一种重组人TNFR-Fc融合蛋白突变体的用途的制作方法
本发明涉及生物制药领域,特别涉及一种重组人tnfr-fc融合蛋白突变体的新用途。
背景技术
肿瘤坏死因子α(tnfα)是一种有效的前炎症细胞因子,其对不同的细胞类型表现出多重效应且对慢性炎症和自身免疫性疾病的发病机理起到关键作用。目前已有两类经典tnfα拮抗剂在临床上应用,包括:可溶性tnf受体-fc融合蛋白(etanercept——依那西普);抗tnfα单克隆抗体(adalimumab——阿达木单抗,infliximab——英夫利昔单抗,golimumab——戈利木单抗以及certolizumabpegol——赛妥珠单抗),成为治疗类风湿性关节炎、强直性脊柱炎、银屑病、溃疡性结肠炎的重要的药物。
虽然与可溶性tnfα(stnfα)的结合和发挥中和活性是这些抗tnfα药物的关键作用机制(moa),但积累的证据表明,除了stnfα,其前体形式——膜结合tnfα(mtnfα),也参与了炎症反应。在类风湿性关节炎(ra)中,抗tnfα单抗被认为主要通过中和stnfα和mtnfα发挥疗效。在其他情况下,如克罗恩病(cd)和溃疡性结肠炎(uc)这两种主要类型的炎症性肠病(ibd),通过mtnfα和fcγ受体(触发凋亡或adcc)的信号传导可能发挥更重要的作用(horiuchit,mitomah,harashimas,tsukamotoh,shimodat.transmembranetnf-alpha:structure,functionandinteractionwithanti-tnfagents.rheumatology(oxford).2010;49(7):1215-1228.doi:10.1093/rheumatology/keq031;feaganbg,choquetted,ghoshs,gladmandd,hov,meibohmb,zoug,xuz,shankarg,sealeydc,etal.thechallengeofindicationextrapolationforinfliximabbiosimilars.biologicals.2014;42(4):177-183.doi:10.1016/j.biologicals.2014.05.005)。
另一方面,已有的研究表明:现有tnfα拮抗剂药物均可抑制stnfα或mtnfα与tnfr的结合,即发挥中和作用,tnfr-fc融合蛋白依那西普显示比tnfα单抗(英夫利昔单抗、阿达木单抗)更强的stnfα中和能力;然而当这些药物与mtnfα结合时,它们具有的诱导fc介导的adcc或cdc作用的潜力不同,对于mtnfα介导的adcc效应和cdc效应,英夫利昔单抗、阿达木单抗显示出比依那西普更强的效果,原因在于tnfr-fc融合蛋白结合mtnfα和fcγ受体的信号传导的效果差(mitomah,horiuchit,tsukamotoh,tamimotoy,kimotoy,uchinoa,tok,harashimas,hattan,haradam.mechanismsforcytotoxiceffectsofanti-tumornecrosisfactoragentsontransmembranetumornecrosisfactoralpha-expressingcells:comparisonamonginfliximab,etanercept,andadalimumab.arthritisrheum.2008;58(5):1248-1257.doi:10.1002/art.23447;arorat,padakir,liul,hamburgerae,ellisonar,stevenssr,louiejs,kohnot.differencesinbindingandeffectorfunctionsbetweenclassesoftnfantagonists.cytokine.2009;45(2):124-131.doi:10.1016/j.cyto.2008.11.008),因此依那西普没有被批准用于克罗恩病和溃疡性结肠炎的治疗。
本申请人的专利文献cn101591388a公开了一种重组人tnfr-fc融合蛋白突变体(t0001),在tnfr区域具有w89y/e92n突变位点。t0001是基于对tnfα与tnfr的结合表位设计,筛选出的高亲和力突变体,体现出比etanercept高1.5倍的中和活性。在抑制由胶原介导的大鼠关节炎方面具有明显的增效作用。这些数据表明,与依那西普相比,在类风湿性关节炎患者中,高亲和力变体可能有更好的治疗效果(yangt等,ploscomputbiol.2010;6(2):e1000669.)。
基于此,本发明人旨在寻找一种可以作用于mtnfα、具有adcc或者cdc效应,从而可用于制备治疗克罗恩病和溃疡性结肠炎药物的tnfr-fc融合蛋白。
技术实现要素:
本发明所要解决的技术问题是为克服现有技术中存在的tnfr-fc融合蛋白的adcc或者cdc作用弱,无法通过adcc或者cdc发挥炎性肠病治疗作用,提供一种重组人tnfr-fc融合蛋白突变体可作用于mtnfα、增强adcc或者cdc活化效应,从而可用于炎性肠病防治的新用途。
本发明人经过多次尝试后惊喜地发现,重组人tnfr-fc融合蛋白突变体(t0001)(专利文献cn101591388a)相对于依那西普有显著增强的mtnfα结合作用以及adcc效应和cdc效应。
本发明解决上述技术问题所采用的技术方案之一是:一种重组人tnfr-fc融合蛋白突变体的用途,所述的重组人tnfr-fc融合蛋白突变体的tnfrp75部分的氨基酸序列如seqidno.1所示,所述的用途为将所述重组人tnfr-fc融合蛋白突变体用于制备mtnfα拮抗剂、adcc和/或cdc效应的活化剂。其中adcc和/或cdc效应的活化剂指的是具有adcc和/或cdc效应功能或者活性的试剂。
本发明解决上述技术问题的另一技术方案是:一种重组人tnfr-fc融合蛋白突变体用途,所述的重组人tnfr-fc融合蛋白突变体的tnfrp75部分的氨基酸序列如seqidno.1所示,所述的用途为将所述重组人tnfr-fc融合蛋白突变体用于制备治疗炎性肠病的药物。
本发明中,“tnfp75部分”指的是tnfp75受体的细胞外部分,即野生型tnfp75受体氨基酸序列n端第1-257位氨基酸构成的与配体结合的结构域,n段1-22位氨基酸为信号肽;“融合蛋白”为本领域常规概念,指通过dna重组技术得到的两个基因重组后的表达产物;“突变体”意指带有点突变——e92n、w89y的tnfr-fc融合蛋白,即tnfp75受体氨基酸序列的第92位由谷氨酸(e)突变为天冬酰胺(n),第89位由色氨酸(w)突变为酪氨酸(y)。
其中,所述fc段为人免疫球蛋白(igg)的恒定区;较佳地,所述的人重组tnfr-fc融合蛋白突变体的氨基酸序列如seqidno.2所示。
所述的重组人tnfr-fc融合蛋白突变体可以作为mtnfα拮抗剂、adcc和/或cdc效应的活化剂、或者制备治疗炎性肠病的药物的唯一活性成分;还可以与其他活性成分例如现有tnfα拮抗剂共同起作用。
所述炎性肠病包括但不仅限于克罗恩病和溃疡性结肠炎等。
根据本发明,重组人tnfr-fc融合蛋白突变体可以依据文献ploscomputbiol.2010;6(2):e1000669.(第4-6页的“materialsandmethods”部分)及专利cn101591388a(第11页实施例1-7)的记载制备得到;也可以从上海复旦张江生物医药股份有限公司购得。
在符合本领域常识的基础上,上述各优选条件,可任意组合,即得本发明各较佳实例。
本发明所用试剂和原料均市售可得。
本发明的积极进步效果在于:
本发明发现重组人tnfr-fc融合蛋白w89y/e92n突变体(t0001)在有stnfα存在下,与fcγriiia的竞争性结合力是etanercept的1.5倍;t0001对stnfα的中和能力是依那西普的2倍,是英夫利昔单抗和阿达木单抗的约20倍;并且其在与mtnfα的竞争性结合、诱导adcc效应方面效果与阿达木单抗相当,可以将其用于制备治疗炎性肠病,特别是克罗恩病和溃疡性结肠炎的药物。
附图说明
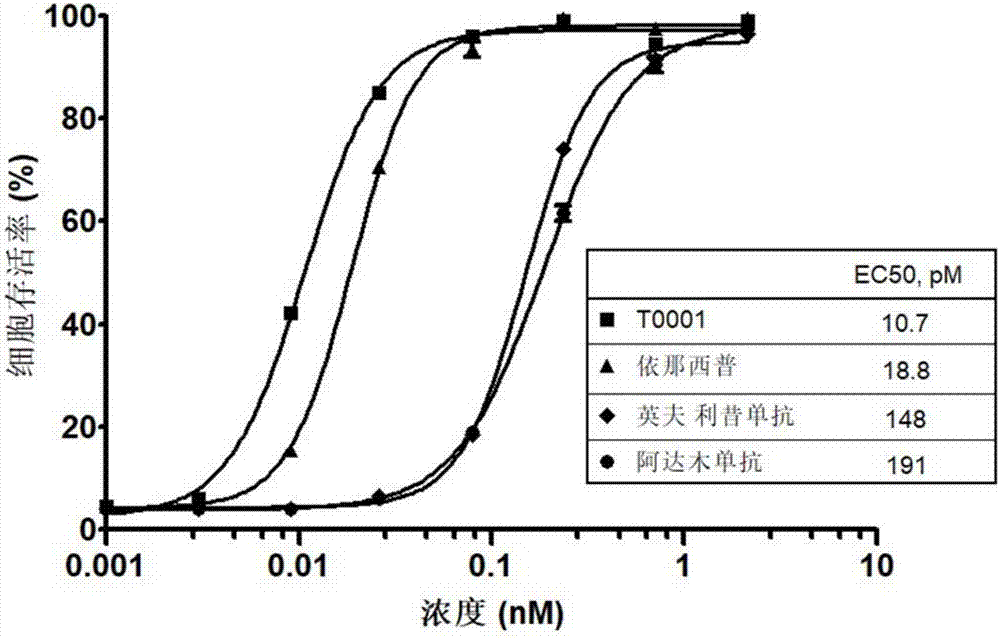
图1为tnfα拮抗剂对小鼠l929细胞中stnfα诱导的细胞毒活性的中和活性。
图2为tnfα拮抗剂对mtnfα转染细胞的竞争性结合。
图3为tnfα拮抗剂与fcγriiia在转染细胞上的竞争性结合,其中:a为t0001,b为依那西普,c为英夫利昔单抗,d为阿达木单抗。
图4为报告基因法测定tnfα拮抗剂介导adcc的能力。
图5为pbmc测定法测定tnfα拮抗剂介导adcc的能力。
图6为tnfα拮抗剂与c1q的竞争性结合,其中:a为t0001,b为依那西普,c为英夫利昔单抗,d为阿达木单抗。
图7为tnfα拮抗剂介导cdc的能力。
具体实施方式
下面通过实施例的方式进一步说明本发明,但并不因此将本发明限制在所述的实施例范围之中。下列实施例中未注明具体条件的实验方法,按照常规方法和条件,或按照商品说明书选择。
本发明为了探讨重组人tnfr-fc融合蛋白w89y/e92n突变体(t0001)在克罗恩病和溃疡性结肠炎中的治疗潜力,下述实施例将t0001与3种代表性的临床可用tnfα拮抗剂:etanercept、infliximab以及adalimumab相比较,评价了t0001的结合特征和fc效应功能。
(1)tnfα拮抗剂
t0001:t0001是etanercept的一个高亲和力变体,包含tnfr区域和fc区域,在tnfr区域具有w89y/e92n突变位点,参见中国发明专利公开号cn101591388a中的seqidno.6(具体请参见本发明序列表中的seqidno.1);t0001的全长氨基酸序列参见中国发明专利公开号cn101591388a中的seqidno.13(具体请参见本发明序列表中的seqidno.2)。t0001由上海复旦张江生物医药股份有限公司制备,制备方法参见文献yangt,等,ploscomputbiol.2010;6(2):e1000669.中的第4-6页的“materialsandmethods”部分及专利cn101591388a中的实施例1-7;也可以从上海复旦张江生物医药股份有限公司购得。
infliximab
etanercept
adalimumab
rituximab
(2)细胞系
l929细胞:tnf敏感的鼠纤维肉瘤细胞系,本发明用做tnfα诱导的细胞毒中和试验的靶细胞。购自中国科学院典型培养物保藏委员会细胞库。
pbmcs:外周血单核细胞,本发明用作adcc试验的效应细胞。购自hemacare,(cat.pb009c-2)。
表达膜型tnfα(mtnfα)的cho细胞:本发明用于tnfα拮抗剂竞争性结合试验,也作为tnfα拮抗剂adcc效应试验的靶细胞,制备方法参见实施例1。
wil2-s细胞:白细胞分化抗原cd20阳性的人b淋巴细胞,全称为wil2-s(
jurkatt细胞:来自人急性t细胞白血病的t淋巴细胞系,全称为jurkat,clonee6-1(
jurkat-nfat荧光素酶报告基因细胞系(jurkat-nfatluciferasereportercellline):稳定转染fcγriiia和nfat-luciferasereportergene报告元件的jurkatt细胞,本发明用于tnfα拮抗剂与fcγriiia竞争性结合试验,也作为tnfα拮抗剂adcc效应报告基因试验的效应细胞。制备方法参见实施例1。
实施例1用于t0001功能学评价的细胞株的构建
1.1表达mtnfα的cho细胞系的构建
参照arorat,等的文献方法构建表达mtnfα的cho细胞系,参见cytokine.2009;45(2):124-131。简单来说,通过定点突变法构建了缺失77至88位氨基酸序列的膜结合tnfα(mtnfα)突变体,其膜外部分可不受tace酶的降解。将mtnfα突变体的dna序列克隆于pires表达载体(clontechlaboratories,inc.cat.631605)。pires载体含有cmvie启动子,在哺乳动物细胞中可驱动mtnfα基因的表达和neomycin抗性标记的筛选。通过将mtnfα/pires转染cho-k1(atcc,ccl-61)细胞构建mtnfα细胞株,转染后的细胞通过500μg/mlg418筛选,并采用facscalibur流式细胞仪(bectondickinson)筛选高表达mtnfα的单克隆细胞株。
1.2jurkat-nfat荧光素酶报告基因细胞株的构建
参照parekhbs等的文献方法构建jurkat-nfat荧光素酶报告基因细胞株,参见mabs.2012;4(3):310-318。简单来说,将表达人fcγriiia/158v的pcdna3.1/hygro(invitrogen,cat.v87020)质粒和表达人fcεriγ的pcdna3.1/zeo(invitrogen,cat.v86020)质粒各取0.8μg共转染密度为8×105个/ml的jurkatt细胞(atcc,tib-152)细胞,流式细胞仪筛选出具有潮霉素(hygromycin)和博莱霉素(zeocin)双重抗性的表达fcγriiia的细胞,并进一步通过fcr抗体诱导的il-2分泌进行细胞株确认。取前一步确认的细胞5×104个/ml,加入50μl滴度为2×107tu/ml的pgmlv-nfat-lu慢病毒悬液(genomeditech,cat.gm-022012),经过嘌呤霉素(puromycin)抗性选择,获得可受nfat调控表达荧光素酶报告基因的细胞株。这些细胞株再通过nfat激活剂(10ng/mlpma和0.5μm离子霉素(ionomycin))进行诱导荧光素酶表达筛选,获得非特异性表达低的理想细胞株。
下述实施例2~8为t0001的功能学评价。
实施例2在小鼠l929细胞中,对可溶性tnfα诱导的细胞毒性的中和测定
小鼠l929细胞在含3%fbs(moregate,cat.fbsfc-500)的dmem(源培,cat.l170)培养基中以1.5×104/孔密度接种96孔细胞培养板,细胞在37℃培养24小时后加入不同浓度(0.001nm、0.003nm、0.009nm、0.026nm、0.079nm、0.237nm、0.711nm以及2.133nm)的上述不同tnfα拮抗剂、1μg/ml放线菌素d(actinomycind)(solarbio,cat.a8030)和20iu/mlstnfα(nibsc,12/154),并在37℃继续培养18小时。将不加tnfα拮抗剂和stnfα的孔作为空白对照,采用mts试剂(promega,cat.g5430)测定细胞的活力。每孔加入mts试剂孵育4小时后通过微孔板酶标仪spectramaxm2e(moleculardevices)采集490nm的读数。细胞存活率(%)=药物处理组的吸光值/对照孔的吸光值×100,并采用graphpadprism5软件的四参数模式来进行反应曲线的拟合。
结果如图1所示,t0001对stnfα诱导的细胞毒活性显示出最强的中和活性。四种tnfα拮抗剂对tnfα诱导的细胞毒性的中和活性的半数有效浓度(ed50)分别为:t0001为10.7pm;etanercept为18.8pm;infliximab为148pm;adalimumab为191pm。两个tnfr-fc融合蛋白(t0001、etanercept)表现出比两个单抗(infliximab、adalimumab)大约10倍的中和活性,而t0001显示大约是etanercept的两倍的活性。
实施例3tnf拮抗剂与mtnfα的结合
t0001进行生物素标记,标记试剂采用
结果如图2所示,当与ec50浓度的生物素化t0001(~13.3nm)竞争时,按照百分比竞争活性,infliximab的ec50值为2.53nm,t0001和adalimumab的ec50值分别为6.44和5.86nm,而etanercept的ec50值为28.9nm。infliximab单抗在四种tnfα拮抗剂中是最具竞争性的,t0001和adalimumab单抗表现出类似的竞争力,它们对mtnfα的结合活性与英夫利昔单抗相比降低了约60%。etanercept的竞争结合能力最差。这些tnfα拮抗剂均不与未转染mtnfα的cho细胞结合。
实施例4tnf拮抗剂与fcγriiia的结合
一株非tnfα靶标的人鼠嵌合igg1单抗(rituximab,人鼠嵌合抗cd20抗体,购自roche)进行生物素标记,作为非tnfα靶标的igg1抗体,用于tnf拮抗剂与fcγriiia的竞争性结合试验。单抗标记试剂采用
结果如图3所示,不同tnfα拮抗剂与转染细胞上的fcγriiia的结合。在stnfα存在下,两种tnfα单抗infliximab和adalimumab比没有stnfα情况下,竞争性结合增加了4倍(图3c、图3d);而对于两个tnfr-fc融合蛋白t0001和etanercept,有stnfα比无stnfα情况下竞争性结合只有2倍的增加(图3a、图3b)。在所测试的药物中,英夫利昔单抗显示出与fcγriiia具有最佳的结合力。t0001在有stnfα存在下,与fcγriiia的竞争性结合力是etanercept的1.5倍。
实施例5adcc-报告基因测定
实施例1.1制备的表达mtnfα的cho细胞在rpmi1640(gibco,cat.a10491)培养基中以5×103/孔密度接种白色不透明96孔细胞培养板。在细胞中加入不同浓度(2.85nm、6.28nm、13.81nm、30.37nm、66.82nm、147nm、323.4nm、711.47nm、1565.24nm、3443.53nm以及7575.76nm)的上述不同tnfα拮抗剂于37℃,5%co2孵育1小时,随后每孔加入1×105实施例1.2制备的jurkat-nfat荧光素酶报告基因细胞。混合液于37℃,5%co2孵育6小时,随后采用bio-glotmluciferaseassaysystem(promega,g7940)测定相对荧光单位(rlus)。adcc效应采用诱导倍数(fi)进行计算,fi=(rlu诱导组–rlu空白组)/(rlu未诱导组–rlu空白组)(chengzj,garvind,paguioa,moravecr,engell,fanf,surowyt.developmentofarobustreporter-basedadccassaywithfrozen,thaw-and-usecellstomeasurefceffectorfunctionoftherapeuticantibodies.jimmunolmethods.2014;414:69-81.doi:10.1016/j.jim.2014.07.010.)。采用graphpadprism5软件的四参数模式来进行反应曲线的拟合。
将rituximab作用于wil2-s(cd20+)细胞组作为系统阳性对照组,将rituximab作用于表达mtnfα的cho细胞组作为系统阴性对照组。
结果如图4,当效应细胞与靶细胞之比(e:t)比为20:1时,四种tnfα拮抗剂的ec50值:infliximab为139nm,adalimumab为140nm,t0001为250nm,etanercept为547nm。英夫利昔单抗显示出最小的ec50和最高的最大fi值,表明比其他三种tnfα拮抗剂具有更强的adcc活性。如所预期的,两种单抗(infliximab和adalimumab)显示比两种fc-融合蛋白(t0001和etanercept)具有更强的adcc作用。在fc融合蛋白组中,t0001显示出超过依那西普2倍的adcc作用,这可能归因于其更强的与靶细胞表面mtnfα的结合。
实施例6经典adcc测定
经典adcc测定采用pbmc作为效应细胞。实施例1制备的表达mtnfα的cho细胞在含5%fbs(pan,cat.p30-2102)的rpmi1640培养基中以1×104/孔密度接种透明平底96孔细胞培养板。在细胞中加入不同浓度(10nm、100nm以及1000nm)的上述不同tnfα拮抗剂,随后每孔加入2×105pbmc(hemacare,cat.pb009c-2),于37℃,5%co2孵育4小时。采用cytotoxicityldhassaykit-wst(dojindo,cat.ck12)测定ldh的释放,在spectramaxm2e酶标仪上读取490nm吸光值,将仅含有靶细胞和效应细胞的孔作为ldh自发释放对照组,另取用裂解液将靶细胞完全裂解的孔作为ldh最大释放对照组。adcc效应采用百分比裂解度进行计算,百分比裂解度(%)=(ldh药物释放组-ldh自发释放组)/(ldh最大释放组-ldh自发释放组)×100。
结果如图5,t0001显示出与报告基因法相似的adcc效应,进一步确认了t0001具有adcc效应功能。t0001的adcc效应接近于adalimumab,远高于etanercept。
实施例7tnf拮抗剂与c1q的结合
在96孔板(nunc,cat.446469)中包被5μg/ml羊抗人补体c1q抗体(goatanti-humanc1qantibody;购自accuratechemical&scientificcorporation,cat.ynghc1q),于4℃孵育16小时,随后采用pbst/3%bsa室温封闭2小时。封闭后每孔加入100μl3μg/ml人补体c1q(购自quidelcorporation,cat.a400)在室温孵育2小时。在板孔中单独加入不同浓度(40nm、200nm、1000nm、2000nm、4000nm、8000nm、16000nm以及32000nm)的上述不同tnfα拮抗和133.3nm生物素化的rituximabigg1抗体,或同时加入0.8倍摩尔比的stnfα。96孔板于室温孵育15小时,随后加入streptavidin-hrp(碧云天,cat.a0303)于室温孵育1小时。在每一部孵育步骤后都采用含0.05%tween-20的pbs进行洗涤。过氧化物酶活性通过tmb试剂(thermoscientific,cat.34028)进行测定,96孔板在室温放置30分钟进行显色,随后加入1mh2so4终止显色反应,在spectramaxm2e酶标仪上读取450nm吸光值(需扣除650nm的背景值。仅加入生物素化igg1的细胞作为对照组,百分比竞争活性(%)=(对照组吸光值–药物组吸光值)/对照组吸光值×100。
结果如图6所示,所有四种tnfα拮抗剂在缺乏外源性stnfα的情况下显示出对c1q的差的竞争性结合。
然而,在stnfα存在时,infliximab和adalimumab显示出对c1q的显著竞争性结合活性(图6c、图6d)。infliximab单抗在这些拮抗剂中的竞争活性提升最大,而t0001和etanercept两种fc-融合蛋白在低于4000nm浓度时没有获得它们的竞争性变化(图6a、图6b)。
实施例8cdc测定
实施例1制备的表达mtnfα的cho细胞在gmem(gibco,cat.11710-035)培养基中以5×104/孔密度接种黑色96孔细胞培养板(corning,cat.3603)。在细胞中加入不同浓度(4.17nm、104.17nm、208.33nm、416.67nm、833.33nm、1666.67nm以及3333.33nm)的tnfα拮抗剂于4℃孵育1小时,随后加入人补体血清(quidel,cat.a113)至终浓度8%,于37℃,5%co2孵育2小时。每孔加入50μlalamarbluetm(invitrogen,cat.dal1100)后继续在相同的条件下孵育17小时。将不加tnfα拮抗剂和补体血清的细胞作为对照组,96孔板在spectramaxm2e酶标仪上读取530/590nm(激发/发射波长)。细胞死亡率(%)=(对照组吸光值–药物组吸光值)/对照组吸光值×100。采用graphpadprism5软件的四参数模式来进行反应曲线的拟合。将rituximab作用于wil2-s(cd20+)细胞组作为系统阳性对照组,将rituximab作用于表达mtnfα的cho细胞组作为系统阴性对照组。
结果如图7示,在该测定中,两种单抗infliximab和adalimumab比t0001和etanercept可更有效地诱导cdc活性,t0001具有比etanercept高2.4倍的cdc效力,这同样可以通过其对靶细胞更强的结合力来解释。在用缺乏补体成分c5的血清或使用热灭活的补体孵育的培养组中未观察到tnfα拮抗剂依赖性细胞死亡,采用对照抗体rituximab单抗作为系统对照进行测试,利妥昔单抗在表达mtnfα的cho细胞上没有检测到cdc活性,但在使用wil2-s(cd20+)细胞作为靶细胞时,利妥昔单抗的剂量-反应曲线显示出其具有很强的cdc活性。
显然,本领域的技术人员可以对本发明进行各种改动和变型而不脱离本发明的精神和范围。这样,倘若本发明的这些修改和变型属于本发明权利要求及其等同技术的范围之内,则本发明也意图包含这些改动和变型在内。
<110>上海复旦张江生物医药股份有限公司
<120>一种重组人tnfr-fc融合蛋白突变体的用途
<130>p1710538c
<160>2
<170>patentinversion3.5
<210>1
<211>257
<212>prt
<213>人工序列
<220>
<223>重组人tnfr-fc融合蛋白突变体的tnfrp75部分的氨基酸序列
<400>1
metalaprovalalavaltrpalaalaleualavalglyleugluleu
151015
trpalaalaalahisalaleuproalaglnvalalaphethrprotyr
202530
alaprogluproglyserthrcysargleuargglutyrtyraspgln
354045
thralaglnmetcyscysserlyscysserproglyglnhisalalys
505560
valphecysthrlysthrseraspthrvalcysaspsercysgluasp
65707580
serthrtyrthrglnleutrpasntyrvalproasncysleusercys
859095
glyserargcysserseraspglnvalgluthrglnalacysthrarg
100105110
gluglnasnargilecysthrcysargproglytrptyrcysalaleu
115120125
serlysglngluglycysargleucysalaproleuarglyscysarg
130135140
proglypheglyvalalaargproglythrgluthrseraspvalval
145150155160
cyslysprocysalaproglythrpheserasnthrthrserserthr
165170175
aspilecysargprohisglnilecysasnvalvalalaileprogly
180185190
asnalasermetaspalavalcysthrserthrserprothrargser
195200205
metalaproglyalavalhisleuproglnprovalserthrargser
210215220
glnhisthrglnprothrprogluproserthralaproserthrser
225230235240
pheleuleuprometglyproserproproalagluglyserthrgly
245250255
asp
<210>2
<211>489
<212>prt
<213>人工序列
<220>
<223>重组人tnfr-fc融合蛋白突变体的全长氨基酸序列
<400>2
metalaprovalalavaltrpalaalaleualavalglyleugluleu
151015
trpalaalaalahisalaleuproalaglnvalalaphethrprotyr
202530
alaprogluproglyserthrcysargleuargglutyrtyraspgln
354045
thralaglnmetcyscysserlyscysserproglyglnhisalalys
505560
valphecysthrlysthrseraspthrvalcysaspsercysgluasp
65707580
serthrtyrthrglnleutrpasntyrvalproasncysleusercys
859095
glyserargcysserseraspglnvalgluthrglnalacysthrarg
100105110
gluglnasnargilecysthrcysargproglytrptyrcysalaleu
115120125
serlysglngluglycysargleucysalaproleuarglyscysarg
130135140
proglypheglyvalalaargproglythrgluthrseraspvalval
145150155160
cyslysprocysalaproglythrpheserasnthrthrserserthr
165170175
aspilecysargprohisglnilecysasnvalvalalaileprogly
180185190
asnalasermetaspalavalcysthrserthrserprothrargser
195200205
metalaproglyalavalhisleuproglnprovalserthrargser
210215220
glnhisthrglnprothrprogluproserthralaproserthrser
225230235240
pheleuleuprometglyproserproproalagluglyserthrgly
245250255
aspgluprolyssercysasplysthrhisthrcysproprocyspro
260265270
alaprogluleuleuglyglyproservalpheleupheproprolys
275280285
prolysaspthrleumetileserargthrprogluvalthrcysval
290295300
valvalaspvalserhisgluaspprogluvallyspheasntrptyr
305310315320
valaspglyvalgluvalhisasnalalysthrlysproarggluglu
325330335
glntyrasnserthrtyrargvalvalservalleuthrvalleuhis
340345350
glnasptrpleuasnglylysglutyrlyscyslysvalserasnlys
355360365
alaleuproalaproileglulysthrileserlysalalysglygln
370375380
proarggluproglnvaltyrthrleuproproserarggluglumet
385390395400
thrlysasnglnvalserleuthrcysleuvallysglyphetyrpro
405410415
seraspilealavalglutrpgluserasnglyglnprogluasnasn
420425430
tyrlysthrthrproprovalleuaspseraspglyserphepheleu
435440445
tyrserlysleuthrvalasplysserargtrpglnglnglyasnval
450455460
phesercysservalmethisglualaleuhisasnhistyrthrgln
465470475480
lysserleuserleuserproglylys
485
- 还没有人留言评论。精彩留言会获得点赞!