一株青霉素酰化酶及其编码基因、产生菌和应用的制作方法
本发明涉及了一株青霉素酰化酶产生菌株,该菌产的一种青霉素酰化酶及其基因,以及酶法合成青霉素G类以及头孢菌素类半合成抗生素的方法,属于生物制药技术领域。
背景技术:
青霉素酰化酶(penicillin acylase,PGA)又称为青霉素氨基酰化酶,是一种N末端亲核丝氨酸水解酶。该酶在酸性以及中性条件下,可用于催化6-氨基青霉烷酸(6-APA)和7-氨基脱乙酰头孢烷酸(7-ADCA)与酰基供体发生酰基化反应,制备半合成抗生素;而在偏碱性条件下,催化青霉素钾盐等水解成为6-APA和苯乙酸(PAA),而6-APA是大量制备半合成β-内酰胺抗生素所需要的重要母核。并且PGA能合成的底物范围广,可以催化制备高效、广谱、适用于不同用途的新型β-内酰胺抗生素。因此,青霉素酰化酶是工业上应用价值很高的酶。
目前生产最多、应用最广的半合成抗生素是半合成青霉素类和头孢菌素类抗生素。其中半合成青霉素类抗生素是以6-氨基青霉烷酸为母核,对其侧链进行化学修饰等产生的,主要有氨苄青霉素、阿莫西林、阿洛西林、氨苄西林等;头孢菌素类抗生素是以7-氨基去乙酰氧基头孢烷酸为母核形成的抗生素药物,主要有头孢氨苄、头孢羟氨苄、头孢拉定、头孢克洛等。
另外,头孢替唑是第一代头孢菌素类抗生素,通过抑制细菌细胞壁的合成而发挥抗菌活性。临床上主要用于治疗呼吸系统感染、泌尿系统感染、败血症、腹膜炎等。据现有文献报道,头孢替唑的合成方法主要是化学法,酶法合成即以7-氨基-3- (2-氨基-1,3,4-噻二唑)硫甲基头孢烷酸为母核形成头孢替唑尚未有文献报道。
尽管青霉素酰化酶的研究已取得重要进展,但性能优良并能够适用于多种新一代β-内酰胺抗生素合成的酶类仍然难以满足需求,因此筛选性能更优越的青霉素酰化酶,如pH耐受性好、合成底物范围广泛的酶更适用于工业催化等,对酶法催化半合成β-内酰胺类抗生素具有重要意义。
技术实现要素:
本发明公开了一种青霉素酰化酶产生菌,该菌产的一种青霉素酰化酶及其基因,该青霉素酰化酶表达载体构建和该酶在大肠杆菌(E.coli)BL21(DE3)中的异源表达获得重组酶,该青霉素酰化酶合成以6-氨基青霉烷酸、7-氨基去乙酰氧基头孢烷酸、7-氨基-3- (2-氨基-1,3,4-噻二唑)硫甲基头孢烷酸为母核的β-内酰胺类抗生素。
本发明的目的之一是提供一种青霉素酰化酶产生菌,该菌株为木糖氧化无色杆菌,命名为Achromobacter xylosoxidans PX02,保藏登记号为CCTCC NO:M 2016392。
本发明的又一目的是提供所述青霉素酰化酶产生菌产生的青霉素酰化酶,其特征在于所述青霉素酰化酶具有SEQ ID NO:2所示的氨基酸序列。
本发明的再一目的是提供所述青霉素酰化酶的编码基因、重组载体或重组菌,所述编码基因的核苷酸序列如SEQ ID NO:1所示,所述重组菌由所述编码基因与载体pET-28a构建的表达载体转化到大肠杆菌BL21(DE3)所得。
本发明的目的之一是提供一种青霉素酰化酶产生菌在合成β-内酰胺类抗生素中的应用。
为了实现本发明的目的,本发明首先从南京周边药厂土壤中筛选获得青霉素酰化酶菌株PX02,通过16s rDNA基因序列对比分析,该菌株被鉴定为Achromobacter xylosoxidan,命名为Achromobacter xylosoxidans PX02,其保藏登记号为CCTCC NO:M 2016392。
本发明分离克隆到Achromobacter xylosoxidans PX02 菌株来源青霉素酰化酶PGAPX02的编码基因,它具有SEQ ID NO:1所示的核苷酸序列,为此基因的改造并在各种外源基因表达系统中高效表达提供了优良的基因材料。通过PCR法分离克隆了这一青霉素酰化酶PGAPX02基因,DNA全序列分析结果表明,该青霉素酰化酶PGAPX02基因全长2583个核苷酸,编码861个氨基酸,NCBI序列比对后发现与Achromobacter xylosoxidans A8 的序列相似性90%,成熟肽氨基酸同源性为94.7%。其全部氨基酸序列示于SEQ ID NO:2。
本发明构建了青霉素酰化酶PGAPX02基因的表达载体。它可以通过本领域常规方法将本发明所述基因连接于各种载体上构建而成。所述的载体可为本领域常规的各种载体,如市售的质粒、粘粒、噬菌体或病毒载体等,优选pET28a。较佳的,可通过下述方法制得本发明的重组表达载体:将通过PCR扩增所得的产物用限制性内切酶Nhe I和Xho I进行双酶切,形成互补的粘性末端,同时将载体pET28a用限制性内切酶Nhe I和Xho I进行双酶切经T4 DNA连接酶连接,构建了含有本发明的青霉素酰化酶PGAPX02基因的重组表达载体pET28a-pgapx02。
本发明将上述重组表达载体转化至宿主细胞制得重组表达转化体。所述的宿主可为本领域常规的各种宿主,只要能满足重组表达载体可稳定地自行复制,且所携带的本发明的青霉素酰化酶PGAPX02基因能被有效表达即可。本发明优选大肠杆菌,更优选大肠埃希氏菌(E.coli)BL21(DE3)。将前述重组表达质粒pET28a-pgapx02转化至(E.coli)BL21(DE3),即可得本发明优选的基因工程菌株,即大肠埃希氏菌(E.coli) BL21(DE3)/pET28a-pgapx02。并将表达载体成功导入大肠杆菌表达菌BL21中,加入IPTG诱导表达后,经SDS-PAGE验证在BL21中成功表达了青霉素酰化酶PGAPX02,经检测,该表达酶的活力达到4000U/L。
本发明对青霉素酰化酶PGAPX02进行了酶学性质的研究。青霉素酰化酶PX02的最适反应pH为8.0,在pH 6.5~9的范围具有较高的稳定性(相对酶活90%以上)。其最佳反应温度为55℃,在25℃-45℃下温育2 h后都能保持较高的青霉素酰化酶活力(相对酶活90%以上)。实验证明该青霉素酰化酶对多种有机溶剂都具有良好的耐受性,在部分有机溶剂中青霉素酰化酶的稳定性提高。
本发明还提供青霉素酰化酶在合成β-内酰胺类抗生素中的应用。所述青霉素酰化酶PGAPX02催化合成青霉素G类半合成抗生素如阿莫西林、氨苄西林和头孢菌素类半合成抗生素如头孢氨苄、头孢羟氨苄、头孢替唑。
所述的酶催化反应分别以6-氨基青霉烷酸、7-氨基去乙酰氧基头孢烷酸和7-氨基-3- (2-氨基-1,3,4-噻二唑)硫甲基头孢烷酸为母核,以对应的侧链为酰基供体,在青霉素酰化酶PGAPX02的催化作用下进行缩合反应,以6-氨基青霉烷酸为母核合成阿莫西林、氨苄西林和以7-氨基去乙酰氧基头孢烷酸为母核合成头孢氨苄、头孢羟氨苄,以及以7-氨基-3- (2-氨基-1,3,4-噻二唑)硫甲基头孢烷酸为母核合成头孢替唑,阿莫西林和氨苄西林的合成率分别为65%-70%和67%-72%,头孢氨苄和头孢羟氨苄的合成率分别为72%-76%和75%-80%,头孢替唑的合成率为63%-69%。所述的母核与酰基供体的摩尔比为1:2~1:4,其中母核的浓度为20~100mmol/L,酰基供体的浓度为40~300 mmol/L。所述的体系为有机溶剂和水组成的混合溶剂,所述有机溶剂选自丙三醇、DMSO、聚乙二醇中的任意一种,其体积百分含量为10~80%。所述的酰基供体为羧酸及其甲酯、酰胺、酸酐及其衍生物等,所述羧酸可以为脂肪酸或芳香酸。
本发明的有益效果在于与已报道的木糖氧化无色杆菌相比,菌株Achromobacter xylosoxidans PX02的青霉素酰化酶PGAPX02单位菌体产酶量较高,发酵24 h青霉素酰化酶活力达到4000 U/L,比活力达到400 U/L/OD600;该青霉素酰化酶有耐高温、pH稳定性好、有机溶剂耐受性强、底物范围广泛等优点,该青霉素酰化酶成功催化合成五种β-内酰胺类抗生素,并且具有较好的合成头孢替唑的能力,表明其在催化合成β-内酰胺类抗生素方面有很好的应用前景。
附图说明
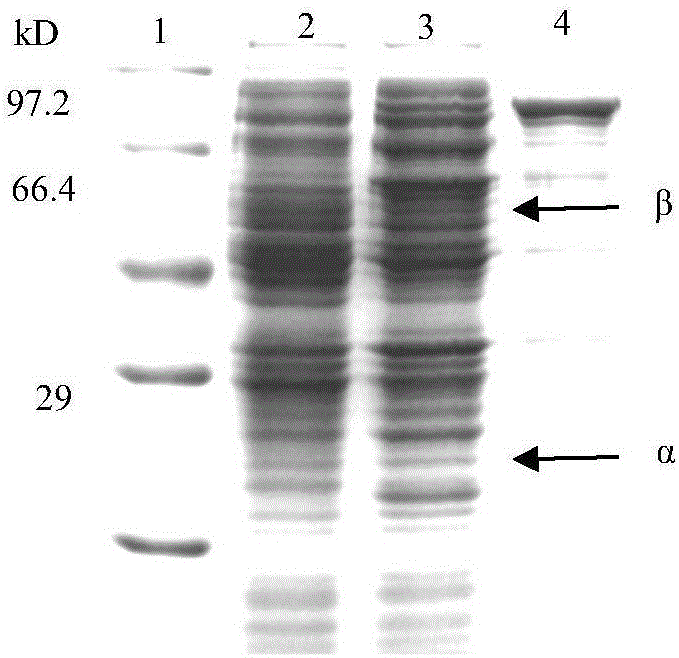
图1显示青霉素酰化酶PGAPX02的SDS-PAGE电泳分析。
图2显示青霉素酰化酶PGAPX02的最适反应pH。
图3显示青霉素酰化酶PGAPX02的pH稳定性(在各pH条件下37℃温育2h后的相对酶活)。
图4显示青霉素酰化酶PGAPX02的最适反应温度。
图5显示青霉素酰化酶PGAPX02的温度稳定性(在各温度条件下温育2h后的相对酶活)。
图6显示20%有机溶剂对青霉素酰化酶PGAPX02的影响。
本发明所述的生物材料,其分类命名为 Achromobacter xylosoxidans PX02,已保藏于中国典型培养物保藏中心(简称CCTCC),保藏编号为:CCTCC NO:M 2016392,保藏日期为:2016年7月13日,保藏地址为:中国. 武汉. 武汉大学。
具体实施方式
实施例一
本实施例说明产青霉素酰化酶菌株的筛选程序。
初筛采用如下方法:以高浓度青霉素G为筛选压力从药厂周边土壤等样品中筛选获得耐高浓度青霉素微生物。具体培养基配方为:酵母膏5 g/L,蛋白胨 5 g/L,NaCl 2 g/L,苯乙酸2 g/L,青霉素 106U/ml ,pH 7.0。培养温度为30℃,培养时间为24~48 h,摇床转速为180 rmp。此方法可筛选到大量耐高浓度青霉素微生物。
对初筛的候选菌株进行复筛,采用如下方法:用50mM磷酸缓冲液(pH7.5)配置2mg/ml底物类似物2-硝基-5-苯乙酰胺苯甲酸2-nitro-5-phenylacetamidobenzoic acid(NIPAB),并加到透明的48孔板中,将候选菌株挑到48孔板中,以不加菌株的为空白对照,37℃反应,可直接观察反应液的颜色变化,如有产青霉素酰化酶菌株,反应液会变为黄色。将所筛选到的产青霉素酰化酶菌株接种到产酶发酵培养基,具体配方为:酵母膏5 g/L,蛋白胨 5 g/L,NaCl 2 g/L,苯乙酸2 g/L ,pH 7.0。培养温度为30℃,培养时间为48 h,摇床转速为180 rmp。发酵结束后,测定其青霉素酰化酶活力,选取酶活力最高的菌株作为产青霉素酰化酶菌株。将该菌保藏于中国典型培养物保藏中心(简称CCTCC),保藏编号为:CCTCC NO:M 2016392。
青霉素酰化酶活力检测方法(以NIPAB为底物)为:取发酵液于12000 rpm、4℃离心10 min,弃上清,菌体用等体积的50mM磷酸缓冲液(pH7.5)重悬,超声破碎细胞后,12000 rpm,4℃离心10 min,取上清为酶液;将酶液用50mM磷酸缓冲液(pH7.5)稀释适当倍数,并用相应的缓冲液配置2mg/ml NIPAB底物溶液;反应体系中先加入100 µL酶液,以灭活的酶液为空白对照,再加入100 µL底物溶液,在酶标仪中进行反应,反应温度为37℃,反应时间为4min,在405 nm波长下检测反应结束时生产的5-氨基-2-硝基苯甲酸5-Amino-2-nitrobenzoic acid(ANBA)的量。每1个单位(U)青霉素酰化酶定义为,在相应条件下,每分钟催化水解NIPAB分解产生1 µmol ANBA所需的酶量。
实施例二
本实验说明青霉素酰化酶PGAPX02编码基因的分离克隆程序。
采用酚-氯仿法抽提菌体总DNA。选取Achromobacter xylosoxidans A8(NCBI Reference Sequence:NC_014640.1)的全基因测序结果进行分析,获得编码无色杆菌来源的青霉素酰化酶的基因,根据该基因序列设计引物:PX02-F1和PX02-R1。
PX02-F1(SEQ ID:3):5′-ATGAAGCAGCAATGGTTGTCGGC-3′;
PX02-R1(SEQ ID:4):5′-TCAGTAGCGCAACGTCTCTACCG-3′。
以木糖氧化无色杆菌Achromobacter xylosoxidans PX02总DNA为模板,以引物PX02-F1和PX02-R1扩增青霉素酰化酶PGAPX02的CDS编码序列。将扩增到的2500 bp的PCR片段连接到pMD18-T载体,进行序列测定和分析,青霉素酰化酶的基因序列如SEQ ID NO:1所示,氨基酸序列如SEQ ID NO:2所示。木糖氧化无色杆菌Achromobacter xylosoxidans PX02与Achromobacter xylosoxidans A8青霉素酰化酶基因同源性为 90%,成熟肽氨基酸序列同源性为94.7 %。
实施例三
本实验说明青霉素酰化酶PGAPX02的表达载体的构建及其在大肠杆(E.coli)BL21(DE3)中的表达。
以PGAPX02总DNA为模板,设计引物,如下:
PX02-F2(SEQ ID:5):5′-ATAGCTAGCATGAAGCAGCAATGGTTGTCGGC-3′(下划线部分为Nhe I酶切位点);
PX02-R2(SEQ ID:6):5′-ATA CTCGAGTCAGTAGCGCAACGTCTCTACCG-3′(下划线部分为Xho I酶切位点)
以PX02-F2和PX02-F2扩增青霉素酰化酶PGAPX02基因,经纯化后用限制性内切酶Nhe I与Xho I进行双酶切,同时以相同限制性内切酶酶切载体pET28a(购自Novagen公司),将酶切产物进行琼脂糖凝胶电泳,回收基因片段和载体,并将基因片段与载体通过T4 DNA连接酶连接,经酶切鉴定及PCR筛选出阳性克隆命名为pET28a-pgapx02,从而构建了表达载体pET28a-pgapx02。
将构建的重组质粒pET28a-pgapx02通过热休克转化法转化大肠杆菌(E.coli)BL21(本实验室保存,特征在于蛋白酶缺陷,T7RNA聚合酶基因)感受态细胞,得到重组菌BL21/ pET28a-pgapx02。同时将pET28a质粒同样转化入BL21感受态细胞,命名为BL21/pET28a,用于青霉素酰化酶PGAPX02表达时的对照菌株。将重组菌BL21/ pET28a-pgapx02及BL21/pET28a接种至含100mg mL-1卡那霉素的LB液体培养基,37℃,180rpm培养过夜后,按2%接种量接种至新鲜LB培养基中(含100mg mL-1卡那霉素),37℃,180rpm培养OD600为0.8~1.0时,加入诱导剂IPTG(终浓度0.1mmol L-1),诱导24 h。分别将诱导后的青霉素酰化酶PGAPX02重组菌破碎后离心,分别取上清和沉淀进行SDS-PAGE,以同样条件下诱导的BL21/pET28a破碎液作为对照,电泳结果如图1所示,其中第一栏为Marker,第二栏为BL21/pET28a加IPTG诱导后细胞破碎液,第三为重组菌BL21/ pET28a-pgapx02加IPTG诱导表达产物破碎上清,第四为重组菌BL21/pET28a-pgapx02加IPTG诱导表达产物破碎沉淀。由此可见经诱导后的重组菌BL21/pET28a-pgapx02破碎后的上清中在27 kD和65 kD左右处均有一明显条带,分别对应α和β亚基。同时分别测定破碎液上清中青霉素酰化酶PGAPX02的酶活,加入诱导剂IPTG诱导后重组菌BL21/pET28a-pgapx02的酶活力为4000U/L,而BL21/pET28a基本不存在青霉素酰化酶PGAPX02酶活。
实施例四
本实验说明青霉素酰化酶PGAPX02的酶学性质。
青霉素酰化酶PGAPX02最适反应pH和pH稳定性的检测:分别配制4.0、5.0、6.0、6.5、7.0、7.5、8.0、8.5、9.0、10.0的缓冲液,将酶液与不同pH缓冲液溶解的NIPAB反应液反应,以pH8.0条件下的青霉素酰化酶活力为对照(100%),分别检测青霉素酰化酶PGAPX02活力,不同pH体系中的酶活如图2所示。从图2中可以看出pH 6.5-8.5范围内具有较高的酶活力,其最适反应pH为8.0。pH稳定性测定:将青霉素酰化酶PGAPX02在不同pH的缓冲溶液中37℃保温2 h后,以最高酶活为100%,测残余酶活,从图3中可以看出该青霉素酰化酶在pH 5.0~10的范围内具有较高的稳定性,在pH 10的溶液中保温2h后,仍然保留最高活力的82%。
青霉素酰化酶PGAPX02最适反应温度和热稳定性的检测:最适反应温度的测定在0.05 M Na2HPO4•12H2O-NaH2PO4•2H2O缓冲体系(pH 7.5)中进行,在不同的温度下以NIPAB为底物进行酶促反应。结果如图4所示,该酶的最适反应温度为55℃。热稳定性测定:青霉素酰化酶PGAPX02在25 °C,35 °C,45 °C,55 °C,65 °C,75 °C下处理2h以后,以最高酶活为100%,测定残留酶活。由图5可以看出该青霉素酰化酶具有较好的热稳定性,该酶在25℃-45℃范围内都能保持较高的青霉素酰化酶活力(相对酶活90%以上)。
青霉素酰化酶PGAPX02的有机溶剂耐受性:考察8种有机溶剂对青霉素酰化酶PGAPX02稳定性的影响,分别将8种有机溶剂缓缓加入酶液中至有机溶剂终浓度为20%(V/V),37℃,180 rpm振荡,定时取样,以NIPAB为底物检测青霉素酰化酶活力。以0h不添加有机溶剂的青霉素酰化酶酶活为对照(100%),测定酶的相对酶活。有机溶剂对青霉素酰化酶PGAPX02酶活影响结果如图6所示。图6结果表明该酶在不添加有机溶剂时,24h后酶活剩80%,而在聚乙二醇和丙三醇中,于37℃处理24h后,酶活分别为101%和112%,说明这两种有机溶剂对酶有一定的激活作用,在DMSO中也表现出一定的耐受性,24h后酶活为60%,由实验结果说明,青霉素酰化酶PGAPX02具有良好的有机溶剂耐受性。
实施例六
本实验说明青霉素酰化酶PGAPX02在催化合成β-内酰胺类抗生素的应用。
青霉素酰化酶PGAPX02催化合成青霉素G类半合成抗生素:以40%丙三醇-0.05 mM磷酸缓冲液(pH 7.0)为反应介质,反应总体积为2mL,其中含有20 mM 6-氨基青霉烷酸,50 mM对羟基苯甘氨酸甲酯(或苯甘氨酸甲酯),加入青霉素酰化酶PGAPX02酶液(实施例三制备)200 µL(8000U/L),在25℃,转速200 rpm条件下反应,定时取样。样品用溶剂(水︰甲醇︰乙酸=60︰32︰8(V/V))稀释10倍,采用HPLC进行检测。结果反应6 h(或4 h),阿莫西林的最大合成率可达65%-70%(或氨苄西林最大合成率可达67%-72%)。
实施例七
本实验说明青霉素酰化酶PGAPX02在催化合成β-内酰胺类抗生素的应用。
青霉素酰化酶PGAPX02催化合成头孢菌素类半合成抗生素:以40%丙三醇-0.05 mM磷酸缓冲液(pH 7.0)为反应介质,反应总体积为2 mL,其中含有20 mM 7-氨基去乙酰氧基头孢烷酸,50 mM对羟基苯甘氨酸甲酯(或苯甘氨酸甲酯),加入青霉素酰化酶PGAPX02酶液(实施例三制备)200 µL(8000U/L),在25℃,转速200 rpm条件下反应,定时取样。样品用溶剂(水︰甲醇︰乙酸=60︰32︰8(V/V))稀释10倍,采用HPLC进行检测。结果反应2 h,头孢氨苄的最大合成率可达72%-76%(或头孢羟氨苄最大合成率可达75%-80%)。
以40%丙三醇-0.05 mM磷酸缓冲液(pH 7.0)为反应介质,反应总体积为2 mL,其中含有20 mM 7-氨基-3- (2-氨基-1,3,4-噻二唑)硫甲基头孢烷酸,50 mM四氮唑乙酸甲酯,加入青霉素酰化酶PGAPX02酶液(实施例三制备)200 µL(8000U/L),在25℃,转速200 rpm条件下反应,定时取样。样品用溶剂(水︰甲醇︰乙酸=60︰32︰8(V/V))稀释10倍,采用HPLC进行检测。结果反应2 h,头孢替唑的最大合成率可达42%-47%。
实施例八
本实验说明扩大反应体系青霉素酰化酶PGAPX02催化合成头孢替唑。
以40%丙三醇-0.05 mM磷酸缓冲液(pH 7.0)为反应介质,反应总体积为20 mL,其中含有100 mM 7-氨基-3- (2-氨基-1,3,4-噻二唑)硫甲基头孢烷酸,250 mM四氮唑乙酸甲酯,加入青霉素酰化酶PGAPX02酶液(实施例三制备)2 mL(8000U/L),在25℃,转速400 rpm条件下反应,自动电位滴定仪检测并用10%的氨水滴定,维持pH 7.0,定时取样。样品用溶剂(水︰甲醇︰乙酸=60︰32︰8(V/V))稀释10倍,采用HPLC进行检测。结果反应6h,头孢替唑的合成率最高可达63%-69%。
结果表明青霉素酰化酶PGAPX02具有良好的合成青霉素G类以及头孢菌素类半合成抗生素的能力,表明该酶将来在合成多种新型β-内酰胺类抗生素中具有巨大的潜力,在生物制药工业中具有广阔的应用前景。
序列表
<110> 南京工业大学
<120> 一株青霉素酰化酶及其编码基因、产生菌和应用
<130> xb16092301
<160> 2
<170> PatentIn version 3.3
<210> 1
<211> 2583
<212> DNA
<213> Achromobacter xylosoxidans PX02
<400> 1
atgaagcagc aatggttgtc ggccgccatc ctggcggcct gtgcggccgt gggcgctcat 60
gcccaggccc aggaaatccc gcagaagact tcgccgcagg ccggtgcctc ggcgcgtccc 120
gctggagcca ccagcggaca ggtcacgata cggcgcgatg gttatggcat gccgcacgtg 180
tacgccaaca ccgtgtatgg cattttctat ggctatggct acgcggtggc gcaggatcgc 240
ccgttccaga tggagatggc gcggcgcagc acccagggcc gcgtggctga agtgcttgga 300
cagcccatgg tggcgttcga caagtccatc cggggaaatt tctcgccaga gcgcatccag 360
cgccagctcg cggcgttgcc ggcatcggag cgccagatcc tggacggcta cgcggccggc 420
atgaatgcct ggatcgcgcg ggtgcgcgcc gagcctggcg gactgatgcc caaggaattc 480
aacgacctgc gcttccagcc ggcggactgg acggcctacg acgtggcgat ggtgttcgtg 540
ggcaccatgg ccaaccgctt ctccgacgcc aatagcgaga tcgacaatct ggcgctgctg 600
accgcgctga aggacaagca cggcgatgag cgcgccatgc agatcttcaa ccagctgcgc 660
tggatgaccg acagccgcgc gcccaccacc gtgccggagg aagagggggt gtaccaaccg 720
gacgccgctc gtccttccgc caggctctcc tatgcgttgc cgcgctacga gggcacgccg 780
cccatgctgg aacgcgtggc gcgcgatccg caaacccgcg gtgtgctgga cgaagcgcct 840
gccgcggtgc cggcgcgact gctggcccag tttgcggaat cggggcaacc gggcatcgcc 900
ggctttccca cgaccagcaa catgtggatc gtgggccgcg accatgccaa ggacgcgcgc 960
gccatcctgc tgaacggacc gcagttcggc tggtggaacc cggcgtatac ctacgggatc 1020
ggcttgcacg gggccggatt cgacgtggtc ggcaatacgc cgtttgccta tcccagcatc 1080
ttgttcggcc acaatgcgca cgtgacctgg ggctcgactg ccggctttgg cgatgacgtg 1140
gacatctacg ccgaaaagct ggatcccaac gaccgcaccc gctatttcca tgatggcgtc 1200
tggaagacga tggaaaagcg caccgagctc atcgaggtga aggacgcgca gccggtggtg 1260
atggatgtct accgcaccgt gcacggcatc gtcacgaagt tcgacgacaa gcagcgcgtg 1320
gcctacgcca aggcgcgcgc ctgggagggc tatgagctgc aatcgctgat ggcctggacc 1380
cacaaggcgc agtcccgcaa ctgggaccag tggaaacagc aggccgcgcg ccacgccctg 1440
accatcaact ggtattacac ggacgacaag ggcaatatcg gctacgcgca tacggggttc 1500
taccccaaac gccgtccggg ccacgatccg cgcctgcccg tgcctggcac cggcgagatg 1560
gattgggacg gcatactgcc tttctcgacc aacccgcagg tgtacaaccc gcgccagggt 1620
tttatcgcca actggaacaa ccagcccatg cgcggctatc cctccaccga ccttttcgcc 1680
atcgtgtggg gccaggcgga tcgctatgcc gaaatcgaga cgcgcctgaa ggccatgaca 1740
gccaatggcg gcaaggtcag cgcgcaacag atgtgggacc tgatccgcac gaccagctat 1800
gccgacgtga accgccgcca tttcctgccg ttcctgcagc aggcggtgca agggctgccc 1860
gcggatgacg cgcgggcgcg gctggttgcc gggctggcat cgtgggacgg catgggtacc 1920
agcgacaagc aaccgggcta ctacgaccac acgggccccg cggtgatgga cgcctggctg 1980
cgcgccatgc tcaagcgcgc gctggccgac gagatgcccg ccgacttctt caagtggtac 2040
agcgccaccg gttatccgac gcaggcggcg cctgctaccg ggtcggtcaa cctgacggtg 2100
ggggtgaagg tgttgttcaa tgcgctggcg ggtcgcgatg cgggcgtgcc acagcagtac 2160
gatttcttca acggccaacg accccaggat gtaacgctgg cggccctgga cgacgcgctg 2220
gcggctttgc gcaaggccta tggcgaggat cccgcccagt ggcgcatacc tgcgcctccg 2280
atggtgttcg cgcccaagaa cttcctgggc gtgccgcagg cggacgacaa ggccgtgttg 2340
agcttcccgg ccacccagaa ccgcggcacc gagaacaaca tgacggtgtt cgatggcaag 2400
ggcgtgcgcg cggtggacgt ggtggcgccg ggacaaagcg gttttgtcgc cccggacggc 2460
accccgtcac cccacgcgcg cgaccagttc gacctttaca ccagtttcgg cagcaagcgg 2520
gtctggttca cggatgcgga agtgcgcgcc cacgctaagt cggtagagac gttgcgctac 2580
tga 2583
<210> 2
<211>860
<212>PRT
<213> Achromobacter xylosoxidans PX02
<400> 2
Met Lys Gln Gln Trp Leu Ser Ala Ala Ile Leu Ala Ala Cys Ala Ala
1 5 10 15
Val Gly Ala His Ala Gln Ala Gln Glu Ile Pro Gln Lys Thr Ser Pro
20 25 30
Gln Ala Gly Ala Ser Ala Arg Pro Ala Gly Ala Thr Ser Gly Gln Val
35 40 45
Thr Ile Arg Arg Asp Gly Tyr Gly Met Pro His Val Tyr Ala Asn Thr
50 55 60
Val Tyr Gly Ile Phe Tyr Gly Tyr Gly Tyr Ala Val Ala Gln Asp Arg
65 70 75 80
Pro Phe Gln Met Glu Met Ala Arg Arg Ser Thr Gln Gly Arg Val Ala
85 90 95
Glu Val Leu Gly Gln Pro Met Val Ala Phe Asp Lys Ser Ile Arg Gly
100 105 110
Asn Phe Ser Pro Glu Arg Ile Gln Arg Gln Leu Ala Ala Leu Pro Ala
115 120 125
Ser Glu Arg Gln Ile Leu Asp Gly Tyr Ala Ala Gly Met Asn Ala Trp
130 135 140
Ile Ala Arg Val Arg Ala Glu Pro Gly Gly Leu Met Pro Lys Glu Phe
145 150 155 160
Asn Asp Leu Arg Phe Gln Pro Ala Asp Trp Thr Ala Tyr Asp Val Ala
165 170 175
Met Val Phe Val Gly Thr Met Ala Asn Arg Phe Ser Asp Ala Asn Ser
180 185 190
Glu Ile Asp Asn Leu Ala Leu Leu Thr Ala Leu Lys Asp Lys His Gly
195 200 205
Asp Glu Arg Ala Met Gln Ile Phe Asn Gln Leu Arg Trp Met Thr Asp
210 215 220
Ser Arg Ala Pro Thr Thr Val Pro Glu Glu Glu Gly Val Tyr Gln Pro
225 230 235 240
Asp Ala Ala Arg Pro Ser Ala Arg Leu Ser Tyr Ala Leu Pro Arg Tyr
245 250 255
Glu Gly Thr Pro Pro Met Leu Glu Arg Val Ala Arg Asp Pro Gln Thr
260 265 270
Arg Gly Val Leu Asp Glu Ala Pro Ala Ala Val Pro Ala Arg Leu Leu
275 280 285
Ala Gln Phe Ala Glu Ser Gly Gln Pro Gly Ile Ala Gly Phe Pro Thr
290 295 300
Thr Ser Asn Met Trp Ile Val Gly Arg Asp His Ala Lys Asp Ala Arg
305 310 315 320
Ala Ile Leu Leu Asn Gly Pro Gln Phe Gly Trp Trp Asn Pro Ala Tyr
325 330 335
Thr Tyr Gly Ile Gly Leu His Gly Ala Gly Phe Asp Val Val Gly Asn
340 345 350
Thr Pro Phe Ala Tyr Pro Ser Ile Leu Phe Gly His Asn Ala His Val
355 360 365
Thr Trp Gly Ser Thr Ala Gly Phe Gly Asp Asp Val Asp Ile Tyr Ala
370 375 380
Glu Lys Leu Asp Pro Asn Asp Arg Thr Arg Tyr Phe His Asp Gly Val
385 390 395 400
Trp Lys Thr Met Glu Lys Arg Thr Glu Leu Ile Glu Val Lys Asp Ala
405 410 415
Gln Pro Val Val Met Asp Val Tyr Arg Thr Val His Gly Ile Val Thr
420 425 430
Lys Phe Asp Asp Lys Gln Arg Val Ala Tyr Ala Lys Ala Arg Ala Trp
435 440 445
Glu Gly Tyr Glu Leu Gln Ser Leu Met Ala Trp Thr His Lys Ala Gln
450 455 460
Ser Arg Asn Trp Asp Gln Trp Lys Gln Gln Ala Ala Arg His Ala Leu
465 470 475 480
Thr Ile Asn Trp Tyr Tyr Thr Asp Asp Lys Gly Asn Ile Gly Tyr Ala
485 490 495
His Thr Gly Phe Tyr Pro Lys Arg Arg Pro Gly His Asp Pro Arg Leu
500 505 510
Pro Val Pro Gly Thr Gly Glu Met Asp Trp Asp Gly Ile Leu Pro Phe
515 520 525
Ser Thr Asn Pro Gln Val Tyr Asn Pro Arg Gln Gly Phe Ile Ala Asn
530 535 540
Trp Asn Asn Gln Pro Met Arg Gly Tyr Pro Ser Thr Asp Leu Phe Ala
545 550 555 560
Ile Val Trp Gly Gln Ala Asp Arg Tyr Ala Glu Ile Glu Thr Arg Leu
565 570 575
Lys Ala Met Thr Ala Asn Gly Gly Lys Val Ser Ala Gln Gln Met Trp
580 585 590
Asp Leu Ile Arg Thr Thr Ser Tyr Ala Asp Val Asn Arg Arg His Phe
595 600 605
Leu Pro Phe Leu Gln Gln Ala Val Gln Gly Leu Pro Ala Asp Asp Ala
610 615 620
Arg Ala Arg Leu Val Ala Gly Leu Ala Ser Trp Asp Gly Met Gly Thr
625 630 635 640
Ser Asp Lys Gln Pro Gly Tyr Tyr Asp His Thr Gly Pro Ala Val Met
645 650 655
Asp Ala Trp Leu Arg Ala Met Leu Lys Arg Ala Leu Ala Asp Glu Met
660 665 670
Pro Ala Asp Phe Phe Lys Trp Tyr Ser Ala Thr Gly Tyr Pro Thr Gln
675 680 685
Ala Ala Pro Ala Thr Gly Ser Val Asn Leu Thr Val Gly Val Lys Val
690 695 700
Leu Phe Asn Ala Leu Ala Gly Arg Asp Ala Gly Val Pro Gln Gln Tyr
705 710 715 720
Asp Phe Phe Asn Gly Gln Arg Pro Gln Asp Val Thr Leu Ala Ala Leu
725 730 735
Asp Asp Ala Leu Ala Ala Leu Arg Lys Ala Tyr Gly Glu Asp Pro Ala
740 745 750
Gln Trp Arg Ile Pro Ala Pro Pro Met Val Phe Ala Pro Lys Asn Phe
755 760 765
Leu Gly Val Pro Gln Ala Asp Asp Lys Ala Val Leu Ser Phe Pro Ala
770 775 780
Thr Gln Asn Arg Gly Thr Glu Asn Asn Met Thr Val Phe Asp Gly Lys
785 790 795 800
Gly Val Arg Ala Val Asp Val Val Ala Pro Gly Gln Ser Gly Phe Val
805 810 815
Ala Pro Asp Gly Thr Pro Ser Pro His Ala Arg Asp Gln Phe Asp Leu
820 825 830
Tyr Thr Ser Phe Gly Ser Lys Arg Val Trp Phe Thr Asp Ala Glu Val
835 840 845
Arg Ala His Ala Lys Ser Val Glu Thr Leu Arg Tyr
850 855 860
- 还没有人留言评论。精彩留言会获得点赞!