旋转性隔离的翻译元件的制作方法
本申请为2015年07月08日进入中国国家阶段、申请号为201480004301.x、申请日为2014年01月16日、发明名称为“旋转性隔离的翻译元件”的发明专利申请的分案申请。相关申请的交叉引用本申请根据35u.s.c.§119(e)要求2013年1月18日提交的美国临时申请61/754,339号的优先权,并根据35u.s.c.§120要求2013年3月13日提交的美国临时申请13/801,762号的优先权,通过引用将每篇申请的内容整体合并至本文中。
背景技术:
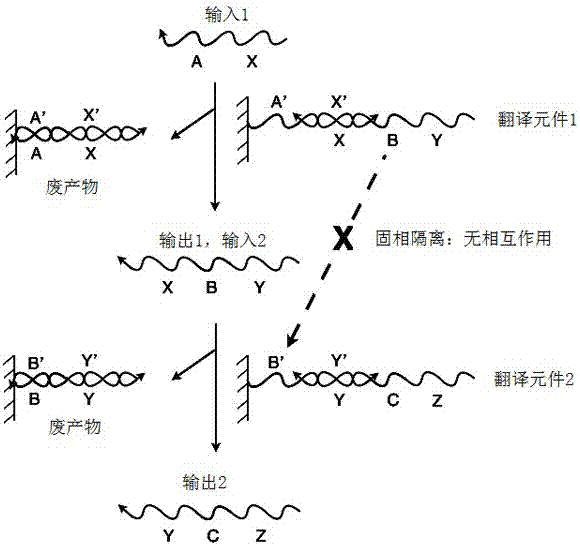
:以分子来制造纳米级计算机具有很大的发展潜力,部分原因在于这种计算机可很好地解决某些计算问题。特别地,使用生物分子的计算机能够与生物学环境相容,使得它们能够用于复杂疾病的诊断,甚至治疗。将一种核酸序列翻译成另一种核酸序列的能力基本上使人们能够利用核酸建立逻辑门和网络。这些门和网络通过两个事件驱动,即杂交和链置换,这两者通常是热动力学上有利的,即它们涉及从较高能量状态到较低能量状态的转变。因此,两个事件可在系统内自发进行。杂交涉及核酸的游离的单链片段。因此,可通过这些游离片段的可获得性来调节核酸网络。“隔离事件(sequesteringevent)”允许某些序列有条件地用于网络的其余部分。这些事件赋予了翻译元件(translators)的构造,翻译元件将一个单链核酸序列转化为一个不同的单链核酸序列。这些翻译元件是利用核酸建立基本逻辑算子的基础,这些基本逻辑算子例如是与(and)、非(not)、或(or)、与非(nand)、或非(nor)和异或非(xnor)。通过这些以及其他逻辑组件,可以建造包括例如放大元件的组件的较大网络。因此,这些翻译事件对于利用核酸的信息处理以及分子计算是重要的。技术实现要素:根据本发明的一个方面,本发明提供一种组合物,其包含第一核酸复合物和第二核酸复合物,每一个均包含第一、第二、第三和第四核酸链,每个核酸链按顺序包含第一、第二和第三片段,其中第一复合物的第一、第二、第三和第四链的核酸链分别被定义为b-x-d,和f-y-c,第二复合物的第一、第二、第三和第四链分别被定义为e-y-d,f-z-g,其中每个字母表示一个片段,每个以“-”连接的一串字母表示一个链,并且其中:第一复合物和第二复合物的每一个都包含形成在第一链的第二片段和第二链的第二片段之间的第一双链区(在第一复合物和第二复合物中分别为x::和::y),形成在第三链的第二片段和第四链的第二片段之间的第二双链区(y::和z::),形成在第一链的第三片段和第三链的第一片段之间的第三双链区(d::和::f),以及形成在第二链的第一片段和第四链的第三片段之间的第四双链区(::c和e::);在第一复合物和第二复合物中的每一个中,第一链的第一片段(在第一复合物和第二复合物中分别为b和)、第二链的第三片段(和d)、第三链的第三片段(和g)以及第四链的第一片段(f和)是单链的;第一复合物的第三链具有与第二复合物的第二链(e-y-d)适当的序列互补性,从而允许它们之间在杂交条件下结合;并且第一复合物的第四链(f-y-c)具有与第二复合物的第一链适当的序列互补性,从而允许它们之间在杂交条件下结合。在一个方面,该组合物进一步包括第三核酸复合物,该第三核酸复合物包含第一和第二核酸链,每个链按顺序包括第一片段、第二片段和第三片段,其中第一链和第二链分别被定义为e-z-h和其中该第三核酸复合物包含形成在第一链的第二片段和第二链的第二片段之间的双链区并且第一链的第一片段(e)和第二链的第三片段是单链的,并且其中:第二复合物的第三链(f-z-g)具有与第三复合物的第二链适当的序列互补性,从而允许它们之间在杂交条件下结合;第二复合物的第四链具有与第三复合物的第一链(e-z-h)适当的序列互补性,从而允许它们之间在杂交条件下结合。在一些方面,第一复合物的第三链的第三片段与第三复合物的第一链的第一片段(e)具有序列互补性,但在正常杂交条件下,由于对这两个片段的任何一个或两个进行化学修饰而使彼此不会稳定结合;并且第一复合物的第四链的第一片段(f)与第三复合物的第一链的第三片段具有序列互补性,但在正常杂交条件下,由于对这两个片段的任何一个或两个进行化学修饰而使彼此不会稳定结合。适于本发明技术的化学修饰包括,但不限于,以结合了mini-peg的丝氨酸来源的γ-pna替换核酸片段的糖-磷酸二酯骨架。在一些方面,化学修饰包括用三环胞嘧啶类似物取代核酸片段中的至少一个核苷的含氮碱基。在一个方面,化学修饰包括在核苷酸的糖基部分的2’位置引入杂原子。这些复合物的图示及其相互关系见图6a,其中翻译元件102对应于第一复合物,翻译元件103对应于第二复合物,翻译元件104对应于第三复合物。类似地,图10-13、14a-d和15-16也图示了本发明范围内的翻译元件(核酸复合物),包括每个单独的翻译元件以及它们的作为翻译元件群组形式(或组合物形式)的组合。在一个方面,每个片段的长度为约5个碱基至约50个碱基。在另一个方面,每个单链片段的长度为约5个碱基至约30个碱基。在本发明的范围内,每种组合物可进一步任选地包含药学上可接受的载体。本发明的一些实施方式进一步提供一种包含本发明的组合物或复合物的细胞。本发明还提供适于模拟、设计、记录、报道或分析本发明公开的技术的某些方面的计算机执行的方法以及非瞬时性计算机可读介质。附图说明附图作为本发明的实施方式来提供,仅作为举例图示,并非作为限制。图1a图示核酸翻译元件的固相隔离实施方式,其中单个区段(a、b等)代表任意长度和序列的寡核苷酸片段。图1b图示核酸翻译元件的固相隔离实施方式如何隔离寡核苷酸片段。图2a图示核酸翻译元件的“立足点(toe-hold)”隔离实施方式。同样地,单个区段代表任意长度和序列的寡核苷酸片段。图2b图示图2a中所示的立足点隔离实施方式如何隔离寡核苷酸片段。图3a显示包含三个立足点隔离的核酸翻译元件的系统。所显示的反应都是链置换反应,都通过图2a所示的分支迁移机制进行。图3b显示具有与图3a中所示的寡核苷酸相同的寡核苷酸系统,但不具有链置换反应,而是显示立足点“碰撞”,其中立足点被不能产生链置换反应的序列结合。这种结合事件占据了立足点,使得期望的链不能结合。图4a-b显示37℃时pbs缓冲液(137mmnacl,2.7mmkcl,10mmna2hpo4,1.76mmkh2po4,ph7.4)中具有立足点(a)或不具有立足点(b)时的交换反应。利用标准曲线将荧光信号转化为浓度。a)双链与10-200nm[(a24)]0滴定液(10nm的增量)孵育(蓝色变红色),通过荧光共振能量转移法(fret)监测游离[f-(a24)]t的浓度。b)双链与滴定液(10nm的增量)(从低到高)混合,的浓度通过fret监测。图5显示用于设计翻译元件的立足点交换法。图6a-6b图示旋转隔离系统中使用的核酸复合物(翻译元件)。图7显示核酸类似物的经修饰的骨架结构,其中“b”代表任意核碱基,(a)显示在dna中发现的天然磷酸二酯骨架,(b)显示肽核酸,(c)显示胍基肽核酸,(d)显示l-丝氨酸来源的γ-pna,(e)显示二酰胺基磷酸酯(在本文中具有吗啉代糖),(f)和)g)显示结合了mini-peg的丝氨酸来源的γ-pna。图8图示了核酸类似物的经修饰的糖结构,其中“b”代表任意核碱基,(a)显示在dna中发现的天然脱氧核糖,(b)显示吗啉基,(c)显示锁核酸,(d)显示氟修饰的rna衍生物。图9图示核酸类似物的经修饰的核碱基结构,(a)甲基胞嘧啶,(b)二氨基嘌呤,(c)吩恶嗪,(d)g形夹。图10示出mrna适配元件(adaptor)。图11显示rnai适配元件。图12图示反义适配元件。图13显示扇入翻译元件群组。图14a-14d示出四个扇出翻译元件群组。这些群组中的两个产生两个或更多个相同的翻译元件,因此起到扩增元件的作用(a和b),而另外两个群组产生两个或更多个不同的下游翻译元件(c和d)。图15图示一个复合条件性翻译元件群组。图16显示一个反相元件(inverter)。图17a-m图示旋转隔离翻译元件(rst)网络和立足点隔离翻译元件(thst)网络关于前景信号和背景信号的模拟结果,其中使用了不同尺寸的立足点,尺寸范围为3个核苷酸(nt)至15nt(如图所示)。所有模拟都以100nm翻译元件进行3个阶段。图18a-d显示rst网络和thst网络在不同网络深度(即涉及的翻译元件/阶段的数量,如图所示)时的模拟结果。所有模拟均以8-nt立足点在100nm翻译元件浓度处进行。图19a-n显示以中等大小网络(5个阶段)进行的长立足点(13nt)和短立足点(8nt)的模拟结果,并且显示网络行为如何随着所示的翻译元件的浓度的改变而变化。某些附图或全部附图都是示例性的图示,因此它们不必然示出所示元件的实际相对尺寸或位置。附图仅为了说明一个或多个实施方式的目的而提供,应当明确理解的是它们不能被用来限制本发明的范围。具体实施方式固相隔离、立足点隔离和立足点交换等多种方法可被用来翻译核酸序列,从而建立逻辑算子和网络。以下更详细描述的这三种具体方法通过利用三程(three-way)、立足点介导的分支迁移反应的几何学得以示例。然而,对于分支迁移反应,其他机制也是可能的,包括但不限于,四程分支迁移、四程加速迁移和多链复合物迁移。因此,虽然为了说明目的以下所述实施方式利用了三程分支迁移,但是本发明预期到dna逻辑门和网络被建立以利用其它分支迁移途径。相反,本发明的实施方式可被应用于任何分支迁移反应。在说明书和附图中,大写字母,例如a、b、c、x、y、z(任选地具有下标或上标),被用来表示任意长度的寡核苷酸的片段。相应的a’、b’、c’、x’、y’、z’,或可互换使用的表示各自的反向互补序列。术语“寡核苷酸”、“多核苷酸”和“核酸”包括所有形式的核酸分子。该类别包括,但不限于,核糖核酸(rna)、脱氧核糖核酸(dna)、肽核酸(pna)及其衍生物,分别可具有和不具有修饰。固相隔离固相隔离通过珠、纳米颗粒或表面将空间内的相关序列/链物理性隔开。这种方法利用了位点分离的原理,其广泛应用于有机化学中。固相隔离几何学中的这些置换事件的发生时序可通过调节所需的链是处于系统的溶液相还是固相中而受控制。图1a显示翻译元件的基本固相隔离装置,翻译元件是允许系统将一种核酸序列替换为另一种核酸序列的组件。对于链a’-x’,a和x的每一个代表任意长度和序列的一段寡核苷酸,x’和a’代表它们各自的反向互补序列。链a’-x’结合至固体载体并且最初与y-b-x杂交,形成不完全碱基配对双链体形式的核酸结构,其可用作翻译元件。在该构造中,链y-b-x是被固相隔离的,因此不能与系统的其余部分相互作用。但是,在链x-a(也称为多核苷酸置换分子)的存在下,链y-b-x可从固体载体被置换出来并暴露于系统的溶液相,而链x-a被结合至载体。该操作包括两个步骤:第一个步骤是互补序列a和a’的杂交(通常称为“立足点结合”)。在第二个步骤中,链x-a的x区结合至链a’-x’的x’区,置换y-b-x的x区,并将该链释放到溶液中,而让x-a与固体载体结合(该步骤通常称为“分支迁移反应”)。这个两步骤程序有效地将游离x-a链翻译成游离y-b-x链。图1b显示的系统具有输入链x-a,其与不完全碱基配对的双链体(“翻译元件1”)相互作用。输出包括完全碱基配对双链体a’-x’/x-a(被认为是“废”产物),以及输出链y-b-x(被称为“输出1”并且可被作为进一步反应中的“输入2”)。“输入2”与“翻译元件2”相互作用,从而产生“输出2”和另一种废产物。在该图中,链y-b-x的b区图示序列在该网络中的隔离。开始时,y-b-x不能与翻译元件2的b’区杂交,因为两者被分离在单独的固体载体上。当输入1结合至翻译元件1并释放y-b-x到溶液中时,y-b-x随后可与翻译元件2相互作用。因此,y-b-x和翻译元件2相互作用的能力取决于输入1的存在。由于位阻效应,结合至一个固体表面的链与结合至另一固体表面的链极其缓慢地相互作用。因此,溶液相中的链是能够与固相算子相互作用的唯一组件。立足点隔离立足点隔离与固相翻译元件实现相同的操作,但通过将多段序列保持结合在双链体中而起作用。正如在固相翻译元件中一样,置换事件可游离出感兴趣的序列。所有链都可以共同在溶液中,其结果是,置换时间通过立足点的可获得性而被调节。术语“立足点”在本文中是指提供置换事件的起点的单链核酸序列的短片段。图2a显示立足点隔离的翻译元件,其类似于图1a中的翻译元件,但是其基于立足点隔离而非固相隔离。在该实施例中,翻译元件的a’区是立足点,其结合输入链并允许链置换反应进行。图2b显示基于立足点的系统,其具有与不完全碱基配对双链体“翻译元件1”相互作用的输入链a-x-b。输出包括废产物(即完全碱基配对双链体b’-x’-a’/a-x-b)和“输出1”链x-b-y-c(可用作进一步反应的“输入2”)。“输入2”与“翻译元件2”相互作用,并产生“输出2”和另一废产物。在该图中,x-b-y-c的b区在翻译元件1中通过杂交至互补的b’区而被隔离,因而不能与翻译元件2的b’区相互作用。x-b-y-c与翻译元件2的相互作用能力取决于系统内输入1(a-x-b)的存在。虽然立足点几何学可能非常有用,但是其用途到目前为止受到包含这种立足点几何学的系统能够传播信息的速度的限制。常规立足点隔离法中的固有限制常常可以将形成的传播降低至生物学上有用的时间尺度以下。更具体地说,由于碰撞相互作用,立足点隔离的翻译元件仅在很窄的浓度动态范围内以适宜的速度起作用。通常调整立足点的长度,可使碰撞相互作用在较低浓度时成为可逆的。但是,当浓度增加时,碰撞将占主要部分,系统将会慢慢停止。另一方面,当浓度降低时,立足点介导的置换也将慢慢停止。在这个意义上,要注意的是,在生物体内,浓度变化很大,因而这样的翻译元件很难适用于试剂的生物体环境。动力学瓶颈被认为是由非生产性反应造成的,非生产性反应在本文也称作“立足点碰撞”,其发生于立足点被不能产生置换反应的具有互补序列或“碰撞链”的分子结合时。例如,图3a显示了具有三个立足点隔离的核酸翻译元件的系统,其与图2a中的系统非常类似。但是,如果三个链都共同处于溶液中,可能发生其他结合事件。图3b图示了可能发生的一些非生产性结合事件或碰撞。通过引入“多核苷酸碰撞分子”,这些事件不会导致置换反应,但可使系统减缓,因为碰撞链的发生阻碍可产生置换反应的链结合。当核酸翻译元件的浓度在系统内升高时,这种碰撞事件可变得更加占优势。这种方法的另一个限制在于由翻译元件的大的单链区的暴露导致的潜在毒性。例如,当单链区结合至细胞的内源性dna或rna时,它可改变该dna或rna分子的转录、翻译或其他功能。这种“非靶标”事件可导致细胞内的不期望的后果。还有一个限制在于潜在的信号泄漏,这是由翻译元件在不存在暴露的立足点的情况下的背景反应导致的,这也可能导致不期望的后果。例如,图4a显示了翻译元件和立足点之间的反应性随着立足点浓度的升高而上升。但是,在不存在立足点的情况下,也观察到了背景反应性(图4b)。一种建议的解决方案是保持立足点较短从而减轻碰撞对系统的影响:立足点越短,互补序列的开关速度越快。因此,常见的是5个或6个核苷酸长度的立足点,因为在这些长度时,如果非生产性结合事件发生,双链的“碰撞”状态持续的时间则短。但是,该方法造成了前述的动力学瓶颈,因为生产性结合事件受到相同的热动力学参数的限制;因此,加入的链同样不会牢固地结合至这些立足点。因此,当结合了正确的加入链时,期望的置换并非总是出现,因为它需要处于结合状态足够长的时间,以启动置换反应。因此,短立足点的使用增加了出现特定操作和产生输出所需的时间量。换言之,在发生许多结合事件(包括碰撞链结合和期望链结合)之前,置换反应不会发生。这种低效率限制了该系统的实用性,其将信息的传播减慢到太长而无不能利用的时间尺度。较短的立足点,联合翻译元件上的较短的单链区,也可帮助降低毒性。但是,较短的立足点不能解决信号泄漏的问题。立足点交换“立足点交换”使用于立足点隔离类似的配对相互作用,但使用不同的几何学。立足点交换尝试解决由立足点隔离途径造成的立足点碰撞问题。参考图5,初始立足点a-b仅与该立足点正下方的双链体的单链区部分地互补(即a::)。类似地,当b-c-d立足点通过与a-b立足点的交换反应而产生时,b-c-d立足点也仅与其下方的双链体的单链区部分地互补。这样将不会发生立足点碰撞。但是,立足点交换法也具有固有限制。首先,与立足点隔离类似,翻译元件的长单链区可导致不期望的毒性作用(由与细胞内的内源性核酸非靶标结合导致)。另一个问题涉及翻译的动态浓度,这与立足点碰撞中遇到的问题类似,但不相同。但该问题对于立足点交换而言不太严重,因为相比于产率,它对翻译的速度影响更大。在这种情况下,立足点的长度决定了自发解离的速率,较短的立足点自发解离速度较快。另一方面,长度还决定了正向置换速率,但较长的立足点使正向置换更快。因此,在这些限制因素之间存在一个依赖于浓度的方式的平衡。立足点交换法的一个重要限制在于低产率,因为所有的反应都是双向的,导致对终产物的无青睐性,除非例如整个翻译最终被结合至不可逆反应。旋转隔离的翻译元件本发明提供了一系列翻译元件,其解决了以上详细描述的各种方法带来的问题。利用这些翻译元件,使得非靶标的毒性作用和背景反应性最小化,可避免立足点碰撞,整体产率接近于完全,并且翻译速度被显著提高。图6a图示了“旋转隔离翻译元件”系统中的某些基本组件。在顶部的是形成在两个链a-x-c和之间的核酸复合物(101)。借助它们的序列互补性,形成有双链区x::而a、c、和仍为单链。这种部分双链体下文被称为“双立足点”。图6a中显示的第二核酸复合物(102)由四个核酸链构成:b-x-d、和f-y-c。如图所示,这些链在它们之间形成四个双链区,在端部形成四个单链区。这四个链中的一个,与第一复合物中的其中一个链a-x-c互补,另一个链b-x-d与第一复合物中的另一个链互补。借助于突出的单链区的序列互补性,第一复合物(101)具有适当的序列互补性,从而能够结合第二复合物(102),这启动了涉及6个链的链置换反应。链置换反应的输出包括两个完全退火的双链体(105和106)以及一个新的双立足点(107),该新的双立足点具有在中部的双链区以及四个独立的单链区(图6b)。图6b显示输出核酸复合物(105、106和107)处于较低能量状态,因为存在两个完全的双链体a-x-c::(105)和b-x-d::(106)。换言之,这种链置换强烈地倾向于正向方向,因此不容易逆转。新形成的双立足点,由和f-y-c构成(107),随后可启动与图6a中的第三核酸复合物(103)的另一个链置换反应(箭头所示),从而继续翻译程序。类似地,从该置换反应生成的新的双立足点可靶定第四复合物(104),依此类推。如以上所解释的,这种方式的链置换反应达到了高的产出速度,因为每个反应以比输入核酸分子低得多的能量状态产生输出产物。而且,如表1所示,实验数据显示这种双立足点介导的反应也可以很快。将“双立足点”栏与“3’立足点”或“5’立足点”中的那些进行比较。表1.单立足点和双立足点介导的链置换反应的反应速度比较根据这些发现,本发明提供大量的翻译元件和翻译元件群组(有时也称作“适配元件”)。翻译元件或翻译元件群组利用一个核酸分子(例如单链核酸、完全或部分双链核酸、或更复杂的核酸结构或翻译元件)作为输入,并进行一个或多个链置换反应,以产生一个或多个核酸分子作为输出。每个输入或输出可作为系统内其他翻译元件或翻译元件群组的输出或输入。因此,每个翻译元件和翻译元件群组起到一个逻辑算子的作用,它们组合起来执行期望的计算和信息传送。因此,本发明的一个实施方式提供一种组合物,其包含第一核酸复合物和第二核酸复合物,每个均包含第一、第二、第三和第四核酸链。每个链按顺序(即从5’至3’,连续地或用中间核苷酸隔开)第一片段、第二片段和第三片段,其中对于第一复合物的第一、第二、第三和第四链,核酸链分别被定义为:b-x-d、和f-y-c,对于第二复合物的第一、第二、第三和第四链,核酸链分别被定义为:e-y-d、f-z-g、每个字母表示一个片段,以“-”连接的每个字母串表示一个链。在一个方面,第一复合物和第二复合物的每一个包含形成于第一链的第二片段和第二链的第二片段之间的第一双链区(在第一复合物和第二复合物中分别为x::和::y)、形成于第三链的第二片段和第四链的第二片段之间的第二双链区(y::和z::)、形成于第一链的第三片段和第三链的第一片段之间的第三双链区(d::和::f)、以及形成于第二链的第一片段和第四链的第三片段之间的第四双链区(::c和e::)。在第一复合物和第二复合物的每一个中,第一链的第一片段(在第一复合物和第二复合物中分别为b和)、第二链的第三片段(和d)、第三链的第三片段(和g)和第四链的第一片段(f和)都是单链的。而且,第一复合物的第三链具有与第二复合物的第二链(e-y-d)合适的序列互补性,从而允许它们在杂交条件下结合;并且第一复合物的第四链(f-y-c)具有与第二复合物的第一链合适的序列互补性,从而允许它们在杂交条件下结合。在一个方面,所述组合物进一步包含第三核酸复合物,其包含第一核酸链和第二核酸链,每个链按顺序包含第一、第二和第三片段,其中第一链和第二链分别被定义为e-z-h和其中第三核酸复合物包含形成于第一链的第二片段和第二链的第二片段之间的双链区(z::),并且第一链的第一片段(e)和第二链的第三片段是单链的,并且其中:第二复合物的第三链(f-z-g)具有与第三复合物的第二链合适的序列互补性,从而允许它们在杂交条件下结合;并且第二复合物的第四链具有与第三复合物的第一链(e-z-h)合适的序列互补性,从而允许它们在杂交条件下结合。化学修饰以避免立足点碰撞本发明还提供了防止立足点碰撞的方法。如图6a所示,由于暴露出的单链区c/和/d,立足点碰撞仍可发生于例如立足点(101)和复合物(103)之间。在一个这样的方法中,每个可能在翻译过程中以单链被暴露出来的核酸片段都经受潜在的化学修饰。除了其自然状态(状态n,在例如cn中以下标表示),每个这样的片段都可被修饰而形成两种状态,分别为状态r(例如右旋)和状态l(左旋),只要这样的修饰允许(或促进)和阻止(或抑制)特定状态的互补链之间的结合,如下表2所示。表2.化学修饰的结合目的其中一个链的修饰互补链的修饰允许/抑制结合nn允许nr允许nl允许rr允许rl抑制ll允许通过这样的修饰,在一个方面,第一复合物102的第三链的尾部片段具有与第三复合物(104)的第一链的第一片段的头部片段(er)相反的序列互补性,但不会稳定地与其结合。同样,在另一个方面,第一复合物(102)的第四链的头部片段(fl)具有与第三复合物(104)的第一链的第一片段的尾部片段相反的序列互补性,但不会稳定地与其结合。因此,不会发生立足点碰撞,并且这些修饰保证了置换反应以受控的方式进行。因此,在一个方面,术语“旋转”是指调节隔离的修饰(l和r),而非所使用的交换反应。如本文所使用的,术语“不会稳定地与其结合”表示两个核酸链不会形成稳定的双链体结构,即使它们共享序列互补性。在一些方面,“稳定的”双链体结构是指能够以非瞬时的形式存在的双链体并且在反应溶液中的浓度高于每个单独的链的总浓度的0.1%、或1%、或5%、或10%。如下文进一步描述的,能够实现表2中定义的结合目的的化学修饰是已知的。“化学修饰”在本文中并不限于在核酸分子产生后对其进行化学性的修饰;该术语还包括在产生该核酸分子期间引入经化学修饰的核苷酸。有许多已充分研究的核酸修饰可被用来调节与天然dna或rna结合的热动力学性质。它们包括对寡核苷酸的骨架、糖或核碱基的改变。这些修饰可单独使用或彼此结合使用;即,使用经修饰的骨架并不排除在相同的链中使用经修饰的核碱基。核酸骨架类似物可被用来改善能够产生置换反应的链的结合。有许多不同的类似物可供使用,所有这些都提供比天然核酸更加紧密的对dna和rna的结合亲和性。这些类似物包括但不限于具有不带电的骨架(肽核酸或二酰胺基磷酸酯)、带正电的骨架(胍基肽核酸)、允许预组构(pre-organization)的氢结合基团(γ肽核酸)的核酸。在一个方面,骨架包括结合了mini-peg的丝氨酸衍生的γ-pna,如图7f-g所示。某些类似物的大体结构见图7。这些类似物都提高核酸杂交反应的热动力学,允许更紧密的立足点结合,因而允许更快的置换。通过在逻辑网络的特定位置使用这些类似物,与碰撞相互作用相比,可强有力地朝期望的置换反应方向进行,因而加快期望的链置换反应的速率。使用经修饰的糖环也可改变寡核苷酸结合dna或rna的热动力学。使用最广泛的类似物是吗啉代、锁核酸(lna)和lna衍生物。其他经修饰的糖在文献中被证明在改变结合热动力学方面也可以产生类似效果。它们的例子为在1’、2’、3’或4’位具有修饰的糖以及核糖环戊烷环中的氧被不同原子取代的糖。这些类似物图示于图8中。核碱基修饰也可用来实现与骨架和糖类似物相同的作用;即,改变特定杂交反应的热动力学。这些碱基包括甲基胞嘧啶、二氨基蝶呤、g形夹和吩恶嗪(图9),它们均提高链对其反向互补序列的结合亲和性。核碱基修饰存在的另一个机会涉及伪互补碱基。这类碱基类似物彼此形成弱碱基对,但与标准碱基形成强碱基对。一个这样的碱基对是2-氨基腺嘌呤(na)和2-硫代胸腺嘧啶(st)。这些碱基可用于有利于一条链结合同时不利于另一条链结合,这是提高生产性结合事件几率同时降低碰撞几率的一个例子。可用于改变结合相互作用的热动力学的另一种化学修饰是引入荷电聚合物,如壳聚糖,其已在文献中被显示促进置换反应的速率。但是,因为这些聚合物非特异性地加速反应,所以它们必须结合上述其他修饰中的一种来应用,以能够区别期望的和不期望的结合事件。可使用不同的方法来合成具有上述化学修饰的分子。例如,设计经修饰的分子时可以考虑骨架化学。骨架化学用于将单个单体结合至较长的链中。涉及核碱基或糖但保持dna/rna的天然磷酸二酯骨架的修饰可以通过如用于天然单体的标准亚磷酰胺化学合成。这些方法的例子见于例如beaucage,s.,andr.iyer,tetrahedron48:2223(1992),inbrown,d.m.a,“briefhistoryofoligonucleotidesynthesis,”20methodsinmolecularbiology(protocolsforoligonucleotidesandanalogs)1–17(1993),inreese,colinb.,organic&biomolecularchemistry3:3851(2005),和iyer,r.p.;ands.l.beaucage,“7.05.oligonucleotidesynthesis,”7comprehensivenaturalproductschemistry(dnaandaspectsofmolecularbiology)105-52(1999)。前述公开文献的各自内容通过引用以整体形式合并至本文中。如果骨架以特定修饰被改变,则使用不同的化学。这种修饰化学在科学文献中有描述。因此,肽核酸(pna)及其衍生物依赖于酰胺键而使各个单体连接在一起。因此,并非利用亚磷酰胺化学,而是通过酰胺结合形成条件和偶联剂(如hbtu)来制备这些单体的链。制备pna或pna样寡核苷酸的方法可见于例如f.beck,“solidphasesynthesisofpnaoligomers,”methodsinmolecularbiologyseries(peptidenucleicacids),humanapress,http://www.springerlink.com/content/mr571738x7t65067/。这些化学修饰包括向dna或pna片段中引入旋向性。dna(特别是pna)的旋向性是一个已充分表征的特性,产生旋向性的方法也是已知的。参见例如corradinietal.“controlofhelicalhandednessindnaandpnananostructures,”methodsmolbiol.749:79-92(2011)。左旋dna或pna被阻止与右旋dna或pna结合,然而两者都可结合至它们的天然副本。另一种骨架修饰方法涉及嵌合寡核苷酸。这些是在相同分子中包含不同骨架化学的寡核苷酸链。例如,如果需要一半是pna骨架、一半是dna骨架的链,则需要一种方式来将这两种骨架化学连接起来。制备这些嵌合链在本领域中通常是已知的。在上面pna/dna嵌合体的例子中,化学差异可通过利用修饰的nda或pna单体被桥接。对于dna,5’-二甲氧基三苯甲基(dmt)保护的羟基被单甲氧基三苯甲基(mmt)保护的胺(在去保护后可以与pna的羧酸反应)取代。对于pna,受保护的n端氮被dmt保护的羟基(在去保护后可以与dna上的亚磷酰胺基团反应)取代。这些方法在例如e.uhlmannetal.,angew.chem.(int’led.)37:2796–823(1998)中有进一步描述。所有这些修饰,无论单独使用或彼此结合使用,都能够影响任意核酸网络中的特定相互作用的热动力学条件,从而在不改变序列组成的情况下,相对碰撞相互作用,有利于期望链或复合物的结合。所有这些相互作用可应用至任何分支介导的迁移反应,无论它们是3程分支迁移(如固相隔离、立足点隔离、或立足点交换)或是通过其他机制进行的分支迁移(例如四程分支迁移、四程加速分支迁移或多链复合物迁移)。适配元件和翻译元件群组在本发明中提供的翻译元件可被用来构建核酸计算机,其可用于医学用途。例如,翻译元件可“感测”到病毒核酸的存在,随后进行一系列翻译事件,导致调节rna(如sirna、反义rna)的释放,其可被用来启动宿主细胞的抗病毒响应或凋亡,从而消除病毒。i.mrna适配元件“感测”可实施为启动一个或多个链置换反应,利用一个核酸(如病毒dna/rna、肿瘤dna/rna)作为输入,并释放出一个或多个立足点作为输出。进行这样的一个或多个链置换反应的翻译元件被定义为“适配元件”。例如,在感测肿瘤mrna的情况下,这种适配元件可被称为“mrna适配元件”。图10图示了一个mrna适配元件。在该mrna(201)上有一个特别的序列,其具有一段片段或该mrna的第一核酸复合物(202)被设计以包括两条链,其中一条具有与该mrna的互补序列(s-q-p)。另一条链,如图10所示,与第一条链形成双链区(q-p::)并且具有一个单链区a-该mrna适配元件中的第二复合物(203)包括至少三条链。第一链(p-b-)与复合物202的第二链部分地互补,从而与第二链和第三链形成两个独立的双链区(::a和b::)。在缺少第一链的情况下,第二链和第三链以上述双立足点的形式存在。因此,在包含具有复合物202和203的mrna适配元件的细胞中,一旦出现mrna分子201,该mrna启动与复合物202的链置换反应,从而释放出第二链(a-)。所释放的第二链随后靶定复合物203,导致链置换反应,释放出双立足点形式的复合物(204),其包括第二链和第三链。该双立足点(204)随后能够启动如本文所述的额外链置换反应。参见例如图6a。这种“mrna适配元件”并不仅仅感测mrna分子。它可以感测任何单链核酸,包括但不限于病毒dna和突变的病毒dna。因此,本发明提供一种组合物,其包含第一核酸复合物(例如图10中的202)和第二核酸复合物(203)。该第一复合物(202)包含(a)第一链,按顺序由第一片段(s)、第二片段(q)、第三片段(p)和第四片段(q)组成,和(b)第二链,按顺序由第一片段(a)、第二片段第三片段和第四片段组成。该第一复合物包含形成于第一链的第二及第三片段(q-p)和第二链的第三及第四片段之间的双链区。并且,该第二复合物(203)包含(a)第一链,按顺序由第一片段(p)、第二片段(b)和第三片段组成,它们分别具有与第一复合物的第二链的第一片段(a)、第二片段和第三片段适当的序列互补性,从而允许它们在杂交条件下结合,(b)第二链,按顺序由第一片段(a)、第二片段(x)和第三片段(c)组成,以及(c)第三链,按顺序由第一片段第二片段和第三片段组成。该第二复合物(203)包含形成于第一链的第三片段和第二链的第一片段(a)之间的第一双链区,形成于第一链的第二片段(b)和第三链的第三片段之间的第二双链区,以及形成于第二链的第二片段(x)和第三链的第二片段之间的第三双链区。如图10所示,可预期到对可暴露作为单链的核酸片段的化学修饰,以降低或避免立足点碰撞。根据本发明的一个方面,本发明的一个实施方式提供一种组合物,其包含:(a)第一核酸复合物,包含(i)第一核酸链,按顺序包括第一片段、第二片段和第三片段并且被定义为s-q-p,其中每个字母表示一个片段,以“-”连接的字母串表示一个链,和(ii)第二链,按顺序包括第一片段、第二片段、第三片段和第四片段并且被定义为a-其中第一复合物包含形成于第一链的第二及第三片段和第二链的第三及第四片段之间的双链区(q-p::);和(b)第二核酸复合物,包含(i)第一核酸链,按顺序包含第一片段、第二片段和第三片段并且被定义为p-b-(ii)第二核酸链,按顺序包含第一片段、第二片段和第三片段并且被定义为a-x-c,和(iii)第三链,按顺序包含第一片段、第二片段和第三片段并被定义为其中该第二复合物包含形成于第一链的第三片段和第二链的第一片段之间的第一双链区(::a),形成于第一链的第一片段和第三链的第三片段之间的第二双链区(b::),和形成于第二链的第二片段和第三链的第二片段之间的第三双链区(x::),其中第一复合物的第一链的第一片段(s)、第一复合物的第二链的第一(a)和第二片段、第二复合物的第三链的第一片段(p)、第二复合物的第二链的第三片段(c)、以及第二复合物的第三链的第一片段是单链的;并且其中第一复合物的第二链的第一、第二和第三片段(a,)具有与第二复合物的第一链的第三、第二和第一片段(b,p)适当的序列互补性,从而分别允许它们在杂交条件下结合。在一个方面,第一复合物的第一链(s-q-p-r)具有与病原性核酸的核酸片段适当的序列互补性,从而允许它们在杂交条件下结合。在一个方面,病原性核酸是病毒dna、病毒rna、细菌dna、细菌rna、突变肿瘤dna或肿瘤rna。ii.rnai适配元件本发明进一步提供一种适配元件,其利用一个核酸分子作为输入,并将rnai(例如sirna)分子作为输出,因而当释放后,该sirna可进行预期的生物学功能。这种适配元件图示于图11中并包括复合物303和304。在图11中,复合物301代表双立足点,其可通过上游翻译元件产生,并且可靶定复合物302以进行链置换反应,从而导致新的双立足点的释放,该新的双立足点包含该复合物的远端半部的两个核酸链。复合物303类似于图6a中的翻译元件102-104,但与它们不同的是,复合物303包括两个rnai分子(和irna),这两个rnai分子在远端(即图中所示右侧)结合至单链。另一方面,另一复合物(304)类似于一个双立足点(例如图6a中的立足点101),但区别在于在远端包括两个rnai分子(和j)。因此,当将复合物302的远端(即右侧)半部作为新的双立足点释放后,该新的双立足点启动与复合物303的链置换反应,导致释放出包含该复合物的远端部分以及rnai链的复合物。这种新的复合物随后可启动与复合物304的链置换反应,导致释放出所有的rnai分子(和j)。因此,在一个实施方式中,本发明提供一种组合物,其包含第一核酸复合物,包含(a)第一、第二、第三和第四核酸链,每个链按顺序包含第一、第二和第三片段,以及(b)第一和第二rna链,其中第一复合物包含:(i)形成于第一链的第二片段和第二链的第二片段之间的第一双链区;(ii)形成于第三链的第二片段和第四链的第二片段之间的第二双链区;(iii)形成于第一链的第三片段和第三链的第一片段之间的第三双链区;(iv)形成于第二链的第一片段和第四链的第三片段之间的第四双链区;(v)形成于第三链的第三片段和第一rna之间的第五双链区;和(vi)形成于第四链的第一片段和第二rna之间的第六双链区,并且第一链的第一片段和第二链的第三片段(d)是单链的。在一个方面,所述组合物进一步包含第二核酸复合物,包含第一和第二核酸链以及第一和第二rna链,其中:第二复合物包含形成于第一链的第二片段和第二链的第二片段之间的第一双链区、形成于第一链的第三片段和第一rna链之间的第二双链区、以及形成于第二链的第一片段和第二rna链之间的第三双链区,其中第一链的第一片段(e)和第二链的第三片段是单链的;第一复合物的第三链与第二复合物的第二链具有合适的序列互补性,从而允许它们在杂交条件下结合;并且第一复合物的第四链与第二复合物的第一链具有合适的序列互补性,从而允许它们在杂交条件下结合。iii.反义适配元件适配元件也被用来接收来自上游翻译元件的信号并释放出反义rna。参考图12,复合物401代表典型的双立足点(如上所述),复合物402类似于典型的旋转隔离翻译元件(如图6a中的102-104),只是任选地,其中一个链具有较长的单链区(f)。这种较长的单链区(f)有助于维持翻译元件403的稳定性,同时允许其他单链区足够的短。同样,它也允许翻译元件402的单链区足够的短,以使翻译元件402和靶标404之间的不期望的反应最小化。因此,当通过由立足点401启动的链置换反应从复合物402释放出后,该复合物(402)的远端半部作为新的双立足点靶定复合物403并释放出包括f-z和的部分双链体。因此,当接触包含e-z片段的细胞rna分子,链随后自由地结合至细胞rna,进行链置换,并让f-z链留在溶液中。与细胞rna之间的结合可导致细胞rna的活性抑制(例如翻译和剪切),其中片段起到了反义核酸的作用。这种设计的独特优点在于链可以很长。在反义应用中已知的是,反义核酸越长,其阻止翻译越有效。但是,不幸的是,在常规技术中,较长的反义核酸导致更严重的非靶标效应(例如毒性)。这是因为这些反义核酸是单链的。本发明提供一种部分双链体形式(即任何一条链或两条链的一部分是单链的)的反义核酸(即链)。因此,该反义核酸即使长度很长,在该反义核酸结合至靶标细胞rna之前,仅有小部分以单链形式暴露于溶液中。因此,这种减少的暴露降低了潜在的非靶标效应。因此,在本发明中,具有大于例如30nt、35nt、40nt、45nt、50nt、55nt、60nt、65nt、70nt、80nt、90nt或100nt的长度的反义核酸可适合引入至生物学系统中。因此,一个实施方式提供了一种组合物,其包含:(a)第一核酸复合物,包含(i)第一核酸链,按顺序包含第一、第二和第三片段并被定义为b-x-d,(ii)第二核酸链,按顺序包含第一、第二和第三片段并被定义为(iii)第三链,按顺序包含第一、第二和第三片段并被定义为和(iv)第四链,按顺序包含第一、第二和第三片段并被定义为f-y-c,其中第一复合物包含形成于第一链的第二片段和第二链的第二片段之间的第一双链区(x::)、形成于第三链的第二片段和第四链的第二片段之间的第二双链区(::y)、形成于第一链的第三片段和第三链的第一片段之间的第三双链区(d::)、以及形成于第二链的第一片段和第四链的第三片段之间的第四双链区(::c);以及(b)第二核酸复合物,包含(i)第一核酸链,按顺序包含第一、第二和第三片段并被定义为(ii)第二核酸链,按顺序包含第一、第二和第三片段并被定义为e-y-d,(iii)第三链,按顺序包含第一和第二片段,并被定义为f-z,和(iv)第四链,按顺序包含第一和第二片段并被定义为其中第二复合物包含形成于第一链的第二片段和第二链的第二片段之间的第一双链区(::y)、形成于第三链的第二片段和第四链的第一片段之间的第二双链区(z::)、形成于第一链的第三片段和第三链的第一片段之间的第三双链区(::f)、以及形成于第二链的第一片段和第四链的第二片段之间的第四双链区(e::),其中第一复合物的第一链的第一片段(b)、第二链的第三片段第三链的第三片段以及第四链的第一片段(f),以及第二复合物的第一链的第一片段和第二链的第三片段(d)都是单链的;其中第一复合物的第三链的第一、第二及第三片段(和)具有与第二复合物的第二链的第三、第二和第一片段(d、y和e)合适的序列互补性,从而分别允许它们在杂交条件下结合;并且其中第一复合物的第四链的的第三、第二和第一片段(c、y和f)具有与第二复合物的第一链的第一、第二和第三片段(和)合适的序列互补性,从而分别允许它们在杂交条件下结合。在一些方面,第一复合物的第四链的第一片段(f)比第一复合物的第三链的第三片段长至少5个核苷酸(nt)、或6nt、7nt、8nt、9nt、10nt、15nt、20nt。在一些方面,第二复合物的第四链的第一片段的长度至少为30个核苷酸(nt)、或35nt、40nt、45nt、50nt、55nt、60nt、65nt、70nt或80nt。iv.扇入本发明还提供了一种“扇入(fan-in)”翻译元件群组。一个扇入翻译元件群组可将两个或更多个不同的核酸输入作为输入信号并产生相同的输出。换言之,扇入翻译元件群组可执行or操作。图13显示了一个扇入群组。在所图示的扇入翻译元件群组中,翻译元件502-503以及505-506每个类似于图6a中的102-103并且彼此类似。由于序列差异,它们之间没有干扰。但是,在翻译元件503和506之间,它们共享相同的远端半部(f-z-g/)。这意味着,当它们的上游翻译元件执行一系列的链置换反应时,翻译元件503和506产生相同的靶定复合物507的输出立足点。因此,无论输入是立足点501或504,链置换反应都将在翻译元件507处被触发。因此,在一个实施方式中,本发明提供一种组合物,其包含第一核酸复合物和第二核酸复合物,每个复合物包含第一、第二、第三和第四核酸链,每个链按顺序包含第一、第二和第三片段,其中第一复合物的第一、第二、第三和第四链的核酸链分别被定义为:b-x-d、和f-y-c,第二复合物的第一、第二、第三和第四链的核酸链分别被定义为e-y-d、f-z-g、其中每个字母表示一个片段,以“-”连接的字母串表示一个链,并且其中:第一复合物和第二复合物的每一个都包含形成于第一链的第二片段和第二链的第二片段之间的第一双链区(在第一复合物和第二复合物中分别为x::和::y)、形成于第三链的第二片段和第四链的第二片段之间的第二双链区(y::和z::)、形成于第一链的第三片段和第三链的第一片段之间的第三双链区(d::和::f)、以及形成于第二链的第一片段和第四链的第三片段之间的第四双链区(::c和e::);在第一复合物和第二复合物的每一个中,第一链的第一片段(在第一复合物和第二复合物中分别为b和)、第二链的第三片段(和d)、第三链的第三片段(和g)以及第四链的第一片段(f和)是单链的;第一复合物的第三链具有与第二复合物的第二链(e-y-d)合适的序列互补性,从而允许它们在杂交条件下结合;第一复合物的第四链(f-y-c)具有与第二复合物的第一链合适的序列互补性,从而允许它们在杂交条件下结合;所述组合物进一步包含第三核酸复合物,其包含第一核酸链和第二核酸链,每个链按顺序包含第一、第二和第三片段,其中第一和第二链分别被定义为e-z-h和其中所述第三核酸复合物包含形成于第一链的第二片段和第二链的第二片段之间的双链区(z::),并且第一链的第一片段(e)和第二链的第三片段是单链的,并且其中:第二复合物的第三链(f-z-g)具有与第三复合物的第二链合适的序列互补性,以允许它们之间在杂交条件下结合;并且第二复合物的第四链具有与第三复合物的第一链合适的序列互补性,以允许它们在杂交条件下结合;并且所述组合物进一步包含第四核酸复合物和第五核酸复合物,每个复合物包含第一、第二、第三和第四核酸链,每个链按顺序包含第一、第二和第三片段,其中第四复合物的第一、第二、第三和第四链分别被定义为:k-r-m,和f-s-n,第五复合物的第一、第二、第三和第四链分别被定义为:e-s-m,f-z-g和其中:第四复合物和第五复合物的每一个都包含形成于第一链的第二片段和第二链的第二片段之间的第一双链区(在第四复合物和第五复合物中分别为r::和::s)、形成于第三链的第二片段和第四链的第二片段之间的第二双链区(::s和z::)、形成于第一链的第三片段和第三链的第一片段之间的第三双链区(m::和::f)、以及形成于第二链的第一片段和第四链的第三片段之间的第四双链区(::n和e::);在第四复合物和第五复合物的每一个中,第一链的第一片段(在第一和第二复合物中分别为k和)、第二链的第三片段(和m)、第三链的第三片段(和g)以及第四链的第一片段(f和)都是单链的;第四复合物的第三链具有与第五复合物的第二链(e-s-m)合适的序列互补性,从而允许它们在杂交条件下结合;第四复合物的第四链(f-s-n)具有与第五复合物的第一链合适的序列互补性,从而允许它们在杂交条件下结合;第二复合物的第三链(f-z-g)和第五复合物的第三链(f-z-g)具有合适的序列同一性从而允许它们在杂交条件下结合至相同的靶标核酸;第二复合物的第四链和第五复合物的第四链具有合适的序列同一性从而允许它们在杂交条件下结合至相同的靶标核酸。v.扇出与扇入翻译元件群组相反,“扇出(fan-out)”翻译元件群组采用单个输入并产生多个输出核酸。在一个方面,该多个输出核酸具有不同的序列,因而它们可进行不同的功能或下游反应。在另一个方面,该多个输出核酸是相同的,因而该扇出翻译元件群组可作为放大元件。图14a显示了以一个立足点作为输入并产生两个相同的新立足点的扇出翻译元件群组。类似地,图14b中的扇出群组产生四个相同的新立足点。另一方面,图14c-d表示产生两个或更多个不同的新立足点的扇出翻译元件群组。在图14a中,核酸601是如本文所述的典型双立足点。复合物602与图6a的翻译元件102和103的区别在于远端上的单链区更长,该单链区含有两个(而不是一个)连续片段。如复合物603中可明显看出的,每个连续片段群组(a-b)都能够释放新立足点,一旦释放后,该新的立足点具有位于单独的链上的两个不同的单链区a和b。因此,这些新的立足点中的每一个可与翻译元件604启动一个新的链置换反应。图14b的翻译元件类似于图14a中的翻译元件,只是翻译元件606中的单链区各个都具有能够释放出两个新立足点的四个连续片段。图14c中的翻译元件群组可利用与图14a-b(601和605)相同的立足点输入(609),但翻译元件610一旦接触该输入后释放出不同于图14a-b的立足点。如翻译元件611中的相应链所反映出的,这种不同是更加明显的。即,当接触翻译元件610释放的立足点并进行链置换反应时,翻译元件611释放出两个不同的立足点。这两个立足点中的一个能够启动与翻译元件612的新的链置换反应,另一个与翻译元件613启动新的链置换反应。因此,图14b中的翻译元件群组能够由一个立足点输入(609)分别产生来自翻译元件612和613的两个不同的新立足点输出。类似于图14c中的翻译元件群组,图14d的翻译元件群组也可用一个立足点(614)作为输入并产生四个不同的新立足点作为输出,每个分别由翻译元件617-620释放。因此,本发明的一个实施方式提供一种核酸复合物,其包含第一、第二、第三、第四、第五和第六核酸链,其中:第一链按顺序包含第一、第二、第三和第四片段并被定义为其中每个字母表示一个片段,以“-”相连的字母串表示一个链;第二链按顺序包含第一、第二、第三和第四片段并被定义为-p;第三链和第五链每一个按顺序包含第一、第二和第三片段并被定义为a-y-c;第四链和第六链每一个按顺序包含第一、第二和第三片段并被定义为-b;并且其中该复合物包含:形成于第一链的第二片段和第二链的第三片段之间的第一双链区(:x);形成于第一链的第四片段和第三链的第一链之间的第二双链区(::a);形成于第一链的第三片段和第四链的第三片段之间的第三双链区(::b);形成于第二链的第二片段和第五链的第一片段之间的第四双链区(::a);形成于第二链的第一片段和第六链的第三片段之间的第五双链区(::b);形成于第三链的第二片段和第四链的第二片段之间的第六双链区(y::);和形成于第五链的第二片段和第六链的第二片段之间的第七双链区(y::),并且第一链的第一片段第二链的第四片段(p)、第三和第五链的第三片段(c)以及第四和第六链的第一片段都是单链的。类似地,本发明还提供了扇出两个或更多个不同立足点的核酸复合物。vi.复合条件性“复合条件性”翻译元件群组执行and操作。复合条件性群组需要两个单独的输入信号以激活和释放出输出核酸。图15显示了一个复合条件性翻译元件群组,其包括至少三个翻译元件核酸复合物703、704和705。翻译元件703以立足点701作为输入并产生含有703的远端半部的第一新立足点。类似地,翻译元件704以立足点702作为输入并产生含有704的远端半部的第二新立足点。翻译元件705含有至少六条链,从左至右,由三个双链对构成。仅当接触由翻译元件703释放的第一新立足点时,翻译元件705才能进行链置换反应,导致中间双链的两个单链区(和h)的暴露。随后,当与翻译元件704产生的第二新立足点接触时,发生另一个链置换反应,并释放出作为第三新立足点的705的第三双链对。该第三新立足点随后可任选地通过例如翻译元件706出发进一步的反应。因此,在一个实施方式中,本发明提供一种组合物,其包含第一、第二和第三核酸复合物,其中:第一复合物包含第一、第二、第三和第四核酸链,每个链按顺序包含第一、第二和第三片段并且分别被定义为b-w-d,-h和-y-c,其中每个字母表示一个片段,每个以“-”相连的字母串表示一个链,并且其中第一复合物包含形成于第一链的第二片段和第二链的第二片段之间的第一双链区(w::)、形成于第三链的第二片段和第四链的第二片段之间的第二双链区(::y)、形成于第一链的第三片段和第三链的第一片段之间的第三双链区(d::)和形成于第二链的第一片段和第四链的第三片段之间的第四双链区(::c);第二复合物包含第一、第二、第三和第四核酸链,每个链按顺序包含第一、第二和第三片段并分别被定义为f-x-h,和j-z-g,其中第二复合物包含形成于第一链的第二片段和第二链的第二片段之间的第一双链区(x::)、形成于第三链的第二片段和第四链的第二片段之间的第二双链区(::z)、形成于第一链的第三片段和第三链的第一片段之间的第三双链区(h::)以及形成于第二链的第一片段和第四链的第三片段之间的第四双链区(::g);第三复合物包含第一、第二、第三、第四、第五和第六核酸链,每个链按顺序包含第一、第二、第三片段并分别被定义为i-z-h,j-q-m和其中第三复合物包含形成于第一链的第二片段和第二链的第二片段之间的第一双链区(::y)、形成于第三链的第二片段和第四链的第二片段之间的第二双链区(::z)、形成于第五链的第二片段和第六链的第二片段之间的第三双链区(q::)、形成于第一链的第三片段和第三链的第一片段之间的第四双链区(g::)、形成于第二链的第一片段和第四链的第三片段之间的第五双链区(::h)、形成于第三链的第三片段和第五链的第一片段之间的第六双链区(::j)、以及形成于第四链的第一片段和第六链的第三片段之间的第七双链区(i::);其中,在第一复合物中,第一链的第一片段(b)、第二片段的第三片段第三链的第三片段(h)、第四链的第一片段是单链的;在第二复合物中,第一链的第一片段(f)、第二片段的第三片段第三链的第三片段和第四链的第一片段(j)是单链的;在第三复合物中,第一链的第一片段第二链的第三片段(d)、第五片段的第三片段(m)和第六链的第一片段是单链的,并且其中:第一复合物的第三链(-h)和第三复合物的第二链(-y-d)具有合适的序列互补性,从而允许它们在杂交条件下结合;第一复合物的第四链(-y-c)和第三复合物的第一链(-g)具有合适的序列互补性,从而允许它们在杂交条件下结合;第二复合物的第三链和第三复合物的第四链(i-z-h)具有合适的序列互补性,从而允许它们在杂交条件下结合;并且第二复合物的第四链(j-z-g)和第三复合物的第三链具有合适的序列互补性,从而允许它们在杂交条件下结合。vii.反相元件“反相元件”构成一个或多个翻译元件,一旦与输入信号接触,其产生阻止链置换反应的核酸(“停止元件”),否则的话,该链置换反应会通过系统内不同的输入信号启动而发生。如图16所示,反相核酸复合物804由三个链组成,其中两个f-y-d和在中部片段之间形成双链区(y::)。第三链包括两个片段c和每个与其他两个链的一个片段互补。一旦接触立足点802,将发生链置换反应,导致第三链c-的释放。核酸复合物803作为一个翻译元件以立足点801作为输入并产生该复合物的远端半部作为输出(新立足点)。该新立足点如果接触翻译元件805则可以进行后续的链置换反应,可能与翻译元件806进行后续的反应。但是,如果在立足点801与翻译元件803之间的链置换反应发生之前,立足点802接触了反相元件804并释放出停止链c-801-803-805-806的信号处理路径就不会发生。这是因为停止链c-结合至暴露的单链区和d,从而阻止805与由翻译元件803产生的立足点之间的链置换反应。因此,本发明的一个实施方式提供一种组合物,其包含第一和第二核酸复合物,其中:第一复合物包含第一、第二、第三和第四核酸链,每个链按顺序包含第一、第二和第三片段并分别被定义为b-x-d,和j-z-c,其中每个字母表示一个片段,每个以“-”相连的字母串表示一个链,其中第一复合物包含形成于第一链的第二片段和第二链的第二片段之间的第一双链区(x::)、形成于第三链的第二片段和第四链的第二片段的第二双链区(::z)、形成于第一链的第三片段和第三链的第一片段之间的第三双链区(d::)和形成于第二链的第一片段和第四链的第三片段之间的第四双链区(::c);第二复合物包含第一和第二核酸链,每个链按顺序包含第一、第二和第三片段并且分别被定义为f-y-d和并且包含第三核酸,其按顺序包含第一和第二片段并且被定义为-c,其中第二复合物包含形成于第一链的第二片段和第二链的第二片段之间的第一双链区(y::)、形成于第一链的第三片段和第三链的第一片段之间的第二双链区(d::)、形成于第二链的第一片段和第三链的第二片段之间的第三双链区(::c);其中第一复合物的第一链的第一片段(b)、第一复合物的第二链的第三片段第一复合物的第三链的第三片段第一复合物的第四链的第一片段(j)、第二复合物的第一链的第一片段(f)、第二复合物的第二链的第三片段是单链的;并且其中:第二复合物的第三链(-c)和第一复合物的第三链的第一片段及第一复合物的第四链的第三片段(c)的按顺序的组合具有合适的序列同一性以允许它们靶定至相同的靶标核酸序列。对于上述任何翻译元件和翻译元件群组,预期的是,化学修饰可被用来降低或消除立足点碰撞,即,系统内(例如细胞)立足点与任何翻译元件的单链区的非预期结合。这种修饰可利用本发明提供的信息而轻易设计。而且,对于每个翻译元件群组,响应的附图仅提供示例性的修饰(如下标n、r和l所标注的)。这些修饰的期望的性质见表2。在一些方面,要注意的是,如果未明确表示为双链区的一部分,那么核酸片段是指单链的。单链的表示在附图中也是明显的。每个核酸链或片段的长度可通过计算机或实验来确定。表1显示了长度的影响(特别是对立足点的影响)。在一个方面,每个片段为约3个碱基至约50个碱基长。或者,片段为至少约3、4、5、6、7、8、9、10、11、或12个碱基长。在另一个方面,片段不长于约50、45、40、35、30、29、28、27、26、25、24、23、22、21、20、19、28、17、16、15、14、13、12、11、10、9、8、7、6或5个碱基。而且,以单链形式存在的片段(例如立足点)可相对较短,例如约3、4、5、6、7、8、9、10个碱基至约30、29、28、27、26、25、24、23、22、21、20、19、28、17、16或15个碱基。在一个方面,立足点片段为至少约3、4、5、6、7、8、9、10、11或12个碱基长。在另一个方面,立足点片段不长于约20、19、28、17、16、15、14、13、12、11、10、9、8、7、6或5个碱基。相反,形成翻译元件内的双链区的片段可相对较长。方法和计算机建模本发明进一步提供利用本发明的翻译元件进行计算,特别是在生物学环境中传播信息的方法。例如,mrna适配元件可被用来检测病原性核酸或瘤性核酸的存在并产生例如立足点形式的输出信号。如果需要,可使用对多种核酸具有特异性的多个mrna适配元件以确保可靠的检测。系统内的其他翻译元件,例如扇入翻译元件、扇出翻译元件和反相翻译元件,随后可被用来处理mrna适配元件生成的信号,最终导致利用相应的rnai和反义适配元件产生调节核酸,例如rnai和反义rna。通过谨慎地调整处理过程,可精确地控制这些调节rna的严格序列和量。因此,生物学反应(例如凋亡和免疫响应)可通过这些调节rna来出发,带来治疗或诊断效果。容易意识到的是,本文所述的翻译元件、组合物和系统可包括生物学材料,但它们也可利用计算机被数学建模。因此,本发明还提供用于执行这种建模的计算机方法、系统和程序植入介质。因此,对应于本发明的每个翻译元件、组合物或系统,本发明提供了一种计算机执行的方法,其中计算机被配置用来代表它们的序列、结构、合成和链置换反应。本发明描述的方法可在计算机系统或网络上执行。合适的计算机系统可包括至少一个处理器和存储器;任选地,还包括计算机可读介质,用于存储处理器所执行的计算机代码。一旦代码被执行,计算机系统即进行所述方法。关于这点,“处理器”是一种电子回路,其可执行计算机程序。合适的处理器的例子包括但不限于中央处理单元、微处理器、图形处理单元、物理处理单元、数字信号处理器、网络处理器、前端处理器、协处理器、数据处理器和音频处理器。术语“存储器”是指存储数据用于存取的电子装置。因此,在一个方面,合适的存储器为保存数据并辅助计算的计算机单元。更广义上来说,用于提供必需的网络数据传输的合适方法和装置是已知的。本发明还预期到一种非瞬时性计算机可读介质,其包括用于进行本文所述方法的可执行代码。在某些实施方式中,该介质进一步包含该方法所需的数据或数据库。实施例本发明进一步通过该实施例来阐释,该实施例代表了本发明的旋转隔离翻译元件(rst)网络的时序行为的动力学模拟结果,并将其与立足点隔离的翻译元件(thst)网络的动力学模拟进行并行比较,后者在以上“立足点隔离”的子标题下结合图2和图3提到。生成包括系列个n翻译的机制,称作“阶段数目”,其从frezza,b.m.,orchestrationofmolecularinformationthroughhigherorderchemicalrecognition:athesispresented,thescrippsresearchinstitute,lajolla,california(2010),atchapter3中衍生得到它们的正向速率常数。该机制也包括翻译元件的背景“泄漏”(也具有根据frezza(2010)的速率常数)和“碰撞”相互作用。后两者从最邻近的结合自由能衍生得到它们的速率常数,参见allawiandsantalucia,biochemistry36:10581-94(1997),其对于杂交使用标准正向速率常数(每秒每摩尔105)并且利用细致平衡得到反向速率常数。对于每个模拟,对rst网络和thst网络进行前景信号模拟和背景信号模拟。“前景信号”是指,假设初始输入序列与模拟中的其他翻译元件处于相同浓度,所产生的最终输出序列的浓度。“背景信号”表示,假设没有初始输入序列,所产生的最终输出序列的浓度。这些模拟证实了每个网络设计在改变立足点长度、翻译元件浓度、网络规模(阶段数量)的条件下提供的理想化的响应。图17a-m显示了对于不同尺寸的立足点(3至15个核苷酸(nt)),rst和thst的前景信号和背景信号的模拟结果。所有模拟都以100nm翻译元件进行3个阶段。图17显示出,在极短的立足点时,rst实际上产生非常显著的背景信号。但是,随着立足点长度的增加,该背景下降至接近零(8nt和更长时;例如见图17f),然而在所有情况下,thsts具有相当一致并且相对高的背景翻译速率,也成为“泄漏”。而且,随着立足点长度增加超出11个nt,thst上的碰撞相互作用开始主导系统,并急剧地减慢前景活动,而rst名义上以较长的立足点速率操作(参见例如图17i-m)。在另一个系列的模拟中,在100nm翻译元件浓度,对于rst和thst,8nt立足点产生相似的前景信号。该系统使用逐渐增加的网络深度(就阶段数量而言,即翻译元件的数量)来检验。如图18a-d所示,当阶段数量增加时,rst和thst的背景信号都显著增加。但是,这种增加的程度在thst的情况下严重得多。另一个系列的模拟检验了中等规模的网络(5个阶段),使用长(13nt)和短(8nt)立足点,并显示了它们的行为如何随翻译元件的浓度的变化而改变。结果显示于图19a-n中。较高浓度的翻译(10μm)进行得非常快,非常低浓度的翻译(100pm)进行得非常慢。这种行为与rst和thst系统采用二级反应动力学的事实一致,因此,达到平衡的时间取决于系统的总浓度。在长立足点的情况下,对于rst,直至非常低的浓度时,反应时间仍保持生物学相关性,在低至100pm时其仍显示出温和的反应性,然而thst需要高得多的浓度(10μm)来获得任何有意义的活性(图19a-g)。在短立足点的情况下,rst的表现类似,即使是具有显著较高的背景信号(图19h-n)。使用较短的立足点时,thst在低浓度处获得有意义的反应性,但在高浓度时,导致thst背景信号的泄漏却呈灾难性地高。这些数据一起证实了rst提供了设计可能性,利用较长的立足点可将背景(泄漏)活性降低至近乎不可检测的水平,通过极端抑制而不丧失反应性,而thst却面临这个问题。而且,它们显示出,rst可在巨大的动态浓度范围内以理想的前景:背景反应性来起作用,使得它们理想地适用于生物学环境。**************************虽然已描述了本发明的具体实施方式,但是它们仅是本发明的说明,并不限制本发明。阅读本说明书后,本领域技术人员可容易地作出许多变化。本发明的完整范围应当参考权利要求书及其等同的完整范围以及说明书(连同这些变化)来确定。当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1