序列标有下划线)。
引物2:
(SEQ ID NO:134;CBD编码序列标有下划线;突变(NY-ESO-1)的密码子为加粗斜体)。
引物3:
(SEQ ID NO:135;CBD编码序列标有下划线;突变(NY-ESO-1)的密码子为加粗斜体)。
引物4:GGATCCTTATTAGTGGTGGTGGTGGTGGTGTTCGATTGG(SEQ ID NO:129;BamHI序列标有下划线)。
所得的NheI/BamHI片段的序列如下:
实例40:双重CHER2-CA9李斯特菌疫苗对植入BALB/C小鼠乳腺中的4T1肿瘤生长的抗肿瘤效果。
实验细节:
产生重组Lm(LmddA-cHer2/CA9)。该Lm菌株表达并分泌嵌合Her2(cHer2)蛋白(在染色体上作为与基因组李斯特菌溶血素O(LLO)的融合蛋白)以及人碳酸酐酶9(CA9)的片段(采用质粒,作为融合至截短的LLO(tLLO)的融合蛋白),以便多重靶向肿瘤细胞。
疫苗效价:
LmddA-PSA–6.5×108
LmddA-CA9–1.4×1010
LmddA-cHER2–1.05×1010
双重cHer2-CA9(LmddA)–1.5×109
试验方案:
让4T1细胞在含有10%FBS、2mM L-Glu、1.5g/L碳酸氢钠、4.5g/L葡萄糖、1mM丙酮酸钠和10mM HEPES的RPMI中生长。在注射当天,用胰蛋白酶处理细胞,然后PBS中洗涤2次。对细胞计数并以7×103个细胞/50μl重悬。
将肿瘤植入每只小鼠的乳腺中。每组16只小鼠。3天后接种小鼠。在第4天,对每组中4只小鼠实施安乐死,并检查肿瘤生长。在第10天向小鼠给予每种疫苗的强化。在第11天,对每组中4只小鼠实施安乐死,并测量肿瘤。在第18天,对每组中4至5只小鼠实施安乐死,并测量肿瘤。在第21天,对每组中的剩余小鼠实施安乐死,并测量肿瘤。
结果
在第4天,几乎感知不到肿瘤,所以没有进行测量。
1/20/2012-
第11天
1/27/2012-
第18天
1/30/2012-
第21天
数值显示,双重疫苗(表达两种异源抗原的重组李斯特菌)最初(第11天)对肿瘤块有巨大影响(图70)。两只安乐死的小鼠没有肿瘤,另外的比对照小,与单独CA9和cHER2接种小鼠的尺寸类似。到第18天,在几个组的一些小鼠中测量到多个肿瘤。PBS和PSA对照小鼠具有比单独CA9和cHER2或双重疫苗组大得多的肿瘤。双重疫苗组具有一个异常值,具有大的肿瘤负荷,否则该组的均值应该是最小。该实验提前结束,因为几个组中的小鼠看起来非常病态并不断死亡。但是,在最后的测量中,双重疫苗组中的小鼠具有最小的肿瘤(图70)。这可能是由于早期看到的对肿瘤生长的控制水平。
总之,双重疫苗在4T1模型中显示了初始水平的肿瘤控制,其高于以单价疫苗或对照小鼠获得的水平,因为双重疫苗组在实验结束时具有最小的肿瘤负荷(参见图70)。
提供前述实例是为了更充分地阐述本发明的实施例。但是,它们绝不应解释为限制本发明的广泛范围。
实例41:通过同时表达和分泌LLO-PSA和TLLO-HMW-MAA2160-2258融合蛋白、引起对两种异源抗原的免疫应答而具有增强的抗肿瘤活性的重组单核细胞增多性李斯特菌载体的开发。
材料与方法:
pADV168质粒的构建。HMW-MAA-C片段通过XhoI和XmaI限制性内切酶双重消化而从pCR2.1-HMW-MAA2160-2258质粒切离。将该片段克隆进已经用XhoI和XmaI消化而切下E7基因的pADV134质粒中。将pADV168质粒电穿孔进电转化感受态dal(-)dat(-)大肠杆菌菌株MB2159,并筛选阳性克隆用于RFLP和序列分析。
Lmdd-143/168、LmddA-143/168和对照菌株LmddA-168、Lmdd-143/134和LmddA-143/134的构建。以pADV168或pADV134质粒转化Lmdd、Lmdd-143和LmddA-143。在补加了链霉素(250μg/ml)并且无D-丙氨酸的脑心浸液(BHI培养基)琼脂板上选择转化子。采用抗LLO、抗PSA或抗E7抗体通过Western印迹在细菌培养上清液中对各个克隆筛选LLO-PSA、tLLO-HMW-MAA2160-2258和tLLO-E7分泌。将对来自每个菌株的所选克隆在体外和体内评估毒力。每个菌株在体内传代两次以选择最稳定的重组克隆。简而言之,使来自每个构建体的所选克隆生长,并以1×108个CFU/小鼠腹膜内注射至4只小鼠的组。在第1和第3天收获脾,匀化并在BHI琼脂板上铺板。在第一次传代后,选择来自每个菌株的一个克隆,并在体内再传代一次。为了防止载体进一步减毒至损害其存活力的水平,在具有不同减毒水平的两个载体(Lmdd-143/168、LmddA-143/168)中产生构建体。
表达和分泌作为融合蛋白的两种抗原的工程化李斯特菌菌株LmddA244G的构建。将抗原Her2嵌合体在基因上融合至基因组李斯特菌溶血素O,将第二抗原HMW-MAA-C(HMC)融合至质粒中截短的李斯特菌溶血素O(图71A)。采用抗LLO和抗FLAG抗体通过Western印迹分别检测融合蛋白LLO-ChHer2和tLLO-HMC的分泌(参见图71B)。
溶血分析。为了确定基因组LLO引起吞噬溶酶体逃逸的能力,采用对照野生型10403S和LmddA244G-168的分泌上清液和作为靶细胞的山羊血红细胞进行溶血测定。
J774细胞中的体外胞内复制。采用鼠巨噬细胞样J774细胞系进行体外细胞内生长测定。简而言之,以任一种单价疫苗(LmddA164和LmddA168)或者二价免疫治疗以1:1MOI在无抗生素的培养基中感染J774细胞1小时。在1h感染后,用10μg/ml庆大霉素处理细胞以杀死细胞外细菌。以定期时间间隔收获样品,用水溶解细胞以对细胞内CFU的数量定量。将裂解液的十倍系列稀释液在BHI板上一式双份铺板,并对每个样品中的集落形成单位(CFU)计数。
体内毒力研究。以两种不同剂量(1×108和1×109个CFU/剂)的Lmdd-143/168、LmddA-143/168、LmddA-168、Lmdd-143/134或LmddA-143/134菌株腹膜内注射四只C57BL/6小鼠(7周龄)的组。跟踪小鼠2周,用于存活和LD50评估。根据其他的基于Lm的疫苗的前期经验,>1×108的LD50构成了可接受的值。
结果
在成功构建pADV168质粒后,为正确HMW-MAA序列的存在进行测序。这些新菌株中的该质粒表达和分泌特异于每个构建体的LLO融合蛋白。这些菌株是高度减毒的,LD50至少为1×108个CFU,对于actA缺陷型(LmddA)菌株可能高于1×109个CFU,该菌株缺乏actA基因,因而缺乏细胞间扩散的能力。
产生了重组Lm(LmddA-cHer2/HMC)。这种Lm菌株表达和分泌嵌合Her2(cHer2)蛋白(从染色体上,作为与基因组李斯特菌溶血素O(LLO)的融合蛋白)和HMW-MAA2160-2258(也称为HMW-MAA C或HMC)片段(采用质粒,作为与截短的LLO(tLLO)的融合蛋白),以同时靶向肿瘤细胞和肿瘤脉管系统,称作LmddA244G-168。采用抗FLAG和抗-LLO特异性抗体通过Western印迹检测两种融合蛋白tLLO-HMC和LLO-cHer2从LmddA244G-168的表达和分泌(图71B)。此外,将疫苗LmddA244G-168在小鼠体内传代两次以稳定LmddA-244G的毒力并证实其保持了重组融合蛋白的表达(图71B)。疫苗LmddA244G-168在小鼠体内传代两次后保持了其表达和分泌两种融合蛋白tLLO-HMC和LLO-cHer2的能力。
菌株LmddA244G-168将染色体LLO作为融合蛋白LLO-cHer2表达,该融合蛋白可影响LLO的功能性能力以引起吞噬溶酶体逃逸。为了确定这一点,采用对照野生型10403S和LmddA244G-168的分泌上清液和作为靶细胞的山羊血红细胞进行溶血测定。如图72A中所示,当与野生型高毒力的Lm菌株10403S相比,LmddA244G-168的溶血能力有1.5倍的降低。
此外,为了考查融合蛋白LLO-cHer2的表达是否不对LmddA-cHer2/HMC感染巨噬细胞及其细胞内生长有任何有害效应,采用小鼠巨噬细胞样细胞J774进行了细胞感染测定。图72B中指定的结果显示:表达单一或双重抗原的不同的基于李斯特菌的免疫治疗剂的细胞内生长行为是相似的,提示两种抗原的共表达不引起LmddA244G-168呈递细胞内靶蛋白用于免疫应答的能力的任何改变。
实例42:检测以Lmdd-244G/168免疫时引起的免疫应答和抗肿瘤效果。
在小鼠中研究了以Lmdd-244G-168菌株免疫时对cHer2和HMW-MAA的免疫应答,其中采用标准方法,例如检测针对这些抗原的IFN-γ生成。在NT2乳腺肿瘤模型中测试了双重表达载体的治疗功效。
IFN-γELISpot。我们在FvB小鼠中评估了二价免疫疗法产生特异于两种抗原Her2和HMW-MAA的免疫应答的能力。将小鼠(3只/组)以不同的免疫疗法例如LmddA134(Lm-对照)、LmddA164和LmddA244G/168在第0天免疫,在第14天强化。在第21天收获的脾中检测到Her2/neu特异性免疫应答。按照试剂盒说明进行IFN-γElispot测定,用特异于细胞内区域的肽表位(RLLQETELV)(SEQ ID NO.76)刺激脾细胞。
IFN-γELISA。在经免疫小鼠的脾细胞中HMW-MAA-C特异性免疫应答的产生通过以HMA-MAA-C蛋白刺激细胞2天来确定。通过采用小鼠干扰素-γELISA试剂盒进行的ELISA检测IFN-γ的释放。
抗肿瘤功效。采用小鼠NT2乳腺肿瘤模型考察抗肿瘤功效。将1×106个NT2细胞在第0天植入FvB小鼠,在右胁腹上建立的肿瘤在第6天开始用不同的免疫疗法处理,免疫三次,每次间隔一周。每周监测肿瘤两次,直至研究结束。如果肿瘤直径大于1.5cm,则让小鼠安乐死。
结果
接下来,采用小鼠NT2乳腺肿瘤模型考察了LmddA244G的抗肿瘤治疗功效。用表达单抗原LmddA164(ChHer2)、LmddA168(HMC)或二价免疫治疗剂LmddA244G-168的不同免疫疗法处理带有在右胁腹上建立的NT2肿瘤的FvB小鼠,免疫三次,每次间隔一周。单价和二价免疫疗法处理引起NT2肿瘤减小,如图73A和73C所示。但是,在二价免疫疗法处理后,观察到控制NT2肿瘤生长的更强的影响。另外的对每组中无肿瘤小鼠百分比的分析证实,当与单价免疫疗法(小于40%)处理组相比时,用二价免疫疗法处理产生了最大量的无肿瘤小鼠(70%)。这些观察结果支持:使用单核细胞增多性李斯特菌作为载体同时靶向两种抗原进行治疗产生了增强的抗肿瘤功效。
评估了二价免疫疗法在FvB小鼠中产生特异于两种抗原Her2和HMW-MAA的免疫应答的能力。将小鼠以不同的免疫疗法例如LmddA134(不相关对照)、LmddA164和LmddA244G/168在第0天免疫,在第14天强化。采用特异于细胞内区域的肽表位使用基于ELISpot的测定法检测Her2/neu特异性免疫应答。采用基于ELISpot的测定法检测,表达Her2的单价和二价免疫疗法都产生了可比水平的免疫应答(参见图74)。
采用ELISA检测免疫小鼠脾细胞中HMW-MAA-C特异性免疫应答的产生。采用基于单拷贝(单价免疫疗法)或多拷贝(二价免疫疗法)的表达从Lm表达肿瘤抗原产生可比水平的抗原特异性免疫应答(参见图74)。
实例43:双重cHER2-HMW-MAA李斯特菌疫苗对植入Balb/c小鼠乳腺中的4T1肿瘤生长的抗肿瘤功效。
实验细节:
产生了重组Lm(LmddA-cHer2/HMW-MAA)。这种Lm菌株表达并分泌嵌合Her2(cHer2)蛋白(在染色体上作为与基因组李斯特菌溶血素O(LLO)的融合蛋白)以及高分子量黑素瘤相关抗原(HMW-MAA)(采用质粒,作为融合至截短的LLO(tLLO)的融合蛋白),以便多重靶向肿瘤细胞。
表4
疫苗效价:
LmddA-PSA–6.5×108
LmddA-HMW-MMA–1.4×1010
LmddA-cHER2–1.05×1010
双重cHer2-HMW-MMA(LmddA)–1.5×109
试验方案:
让4T1细胞在含有10%FBS、2mM L-Glu、1.5g/L碳酸氢钠、4.5g/L葡萄糖、1mM丙酮酸钠和10mM HEPES的RPMI中生长。在注射当天,用胰蛋白酶处理细胞,然后PBS中洗涤2次。对细胞计数并以7×103个细胞/50μl重悬。
将肿瘤植入每只小鼠的乳腺中。每组16只小鼠。3天后接种小鼠。在第8天,对每组中4只小鼠实施安乐死,并检查肿瘤生长。在第8天向小鼠给予每种疫苗的强化。在第15天,对每组中4只小鼠实施安乐死,并测量肿瘤。在第15天向小鼠给予每种疫苗的再一次强化。在第15、第21、第28和第35天,对每组中4至5只小鼠实施安乐死,并测量肿瘤。在第42天,对每组中的剩余小鼠实施安乐死,并测量肿瘤。
结果
结果汇总在图75中。该图显示,双重疫苗(表达两种异源抗原的重组李斯特菌)对肿瘤体积有重大影响(图75)。在接受二价疗法的小鼠中肿瘤体积小于对照和单价HMW-MMA和cHER2接种小鼠的肿瘤体积。PBS和PSA对照小鼠具有在体积上与单价HMW-MMA和cHER2组可比的肿瘤。
总之,双重疫苗在4T1模型中显示了初始水平的肿瘤控制,其高于以单价疫苗或对照小鼠获得的水平,因为双重疫苗组在实验结束时具有最小的肿瘤负荷(参见图75)。
实例44:双重和顺序CHER2-HMW-MAA李斯特菌疫苗对NT2乳腺肿瘤模型生长的抗肿瘤功效的比较研究。
实验细节:
采用小鼠NT2乳腺肿瘤模型考察抗肿瘤功效。将1×106个NT2细胞在第0天植入FvB小鼠,在右胁腹上建立的肿瘤在第6天开始用不同的免疫疗法处理,免疫三次,每次间隔一周。每周监测肿瘤两次,直至研究结束。如果肿瘤直径大于1.5cm,则让小鼠安乐死。
表5
结果
采用小鼠NT2乳腺肿瘤模型考察了不同李斯特菌疫苗方案的抗肿瘤治疗功效。用表达单抗原LmddA164(ChHer2)、LmddA168(HMC)或表达同时给药的两种抗原的疗法组合(二价疗法)的不同免疫疗法处理带有在右胁腹上建立的NT2肿瘤的FvB小鼠,1×108免疫五次,每次间隔一周。另外,用以下不同的免疫疗法进行了组合疗法与顺序疗法的比较:表达单抗原LmddA164(ChHer2)、LmddA168(HMC)的免疫疗法,表达同时给药的两种抗原的组合疗法(二价疗法),或者每种单抗原顺序给药的组合(cHer2,然后是HMW-MAA)。在后者中,3次每周一剂的LmddA164(ChHer2)给药,然后是3次每周一剂的LmddA168(HMC)。结果汇总在图76中。所有的方案都引起NT2肿瘤体积大概等量的减小,如图76中所示。这些观察结果显示,两单价构建体的同时或者顺序给药在控制肿瘤生长方面与二价构建体至少是可比的(图76)。
已参考附图对本发明的优选实施例进行了描述,应当理解,本发明不限于精确的实施例,本领域技术人员可在不脱离所附权利要求书中定义的本发明范围和精神的情况下对其进行各种改动和修饰。
- 还没有人留言评论。精彩留言会获得点赞!