产生油脂的酵母和油脂制造方法与流程
本发明涉及产生油脂的酵母和油脂制造方法。更具体地,本发明涉及在菌体外蓄积油脂的酵母株以及使用该酵母株的油脂制造方法。
背景技术:
为了摆脱对化石燃料的依赖并且抑制温室气体的排放,需要对碳中和资源(即生物质)的有效利用。关于生物质的利用,专利文献1公开了利用诸如小球藻的藻类制造油脂的技术。该文献公开的技术通过在压力容器内分解包含藻类和水的混合物,从而在不使用有机溶剂的情况下从藻体中回收油脂。
本发明相关的专利文献2公开了从碳水化合物中产生油脂的丝孢酵母(trichosporon)属酵母。
现有技术文献
专利文献
1.专利文献特开2012-5398号公报
2.专利文献特开平7-236492号公报
技术实现要素:
发明待解决的问题
由于专利文献2所公开的丝孢酵母属酵母在菌体外产生油脂,因此可在不进行从菌体内提取油脂的操作的情况下获得油脂,而且所述丝孢酵母属酵母可能用于由生物质制造油脂。但是,对于丝孢酵母属酵母,尚未进行关于糖同化性的详细研究。另外,由于进行丝状真菌式生殖,所以丝孢酵母属酵母大多生长缓慢,并存在病原性菌种。
本发明的主要目的在于提供一种可用于由生物质制造油脂的新型酵母。
解决问题的手段
为了解决如上问题,本发明提供以下1-4。
1.一种油脂的制造方法,其包括在含有碳源的培养基中培养红冬孢酵母(rhodosporidiumtoruloides)ipm33-18株(nitebp-01900)或其基因改造体,以在培养物中蓄积油脂的步骤;
从所述培养物中回收所述油脂的步骤。
2.根据1所述的制造方法,其包括从所述培养物的液体部分中回收油脂的步骤。
3.根据1或2所述的制造方法,其特征在于不包括从菌体中提取油脂的步骤。
4.根据1或2所述的制造方法,其包括从菌体中回收油脂的步骤。
5.根据1~4中任一项所述的制造方法,所述基因改造为导入2-脱氧葡萄糖耐受突变。
6.红冬孢酵母(rhodosporidiumtoruloides)ipm33-18株(nitebp-01900)或其基因改造体。
在本发明中,术语“培养基”、“培养物”和“培养物的液体部分”具有下述含义。
“培养基”意指包含酵母生长中所需的营养成分的液体。“培养基”不应包含菌体和由菌体产生的油脂。
“培养物”包括培养基和在培养基中生长的菌体。此外,“培养物”应包含由在培养基中生长的菌体产生的油脂。
“培养物的液体部分”意指在培养物中除了菌体以外的液相。“培养物的液体部分”应包含由菌体产生的油脂中的在该液体部分中所蓄积的那些。即,由菌体产生的油脂中的在菌体内蓄积的那些不包含在“培养物的液体部分”中。另外,“培养物的液体部分”由以下成分组成:包含培养基的水层和包含由菌体在液体部分中产生的油脂的油层。
此外,在本发明中,“在培养物中的油脂蓄积”意指由酵母产生的油脂在“菌体”内和“培养物的液体部分”中的蓄积。其中,“在培养物的液体部分中蓄积”包括由酵母产生的油脂释放至该液体部分中,由酵母产生的油脂分泌至菌体外的情况,以及在菌体内的产生油脂的酵母通过细胞死亡而分解,油脂被释放到菌体外的情况。另外,术语“在菌体外蓄积”与“在培养物的液体部分中蓄积”同义使用。
发明效果
根据本发明,提供了一种可用于从生物质中制造油脂的新型酵母。
附图说明
图1为显示了在ipm33-18株的菌体内和菌体外的油脂蓄积量的测量结果的图(实施例1)。
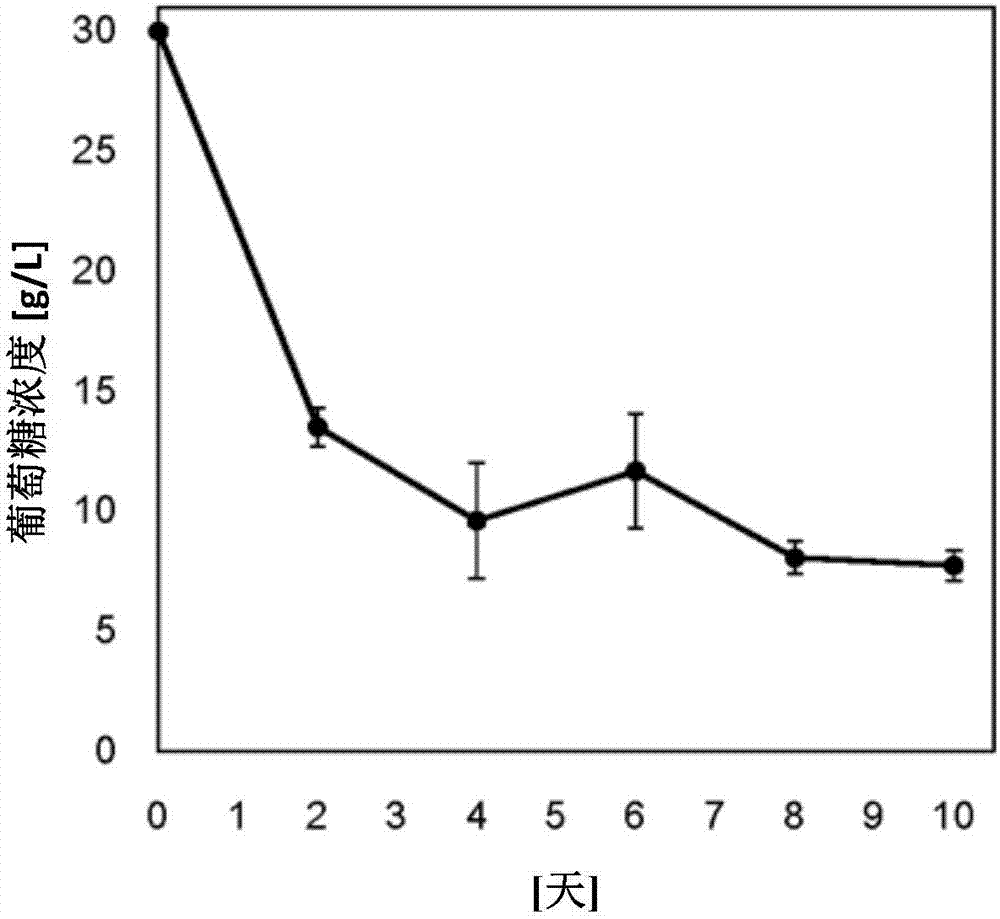
图2为显示了在ipm33-18株的培养上清液中的葡萄糖浓度的测量结果的图(实施例1)。
图3为显示了在ipm33-18株的培养上清液中的氮浓度的测量结果的图(实施例1)。
图4为显示了ipm33-18株的菌体重量(干燥重量)的测量结果的图(实施例1)。
图5为显示了在培养开始10天时,在ipm33-18株的菌体外蓄积的油脂的脂肪酸组成的图(实施例1)。
图6为显示了在ipm33-18株的基因改造体的菌体外的油脂蓄积量的测量结果的图(实施例3)。
具体实施方式
以下,对用于实施本发明的优选方式进行说明。另外,以下说明的实施方式只表示本发明的代表性实施方式的一个例子,因此并不构成对本发明的范围进行的狭义解释。
本发明人从日本的亚热带地区(西表岛)和寒冷地区(利尻岛)的土壤和植物中分离酵母株,并构建了包含1000株以上酵母的库(plosone,2012,vol.7,no.11,e50784)。本发明人使用该酵母库对具有能够在菌体外蓄积油脂的能力的株进行筛选,结果新发现了红冬孢酵母(rhodosporidiumtoruloides)ipm33-18株具有该能力。基于该发现,本发明提供了使用该菌株制造油脂的方法。
ipm33-18株已被国际保藏在独立行政法人制品评价技术基础机构专利微生物保藏中心(千叶县木更津市上总镰足2-5-8122号室),登录号为nitebp-01900。
ipm33-18株在10~37℃的温度下生长,并具有广泛的糖同化性。因此,在通常环境下,其能够用于从多种生物质中产生油脂。此外,不同于在菌体内蓄积产生的油脂的通常的油酵母(蓄积油脂的酵母),ipm33-18株在菌体外蓄积产生的油脂,因此根据ipm33-18株,可在不进行从菌体内提取油脂的操作的情况下制造油脂。
ipm33-18株的菌学性质如下。
分类学上的位置
红冬孢酵母(rhodosporidiumtoruloides)
科学性质
基因组dna的d1/d2区域的碱基序列与上述种子的标准株一致。培养基上的形态等也显示与红冬孢酵母相同的性质。
培养条件
有氧、静置培养
培养基组成
另外,在本发明中,ipm33-18株的基因改造体可代替ipm33-18株使用或与ipm33-18株组合使用。可适当选择本领域技术人员公知的方法用于基因改造,例如使用突变原性物质的ipm33-18株的培养和使用载体的基因导入法。
作为基因改造,优选的是用于提高糖的同化性的改造或用于提高油脂在培养物中的蓄积量的改造。作为用于提高糖的同化性的改造,可以提及例如导入2-脱氧葡萄糖耐受突变。
或者,在本发明中,ipm33-18株的提取液可代替ipm33-18株使用。在该提取液中可以使用经纯化的所需酶的一部分或全部。例如,当使用衍生自ipm33-18株的脂肪酶时,根据lukaszewicz等人的文献(1stannualinternationalinterdisciplinaryconference,aiic2013年4月24-26,azores,portugal,第441-449页)中所述的方法,可以通过硫酸铵分级纯化。
本发明的油脂的制造方法包括在含有碳源的培养基中培养ipm33-18株或其基因改造体,以在培养物中蓄积油脂的步骤;从所述培养物中回收所述油脂的步骤。如下所述,从培养物中回收油脂的步骤可以是如下步骤的任一者:从培养物的液体部分中回收油脂的步骤,或者除了从所述液体部分中回收油脂之外还从菌体中回收油脂的步骤。
可以使用含有碳源的培养液通过现有公知的方法培养ipm33-18株。由于ipm33-18株具有广泛的糖同化性,因此可以使用(而无特别限定)糖类、糖醇和酸性糖或包含它们的生物质作为碳源。这里,在本发明中,“生物质”意指含有上述碳源的可再生材料。
糖类可以是单糖类、低聚糖类以及多糖类。低聚糖类指二~十糖类,它们可以是均质低聚糖类或异质低聚糖类。另外,多糖指具有比低聚糖类更大单糖单元数的糖类,它们可以是均质多糖或异质多糖。具体而言,单糖类可以是戊糖(如l-阿拉伯糖、d-木糖、d-核糖等)、己糖(如d-葡萄糖、d-半乳糖、d-果糖、d-甘露糖等)、6-脱氧己糖(如l-鼠李糖等)等。低聚糖类可以是二糖(如蔗糖、麦芽糖、乳糖、纤维二糖、海藻糖、蜜二糖等)、三糖(如棉子糖等)等。多糖类可以是淀粉、纤维素、肝糖、葡聚糖、甘露聚糖、木聚糖等。上述糖类可以单独使用或适当组合使用。淀粉水解物等也可包含在上述组合中。此外作为糖类,也可以使用包含糖类为主要成分的原料,例如废糖蜜、豆渣等。
糖醇可以是d-山梨糖醇、d-甘露糖醇、半乳糖醇、麦芽糖醇等。酸性糖可以是葡萄糖醛酸、半乳糖醛酸等。
培养基中的碳源的量不受特别限制,但通常为约3~15%(w/w)。
除了碳源以外,培养基可以包含氮源、无机物和其他营养素。作为氮源,可以使用氨、氯化铵、硫酸铵、碳酸铵、乙酸铵、硝酸钠、尿素等无机有机氮化合物。进一步,作为氮源,也可以使用诸如蛋白胨、肉提取物、酵母提取物、玉米浆、酪蛋白水解物、鱼粉或其消化物、脱脂大豆饼或其消化物的含氮天然产物。作为无机物,可以使用磷酸二氢钾、磷酸氢二钾、硫酸镁、氯化钠、硫酸亚铁、氯化亚铁、硫酸锰、氯化钙、碳酸钙、硫酸锌、硫酸铜、硼酸钼酸铵、碘化钾等。
培养在振荡培养或深部搅拌培养等的有氧条件下进行。培养温度一般优选为20~35℃,但只要是细菌可以生长的温度,也可使用其他温度条件。培养中培养基的ph通常为4.0~7.2。培养开始后通常2~4天内,油脂在菌体外产生蓄积。
可以使用亲油性溶剂通过现有公知的方法从培养物中回收油脂。具体地,向培养物中添加溶剂,将培养物的液体部分中所包含的油脂回收到溶剂中。可以在从培养物中分离菌体而得到液体部分之后,向液体部分中添加溶剂来进行油脂的回收。可通过诸如离心分离和静置沉降等的操作以及诸如分离器、倾析器和过滤器等的装置从培养物中分离菌体,从而获得培养物的液体部分。其中,可根据常规方法提取在菌体内蓄积的油脂。作为溶剂,优选使用溶解油脂,但与水不可溶混或与水较差溶混、在常温下为液状的有机溶剂,例如卤代低级烷烃(氯仿、二氯甲烷、四氯化碳、1,2-二氯乙烷)、正己烷、乙醚、乙酸乙酯、芳香族烃(苯、甲苯、二甲苯)等。不特别限定提取溶剂的添加量,只要其能够充分地回收在培养物中或液体部分中产生蓄积的油脂的量即可。
本发明的制造方法得到的油脂可以是:由脂肪族羧酸和脂肪族醇组成的脂肪族酯化合物。不特别限定脂肪族羧酸,只要其由酵母产生即可,其可以是例如,碳原子数为8~24,优选12~24的脂肪族羧酸,具体为肉豆蔻酸、棕榈酸、棕榈油酸、硬脂酸、油酸、亚油酸、亚麻酸、山嵛酸和二十四烷酸等。另外,作为油脂,也可以包含诸如磷脂质、游离脂肪酸、糖脂质、类固醇化合物、类胡萝卜素等的光合色素等。
根据本发明的制造方法,可在不进行从菌体中提取油脂的操作的情况下从培养物或其液体部分中回收油脂。以往为了提取菌体内的油脂,破坏坚固的细胞壁的操作是必要的。与此相对,根据本发明的制造方法,有可能仅通过混合培养物或其液体部分与亲油性溶剂来回收油脂。另外,如上所述,本发明的ipm33-18株具有广泛的糖同化性。因此,本发明可用于从生物质中制造油脂,并且可用于例如生物柴油生产或利用未使用的生物质的生物炼制等的应用。
实施例
<实施例1:ipm33-18株的油脂菌体外蓄积能力>
按以下步骤,培养ipm33-18株,从培养物的液体部分中回收油脂。
培养
培养在以下条件下进行。
用铂环刮取ym琼脂培养基上的菌落,并将其移入3ml的ym培养基,在27℃和150rpm下预培养过夜。用分光光度计测量600nm处的吸光度,确定浊度(od600)。使用25ml的ss2培养基(3%葡萄糖、0.5%硫酸铵、0.05%硫酸镁、0.01%氯化钠、0.01%氯化钙、0.01%酵母提取物)接种预培养液,使得od600变成0.2,主培养在27℃和150rpm下进行。
采样
在主培养开始后的第2、4、6、8、10天时采集3ml培养物,在12,000rpm下离心分离5分钟,并回收菌体。使用培养上清液(培养物的液体部分)用于测量葡萄糖浓度和氮浓度。菌体用蒸馏水洗涤1次后,再次离心分离并冷冻干燥。
另外,将5ml的己烷添加至剩余量的培养上清液中并搅拌,分离出上层(己烷层)。用蒸发器去除己烷,得到红色油状残渣。
在培养开始后的第2、4、6、8、10天时测定以下项目。
葡萄糖浓度
使用高效液相色谱进行培养上清液中的葡萄糖浓度的测定。高效液体色谱分析在以下条件下进行。
高效液相色谱仪:由岛津制作所制造的prominence
柱:由bio-radlaboratories公司制造的aminexfermentationmonitoringcolumn
保护柱:由bio-radlaboratories公司制造的micro-guardcationhrefillcartridges
检测器:由岛津制作所制造的rid10
柱式加热炉:由岛津制作所制造的cto20a
自动进样器:由岛津制作所制造的sil-20a
流动相:5mm硫酸水溶液,每分钟0.6ml
柱温:60℃
由浓度已知的d-葡萄糖(和光纯药工业)的相对保留值和峰面积来鉴定葡萄糖,并确定其浓度。
氮浓度
根据文献(bioresour.technol.,2012,vol.114,p.443-449)中所述的方法进行培养上清液中的氮浓度的测量。向100μl的上清中加入500μl的碱性次氯酸盐溶液、500μl的酚硝普盐(フェノールニトルプルシド)溶液和3ml的水。在25℃下放置1小时后,测量570nm处的吸光度。基于用20、40、80mg/l的硫酸铵水溶液制成的校准线来确定氮浓度。
油脂量
根据文献(j.lipidres,2010,vol.51,no.3,p635-640)中所述的方法将通过如上采样获得的干燥菌体和红色油状残渣甲基化。在将300μl甲苯和1.5ml甲醇添加至干燥菌体或红色油状残渣中后,仅对干燥菌体进行15分钟的超声破碎。接着,加入含8%盐酸的85%的甲醇300μl,搅拌后,在45℃下静置过夜。最后,加入1ml己烷和1ml蒸馏水,搅拌后,挑取上层(己烷层),供接下来的气相色谱分析用。
通过气相色谱分析,解析脂肪酸甲酯的组成。气相色谱分析在以下条件下进行。
气相色谱仪:由岛津制作所制造的gc-2010plus
检测器:fid
自动进样器:由岛津制作所制造的aoc20
柱:由agilenttechnologies公司制造的db-23毛细管柱(30m×0.25mm×0.25μm)
温度程序:在50℃下保持2分钟,以每分钟10℃升温至180℃,保持5分钟,以每分钟5℃升温至240℃,保持3分钟
载气:氦气(每分钟1ml)
补气:氮气
注射器温度:250℃
检测器温度:300℃
分流比:50:1
由标准物质(花生酸甲酯、山嵛酸甲酯、癸酸甲酯、顺式-13-二十二烯酸甲酯、月桂酸甲酯、亚油酸甲酯、亚麻酸甲酯、肉豆蔻酸甲酯、辛酸甲酯、油酸甲酯、棕榈酸甲酯、棕榈油酸甲酯、硬脂酸甲酯、二十四烷酸甲酯、和光纯药工业)的相对保留值鉴定各脂肪酸甲酯。由脂肪酸甲酯混合物(18918-1amp,supelco)的峰面积来确定各脂肪酸甲酯的浓度。
图1示出了培养期间在菌体内和菌体外的油脂蓄积量的变化。图1的纵轴示出了在1l培养物所包含的菌体中蓄积的油脂量(细胞内)以及在液体部分中蓄积的油脂量(细胞外)。另外,图2~4示出了培养上清液中的葡萄糖浓度、氮浓度和干燥菌体重量的变化。
随着培养的进行,培养基中的葡萄糖和氮被消耗而产生油脂,其中可以明确的是:一定量的油脂在细胞外蓄积。特别地,当菌体的增殖达到平稳在培养第4天之后,油脂的菌体外蓄积增加,菌体外蓄积量/菌体内蓄积量的比例显著增加。
图5示出了菌体外蓄积的油脂的脂肪酸组成(培养开始第10天)。菌体外(10dextra)和菌体内(10intra)蓄积的油脂的组成无明显差异,两者均包含肉豆蔻酸(c14:0)、棕榈酸(c16:0)、棕榈油酸(c16:1)、硬脂酸(c18:0)、油酸(c18∶1)、亚油酸(c18∶2)、亚麻酸(c18∶3)、山嵛酸(c22:0)、二十四烷酸(c24:0)之类的主要脂肪酸。
<实施例2:ipm33-18株的糖同化性>
用其它糖类替换实施例1中用于碳源的葡萄糖,通过检测油脂的菌体外蓄积来分析ipm33-18株的糖同化性。使用木糖、阿拉伯糖、甘油、纤维二糖、淀粉、木聚糖作为糖类。结果,在将任何糖用作碳源的情况下,都能够确定脂质的菌体外蓄积与葡萄糖类似。很明显,ipm33-18株具有广泛的糖同化性。
<实施例3:ipm33-18株的基因改造>
将2-脱氧葡萄糖耐受突变导入ipm33-18株中,以分析油脂的菌体外蓄积量的变化。
根据文献(appl.microbiol.biotechnol.2011,vol.90,no.4,p.1573-1586)中所述的方法,将2-脱氧葡萄糖耐受突变导入ipm33-18株中以制备基因改造体。在合成木糖培养基(0.17%ynb,0.5%as,6%木糖)中培养基因改造体。除了培养基以外的培养条件与实施例1相同。
图6示出了培养期间菌体外的油脂蓄积量的变化。通过导入2-脱氧葡萄糖耐受突变,油脂的产量显著增加。
pct
受理局填写栏
国际局填写栏
pct/ro/134表
- 还没有人留言评论。精彩留言会获得点赞!