降低系统性调节性T细胞水平或活性来治疗CNS疾病和损伤的制作方法
本发明一般性地涉及用于通过短暂地降低循环中系统性免疫抑制的水平来治疗中枢神经系统(Central Nervous System,CNS)的疾病、障碍、病症或损伤的方法和组合物。
背景技术:
大部分的中枢神经系统(CNS)病理共有共同的神经炎性组分,其是疾病发展的一部分并且导致疾病恶化。在这些病理中,阿尔茨海默病(Alzheimer’s disease,AD)(一种年龄相关性神经变性疾病)以记忆和认知功能的进行性丧失为特征,其中提出淀粉样蛋白-β(Aβ)聚集体的累积在CNS内的炎性级联反应中起关键作用,最终导致神经元损伤和组织破坏(Akiyama等,2000;Hardy和Selkoe,2002;Vom Berg等,2012)。尽管神经变性疾病中发生慢性神经炎性应答,但是在过去十年内研究基于免疫抑制的神经变性疾病治疗的临床和临床前研究已提出为什么抗炎性药物达不到要求的问题(Breitner等2009;Group等,2007;Wyss-Coray和Rogers,2012)。我们提供了新的解决方案,其克服了AD以及类似CNS疾病和损伤的现有治疗的缺点;该方法基于我们对不同的系统性和中枢免疫系统组分在CNS维持和修复中的作用的独特了解。
技术实现要素:
在一个方面,本发明提供了用于治疗不包括自身免疫性神经炎性疾病—复发-缓解型多发性硬化(relapsing-remitting multiple sclerosis,RRMS)的CNS的疾病、障碍、病症或损伤的药物组合物,其包含引起个体中系统性免疫抑制的水平降低的活性剂,其中所述药物组合物用于通过包括至少两个治疗过程的给药方案来施用,每个治疗过程顺序地包括治疗期,之后是非治疗的间隔期。
在另一个方面,本发明提供了用于治疗不包括自身免疫性神经炎性疾病—复发-缓解型多发性硬化(RRMS)的中枢神经系统(CNS)的疾病、障碍、病症或损伤的方法,所述方法包括向有此需要的个体施用根据权利要求1至24中任一项所述的药物组合物,其中所述药物组合物通过包括至少两个治疗过程的给药方案来施用,每个治疗过程顺序地包括治疗期,之后是非治疗期的间隔期。
附图说明
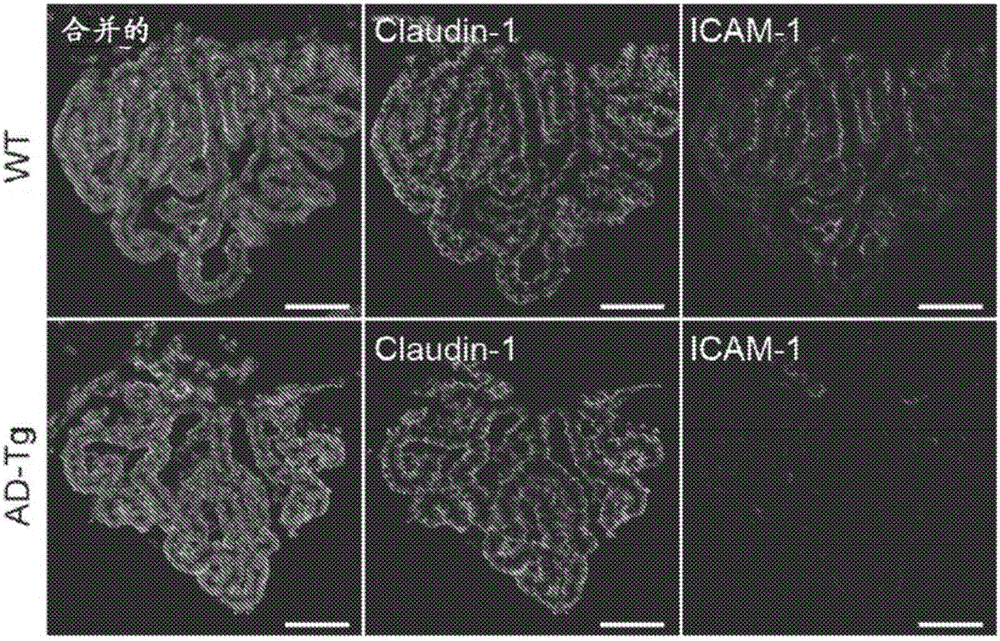
图1A至1B描绘了在AD的5XFAD转基因小鼠模型(AD-Tg)中随疾病进程的脉络丛(choroid plexus,CP)活性。(A)从1、2、4和8个月大AD-Tg小鼠分离的CP中通过RT-qPCR测量的基因icam1、vcam1、cxcl10和ccl2的mRNA表达水平,示为与年龄匹配WT(野生型)对照相比的倍数变化(n=6-8/组;学生t检验,针对每个时间点)。(B)经针对上皮紧密连接分子Claudin-1(Hoechst核染色)和整联蛋白配体ICAM-1免疫染色的8个月大AD-Tg小鼠和年龄匹配WT对照的CP的代表性显微镜图像(比例尺,50μm)。在所有图中,误差棒均表示平均值±s.e.m.;*:P<0.05;**:P<0.01;***:P<0.001。
图2A至2C示出了(A)未患CNS疾病的年轻人和年长人以及AD患者的人死后CP中ICAM-1免疫反应性的量化(n=5/组;单向ANOVA,接着是Newman–Keuls事后分析);(B)8个月大AD-Tg小鼠和年龄匹配WT对照的CP中表达IFN-γ的免疫细胞的流式细胞术分析(细胞内染色,并且对CD45预先门控)。阴影的直方图表示同型对照(n=4-6/组;学生t检验);以及(C)与年龄匹配WT对照相比的、从4和8个月大AD-Tg小鼠分离的CP组织中通过RT-qPCR测量的ifn-γ的mRNA表达水平(n=5至8/组;学生t检验针对每个时间点)。在所有图中,误差棒均表示平均值±s.e.m.;*:P<0.05;**:P<0.01;***:P<0.001。
图3A至3B描绘了(A)8个月大AD-Tg小鼠和WT对照小鼠中CD4+Foxp3+脾细胞频率(对TCRβ预先门控)的代表性流式细胞术图;以及(B)对来自1、2、4和8个月大AD-Tg小鼠和WT对照小鼠的脾细胞的定量分析(n=6-8/组;学生t检验针对每个时间点)。在所有图中,误差棒均表示平均值±s.e.m.;*:P<0.05;**:P<0.01;***:P<0.001。
图4示出了门控策略和在最后一次注射DTx后1天来自AD-Tg/Foxp3-DTR+/-小鼠的脾细胞的代表性流式细胞术图。连续4天腹膜内(i.p.)注射DTx,实现Foxp3+细胞的约99%消耗。
图5A至5G示出了在AD-Tg小鼠中短暂地消耗Treg的作用。(A)将AD-Tg/Foxp3-DTR+(其表达DTR转基因)和非DTR表达AD-Tg同窝出生小鼠(AD-Tg/Foxp3-DTR-)对照组用DTx处理连续4天。在最后一次DTx注射后1天,6个月大经DTx处理的AD-Tg小鼠中通过RT-qPCR测量的基因icam1、cxcl10和ccl2的CP mRNA表达水平(n=6-8/组;学生t检验)。(B-D)在最后一次DTx注射后3周,6个月大经DTx处理的AD-Tg小鼠和对照的脑实质(不包括脉络丛,将其单独切除)的流式细胞术分析。经DTx处理的AD-Tg/Foxp3-DTR+小鼠和AD-Tg/Foxp3-DTR-对照的脑实质中CD11b高/CD45高mo-MΦ细胞和CD4+T细胞数目增加的定量流式细胞术分析(B)以及CD4+Foxp3+Treg频率的代表性流式细胞术图(C)和定量分析(D)(n=3-7/组;学生t检验)。(E)在最后一次DTx注射后3周,6个月大经DTx处理的AD-Tg AD-Tg/Foxp3-DTR+和AD-Tg/Foxp3-DTR-对照的脑实质中foxp3和il10的mRNA表达水平(n=6-8/组;学生t检验)。(F)GFAP免疫染色的定量分析,其显示在最后一次DTx注射后3周,来自6个月大经DTx处理的AD-Tg/Foxp3-DTR+小鼠和AD-Tg/Foxp3-DTR-对照小鼠的海马切片中的星形神经胶质增生(astrogliosis)减轻(比例尺,50μm;n=3-5/组;学生t检验)。(G)在最后一次DTx注射后3周,脑实质中il-12p40和tnf-α的mRNA表达水平(n=6-8/组;学生t检验)。在所有图中,误差棒均表示平均值±s.e.m.;*:P<0.05;**:P<0.01;***:P<0.001。
图6A至6E示出了短暂地消耗Treg对Aβ斑学习/记忆表现的作用。在最后一次DTx注射后3周,5个月大经DTx处理的AD-Tg/Foxp3-DTR+小鼠和AD-Tg/Foxp3-DTR-对照小鼠的脑的(A)代表性显微镜图像和(B)定量分析,针对Aβ斑和Hoechst核染色来进行免疫染色(比例尺,250μm)。量化海马齿状回(dentate gyrus,DG)和大脑皮质的第5层中的平均Aβ斑面积和数目(在6μm脑切片中;n=5至6/组;学生t检验)。图6C至6E示出了在最后一次DTx注射后3周,6个月大经DTx处理的AD-Tg/Foxp3-DTR+小鼠和对照小鼠的Morris水迷宫(Morris water maze,MWM)测试表现。在短暂的Treg消耗之后,AD-Tg小鼠相对于AD-Tg对照在MWM的(C)获得、(D)探索和(E)反向相位中表现出较佳的空间学习/记忆表现(n=7-9/组;针对各成对比较双向重复测量ANOVA接着是Bonferroni事后分析;*:对于整体的获得、探索和反向P<0.05,)。在所有图中,误差棒均表示平均值±s.e.m.;*:P<0.05;**:P<0.01;***:P<0.001。
图7示出了与年龄匹配WT对照相比的、从6个月大和12个月大APP/PS1AD-Tg小鼠(阿尔茨海默病的小鼠模型(参见材料和方法))分离的CP中通过RT-qPCR测量的ifn-γ的mRNA表达水平(n=5-8/组;学生t检验)。误差棒表示平均值±s.e.m.;*:P<0.05。
图8A至8I示出了在AD-Tg小鼠中施用每周醋酸格拉替雷(Glatiramer acetate,GA)的治疗效果。(A)每周GA治疗方案的示意图。向小鼠(5个月大)皮下(s.c.)注射GA(100μg),在第一周期间注射两次(第1和4天),此后每周一次地注射,持续总共4周的时间。在最后一次注射后1周(MWM)、1个月(RAWM)和2个月(RAWM,使用不同的实验空间设置)检测小鼠的认知表现,并且检测海马炎症。图8B至8D示出了在6m龄时,未经治疗的AD-Tg小鼠和经每周-GA治疗的AD-Tg小鼠海马中基因的mRNA表达水平,其显示在经每周-GA治疗的小鼠中(B)诸如TNF-α、IL-1β和IL-12p40的促炎细胞因子的表达降低;(C)抗炎细胞因子IL-10和TGF-β升高,以及(D)神经营养因子IGF-1和BDNF升高(n=6-8/组;学生t检验)。在图8E至8G中,用每周-GA或者用载剂(PBS)处理AD-Tg小鼠(5个月大)并在6m龄时与年龄匹配的WT同窝出生小鼠在MWM任务中进行比较。相对于对照,经治疗的小鼠在MWM的(E)获得、(F)探索和(G)反向相位中表现出较佳的空间学习/记忆表现(n=6-9/组;针对各成对比较双向重复测量ANOVA接着是Bonferroni事后分析)。图8H至8I示出了在最后一次GA注射后1个月(H)或2个月(I),相同小鼠在RAWM任务中的认知表现(n=6-9/组;针对各成对比较双向重复测量ANOVA接着是Bonferroni事后分析)。数据是至少三次独立实验的代表。所有图中,误差棒均表示平均值±s.e.m.;*:P<0.05;**:P<0.01;***:P<0.001。
图9A至9H进一步示出了在AD-Tg小鼠中施用每周-GA的治疗效果。A至B示出了经每周-GA或载剂(PBS)处理并在施用方案的第1周结束时(在总共两次GA注射之后)检测的5XFAD AD-Tg小鼠。与年龄匹配WT对照相比的、经处理的6个月大AD-Tg小鼠中CD4+Foxp3+脾细胞频率(A)和CP IFN-γ表达免疫细胞(B;经细胞内染色并对CD45预先门控)的流式细胞术分析(n=4-6/组;单向ANOVA,之后是Newman–Keuls事后分析)。(C)4个月大AD-Tg小鼠的CP中通过RT-qPCR测量的基因icam1、cxcl10和ccl2的mRNA表达水平,所述小鼠用每周-GA或载剂处理并在每周-GA方案的第1周或第4周结束时进行检测(n=6-8/组;单向ANOVA,之后是Newman–Keuls事后分析)。图9D至9E示出了在每周-GA之后、来自6个月大AD-Tg/CX3CR1GFP/+BM嵌合体的脑切片的代表性图像。CX3CR1GFP细胞位于经每周-GA处理的AD-Tg小鼠中的第三脑室的CP(3V;i)、邻近脑室间隙(ii)和侧脑室的CP(LV;iii)(D;比例尺,25μm)。共聚焦z轴叠加的代表性正交投影,其显示在经每周-GA处理的7个月大AD-Tg/CX3CR1GFP/+小鼠的CP中GFP+细胞与髓系标志物CD68共存,而在对照经PBS处理的AD-Tg/CX3CR1GFP/+小鼠中不共存(E;比例尺,25μm)。(F)CX3CR1GFP细胞与髓系标志物IBA-1共位于经GA处理的AD-Tg/CX3CR1GFP/+小鼠的脑中的Aβ斑附近,并且共表达髓系标志物IBA-1(比例尺,25μm)。图9G至9H示出了在每周-GA方案的第2周从4个月大WT小鼠、未经处理AD-Tg小鼠和AD-Tg小鼠的海马分离的细胞的代表性流式细胞术图。对CD11b高/CD45高mo-MΦ门控(G)并量化(H;n=4-5/组;单向ANOVA,之后是Newman–Keuls事后分析)。在所有图中,误差棒均表示平均值±s.e.m.;*:P<0.05;**:P<0.01;***:P<0.001。
图10A至10H描绘了在AD-Tg小鼠中施用p300抑制剂(C646)的治疗效果。在图10A至10B中,用p300i或载剂(DMSO)处理年长小鼠(18个月)持续1周的时间并在终止处理后1天进行检测。显示在p300i处理之后脾中表达IFN-γ的CD4+T细胞的频率升高(A)和CP中表达IFN-γ的免疫细胞数目升高(B)的代表性流式细胞术图。图10C至10E示出了10个月大AD-Tg小鼠脑中的Aβ斑负荷的代表性显微镜图像(C)和定量分析,所述小鼠接受p300i或载剂(DMSO)持续1周的时间并且随后在另外的3周之后进行检测。针对Aβ斑并通过Hoechst核染色对脑进行免疫染色(n=5/组;比例尺,250μm)。在海马DG(D)和大脑皮质的第5层(E)中量化平均Aβ斑面积和斑数目(在6μm脑切片中;n=5-6/组;学生t检验)。(F)对不同组的7月龄AD-Tg小鼠的p300i处理(或作为载剂的DMSO)施用方案(1或2个时期)的示意图。图10G至10H示出了相对于未经处理AD-Tg组的大脑皮质(第5层)的Aβ斑覆盖百分比的变化平均值(G)以及大脑可溶性Aβ1-40和Aβ1-42蛋白平均水平的变化(H)(未处理组中的Aβ1-40和Aβ1-42平均水平分别为90.5±11.2和63.8±6.8pg/mg总分数;n=5-6/组;单向ANOVA,之后是Newman–Keuls事后分析)。在所有图中,误差棒均表示平均值±s.e.m.;*:P<0.05;**:P<0.01;***:P<0.001。
图11A至11B示出了在AD-Tg小鼠中施用抗PD1抗体的治疗效果。(A)实验组的小鼠、其年龄、处理施用方案以及检测小鼠的时间点的示意图。在10月龄时,在实验的第1天和第4天向5XFAD阿尔茨海默病(AD)转基因(Tg)小鼠腹膜内(i.p.)注射250μg抗PD1抗体(RMP1-14)或对照IgG(大鼠)抗体并在3周后通过放射臂水迷宫(radial arm water maze,RAWM)空间学习和记忆任务来检测它们的认知表现。使用年龄匹配的未经处理WT和AD-Tg小鼠作为对照。(B)示出了通过放射臂水迷宫(RAWM)空间学习和记忆任务评估的认知表现。通过双向重复测量ANOVA来分析数据并使用Bonferroni事后程序来进行后续成对比较。n=6-12/组。在所有图中,误差棒均表示平均值±s.e.m.;*:P<0.05;**:P<0.01;***:P<0.001。
图12A至12B示出了在AD-Tg小鼠中施用抗PD1抗体对产生IFN-γ+的T细胞的系统性作用。(A)实验组的小鼠、其年龄、处理施用方案以及检测小鼠的时间点的示意图。在实验的第1天和第4天向小鼠腹膜内注射250μg抗PD1抗体(RMP1-14)或对照IgG(大鼠)抗体并在第7天检测。(B)经PD-1或IgG处理的AD-Tg小鼠以及未经处理的AD-Tg和WT对照中CD4+IFN-γ+T细胞脾细胞频率(经细胞内染色并且对CD45和TCR-β预先门控)的流式细胞术分析(n=4-6/组;单向ANOVA,之后是Newman–Keuls事后分析;**:P<0.01,指定的处理组之间;误差棒表示平均值±s.e.m.)。
图13A至13B示出了在AD-Tg小鼠中进行抗PD1处理之后对CP的作用。将10月龄的AD-Tg小鼠用PD-1、IgG进行处理或者不予处理。在实验的第1天和第4天向小鼠腹膜内注射250μg抗PD1抗体(RMP1-14)或对照IgG(大鼠)抗体并在第7天检测。(A)通过实时定量PCR(RT-qPCR)测量的CP IFN-γ水平,与通过流式细胞术测量的CD4+IFN-γ+T细胞脾细胞频率成正相关(皮尔逊r=0.6284,P<0.05)和负相关。当将CP IFN-γ水平与CD4+Foxp3+CD25+Treg脾细胞频率(n=3-4/组)进行比较时,在相同小鼠中观察到相反的负趋势。(B)相同小鼠的CP中通过RT-qPCR测量的基因cxcl10和ccl2的mRNA表达水平(n=3-4/组;单向ANOVA,之后是Newman–Keuls事后分析)。在所有图中,误差棒均表示平均值±s.e.m.;*:P<0.05。
图14A至14B示出了当比较1个处理疗程相对于2个处理疗程时,在AD-Tg小鼠中施用抗PD1抗体的治疗效果。使接受了1个抗PD1处理疗程的图11A至11B中所述的小鼠组中的一半小鼠在第一RAWM任务之后再接受一个抗PD1处理疗程,或者不予处理。在另外的3周之后,通过RAWM使用用于认知学习和记忆的空间线索的不同且新的实验设置来测试所有小鼠。(A)实验组的小鼠、其年龄、处理施用方案以及检测小鼠的时间点的示意图。对于每个处理过程,向小鼠腹膜内注射250μg抗PD1抗体(RMP1-14)或对照IgG(大鼠)抗体。(B)示出了通过RAWM空间学习和记忆任务评估的认知表现。使用双向重复测量ANOVA来分析数据并使用Bonferroni事后程序来进行后续成对比较。n=6-12/组。在所有图中,误差棒均表示平均值±s.e.m.;*:P<0.05;**:P<0.01;***:P<0.001。
图15A至15H示出了通过全反式视黄酸(all-trans retinoic acid,ATRA)提高的系统性Treg水平对AD病理的不利影响。图15A至15B示出了显示5个月大AD-Tg小鼠中CD4+/Foxp3+/CD25+Treg脾细胞的频率升高的代表性流式细胞术图(A)和定量分析(B),所述小鼠接受ATRA或载剂(DMSO)持续1周的时间(n=5/组;学生t检验)。图15C至15F示出了AD-Tg小鼠脑中的Aβ斑负荷和星形神经胶质增生的代表性显微镜图像(C)和定量分析(D、E、F),所述小鼠在5月龄时接受ATRA或载剂(DMSO)持续1周时间的处理,并且随后在另外的3周之后进行检测。针对Aβ斑、GFAP(标记星形神经胶质增生)并通过Hoechst核染色对脑进行免疫染色(n=5/组;比例尺,250μm)。在海马DG和大脑皮质的第5层中量化平均Aβ斑面积和斑数目,并在海马中测量GFAP免疫反应性(在6μm脑切片中;n=5-6/组;学生t检验)。(G)AD-Tg小鼠大脑脑实质中通过ELISA量化的可溶性Aβ1-40和Aβ1-42的水平,所述小鼠在5月龄时用ATRA或载剂(DMSO)处理1周的时间并且随后在另外的3周之后进行检测(n=5-6/组;学生t检验)。(H)AD-Tg小鼠在RAWM任务中的认知表现,所述小鼠在5月龄时用ATRA或载剂(DMSO)处理1周的时间并且随后在另外的3周之后进行检测(n=5/组;双向重复测量ANOVA,之后是针对各成对比较的Bonferroni事后分析)。在所有图中,误差棒均表示平均值±s.e.m.;*:P<0.05;**:P<0.01;***:P<0.001。
图16A至16F示出了通过每周-GA施用提高的系统性Treg水平对AD病理的不利影响。(A)与每周-GA方案相比的每日-GA处理方案的图示。在每日-GA处理组中,每日向小鼠皮下注射100μg GA,持续1个月的时间。(B)与年龄匹配WT和未经处理AD-Tg小鼠相比,经每日-GA和每周-GA处理的7个月大AD-Tg小鼠的认知表现,其如通过在RAWM学习和记忆任务中每天的平均错误次数所评估(n=6-8/组;单向ANOVA,之后是Newman–Keuls事后分析)。(C)未经处理AD-Tg小鼠和经每日或每周-GA处理的AD-Tg小鼠的大脑皮质和海马(HC)的代表性显微镜图像,所述小鼠经针对Aβ斑和Hoechst核染色进行免疫染色(比例尺,250μm)。图16D-16F示出了经GA处理的AD-Tg小鼠(每日-GA和每周-GA组)和未经处理AD-Tg小鼠中的Aβ斑尺寸和数目(每6μm切片)的量化。经每周-GA处理的AD-Tg小鼠显示作为其海马齿状回(DG)总面积百分比的Aβ斑负荷降低,以及平均Aβ斑数目减少(n=6/组;单向ANOVA,之后是Newman–Keuls事后分析)。在所有图中,误差棒均表示平均值±s.e.m.;*:P<0.05;**:P<0.01;***:P<0.001。
具体实施方式
根据本发明已发现,在阿尔茨海默病的小鼠模型(AD-Tg小鼠)中短期地暂时消耗Foxp3+调节性T细胞(Treg)导致白细胞通过脑脉络丛向CNS的募集提高、CNS浸润性抗炎单核细胞源性巨噬细胞mo-MΦ和CD4+T细胞的数量升高并且在脑中聚集的Foxp3+Treg显著富集。此外,单一治疗期的长期作用导致海马神经胶质增生减轻并且脑中促炎细胞因子的mRNA表达水平降低。重要的是,对疾病病理的作用包括AD-Tg小鼠中表现出稳健Aβ斑病理的两个脑区域海马齿状回和大脑皮质(第5层)中的大脑淀粉样蛋白β(Aβ)斑负荷减小。最重要的是,在Treg的短期暂时消耗之后空间学习和记忆显著改善,达到与野生型小鼠近似的认知表现(实施例2和3)。总而言之,这些发现证明,短期Treg消耗接着无干扰时期导致短暂地破坏AD-Tg小鼠中Treg介导的系统性免疫抑制,其使得消炎细胞mo-MΦ和Treg能够向脑募集并且导致神经炎性应答消退、Aβ清除和认知下降逆转。这些发现强烈反驳该研究领域中的常识,根据所述常识,提高系统性免疫抑制将导致神经炎性应答减轻。相反,我们的发现显示需要通过短期地、短暂地且暂时地降低Treg介导的系统性抑制来加强系统性应答以在脑中实现消炎免疫细胞聚集(包括Treg自身),由此抵抗AD病理。
如下文所详述的,已通过使用数个独立的实验范例来证明本文中提供的发明人方法的特异性。简言之,首先发明人在两个不同的施用方案中使用了免疫调节性化合物,所述施用方案对外周Treg水平、CP活化和疾病病理产生相反的作用:每日施用方案,其增强外周Treg水平(Weber等,2007);和每周施用方案,发明人发现其降低外周Treg水平(实施例3和实施例5)。当在AD-Tg小鼠中通过Treg的短暂体内基因消耗(实施例2)或者通过其Foxp3功能的药理学抑制(实施例3和4)来证明这些操作导致CP活化促进白细胞向CNS运输、在病理部位发生消炎免疫细胞聚集、大脑Aβ斑清除和脑实质的免疫环境偏向炎症消退时,发明人还提供了外周Treg水平和疾病病理之间的直接功能性联系。
根据本发明还已发现,不频繁地施用通用抗原Copolymer-1持续有限的时间(代表1个治疗期)降低Treg介导的系统性免疫抑制并且通过提高脑脉络丛门活性而改善白细胞向CNS中的选择性浸润,从而对阿尔茨海默病病理产生显著的有益作用(实施例3),而增强Treg免疫抑制的Copolymer1的每日施用(Hong等,2005;Weber等,2007)显示对疾病病理无有益作用或者甚至有一定程度的不利影响(实施例5)。本发明的发明人在本文中还表明,通过用特异性小分子抑制剂(p300i)抑制p300或者使PD-1-抗体与PD-1受体相互作用来直接干扰Foxp3Treg活性提高AD-Tg小鼠中的脉络丛门活性并且减轻阿尔茨海默病病理(实施例4)。
重要的是,发明人提供的这些实例分别证明了导致系统性免疫抑制短期减低的不同干预:Copolymer-1充当免疫调节性化合物,p300i充当降低Foxp3乙酰化和Treg功能的小分子,而抗PD-1用作Treg上表达的PD-1的中和抗体。这些治疗方法用于短暂地增强外周中的免疫应答的短期治疗,其主要通过升高外周IFN-γ水平和IFN-γ产生细胞,由此活化脑脉络丛,从而允许T细胞和单核细胞向CNS中选择性地浸润并且这些细胞向病理和神经炎症部位归巢。在本文中还发现,间插有非治疗间隔期的重复治疗期相对于单一治疗期而言显著提高治疗效力(实施例4)。紧接着的非治疗时间间隔允许脑中的Treg水平和活性短暂地增强,促进神经炎症消退并且诱导有利于CNS愈合和修复的环境条件,随后导致组织恢复。在所有这些情况下,对脑病理的作用均是稳健的,涉及神经炎性应答消退、淀粉样蛋白β斑从AD小鼠脑清除和认知下降逆转。还已在AD的转基因小鼠模型中使用Foxp3+调节性T细胞的短暂消耗的遗传模型来证明当前方法的特异性(实施例2)。
因此,根据本发明已发现Foxp3+CD4+Treg介导的系统性免疫抑制干扰抵抗AD病理的能力,其至少部分地通过抑制协调消炎白细胞向CNS募集所需的IFN-γ依赖性CP活化来起作用(Schwartz和Baruch,2014b)。系统性Treg是维持自身免疫稳态并保护免受自身免疫性疾病的关键(Kim等,2007)。然而,我们的发现表明:在神经变性病症下,当脑中需要修复性免疫应答时,引发该应答的能力被系统性Treg干扰。尽管如此,根据我们的结果,脑中需要Treg,其向神经病理部位归巢并且局部执行抗炎活性。本发明代表针对在抵抗如AD的进行性神经元死亡方面的明显矛盾需求的独特且意料之外的解决方案:为了增加患病脑中的Treg而短暂地降低/抑制循环中的Treg。因此,短期且暂时降低外周免疫抑制对病理产生长期作用,所述降低允许抗炎细胞(包括Treg和mo-MΦ)向脑斑部位募集。然而,显著的是,短暂地降低系统性Treg水平或活性可有助于经由另外机理的疾病减轻,所述机理包括支持CNS特异性的保护性自身免疫应答(Schwartz和Baruch,2014a),或提高在清除血管Aβ中发挥作用的循环单核细胞的水平(Michaud等,2013)。
尽管病因不同的神经变性疾病共有共同的局部神经炎性组分,但是我们的结果强烈反驳将所有的CNS病理简单化地表征为会一律从系统性抗炎治疗受益的疾病。因此,尽管例如复发-缓解型多发性硬化(RRMS)的自身免疫性炎性脑病理受益于连续地系统性施用抗炎药物和免疫抑制性药物来实现持久的外周免疫抑制,但在慢性神经变性疾病例如在AD、原发性进行性多发性硬化(primary progressive multiple sclerosis,PP-MS)和继发性进行性多发性硬化(secondary-progressive multiple sclerosis,SP-MS)的情况下这将是无效的或者对病理产生不利影响(实施例5)。此外,我们的发现揭示了关于系统性Treg相对于组织相关性Treg在这些病理中的作用的错误认知(He和Balling,2013)。由于免疫-脑轴是终身脑可塑性的一部分(Baruch等,2014)并且神经变性疾病主要是年龄相关性的,因此我们的本发现还表明更具一般性的现象,其中系统性免疫抑制干扰脑功能。因此,短期的周期性降低系统性免疫抑制过程可代表适用于广泛范围脑病理(包括AD和年龄相关性痴呆)的治疗方法或者甚至预防方法。
重要的是,本文中在AD小鼠模型中提供的发明人方法和发现不是直接地靶向AD中的任何疾病特异性因子(例如淀粉样蛋白β或tau病理学)而是证明了预期在临床上适用于广泛范围CNS病理的新方法-短暂地降低Treg介导的系统性免疫抑制以增强消炎免疫细胞向CNS内的病理部位募集。
鉴于上述意想不到的结果,本发明提供了一种用于治疗不包括自身免疫性神经炎性疾病—复发-缓解型多发性硬化(RRMS)的CNS的疾病、障碍、病症或损伤的药物组合物,其包含导致个体中系统性免疫抑制的水平降低的活性剂,其中所述药物组合物用于通过包括至少两个治疗过程的给药方案来施用,每个治疗过程顺序地包括治疗期,之后是非治疗的间隔期。
在某些实施方案中,对所述给药方案进行校正使得短暂地降低系统性免疫抑制的水平。
本文中使用的术语“治疗”指获得期望生理效应的手段。就部分或完全地治疗疾病和/或由该疾病引起的症状而言,所述效应可以是治疗性的。该术语指抑制疾病,即停止或减慢其发展;或者改善疾病,即促使疾病消退。
术语“非治疗期”在本文中可与术语“无治疗时期”互换使用,并且指在此期间不向所治疗的个体施用活性剂的时期。
本文中使用的术语调节性T细胞的“系统性存在”指调节性T细胞在循环免疫系统(即,血液、脾和淋巴结)中的存在(如通过其水平或活性所测量的)。免疫学领域中众所周知的事实是:脾中的细胞群谱反映在血液中的细胞群谱(Zhao等,2007)。
本发明的治疗适用于表现出系统性免疫抑制升高的患者和未表现出这样的升高的患者二者。有时,需要根据本发明的治疗的个体具有一定的外周免疫抑制水平,其反映在循环中Treg的频率或数目升高,和/或Treg的功能活性增强和/或IFNγ产生白细胞减少和/或响应于刺激的白细胞增殖降低。Treg频率或数目的升高可以以总数目计或者作为总CD4细胞的百分比计。例如,根据本发明已发现,与野生型小鼠相比,阿尔茨海默病动物模型的CD4细胞中具有较高的Foxp3频率。然而,在所述个体中,即使系统性Treg细胞的水平未升高、其功能活性未增强、IFNγ产生白细胞的水平未降低或者响应于刺激的白细胞增殖未降低,降低系统性免疫抑制水平或活性的本发明方法仍有效地治疗不包括自身免疫性神经炎性疾病—RRMS的CNS的疾病、障碍、病症或损伤。重要的是,所述系统性免疫抑制还可涉及除Treg之外的另外免疫细胞类型,例如髓源性抑制细胞(myeloid-derived suppressor cell,MDSC)(Gabrilovich和Nagaraj,2009)。
系统性免疫抑制的水平可通过本领域普通技术人员公知的多种方法来进行检测。例如,可通过如下对外周血单核细胞和T淋巴细胞进行流式细胞术来测量Treg的水平:针对Treg的细胞表面标志物或核细胞内标志物(Chen&Oppenheim,2011)、CD45、TCR-β或淋巴细胞的CD4标志物进行免疫染色并测量与所述细胞特异性结合的抗体的量。Treg的功能活性可通过多种测定来测量;例如,常用的是胸苷掺入测定,其中通过[3H]胸苷掺入或者通过使用CFSE(5-(和6)-羧基荧光素二乙酸琥珀酰亚胺酯(其能够进入细胞)来测量针对抗CD3mAb刺激的CD4+CD25-T细胞(传统T细胞)增殖的抑制;以CFSE荧光强度的连续减半来测量细胞分裂)。IFNγ产生白细胞的数目或者其活性或其增殖能力可容易地由技术人员使用本领域中已知的方法来评估;例如,可如下测量IFNγ产生白细胞的水平:在短暂的离体刺激和golgi-stop并通过IFNγ细胞内染色进行免疫染色(使用例如BD Biosciences Cytofix/cytopermTM固定/透化试剂盒)之后对外周血单核细胞进行流式细胞术分析;收集这些细胞的条件培养基并使用ELISA来量化分泌的细胞因子的水平;或者比较条件培养基中不同细胞因子(例如IL2/IL10、IL2/IL4、INFγ/TGFβ)的比率等。人外周血中MDSC的水平可容易地由技术人员例如通过使用DR-/LIN-/CD11b+、DR-/LIN-/CD15+、DR-/LIN-/CD33+和DR(-/低)/CD14+细胞频率的流式细胞术分析来评估,如(Kotsakis等,2012)所述的。
在人中,当循环中Treg的总数比健康对照群体中高10%、20%、30%、40%、50%、60%、70%、80%、90%或100%或者更多;与健康对照群体中相比Treg细胞在总CD4+细胞中的百分比升高10%、20%、30%、40%、50%、60%、70%、80%、90%或100%或者更多;或者与健康对照群体中相比Treg的功能活性升高10%、20%、30%、40%、50%、60%、70%、80%、90%或100%或者更多时,可认为外周/系统性免疫抑制是升高的。或者,当IFNγ产生白细胞的水平或其活性相对于健康对照群体降低10%、20%、30%、40%、50%、60%、70%、80%、90%或100%;或者响应于刺激的白细胞增殖相对于健康对照群体降低10%、20%、30%、40%、50%、60%、70%、80%、90%或100%时,可认为外周/系统性免疫抑制是升高的。
当将药剂施用于个体之后,该个体的循环中Treg的总数与施用该药剂之前的水平相比降低10%、20%、30%、40%、50%、60%、70%、80%、90%或100%;Treg细胞在总CD4+细胞中的百分比相对于健康对照群体下降10%、20%、30%、40%、50%、60%、70%、80%、90%或100%,或者Treg的功能活性与施用该药剂之前的水平相比降低10%、20%、30%、40%、50%、60%、70%、80%、90%或100%时,可认为该药剂是导致系统性免疫抑制的水平降低的药剂。或者,当将药剂施用于个体之后,相对于健康对照群体IFNγ产生白细胞的总数或其活性增加10%、20%、30%、40%、50%、60%、70%、80%、90%或100%;或者响应于刺激的白细胞增殖增加10%、20%、30%、40%、50%、60%、70%、80%、90%或100%时,可认为该药剂是导致系统性免疫抑制的水平降低的药剂。
根据本发明使用的药剂可以是下调调节性T细胞的水平或活性或者干预其活性的任何药剂,但是可替代性地限于不包括选自以下组成组的药剂:(i)多巴胺或其药学可接受盐;(ii)多巴胺前体或其药学可接受盐;(iii)多巴胺受体1型家族的激动剂(D1-R激动剂)或其药学可接受盐;和(iv)多巴胺受体2型家族的拮抗剂(D2-R拮抗剂)或其药学可接受盐,即使不知道这些药剂依照根据本发明的治疗过程来使用。
在某些实施方案中,治疗期包括向个体施用药物组合物,并且治疗期被维持至少直至所述水平下降至参考值以下,在间隔期期间暂停施用,并且只要所述水平低于参考值就一直维持间隔期。所述参考值可选自(a)在于所述施用之前从所述个体获得的最新血样中测量的调节性T细胞或髓源性抑制细胞的系统性存在水平或活性水平;或者(b)患有CNS的疾病、障碍、病症或损伤的个体群特征性的调节性T细胞或髓源性抑制细胞的系统性存在水平或活性水平。
或者,治疗期包括向个体施用药物组合物并且治疗期被维持至少直至IFNγ产生白细胞的系统性存在或水平或者响应于刺激的白细胞增殖速率上升至参考值以上,在间隔期期间暂停施用,并且只要所述水平高于所述参考值就一直维持间隔期,其中所述参考值选自(a)在于所述施用之前从所述个体获得的最新血样中测量的IFNγ产生白细胞的系统性存在水平或活性水平或者响应于刺激的白细胞增殖速率;或者(b)患有CNS的疾病、障碍、病症或损伤的个体群特征性的IFNγ产生白细胞的系统性存在水平或活性水平或者响应于刺激的白细胞增殖速率。
治疗期和间隔期的时长可由医生在临床试验中针对特定患者群来确定,并且随后可一致地适用于该患者群而无需根据个人监测免疫抑制的水平。
在某些实施方案中,治疗期可时长为3天至4周,例如时长1周至4周。
在某些实施方案中,间隔期可为1周至6个月,例如时长2周至6个月,特别是时长3周至6个月。
在治疗期中,药物组合物的施用可以是重复施用,例如药物组合物可每日或者每2、3、4、5或6天一次地,每周一次地,每两周一次地,每三周一次地或者每4周一次地施用。这些频率适用于任何活性剂、可基于本领域中的常用实践并且可最终由医生在临床试验中确定。或者,可根据活性剂的性质来调整治疗期中重复施用的频率,其中例如小分子可每日施用;抗体可每3天一次地施用;而copolymer 1可每周、每两周一次地、每三周一次地或每四周一次地施用。应理解,当在治疗期期间以相对低的频率(例如在1个月的治疗期期间每周一次或者在六个月的治疗期期间每月一次)施用诸如copolymer 1的药剂时,在该治疗期之后是非治疗间隔期,其时长长于治疗期期间的重复施用之间的时间(即,在该实例中分别长于1周或1个月)。该实例中治疗期期间的施用之间的一周或一个月暂停不视为间隔期。
可相对于施用频率来调整治疗期和间隔期的时长使得例如每3天一次地施用活性剂的频率可产生6天或9天的治疗期和相应开始的间隔期。
作为预定一般性治疗方案的替代方案,可如下针对所治疗的每个患者将免疫抑制水平校正至期望的水平(个性化医疗):个别地监测Treg细胞的水平或活性(或者IFN-γ产生白细胞的水平或活性,或者响应于刺激的白细胞增殖速率)并如由监测结果所确定的凭经验且个性地调整治疗期、施用频率和间隔期。
因此,可如下确定治疗期的时长:(a)通过测量在所述施用之后的预定时间段内从个体获得的血样中的水平来监测该个体中调节性T细胞的系统性存在水平或活性水平;(b)将(a)中测量的水平与上述参考值进行比较并确定该水平是否与参考值不同;(c)基于(a)中测量的所述水平与所述参考值的关系来决定是通过重复施用继续治疗期还是通过制止重复施用来开始接下来的间隔期;以及(d)根据(c)中的决定来重复施用或开始接下来的间隔期。或者,可监测IFN-γ产生白细胞的水平或响应于刺激的白细胞增殖速率并与上述合适的参考值进行比较。
类似地,可如下确定间隔期的时长:(a)通过测量在所述施用之后的预定时间段内从个体获得的血样中的水平来监测该个体中调节性T细胞的系统性存在水平或活性水平;(b)将(a)中测量的水平与上述参考值进行比较并确定该水平是否与参考值不同;(c)基于(a)中测量的所述水平与所述参考值的关系来决定是通过重复施用以及步骤(a)和(b)来开始新的治疗过程还是通过仅重复骤(a)和(b)来延长间隔期;以及(d)根据(c)中的决定来重复施用以及步骤(a)和(b)或者仅重复步骤(a)和(b)。或者,可监测IFN-γ产生白细胞的水平或响应于刺激的白细胞增殖速率并与上述合适的参考值进行比较。
在任何情况下,均将给药方案(即治疗期和间隔期的时长)校正成使得免疫抑制水平的降低(例如,如通过个体中调节性T细胞的系统性存在水平或活性水平的降低所测量的)是短暂的。
在某些实施方案中,所述预定时间段,即在最近施用活性剂和监测步骤之间经过的时间为2天至6个月。
在某些实施方案中,所监测的调节性T细胞是选自以下的CD4+细胞:表达CD25、CD127、GITR、CTLA-4或PD-1中一种或更多种的FoxP3+细胞;或者表达CD25、CD127、GITR、CTLA-4或PD-1表面分子中一种或更多种的FoxP3-细胞。特别地,常见的调节性T细胞表型是CD4+CD25+FoxP3+细胞或CD4+CD25+FoxP3-细胞。
能够降低调节性T细胞的水平的药剂在本领域中是已知的(Colombo和Piconese,2007)并且这些药剂可根据本发明使用。下文所引用的出版物各自通过引用并入,如同在本文中完全公开一样。
因此,所述药剂可选自但并非必需限于:(i)诸如以下的抗体:(a)抗PD-1、(b)抗PD-L1、(c)抗PD-L2(Coyne和Gulley,2014;Duraiswamy等,2014;Zeng等,2013)、(d)抗CTLA-4(Simpson等,2013;Terme等,2012)、(e)与干扰素α组合的抗PD-1(Terawaki等,2011)、(f)与抗CTLA4组合的抗PD-1、(g)抗CD47(Tseng等,2013)、(h)抗OX40(Voo等,2013)、(i)抗VEGF-A(贝伐单抗)(Terme等,2013)、(j)抗CD25(Zhou等,2013)、(k)抗GITR(触发GITR的mAb(DTA-1))(Colombo和Piconese,2007)、(l)抗CCR4、(m)抗TIM-3/半乳凝集素9(Ju等,2014)、(n)抗-杀伤细胞免疫球蛋白样受体(killer-cell immunoglobulin-like receptor,KIR)、(o)抗LAG-3;或(p)抗4-1BB;(ii)(a)至(p)的任意组合;(iii)与佐剂组合的(a)至(p)的任意组合,例如与抗OX40抗体和TLR9配体(例如CpG)组合的抗CTLA-4抗体(Marabelle等,2013);(iv)选自以下的小分子:(a)p300抑制剂(Liu等,2013),例如吉西他滨(低剂量)(Shevchenko等,2013),或者C646或其类似物,即式I的化合物:
其中
R1选自H、-CO2R6、-CONR6R7、-SO3H或-SO2NR6R7;
R2选自H、-CO2R6或卤素,优选Cl;
R3选自卤素(优选F)、-NO2、-CN、-CO2R6(优选CO2CH3或CO2CH2CH3)或-CH2OH;
R4和R5各自独立地是H或-C1-C6烷基,优选甲基;
R6是H或-C1-C6烷基,优选H、甲基或乙基;并且
R7是H或-C1-C6烷基,优选H或甲基[参见(Bowers等,2010)];
(b)舒尼替尼(Terme等,2012);(c)多金属氧酸盐-1(Polyoxometalate-1,POM-1)(Ghiringhelli等,2012);(d)α,β-亚甲基腺苷5’–二磷酸(α,β-methyleneadenosine 5’-diphosphate,APCP)(Ghiringhelli等,2012);(e)三氧化二砷(As2O3)(Thomas-Schoemann等,2012);(f)GX15-070(Obatoclax)(Kim等,2014);(g)视黄酸拮抗剂,例如Ro 41-5253(一种合成的类视黄醇和选择性小分子拮抗剂)(Galvin等,2013)或LE-135(Bai等,2009);(h)SIRPα(CD47)拮抗剂,例如CV1-hIgG4(SIRPα变体),其作为单一药剂或与抗CD47抗体组合(Weiskopf等,2013);(i)CCR4拮抗剂,例如AF399/420/18025,其作为单一药剂或与抗CCR4抗体组合(Pere等,2011);(j)腺苷A2B受体拮抗剂,例如PSB603(Nakatsukasa等,2011);(k)吲哚胺-2,3-加双氧酶(indoleamine-2,3-dioxygenase,IDO)的拮抗剂;或(l)HIF-1调节剂;(v)选自以下的蛋白质:(a)印楝叶糖蛋白(Neem leaf glycoprotein,NLGP)(Roy等,2013);或(b)sCTLA-4(CTLA-4的可溶性同工型);(vi)沉默分子,例如miR-126反义分子(Qin等,2013)和抗-半乳凝集素-1(galectin-1,Gal-1)(Dalotto-Moreno等,2013);(vii)OK-432(酿脓链球菌(Streptococcus pyogenes)的冻干制剂)(Hirayama等,2013);(viii)IL-12和抗CTLA-4的组合;(ix)Copolymer 1或者调节Treg活性或水平的共聚物;(x)抗生素剂,例如万古霉素(Brestoff和Artis,2013;Smith等,2013);或者(xi)(i)至(x)的任意组合。
在某些实施方案中,所述药剂是抗PD-1抗体,即对PD-1具有特异性的抗体。
很多抗PD-1抗体在本领域中是已知的。例如,根据本发明使用的抗PD-1抗体可选自Ohaegbulam等(Ohaegbulam等,2015)(其全部内容均在此通过引用并入本文)中公开的那些,即CT-011(pidilizumab;人源化IgG1;Curetech)、MK-3475(lambrolizumab、派姆单抗(pembrolizumab);人源化IgG4;Merck)、BMS-936558(纳武单抗(nivolumab);人IgG4;Bristol-Myers Squibb)、AMP-224(PD-L2IgG2a融合蛋白;AstraZeneca)、BMS-936559(人IgG4;Bristol-Myers Squibb)、MEDI4736(人源化IgG;AstraZeneca)、MPDL3280A(人IgG;Genentech)、MSB0010718C(人IgG1;Merck-Serono),或者根据本发明使用的抗体可以是MEDI0680(AMP-514;AstraZeneca),一种人源化IgG4mAb。
在某些实施方案中,CT-011抗体可以以0.2mg/kg至6mg/kg或以1.5mg/kg至6mg/kg的剂量施用于人;MK-3475抗体可以以1mg/kg至10mg/kg的剂量施用于人;BMS-936558可以以0.3mg/kg至20mg/kg、0.3mg/kg至10mg/kg、1mg/kg至10mg/kg的剂量或者以1mg/kg或3mg/kg的剂量施用于人;BMS-936559可以以0.3mg/kg至10mg/kg的剂量施用于人;MPDL3280A可以以1mg/kg至20mg/kg的剂量施用于人;MEDI4736可以以0.1mg/kg至15mg/kg的剂量施用于人;MSB0010718C可以以1mg/kg至20mg/kg的剂量施用于人。
抗CTLA4抗体可以是Tremelimumab(Pfizer),一种全人IgG2单克隆抗体;或易普利姆玛(ipilimumab),一种全人人IgG1单克隆抗体。
抗-杀伤细胞免疫球蛋白样受体(KIR)抗体可以是Lirilumab(BMS-986015;由Innate Pharma开发并特许给Bristol-Myers Squibb),一种全人单克隆抗体。
抗LAG-3抗体针对淋巴细胞活化基因-3。可根据本发明使用的一种这样的抗体是单克隆抗体BMS-986016(派姆单抗);人源化IgG4;Merck)。
抗4-1BB抗体可以是靶向4-1BB的PF-05082566(Pfizer Oncology),一种全人源化IgG2激动剂单克隆抗体;或Urelumab(BMS-663513;Bristol-Myers Squibb),一种全人IgG4单克隆抗体。
在某些实施方案中,可使用抗体组合,其例如但不限于与利妥昔单抗(商品名Rituxan、MabThera和Zytux)(一种针对CD20蛋白的嵌合单克隆抗体)组合的CT-011,例如各自为3mg/kg;与易普利姆玛(例如3mg/kg)组合的BMS-936558(例如1mg/kg);或者与HLA-A*0201限制性多肽疫苗组合的BMS-936558(例如1mg/kg至10mg/kg)(Weber等,2013)。
表1*
*基于Bowers等(2010)
在某些实施方案中,所述药剂是p300抑制剂,其式列于表1中,即C646(4-(4-((5-(4,5-二甲基-2-硝基苯基)呋喃-2-基)亚甲基)-3-甲基-5-氧代-4,5-二氢-1H-吡唑-1-基)苯甲酸)、C146(4-羟基-3-(((2-(3-碘苯基)苯并[d]唑-5-基)亚氨基)甲基)苯甲酸)或C375(2-氯-4-(5-((2,4-二氧代-3-(2-氧代-2-(对甲苯基氨基)乙基)噻唑烷-5-亚基)甲基)呋喃-2-基)苯甲酸)。特别地,p300抑制剂是C646。
在某些实施方案中,吲哚胺-2,3-双加氧酶通路的小分子抑制剂可以是Indoximod(NLG-9189;NewLink Genetics)、INCB024360(Incyte)或NLG-919(NewLink Genetics)。
HIF-1调节剂可以是Zheng等(Zheng等,2015)中所述的M30,即5-[N-甲基-N-炔丙基氨基甲基]-8-羟基喹啉。
在某些实施方案中,所述药剂可来源于靶向革兰氏阳性细菌和革兰氏阴性细菌并且从而有利于Treg的免疫调节的广谱抗生素,例如靶向革兰氏阳性细菌并且已显示降低Treg水平/活性的万古霉素(Brestoff和Artis,2013;Smith等,2013)。
在某些实施方案中,所述药剂可以是在某一方案中将导致Treg下调的任何共聚物,例如YFAK、VYAK、VWAK、VEAK、FEAK、FAK、VAK或WAK。本文中使用的术语“Cop-1”和“Copolymer 1”可互换使用。
本发明的药物组合物可包含调节Treg活性或水平的无规共聚物作为活性剂,所述无规聚合物包含适量的带正电荷氨基酸(例如赖氨酸或精氨酸),其与带负电荷氨基酸(优选较少量)(例如谷氨酸或天冬氨酸)组合、任选地与不带电荷的中性氨基酸(例如丙氨酸或甘氨酸)(充当填料)并且任选地与适于赋予共聚物免疫原性的氨基酸(例如芳族氨基酸,如酪氨酸或色氨酸)组合。这样的组合物可包括WO 00/05250中公开的任意那些共聚物,其全部内容均在此通过引用并入本文。
更特别地,用于本发明的组合物包含选自由无规共聚物组成的组的至少一种共聚物,所述无规共聚物含有选自以下至少三组中每一组的一个氨基酸:(a)赖氨酸和精氨酸;(b)谷氨酸和天冬氨酸;(c)丙氨酸和甘氨酸;以及(d)酪氨酸和色氨酸。
用于本发明的共聚物可由L-氨基酸或D-氨基酸或者其混合物构成。本领域技术人员知晓,L-氨基酸存在于大多数天然蛋白质中。然而,D-氨基酸可商购获得并且可替换用于制备用于本发明的三元共聚物及其他共聚物的一些或全部氨基酸。本发明考虑使用包含D-和L-氨基酸二者的共聚物以及基本由L-或D-氨基酸组成的共聚物。
在某些实施方案中,本发明的药物组合物包含Copolymer 1,其是基本上由氨基酸L-谷氨酸(E)、L-丙氨酸(A)、L-酪氨酸(Y)和L-赖氨酸(K)以1.5:4.8:1:3.6的近似比组成的无规多肽的混合物、具有净总正电荷并且分子量为约2KDa至约40KDa。在某些实施方案中,Cop 1的平均分子量为约2KDa至约20KDa、约4,7KDa至约13K Da、约4KDa至约8.6KDa、约5KDa至9KDa或者约6.25KDa至8.4KDa。在另一些实施方案中,Cop 1的平均分子量为约13KDa至约20KDa、约13KDa至约16KDa或者约15KDa至约16KDa。本发明还涵盖Cop 1的低于40KDa的其他平均分子量。所述分子量范围的Copolymer 1可通过本领域中已知的方法,例如通过美国专利No.5,800,808中所述的方法来制备,其全部内容均在此通过引用整体并入本文。Copolymer 1可以是长度上包含约15至约100或者约40至约80个氨基酸的多肽。在某些实施方案中,Cop 1为以通用名称醋酸格拉替雷为人知晓的其醋酸盐的形式,其已以商品名(Teva Pharmaceuticals Ltd.,Petach Tikva,Israel的商标)在数个国家被批准用于治疗多发性硬化(MS)。如果进行以下一种或更多种替换,预期Copolymer 1对本文所公开药物组合物的活性仍得以保留:天冬氨酸替换谷氨酸、甘氨酸替换丙氨酸、精氨酸替换赖氨酸和色氨酸替换酪氨酸。
在本发明的某些实施方案中,调节Treg活性或水平的共聚物是三种不同氨基酸的共聚物,所述不同氨基酸各自来自于组(a)至(d)中三个组的不同组。本文中将这些共聚物称为三元共聚物。
在一个实施方案中,调节Treg活性或水平的共聚物是包含酪氨酸、丙氨酸和赖氨酸的三元共聚物(在下文命名为YAK),其中所述氨基酸的平均摩尔分数可改变:酪氨酸可以以约0.05至0.250的摩尔分数存在、丙氨酸以约0.3至0.6的摩尔分数存在;并且赖氨酸以约0.1至0.5的摩尔分数存在。酪氨酸、丙氨酸和赖氨酸的摩尔比可分别为约0.10:0.54:0.35。用精氨酸替换赖氨酸、甘氨酸替换丙氨酸和/或色氨酸替换酪氨酸是可能的。
在某些实施方案中,调节Treg活性或水平的共聚物是包含酪氨酸、谷氨酸和赖氨酸的三元共聚物(在下文命名为YEK),其中所述氨基酸的平均摩尔分数可改变:谷氨酸可以以约0.005至0.300的摩尔分数存在、酪氨酸可以以约0.005至0.250的摩尔分数存在并且赖氨酸可以以约0.3至0.7的摩尔分数存在。谷氨酸、酪氨酸和赖氨酸的摩尔比可分别为约0.26:0.16:0.58。用天冬氨酸替换谷氨酸、精氨酸替换赖氨酸和/或色氨酸替换酪氨酸是可能的。
在某些实施方案中,调节Treg活性或水平的共聚物是包含赖氨酸、谷氨酸和丙氨酸的三元共聚物(在下文命名为KEA),其中所述氨基酸的平均摩尔分数可改变:谷氨酸可以以约0.005至0.300的摩尔分数存在、丙酸以约0.005至0.600的摩尔分数存在并且赖氨酸以约0.2至0.7的摩尔分数存在。谷氨酸、丙氨酸和赖氨酸的摩尔比可分别为约0.15:0.48:0.36。用天冬氨酸替换谷氨酸、甘氨酸替换丙氨酸和/或精氨酸替换赖氨酸是可能的。
在某些实施方案中,调节Treg活性或水平的共聚物是包含酪氨酸、谷氨酸和丙氨酸的三元共聚物(在下文命名为YEA),其中所述氨基酸的平均摩尔分数可改变:酪氨酸可以以约0.005至0.250的摩尔分数存在、谷氨酸可以以约0.005至0.300的摩尔分数存在并且丙氨酸以约0.005至0.800的摩尔分数存在。谷氨酸、丙氨酸和酪氨酸的摩尔比可分别为约0.21:0.65:0.14。用色氨酸替换酪氨酸、天冬氨酸替换谷氨酸和/或甘氨酸替换丙氨酸是可能的。
三元共聚物YAK、YEK、KEA和YEA的平均分子量变化范围可为约2KDa至40KDa,优选约3KDa至35KDa,更优选约5KDa至25KD不等。
Copolymer 1及调节Treg活性或水平的其他共聚物可通过本领域中已知的方法来制备,例如在缩合条件下在溶液中使用期望摩尔比的氨基酸或者通过固相合成操作。缩合条件包括用于使一个氨基酸的羧基与另一氨基酸的氨基缩合形成肽键的合适的温度、pH和溶剂条件。可使用缩合剂(例如二环己基碳二亚胺)来促进肽键的形成。可使用封闭基团来保护官能团(例如侧链部分和一些氨基或羧基)免受不期望的副反应。
例如,可通过美国专利3,849,550中公开的方法来制备共聚物,其中在环境温度下(20℃至26℃)在无水二氧六环中以二乙胺作为引发剂使酪氨酸、丙氨酸、谷氨酸γ-苄酯和Nε-三氟乙酰基-赖氨酸的N-羧酸酐聚合。谷氨酸的γ-羧基可通过溴化氢在冰醋酸中去封闭。三氟乙酰基通过1M哌啶来从赖氨酸中脱除。本领域技术人员容易理解,可通过选择性地消除涉及谷氨酸、丙氨酸、酪氨酸或赖氨酸中任一个的反应来对所述方法进行调整以制备包含期望氨基酸(即,Copolymer 1中四种氨基酸中的三种)的肽和多肽。
共聚物的分子量可在多肽合成期间或者在共聚物已制备之后进行调节。为了在多肽合成期间调节分子量,对合成条件或氨基酸量进行调整使得当多肽达到期望的近似长度时合成停止。在合成之后,可通过任何可利用的尺寸选择操作(例如,在分子量筛分柱或凝胶上对多肽进行色谱)并收集期望的分子量范围来获得具有期望分子量的多肽。还可例如通过酸或酶促水解来使共聚物部分水解以消除高分子量物类,然后纯化以除去酸或酶。
在一个实施方案中,可通过包括以下的方法来制备具有期望分子量的共聚物:使受保护的多肽与氢溴酸反应以形成具有期望分子量谱的三氟乙酰基-多肽。在通过一个或更多个测试反应预先确定的时间和温度下进行该反应。在测试反应期间,改变时间和温度并确定给定受试多肽批次的分子量范围。将对所述多肽批次提供最佳分子量范围的测试条件用于该批次。因此,可通过包括以下的方法来合成具有期望分子量谱的三氟乙酰基-多肽:在通过测试反应预先确定的时间和温度下使受保护的多肽与氢溴酸反应。然后,用哌啶水溶液进一步处理具有期望分子量谱的三氟乙酰基-多肽以形成具有期望分子量的低毒性多肽。
在某些实施方案中,使来自给定批次的受保护多肽的受试样品与氢溴酸在约20℃至28℃的温度下反应约10小时至50小时。用于该批次的最佳条件通过进行数个测试反应来确定。例如,在一个实施方案中,使受保护的多肽与氢溴酸在约26℃的温度下反应约17小时。
由于Cop 1与MS相关性HLA-DR分子的结合基序是已知的(Fridkis-Hareli等,1999),因此可容易地制备具有限定序列的多肽并测试与HLA-DR分子的肽结合槽的结合,如Fridkis-Hareli等(1999)出版物中所述的。这样的肽的实例是WO 00/05249和WO 00/05250(其全部内容均在此通过引用并入本文)中公开的那些并且包括SEQ ID NO.1-32的肽(表2)。
预期这样的肽及其他类似肽与Cop1具有类似的活性。这样的肽及其他类似肽还被视为在与CNS髓鞘抗原交叉反应的共聚物的定义之内并且认为其使用是本发明的一部分。
表2
根据本发明的“调节Treg活性或水平的共聚物”的定义意指涵盖其他的合成氨基酸共聚物,例如Fridkis-Hareli等,2002和美国专利8,017,125所述的无规四氨基酸共聚物(作为用于治疗多发性硬化的候选物),即以下共聚物:包含氨基酸缬氨酸(V)、苯丙氨酸(F)、丙氨酸(A)和赖氨酸(K)的VFAK;包含氨基酸缬氨酸(V)、酪氨酸(Y)、丙氨酸(A)和赖氨酸(K)的VYAK;包含氨基酸缬氨酸(V)、色氨酸(W)、丙氨酸(A)和赖氨酸(K)的VWAK;包含氨基酸缬氨酸(V)、谷氨酸(E)、丙氨酸(A)和赖氨酸(K)的VEAK;包含氨基酸苯丙氨酸(F)、谷氨酸(E)、丙氨酸(A)和赖氨酸(K)的FEAK;包含氨基酸苯丙氨酸(F)、丙氨酸(A)和赖氨酸(K)的FAK;包含氨基酸缬氨酸(V)、丙氨酸(A)和赖氨酸(K)的VAK;以及包含氨基酸色氨酸(W)、丙氨酸(A)和赖氨酸(K)的WAK。
根据本发明的药物组合物可用于治疗为选自以下的神经变性疾病、障碍或病症的CNS疾病、障碍或病症:阿尔茨海默病(Alzheimer’s disease);肌萎缩侧索硬化;帕金森病(Parkinson’s disease);亨廷顿病(Huntington’s disease);原发性进行性多发性硬化;继发性进行性多发性硬化;皮质基底节变性(corticobasal degeneration);雷特综合征(Rett syndrome);选自年龄相关性黄斑变性和视网膜色素变性的视网膜变性病症;前部缺血性视神经病变;青光眼;葡萄膜炎;抑郁;创伤相关性应激或创伤后应激障碍;额颞痴呆;路易体痴呆(Lewy body dementias);轻度认知受损;后大脑皮质萎缩;原发性进行性失语或进行性核上性麻痹。在某些实施方案中,所述CNS病症是年龄相关性痴呆。
在某些实施方案中,所述CNS病症是阿尔茨海默病、肌萎缩侧索硬化、帕金森病、亨廷顿病。
根据本发明的药物组合物还可用于治疗选自以下的CNS损伤:脊髓损伤、闭合性头部损伤、钝伤、穿透性创伤、出血性卒中、缺血性卒中、脑缺血、视神经损伤、心肌梗死、有机磷中毒和由肿瘤切除引起的损伤。
如上所述,发明人已发现,本发明改善模拟阿尔茨海默病的小鼠中的认知功能。因此,所述药物组合物可用于改善CNS运动和/或认知功能,例如用于减轻可发生于无诊断疾病的个体中以及患有神经变性疾病的人中的年龄相关性认知功能丧失。此外,所述药物组合物可用于减轻由急性应激或创伤性事件引起的认知功能丧失。上文中提及的认知功能可包括学习、记忆或二者。
本文中使用的术语“CNS功能”尤其指接收和处理感觉信息、思考、学习、记忆、感觉、生成和理解语言、控制运动功能以及听觉和视觉响应、维持平衡和均衡、运动协调、传导感觉信息和控制诸如呼吸、心率和消化的自主功能。
术语“认知”、“认知功能”和“认知表现”在本文中可互换使用并且与涉及但不限于以下的任何智力过程或状况相关:学习、记忆、意象生成、思考、意识、推理、空间能力、说话和语言技能、语言获取和判断注意能力。认知形成于多个脑区域,例如海马、皮质及其他脑结构中。然而,认为长期记忆至少部分地储存于皮质中,并且已知感觉信息通过特定的皮质结构(存在于岛叶皮质中的味觉皮质)来获取、巩固和检索。
在人中,认知功能可通过任何已知的方法来测定,所述方法例如但不限于通过临床总体印象变化量表(CIBIC-补充量表);简易智力状况检查(Mini Mental State Exam,MMSE);神经精神症状问卷(Neuropsychiatric Inventory,NPI);临床痴呆评定量表(Clinical Dementia Rating Scale,CDR);剑桥自动化成套神经心理测试(Cambridge Neuropsychological Test Automated Battery,CANTAB)或Sandoz老年临床评估(Sandoz Clinical Assessment-Geriatric,SCAG)。还可使用诸如以下成像技术来间接测量认知功能:正电子发射断层显像(Positron Emission Tomography,PET)、功能性磁共振成像(functional magnetic resonance imaging,fMRI)、单光子发射计算机断层显像(Single Photon Emission Computed Tomography,SPECT)或允许测量脑功能的任何其他成像技术。
患者中影响认知的一个或更多个过程的改善将意味着所述患者中认知功能的改善,因此在某些实施方案中,改善认知包括改善学习、可塑性和/或长期记忆。术语“改善”和“增强”在本文中可互换使用。
术语“学习”涉及获取或获得新的知识、行为、技能、价值或偏好,或者改变和增强现有的知识、行为、技能、价值或偏好。
术语“可塑性”涉及与脑随学习而改变和改变已获取记忆的能力相关的突触可塑性、脑可塑性或神经可塑性。反映可塑性的一个可测量参数是记忆消退。
术语“记忆”涉及其中编码、储存和检索信息的过程。记忆具有三种可区别的类别:感觉记忆、短期记忆和长期记忆。
术语“长期记忆”是长时间或无限时间地保持信息的能力。长期记忆包括两个主要部分:外显记忆(陈述性记忆)和内隐记忆(非陈述性记忆)。长期记忆通过记忆巩固来实现,记忆巩固是在记忆痕迹初始获取之后使其稳定的一类过程。巩固区分为两个特定过程:突触巩固,其发生于学习之后的前几个小时;和系统巩固,其中海马依赖性记忆历经数周至数年的时间变得不依赖于海马。
在另一个方面,本发明涉及用于治疗不包括自身免疫性神经炎性疾病—复发-缓解型多发性硬化(RRMS)的中枢神经系统(CNS)的疾病、障碍、病症或损伤的方法,所述方法包括向有此需要的个体施用如上限定的根据本发明的药物组合物,其中所述药物组合物通过包括至少两个治疗过程的给药方案来施用,每个治疗过程顺序地包括治疗期,之后是间隔期。
在某些实施方案中,治疗期包括向个体施用药物组合物并且治疗期被维持至少直至所述水平下降到参考值以下,在间隔期期间暂停施用,并且只要所述水平低于参考值就一直维持间隔期。所述参考值可选自(a)在于所述施用之前从所述个体获得的最新血样中测量的调节性T细胞或髓源性抑制细胞的系统性存在水平或活性水平;或者(b)患有CNS的疾病、障碍、病症或损伤的个体群特征性的调节性T细胞或髓源性抑制细胞的系统性存在水平或活性水平。
或者,治疗期包括向个体施用药物组合物并且治疗期被维持至少直至IFNγ产生白细胞的系统性存在或水平或者响应于刺激的白细胞增殖速率上升至参考值以上,在间隔期期间暂停施用,并且只要所述水平高于所述参考值就一直维持间隔期,其中所述参考值选自(a)在于所述施用之前从所述个体获得的最新血样中测量的IFNγ产生白细胞的系统性存在水平或活性水平或者响应于刺激的白细胞增殖速率;或者(b)患有CNS的疾病、障碍、病症或损伤的个体群特征性的IFNγ产生白细胞的系统性存在水平或活性水平或者响应于刺激的白细胞增殖速率。
描述本发明药物组合物的不同特征的上述实施方案也与本发明的方法相关,因为所述方法采用相同的药物组合物。
在又一个方面,本发明提供了用于治疗不包括自身免疫性神经炎性疾病—RRMS的CNS的疾病、障碍、病症或损伤的药物组合物,所述药物组合物包含引起个体中系统性免疫抑制的水平降低的活性剂,所述活性剂选自:(i)对(a)CD47、(b)OX40、(c)VEGF-A(贝伐单抗)、(d)CD25、(e)GITR(触发GITR的mAb(DTA-1))、(f)CCR4或(g)TIM-3/半乳凝集素9具有特异性的抗体;(h)抗杀伤细胞免疫球蛋白样受体、(i)抗LAG-3或(j)抗4-1BB;(ii)(a)至(j)的任意组合;(iii)与佐剂组合的(a)至(j)的任意组合,所述佐剂例如TLR9配体,例如CpG;(iv)选自以下的蛋白质:(a)印楝叶糖蛋白(NLGP)或(b)sCTLA-4;(v)选自以下的小分子:(a)舒尼替尼;(b)多金属氧酸盐-1(POM-1);(c)α,β-亚甲基腺苷5’-二磷酸(APCP);(d)三氧化二砷(As2O3);(e)GX15-070(Obatoclax);(f)视黄酸拮抗剂,例如Ro 41-5253或LE-135;(g)SIRPα(CD47)拮抗剂,例如CV1-hIgG4,其作为单一药剂或与抗CD47抗体组合;(h)CCR4拮抗剂,例如AF399/420/18025,其作为单一药剂或与抗CCR4抗体组合;或者(i)腺苷A2B受体拮抗剂,例如PSB603;(j)吲哚胺-2,3-加双氧酶的拮抗剂;(k)HIF-1调节剂;(vi)沉默分子,例如miR-126反义分子和抗半乳凝集素-1(Gal-1);(vii)OK-432;(viii)IL-12和抗CTLA-4的组合;(ix)抗生素剂,例如万古霉素;或者(x)(i)至(ix)的任意组合。
根据本发明使用的药物组合物可使用一种或更多种生理学上可接受的载体或赋形剂以常规方式配制。所述载体就与组合物的其他成分相容并且对其赋形剂无害而言必需是“可接受的”。
列举了载体、施用模式、剂型等的以下实例作为从其可选择用于本发明的载体、施用模式、剂型等的已知可能性。然而,本领域技术人员将了解,所选的任何给定制剂和施用模式均应首先进行测试以确定其获得期望的结果。
施用方法包括但不限于胃肠外,例如静脉内、腹膜内、肌内、皮下、经粘膜(例如,经口、鼻内、经颊、经阴道、经直肠、眼内)、鞘内、局部和皮内途径。施用可以是系统性的或局部的。
术语“载体”指与活性剂一起施用的稀释剂、佐剂、赋形剂或载剂。药物组合物中的载体可包含粘合剂,例如微晶纤维素、聚乙烯吡咯烷酮(聚维酮或聚乙烯吡咯酮)、黄蓍胶、明胶、淀粉、乳糖或乳糖单水合物;崩解剂,例如褐藻酸、玉米淀粉等;润滑剂或表面活性剂,例如硬脂酸镁或月桂基硫酸钠;以及助流剂,例如胶体二氧化硅。
对于经口施用,所述药物制剂可以是液体形式,例如溶液、糖浆或混悬剂;或者可提供为在使用之前与水或其他合适载剂重构的药物产品。这样的液体制剂可使用诸如以下药学可接受的添加剂通过常规方式来制备:助悬剂(例如,山梨糖醇糖浆、纤维素衍生物或氢化可食用脂肪)、乳化剂(例如,卵磷脂或阿拉伯胶)、非水性载剂(例如,杏仁油、油性脂或分级植物油)和防腐剂(例如,对-羟基苯甲酸甲酯或对-羟基苯甲酸丙酯或者山梨酸)。所述药物组合物可采用例如片剂或胶囊剂,其使用诸如以下药学可接受的赋形剂通过常规方式制备:粘合剂(例如,预胶化玉米淀粉、聚乙烯吡咯烷酮或羟丙基甲基纤维素)、填料(例如,乳糖、微晶纤维素或磷酸氢钙)、润滑剂(例如,硬脂酸镁、滑石或二氧化硅)、崩解剂(例如,马铃薯淀粉或羟乙酸淀粉钠)或润湿剂(例如月桂基硫酸钠)。可通过本领域中已知的方法对片剂进行包被。
用于经口施用的制剂可适当地配制成提供活性化合物的受控释放。
对于经颊施用,所述组合物可采用以常规方式配制的片剂或锭剂的形式。
所述组合物可配制成用于通过注射,例如通过推注或连续输注来肠胃外施用。注射用制剂可以以单位剂形,例如在已添加有防腐剂的安瓿或多剂量容器中提供。所述组合物可采用诸如在油性或水性载剂中的混悬剂、溶液剂或乳剂的形式,并且可包含配制剂,例如助悬剂、稳定剂和/或分散剂。或者,活性成分可以是在使用之前与合适载剂(例如,无菌的无热原的水)重构的粉末形式。
所述组合物还可配制成经直肠组合物,例如栓剂或保留型灌肠剂,例如其含有常规的栓剂基质,例如可可油或其他甘油酯。
对于通过吸入的施用,使用合适的抛射剂由加压包装或喷雾器以气雾喷雾剂的形式来方便地递送根据本发明使用的组合物,所述合适的抛射剂例如二氯二氟甲烷、三氯氟甲烷、二氯四氟乙烷、二氧化碳或其他合适的气体。在加压气雾剂的情况下,可通过设置递送计量量的阀来决定剂量单位。用于吸入器或吹入器中的胶囊剂或药筒(例如明胶的胶囊剂或药筒)可配制成包含化合物和合适粉末基质(例如乳糖或淀粉)的粉末混合物。
用于人使用的活性成分剂量的确定基于本领域中的常用实践,并且最终将由医生在临床试验中确定。用于施用于人的预期大约当量剂量可使用已知的公式基于下文中公开的体内实验证据来计算(例如,Reagan-Show等(2007)Dose translation from animal to human studies revisited.The FASEB Journal 22:659-661)。根据该范例,成人的当量剂量(mg/kg体重)等于给予小鼠的剂量(mg/kg体重)乘以0.081。
现在将通过以下非限制性实施例对本发明进行举例说明。
实施例
材料和方法
动物.5XFAD转基因小鼠(Tg6799)和AD双转基因B6.Cg-Tg(APPswe,PSEN1dE9)85Dbo/J小鼠(Borchelt等,1997)购买自杰克逊实验室(The Jackson Laboratory),所述5XFAD转基因小鼠在神经元特异性小鼠Thy-1启动子(Oakley等,2006)的转录控制之下共-过表达人APP(Swedish型突变:K670N/M671L;佛罗里达型突变:I716V;和伦敦型突变:V717I)和PS1(M146L/L286V)转基因的家族性AD突变体形式。基因分型如先前所述(Oakley等,2006)通过尾DNA的PCR分析来进行。使用杂合突变体cx3cr1GFP/+小鼠(Jung等,2000)(B6.129P-cx3cr1tm1Litt/J,其中CX3CR1趋化因子受体等位基因之一被替换成编码GFP的基因)作为BM嵌合体的供体。使Foxp3.LuciDTR小鼠(Suffner等,2010)与5XFAD小鼠交配以能够实现Foxp3+Treg的条件性消耗。通过魏茨曼科学研究所的动物繁育中心(Animal Breeding Center of the Weizmann Institute of Science)来繁育和维持动物。本文中详述的所有实验均遵守由魏茨曼科学研究所的机构动物照管及使用委员会制定(Institutional Animal Care and Use Committee,IACUC)的规则。
RNA纯化、cDNA合成和定量实时PCR分析.海马齿状回(DG)的总RNA用TRI试剂(Molecular Research Center)提取并使用RNeasy试剂盒(Qiagen)从裂解液纯化。脉络丛的总DNA使用RNA MicroPrep试剂盒(Zymo Research)来提取。使用高容量cDNA逆转录试剂盒(Applied Biosystems)来将mRNA(1μg)转化为cDNA。特定mRNA的表达使用基于荧光的定量实时PCR(real-time PCR,RT-qPCR)来测定。RT-qPCR反应使用Fast-SYBR PCR Master Mix(Applied Biosystems)来进行。量化反应使用标准曲线方法对每份样品以一式三份方式进行。选择肽酰脯氨酰异构酶A(peptidylprolyl isomerase A,ppia)作为参照(管家)基因。扩增循环为95℃5秒、60℃20秒和72℃15秒。在测定结束时,构建熔解曲线以评价反应的特异性。对于ifn-γ和ppia基因分析,根据生产商的方案(PreAmp Master Mix Kit;Applied Biosystems),用非随机PCR引物预扩增cDNA 14个PCR循环,从而提高后续实时PCR分析的灵敏度。mRNA表达使用TaqMan RT-qPCR根据生产商的说明(Applied Biosystems)来确定。所有的RT-qPCR均使用StepOne软件V2.2.2(Applied Biosystems)来进行并分析。使用下述TaqMan Assays-on-DemandTM探针:Mm02342430_g1(ppia)和Mm01168134_m1(ifn-γ)。
对于所有其他的检测基因,使用以下引物:
ppia正向5’-AGCATACAGGTCCTGGCATCTTGT-3'(SEQ ID NO:33)和反向5'-CAAAGACCACATGCTTGCCATCCA-3’(SEQ ID NO:34);
icam1正向5’-AGATCACATTCACGGTGCTGGCTA-3’(SEQ ID NO:35)和反向5’-AGCTTTGGGATGGTAGCTGGAAGA-3’(SEQ ID NO:36);
vcam1正向5’-TGTGAAGGGATTAACGAGGCTGGA-3’(SEQ ID NO:37)和反向5’-CCATGTTTCGGGCACATTTCCACA-3’(SEQ ID NO:38);
cxcl10正向5'-AACTGCATCCATATCGATGAC-3'(SEQ ID NO:39)和反向5'-GTGGCAATGATCTCAACAC-3'(SEQ ID NO:40);
ccl2正向5’-CATCCACGTGTTGGCTCA-3’(SEQ ID NO:41)和反向5'-GATCATCTTGCTGGTGAATGAGT-3’(SEQ ID NO:42);
tnf-γ正向5’-GCCTCTTCTCATTCCTGCTT-3’(SEQ ID NO:43)和反向CTCCTCCACTTGGTGGTTTG-3’(SEQ ID NO:44);
il-1β正向5’-CCAAAAGATGAAGGGCTGCTT-3’(SEQ ID NO:45)和反向5’-TGCTGCTGCGAGATTTGAAG-3’(SEQ ID NO:46);
il-12p40正向5’-GAAGTTCAACATCAAGAGCA-3’(SEQ ID NO:47)和反向5’-CATAGTCCCTTTGGTCCAG-3’(SEQ ID NO:48);
il-10正向5’-TGAATTCCCTGGGTGAGAAGCTGA-3’(SEQ ID NO:49)和反向5’-TGGCCTTGTAGACACCTTGGTCTT-3’(SEQ ID NO:50);
tgfβ2正向5’-AATTGCTGCCTTCGCCCTCTTTAC-3’(SEQ ID NO:51)和反向5’-TGTACAGGCTGAGGACTTTGGTGT-3’(SEQ ID NO:52);
igf-1正向5’-CCGGACCAGAGACCCTTTG(SEQ ID NO:53)和反向5’-CCTGTGGGCTTGTTGAAGTAAAA-3’(SEQ ID NO:54);
bdnf正向5’-GATGCTCAGCAGTCAAGTGCCTTT-3’(SEQ ID NO:55)和反向5’-GACATGTTTGCGGCATCCAGGTAA-3’(SEQ ID NO:56)。
免疫组织化学.对石蜡包埋的经切片小鼠脑(6μm厚)和人脑(10μm厚)进行组织处理和免疫组织化学。对于人ICAM-1染色,使用第一小鼠抗ICAM(1:20Abcam;ab2213)抗体。将载玻片用3%H2O2温育10分钟,并使用第二生物素缀合的抗小鼠抗体,然后用Vectastain ABC试剂盒(Vector Laboratories)进行生物素/抗生物素扩增。随后,施加3,3’-二氨基联苯胺(DAB底物)(Zytomed试剂盒);将玻片脱水并用基于二甲苯的封固溶液封固。对于组织染色,在组织切除和固定之前,经心脏向小鼠灌注PBS。在解剖显微镜下(Stemi DV4;Zeiss)从脑的侧脑室、第三脑室和第四脑室分离CP组织。对于全标本CP染色(whole mount CP staining),将组织用2.5%多聚甲醛(PFA)在4℃下固定1小时,随后转移至含有0.05%叠氮化钠的PBS。在染色之前,将解剖的组织用PBS洗涤并在室温下封闭(20%马血清、0.3%Triton X-100和PBS)1小时。在室温下进行第一抗体(在含有2%马血清和0.3%Triton X-100的PBS中)或第二抗体的全标本染色1小时。在每个步骤之后在PBS中进行三次洗涤。将组织施加至载玻片,用Immu-mount(9990402,来自于Thermo Scientific)封固并用盖玻片密封。对于经切片脑的染色,如先前所述(Baruch等,2013;Kunis等,2013)采用两种不同的组织制备方案(石蜡包埋的切片或用切片机切成的自由浮动切片)。使用以下第一抗体:小鼠抗-Aβ(1:300,Covance,#SIG-39320);兔抗-GFP(1:100,MBL,#598);大鼠抗-CD68(1:300,eBioscience,#14-0681);大鼠抗-ICAM-1(1:200,Abcam,#AB2213);山羊抗-GFP(1:100,Abcam,#ab6658);兔抗-IBA-1(1:300,Wako,#019-19741);山羊抗-IL-10(1:20,R&D systems,#AF519);大鼠抗-Foxp3(1:20,eBioscience,#13-5773-80);兔抗-CD3(1:500,Dako,#IS503);小鼠抗-ZO-1、小鼠抗-E-Cahedrin和兔抗-Claudin-1(全部均为1:100,Invitrogen,#33-9100,#33-4000,#51-9000);兔抗-GFAP(1:200,Dako,#Z0334)。第二抗体包括Cy2/Cy3/Cy5-缀合的驴抗-小鼠/山羊/兔/大鼠抗体(1:200;全部均来自于Jackson Immunoresearch)。对载玻片进行Hoechst核染色(1:4000;Invitrogen Probes)1分钟。在免疫染色操作中常规使用两个阴性对照:用同种型对照抗体染色接着用第二抗体染色;或者仅用第二抗体染色。对于Foxp3细胞内染色,使用Retreivagen试剂盒(#550524,#550527;BD PharmingenTM)来进行经石蜡包埋载玻片的抗原修复。使用荧光显微镜(E800;Nikon)或激光扫描共聚焦显微镜(Carl Zeiss,Inc.)来进行显微分析。荧光显微镜装备有数码像机(DXM 1200F;Nikon)并且装备有20×NA 0.50物镜或40×NA0.75物镜(Plan Fluor;Nikon)。共聚焦显微镜装备有LSM 510激光扫描能力(三种激光:Ar 488、HeNe 543和HeNe 633)。使用采集软件(NIS-Elements,F3[Nikon]或LSM[Carl Zeiss,Inc.])来对固定后的组织进行记录。对于染色强度的量化,如先前所述(Burgess等,2010),使用ImageJ软件(NIH)来测量总细胞染色和背景染色并计算特异性染色的强度。将图像使用Photoshop CS6 13.0(Adobe)剪裁、合并和优化,并使用Illustrator CS5 15.1(Adobe)进行排列。
人CP的经石蜡包埋切片.年轻和年长的死后非CNS患病个体以及AD患者的人脑切片在正当同意和伦理委员会批准(TW220)之下从Oxford Brain Bank(之前称为Thomas Willis Oxford Brain Collection(TWOBC))获得。涉及这些切片的实验均经魏茨曼科学研究所生物伦理委员会(Weizmann Institute of Science Bioethics Committee)批准。
流式细胞术、样品制备和分析.经心脏向小鼠灌注PBS并如先前所述(Baruch等,2013)处理组织。在PBS中在解剖显微镜下(Stemi DV4;Zeiss)将脑解剖并取出不同的脑区域,使用gentleMACSTM解离器(Miltenyi Biotec)解离组织。从脑的侧脑室、第三脑室和第四脑室分离脉络丛组织,在含有400U/ml IV型胶原酶(Worthington Biochemical Corporation)的PBS(具有Ca2+/Mg2+)中在37℃下孵育45分钟,然后通过吹吸人工均化。将脾用注射器的柱塞捣碎并用ACK(氯化铵钾)裂解缓冲液处理以除去红细胞。在所有情况下,均根据生产商的方法来染色样品。将所有的样品通过70μm尼龙网过滤并用抗Fc CD16/32(1:100;BD Biosciences)封闭。对于IFN-γ的细胞内染色,将细胞与对甲氧基安非他明(10ng/ml;Sigma-Aldrich)和离子霉素(250ng/ml;Sigma-Aldrich)一起孵育6小时,添加布雷菲德菌素-A(10μg/ml;Sigma-Aldrich)用于最后4小时。用BD Cytofix/CytopermTM Plus固定/透化试剂盒(目录号555028)来进行细胞因子的细胞内标记。对于Treg染色,使用eBioscience FoxP3染色缓冲液套装(目录号00-5523-00)。以下经荧光染料标记的单克隆抗体购买自BD Pharmingen,BioLegend,R&D Systems或eBiosciences,并且根据生产商的方案使用:PE或Alexa Fluor 450缀合的抗-CD4;PE缀合的抗-CD25;PerCP-Cy5.5-缀合的抗-CD45;FITC-缀合的抗-TCRβ;APC-缀合的抗-IFN-γ;APC-缀合的抗-FoxP3;亮紫缀合的抗-CD45。使用FlowJo软件在LSRII细胞计数器(BD Biosciences)上对细胞进行分析。在每个实验中,使用相关的阴性对照组、阳性对照和每个组织的单染色样品来鉴定目的群体并排除其他群体。
BM嵌合体的制备.BM嵌合体如先前所述(Shechter等,2009;Shechter等,2013)来制备。简言之,使性别匹配的受体小鼠在防护头部的同时经受致死性全身照射(950拉德)(Shechter等,2009)。然后,向小鼠静脉内注射来自CX3CR1GFP/+供体的5×106个BM细胞。在BM移植之后,使小鼠静置8周至10周以使得能够重建造血谱系,之后将其用于实验。通过对血样进行FACS分析根据GFP表达细胞在循环单细胞(CD11b+)中的百分比来确定嵌合性百分比。在这一防护头部的模型中,实现平均60%嵌合性,并且证实CNS浸润性GFP+骨髓细胞是CD45高/CD11b高,表示单核细胞源性巨噬细胞而非小胶质细胞(Shechter等,2013)。
Morris水迷宫.每天对小鼠进行三次试验,持续连续4天以学习发现池(直径为1.1m)中位于水表面1.5cm下的隐藏平台。使水温保持在21℃至22℃。用奶粉使水变成不透明的。在测试室内,小鼠仅可利用远端的视觉形状和物体线索来帮助定位浸没的平台。记录逃避潜伏期(即,发现并爬上平台所需的时间),持续长达60秒。允许每只小鼠在平台上停留15秒,然后从迷宫移至其养笼中。如果小鼠在60秒内未发现平台,则人工将其放置在平台上并在15秒后返回至其养笼中。每只小鼠的试验间间隔时间为10分钟。在第5天,移出平台,并在无法逃避下对小鼠进行单独实验,持续60秒。在第6天和第7天,将平台放置在与原始训练象限相对的象限内,并每天重新训练小鼠三次。使用EthoVision V7.1自动化追踪系统(Noldus Information Technology)来记录数据。使用方差分析(ANOVA)和Bonferroni事后检验来进行统计学分析。所有的MWM测试均在关灯阶段期间在10a.m.和5p.m.期间进行。
放射臂水迷宫.使用放射臂水迷宫(RAWM)来测试空间学习和记忆,如先前详细描述的(Alamed等,2006)。简言之,将六个不锈钢插入物放置在箱中,形成从敞开中心区域发散的六个游泳臂。将逃避平台放置在直径为1.1m的池中水平面1.5cm下一个臂(目标臂)的端部。使水温保持在21℃至22℃。用奶粉使水变成不透明的。在测试室内,小鼠仅可利用远端的视觉形状和物体线索来帮助定位浸没的平台。对于给定的小鼠,目标臂的位置保持不变。在第1天,对小鼠训练15次试验(历经3小时),其中试验在可见平台和隐藏平台之间交替进行,最后4次试验仅用隐藏平台进行。在第2天,用隐藏平台对小鼠训练15次试验。将进入不正确臂或者在15秒内未能选择臂记录为错误。通过计数小鼠在每次试验中的臂进入错误次数和逃避潜伏期来测量空间学习和记忆。训练数据分析为连续三次试验对训练块的平均错误数或逃避潜伏期。
GA施用.向每只小鼠皮下(s.c.)注射溶解于200μl PBS中的100μg总剂量的GA(批号P53640;Teva Pharmaceutical Industries,Petah Tiqva,Israel)。根据每周-GA方案(Butovsky等,2006)或每日-GA施用来注射小鼠(图8和图16)。根据每个实验的指示,在最后一次GA注射后1周或者在处理之后1个月对小鼠实施安乐死。
Treg的条件性消融.每天向Foxp3.LuciDTR小鼠(Suffner等,2010)腹膜内(i.p.)注射白喉毒素(DTx;8ng/g体重;Sigma),持续连续4天。通过对血液和脾中的免疫细胞进行流式细胞术分析来确定DTx的效力,实现GFP表达FoxP3+CD4+Treg细胞的几乎完全(>99%)消耗(图4)。
P300抑制.小鼠中p300的抑制类似于先前所述的来进行(Liu等,2013)。将p300i(C646;Tocris Bioscience)溶解于DMSO中并每天i.p.注射(8.9mg kg-1d-1,i.p.),持续1周。类似地,向载剂处理的小鼠注射DMSO。
ATRA处理.向小鼠的全反式视黄酸(all-trans retinoic acid,ATRA)施用类似于先前所述的来进行(Walsh等,2014)。将ATRA(Sigma)溶解于DMSO中并每隔一天地i.p.注射(8mg kg-1d-1),历经1周的疗程。类似地,向载剂处理的小鼠注射DMSO。
可溶性Aβ(sAβ)蛋白的分离和量化.组织均化和sAβ蛋白提取如先前所述来进行(Schmidt等,2005)。简言之,将大脑脑实质解剖,急速冷冻并保存在-80℃直至均化。顺序地从样品提取蛋白质以获得包含溶解度不同的蛋白质的分离级分。将样品在Dounce均化器中使用研磨用玻璃杵在10体积的冰冷组织均化缓冲液中进行均化,所述缓冲液包含250mM蔗糖、20mM Tris碱、1mM乙二胺四乙酸(EDTA)和1mM乙二醇四乙酸(pH7.4)。在六个冲程之后,将均浆与100-mM NaCl溶液中的0.4%二乙胺(DEA)1:1混合,之后再进行六个冲程,然后在4℃下以135,000g离心45分钟。收集上清液(含有细胞外蛋白质和细胞溶质蛋白质的DEA可溶性级分)并用10%的0.5M Tris-HCl(pH 6.8)中和。通过酶联免疫吸附测定(ELISA)使用可商购获得的试剂盒(Biolegend;分别为#SIG-38954和#SIG-38956)根据生产商的说明从可溶性级分单独测量Aβ1-40和Aβ1-42。
Aβ斑量化.从每个脑收集6μm的冠状切片,并对每只小鼠来自贯穿目的区域(齿状回或脉络丛)的4个不同预定深度的8个切片进行免疫染色。使用Image-Pro Plus软件(Media Cybernetics,Bethesda,MD,USA)来进行阳性染色像素的基于直方图的分割。将分割算法人工地应用于齿状回区域中或皮层V中的每个图像,并确定总Aβ免疫染色所占据面积的百分比。由相同的6μm冠状脑切片来量化斑数目并表示为每个脑区域的平均斑数目。在量化之前,对切片进行编码以掩盖实验组的身份,并通过对组身份盲态的观察者来量化斑负荷。
统计学分析.用于分析每个实验组的特定测试在图例中进行了说明。在两个组之间进行比较使用双尾学生t检验来分析数据,单向ANOVA用于比较数个组,接着在拒绝无效假设(P<0.05)之后进行Newman–Keuls事后程序以对组进行逐对比较。行为测试的数据使用双向重复测量ANOVA来进行分析,并使用Bonferroni事后程序来进行后续逐对比较。基于文献和过往经历来选择具有充分统计学效力的样品尺寸并根据年龄、性别和基因型将小鼠分配至实验组。研究者对实验期间的组身份和结果评估呈盲态。所有的入选标准和排除标准都根据IACUC指导方针预先确立。结果表示为平均值±s.e.m。在图表中,y轴误差棒表示s.e.m。统计学计算使用GraphPad Prism软件(GraphPad Software,San Diego,CA)来进行。
简介.阿尔茨海默病(AD)是一种年龄相关性神经变性疾病,其以中枢神经系统(CNS)中的神经元损伤、淀粉样蛋白β(Aβ)斑形成和慢性炎症为特征,导致认知功能逐渐丧失和脑组织破坏(Akiyama等,2000;Hardy和Selkoe,2002)。在这些病症下,CNS的循环骨髓细胞和驻留骨髓细胞、小胶质细胞在减缓神经炎性应答方面起非冗余作用(Britschgi和Wyss-Coray,2007;Cameron和Landreth,2010;Lai和McLaurin,2012)。特别地,当小胶质细胞未能最终清除Aβ沉积物,则CNS浸润性单核细胞源性巨噬细胞(mo-MΦ)在限制Aβ斑形成和抵抗AD样病理方面发挥有益作用(Butovsky等,2007;Koronyo-Hamaoui等,2009;Mildner等,2011;Simard等,2006;Town等,2008)。已鉴定上皮层形成血液-CSF-屏障(blood-CSF-barrier,BCSFB)的脑脉络丛(CP)为白细胞进入CNS使得能够在神经组织损伤之后募集mo-MΦ和T细胞的选择性门(Kunis等,2013;Shechter等,2013)。在此,我们假设:在AD中,消炎免疫细胞向患病实质的次最佳募集是涉及CP门功能障碍的系统性免疫失效的结果。
实施例1.小鼠AD模型中随疾病进程的脉络丛(CP)门活性
我们首先对AD的5XFAD转基因小鼠模型(AD-Tg)中随疾病进程的CP活性进行了检测;这些小鼠共表达与家族性AD相关的五种突变并且在早至2月龄时发生脑Aβ病理和神经胶质增生(Oakley等,2006)。我们发现:随着疾病病理的进展期,与年龄匹配的野生型(WT)对照相比,AD-Tg小鼠的CP表达显著更低水平的白细胞归巢和运输决定簇,包括icam1、vcam1、cxcl10和ccl2(图1A),所述决定簇显示响应于急性CNS损伤而被CP上调并且是白细胞经上皮迁移所需要的(Kunis等,2013;Shechter等,2013)。对整联蛋白配体ICAM-1进行免疫组织化学染色确定其经由AD-Tg小鼠CP上皮的降低表达(图1b)。另外,对人死后脑中的ICAM-1进行染色显示其在CP上皮中的年龄相关性降低,这与我们之前的观察结果一致(Baruch等,2014),并且对该效应进行量化评估揭示与未患CNS疾病的年长个体相比在AD患者中进一步降低(图2A)。由于CP对白细胞运输决定簇的诱导依赖于上皮干扰素(IFN)-γ信号传导(Kunis等,2013),因此我们接下来测试了所观察到的效应是否可反映CP处丧失IFN-γ可用性。使用流式细胞术细胞内染色对5XFAD AD-Tg小鼠的CP进行检测揭示该隔室中具有显著更低数目的IFN-γ产生细胞(图2B),并且定量实时PCR(RT-qPCR)分析确定与年龄匹配的WT对照相比,AD-Tg小鼠CP处ifn-γ的mRNA表达水平较低(图2C)。
实施例2.Treg介导的系统性免疫抑制、CP门活性和AD病理之间的功能关系
调节性T细胞(Treg)在抑制系统性效应免疫应答中起关键作用(Sakaguchi等,2008)。我们设想Treg介导的系统性免疫抑制影响CP处的IFN-γ可用性,并且因此集中于Treg在AD病理中的参与。与AD患者中Treg水平和抑制活性升高的先前报道一致(Rosenkranz等,2007;Saresella等,2010;Torres等,2013),相对于其年龄匹配的WT同窝出生小鼠来评价5XFAD AD-Tg小鼠脾细胞中的Foxp3+Treg频率揭示其水平随疾病进程升高(图3A、3B)。为了研究Treg介导的系统性免疫抑制、CP门活性和AD病理之间的功能关系,我们使5XFAD AD-Tg小鼠与Foxp3-白喉毒素受体(DTR+)小鼠杂交,使得通过施用白喉毒素(DTx)来实现AD-Tg/DTR+小鼠中Foxp3+Treg的短暂条件性体内消耗(图4A)。Treg的短暂消耗导致相对于经DTx处理的AD-Tg/DTR-同窝出生小鼠,AD-Tg/DTR+小鼠中CP对白细胞运输分子的mRNA表达升高(图5A)。分析短暂Treg消耗对脑实质的长期作用(3周后)揭示脑中的免疫细胞累积,包括代表浸润性mo-MΦ的CD45高/CD11b高骨髓细胞(Shechter等,2013)和CD4+T细胞的数量升高(图5B)。另外,Treg的短期暂时消耗导致聚集在脑中的CD4+T细胞中Foxp3+Treg显著富集,如通过流式细胞术所评估的(图5C、5D)。对海马的RT-qPCR分析显示foxp3和il10mRNA的表达增加(图5E)。
接下来,我们研究了短期Treg消耗、之后免疫调节细胞在脑病理部位聚集是否对脑功能产生长期效果。我们观察到海马神经胶质增生减轻(图5F)并且促炎细胞因子(例如il-12p40和tnf-α)的mRNA表达水平降低(图5G)。此外,5XFAD AD-Tg小鼠中表现出稳健Aβ斑病理的两个脑区域海马齿状回和大脑皮质(第5层)(Oakley等,2006)中的大脑Aβ斑负荷减小(图6A、6B)。使用Morris水迷宫(MWM)测试来评价对认知功能的作用揭示:相对于经DTx处理的AD-Tg/DTR-年龄匹配小鼠,在Treg消耗之后AD-Tg/DTR+小鼠中的空间学习和记忆得到显著改善,达到与WT小鼠近似的表现(图6C至6E)。总而言之,这些数据表明在AD-Tg小鼠中短暂地破坏Treg介导的系统性免疫抑制导致消炎细胞(包括mo-MΦ和Treg)在脑中聚集,并且在其之后神经炎性应答消退、Aβ清除且认知下降逆转。
实施例3.每周施用Copolymer-1降低Treg介导的系统性免疫抑制、提高CP门活性并且减轻AD病理。
为了进一步证实系统性免疫抑制、CP功能和AD病理之间的相反关系的因果性质,我们接下来使用了免疫调节性化合物醋酸格拉替雷(GA;还称为Copolymer-1或在每周施用方案中发现其在AD的APP/PS1小鼠模型(Butovsky等,2006)中具有治疗效果;该效果与mo-MΦ向大脑疾病病理部位的募集功能性相关(Butovsky等,2007)。在此,我们首先检验了:是否与我们在5XFAD AD-Tg小鼠中的观察结果类似,APP/PS1AD-Tg小鼠中的CP也在IFN-γ表达水平方面具有缺陷。我们发现:相对于年龄匹配的WT对照,在APP/PS1AD-Tg小鼠中CP处的IFN-γ水平降低(图7A)。这些结果激励我们测试每周-GA在APP/PS1小鼠中的治疗效果(Butovsky等,2006)是否可在5XFAD AD-Tg中重现,并且倘若如此,则其是否会影响系统性Treg和CP对mo-MΦ运输的活化。因此,我们用GA的每周施用方案来处理5XFAD AD-Tg小鼠,历经4周的时间(在下文为“每周-GA”,示意性地描绘于图8A中)。我们发现,经每周-GA处理的5XFAD AD-Tg小鼠表现出减轻的神经炎症(图8B至8D)和改善的认知表现,这在处理之后持续长达2个月(图8E至8I)。通过流式细胞术来检测每周-GA对系统性免疫和CP的作用,我们发现在经处理5XFAD AD-Tg小鼠中脾细胞Foxp3+Treg水平降低(图9A)并且CP处的IFN-γ产生细胞增加,达到与在WT对照中所观察到的那些的近似水平(图9B)。在经每周-GA处理的小鼠中,CP处IFN-γ表达细胞的水平升高伴随着白细胞运输分子的上皮表达上调(图9C)。
为了检测浸润性mo-MΦ向CNS的进入,我们使用了5XFAD AD-Tg/CX3CR1GFP/+骨髓(BM)嵌合小鼠(利用头部保护来制备),从而使循环(绿色荧光蛋白(GFP)+标记的)骨髓细胞能够可视化(Shechter等,2009;Shechter等,2013)。我们发现:与经载剂处理的AD-Tg/CX3CR1GFP/+对照相比,在每周-GA处理之后GFP+mo-MΦ向CP以及向邻近脑室间隙的归巢增加(图9D至9E)。脑实质的免疫组织化学揭示,在脑斑形成部位存在GFP+mo-MΦ聚集(图9F),并且通过对AD-Tg非嵌合小鼠中的海马进行流式细胞术分析来量化浸润性骨髓细胞显示,CD11b高CD45高表达细胞的数目增加(图9G、9H)。总而言之,这些结果证实了mo-MΦ向AD病理部位募集、系统性Treg水平降低和IFN-γ依赖性CP活化之间的功能联系。
实施例4.短期地直接干预Treg活性提高CP门活性并且减轻AD病理。
4.1使用小分子组蛋白乙酰转移酶抑制剂的Treg活性干预。表明Treg介导的系统性免疫抑制干预抵抗AD病理的能力的上述发现使人想到Treg在癌症免疫治疗中所贡献的功能,其中这些细胞阻碍免疫系统引发有效的抗肿瘤应答的能力(Bos和Rudensky,2012;Nishikawa和Sakaguchi,2010)。因此,我们考虑,直接干预Foxp3+Treg细胞活性的处理在AD方面可能是有利的。我们测试了p300i(C646(Bowers等,2010)),即p300的非肽抑制剂,p300是一种调节Treg功能的组蛋白乙酰转移酶(Liu等,2013);该抑制剂显示影响Treg的抑制活性而同时使保护性T效应细胞应答不受干扰(Liu等,2013)。我们发现:与经载剂(DMSO)处理的对照相比,经p300i处理的小鼠显示脾(图10A)以及CP(图10B)中系统性IFN-γ表达细胞的水平升高。接下来,我们用p300i或载剂处理AD-Tg小鼠,历经1周的疗程,并在3周后检测其大脑Aβ斑负荷。免疫组织化学分析揭示,在经p300i处理的AD-Tg小鼠中大脑Aβ斑载荷显著减小(图10C至10E)。我们还测试了一个处理疗程之后对斑病理的效应是否能够持续超过3周,并且倘若如此,额外的处理疗程是否有助于持久效应。因此,我们将接受单个p300i处理疗程并在2个月后检测的AD-Tg小鼠与在此期间接受两个处理疗程(之间具有1个月的间隔时间)的年龄匹配小组进行了比较(示意性地描绘于图10F中)。我们发现,脑斑的减小甚至在单个处理疗程之后两个月仍明显,但是在接受两个处理疗程(之间具有1个月的间隔时间)的小鼠中更强(图10G)。由于AD中的突触可塑性和记忆受损与可溶性Aβ1-40/Aβ1-42(sAβ)水平的大脑水平升高有关(Shankar等,2008),因此我们还测量了在单个和重复周期的p300i处理之后的sAβ水平。同样地,我们发现1个疗程和两个疗程(之间具有1个月的间隔之间)二者均有效地降低大脑sAβ,但是就对sAβ1-42的效应而言,在重复疗程之后,该效应较为更强(图10H)。这些结果表明,尽管单个的短期处理疗程有效,但是重复的治疗疗程将有利于维持持久的治疗效果,这与我们在每周-GA处理之后的观察结果类似。
4.2.使用抗PD1抗体的Treg活性干预。在10月龄时,在实验的第1天和第4天,向5XFAD阿尔茨海默病(AD)转基因(Tg)小鼠i.p.注射250μg的抗PD1抗体(RMP1-14;#BE0146;Bioxcell Lifesciences Pvt.LTD.)或对照IgG抗体(IgG2a;#BE0089;Bioxcell Lifesciences Pvt.LTD.),并在3周后(示意性地描绘于图11A中)通过放射臂水迷宫(RAWM)空间学习和记忆任务来检测其认知表现,如先前所详细描述的(Alamed等,2006)。简言之,在RAWM任务的第1天,对小鼠训练15次试验(历时3小时),其中试验在可见平台和隐藏平台之间交替进行并且最后4次试验仅用隐藏平台进行。在第2天,用隐藏平台对小鼠训练15次试验。将进入到不正确臂中或者在15秒内未能选择臂评分为错误。通过计数小鼠在每次试验中的臂进入错误次数和逃避潜伏期来测量空间学习和记忆。使用年龄匹配的未经处理WT和AD-Tg小鼠作为对照。我们发现:如在3周后所评估的,处理1个治疗期的5XFAD AD-Tg小鼠显示在RAWM中的空间认知表现显著改善(图11B),所述1个治疗期包括两次抗PD1注射(第1天和第4天)。
接下来,我们检验了对疾病病理的效果是否与系统性免疫抑制的降低有关。我们重复了上述实验,但这次在处理期结束时对小鼠进行检测(实验的第7天;示意性地描绘于图12A中)。我们观察到:在该时间点,经PD-1处理的AD-Tg小鼠中系统性免疫抑制的减弱伴随着IFN-γ产生CD4脾细胞升高的系统性效应(图12B),其与CP处IFN-γmRNA水平升高(图13A)和CP白细胞运输分子(趋化因子CCL2和CXCL10)的表达升高(图13B)的局部效应相关。这些数据表明,在AD-Tg小鼠中的短时期抗PD-1处理与如所预期的减弱Treg介导之免疫抑制的系统性应答(Naidoo等,2014)以及白细胞向CNS运输之CP门活性的活化相关。
最后,我们检验了对AD-Tg小鼠中疾病病理的作用以及额外的处理期是否有利于其对病理的作用。为此,如上所述使10个月大的AD-Tg小鼠接受1个时期的抗PD-1处理,或者间隔3周再接受处理。将对照组用IgG进行处理或者保持不处理,并在3周之后测试所有组小鼠的认知表现(示意性地描绘于图14A中)。我们发现,尽管处理1个抗-PD-1时期(“AD-Tg+PD-1X1”)并且在2个月后检测的AD-Tg小鼠相比较于经IgG处理和未经处理的AD-Tg小鼠表现出显著的认知改善,但是与提前一个月评估认知表现的相同小鼠相比,该效果较不稳健。相比之下,与接受1个时期的AD-Tg小鼠相比以及与经IgG处理和未经处理的AD-Tg小鼠相比,接受第二时期抗PD-1处理(“AD-Tg+PD-1X2”)的AD-Tg小鼠在RAWM中表现出显著更佳的空间学习和记忆能力(图14B)。这些发现揭示:为了维持持久的治疗效果,需要重复的时期。
4.3使用抗PD1抗体和抗CTLA4抗体的组合的Treg活性干预
在10月龄时,在实验的第1天和第4天向5XFAD阿尔茨海默病(AD)转基因(Tg)小鼠i.p.注射250μg抗PD1抗体(RMP1-14;#BE0146;Bioxcell Lifesciences Pvt.LTD.)和250μg抗CTLA4抗体(InVivoMAb抗-mCD152;#BE0131;Bioxcell Lifesciences Pvt.LTD.)或者对照IgG抗体(IgG2a,#BE0089或Polyclonal Syrian Hamster IgG,#BE0087;Bioxcell Lifesciences Pvt.LTD.),并在3周后通过放射臂水迷宫(RAWM)空间学习和记忆任务来检测其认知表现,如上所述。
使一些小鼠以3周的间隔期再接受一个处理期。将对照组用IgG进行处理或者不作处理,并且在3周后测试所有组小鼠的认知表现。
预期,与经IgG处理和未经处理的AD-Tg小鼠相比,经抗体组合处理的小鼠表现出显著的认知改善以及显著的脑斑载荷减小。
实施例5.Treg活性增强对AD病理具有不利影响。
为了确定Treg介导的系统性免疫抑制在AD中的不利影响,我们接下来研究了增强系统性Treg水平是否可对AD病理具有相反的不利影响。为了对此进行测试,我们通过施用全反式视黄酸(ATRA)来增强AD-Tg小鼠中的Treg抑制功能,全反式视黄酸诱导Treg分化(Mucida等,2007),稳定Treg表型(Zhou等,2010)并且使Treg更具抑制性(Zhou等,2010)。我们使用了处于疾病进程的相对早期阶段的5XFAD AD-Tg小鼠,并且用ATRA或载剂(DMSO)对其进行处理。经ATRA处理的AD-Tg小鼠显示出显著更高的脾细胞Foxp3+CD25+Treg频率(图15A、15B)。在最后一次ATRA注射后3周对小鼠进行检测揭示大脑Aβ斑负荷和神经胶质增生较高(约增加2倍至3倍;图15C至15E),并且评估sAβ揭示在增强系统性Treg之后大脑sAβ1-40和sAβ1-42水平升高(图15F至15G)。使用RAWM来评估认知表现显示:相对于经载剂处理的AD-Tg小鼠,在经ATRA处理的AD-Tg小鼠中空间记忆缺损加重(图15H)。
鉴于我们在系统性Treg对AD病理的不利影响方面的目前发现,综合已知每日GA施用诱导Treg并且在临床上用于治疗多发性硬化(multiple sclerosis,MS)的事实(Haas等,2009;Hong等,2005;Weber等,2007),我们测试了每日GA方案(在1个月的时期内)相比较于每周-GA是否可能对AD-Tg小鼠的疾病病理具有不利影响。我们比较了每日-GA施用相对于每周-GA施用(示意性地描绘于图16A中)在5XFAD AD-Tg小鼠中的效果。通过RAWM任务来评估认知表现揭示,相比较于每周-GA处理的有益效果,在接受每日-GA的AD-Tg小鼠中,或者未观察到对空间记忆的有益作用,或者未观察到效应恶化的倾向(图16B)。另外,不同于每周-GA施用对斑清除的稳健作用,经每日-GA处理的AD-Tg小鼠未表现出任何有益效应或者表现出对斑载荷的适度不利作用(图16C至16F)。这些发现突出了,用每日-GA进行相同的免疫调节治疗可如何相对且独特地影响着与神经炎症相关的两种CNS病理MS和AD(Schwartz和Baruch,2014a)。
实施例6.直接干预Treg活性提高CP门活性并且防止或减轻PTSD病理。
严重应激条件或慢性应激可导致创伤后应激障碍(posttraumatic stress disorder,PTSD)和抑郁。我们之前表明CP门活性对处理精神性应激可至关重要,并且在CP的次最佳功能的情况下,精神创伤性事件可导致PTSD(Schwartz和Baruch,2012)。我们还假设,在创伤后进行有助于调节CP应答的及时系统性干预可防止PTSD的慢性病症的发生。我们发现短期地减弱Treg介导的系统性免疫抑制对脑病理具有长期作用,所述发现启示在创伤性事件之后立即进行这种干预可防止PTSD发生。
为了测试我们的工作假设:CP参与处理创伤性应激并且在创伤性应激导致发生PTSD的情况下其可能功能障碍,我们采用了生理性PTSD样动物模型,在该模型中小鼠表现出高度警戒行为、注意力缺损、风险评估增加和睡眠不良(Lebow等,2012)。在PTSD诱发的这一实验模型中,使小鼠习惯颠倒的昼/夜循环10天,施以两个电击事件(创伤和触发)(称为“PTSD诱发”)并在创伤之后的不同时间点进行评价。在创伤性事件之后,向小鼠注射所述化合物,其短暂地降低外周免疫抑制。根据以下一种或更多种方案来对小鼠进行处理:
·在创伤性事件后的第1天和第4天向小鼠i.p.注射250μg的抗PD1抗体(RMP1-14;#BE0146;Bioxcell Lifesciences Pvt.LTD.)或对照IgG抗体(IgG2a;#BE0089;Bioxcell Lifesciences Pvt.LTD.),并在另外的两周间隔期之后进行检测;
·在实验的第1天和第4天向小鼠i.p.注射250μg抗PD1抗体(RMP1-14;#BE0146;Bioxcell Lifesciences Pvt.LTD.)和250μg抗CTLA4抗体(InVivoMAb抗-mCD152;#BE0131;Bioxcell Lifesciences Pvt.LTD.)或者对照IgG抗体(IgG2a,#BE0089或Polyclonal Syrian Hamster IgG,#BE0087;Bioxcell Lifesciences Pvt.LTD.),并在两周的间隔期之后进行检测;
·在创伤性事件之后如上所述向小鼠i.p.注射每周-GA,并在两周的间隔期之后进行检测;
·在创伤性事件之后用p300i或载剂处理小鼠,历经一周的时间,并如上所述在3周后进行检测。
使一些小鼠以适当的间隔期再接受一个处理期。
在预期之内,接受处理的小鼠在该实验模型中不表现与PTSD相关的焦虑行为,如在(Lebow等,2012)中所述的通过在昼/夜迷宫中探索和风险评估所花费的时间或其他行为任务所评估。
实施例7.短暂地降低系统性免疫抑制减轻帕金森病病理。
在这些实验中使用帕金森病(PD)转基因(Tg)小鼠。根据以下一种或更多种方案在疾病的进展期对小鼠进行处理:
·在创伤性事件后的第1天和第4天向小鼠i.p.注射250μg的抗PD1抗体(RMP1-14;#BE0146;Bioxcell Lifesciences Pvt.LTD.)或对照IgG抗体(IgG2a;#BE0089;Bioxcell Lifesciences Pvt.LTD.),并在另外的两周间隔期之后进行检测;
·在实验的第1天和第4天向小鼠i.p.注射250μg抗PD1抗体(RMP1-14;#BE0146;Bioxcell Lifesciences Pvt.LTD.)和250μg抗CTLA4抗体(InVivoMAb抗-mCD152;#BE0131;Bioxcell Lifesciences Pvt.LTD.)或者对照IgG抗体(IgG2a,#BE0089或Polyclonal Syrian Hamster IgG,#BE0087;Bioxcell Lifesciences Pvt.LTD.),并在两周的间隔期之后进行检测;
·在创伤性事件之后如上所述向小鼠i.p.注射每周-GA,并在两周的间隔期之后进行检测;
·在创伤性事件之后用p300i或载剂处理小鼠,历经一周的时间,并如上所述在3周后进行检测。
使一些小鼠以适当的间隔期(约3周至1个月)再接受一个处理期。
使用例如评估小鼠停留在旋转杆上的能力的旋转杆表现测试来评价运动神经功能。
在预期之内,与经IgG处理或经载剂处理的对照组或者未处理组相比,处理一个治疗期的PD-Tg小鼠表现出显著改善的运动表现。接受两个治疗过程并且在适当的间隔期之后检测的PD-Tg小鼠预期表现出持久的治疗效果。为了维持这一治疗效果,使小鼠接受活动的处理期,其中每个时期之间具有适当的间隔期。
实施例8.短暂地降低系统性免疫抑制减轻亨廷顿病病理。
这些实验中使用的模型可以是亨廷顿病(HD)R6/2转基因小鼠(Tg)测试系统。R6/2转基因小鼠在疾病的进展期过表达突变的人亨廷顿基因,其包括多个CAG重复片段的插入。这些小鼠从早至5周至6周龄开始表现出进行性行为-运动缺损,并且在10周至13周时导致过早死亡。症状包括体重轻、抱拢、震颤和惊厥。
在小鼠45天大时根据以下一种或更多种方案对其进行处理:
·在创伤性事件后的第1天和第4天向小鼠i.p.注射250μg的抗PD1抗体(RMP1-14;#BE0146;Bioxcell Lifesciences Pvt.LTD.)或对照IgG抗体(IgG2a;#BE0089;Bioxcell Lifesciences Pvt.LTD.),并在另外的两周间隔期之后进行检测;
·在实验的第1天和第4天向小鼠i.p.注射250μg抗PD1抗体(RMP1-14;#BE0146;Bioxcell Lifesciences Pvt.LTD.)和250μg抗CTLA4抗体(InVivoMAb抗-mCD152;#BE0131;Bioxcell Lifesciences Pvt.LTD.)或者对照IgG抗体(IgG2a,#BE0089或Polyclonal Syrian Hamster IgG,#BE0087;Bioxcell Lifesciences Pvt.LTD.),并在两周的间隔期之后进行检测;
·在创伤性事件之后如上所述向小鼠i.p.注射每周-GA,并在两周的间隔期之后进行检测;
·在创伤性事件之后用p300i或载剂处理小鼠,历经一周的时间,并如上所述在3周后进行检测。
使一些小鼠以适当的间隔期(约3周至1个月)再接受一个处理期。
使用例如评估小鼠停留在旋转杆上的能力的旋转杆表现测试来评价运动神经功能。
在预期之内,与经IgG处理或经载剂处理的对照组或者未处理组相比,处理一个治疗期的HD-Tg小鼠表现出显著改善的运动表现。接受两个治疗过程并且在适当的间隔期之后检测的HD-Tg小鼠预期表现出持久的治疗效果。为了维持这一治疗效果,使小鼠接受活动的处理期,其中每个时期之间具有适当的间隔期。
实施例9.短暂地降低系统性免疫抑制减轻肌萎缩侧索硬化病理。
该实验中使用的模型可以是过表达含有Gly93→Ala(G93A)基因的缺陷型人突变体SOD1等位基因的转基因小鼠(B6SJL-TgN(SOD1-G93A)1Gur(本文中为“ALS小鼠”)。该模型发生运动神经元疾病并且因此构成用于测试ALS的公认动物模型。
在小鼠75天大时根据以下一种或更多种方案对其进行处理:
·在创伤性事件后的第1天和第4天向小鼠i.p.注射250μg的抗PD1抗体(RMP1-14;#BE0146;Bioxcell Lifesciences Pvt.LTD.)或对照IgG抗体(IgG2a;#BE0089;Bioxcell Lifesciences Pvt.LTD.),并在另外的两周间隔期之后进行检测;
·在实验的第1天和第4天向小鼠i.p.注射250μg抗PD1抗体(RMP1-14;#BE0146;Bioxcell Lifesciences Pvt.LTD.)和250μg抗CTLA4抗体(InVivoMAb抗-mCD152;#BE0131;Bioxcell Lifesciences Pvt.LTD.)或者对照IgG抗体(IgG2a,#BE0089或Polyclonal Syrian Hamster IgG,#BE0087;Bioxcell Lifesciences Pvt.LTD.),并在两周的间隔期之后进行检测;
·在创伤性事件之后如上所述向小鼠i.p.注射每周-GA,并在两周的间隔期之后进行检测;
·在创伤性事件之后用p300i或载剂处理小鼠,历经一周的时间,并如上所述在3周后进行检测。
使一些小鼠以适当的间隔期(约3周至1个月)再接受一个处理期。
评价运动神经功能,所述评价使用例如评估小鼠停留在旋转杆上的能力的旋转杆表现测试,或者允许小鼠抓在和握在下端带有小环的垂直线(直径2mm)上。垂直线允许小鼠利用前肢和后肢二者抓住线。使线维持24rpm的垂直取向的圆周运动(圆半径为10cm)。用计时器记录小鼠能够悬挂到线上的时间。
在预期之内,与经IgG处理或经载剂处理的对照组或者未处理组相比,处理一个治疗期的ALS小鼠表现出显著改善的运动表现。接受两个治疗过程并且在适当的间隔期之后检测的ALS小鼠预期表现出持久的治疗效果。为了维持这一治疗效果,使小鼠接受活动的处理期,其中每个时期之间具有适当的间隔期。
参考文献
Akiyama H,Barger S,Barnum S,Bradt B,Bauer J,Cole GM,Cooper NR,Eikelenboom P,Emmerling M,Fiebich BL,Finch CE,Frautschy S,Griffin WS,Hampel H,Hull M,Landreth G,Lue L,Mrak R,Mackenzie IR,McGeer PL,O'Banion MK,Pachter J,Pasinetti G,Plata-Salaman C,Rogers J,Rydel R,Shen Y,Streit W,Strohmeyer R,Tooyoma I,Van Muiswinkel FL,Veerhuis R,Walker D,Webster S,Wegrzyniak B,Wenk G,Wyss-Coray T(2000)Inflammation and Alzheimer's disease.Neurobiology of aging 21:383-421
Alamed J,Wilcock DM,Diamond DM,Gordon MN,Morgan D(2006)Two-day radial-arm water maze learning and memory task;robust resolution of amyloid-related memory deficits in transgenic mice.Nature protocols 1:1671-1679
Bai A,Lu N,Guo Y,Liu Z,Chen J,Peng Z(2009)All-trans retinoic acid down-regulates inflammatory responses by shifting the Treg/Th17profile in human ulcerative and murine colitis.Journal of leukocyte biology 86:959-969
Baruch K,Deczkowska A,David E,Castellano JM,Miller O,Kertser A,Berkutzki T,Barnett-Itzhaki Z,Bezalel D,Wyss-Coray T,Amit I,Schwartz M(2014)Aging.Aging-induced type I interferon response at the choroid plexus negatively affects brain function.Science 346:89-93
Baruch K,Ron-Harel N,Gal H,Deczkowska A,Shifrut E,Ndifon W,Mirlas-Neisberg N,Cardon M,Vaknin I,Cahalon L,Berkutzki T,Mattson MP,Gomez-Pinilla F,Friedman N,Schwartz M(2013)CNS-specific immunity at the choroid plexus shifts toward destructive Th2inflammation in brain aging.Proceedings of the National Academy of Sciences of the United States of America 110:2264-2269
Borchelt DR,Ratovitski T,van Lare J,Lee MK,Gonzales V,Jenkins NA,Copeland NG,Price DL,Sisodia SS(1997)Accelerated amyloid deposition in the brains of transgenic mice coexpressing mutant presenilin 1and amyloid precursor proteins.Neuron 19:939-945
Bos PD,Rudensky AY(2012)Treg cells in cancer:a case of multiple personality disorder.Science translational medicine 4:164fs144
Bowers EM,Yan G,Mukherjee C,Orry A,Wang L,Holbert MA,Crump NT,Hazzalin CA,Liszczak G,Yuan H,Larocca C,Saldanha SA,Abagyan R,Sun Y,Meyers DJ,Marmorstein R,Mahadevan LC,Alani RM,Cole PA(2010)Virtual ligand screening of the p300/CBP histone acetyltransferase:identification of a selective small molecule inhibitor.Chemistry&biology 17:471-482
Breitner JC,Haneuse SJ,Walker R,Dublin S,Crane PK,Gray SL,Larson EB(2009)Risk of dementia and AD with prior exposure to NSAIDs in an elderly community-based cohort.Neurology 72:1899-1905
Brestoff JR,Artis D(2013)Commensal bacteria at the interface of host metabolism and the immune system.Nature immunology 14:676-684
Britschgi M,Wyss-Coray T(2007)Immune cells may fend off Alzheimer disease.Nature medicine 13:408-409
Burgess A,Vigneron S,Brioudes E,Labbe JC,Lorca T,Castro A(2010)Loss of human Greatwall results in G2 arrest and multiple mitotic defects due to deregulation of the cyclin B-Cdc2/PP2A balance.Proceedings of the National Academy of Sciences of the United States of America 107:12564-12569
Butovsky O,Koronyo-Hamaoui M,Kunis G,Ophir E,Landa G,Cohen H,Schwartz M(2006)Glatiramer acetate fights against Alzheimer's disease by inducing dendritic-like microglia expressing insulin-like growth factor 1.Proceedings of the National Academy of Sciences of the United States of America 103:11784-11789
Butovsky O,Kunis G,Koronyo-Hamaoui M,Schwartz M(2007)Selective ablation of bone marrow-derived dendritic cells increases amyloid plaques in a mouse Alzheimer's disease model.The European journal of neuroscience 26:413-416
Cameron B,Landreth GE(2010)Inflammation,microglia,and Alzheimer's disease.Neurobiology of disease 37:503-509
Chen X,Oppenheim JJ(2011)Resolving the identity myth:key markers of functional CD4+FoxP3+regulatory T cells.International immunopharmacology 11:1489-1496
Colombo MP,Piconese S(2007)Regulatory-T-cell inhibition versus depletion:the right choice in cancer immunotherapy.Nature reviews Cancer 7:880-887
Coyne GO,Gulley JL(2014)Adding fuel to the fire:Immunogenic intensification.Human vaccines&immunotherapeutics 10:3306-3312
Dalotto-Moreno T,Croci DO,Cerliani JP,Martinez-Allo VC,Dergan-Dylon S,Mendez-Huergo SP,Stupirski JC,Mazal D,Osinaga E,Toscano MA,Sundblad V,Rabinovich GA,Salatino M(2013)Targeting galectin-1 overcomes breast cancer-associated immunosuppression and prevents metastatic disease.Cancer research 73:1107-1117
Duraiswamy J,Freeman GJ,Coukos G(2014)Dual blockade of PD-1 and CTLA-4 combined with tumor vaccine effectively restores T-cell rejection function in tumors--response.Cancer research 74:633-634;discussion 635
Gabrilovich DI,Nagaraj S(2009)Myeloid-derived suppressor cells as regulators of the immune system.Nature reviews Immunology 9:162-174
Galvin KC,Dyck L,Marshall NA,Stefanska AM,Walsh KP,Moran B,Higgins SC,Dungan LS,Mills KH(2013)Blocking retinoic acid receptor-alpha enhances the efficacy of a dendritic cell vaccine against tumours by suppressing the induction of regulatory T cells.Cancer immunology,immunotherapy:CII 62:1273-1282
Ghiringhelli F,Bruchard M,Chalmin F,Rebe C(2012)Production of adenosine by ectonucleotidases:a key factor in tumor immunoescape.Journal of biomedicine&biotechnology 2012:473712
Group AR,Lyketsos CG,Breitner JC,Green RC,Martin BK,Meinert C,Piantadosi S,Sabbagh M(2007)Naproxen and celecoxib do not prevent AD in early results from a randomized controlled trial.Neurology 68:1800-1808
Haas J,Korporal M,Balint B,Fritzsching B,Schwarz A,Wildemann B(2009)Glatiramer acetate improves regulatory T-cell function by expansion of naive CD4(+)CD25(+)FOXP3(+)CD31(+)T-cells in patients with multiple sclerosis.Journal of neuroimmunology 216:113-117
Hardy J,Selkoe DJ(2002)The amyloid hypothesis of Alzheimer's disease:progress and problems on the road to therapeutics.Science 297:353-356
He F,Balling R(2013)The role of regulatory T cells in neurodegenerative diseases.Wiley interdisciplinary reviews Systems biology and medicine 5:153-180
Hirayama M,Nishikawa H,Nagata Y,Tsuji T,Kato T,Kageyama S,Ueda S,Sugiyama D,Hori S,Sakaguchi S,Ritter G,Old LJ,Gnjatic S,Shiku H(2013)Overcoming regulatory T-cell suppression by a lyophilized preparation of Streptococcus pyogenes.European journal of immunology 43:989-1000
Hong J,Li N,Zhang X,Zheng B,Zhang JZ(2005)Induction of CD4+CD25+regulatory T cells by copolymer-I through activation of transcription factor Foxp3.Proceedings of the National Academy of Sciences of the United States of America 102:6449-6454
Ju Y,Shang X,Liu Z,Zhang J,Li Y,Shen Y,Liu Y,Liu C,Liu B,Xu L,Wang Y,Zhang B,Zou J(2014)The Tim-3/galectin-9 pathway involves in the homeostasis of hepatic Tregs in a mouse model of concanavalin A-induced hepatitis.Molecular immunology 58:85-91
Jung S,Aliberti J,Graemmel P,Sunshine MJ,Kreutzberg GW,Sher A,Littman DR(2000)Analysis of fractalkine receptor CX(3)CR1 function by targeted deletion and green fluorescent protein reporter gene insertion.Molecular and cellular biology 20:4106-4114
Kim JM,Rasmussen JP,Rudensky AY(2007)Regulatory T cells prevent catastrophic autoimmunity throughout the lifespan of mice.Nature immunology8:191-197
Kim PS,Jochems C,Grenga I,Donahue RN,Tsang KY,Gulley JL,Schlom J,Farsaci B(2014)Pan-Bcl-2 inhibitor,GX15-070(obatoclax),decreases human T regulatory lymphocytes while preserving effector T lymphocytes:a rationale for its use in combination immunotherapy.Journal of immunology 192:2622-2633
Koronyo-Hamaoui M,Ko MK,Koronyo Y,Azoulay D,Seksenyan A,Kunis G,Pham M,Bakhsheshian J,Rogeri P,Black KL,Farkas DL,Schwartz M(2009)Attenuation of AD-like neuropathology by harnessing peripheral immune cells:local elevation of IL-10 and MMP-9.Journal of neurochemistry 111:1409-1424
Kotsakis A,Harasymczuk M,Schilling B,Georgoulias V,Argiris A,Whiteside TL(2012)Myeloid-derived suppressor cell measurements in fresh and cryopreserved blood samples.Journal of immunological methods 381:14-22
Kunis G,Baruch K,Rosenzweig N,Kertser A,Miller O,Berkutzki T,Schwartz M(2013)IFN-gamma-dependent activation of the brain's choroid plexus for CNS immune surveillance and repair.Brain:a journal of neurology 136:3427-3440
Lai AY,McLaurin J(2012)Clearance of amyloid-beta peptides by microglia and macrophages:the issue of what,when and where.Future neurology 7:165-176
Lebow M,Neufeld-Cohen A,Kuperman Y,Tsoory M,Gil S,Chen A (2012)Susceptibility to PTSD-like behavior is mediated by corticotropin-releasing factor receptor type 2 levels in the bed nucleus of the stria terminalis.The Journal of neuroscience:the official journal of the Society for Neuroscience 32:6906-6916
Liu Y,Wang L,Predina J,Han R,Beier UH,Wang LC,Kapoor V,Bhatti TR,Akimova T,Singhal S,Brindle PK,Cole PA,Albelda SM,Hancock WW(2013)Inhibition of p300 impairs Foxp3(+)T regulatory cell function and promotes antitumor immunity.Nature medicine 19:1173-1177
Marabelle A,Kohrt H,Sagiv-Barfi I,Ajami B,Axtell RC,Zhou G,Rajapaksa R,Green MR,Torchia J,Brody J,Luong R,Rosenblum MD,Steinman L,Levitsky HI,Tse V,Levy R(2013)Depleting tumor-specific Tregs at a single site eradicates disseminated tumors.The Journal of clinical investigation 123:2447-2463
Michaud JP,Bellavance MA,Prefontaine P,Rivest S(2013)Real-time in vivo imaging reveals the ability of monocytes to clear vascular amyloid beta.Cell reports 5:646-653
Mildner A,Schlevogt B,Kierdorf K,Bottcher C,Erny D,Kummer MP,Quinn M,Bruck W,Bechmann I,Heneka MT,Priller J,Prinz M(2011)Distinct and non-redundant roles of microglia and myeloid subsets in mouse models of Alzheimer's disease.The Journal of neuroscience:the official journal of the Society for Neuroscience 31:11159-11171
Mucida D,Park Y,Kim G,Turovskaya O,Scott I,Kronenberg M,Cheroutre H(2007)Reciprocal TH17 and regulatory T cell differentiation mediated by retinoic acid.Science 317:256-260
Naidoo J,Page DB,Wolchok JD(2014)Immune modulation for cancer therapy.British journal of cancer 111:2214-2219
Nakatsukasa H,Tsukimoto M,Harada H,Kojima S(2011)Adenosine A2B receptor antagonist suppresses differentiation to regulatory T cells without suppressing activation of T cells.Biochemical and biophysical research communications 409:114-119
Nishikawa H,Sakaguchi S(2010)Regulatory T cells in tumor immunity.International journal of cancer Journal international du cancer 127:759-767
Oakley H,Cole SL,Logan S,Maus E,Shao P,Craft J,Guillozet-Bongaarts A,Ohno M,Disterhoft J,Van Eldik L,Berry R,Vassar R(2006)Intraneuronal beta-amyloid aggregates,neurodegeneration,and neuron loss in transgenic mice with five familial Alzheimer's disease mutations:potential factors in amyloid plaque formation.The Journal of neuroscience:the official journal of the Society for Neuroscience 26:10129-10140
Ohaegbulam KC,Assal A,Lazar-Molnar E,Yao Y,Zang X(2015)Human cancer immunotherapy with antibodies to the PD-1 and PD-L1 pathway.Trends in molecular medicine 21:24-33
Pere H,Montier Y,Bayry J,Quintin-Colonna F,Merillon N,Dransart E,Badoual C,Gey A,Ravel P,Marcheteau E,Batteux F,Sandoval F,Adotevi O,Chiu C,Garcia S,Tanchot C,Lone YC,Ferreira LC,Nelson BH,Hanahan D,Fridman WH,Johannes L,Tartour E(2011)A CCR4 antagonist combined with vaccines induces antigen-specific CD8+T cells and tumor immunity against self antigens.Blood 118:4853-4862
Qin A,Wen Z,Zhou Y,Li Y,Li Y,Luo J,Ren T,Xu L(2013)MicroRNA-126 regulates the induction and function of CD4(+)Foxp3(+)regulatory T cells through PI3K/AKT pathway.Journal of cellular and molecular medicine 17:252-264
Rosenkranz D,Weyer S,Tolosa E,Gaenslen A,Berg D,Leyhe T,Gasser T,Stoltze L(2007)Higher frequency of regulatory T cells in the elderly and increased suppressive activity in neurodegeneration.Journal of neuroimmunology 188:117-127
Roy S,Barik S,Banerjee S,Bhuniya A,Pal S,Basu P,Biswas J,Goswami S,Chakraborty T,Bose A,Baral R(2013)Neem leaf glycoprotein overcomes indoleamine 2,3 dioxygenase mediated tolerance in dendritic cells by attenuating hyperactive regulatory T cells in cervical cancer stage IIIB patients.Human immunology 74:1015-1023
Sakaguchi S,Yamaguchi T,Nomura T,Ono M(2008)Regulatory T cells and immune tolerance.Cell 133:775-787
Saresella M,Calabrese E,Marventano I,Piancone F,Gatti A,Calvo MG,Nemni R,Clerici M(2010)PD1 negative and PD1 positive CD4+T regulatory cells in mild cognitive impairment and Alzheimer's disease.Journal of Alzheimer's disease:JAD 21:927-938
Schmidt SD,Nixon RA,Mathews PM(2005)ELISA method for measurement of amyloid-beta levels.Methods in molecular biology 299:279-297
Schwartz M,Baruch K(2012)Vaccine for the mind:Immunity against self at the choroid plexus for erasing biochemical consequences of stressful episodes.Human vaccines&immunotherapeutics 8:1465-1468
Schwartz M,Baruch K(2014a)Breaking peripheral immune tolerance to CNS antigens in neurodegenerative diseases:boosting autoimmunity to fight-off chronic neuroinflammation.Journal of autoimmunity 54:8-14
Schwartz M,Baruch K(2014b)The resolution of neuroinflammation in neurodegeneration:leukocyte recruitment via the choroid plexus.The EMBO journal 33:7-22
Shankar GM,Li S,Mehta TH,Garcia-Munoz A,Shepardson NE,Smith I,Brett FM,Farrell MA,Rowan MJ,Lemere CA,Regan CM,Walsh DM,Sabatini BL,Selkoe DJ(2008)Amyloid-beta protein dimers isolated directly from Alzheimer's brains impair synaptic plasticity and memory.Nature medicine 14:837-842
Shechter R,London A,Varol C,Raposo C,Cusimano M,Yovel G,Rolls A,Mack M,Pluchino S,Martino G,Jung S,Schwartz M(2009)Infiltrating blood-derived macrophages are vital cells playing an anti-inflammatory role in recovery from spinal cord injury in mice.PLoS medicine 6:e1000113
Shechter R,Miller O,Yovel G,Rosenzweig N,London A,Ruckh J,Kim KW,Klein E,Kalchenko V,Bendel P,Lira SA,Jung S,Schwartz M(2013)Recruitment of beneficial M2 macrophages to injured spinal cord is orchestrated by remote brain choroid plexus.Immunity 38:555-569
Shevchenko I,Karakhanova S,Soltek S,Link J,Bayry J,Werner J,Umansky V,Bazhin AV(2013)Low-dose gemcitabine depletes regulatory T cells and improves survival in the orthotopic Panc02 model of pancreatic cancer.International journal of cancer Journal international du cancer 133:98-107
Simard AR,Soulet D,Gowing G,Julien JP,Rivest S(2006)Bone marrow-derived microglia play a critical role in restricting senile plaque formation in Alzheimer's disease.Neuron 49:489-502
Simpson TR,Li F,Montalvo-Ortiz W,Sepulveda MA,Bergerhoff K,Arce F,Roddie C,Henry JY,Yagita H,Wolchok JD,Peggs KS,Ravetch JV,Allison JP,Quezada SA(2013)Fc-dependent depletion of tumor-infiltrating regulatory T cells co-defines the efficacy of anti-CTLA-4 therapy against melanoma.The Journal of experimental medicine 210:1695-1710
Smith PM,Howitt MR,Panikov N,Michaud M,Gallini CA,Bohlooly YM,Glickman JN,Garrett WS(2013)The microbial metabolites,short-chain fatty acids,regulate colonic Treg cell homeostasis.Science 341:569-573
Suffner J,Hochweller K,Kuhnle MC,Li X,Kroczek RA,Garbi N,Hammerling GJ(2010)Dendritic cells support homeostatic expansion of Foxp3+regulatory T cells in Foxp3.LuciDTR mice.Journal of immunology 184:1810-1820
Terawaki S,Chikuma S,Shibayama S,Hayashi T,Yoshida T,Okazaki T,Honjo T(2011)IFN-alpha directly promotes programmed cell death-1transcription and limits the duration of T cell-mediated immunity.Journal of immunology 186:2772-2779
Terme M,Colussi O,Marcheteau E,Tanchot C,Tartour E,Taieb J(2012)Modulation of immunity by antiangiogenic molecules in cancer.Clinical&developmental immunology 2012:492920
Terme M,Tartour E,Taieb J(2013)VEGFA/VEGFR2-targeted therapies prevent the VEGFA-induced proliferation of regulatory T cells in cancer.Oncoimmunology 2:e25156
Thomas-Schoemann A,Batteux F,Mongaret C,Nicco C,Chereau C,Annereau M,Dauphin A,Goldwasser F,Weill B,Lemare F,Alexandre J(2012)Arsenic trioxide exerts antitumor activity through regulatory T cell depletion mediated by oxidative stress in a murine model of colon cancer.Journal of immunology 189:5171-5177
Torres KC,Araujo Pereira P,Lima GS,Bozzi IC,Rezende VB,Bicalho MA,Moraes EN,Miranda DM,Romano-Silva MA(2013)Increased frequency of T cells expressing IL-10 in Alzheimer disease but not in late-onset depression patients.Progress in neuro-psychopharmacology&biological psychiatry 47:40-45
Town T,Laouar Y,Pittenger C,Mori T,Szekely CA,Tan J,Duman RS,Flavell RA(2008)Blocking TGF-beta-Smad2/3 innate immune signaling mitigates Alzheimer-like pathology.Nature medicine 14:681-687
Tseng D,Volkmer JP,Willingham SB,Contreras-Trujillo H,Fathman JW,Fernhoff NB,Seita J,Inlay MA,Weiskopf K,Miyanishi M,Weissman IL(2013)Anti-CD47 antibody-mediated phagocytosis of cancer by macrophages primes an effective antitumor T-cell response.Proceedings of the National Academy of Sciences of the United States of America 110:11103-11108
Vom Berg J,Prokop S,Miller KR,Obst J,Kalin RE,Lopategui-Cabezas I,Wegner A,Mair F,Schipke CG,Peters O,Winter Y,Becher B,Heppner FL(2012)Inhibition of IL-12/IL-23 signaling reduces Alzheimer's disease-like pathology and cognitive decline.Nature medicine 18:1812-1819
Voo KS,Bover L,Harline ML,Vien LT,Facchinetti V,Arima K,Kwak LW,Liu YJ(2013)Antibodies targeting human OX40 expand effector T cells and block inducible and natural regulatory T cell function.Journal of immunology 191:3641-3650
Walsh JT,Zheng J,Smirnov I,Lorenz U,Tung K,Kipnis J(2014)Regulatory T cells in central nervous system injury:a double-edged sword.Journal of immunology 193:5013-5022
Ward FJ,Dahal LN,Wijesekera SK,Abdul-Jawad SK,Kaewarpai T,Xu H,Vickers MA,Barker RN(2013)The soluble isoform of CTLA-4 as a regulator of T-cell responses.European journal of immunology 43:1274-1285
Weber JS,Kudchadkar RR,Yu B,Gallenstein D,Horak CE,Inzunza HD,Zhao X,Martinez AJ,Wang W,Gibney G,Kroeger J,Eysmans C,Sarnaik AA,Chen YA(2013)Safety,efficacy,and biomarkers of nivolumab with vaccine in ipilimumab-refractory or-naive melanoma.Journal of clinical oncology:official journal of the American Society of Clinical Oncology 31:4311-4318
Weber MS,Hohlfeld R,Zamvil SS(2007)Mechanism of action of glatiramer acetate in treatment of multiple sclerosis.Neurotherapeutics:the journal of the American Society for Experimental NeuroTherapeutics 4:647-653
Weiskopf K,Ring AM,Schnorr PJ,Volkmer JP,Volkmer AK,Weissman IL,Garcia KC(2013)Improving macrophage responses to therapeutic antibodies by molecular engineering of SIRPalpha variants.Oncoimmunology 2:e25773
Wyss-Coray T,Rogers J(2012)Inflammation in Alzheimer disease-a brief review of the basic science and clinical literature.Cold Spring Harbor perspectives in medicine 2:a006346
Zeng J,See AP,Phallen J,Jackson CM,Belcaid Z,Ruzevick J,Durham N,Meyer C,Harris TJ,Albesiano E,Pradilla G,Ford E,Wong J,Hammers HJ,Mathios D,Tyler B,Brem H,Tran PT,Pardoll D,Drake CG,Lim M(2013)Anti-PD-1 blockade and stereotactic radiation produce long-term survival in mice with intracranial gliomas.International journal of radiation oncology,biology,physics 86:343-349
Zhao L,Sun L,Wang H,Ma H,Liu G,Zhao Y(2007)Changes of CD4+CD25+Foxp3+regulatory T cells in aged Balb/c mice.Journal of leukocyte biology 81:1386-1394
Zheng H,Fridkin M,Youdim M(2015)New approaches to treating Alzheimer's disease.Perspectives in medicinal chemistry 7:1-8
Zhou S,Chen L,Qin J,Li R,Tao H,Zhen Z,Chen H,Chen G,Yang Y,Liu B,She Z,Zhong C,Liang C(2013)Depletion of CD4+CD25+regulatory T cells promotes CCL21-mediated antitumor immunity.PloS one 8:e73952
Zhou X,Kong N,Wang J,Fan H,Zou H,Horwitz D,Brand D,Liu Z,Zheng SG(2010)Cutting edge:all-trans retinoic acid sustains the stability and function of natural regulatory T cells in an inflammatory milieu.Journal of immunology 185:2675-2679
165282PCT-CN-ELLA序列表
<110> 耶达研究与开发有限公司
M·埃森巴赫-施瓦茨
K·巴鲁克
N·罗森茨魏希
<120> 降低系统性调节性T细胞水平或活性来治疗CNS疾病和损伤
<130> YEDA-127 PCT
<150> 61/951,783,
<151> 2014-03-12
<150> 62/030,164
<151> 2014-07-29
<160> 56
<170> PatentIn 3.5版
<210> 1
<211> 15
<212> PRT
<213> 人工序列
<220>
<223> 合成的
<400> 1
Ala Ala Ala Tyr Ala Ala Ala Ala Ala Ala Lys Ala Ala Ala Ala
1 5 10 15
<210> 2
<211> 15
<212> PRT
<213> 人工序列
<220>
<223> 合成的
<400> 2
Ala Glu Lys Tyr Ala Ala Ala Ala Ala Ala Lys Ala Ala Ala Ala
1 5 10 15
<210> 3
<211> 15
<212> PRT
<213> 人工序列
<220>
<223> 合成的
<400> 3
Ala Lys Glu Tyr Ala Ala Ala Ala Ala Ala Lys Ala Ala Ala Ala
1 5 10 15
<210> 4
<211> 15
<212> PRT
<213> 人工序列
<220>
<223> 合成的
<400> 4
Ala Lys Lys Tyr Ala Ala Ala Ala Ala Ala Lys Ala Ala Ala Ala
1 5 10 15
<210> 5
<211> 15
<212> PRT
<213> 人工序列
<220>
<223> 合成的
<400> 5
Ala Glu Ala Tyr Ala Ala Ala Ala Ala Ala Lys Ala Ala Ala Ala
1 5 10 15
<210> 6
<211> 15
<212> PRT
<213> 人工序列
<220>
<223> 合成的
<400> 6
Lys Glu Ala Tyr Ala Ala Ala Ala Ala Ala Lys Ala Ala Ala Ala
1 5 10 15
<210> 7
<211> 15
<212> PRT
<213> 人工序列
<220>
<223> 合成的
<400> 7
Ala Glu Glu Tyr Ala Ala Ala Ala Ala Ala Lys Ala Ala Ala Ala
1 5 10 15
<210> 8
<211> 15
<212> PRT
<213> 人工序列
<220>
<223> 合成的
<400> 8
Ala Ala Glu Tyr Ala Ala Ala Ala Ala Ala Lys Ala Ala Ala Ala
1 5 10 15
<210> 9
<211> 15
<212> PRT
<213> 人工序列
<220>
<223> 合成的
<400> 9
Glu Lys Ala Tyr Ala Ala Ala Ala Ala Ala Lys Ala Ala Ala Ala
1 5 10 15
<210> 10
<211> 15
<212> PRT
<213> 人工序列
<220>
<223> 合成的
<400> 10
Ala Ala Lys Tyr Glu Ala Ala Ala Ala Ala Lys Ala Ala Ala Ala
1 5 10 15
<210> 11
<211> 15
<212> PRT
<213> 人工序列
<220>
<223> 合成的
<400> 11
Ala Ala Lys Tyr Ala Glu Ala Ala Ala Ala Lys Ala Ala Ala Ala
1 5 10 15
<210> 12
<211> 15
<212> PRT
<213> 人工序列
<220>
<223> 合成的
<400> 12
Glu Ala Ala Tyr Ala Ala Ala Ala Ala Ala Lys Ala Ala Ala Ala
1 5 10 15
<210> 13
<211> 15
<212> PRT
<213> 人工序列
<220>
<223> 合成的
<400> 13
Glu Lys Lys Tyr Ala Ala Ala Ala Ala Ala Lys Ala Ala Ala Ala
1 5 10 15
<210> 14
<211> 15
<212> PRT
<213> 人工序列
<220>
<223> 合成的
<400> 14
Glu Ala Lys Tyr Ala Ala Ala Ala Ala Ala Lys Ala Ala Ala Ala
1 5 10 15
<210> 15
<211> 15
<212> PRT
<213> 人工序列
<220>
<223> 合成的
<400> 15
Ala Glu Lys Tyr Ala Ala Ala Ala Ala Ala Ala Ala Ala Ala Ala
1 5 10 15
<210> 16
<211> 15
<212> PRT
<213> 人工序列
<220>
<223> 合成的
<400> 16
Ala Lys Glu Tyr Ala Ala Ala Ala Ala Ala Ala Ala Ala Ala Ala
1 5 10 15
<210> 17
<211> 15
<212> PRT
<213> 人工序列
<220>
<223> 合成的
<400> 17
Ala Lys Lys Tyr Glu Ala Ala Ala Ala Ala Ala Ala Ala Ala Ala
1 5 10 15
<210> 18
<211> 15
<212> PRT
<213> 人工序列
<220>
<223> 合成的
<400> 18
Ala Lys Lys Tyr Ala Glu Ala Ala Ala Ala Ala Ala Ala Ala Ala
1 5 10 15
<210> 19
<211> 15
<212> PRT
<213> 人工序列
<220>
<223> 合成的
<400> 19
Ala Glu Ala Tyr Lys Ala Ala Ala Ala Ala Ala Ala Ala Ala Ala
1 5 10 15
<210> 20
<211> 15
<212> PRT
<213> 人工序列
<220>
<223> 合成的
<400> 20
Lys Glu Ala Tyr Ala Ala Ala Ala Ala Ala Ala Ala Ala Ala Ala
1 5 10 15
<210> 21
<211> 15
<212> PRT
<213> 人工序列
<220>
<223> 合成的
<400> 21
Ala Glu Glu Tyr Lys Ala Ala Ala Ala Ala Ala Ala Ala Ala Ala
1 5 10 15
<210> 22
<211> 15
<212> PRT
<213> 人工序列
<220>
<223> 合成的
<400> 22
Ala Ala Glu Tyr Lys Ala Ala Ala Ala Ala Ala Ala Ala Ala Ala
1 5 10 15
<210> 23
<211> 15
<212> PRT
<213> 人工序列
<220>
<223> 合成的
<400> 23
Glu Lys Ala Tyr Ala Ala Ala Ala Ala Ala Ala Ala Ala Ala Ala
1 5 10 15
<210> 24
<211> 15
<212> PRT
<213> 人工序列
<220>
<223> 合成的
<400> 24
Ala Ala Lys Tyr Glu Ala Ala Ala Ala Ala Ala Ala Ala Ala Ala
1 5 10 15
<210> 25
<211> 15
<212> PRT
<213> 人工序列
<220>
<223> 合成的
<400> 25
Ala Ala Lys Tyr Ala Glu Ala Ala Ala Ala Ala Ala Ala Ala Ala
1 5 10 15
<210> 26
<211> 15
<212> PRT
<213> 人工序列
<220>
<223> 合成的
<400> 26
Glu Lys Lys Tyr Ala Ala Ala Ala Ala Ala Ala Ala Ala Ala Ala
1 5 10 15
<210> 27
<211> 15
<212> PRT
<213> 人工序列
<220>
<223> 合成的
<400> 27
Glu Ala Lys Tyr Ala Ala Ala Ala Ala Ala Ala Ala Ala Ala Ala
1 5 10 15
<210> 28
<211> 15
<212> PRT
<213> 人工序列
<220>
<223> 合成的
<400> 28
Ala Glu Tyr Ala Lys Ala Ala Ala Ala Ala Ala Ala Ala Ala Ala
1 5 10 15
<210> 29
<211> 15
<212> PRT
<213> 人工序列
<220>
<223> 合成的
<400> 29
Ala Glu Lys Ala Tyr Ala Ala Ala Ala Ala Ala Ala Ala Ala Ala
1 5 10 15
<210> 30
<211> 15
<212> PRT
<213> 人工序列
<220>
<223> 合成的
<400> 30
Glu Lys Tyr Ala Ala Ala Ala Ala Ala Ala Ala Ala Ala Ala Ala
1 5 10 15
<210> 31
<211> 15
<212> PRT
<213> 人工序列
<220>
<223> 合成的
<400> 31
Ala Tyr Lys Ala Glu Ala Ala Ala Ala Ala Ala Ala Ala Ala Ala
1 5 10 15
<210> 32
<211> 15
<212> PRT
<213> 人工序列
<220>
<223> 合成的
<400> 32
Ala Lys Tyr Ala Glu Ala Ala Ala Ala Ala Ala Ala Ala Ala Ala
1 5 10 15
<210> 33
<211> 24
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 33
agcatacagg tcctggcatc ttgt 24
<210> 34
<211> 24
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 34
caaagaccac atgcttgcca tcca 24
<210> 35
<211> 24
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 35
agatcacatt cacggtgctg gcta 24
<210> 36
<211> 24
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 36
agctttggga tggtagctgg aaga 24
<210> 37
<211> 24
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 37
tgtgaaggga ttaacgaggc tgga 24
<210> 38
<211> 24
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 38
ccatgtttcg ggcacatttc caca 24
<210> 39
<211> 21
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 39
aactgcatcc atatcgatga c 21
<210> 40
<211> 19
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 40
gtggcaatga tctcaacac 19
<210> 41
<211> 18
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 41
catccacgtg ttggctca 18
<210> 42
<211> 23
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 42
gatcatcttg ctggtgaatg agt 23
<210> 43
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 43
gcctcttctc attcctgctt 20
<210> 44
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 44
ctcctccact tggtggtttg 20
<210> 45
<211> 21
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 45
ccaaaagatg aagggctgct t 21
<210> 46
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 46
tgctgctgcg agatttgaag 20
<210> 47
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 47
gaagttcaac atcaagagca 20
<210> 48
<211> 19
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 48
catagtccct ttggtccag 19
<210> 49
<211> 24
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 49
tgaattccct gggtgagaag ctga 24
<210> 50
<211> 24
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 50
tggccttgta gacaccttgg tctt 24
<210> 51
<211> 24
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 51
aattgctgcc ttcgccctct ttac 24
<210> 52
<211> 24
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 52
tgtacaggct gaggactttg gtgt 24
<210> 53
<211> 19
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 53
ccggaccaga gaccctttg 19
<210> 54
<211> 23
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 54
cctgtgggct tgttgaagta aaa 23
<210> 55
<211> 24
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 55
gatgctcagc agtcaagtgc cttt 24
<210> 56
<211> 24
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 56
gacatgtttg cggcatccag gtaa 24
- 还没有人留言评论。精彩留言会获得点赞!