疼痛的治疗的制作方法
本发明涉及化合物及其用途。特别是,涉及抑制核内体蛋白酶激活的受体-2(endosomalprotease-activatedreceptor-2,par2)信号传导的化合物及其治疗疼痛的用途。
背景技术:
g蛋白-偶联受体(gpcr)是细胞表面受体的最大家族,其参与大多数病理生理学过程,且是约30%治疗药物的靶标(audet,m.&bouvier,m.natchembiol2008,4,397–403)。细胞表面gpcr与细胞外配体相互作用并偶联至异源三聚体g蛋白,其触发质膜定界信号(第二信使形成,生长因子受体反式激活,离子通道调节)。配体去除和受体与β-抑制蛋白(βarrs)的关联终止质膜信号。
直到最近,人们普遍认为gpcr的活化,随后的下游信号传导和信号终止仅发生在质膜上。质膜信号传导通过gpcr激酶(grk)(其对活性配体结合的受体构象具有选择性)使受体磷酸化而在活化的几分钟内终止。grk磷酸化gpcr的c-端s/t-富集结构域(sato,p.y.,等人,physiologicalreviews2015,95,377-404)。然后磷酸化的受体与βarr结合,其在空间上阻止受体和g-蛋白之间的偶联,从而终止激动剂-介导的g-蛋白活化。βarrs通过发动蛋白-和网格蛋白-依赖的内吞作用,进一步促进配体结合的受体从细胞表面向早期核内体的转移。一旦发生内吞作用,从gpcr中除去配体和磷酸基团,且受体迅速重新分配至细胞膜或被转运到溶酶体以降解。
然而,最近发现多种gpcr并不总是遵循传统的范例。研究已发现在配体结合和受体活化后,一些细胞表面gpcr内化并重新分配至早期核内体,其中异源三聚体g蛋白信号传导维持较长时间。因此,核内体不仅仅是作为gpcr向再循环或降解途径运输的管道,而且可以成为信号转导的重要部位(murphy,j.e.等人procnatlacadsciusa2009,106,17615-17622)。通过募集gpcr和有丝分裂素激活蛋白激酶类到核内体,βarrs可以介导核内体gpcr信号传导(murphy,j.e.等人procnatlacadsciusa2009,106,17615-17622;defea,k.a.等人procnatlacadsciusa2000,97,11086-11091;defea,k.a.等人jcellbiol2000,148,1267-1281)。
βarrs在血浆和核内体膜上募集多种信号传导蛋白以激活受体,且其是信号传导的必要介质。所述mapk级联反应[erk,c-jun氨基-末端激酶(jnk),p38]是最彻底表征的βarrs-依赖性信号传导通路。βarrs是信号传导的积极参与者的第一个证据是观察到βarr的显性失活突变体抑制β2ar-诱导的erk1/2激活(daakay等人,jbiolchem1998,273,685-688)。随后,发现βarrs将β2ar偶联至c-src并介导erk1/2激活(lutteralll.m.等人science1999,283,655-661)。通过其他gpcr(包括神经激肽-1受体(nk1r)、蛋白酶激活的受体2(par2)、血管紧张素ii1a型受体(at1ar)和血管加压素v2受体(v2r)),βarrs同样参与erk1/2信号传导。这些观察结果导致了这样的观点:βarrs是将激活的gpcr与mapk信号传导复合物偶联的支架。因此βarrs介导gpcr信号传导的第二波,其与在质膜上的g蛋白-依赖性信号传导不同。
技术实现要素:
本发明基于以下发现,即抑制par2的核内体信号传导可以提供治疗蛋白酶诱发的疼痛的新方法。蛋白酶激活的受体-2(par2)是蛋白酶诱发的炎症和疼痛的主要介质。par2由控制神经源性炎症和疼痛传递的初级感觉神经元表达。在损伤和炎症期间产生的多种蛋白酶可激活感觉神经上的par2,包括上皮衍生的胰蛋白酶iv、肥大细胞类胰蛋白酶、巨噬细胞组织蛋白酶s和中性粒细胞弹性蛋白酶。这些蛋白酶在初级感觉神经上断裂并激活par2,导致瞬时受体电位离子通道的敏化和神经肽(物质p和降钙素基因相关肽)的释放。这些肽引起外周炎症和疼痛的中枢传递。胰蛋白酶和类胰蛋白酶通过经典机制激活par2,导致β-抑制蛋白的募集和par2内吞作用。
现在已发现通过防止激活的受体的内吞作用或通过靶向和抑制核内体par2信号传导来抑制par2的核内体信号传导,提供了治疗蛋白酶诱发的疼痛的新的方法。
因此,在一方面本发明提供治疗蛋白酶诱发的疼痛的方法,包括向需要的受试者给药抑制蛋白酶激活的受体-2(par2)的核内体信号传导的化合物。
在一方面,所述抑制核内体par2信号传导的化合物为抑制激活的受体的内吞作用的化合物。
在另一方面,所述抑制核内体par2信号传导的化合物为靶向和抑制核内体par2信号传导的化合物。
在另一方面,所述抑制核内体par2信号传导的化合物为式(i)的抑制核内体par2信号传导的三部分化合物:
其中
a为脂质锚,其促进化合物插入质膜;
l为长度为1nm至50nm的连接基部分;且
x为核内体par2信号传导的抑制剂;
其中该脂质锚分配到在4℃不溶于非离子型洗涤剂的脂质膜中;或
其药学上可接受的盐。
在另一方面,本发明提供治疗蛋白酶诱发的疼痛的方法,包括向需要的受试者给药包含一种或多种抑制受体的内吞作用的化合物和一种或多种靶向和抑制核内体par2信号传导的化合物的组合。
在另一方面,本发明提供抑制par2的核内体信号传导的化合物在制备用于治疗蛋白酶诱发的疼痛的药物中的用途。
在另一方面,本发明提供抑制par2的核内体信号传导的化合物,其用于治疗蛋白酶诱发的疼痛。
在阅读以下发明详述并结合所附实施例和权利要求后,本发明的这些和其他方面对于本领域技术人员将变得更加明显。
附图简述
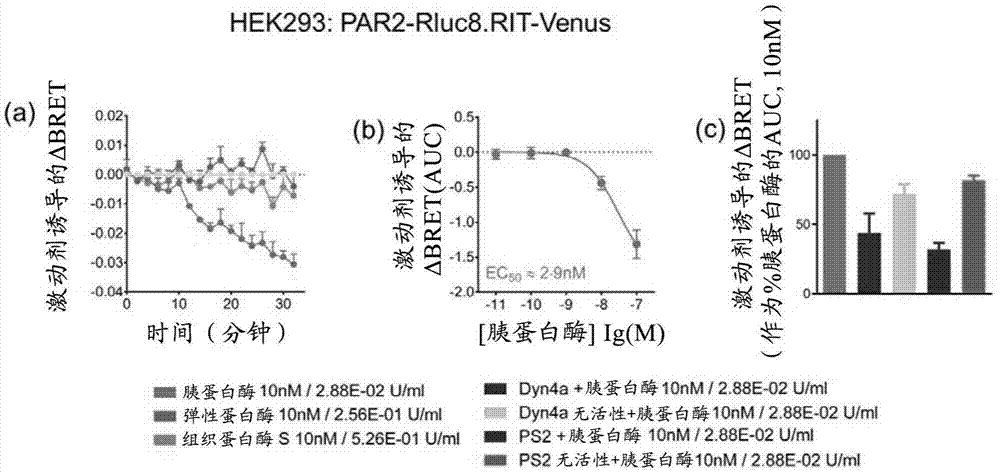
图1:用质膜标记物rit-venus进行bret测定。(a)由三种已知的par2激动剂刺激后的δbret动力学。(b)胰蛋白酶crc(由auc数据产生)。(c)用各种内吞抑制剂和对照化合物预处理30分钟后胰蛋白酶诱导的运输的变化。数据表示为来自至少三个独立实验的平均值±sem,每个实验每个条件包含三个重复。
图2:用早期核内体标记物rab5a-venus的bret测定。(a)由三种已知的par2激动剂刺激后的δbret动力学。(b)胰蛋白酶crc(由auc数据产生)。(c)用各种内吞抑制剂和对照化合物预处理30分钟后胰蛋白酶诱导的运输的变化。数据表示为来自至少三个独立实验的平均值±sem,每个实验每个条件包含三个重复。
图3:使用细胞溶质erk传感器cytoekar进行的fret测试。(a)由两种已知的par2激动剂刺激后的δfret动力学。(b)胰蛋白酶crc(由auc数据产生)。(c)用各种内吞抑制剂和对照化合物预处理30分钟后胰蛋白酶诱导的信号传导的变化。数据表示为来自至少三个独立实验的平均值±sem,每个实验每个条件包含三个重复。
图4:使用细胞核erk传感器nucekar进行的fret测试。(a)由两种已知的par2激动剂刺激后的δfret动力学。(b)胰蛋白酶crc(由auc数据产生)。(c)用各种内吞抑制剂和对照化合物预处理30分钟后胰蛋白酶诱导的信号传导的变化。数据表示为来自至少三个独立实验的平均值±sem,每个实验每个条件包含三个重复。
图5:bret再敏化测定。(a)由两种已知的par2激动剂刺激后的δbret动力学。(b)用各种信号传导抑制剂预处理30分钟后胰蛋白酶诱导的运输方式的变化。数据表示为来自至少三个独立实验的平均值±sem,每个实验每个条件包含两个重复。
图6.分离的drg神经元的神经元兴奋及其用par2内吞作用抑制剂的抑制。
图7.分离的drg神经元的神经元兴奋及其用par2内吞作用抑制剂的抑制。
图8.小鼠中胰蛋白酶诱发的机械性痛觉过敏的抑制。
图9.小鼠中胰蛋白酶诱发的机械性痛觉过敏的抑制。
发明详述
评估蛋白酶激活的受体-2(par2)的内吞作用的贡献,以在亚细胞区室中产生信号,其介导蛋白酶诱发的伤害感受器致敏。生物发光共振能量转移(bret)用于评估hek细胞中par2与质膜(kras)和核内体(rab5a)的驻留蛋白的接近程度。在r36↓s37断裂par2的典型激动剂胰蛋白酶增加par2和rab5a之间的bret并降低par2和kras之间的bret,与内吞作用一致。在e56↓t57和a66↓s67↓v68分别断裂par2的偏向性激动剂组织蛋白酶-s和弹性蛋白酶不诱导par2内吞作用。靶向质膜、细胞溶质或细胞核的
在一方面本发明提供治疗蛋白酶诱发的疼痛的方法,包括向需要的受试者给药抑制par2的核内体信号传导的化合物。
在一方面,所述抑制核内体par2信号传导的化合物为抑制激活的受体的内吞作用的化合物。
应当理解,抑制par2内吞作用的化合物可以在激活的par2的细胞内c-末端的磷酸化与随后的早期核内体受体的发动蛋白和网格蛋白依赖性内吞作用之间的途径中的任何位点或多个位点起作用。
在一个实施方案中,par2内吞作用的抑制可通过向需要的受试者给药β-抑制蛋白抑制剂而实现。如本文所述,术语“β-抑制蛋白抑制剂”表示抑制β-抑制蛋白和激活的par2的细胞内c-端之间的相互作用的化合物。
可以想到在一个实施方案中,抑制βarrs和激活的par2的细胞内c-端之间的相互作用可通过以下实现,向受试者给予β-抑制蛋白抑制剂,其通过提供gpcr激酶-2(grk2)磷酸化的替代位点在激活的par2的细胞内c-端与磷酸化位点竞争,从而减少或改善βarrs与激活的par2的细胞内c-端的结合和随后的受体内吞作用。
在另一实施方案中,可以想到抑制βarrs和激活的par2的细胞内c-端之间的相互作用可通过向受试者给予β-抑制蛋白抑制剂而实现,所述β-抑制蛋白抑制剂抑制激活的par2的细胞内c-端的gpcr激酶-2(grk2)的磷酸化。在一个实施方案中可以想到抑制激活的par2的细胞内c-端的gpcr激酶-2(grk2)磷酸化的β-抑制蛋白抑制剂直接与grk2相互作用,例如,通过在负责受体磷酸化的grk2的中心催化域结合。在另一个实施方案中,可以想到β-抑制蛋白抑制剂可以变构结合grk2,例如,以防止识别激活的par2的细胞内c-端上的磷酸化位点。在另一个实施方案中,可以想到β-抑制蛋白抑制剂将结合至par2的细胞内c-端内的磷酸化位点处或其附近,从而防止grk2的识别和磷酸化。还可以想到抑制βarrs与激活的par2的细胞内c-端之间相互作用的化合物可以通过直接与β-抑制蛋白相互作用起作用,抑制其与par2的细胞内c-末端上的磷酸化位点结合。
在另一实施方案中,par2内吞作用的抑制可通过向需要的受试者给药网格蛋白抑制剂而实现,以抑制激活的par2的网格蛋白-依赖性内吞作用。应理解网格蛋白抑制剂可为抑制激活的par2的网格蛋白-依赖性内吞作用的任何化合物,例如,选择性抑制与网格蛋白的末端结构域的配体缔合的化合物。多种网格蛋白抑制剂是本领域已知的,如pitstop1tm和pitstop2tm。
在另一实施方案中,par2内吞作用的抑制可通过以下实现,向需要的受试者给药发动蛋白抑制剂以抑制激活的par2的发动蛋白-或网格蛋白-依赖性内吞作用。应理解发动蛋白抑制剂可为任何抑制发动蛋白和随后的激活的par2的内吞作用的化合物,例如,抑制l-磷脂酰丝氨酸脂质体-刺激的螺旋发动蛋白的化合物,其中发动蛋白被诱导以围绕脂质体形成螺旋,抑制grb2-刺激的发动蛋白的化合物,或抑制发动蛋白自组装为单个环的化合物。多种发动蛋白抑制剂是本领域已知的,如dyngo4a和dynole2-24。
在另一实施方案中,par2内吞作用的抑制可通过以下实现,向需要的受试者给药显性失活β-抑制蛋白,其通过与内源性抑制蛋白竞争结合至网格蛋白而起作用,从而抑制激活的par2的网格蛋白-依赖性内吞作用。
在另一方面,所述抑制核内体par2信号传导的化合物为靶向和抑制核内体par2信号传导的化合物。
在一个实施方案中,抑制核内体par2信号传导的化合物为式(i)的三部分化合物:
其中
a为脂质锚,其促进化合物插入质膜;
l为长度为1nm至50nm的连接基部分;且
x为核内体par2信号传导的抑制剂;
其中该脂质锚分配到在4℃不溶于非离子型洗涤剂的脂质膜中;或
其药学上可接受的盐。
本文使用的术语“三部分化合物”是指包含与连接基共价结合的核内体gpcr调节剂的化合物,该连接基共价结合至脂质锚,其能够将式(i)化合物锚定在细胞膜的脂质双层上并最终锚定至早期核内体的膜。
本文使用的术语“脂质锚”表示能够分配到脂质膜中并由此将式(i)化合物锚定在脂质膜中的部分。分配到脂质膜中可以直接从细胞外或囊泡腔间隙发生,或者可以从脂质双层侧向发生。脂质锚可以通过其分配到脂质膜中的能力来表征,由此所述脂质膜的特征在于在4℃下在非离子洗涤剂中的不溶性。合适的脂质锚的实例包括但不限于胆固醇,胆甾烷醇,鞘脂,gpi-锚或饱和脂肪酸衍生物。本领域已经描述了许多这样的脂质锚,例如,在wo2005/097199中,其全部内容通过引用并入本文。
在一个实施方案中,脂质锚为选自式(ii)或(iii)的部分:
其中
r1为任选取代的c1-12烷基;
r2和r3独立地为h或c1-3烷基;
r4为-ch2-、-o-、-nh-、-s-、-nh(ch2)aopo3--、-nh(ch2)aso2cf2-、-nh(ch2)aso2nh-、-nhconh-、-nhc(o)o-、-nhch(conh2)(ch2)bc(o)o-、-nhch(cooh)(ch2)bc(o)o-、-nhch(conh2)(ch2)bconh-、-nhch(cooh)(ch2)bconh-、-nhch(conh)(ch2)4nh((co)ch2o)e-或-nhch(cooh)(ch2)4nh((co)ch2o)e-;
a为整数2至3;
b为整数1至2;
e为整数0至1;且
在其它实施方案中所述脂质锚为选自式(vi)、(vii)、(viii)或(ix)的部分:
其中
r4如上所述;
r5的每种情况独立地为-nh-、-o-、-s-、-oc(o)-、-nhc(o)-、-nhconh-、-nhc(o)o-或-nhs(o2)-;
r6的每种情况独立地为任选取代有氟、优选1至4个氟原子的c14-30烷基;
r7的每种情况独立地为nh2、nhch3、oh、h、卤素或o,条件是当r7为nh2、nhch3、oh、h或卤素时则
r8的每种情况独立地为h、oh或当
r9为任选取代有氟、优选1至4个氟原子的c10-30烷基;且
r10的每种情况独立地为任选取代有氟、优选1至4个氟原子的c24-40亚烷基、c24-40亚烯基或c24-40亚炔基。
在其它实施方案中,脂质锚为选自式(x)或(xi)的部分:
其中
r13的每种情况独立地为-o-或-co(ch2)a(co)bo-,其中a为整数1至3且b为整数0至1;
r14为-o-或-oc(o)-;
r15的每种情况独立地选自任选取代有氟、优选1至4个氟原子的c16-30烷基;
r16为-po3-ch2-、-so3ch2-、-ch2-、-co2ch2-或直接键;
r17为-nh-、-o-、-s-、-oc(o)-、-nhc(o)-、-nhconh-、-nhc(o)o-或-nhs(o2)-;
r18为nh2、nhch3、oh、h、卤素或o;
r19为任选取代有氟、优选1至4个氟原子的c16-30烷基;且
各r20为任选取代有下式的基团的c(o)c13-25烷基:
其中
r21为-po3-ch2-、-so3ch2-、-ch2-、-co2ch2-或直接键;
r22为-nh-、-o-、-s-、-oc(o)-、-nhc(o)-、-nhconh-、-nhc(o)o-或-nhs(o2)-;
r23为-o-或-oc(o)-;
r24的每种情况独立地选自任选取代有氟、优选1至4个氟原子的c16-30烷基;
r25为-co(ch2)a(co)bo-或-co(ch2)a(co)bnh-,其中a为整数1至3且b为整数0至1;且
r26为任选取代有氟、优选1至4个氟原子的c4-20烷基。
在其它实施方案中脂质锚为选自式(xii)、(xiii)、(xiv)或(xv)的部分:
其中
r27的每种情况独立地选自-nh-、-o-、-nh(ch2)copo3--、-nh(ch2)cso2nh-、-nhconh-、-nhc(o)o-、-co(ch2)b(co)anh-、-co(ch2)b(co)ao-、-co(ch2)bs-、-co(ch2)bopo3--、-co(ch2)bso2nh-、-co(ch2)bnhconh-、-co(ch2)boconh-、-co(ch2)boso3-或-co(ch2)bnhc(o)o-,其中a为整数0至1,b为整数1至3且c为整数2至3;
r28的每种情况独立地为-ch2-或-o-;
r29的每种情况独立地选自h或任选取代有氟、优选1至4个氟原子的c16-30烷基;
r31的每种情况独立地选自h,或任选取代有氟、优选1至4个氟原子的c1-15烷基,或任选取代有氟、优选1至4个氟原子的c1-15烷氧基;且
n为整数1至2。
本文使用的术语“连接基”涉及将核内体gpcr的调节剂连接至脂质锚的化合物的部分。应该理解连接基应该被选择为使其不与核内体gpcr的调节剂在配体结合位点竞争。连接基也不应该分配到脂质膜中。连接基应当具有1nm至50nm的长度,以便当脂质锚锚定在核内体膜中时,允许核内体gpcr的调节剂与受体相互作用。在一个实施方案中,连接基将包含一个或多个聚乙二醇单元。在另一个实施方案中,预期连接基或连接基的亚基可以是氨基酸残基、衍生化或官能化的氨基酸残基、聚醚、脲、氨基甲酸酯、磺酰胺或其他亚单位,其在核内体gpcr的调节剂和脂质锚之间提供足够的距离而不干扰任一基团的功能。
在一个实施方案中,所述连接基通过式(iv)的部分表示:
其中
z为连接基和脂质锚之间的附接基团且为–c1-c10烷基-、-c2-c10烯基-、-c2-c10炔基-、–c1-c10烷基c(o)-、-c2-c10烯基c(o)–或-c2-c10炔基c(o)-;或
z,与相邻的胺一起,为任选c-末端酰胺化的氨基酸,所述氨基酸选自天冬氨酸、谷氨酸、天冬酰胺、谷氨酰胺、组氨酸、半胱氨酸、赖氨酸、精氨酸、丝氨酸或苏氨酸;其中所述氨基酸通过其侧链官能团连接至脂质锚;
y为连接基和核内体gpcr的调节剂之间的附接基团且为-o-、-nh-、-s-、-c(o)-、-c(o)nh-、-c(o)o-或-c(o)s-;或
y,与相邻的酰胺基一起,为氨基酸,所述氨基酸选自天冬氨酸、谷氨酸、天冬酰胺、谷氨酰胺、组氨酸、半胱氨酸、赖氨酸、精氨酸、丝氨酸或苏氨酸;其中所述氨基酸通过其侧链官能团连接至核内体gpcr的调节剂;
m为1或2;
n为1至20;且
p为1至8。
在另一实施方案中,所述连接基通过式(xx)的部分表示:
其中
r11的每种情况独立地为天然存在的、衍生的或官能化的氨基酸残基的任何侧链;
m为整数3至80;且
n为整数0至1。
在其它实施方案中,所述连接基通过式(xxi)的部分表示:
其中
m为整数0至40;
n为整数0至1;
o的每种情况独立地为整数1至5;
r11的每种情况独立地为天然存在的、衍生的或官能化的氨基酸残基的任何侧链;且
其中所述so2端结合至脂质锚且该n-端结合至核内体gpcr的调节剂。
在另一实施方案中,所述连接基通过式(xxii)的部分表示:
其中
m为整数0至40;
n为整数0至1;
o的每种情况独立地为整数1至5;
各r12独立地为nh或o;
r11的每种情况独立地为天然存在的、衍生的或官能化的氨基酸残基的任何侧链;且
其中所述c(o)-端结合至脂质锚且r12-端结合至核内体gpcr的调节剂。
多种合适的连接基部分已描述于wo2005/097199,将其整体在此引入作为参考。
本文使用的术语“核内体par2信号传导的抑制剂”是指已内吞至核内体中的par2的拮抗剂或抑制剂。核内体par2信号传导的抑制剂可为任何形式,包括,但不限于,有机分子、多肽序列、激素、任何这些的蛋白质片段或衍生物。
本文所用的术语“核内体par2信号传导”是指由已被内吞入核内体(优选地早期核内体)的激活的par2转导的信号。
在一个实施方案中,核内体par2信号传导将是首先在质膜上转导并在受体被内吞入早期核内体时维持的信号传导。
在另一实施方案中,所述核内体par2信号传导将是需要受体内吞作用和/或仅在核内体膜上发生的信号传导,例如β-抑制蛋白介导的信号传导。据信,βarrs与激动剂占据的g蛋白偶联受体激酶(grk)-磷酸化的gpcr在细胞表面相互作用,并通过发动蛋白-和网格蛋白-依赖性内吞作用促进配体结合的受体从细胞表面转移至早期核内体中。最近已发现,该途径可介导第二系列的核内体gpcr信号传导,该信号传导不同于质膜上的g蛋白依赖性信号传导。据信这种机制的重要性取决于gpcr与βarrs相互作用的亲和力,其取决于通过grk的gpcr磷酸化程度而变化。“a类”gpcr(例如,β2ar、α1bar)具有很少的磷酸化位点,并且主要在质膜处与βarr1和βarr2短暂相互作用,对βarr2具有更高的亲和力。“b类”gpcr(例如at1ar、nk1r、par2)在多个位点被磷酸化,并在血浆和核内体膜上长时间以高亲和力与βarr1和2两者相互作用。“c类”gpcr(例如缓激肽b2受体)用βarrs内化到核内体中,然后在除去激动剂后迅速解离。
据信,βarr诱导的mapk信号传导的程度取决于受体对βarr的亲和力,该亲和力取决于受体结构以及七种哺乳动物grk中的哪一种磷酸化该受体。因此,at1ar和v2r的激活导致比激活α1bar和β2ar引起更大的βarr结合的erk1/2的磷酸化,这表明b类受体更强烈地通过该途径进行信号传导。如上所述,par2为b类gpcr。
在本发明一些优选实施方案中,关于通式(i),适用以下一个或多个实施方案:
a)a为选自胆固醇、胆甾烷醇、鞘脂类、gpi-锚或饱和脂肪酸衍生物的脂质锚。
b)a为选自式(ii)、(iii)、(vii)、(viii)、(ix)、(x)、(xi)、(xii)、(xiii)和(ix)的部分的脂质锚。
c)a为选自式(ii)或(iii)的部分的脂质锚。
d)l是包含一个或多个亚基的连接基,所述亚基包含聚乙二醇单元、氨基酸残基、衍生或官能化的氨基酸残基、聚醚、脲、氨基甲酸酯和/或磺酰胺。
e)l为式(iv)、(xx)、(xxi)或(xxii)表示的连接基部分。
f)l为式(iv)表示的连接基部分。
在一个优选实施方案中,a为式(ii)或(iii)表示的脂质锚。
因此,在一个实施方案中,本发明提供式(ia)表示的式(i)的三部分化合物:
其中
a为脂质锚,其促进化合物插入式(ii)或(iii)表示的质膜中:
其中
r1为任选取代的c1-12烷基;
r2和r3独立地为h或c1-3烷基;
r4为c、o、nh或s;且
l为长度为1nm至50nm的连接基;且
x为核内体par2信号传导的抑制剂;或
其药学上可接受的盐。
在另一优选实施方案中,a为式(ii)或(iii)表示的脂质锚且l为式(iv)表示的连接基。
因此,在另一实施方案中,本发明提供式(ib)表示的式(i)的三部分化合物:
其中
a为脂质锚,其促进化合物插入式(ii)或(iii)表示的质膜中:
其中
r1为任选取代的c1-12烷基;
r2和r3独立地为h或c1-3烷基;
r4为c、o、nh或s;
l通过式(iv)表示:
其中
z为连接基和脂质锚之间的附接基团且为–c1-c10烷基–、–c2-c10烯基-、-c2-c10炔基-、–c1-c10烷基c(o)-、-c2-c10烯基c(o)–或-c2-c10炔基c(o)-;或
z,与相邻的胺一起,为任选c-末端酰胺化的氨基酸,所述氨基酸选自天冬氨酸、谷氨酸、天冬酰胺、谷氨酰胺、组氨酸、半胱氨酸、赖氨酸、精氨酸、丝氨酸或苏氨酸;其中所述氨基酸通过其侧链官能团连接至脂质锚;
y为连接基和核内体gpcr的调节剂之间的附接基团且为-o-、-nh-、-s-、-c(o)-、-c(o)nh-、-c(o)o-或-c(o)s-;或
y,与相邻的酰胺基一起,为氨基酸,所述氨基酸选自天冬氨酸、谷氨酸、天冬酰胺、谷氨酰胺、组氨酸、半胱氨酸、赖氨酸、精氨酸、丝氨酸或苏氨酸;其中所述氨基酸通过其侧链官能团连接至核内体gpcr的调节剂;
m为1或2;
n为1至20;
p为1至8;且
x为核内体par2信号传导的抑制剂;或
其药学上可接受的盐。
在一个实施方案中,式(i)的化合物具有以下结构:
其中l和a如本文所定义,或其药学上可接受的盐。
在另一方面,本发明提供化合物:
或其药学上可接受的盐。
在一方面,本发明提供治疗蛋白酶诱发的疼痛的方法,包括向需要的受试者给药有效量的本文定义的式(i)的化合物。
在另一方面,本发明提供本文定义的式(i)的化合物在制备用于治疗蛋白酶诱发的疼痛的药物中的用途。
在另一方面,本发明提供本文定义的式(i)的化合物,其用于治疗蛋白酶诱发的疼痛。
在另一方面,本发明提供治疗蛋白酶诱发的疼痛的方法,包括向需要的受试者给药包含一种或多种抑制受体的内吞作用的化合物和一种或多种靶向和抑制核内体par2信号传导的化合物的组合。
在本发明的范围内,术语“疼痛”包括慢性炎性疼痛(例如与类风湿关节炎、骨关节炎、类风湿性脊柱炎、痛风性关节炎和青少年关节炎有关的疼痛);肌肉骨骼痛,下背部和颈部疼痛,扭伤和拉伤,神经性疼痛,交感神经维持性疼痛(sympatheticallymaintainedpain),肌炎,与癌症和纤维肌痛相关的疼痛,与偏头痛相关的疼痛,与丛集性和慢性每日头痛相关的疼痛,与流感或其它病毒感染如普通感冒、风湿热相关的疼痛,与功能性肠紊乱相关的疼痛如非溃疡性消化不良、非心脏性胸痛和肠易激综合征,与心肌缺血相关的疼痛,手术后疼痛,头痛,牙痛,痛经,神经痛,纤维肌痛综合征,复杂性局部疼痛综合征(crpsi型和ii型),神经性疼痛综合征(包括糖尿病性神经病、化学治疗诱导的神经性疼痛、坐骨神经痛、非特异性下背痛、多发性硬化性疼痛、hiv相关性神经病、疱疹后神经痛、三叉神经痛)以及由物理创伤、截肢术、癌症、毒素或慢性炎症引起的疼痛。在一个优选的实施方案中,所述疼痛是驱体痛或内脏痛。
合成本发明的化合物的一般策略如下所述。
文献中描述了胆甾醇乙醇酸、3-胆甾胺和胆甾醇甘氨酸的合成(hussey,s.l.等人,j.am.chem.soc.2001,123,12712-12713;hussey,s.l.等人,org.lett.2002,4,415-418;martin,s.e.等人,bioconjugatechem.2003,14,67-74)。在类固醇结构的3位上具有酰胺、磺酰胺、脲或氨基甲酸酯官能团的式(iii)的脂质锚可以由3-胆甾胺制备,例如3-胆甾胺可以与琥珀酸酐在dmap存在下反应,得到相应的琥珀酰基取代的化合物。相应的磺酰胺可以通过使3-胆甾胺与氯磺酰乙酸反应获得,其可以如文献中所述制备(hinman,r.l.和locatell,l.j.am.chem.soc.1959,81,5655-5658)。相应的脲或氨基甲酸酯可以根据文献方法通过相应的异氰酸酯来制备(knolker,h.-j.t.等人,angew.chem.int.ed.1995,34,2497;knolker,h.-j.等人,synlett1996,502;.knolker,h.-j.和braxmeier,t.tetrahedronlett.1996,37,5861)。在类固醇结构的3位具有磷酸酯或羧甲基化磷酸酯的化合物(iii)的中间体可以如文献中所述制备(golebriewski,keyes,cushman,bioorg.med.chem.1996,4,1637-1648;cusinato,habeler,等人,j.lipidres.1998,39,1844-1851;himber,missano,等人,j.lipidres.1995,36,1567-1585)。在类固醇结构的3位具有巯基的式(iii)脂质锚可以如文献中所述制备(j.g.parkes,j.g.等人,biochim.biophys.acta1982,691,24-29),相应的羧甲基化硫醇可通过如对于相应的胺和醇所述的简单烷基化获得。在类固醇结构的3位上具有二氟亚甲基砜衍生物的式(iii)的脂质锚可以如文献中所述制备(lapiene,j.等人,bioorg.med.chem.lett.2004,14,151-155)。在式(iii)脂质锚的17位引入各种侧链可以通过使用从去氢异雄酮或孕烯醇酮开始的文献方法来实现(bergmann,e.d.等人,j.am.chem.soc.1959,81,1239-1243和其中的参考文献)。由胆甾烷衍生的式(iii)脂质锚可由相应的前体得到,该前体通过使用文献中的方案(例如在各种过渡金属催化剂存在下氢化)由胆固醇还原5,6-双键得到。
在3位上具有氧衍生的取代基的式(ii)脂质锚按照与对于式(iii)脂质锚所述相似的方式由雌酮制备。在3位上具有氮衍生的取代基的式(ii)脂质锚可按照与对于式(iii)脂质锚所述相似的方式由3-氨基雌酮制备,其可以如文献中所述制备(zhang,x.和sui,z.tetrahedronlett.2003,44,3071-3073;woo,l.w.l.等人,steroidbiochem.molec.biol.1996,57,79-88)。在3位上具有硫衍生的取代基的式(ii)脂质锚可按照与对于式(iii)脂质锚所述相似的方式由3-巯基雌酮(3-thioestrone)制备,其可以如文献中所述制备(woo,l.w.l.等人,j.steroidbiochem.molec.biol.1996,57,79-88)。如文献中所述,在雌酮结构的17位引入各种侧链可以通过魏悌希(wittig)方法,接着氢化所得的双键来实现(peters,r.h.等人,j.org.chem.1966,31,24-26)。通过标准方案(suzuki偶联等)可以实现侧链内的其他操作(例如双键构建、环烷基修饰)。
具有不同烃基的属于神经酰胺类、去氢神经酰胺类和二氢神经酰胺类的式(vi)脂质锚可如文献中所述获得(a.h.merrill,jr.,y.a.hannun(eds.),methodsinenzymology,vol.311,academicpress,1999;.koskinen,p.m和koskinen,a.m.p.synthesis1998,1075)。特别地,鞘氨醇碱可以用作在鞘氨醇主链的1位上具有氧衍生的取代基的式(vi)的所有脂质锚的关键中间体。相应的氨基衍生物可以通过磺酸酯的取代来获得,该磺酸酯可以根据已知的方案由醇制备。1-氨基或1-羟基衍生物的烷基化和酰化可分别通过与溴乙酸和琥珀酸酐反应来实现。该硫代乙酰化衍生物可以通过用巯基乙酸取代磺酸酯来制备。磷酸酯和硫酸酯衍生物可按文献所述获得(a.h.merrill,jr.,yaa.hannun(eds.),methodsinenzymology,vol.311,academicpress,1999;koskinen,p.m.和koskinen,a.m.p.synthesis1998,1075)。酰化、磺酰化、脲和氨基甲酸酯的形成可以通过标准过程来实现。式(vi)脂质锚(其中r5为氨基或氨基衍生的官能团)可使用标准方案由鞘氨醇碱来制备,所述鞘氨醇碱如koskinen公开的那样来获得(koskinen,p.m.和koskinen,a.m.p.synthesis1998,1075)。相应的2-氧取代的鞘脂类可以通过yamanoi公开的策略获得(yamanoi,t.等人,chem.lett.1989,335)。式(vi)脂质锚(其中两个r8都代表羟基)可以通过使用已知方案对相应的烯烃进行双羟基化来获得。相应的单羟基衍生物可以如文献中所述制备(howell,a.r.和ndakala,a.j.curr.org.chem.2002,6,365-391)。式(vi)脂质锚中的取代基r6和r9的修饰可以通过各种综述文章中概述的方案和策略来实现(harwood,h.j.chem.rev.1962,62,99-154;gensler,w.j.chem.rev.1957,57,191-280)。
式(vii)脂质锚可通过文献中描述的方案(
式(viii)脂质锚(其中r4和r5为氧衍生的取代基)可以由市售可得的(r)-(–)-2,2-二甲基-1,3-二氧戊环-4-甲醇开始制备,如fraser-reid中所概述的那样(schlueter,u.lu,j.和fraser-reid,b.org.lett.2003,5,255-257)。式(viii)化合物中的取代基r6的变化可以通过各种综述文章中概述的方案和策略来实现(harwood,h.j.chem.rev.1962,62,99-154;gensler,w.j.chem.rev.1957,57,191-280)。式(viii)脂质锚(其中r4和r5为氮衍生的取代基)可以如下获得:由相应的氧取代的体系开始,通过相应的磺酸酯的亲核取代和如上所述的进一步修饰,或者由如文献中所述可获得的1,2,3-三氨基丙烷开始(henrick,k.等人,jchem.soc.daltontrans.1982,225-227)。
式(ix)脂质锚可通过与式(vii)脂质锚类似的方式获得,或者通过使式(viii)脂质锚的ω-乙烯基化的中间体进行闭环复分解获得。
式(x)和(xi)的脂质锚可以通过文献中描述的合成方案(xue,j.和guo,z.bioorg.med.chem.lett.2002,12,2015-2018;xue,j.和guo,z.j.am.chem.soc.2003,16334-16339;xue,j.等人,j.org.chem.2003,68,4020-4029;shao,n.,xue,j.和guo,z.angew.chem.int.ed.2004,43,1569-1573)以及通过它们与上述制备式(vi)和(viii)的脂质锚的方法的组合来获得。
式(xii)、(xiii)和(xiv)的脂质锚可通过文献中描述的下列合成方案的全合成获得(knolker,h.-j.chem.soc.rev.1999,28,151-157;knolker,h.-j.和reddy,k.r.chem.rev.2002,102,4303-4427;knolker,h.-j.和knoll,j.chem.commun.2003,1170-1171;knolker,h.-j.curr.org.synthesis2004,1)。
式(xv)的脂质锚可以通过nenitzescu-型吲哚合成由4-甲氧基-3-甲基苯甲醛开始制备,得到6-甲氧基-5-甲基吲哚。醚断裂、三氟甲磺酸酯形成和sonogashira偶联导致得到相应的6-炔基取代的5-甲基吲哚。nilsmeier甲酰化反应和随后的硝基甲烷加成反应生成3-硝基乙烯基取代的吲哚衍生物,其经全面氢化,导致形成6-烷基取代的5-甲基色胺。使用琥珀酸酐对氨基进行酰化完成该制备。
已知的固相或溶液相技术可以用于合成本发明的肽,例如将n-或c-末端偶联至固体载体(通常为树脂),然后逐步合成线性肽。用于保护氨基酸残基(包括侧链)的保护基团化学是本领域公知的,并且可以在例如:theodoraw.greene和peterg.m.wuts,protectinggroupsinorganicsynthesis(thirdedition,johnwiley&sons,inc,1999)中发现,其全部内容通过引用并入本文。
用于制备如本文所述的化合物的方法对于本领域技术人员将是显而易见的,并且将包括以下步骤:a)确定脂质锚的一个或多个磷酰基头基或等效头基与par2信号传导的抑制剂的结合和/或相互作用位点之间的距离;b)选择能够横跨(a)中定义的距离的连接基;和c)通过(b)中选择的连接基键合脂质锚和核内体par2信号传导的抑制剂。
本文给出了这种方法的相应操作实例。本领域技术人员能够推导给定的或潜在的核内体par2信号传导的抑制剂的相关结合位点或相互作用位点,并因此确定(a)脂质锚的一个或多个磷酰基头基或等效头基与核内体par2信号传导的抑制剂的结合和/或相互作用位点之间的距离。这些方法包括但不限于分子建模、体外和/或分子相互作用或结合测定(例如酵母双杂交系统或三杂交系统、肽点样(peptidespotting)、覆盖测定、噬菌体展示、细菌展示、核糖体展示),原子力显微镜以及光谱学方法和x射线晶体学。此外,可采用诸如定点诱变的方法来验证给定核内体par2信号传导的抑制剂或候选核内体par2信号传导的抑制剂及其相应靶标的推导的相互作用位点。
本领域技术人员将理解,连接基的选择包括本领域已知的连接基的选择以及新连接基的生成和使用,例如通过分子建模和相应的合成或本领域已知的其他方法。
文中所用的参考步骤b)中的术语“横跨”指的是当脂质锚形成核内体脂质层的一部分时,选择用于将核内体par2信号传导的抑制剂置于受体上的正确位点的连接基的长度。
本领域技术人员能够容易地推导、验证和/或评估给定的三部分化合物以及本文所述的单个部分的亲脂性。本文在实施例中提供了用于确定核内体gpcr靶向作用的相应测试分析。
本领域技术人员将理解,连接基的目的是将脂质锚连接至核内体par2信号传导的抑制剂,以便当脂质锚锚定到核内体膜中时,使得核内体par2信号传导的抑制剂与par2相互作用。该脂质锚和连接基将含有允许两者共价键合的官能团。脂质锚的官能团的性质绝不受限制,并且可以包括例如与连接基形成酰胺键的胺基,或与连接基形成醚键或酯键的羟基或羧酸基。
类似地,本领域技术人员将理解,与核内体par2信号传导的抑制剂相连的连接基末端的官能团的选择主要由选择的核内体par2信号传导的抑制剂上任何可用的官能团决定。例如,如果核内体par2信号传导的抑制剂包含游离胺或羧酸基团,则可以预期连接基的官能团将包含互补的羧酸或胺以形成酰胺键。
应该理解,本发明的化合物可以以一种或多种立体异构形式(例如非对映异构体)存在。本发明在其范围内包括所有这些立体异构形式,其或者是分离的形式(例如,在对映异构体分离中)或组合的形式(包括外消旋混合物和非对映异构体混合物)。本发明预期使用l和d形式的氨基酸,包括使用独立选自l和d形式的氨基酸,例如,其中所述肽包含两个丝氨酸残基,每个丝氨酸残基可以具有相同的或相反的绝对立体化学。除非另有说明,所述氨基酸被认为是l-构型。
因此本发明还涉及相对于氨基酸残基的不对称中心而言基本纯的立体异构体形式的化合物,例如大于约90%de,例如约95%至97%de或大于99%de的化合物及其混合物,包括其外消旋混合物。这样的非对映异构体可以通过不对称合成(例如使用手性中间体)来制备,或者可以通过常规方法如色谱法或使用拆分剂来拆分混合物。
当本发明化合物需要纯化时,可以使用色谱技术如高效液相色谱(hplc)和反相hplc。这些肽可以用质谱法和/或其他适当的方法来表征。
当该化合物包含一个或多个可被质子化或去质子化的官能团(例如在生理ph下)时,该化合物可被制备和/或分离为药学上可接受的盐。应该理解,该化合物在给定的ph下可以是两性离子。如本文所用的表述“药学上可接受的盐”是指给定化合物的盐,其中所述盐适合作为药物给药。该盐可以例如通过分别使酸或碱与胺或羧酸基团反应而形成。
药学上可接受的酸加成盐可以由无机酸和有机酸制备。无机酸的实例包括盐酸、氢溴酸、硫酸、硝酸、磷酸等。有机酸的实例包括乙酸、丙酸、乙醇酸、丙酮酸、草酸、苹果酸、丙二酸、琥珀酸、马来酸、富马酸、酒石酸、柠檬酸、苯甲酸、肉桂酸、扁桃酸、甲磺酸、乙磺酸、对甲苯磺酸、水杨酸等。
药学上可接受的碱加成盐可以由无机碱和有机碱制备。衍生自无机碱的相应抗衡离子包括钠盐、钾盐、锂盐、铵盐、钙盐和镁盐。有机碱包括伯胺、仲胺和叔胺、取代的胺包括天然存在的取代的胺和环状胺、包括异丙胺、三甲胺、二乙胺、三乙胺、三丙胺、乙醇胺、2-二甲基氨基乙醇、氨丁三醇、赖氨酸、精氨酸、组氨酸、咖啡因、普鲁卡因、海巴明、胆碱、甜菜碱、乙二胺、葡糖胺、n-烷基葡萄糖胺、可可碱、嘌呤、哌嗪、哌啶和n-乙基哌啶。
酸/碱加成盐倾向于比相应的游离酸/碱形式更易溶于含水溶剂。
本发明还提供药物组合物,其包含上文定义的治疗有效量的化合物或其药学上可接受的盐以及至少一种药学上可接受的载体或稀释剂。
术语“组合物”旨在包括以包封材料为载体的活性成分的制剂,得到活性成分(有或没有其他载体)被载体包围的胶囊。
尽管如上所述的化合物或其药学上可接受的盐可以是给予受试者的唯一活性成分,但其他一种或多种活性成分与所述化合物一起给药也在本发明的范围内。在一个或多个实施方案中,预期将两种或更多种本发明化合物的组合给药于受试者。预期该一种或多种化合物也可以与一种或多种另外的治疗剂组合给药。该组合可以允许如上所述的一种或多种化合物与其他活性成分分开、顺序或同时给药。该组合可以以药物组合物的形式提供。
如本文所用的术语“组合”是指组合物或试剂盒套件(kitofparts),其中如上定义的组合搭档可以依赖性地或独立地或通过使用与特定量的组合搭档的不同的固定组合(即同时或在不同的时间点)进行给药。然后可将该组合搭档例如同时或按时间顺序交叉给药(对于该试剂盒套件的任何部分,其在不同时间点且具有相同或不同的时间间隔给药)。在组合中施用的组合搭档的总量的比例可以变化,例如,为了应对要治疗的患者亚群的需求或患者的年龄、性别、体重等不同需求可能引起的单个患者的需求。
如本领域技术人员将容易理解的,给药途径和药学上可接受的载体的性质将取决于病症的性质和待治疗的哺乳动物。据信本领域技术人员可以容易地确定特定载体或递送系统的选择和给药途径。在制备含有活性化合物的任何制剂时,应注意确保化合物的活性在该过程中不被破坏,并且化合物能够到达其作用位点而不被破坏。在某些情况下,可能需要通过本领域已知的手段(例如微包封)来保护化合物。类似地,所选择的给药途径应该使得该化合物到达其作用位点。
本领域技术人员可以使用常规方法容易地确定本发明化合物的合适制剂。鉴定优选的ph范围和合适的赋形剂,例如抗氧化剂,在本领域中是常规的。通常使用缓冲系统来提供所需范围的ph值,并且所述缓冲系统包括羧酸缓冲液例如乙酸盐,柠檬酸盐,乳酸盐和琥珀酸盐。多种抗氧化剂可用于这些制剂,包括酚化合物如bht或维生素e、还原剂如甲硫氨酸或亚硫酸盐,以及金属螯合剂如edta。
如上所述的化合物或其药学上可接受的盐可以以肠胃外剂型制备,包括适合于静脉内、鞘内和脑内或硬膜外递送的那些。适于注射使用的药物剂型包括无菌注射溶液或分散体,以及用于即时制备无菌注射溶液的无菌粉末。它们在制造和储存条件下应该是稳定的,并且可以防止还原或氧化以及微生物如细菌或真菌的污染作用。
用于可注射溶液或分散体的溶剂或分散介质可含有用于活性化合物的任何常规溶剂或载体体系,并可含有例如水、乙醇、多元醇(例如甘油、丙二醇和液体聚乙二醇等)、其合适的混合物和植物油。例如,通过使用诸如卵磷脂的包衣,通过在分散的情况下维持所需的粒度和通过使用表面活性剂来维持适当的流动性。可以在必要的情况下通过包含各种抗细菌剂和抗真菌剂,例如对羟基苯甲酸酯、三氯叔丁醇、苯酚、山梨酸、硫柳汞等,防止微生物的作用。在许多情况下,优选包括调节摩尔渗透压浓度的试剂,例如糖或氯化钠。优选地,用于注射的制剂将与血液等渗。可以通过在组合物中使用延迟吸收的试剂例如单硬脂酸铝和明胶来延长可注射组合物的吸收。适于注射使用的药物形式可以通过任何适当的途径递送,包括静脉内、肌肉内、大脑内、鞘内、硬膜外注射或输注。
通过将需要量的本发明化合物与所需的各种其他成分(如上述列举的那些)掺入合适的溶剂中,然后过滤灭菌来制备无菌注射溶液。通常,通过将各种灭菌的活性成分掺入含有碱性分散介质和来自上面列举的那些所需其他成分的无菌媒介物中来制备分散体。在用于制备无菌注射溶液的无菌粉末的情况下,优选的制备方法是真空干燥或冷冻干燥活性成分和任何额外的所需成分的先前无菌过滤的溶液。
其他药物形式包括本发明的口服和肠内制剂,其中所述活性化合物可以与惰性稀释剂或与可吸收的可食用载体一起配制,或者其可以包封在硬或软壳明胶胶囊中,或者可以将其压缩成片剂,或者它可以直接与饮食食物结合。对于口服治疗给药,该活性化合物可以与赋形剂混合并以可摄入片剂、含服或舌下片剂、锭剂、胶囊、酏剂、混悬剂、糖浆剂、薄片剂(wafer)等形式使用。这种治疗上有用的组合物中活性化合物的量是可以获得合适剂量的量。
片剂、锭剂、丸剂、胶囊剂等也可以含有如下所列的组分:粘合剂如树胶、阿拉伯树胶、玉米淀粉或明胶;赋形剂如磷酸二钙;崩解剂如玉米淀粉、马铃薯淀粉、海藻酸等;润滑剂如硬脂酸镁;并可加入甜味剂如蔗糖、乳糖或糖精或调味剂如薄荷、冬青油或樱桃调味剂。当剂量单位形式是胶囊时,除了上述类型的材料之外,它还可以含有液体载体。各种其他材料可作为包衣存在或用于另外改变剂量单位的物理形式。例如,片剂、丸剂或胶囊剂可以涂覆以虫胶、糖或两者。糖浆或酏剂可含有活性化合物、作为甜味剂的蔗糖、作为防腐剂的对羟基苯甲酸甲酯和对羟基苯甲酸丙酯、染料和调味剂如樱桃或橙味调味剂。当然,用于制备任何剂量单位形式的任何材料应该是药用纯的并且在使用量上基本无毒。另外,本发明的化合物可以掺入缓释剂型和制剂中,包括允许将活性肽特异性递送至肠的特定区域的那些剂型和制剂。
液体制剂也可以通过胃或食管气管引导管(oesophagealtube)经肠内给药。肠内制剂可以通过与适宜的基质例如乳化基质或水溶性基质混合而以栓剂的形式制备。本发明化合物还可以(但不是必需)局部、鼻内、阴道内、眼内等给药。
药学上可接受的媒介物和/或稀释剂包括任何和所有溶剂、分散介质、包衣、抗细菌剂和抗真菌剂、等渗剂和吸收延迟剂等。这些用于药物活性物质的介质和药剂的用途在本领域中是公知的。除非任何常规介质或试剂与活性成分不相容,还预期将其用于治疗组合物中。也可以将补充活性成分掺入该组合物中。
配制剂量单位形式的组合物以易于给药和剂量的均匀性是特别有利的。如本文所用的剂量单位形式是指适合作为待治疗的哺乳动物受试者的单位剂量的物理离散单位;每个单位含有经计算可产生所需治疗效果的预定量的活性物质以及所需的药学上可接受的载体。本发明的新剂量单位形式的规格取决于并且直接依赖于(a)活性物质的独特特征和待实现的具体治疗效果,和(b)用于治疗疾病的活性物质配药领域中固有的限制,所述活性物质用于治疗患有身体健康受损的疾病的活体受试者中的疾病,如本文中详细公开的那样。
如上所述,可以将主要活性成分与适合的药学上可接受的媒介物以剂量单位形式混合用于以治疗有效量方便且有效地给药。例如,单位剂型可以包含0.25μg至约2000mg量的主要活性化合物。以比例表示,活性化合物可以存在于约0.25μg至约2000mg/ml的载体中。在含有补充活性成分的组合物的情况下,通过参考所述成分的常规剂量和给药方式来确定剂量。
如本文所用的术语“有效量”是指当根据期望的给药方案施用时提供期望的治疗活性的化合物的量。给药可能发生一次,或者以几分钟或几小时的间隔发生,或者在这些时段中的任何一个时段持续发生。合适的剂量可以在约0.1ng/kg体重至1g/kg体重/剂量的范围内。典型的剂量范围为每剂量1μg至1g/kg体重,例如每剂量1mg至1g/kg体重范围内。在一个实施方案中,剂量可以在每剂量1mg至500mg/kg体重的范围内。在另一实施方案中,所述剂量可以在每剂量1mg至250mg/kg体重的范围内。在又一个实施方案中,所述剂量可以在每剂量1mg至100mg/kg体重的范围内,例如每剂量高达50mg/体重。
如本文所使用的术语“治疗(treatment)”和“治疗(treating)”涵盖对动物,优选哺乳动物(更优选人类)中的病症或疾病的任何治疗,并且包括治疗由核内体par2信号传导介导的疼痛。如本文所用的术语“预防(prevention)”和“预防(preventing)”涵盖对动物,优选哺乳动物(更优选人类)中的病症或疾病的预防或预防性处置,并且包括预防由核内体par2信号传导介导的疼痛。
在整个说明书和随后的权利要求书中,除非上下文另有要求,否则词语“包括”以及诸如“包含”或“含有”的变化将被理解为暗示包括所述的整数或整数或步骤的组,但是不排除任何其他整数或整数的组。
本说明书中对任何在先出版物(或从中得出的信息)或任何已知事项的提及不是也不应被视为确认或承认或以任何形式表明该先前出版物(或从中得出的信息)或已知事物形成本说明书涉及领域中的公知常识的一部分。
现在将参考以下非限制性实施例来描述本发明:
实施例1:受体运输的bret研究
进行受体运输的bret研究以在用不同激动剂刺激时定量par2内化(rit-venus)和运输至早期核内体(rab5a-venus)。
方法
在10cm培养皿中达到约50%融合后,使用pei作为转染剂,用1μgpar2-rluc8和4μgrit-venus或rab5a-venus瞬时转染hek293细胞。转染后24小时,将细胞接种到聚-d-赖氨酸涂覆的96孔isoplate中。第二天将细胞培养基替换为1xhbss(+0·28ghepes/100ml,ph7·40);30μmdyngo4a(dyn4a);30μm无活性的dyn4a对照;30μmpitstop2(ps2)或30μm无活性的ps2对照。当使用时内吞抑制剂在1xhbss+1%二甲基亚砜(dmso)中稀释。更换培养基后,使细胞在37℃下平衡30分钟,然后加入腔肠素h(colenterazineh)[5μm]。再孵育5分钟后,使用lumistaromega微板读取器(bmglabtech,offenburg,germany)评估bret,在535nm和475nm处读取发光。在加入媒介物(1xhbss+/-1%dmso)或蛋白酶之前获得4分钟基线,然后继续测量另外30分钟。使用lumistarmars分析软件计算bret比率,然后在microsoftexcel2013中校正至基线和媒介物处理。
结果
最初的运输实验表明胰蛋白酶是导致par2内化的唯一激动剂。用10nm浓度的胰蛋白酶刺激受体导致具有质膜标记物的bret的时间依赖性减少(图1a)。这与受体和早期核内体标记物之间的bret增加一致(图2a)。这种反应与受体内化一致,并且在用弹性蛋白酶或组织蛋白酶s刺激受体时未观察到。因此认为弹性蛋白酶和组织蛋白酶s激活的par2仅能从质膜发出信号。
通过向细胞施用一系列浓度,进一步探索胰蛋白酶诱导受体内化的能力。发现胰蛋白酶诱导的par2内化是浓度依赖性的(图1b和2b)。类似的ec50浓度来自两种标记物的测定(~2·8nm)。
ps2对网格蛋白的药理学抑制降低了胰蛋白酶刺激后荧光素酶标记受体和两种标记之间bret的程度,与内化的抑制一致。用dyn4a抑制发动蛋白时,观察到胰蛋白酶诱发内化的不太明显的抑制(图1c和2c)。
实施例2:用于评估区室化erk信号传导的全群体fret
方法
在10cm培养皿中达到约50%融合后,使用pei作为转染剂,用2·5μg的par2和n末端ha表位标记(par2-ha)和2·5μg所需的fret生物传感器转染hek293细胞。转染后24小时,将细胞接种到聚-d-赖氨酸涂覆的黑色96孔isoplate中。在过夜血清限制后,在细胞铺板后48小时评估fret。在开始fret实验之前,将细胞培养基替换为1xhbss(+0·28ghepes/100ml,ph7·40);30μmdyngo4a;30μm无活性的dyngo对照;30μmpitstop2或30μm无活性的pitstop2对照。当使用时内吞抑制剂在1xhbss+1%二甲基亚砜(dmso)中稀释。更换培养基后,使细胞在37℃下平衡30分钟。然后使用pherastarfs微板读取器(bmglabtech,offenburg,germany)测定fret,在430nm处依次激发细胞,在550nm和490nm处读取发射。在加入媒介物(1xhbss+/-1%dmso)、蛋白酶或1μmpdbu(用作阳性对照)之前获得4分钟基线。然后每分钟继续测量再保持30分钟。然后计算fret比率并在microsoftexcel2013中校正至基线和媒介物处理。。
结果
胰蛋白酶而不是弹性蛋白酶,能够在细胞质和细胞核内产生erk信号(图3a和4a)。在这些区室中胰蛋白酶诱发的erk信号显示出浓度依赖性(图3b和4b)。当通过用ps2抑制网格蛋白来阻断受体内化时,细胞溶质erk磷酸化减少,但当发动蛋白受到抑制时未减少(图3c)。通过靶向网格蛋白或发动蛋白阻断内化似乎不影响细胞核中erk磷酸化的程度(图4c)。
实施例3:bret再敏化测试
bret再敏化测试用于定量在用多种蛋白酶刺激在细胞表面表达的受体时来自反面高尔基体(trans-golgi)网络(tgn38-venus)的par2的输出。用多种信号传导节点抑制剂重复测定,以验证受体再敏化背后的信号传导机制。
方法
在10cm培养皿中达到约50%融合后,使用pei作为转染剂,用par2-rluc8(1μg)和tgn38-venus(4μg)瞬时转染hek293细胞。转染后24小时,将细胞接种到聚-d-赖氨酸涂覆的96孔isoplate中。第二天进行bret实验。使细胞在1xhbss(+0·28ghepes/100ml,ph7·40)、10μm棓子色素(gallein)(gβγ抑制剂)、100nmcrt0066101(pkd抑制剂)或go6983(pkc抑制剂)中在37℃下平衡30分钟,然后加入腔肠素h[5μm]。孵育5分钟后,使用lumistaromega微板读取器(bmglabtech,offenburg,germany)评估bret,在535nm和475nm处读取发光。在加入媒介物(1xhbss)或蛋白酶之前获得4分钟的基线,然后继续测量另外30分钟。使用lumistarmars分析软件计算bret比率,然后在microsoftexcel2013中校正至基线和媒介物处理。
结果
荧光素酶标记的par2显示在用所有三种激动剂刺激在细胞表面表达的受体时远离反面高尔基体网络(图5a)。这些发现支持通过从反面高尔基体网络中存储的预先存在的受体库中的受体补充来实现par2再敏化。还发现观察到的bret降低是浓度依赖性的(数据未显示)。
用棓子色素抑制gβγ减少了用所有三种蛋白酶刺激后远离反面高尔基体网络的par2运输,暗示这种g蛋白在再敏化过程中。在分别用crt0066101和go6983抑制pkd和pkc后,观察到对正常运输的不太显著的影响。
实施例4:神经元兴奋性的内吞抑制
为了评估蛋白酶对伤害感受器的兴奋性的影响,将drg神经元与胰蛋白酶、组织蛋白酶s、弹性蛋白酶或媒介物(对照)一起预孵育,洗涤,并在洗涤后0分钟或30分钟时测量基强度和在两倍基强度的电流下的动作电位放电。
方法
分离drg神经元。在37℃下通过在胶原酶(1mg/ml,worthingtonbiochemical,lakewood,nj)和分散酶(4mg/ml,rochelifescience,indianapolis,in)中孵育10分钟来消化来自c57bl/6小鼠的drg(t9-t13),用火抛光的巴斯德吸管研磨分散。将细胞接种到涂有层粘连蛋白(0.017mg/ml)和聚-d-赖氨酸(2mg/ml)的24孔板中的盖玻片上。将神经元在含有10%胎牛血清、含有青霉素和链霉素的f12培养基(catn668,批号rnbd5333,sigma-aldrich,stlouis,mousa)中培养,并在37℃、95%空气和5%co2的湿润气氛中保持直到取回(16小时)用于电生理学研究。
电生理学.通过在室温下以电流钳模式从小直径神经元(<30pf电容)用两性霉素b(240μg/ml)进行穿孔膜片钳记录来评估神经元兴奋性。仅分析具有比-40mv更负性的静息膜电位的神经元。通过测量基强度(对引起动作电位的最小电流)和在两倍基强度释放的动作电位的数量来定量兴奋性的变化。使用axopatch200b放大器进行记录,由digidata1322a数字化并使用pclamp10.1软件(moleculardevices,sunnyvale,ca)储存和处理。用2ml/min的外部溶液连续灌注记录室。外部溶液是(mm):140nacl、5kcl、10hepes、10d-葡萄糖、1mgcl2、2cacl2;ph用3mnaoh调节至7.4。移液管溶液是(mm):110k-葡糖酸盐,30kcl,10hepes,1mgcl2,2cacl2;ph用1mkoh调节至7.25。
蛋白酶和抑制剂治疗.将drg神经元与媒介物(对照)或蛋白酶一起预孵育,使用我们报道的引起强烈超兴奋性的条件:胰蛋白酶(50nm,10分钟)、弹性蛋白酶(10u/ml,390nm,30分钟)、cat-s(500nm,60分钟)。然后洗涤神经元并保持在无蛋白酶的f12培养基中。在洗涤后0或30分钟评估神经元兴奋性。在一些实验中,在与蛋白酶孵育之前和期间,将神经元与媒介物(0.3%dmso)、dyngo4a(30μm)或pitstop2(15μm)孵育30分钟。
统计分析.结果表示为平均值±se。使用双向anova和事后tukey检验来分析数据。
结果
所有蛋白酶在0分钟和30分钟时引起基强度的减少,与兴奋性一致。为了评估网格蛋白和发动蛋白对兴奋性的重要性,在与蛋白酶孵育之前和期间,将drg神经元与网格蛋白抑制剂pitstop2、发动蛋白抑制剂dyngo4a或媒介物(0.3%dmso)预孵育30分钟。如图6和7中可见,pitstop2和dymgo4a在0分钟时不影响胰蛋白酶、组织蛋白酶s或弹性蛋白酶对基强度的作用。pitstop2和dyngo4a均在30分钟时阻止胰蛋白酶诱导的基强度减少,但在30分钟时对组织蛋白酶s-或弹性蛋白酶诱导的基强度减少没有影响。单独的媒介物、pitstop2和dyngo4a均未在0或30分钟时影响基强度。
实施例5:内吞抑制剂对蛋白酶诱发的疼痛的作用
方法
伤害感受测定.在实验之前,使小鼠连续2天适应实验装置、室和研究者1-2小时。通过对用标刻度的vonfrey纤维丝(filament)刺激后爪的跖面的缩足来评估机械伤害感受反应。一式三份测量vonfrey评分,以在实验前一天为每只动物建立基线。通过在治疗之前和之后使用数字卡尺测量后爪的厚度来估计爪水肿。用5%异氟烷麻醉小鼠用于足底注射。将dyngo4a(50μm)、pitstop2(50μm)或媒介物(0.2%dmso在0.9%nacl中)(全部10μl)注射到左后爪中。30分钟后,将胰蛋白酶(10nm)、cat-s(2.5μm)或弹性蛋白酶(30u)(均为10μl)注射到相同的左后爪中。在蛋白酶注射后每小时测量vonfrey评分(左和右爪)和爪厚度(左爪)长达4小时。调查人员对测试试剂不知情。
结果
足底注射胰蛋白酶、cat-s和弹性蛋白酶导致同侧爪的机械痛觉过敏持续至少4小时,并且同侧爪的水肿在4小时后最大(图8和9)。先前已报道这些反应依赖于par2和trpv4。dyngo和pitstop抑制胰蛋白酶诱发的机械性痛觉过敏。dyngo不影响cat-s-或弹性蛋白酶诱发的机械性痛觉过敏。dyngo不影响胰蛋白酶-cat-s-或弹性蛋白酶诱发的水肿。内吞抑制剂不影响对侧爪的缩回反应。
这些结果表明,对胰蛋白酶诱导的伤害感受的中枢传递需要par2内吞作用。对组织蛋白酶s或弹性蛋白酶诱发的机械性疼痛不需要par2内吞作用-这些蛋白酶不会引起par2内吞作用。蛋白酶诱发的水肿不需要par2内吞作用,其取决于ca介导的sp和cgrp释放。
实施例6:核内体par2信号传导的抑制
在稳定表达人par2受体的knrk细胞或内源性表达par2的ht-29细胞中,在测量2-糠酰基-ligrl-nh2(2f)刺激的ip1积累的测试中测定各种推定的par2拮抗剂的抑制效力。在测量拮抗剂抑制knrk细胞中atp刺激的ip1积累的能力的测定中确定非特异性活性。
方法
将knrk-hpar2、knrk或ht-29细胞以50x103个细胞/孔的密度接种在透明的聚-d-赖氨酸涂覆的96孔组织培养板中。在37℃和5%co2孵育24小时后,除去培养基并用80μl或90μlip1刺激缓冲液(10mmhepes,1mmcacl2,0.5mmmgcl2,4.2mmkcl,146mmnacl,5.5mm葡萄糖,50mmlicl)替换。添加刺激缓冲液后,含有80μl缓冲液的孔接受10μl10x拮抗剂。将所有平板在37℃、5%co2下进一步孵育30分钟。向平板中加入10μl2f或atp,并进一步孵育40分钟。孵育后,通过抽吸快速除去刺激缓冲液,并用25μl裂解缓冲液(ip-onehtrf测定试剂盒,cisbio)替换。在37℃、5%co2下孵育裂解物10分钟后,将10μl裂解物转移至384孔optiplate(perkinelmer)并使用ip-onehtrf测定试剂盒(cisbio)检测。
将浓度响应曲线范围为3nm至30μm(以10μl体积以10x浓度添加)的par2拮抗剂孵育30分钟,然后加入10μmatp或100nm或300nm2f(ec80,以10x浓度加入10μl体积),然后进一步孵育40分钟。
结果
发现下面示出的par2拮抗剂抑制2f刺激的ip1积累,ic50为5.39±0.13μm。
该拮抗剂对knrk细胞中atp诱导的ip1积累没有影响,表明hpar2-knrk细胞中2f的拮抗作用是par2介导的。拮抗剂也不影响knrk细胞中的基线ip1积累。
- 还没有人留言评论。精彩留言会获得点赞!